Филогенез, филогения - историческое развитие мира живых организмов, как в целом, так и отдельных таксономических групп: царств, типов (отделов), классов, отрядов (порядков), семейств, родов, видов. Термин филогенез введен Э. Геккелем.
Современную систему животных, построенную на принципах родства, можно представить как родословное (или филогенетическое) древо, отражающее основные этапы эволюции животных. Филогенез различных групп организмов изучен неравномерно. Наиболее изучен филогенез позвоночных (особенно высших групп), из беспозвоночных лучше изучен филогенез моллюсков, иглокожих, членистоногих. Слабо изучен филогенез прокариот.
У основания эволюционного древа животных находятся одноклеточные. У одноклеточных все функции организма выполняет одна клетка. Существует несколько гипотез происхождения одноклеточных эукариотов: 1) эукариоты произошли от прокариот путем симбиогенез. В пользу данной гипотезы говорит то, что митохондрии и хлоропласты эукариот имеют собственные ДНК; 2) эукариоты произошли за счет дифференциации прокариотических клеток на цитоплазму и органоиды. Все типы современных простейших происходят от древних саркожгутиковых.
Многоклеточные более высокоорганизованные животные, у которых отдельные функции организма выполняют отдельные клетки или органы. Все многоклеточные в индивидуальном развитии начинаются с одной клетки - зиготы, а начальные стадии зародыша (бластула) напоминают колонии одноклеточных.
Существует ряд гипотез происхождения многоклеточных животных. Наиболее широко известными являются гипотезы Э. Геккеля и И.И. Мечникова. Согласно теории Геккеля, все многоклеточные животные произошли от одной прародительской формы - гастреи. И.И. Мечников предполагал, что предком многоклеточных животных была колония жгутиковых. Первый многоклеточный организм имел шаровидную форму, был покрыт слоем клеток со жгутиками и имел внутри тела пищеварительные амебоидные клетки, свободно плавал в воде. И.И. Мечников назвал его фагоцителлой. Дальнейшая эволюция многоклеточных шла в четырех направлениях. Одно из направлений эволюции привело к образованию Типа Пластинчатых, состоящих из одного слоя клеток и ведущих ползающий образ жизни. Другое направление эволюции привело к формированию Типа Губок с двумя слоями клеток, не оформленных в ткани. У них еще нет нервной системы. Это неподвижные донные животные. Третье направление эволюции привело к формированию Типа Кишечнополостных. Кишечнополостные уже настоящие двуслойные (эктодерма и энтодерма) животные с первичной кишкой, ртом, радиальной симметрией, диффузной нервной системой. Среди них имеются свободно плавающие животные и ведущие прикрепленный образ жизни. Четвертое направление эволюции привело к формированию еще более высокоорганизованных трехслойных животных. Ткани и органы этих животных образовались из трех слоев клеток (зародышевых листков) эктодермы, энтодермы и мезодермы.
Первыми трехслойными были животные похожие на бескишечных планарий. Эволюция трехслойных животных шла в трех направлениях. Одно направление привело к образованию плоских червей, у которых отсутствует полость тела, а между органами расположена паренхима. Плоские черви являются первыми билатеральными животными (все последующие типы имеют такую симметрию). У них впервые появляются мышечная система (гладкая), нервные ганглии. Однако, образовавшийся кишечник слепозамкнутый, не имеет анального отверстия.
Другое направление привело к образованию первичнополостных животных круглых червей. Кишечник у них становится сквозным, состоит из трех отделов с анальным отверстием. Происходит дальнейшая концентрация нервных клеток, образуя окологлоточное нервное кольцо. Общим для плоских и кольчатых червей является наличие кожно-мускульного мешка, пищеварительной, выделительной, половой систем.
Третье направление эволюции трехслойных ведет к формированию вторичнополостных животных. Образование вторичной полости тела обеспечило большую подвижность и устойчивость внутренней среды организма. Вторичнополостные животные более высокоорганизованные: имеют кровеносную систему. К ним относятся все остальные беспозвоночные и хордовые животные.
Эволюция вторичнополостных животных привела к образованию первичноротых и вторичноротых. Эволюция первичноротых . Oт предка первичноротых произошли моллюски и первичные кольчатые черви. В свою очередь от первичных кольчатых червей произошли современные кольчатые черви и членистоногие.
Кольчатые черви имеют прогрессивные черты организации: вторичную полость тела, метамерно расчлененное тело, замкнутую кровеносную систему (у других беспозвоночных незамкнутая), окологлоточное кольцо и нервную цепочку.
Членистоногие достигают большего совершенства: в кровеносной системе возникает сердце; в нервной системе усиливается концентрация ганглиев; образуется наружный скелет (хитиновый покров); характерно многообразие органов дыхания (жабры, трахеи, легкие); образуются членистые конечности и крылья. Более совершенное строение членистоногих позволило им широко заселить Землю.
Моллюски имеют вторичную полость тела, сложно устроенные внутренние органы (образуются почки, трехкамерное сердце, жабры, легкие), у большинства известковую раковину, защищающую тело. Однако у моллюсков кровеносная система незамкнутая, а нервная система разбросано узлового типа.
Эволюции вторичноротых . От предков вторичноротых произошли иглокожие и хордовые.
Иглокожие. Для них характерны: вторичная полость тела, кровеносная система образует лакуны, своеобразная вододвигательная (амбулакральная) система, лучевая симметрия, нервная система в форме кольца с радиальными лучами.
Первичные хордовые, вероятно, произошли от вторичноротых червеобразных предков. Обособление хордовых связано с увеличением подвижности. О филогенетических связях хордовых с беспозвоночными животными свидетельствует следующее: 1) ткани и органы образуются из трех зародышевых листков (впервые образуются у плоских червей); 2) билатеральная симметрия (возникает у плоских червей); 3) вторичная полость тела (возникает у кольчатых червей); 4) вторичный рот (родство с иглокожими); 5) замкнутая кровеносная система (кольчатые черви);
От первичных хордовых произошли современные бесчерепные (ланцетник) и позвоночные животные. Личиночнохордовые - это боковая ветвь хордовых животных, которые в большинстве перейти к сидячему обрату жизни, и потому признаки хордовых у них имеются только на личиночной стадии.
Головохордовые - примитивные хордовые животные. У них имеются нефридии, сходные с метанефридиями кольчатых червей, замкнутая кровеносная система без сердца, как у кольчатых червей.
Предки позвоночных появились около 500 млн. лет назад. Их эволюция шла в двух направлениях: бесчелюстные (круглоротые) и челюстноротые (все остальные позвоночные).
Первые челюстные - панцирные рыбы произошли от панцирных бесчелюстных. Они уже имели прочный внутренний скелет, мозговой и висцеральный череп, челюсти с зубами, парные плавники. Эти рыбы вымерли. Их сменили хрящевые и костные рыбы, тело которых было покрыто чешуей.
Хрящевые рыбы были распространены в морях, а по типу питания являлись хищниками.
Костные рыбы первоначально появились в пресных водоемах. У них развилась жаберная крышка (защищает жабры от засорения) и плавательный пузырь (облегчает плавание в пресной воде). В пересыхающих водоемах (в палеозое) появились лопастеперые рыбы (двоякодышащие и кистеперые), у которых дополнительно к жабрам развились легочные мешки для воздушного дыхания, а образовавшаяся на плавниках мускулатура позволяя опираться о твердый субстрат. В дальнейшем костные рыбы заселили пресные и морские водоемы. В настоящее время костистые рыбы одна из самых процветающих групп позвоночных животных (20 тыс. видов).
Земноводные появились в девоне. Произошли земноводные от древних кистеперых рыб. Первыми земноводными были ихтиостегиды (четвероногие рыбы). У них, как у рыб, был хвостовой плавник, жаберная крышка, чешуя. Но в отличие от рыб имели две пары пятипалых конечностей и легкие. Жили в основном в воде.
От ихтиостег произошли настоящие земноводные - стегоцефалы (панцирноголовые). Голова стегоцефалов была покрыта панцирем из кожных костей. Стегоцефалы жили до начала мелового периода. Вероятно, они дали начало современным беспанцирным земноводным и первым пресмыкающимся.
Выход на сушу сопровождался ароморфозом: жаберное дыхание сменилось на легочное, а плавники на пятипалые конечности, сердце стало трехкамерным, возник второй круг кровообращения, изменились органы чувств, более совершенной стала нервная система.
Первые пресмыкающиеся появились в карбоне. Ископаемые остатки сеймурии имели черты земноводных и пресмыкающихся. Вероятно, от подобных форм и произошли первые пресмыкающиеся - котилозавры. Они еще имели череп в виде костной коробки (сходство со стегоцефалами) с отверстиями для глаз, ноздрей и теменного органа. Расцвет пресмыкающихся был в мезозое.
Происходит: ороговевание эпидермиса и образование чешуй, или щитков (предохраняет тело от высыхания), усложнение строения легких, сердца, совершенствование скелета, образование зародышевых оболочек серозной и амниотической (обеспечило способность размножаться на суше).
Пресмыкающиеся дали начало двум новым классам позвоночных: млекопитающим и птицам.
Птицы произошли от первоптиц из группы ящеротазовых динозавров, бегающих на задних ногах. Из древних первоптиц найдены останки длиннохвостого археоптерикса. Археоптерикс имел признаки пресмыкающихся (длинный хвост, зубы на челюстях, когти на крыльях) и птиц (оперенье, крылья, внешний облик). Настоящие птицы появились от первоптиц в конце мезозоя. Они еще имели мелкие зубы.
Прогрессивные особенности птиц: теплокровность, четырехкамерное сердце, одна дуга аорты (создает полное разделение большого и малого кругов кровообращения).
Млекопитающие появились раньше, чем птицы. Предполагается, что они произошли от зверозубых ящеров, которые имели ряд прогрессивных особенностей: зубы расположены в альвеолах, вторичное костное небо, конечности располагались под туловищем, а не по бокам, произошли изменения в скелете. Наиболее близкими к млекопитающим из зверозубых рептилий были цинодонты. Дальнейшая эволюция млекопитающих связана с изменением зубной системы. Эти изменения привели к образованию двух групп животных. Потомки одной группы - многобугорчатые были мелкими животными, имели на коренных зубах многочисленные бугорки, сильно развитые резцы и не имели клыков. Возможно, что эта группа дала начало однопроходным.
Другая группа развивалась более прогрессивно. Это были мелкие зверьки, которые питались насекомыми, мелкими животными. В дальнейшем происходит разделение данной группы на две самостоятельные ветви – Низшие, Сумчатые и Высшие, Плацентарные.
Сумчатые млекопитающие возникли в меловой период. Наиболее древней группой сумчатых являются семейство опоссумов.
Плацентарные млекопитающие появились также в меловом периоде. Наиболее древней группой плацентарных считают отряд Насекомоядных. Предполагается, что они дали начало большинству групп последующих плацентарных. Возможно, непосредственно от насекомоядных произошли приматы.
Прогрессивная эволюция млекопитающих стлана с приобретением важнейших приспособлений: теплокровность, высокий уровень метаболизма, образование плаценты, вскармливание детеныша молоком, забота о потомстве. Этому способствовали прогрессивные изменения в дыхательной, кровеносной, нервной системах. У млекопитающих четырехкамерное сердце, одна дуга аорты, разделение артериальной и венозной крови, легкие альвеолярного строения, хорошо развит вторичный нервный свод – неопалиум.
13… Строение клетки прокариот. Рост, развитие, размножение бактерий. Типы питания бактеий . Фотосинтез, хемосинтез, фоторедукция. Значение микроорг-змов в природе и народном хозяйстве.К прокариотическим клеткам относят бактерии и сине-зеленые водоросли. У прокариотов ядерное вещество не имеет ядерной мембраны, в состав их плазмалеммы входят углеводы и аминокислоты. У прокариотов лишь одна хромосома, которая состоит только из ДНК. Эта хромосома имеет кольцевидную форму и не содержит щелочных белков - гистонов. Набор хромосом у прокариотов гаплоидный.У этих одноклеточных организмов отсутствует митоз и не наблюдается митотического аппарата. У прокариотов отсутствует мейоз, и при половом процессе передается лишь часть наследственной информации, а гаметой является сам организм. Цитоплазматическая ДНК у прокариотов локализована в плазмидах, а цитоплазматические мембраны отсутствуют. В составе клеточной стенки прокариотов есть пептидогликаны. Митохондрий у прокариотов не наблюдается, дыхательные ферменты локализованы в мезосомах, которые расположены на внутренней поверхности плазмалеммы.Процессы фотосинтеза осуществляются при помощи пигмента бактериохлорофилла, а восстановителями служат соединения серы. Рибосомы прокариотов мельче рибосом эукариотов. Движения цитоплазмы не наблюдается, в ней очень редко встречаются вакуоли. Жгутики прокариотов имеют субмикроскопические размеры, в состав жгутиков входит одна или несколько микротрубочек (фибрилл).Прокариоты чувствительны к антибиотикам и устойчивы к рентгеновскому излучению и повышенной температуре, выдерживая нагревание до +75… + 90°С. Характерной особенностью прокариотов является то, что они не способны к сосуществованию с другими видами клеток. Бактерии -одноклеточные организмы; их можно разглядеть только под микроскопом. Поэтому их и называют микробами или микроорганизмами. Бактерии освоили самые разнообразные среды обитания: они живут в почве, пыли, воде, воздухе, на внешних покровах животных и растений и внутри организма. Их можно обнаружить даже в горячих источниках, где они живут при температуре около 60 °С или выше. Жизнедеятельность микроорганизмов имеет важное значение для всех остальных живых существ, так как бактерии и грибы разрушают органическое вещество и участвуют в круговороте веществ в природе. К тому же бактерии приобретают все большее значение в жизни людей, и не потому, что они вызывают различные заболевания, а потому, что их можно использовать для получения многих необходимых продуктов.
СТРОЕНИЕ КЛЕТКИ. Прокариотическое: нет оформленного ядра, митохондрий, пластид. В центре цитоплазмы - нуклеоид, содержащий одну «хромосому» с кольцевой молекулой ДНК; мембраной от цитоплазмы не отделен. Внутри клетки выросты мембраны выполняют функции ЭПС, аппарата Гольджи; мезосома участвует в дыхании; имеются рибосомы. Цитоплазма покрыта наружной мембраной и плотной клеточной стенкой. Иногда клетка погружена в полужидкую коллоидную капсулу. По строению клеточной стенки бактерий можно разделить на две группы.Одни окрашиваются по Граму, поэтому их называют грамположительными, а другие обесцвечиваются при отмывке красителя и поэтому их называют грамотрицательными. Рост и размножение. Индивидуальный рост и бесполое размножение клеток. Отношение поверхность/объем у бактериальных клеток очень велико. Это способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта. В благоприятных условиях бактерии растут очень быстро. Рост прежде всего зависит от температуры и рН среды, доступности питательных веществ и концентрации ионов. Облигатным аэробам обязательно нужен еще и кислород, а облигатным анаэробам, наоборот, нужно, чтобы его совсем не было. Достигнув определенных размеров, бактерии переходят к бесполому размножению (бинарному делению), т. е. начинают делиться с образованием двух дочерних клеток. Переход к делению диктуется отношением объема ядра к объему цитоплазмы. Перед клеточным делением происходит репликация ДНК, во время которой мезосомы удерживают геном в определенном положении.Мезосомы могут прикрепляться и к новым перегородкам между дочерними клетками и каким-то образом участвовать в синтезе веществ клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20мин.
4 типа питания бактерий и некоторые их характеристики.
Известны три основные группы фотосинтезирующих (бактерии, которые энергию получают от солнца):1. Зеленые серобактерии. Анаэробные бактерии, использующие в качестве доноров водорода и электронов сероводород (H2S) и другие восстановленные соединения серы.
Пример: Свет
2H2S + СО2 →→→→→→→→ [СН2О] + 2S + Н2О.
Бактериохлорофилл
При этом откладывается сера.
2. Пурпурные серобактерии. Клетки имеют темно-красный цвет, так как бактериохлорофилла гораздо меньше, чем красных и коричневых пигментов. Это в основном анаэробы; фотосинтез у них происходит так же, как и у предыдущей группы.
3. Пурпурные несерные бактерии. Для этих бактерий различные органические соединения служат источником водорода, который используется для восстановления СО2 (у фотоавтотрофов) или какого-нибудь органического соединения (у фотогетеротрофов).
Хемосинтезирующие организмы (хемоавтотрофы)-это бактерии, которые используют в качестве источника углерода СО2, но энергию получают не от солнца, а с помощью химических реакций. Энергия может выделяться, скажем, при окислении водорода, сероводорода, серы, железа(П), аммиака, нитрита и других неорганических соединений. У железобактерий: Кислород
Fe2+ →→→→→Fe3+ + Энергия;
У бесцветных серобактерий: Кислород или
S→→→→→→→ SO42- + Энергия;
Сера Сульфат
В анаэробных условиях некоторые виды используют в качестве акцептора водорода нитрат и, таким образом, осуществляют денитрификацию.
У нитрифицирующих бактерий: Кислород
2NH4+→→→→→ NO-2 + Энергия
Аммиак Нитрит
Все рассмотренные только что бактерии - аэробные, так как акцептором электронов (и водорода) служит кислород.
Хемосинтезирующие бактерии играют очень важную роль в биосфере; в основном они участвуют в круговороте азота и таким образом поддерживают плодородие почвы.
Фоторедукция-для зел-х и пурпур-х серобактер., кот.в кач-ве источника исп-ют не Н2О, а Н2 S
ЗНАЧЕНИЕ В ПРИРОДЕ И ХОЗЯЙСТВЕННОЙ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА.
Бактерии молочнокислого брожения (гетеротрофные сапротрофы), расщепляющие углеводы: Положительное: приготовление молочнокислых продуктов (творог, простокваша, масло, сметана), силосование кормов, закваска капусты, засолка огурцов и помидоров. Отрицательное: порча продуктов.
Бактерии уксуснокислого брожения (гетеротрофные сапро-трофы), расщепляющие углеводы:Положительное: окисление спирта в уксусную кислоту, которая применяется для маринования, консервирования плодов и овощей. Отрицательное: порча продуктов.
Бактерии гнилостные (гетеротрофные сапротрофы), расщепляющие белки:Положительное: санитарная роль - минерализация органических остатков. Отрицательное: порча продуктов; во избежание гниения применяют сушку, соление, маринование, стерилизацию, пастеризацию, засахаривание.
Бактерии клубеньковые (гетеротрофные симбионты): Клубеньковые бактерии ризобиум проникают в корни бобовых растений (клевер, люпин, люцерна и др.) и вступают с ними в симбиоз. В результате на корнях образуются опухоли - клубеньки, заполненные бактериями, которые из атмосферного азота синтезируют азотистые соединения, доступные как растению-хозяину, так и другим растениям. Это природное обогащение почвы азотными удобрениями учитывается при составлении полевых севооборотов, куда обязательно включают бобовые растения.
14.Строение эукариотической клетки. мембрана трехслойна Рибосомы Комплекс Гольджи Митохондрии Лизосомы Хлоропласты Хромопласты Лейкопласты Клеточный центр, или центросома Включения Вакуоли Цитоскелет Кариоплазма Ядрышко 1. 2 3 4 5
15.Онтогенез и его периоды. Ранний эмбриогенез. Прямое и непрямое развитие.(51)
Онтогенез - индивид развитие организма с момента рождения и до смерти. Продолжит у разных орг-мов неодинакова и может продолж-ся от неск миндо сотен лет (черепаха). Онтогенез тесно связан с филогенезом но не яв-ся абсолют повторением филогенеза. Под влиянием окруж среды он меняется и может наступить мутац изменчивость. периоды онтогенеза:
1 предзародышевый
2. зародышевый(эмбриональный)
3. постэмбриональный
В 1 период развитие половых Кл и подгот их к дальнейшему развитию после оплодотворения(сперматогенез и овогенез) сперматогенез из 4 периодов: 1. размножение; 2. рост; 3. созревание; 4. формирование
Сперматогенез: Сперматогонии2n«сперматоциты 1порядка2n»сперматоциты2пор2n«сперматозоиды n.
Овогенез:о вогонии 2n»овоциты 1 пор2n «овоциты 2 пор2n „1 яйцеклетка+3 направительных тельца(редуцируются)
Во 2 периоде происх развитие зародыша внутриутробно(в чреве матери) или во внеш среде. Хар-ся быстрыми процессами формообразования, к-рый в короткий срок приводят к появл мнкл орг-ма. Имеет 3 периода: 1. начальный - 1 неделя эмбрионального развития нач-ся с момента оплодотв и заканчивается имплатанцией зародыша в слизистой матери. 2 зародышевый. Время от начала2 до 8 недели от имплантации до завершения органогенеза. 3 плодный - развитие плода с 9 нед до рождения. Усиленный рост, формообразование, питание гематотрофно.
3 период постэбр нач-ся с момента рождения, т.е. выхода организма из зарод оболочек. В этот период развития возник специфич анатомич и функцион особенности, наз-ся возрастными. Периодизация:
1-10 день- новорожденный
11д- 1 год-младенческий
1г-11 л- дети (девочки и мальчики)
11-15л(д) и 12-16(м)- подростковый
16-20(д) и 17-21(м)- юношеский
21-55(ж) и 22- 60(м)- зрелый
56, 61- 74 – пожилой
75-90 старческий, старше 90 долгожители
Прямое развитие (неличиночный тип), связан с > кол-вом желтка в яйцеклетке, либо развитие орг. В материнсом организме харак-но д/рыб, млекопит, птиц, пресмык.
Непрямое развитие-(личиночное) с метаморфозом-харак. д/ беспозв, ракообр, насекомых, червей, характер. Коротким эмбриональным периодом в ходе кот. Формируется личинка способ. Сам-но существовать. Не полное превращение - яйцо-личинка-имаго. Полное превр.- яйцо –личинка -куколка-имаго. РАННИЙ ЭМБРИОГЕНЕЗ: сперматозиод внедряется в яйцеклетку цитоплазма растворяется- стаия синкариона(ядра сливаюся“диплоидное ядро» зигота 2н-дел. На бластомеры-4, 8,16,32,64,128..-обр. бластула(зародыш с полостью внутри)-путем впячивания обр.-двухслойный - гаструла, экт.энт)-(бластоцель, бластопор)- развив. 3 зарод. Листока. энтодерма, мезодерма, эктодерма
Дробление м.б равномерным и неравномерным (дискоидальное)
Гаструляция происходит 4 путями:
1 путем инвагинации(впячивание)
2 эпиболия (обрастание)
3 деляминация(расслоение)
4 имиграция
Нейруляция закладка нервной трубки, хорды, амниотической и туловищной складки
эктодерма обр-ет нерв.пластинку-ст. нейрулы(закладканерв.трубки, хорды), слуха, обоняния, внеш.покровы.Энтодерма дет начало кишечнику, печени, легких, поджелуд железе. Мезодерма формир. Мускулатуру(дермотом, миотом), мышечная тк, кровеносная сист, все хрящевыеи костные элементы скелета, выделит, половая сист.
16.Основные пути филогенеза. Дивергенция, конвергенция, параллелизм.(52)
Эволюционное развитие может идти разными путями в зависимости от объекта и отношения его со средой. Выделяют несколько форм филогенеза.
Филетическая эволюция - это постепенные эволюционные перестройки определенной таксономической группы, которые ведут к возникновению новой группы, отличной от исходной. Примером филетической эволюции может быть ряд предков лошади: эогиппус - эпигиппус - мерик-гиппус - гашогиппус - современная лошадь.
Дивергенция - основной путь эволюции, который был вложен в основу дарвиновской теории. Дивергентная эволюция определяется расхождением таксона на несколько групп под влиянием элементарных факторов эволюции и приспособления к разным условиям существования. При дивергенции сходство определяется общностью происхождения, а различия - приспособлением к разной среде. Начало дивергенции внутри вида при образовании экотипов, экологических рас, географических подвидов. По мере расхождения признаков разновидности становятся видами, виды претерпевают дальнейшую дифференциацию и образуют роды, роды становятся семействами и т. д.
Дивергенция может проходить в нескольких формах: дихотомического ветвления (распадение вида на два дочерних вида), радиации (рай падение, вида на несколько дочерних видов), отщепление (предковый вщ существует одновременно с дочерними, сформированных из обособившихся в разное время популяций). Дивергенция развивается на основе гомологичных органов. Классическим примером дивергенции можно назвать paсхождение признаков у вьюрков Галапагосского архипелага.
Биологическое значение дивергенции заключается в том, что oна приводит к многообразию живых организмов, к равномерному распределению видов по экологическим нишам, сглаживает напряженность борьбы за существование.
Конвергенция - процесс, противоположный дивергенции. Она заключается в независимом приобретении в одинаковых условиях сходных признаков у форм в основном не связанных родством на основе аналогичных органов или структур. Например, роющие конечности крота медведки сходны; китообразные и рыбы имеют одинаковую форму тела. Конвергенцией определяется общее сходство между «голенастыми» болотными птицами (журавли и цапли). К явлениям конвергенции относятся «географические» (климатические) «параллелизмы», которые выражены в более крупной величине теплокровных животных холодных стран по сравнению с ближайшими представителями умеренных областей, в большей величине кладок у птиц и помёта у млекопитающих Арктики, в усилении пигментации тропических птиц.
То конвергенция у далеких групп животных идет за счет внешних условий у близких надо учитывать и родство. В связи с этим конвергенция наблюдается в пределах класса.
Параллелизм. Если ранее дивергировавшие группы попадают в иныв условия существования, у них независимо, параллельно могут формироваться сходные адаптивные признаки. Например, у глубоководных рыбпараллельно и независимо возникли органы свечения; развитие саблезубости у разных кошачьих Иногда параллелизмы охватывают целые системы органов или частьвсей организации. Например, параллельное развитие двудышащих рыб амфибий. Вероятно, сходства между двудышащими и амфибиями определяются родством этих групп. Особенно значительны параллелизмы строении сердца и кровеносной системы. Чем более родственные связи между организмами, тем чаще параллельное развитие охватывает значительные части всей организации. Об этом свидетельствуют и палеонтологические данные, например развитие хоботных. Чем ближе организмы друг к другу, тем более сходны их реакции на одинаковое изменение среды, тем более сходны (параллельны) изменения их строения. Для параллельного развития имеет значение как внешняя среда, так и внутренний факторы (т. е. исторически сложившиеся наследственные структуры организма). Соотношение внутренних факторов с внешними определяет эволюцию организмов. Например, копытные, проживающие в различных климатических зонах, спасаются от хищников бегством. Поэтому у копытных наблюдается развитие крайних степеней пальцесхождения и затем постепенная редукция краевых пальцев.
Генетические основы параллельной эволюции. При определенномизменении факторов внешней среды родственные организмы изменяются аналогично. В борьбе за существование через естественный отбор эти изменения приобретают «наследственную» форму. Материалом для отбора являются индивидуальные уклонения в строении, т. е. мутации. Родственные или близкие формы, как правило, дают одинаковые мутации (например, разные виды дрозофилы). Однако это не значит, что параллельные мутации основаны на одинаковых изменениях одних и тех же генов.
Известно, что разные гены одного и того же организма дают сходные мутации. Особенно, если эти изменения незначительны. А так как в ходе эволюции отбираются лишь комбинации мелких мутаций, то параллельные изменения могут строиться на различной генетической основе.
КУРСОВАЯ РАБОТА
по биологии
“Происхождение эукариотических
Выполнила: Сурова Вера
10-В класс,школа N55
Проверила: Струкова Н.А.
г.Рязань 1999 год
1 . Симбиотическая теория:
1.1 Фундаментальное разграничение между прокариотами и эукариотами;
1.2 Прокариотические организмы;
1.3 Эукариотические клетки;
1.4 Митохондрии и пластиды;
1.5 Способность эукариот к фотосинтезу;
2. Предшественники жизни;
3. Ферментирующие организмы:
3.1 Источники энергии ранних клеток;
3.2 Метаногенез как источник энергии;
3.3 Цианобактерии и хлорооксибактерии;
4. Фотосинтез и воздух;
5. Ассоциации и эукариоты:
5.1 Теории последовательных симбиозов;
5.2 Симбиотическое приобретение подвижности;
5.3 Эукариоты – фотосинтетики;
6. Взгляд на эволюцию клетки:
6.1 Гипотеза прямой филиации;
6.2 Гипотеза: от цианобактерий к водорослям;
6.3 Совместимость 2 –х гипотез: прямой филиации и
эндосимбиоза;
7. Выводы;
8. Литература.
ТЕОРИЯ ЭНДОСИМБИОЗА
Все живые организмы на Земле обычно делят на прокари-от и эукариот. Главной особенностью прокариот в отличие от эукариот является отсутствие у них полноценного клеточного ядра, покрытого мембраной. Клетки прокариот имеют очень не-большие размеры порядка 1 мкм. Объем эукариотических клеток, содержащих полноценное ядро, в 800 – 1000 раз больше объема клеток прокариот. В связи с этим клеткам – эукариотам необходим для выполнения жизненных функций гораздо больший мембранный аппарат. Действительно, в эукариотической клетке выявлены субклеточные структуры, окруженные незави-симыми от плазмалеммы и друг от друга мембранами. Эти структуры называются органеллами. Каждой органелле: митохондрии, хлоропласту, ядру, лизосомам, аппарату Гольджи и т. д. присущи свои специфические функции в жизни клетки.
С помощью молекулярно – биологических подходов получены убедительные доказательства того,что все живые существа должны быть разделены на 3 надцарства: бактерии, археи и эукариоты. Бактерии и археи возникли на Земле от общего предка - так называемого прогенота - около 4 млрд лет назад, а эукариоты по данным палеонтологов появились на Земле через 500 млн лет после прокариот. Наиболее популярной и достаточно обоснованной идеей происхождения эукариотических клеток, в настоящее время являктся идея многократного эндосимбиоза различных прокариот. По представлениям известных микробиологов А. Клюйвера и К. Ван Ниля, именно у древних прокариот произошел отбор клеток с наиболее экономичными и зачастую полифункциональными биохи-мическими реакциями, ставшими основой важнейших путей метаболизма.
Симбиотическая теория происхождения и эволюции клеток основана на двух концепциях, новых для биологии.Согласно первой из этих концепций, самое фундаментальное разграни-чение в живой природе - это разграничение между прокариотами и эукариотами,т.е. между бактериями и организмами, состоящими из клеток с истинными ядрами - протистами, животными, грибами и растениями. Вторая концепция состоит в том, что источником некоторых частей эукариотических клеток была эволюция симбиозов, т.е. формирование постоянных ассоциаций между организмами разных видов. Предполагается, что три класса органелл - митохондрии, реснички и фотосинтезирующие пластиды – произошли от свободно живущих бактерий, которые в результате симбиоза были в определенной последовательности включены в состав клеток прокариот - хозяев. Эта теория в большой мере опирается на неодарвинистские представления, развитые генетиками, экологами, цитологами и другими учеными, которые связали Менделевскую генетику с дар-виновской идеей естественного отбора.Она опирается также на совершенно новые или недавно возрожденные научные направ-ления: на молекулярную биологию,особенно на данные о структуре белков и последовательности аминокислот, на микро палеонтологию, изучающую наиболее ранние следы жизни на Земле, и даже на физику и химию атмосферы, поскольку эти науки имеют отношение к газам биологического происхсждения. Все организмы, состоящие из клеток, могут быть сгруппиро-ваны в пять царств: царство прокариот (Monera, куда относятся бактерии) и четыре царства эукариот (Protoctista, Animalia, Fungi , Plantae). Протоктисты- это эукариотические организмы, не относящиеся к животным, грибам или растениям. В царство протоктистов входят водоросли, протозои, слизевики и другие эукариотические организмы неясной принадлежности. Протисты определены более ограничительно- как однокле-точные эукариоты. Таким образом, царство Protoctista включает не только протистов – одноклеточных эукариот, но и их ближай-ших многоклеточных потомков, таких как красные и бурые водоросли,а также многие микроорганизмы,сходные с грибами,например хитридиевые. Согласно традиционному представ-лению о прямой филиации, такие клеточные органеллы, как митохондрии и пластиды, возникли путем компартментализации самой клетки.
НЕСКОЛЬКО ПРОКАРИОТ ОБРАЗУЮТ
ОДНУ ЭУКАРИОТУ
Клетки - это ограниченные мембранами системы, наименьшие из биологических единиц, способных к саморепродукции. Даже мельчайшие клетки содержат гены в форме молекул ДНК (одной или нескольких) и белоксинтезирующий аппарат, состоящий из нескольких типов РНК и множества белков. Все клетки содержат рибосомы – тельца диаметром около 0,02 мкм, состоящие из РНК по меньшей мере трех типов, и примерно пятидесяти различных белков. Прокариотичесие организмы – наименьшие биологические единицы, которые удовлетворяют этому определению клетки. У них нет ядер. С другой стороны, одиночные эукариотические (имеющие ядро) клетки в известном смысле вообще не являются единицами. Это комплексы различных белоксинтезирующих единиц (ну-клеоцитоплазмы, митохондрий и пластид) , тип и число которых варьирует в зависимости от видовой принадлежности. Таким образом, согласно этой теории, эукариотические клетки возникли в результате кооперации первоначально независимых элементов, объединившихся в определенном порядке. Тэйлор назвал такое представление о раздельном происхождении и развитии частей эукариотической клетки и их последующем объединении - теорией последовательных эндосимбиозов. Условия высокой температуры и кислотности привели к выработке в нуклеоцитоплазме особого класса белков - гистонов - для защиты ее ДНК. Митохондрии были способны расщеплять трехуглеродные соединения до углекислоты и воды. Симбиотическая ассоци-ация нуклеоцитоплазмы и митохондрий начала изменяться в сторону усиления их взаимной зависимости. Этот комплекс включил в себя двигательные органеллы - реснички и жгутики. За этой трансформацией последовало морфогенетическое преобразование, которое состояло в развитии выраженных клеточных асимметрий. С интеграцией взаимозависимых нуклеоцитоплазмы, митохондрий, ундулиподий - завершилось создание полигеномной структуры эукариотической клетки. Первая эукариотическая клетка была гетеротрофной. Она питалась готовыми органическими соединениями. У этих эукариот возникли процессы митоза, а затем и мейоза. И наконец, приобретение эукариотами способности к фотосинтезу, имело место в процессе становления основной эукариотической организации и после его завершения- последнее событие в зтой эпопее. Симбиозы становились более и более интегрированными. В настоящее время зависимость каждой органеллы от продуктов метаболизма других органелл стала настолько полной, что только современные методы анализа позволяют проследить метаболические пути исходных партнеров. Эукариоты разнообразны по своему строению, но метаболически они остаются единообразными. Метаболические ухищрения эукариот были разработаны различными бактериями еще до того времени, когда они объединились, превратившись в хозяина и его органеллы. Прокариоты достигли большого разнообразия в способах получения энергии и в тонкостях биосинтеза. Они оставили след своего существования задолго до того, как на основе эукариотической клетки сформировались такие крупные организмы, как животные, обладающие скелетом.
Схема про исхождения эукариотических клеток путем симбиоза
ПРЕДШЕСТВЕННИКИ ЖИЗНИ
Предки нынешних организмов возникли более трех милли-ардов лет назад из органических соединений, образовавшихся на поверхности Земли. Привлекательность этой концепции отчасти связана с ее доступностью для проверки.
С. Миллер и Х.. Юри показали, что при атмосферных усло-виях, имевших место на юной Земле, могут образовываться аминокислоты и другие органические молекулы. Различные типы органических молекул образовывались спонтанно в течение первого миллиарда лет существования Земли. Многие органические молекулы были обнаружены в межзвездном пространстве и в метеоритах. Значит, органические соединения возникают при взаимодействии легких элементов, в при-сутствии источников энергии. Универсальность химического механизма биорепродукции означает, что все живое на Земле происходит от общих предков: вся нынешняя жизнь произошла от клеток с информационными системами, осно-ванными на репликации ДНК, и на синтезе белков, направляемом информационной РНК.
ФЕРМЕНТИРУЮЩИЕ ОРГАНИЗМЫ
Возможно, что нуклеиновые кислоты с самого начала были отделены от окружающей среды липопротеидными мембранами. Информация стала кодироваться последовательностью нуклеотидов в молекулах ДНК. Древнейшие клетки использовали АТФ и аминокислоты, входящие в состав белков, как прямые источники энергии, и как структурные компоненты. Итак - древнейшая жизнь существовала в форме гетеротрофных бактерий, получавших пищу и энергию из органического материала абиотического происхождения. Для неодарвиновской органической эволюции необходимо три феномена: воспроизведение, мутации и селективное давление среды. Воспроизведение - это неприменное условие биологи-ческой эволюции. При достаточно точной репродукции - мутации и естественный отбор неизбежны. Объяснить проис-хождение такой репродукции - значит понять происхождение самой жизни. Мутирование - первичный источник изменчивости в эволюции, ведет к наследуемым изменениям в самовоспроизводяйщейся системе. Как только выработалось надежное воспроизведение, началась неодарвиновская эволю-ция. Организмы, у которых выработались пути синтеза нужных клетке компонентов, имели преимущество перед другими. Древнейшими автотрофами, то есть организмами способными удовлетворять свои потребности в энергии и органических веществах за счет неорганических источников, были анаэробные метанообразующие бактерии. Способность к фотосинтезу впервые выработалась у бактерий, чувствительных к кислороду. Поглощая солнечный свет с помощью бактериального хлорофилла, они превращали угле-кислоту атмосферы в органические вещества.Появление ана-эробного фотосинтеза необратимо изменило поверхность и атмосферу Земли.
Углекислота удалялась из атмосферы и превращалась в ор-ганическое вещество в соответствии с общим уравнением фотосинтеза: 2H2X+CO2=CH2O+2X+H2O . Фотосинтезирующие бактерии, которые использовали для восстановления СО2 сероводород, откладывали элементарную серу. Они дали начало бактериям, способным к фотосинтезу с выделением кислорода. Вероятно этот тип фотосинтеза зародился в результате накопления мутаций у фотосинтезирующих серных бактерий. Этот путь привел к возникновению сине-зеленых водорослей или цианобактерий. Недавно была открыта и изучена новая группа фотосинтезирующих прокариот, выделяющих О2 - хлороксибактерии. Можно предполагать, что именно циано- и хлорооксибактерии были предками пластид, находящихся в клетках водорослей и растений. По мере накопления О2 в результате микробного фотосинтеза земная атмосфера становилась окислительной, что вызвало кризис всей жизни – до этого времени мир был заселен анаэробными бактериями. Чтобы выжить микробы должны были избегать кислорода, укрываясь в болотах, или должны были выработать обмен веществ, способный предохранить их от возрастающего количества кислорода.
ФОТОСИНТЕЗ И ВОЗДУХ
Примерно 2 миллиарда лет назад один за другим происходили различные эволюционные процессы по созданию метаболи-ческого разнообразия. Способность к выработке специфических ферментов может передаваться в пределах одного вида с помощью вирусоподобных частиц, называемых плазмидами. Поразительны биосинтетические способности прокариот: из углекислоты, нескольких солей и восстановленных неорга-нических соединений, служащих источниками энергии, микро-организмы- хемоавтотрофы могут синтезировать в содержащей кислород неосвещенной воде все сложнейшие макромолекулы, необходимые для роста и размножения. Эти биохимические про-цессы приводили к системам круговорота элементов еще тогда, когда не было растений, грибов, животных.
Ассоциации и эукариоты.
Взгляд на эволюцию клетки.
Прямая филиация.
Традиционный взгляд на происхождение и эволюцию эукарио-тических клеток заключается в том, что все живые организмы произошли прямым путем от единственной предковой популя-ции в результате накопления мутаций под действием естест-венного отбора.
Точечные мутации, дупликации, делеции и другие наследствен-ные изменения, как известно игравшие роль в эволюции живот-ных и растений, ответственны и за дифференцировку эукарио-тических клеток из прокариотических- теория прямой филиации. т. е. представление о несимбиотическом происхождении клеток. Возможно, зеленые водоросли являются потомками родоначаль-ников всех растений. Многие особенности этих водорослей наводят на мыысль, что они и растения имеют общего предка. Филогения животных сложнее, но концепция прямой филиации вероятно приложима также и к их путям эволюции. Губки очевидно произошли от протистов, независимо от остальных групп животных. Итак, по крайней мере для животных и растений гипотеза, по которой сложные эукариотические орга-низмы развились из более просто устроенных эукариот путем накопления отдельных мутаций под действием естественного отбора, вполне правомерна. Эволюционные новшества опирают-ся на сохранение высокосовершенных благоприятных генов. Итак, новые организмы с повышенной способностью контроли-ровать свою среду обитания развивались больше благодаря сохранению дупликации и рекомбинации высокоадаптированных генов,чем путем приобретения новых свойств в результате простых, случайных точечных мутаций. Гипотеза прямой филиации не позволяет объяснить происхождение эукариот от прокариот. Например хиатус между неспособными к митозу цианобактериями, и такими формами с вполне развитым митозом, как красные водоросли, трудно понять с позиций прямой филиации, но он естественно вытекает из симбиотической теории.
Согласно теории прямой филиации и теории симбиоза, все организмы на Земле произошли от биохимически одно-типных бактериальных предков, а растениям и животным дали начало эукариотические микробы. Фотосинтезирующие бактерии дали начало водорослям, и в конце концов расте-ниям, а некоторые водоросли утратили свои пластиды и пре-вратились в предков грибов и животных. Теоретики прямой филиации согласны в том, что пластиды и другие органеллы эукариот, включая ядро, сформировались путем дифферен-циации внутри самих клеток и называют это ботаническим мифом.
Филогения типов растений
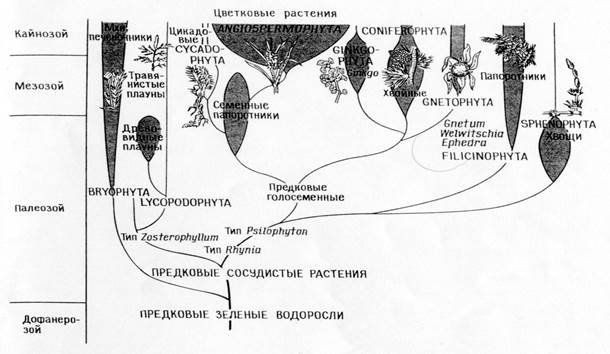
Сопоставление филогений, соответствующих теории прямой филиации (А)
и теории симбиоза (Б)
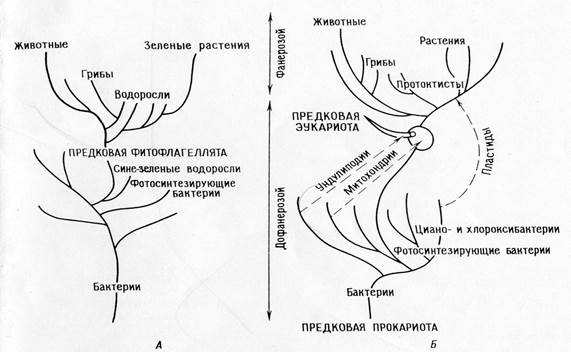
Различия между положениями теории прямой филиации и симбиотической теории:
Теория прямой филиации.
Главная дихотомия: животные – растения
Фотосинтезирующие эукариоты (водоросли и зеленые растения) произошли от фотосинтезирующих прокариот (сине-зеленых водорослей)
Растения и их фотосинтезируюшшие системы в докембрии эволюционировали монофилетически.
Животные и грибы произошли от автотрофов, утративщих пластиды.
Митохондрии дифференцировались внутриклеточно у фотосинтезирующих праводорослей.
У праводорослей внутриклеточно дифференцировались ундолиподии, митотический аппарат и все прочие эукариотические органеллы.
Все организмы произошли от предков – прокариот путем накопления одиночных мутаций.
СЛЕДСТВИЕ: согласно теории, должны существовать праводоросли; трудно связать эту теорию с ископаемой летописью и построить соответствующую филогению.
Теория последовательных эндосимбиозов:
Главная дихотомия: прокариоты – эукариоты.
Фотосинтезирующие эукариоты (ядросодержащие водоросли и растения) и нефотосинтезирующие эукариоты (животные, грибы, простейшие) произошли от общих гетеротрофных предков (амебофлагеллят).
Эволюция фотосинтеза в архее была монофилетической у бактерий (включая циано- и хлорооксибактерий), фотосин-тезирующие протисты возникли внезапно, когда их гетеро-трофные протисты – предки вступили в симбиоз с фото-синтезирующими прокариотами (протопластидами).
Животные и большинство водорослей и грибов произошли непосредственно от гетеротрофных протистов.
Митохондрии уже были у гетеротрофных протистов,когда последние в результате симбиоза приобрели пластиды. Промитохондрии были ранее приобретены симбиотическим путем.
Митоз выработался у гетеротрофных эукариотических протистов путем дифференциации ундолиподиальной системы.
Прокариоты произошли от ранних прокариотических предков путем ряда одиночных мутационных событий, эукариоты тоже произошли от эукариотических предков благодаря одиночным мутационным событиям, эукариотические же предки произошли от прокариот в результате серии симбиозов.
Следствия: теория объясняет биохимические пути и их избыточность у органелл. Она позволяет построить последо-вательную филогению, объясняющую “биологический разрыв “ между временами протерозоя и фанерозоя. Теория предска-зывает, что у органелл должны сохраняться остатки систем репликации.
Ботанический миф
Со времен Жюсье низшие растения называют таллофитами.а высшие - бриофитами и трахеофитами. Согласно ботаническому мифу,общие предки фотосинтезирующих бактерий и сине-зеленых водорослей путем прямой филиации дали начало эукариотическим водорослям в том числе морским. Группы организмов, филогению которых в наибольшшей мере затра-гивает концепция прямой филиации, оказались в области ботаники - это бактерии, водоросли, грибы и растения.
Гипотетические организмы, которые могли бы быть связующим звеном между цианобактериями и зелеными водорослями, были названы родоначальными фито-монадами (ancestral phytomonads) или праводорослями. Некоторые авторы утверждали, что подходящими кандитами на роль праводорослей могут быть эукариоты Cyanidium и Cyanophora, сходные по своим пигментным системам с цианобактериями. У таллофитов не было обнаружено промежуточных форм между немитотическим состоянием цианобактерий, и вполне развитым типичным митозом зеленых водорослей и растений. Хотя грибы и цветковые растения полностью лишены ундулиподий, им свойственны митоз и мейоз, для которых необходимы веретена, состоящие из микротрубочек. Имеется общее согласие в том,что все многоклеточные организмы,обладающие микротрубочками,происходят от эукариотических микроорганизмов с ундолиподиями, но никто не знает, от какой именно группы протистов. Подкреплением ботанического мифа служили древность и сложность фотосинтеза, а токже тот факт, что у многих организмов (у бактерий, евгленовых, пара-зититических растений) как в природе, так и в лаборатории, наблюдалась утрата фотосинтеза в результате мутаций. Поскольку фотосинтез - анаэробный процесс,обеспечивающий в конечном итоге существование всех организмов, то он должен был развиться очень рано в истории жизни на нашей планете, еще до того как возник митоз, для которого небходим кис-лород. в фотосинтезе участвует множество пигментов, липидов и ферментов,упорядоченно расположенных в очень сложных мембранах. При этом детали фотосинтетических механизмов у водорослей и растений, с одной стороны, и у цианобактерий, с другой - поразительно сходны. Все эти организмы освобождают газообразный кислород из воды, которая служит источником атомов водорода, используемых для восстановления СО2 до органических веществ клетки. Многие водоросли и простейщие не имеют ни митоза, ни генетической системы, сходной с таковой животных и растений. Инфузории имеют особую гене-тическую систему с ядрами 2 типов, причем клетки нередко многоядерные. Один тип яядер – макронуклеусы, а другой – намного меньшие микронуклеусы. Микронуклеусы физиологи-чески несущественны. однако они хранят копии генов, а при половом процессе - претерпевают мейоз. Физиологически необходимые макронуклеусы служат местом транскрипции РНК, и, таким образом, управляют физиологией клетки. Одна из трудностей при выяснении отношений между цианобактериями и эукариотическими водорослями связана с отсутствием данных о происхождении ундолиподий: не было найдено никаких промежуточных звеньев между цианобактериями, у которых ни-когда не бывает ундолиподий, и множеством водорослей, которые их имеют. Другая проблема – полное отсутствие полового процесса у цианобактерий: между бесполыми цианобактериями и водорослями, большинству которых свойственен половой процесс, не оказалось никаких промежуточных форм. Обнаружилось, что большинство водо-рослей имеют ундолиподии на какой – либо стадии жизненного цикла. Подвижные структуры у водорослей, простейших, жи-вотных и растений оказались одинаковыми: все они состоят из микротрубочек диаметром 0,024 мкм. Выяснилось,что мито-тическое веретено даже у грибов, у которых нет ундолипдий,состоит из белковых микротрубочек того же диаметра, что и в ундолиподиях. На протяжении полувека казалось чрезвычайно правдоподобным, что эукариотичкские водоросли произошли от цианобактерий. Этот ботанический миф привел к предположе-ниям, которые многие годы служили как стимулом для исследования, так и системой координат,в которой про-изводилась оценка получаемых результатов. Утверждение, что связующие звенья между цианобактериями и эукариотическими водорослями вымерли, не оставив ископаемых следов и живых реликтов, становилось все менее убедительным по мере поступ-ления новых данных о живых микробах и об окаменелостях. Главные промежуточные звенья между фотосинтезирующими прокариотами и эукариотами не вымерли бесследно, как того требует ботанический миф,- они никогда не существовали. Согласно теории последовательных эндосимбиозов, циано-бактерии – действительно родоначальники,но только пластид, а не остальных частей эукариотической клетки, и уж во всяком случае не ее ядра. И все же в вопросе о происхождении ядра гипотеза прямой филиации и теория симбиоза совместимы. Мыслимые модификации крайнего варианта теории симбиоза включают, например,идею о симбиотическом происхождении пластид, но не митохондрий, или о симбиотическом про-исхождении пластид и митохондрий, но не ундолиподий. Сторонники прямой филиации выдвинули также предположение о происхождении ядра путем симбиоза.
Решающие доказательства эндосимбиотического происхождения хлоропластов и митохондрий.
Наиболее существенные доказательства происхождения важней-ших органелл эукариотических клеток - хлоропластов и мито-хондрий из прокариот получены в последние годы при изуче-нии структуры рибосомальных РНК. Молекудярный биолог
К. Воз показал уже в 80-х годах, что структура РНК из рибосом хлоропластов растений почти идентична структуре РНК из некоторых цианобактерий. В то же время группа исследователей установила, что РНК рибосом, полученных из митохондрий разных эукариотов схожа с РНК некоторых бактерий, в частности бактерий рода Paracoccus . Набор дыхательных ферментов указанной бактерии очень похож на
“ дыхательный ансамбль “ митохондрий животных.
Эти данные достаточно ясно свидетельствуют в пользу большей вероятности эндосимбиотического происхождения эукариотических клеток. Явление эндосимбиоза и по сей день чрезвычайно широко распространено в природе. Известно, что на теле некоторых морских рыб часто поселяются светящиеся бактерии, которые не только сами получают большие преимущества для своей жизни, но и очень полезны для организмов хозяев. Все сказанное убедительно свидетельствует в пользу эндосимбиотического происхождения эукариотических клеток из гораздо меньщих по объему клеток прокариот. Однако главный вопрос заключается в установлении природы хозяйской клетки, ведь клетки эукариот обладают хорощо сформированным ядром, имеющим оболочку, а прокариоты такого ядра не имеют. Л. Маргелис, например, считает,что роль хозяйских клеток, возможно, сыграли прокариоты – предшественники современных бактерий, относящиеся к микоплазмам - очень примитивно построенным прокариотам, практически лишенным клеточных стенок. В последнее время появились и широко обсуждаются работы японского ученого
Т. Ошимы. На основании тщательного изучения структуры рибосомных РНК многих про- и эукариот, в том числе и многих представителей до сих пор малоизученного надцарства архей, этот автор приходит к выводу, что хозяйской клеткой являлся один из предков современных архей. Следует сказать, что у представителей именно этого надцарства, в отличие от бактерий, имеется очень много общих с эукариотами биохи-мических и молекулярно – биологических особенностей. В по-следних работах Т. Ошима приводятся доводы в пользу того, что хозяйской клеткой при возникновении эукариот являлся предок архей, подобный одному из современных представителей этих прокариот - термоплазме. Каково будет окончательное решение проблемы установления природы хозяйской клетки, давшей начало эукариотической клетке в процессе множествен-ного эндосимбиоза, покажут дальнейшие исследования.
Наиболее популярной и обоснованной гипотезой происхождения эукариотических клеток является в настоящее время представле-
ние об их образовании путем многократно происходившего в процессе эволюции в течении первых 1.5 – 2 млрд лет существования на Земле живых клеток эндосимбиоза различных прокариот, относящихся к бактериям и археям. Решающие доказательства эндосимбиотического происхождения хлоро-пластов и митохондрий были получены ведущими биологами мира при изучении структуры рибосомальных РНК. Эти данные достаточно ясно свидетельствуют в пользу большой вероятности эндосимбиотического происхождения эукарио-тических клеток из гораздо меньших по объему (на несколько порядков) клеток прокариот.
ЛИТЕРАТУРА
1. Соросовский образовательный журнал. Биология, Химия, Науки о земле, Физика, Математика. N 5 1998г
2 Маргелис Л. Роль симбиоза в эволюции клетки. Пер. с англ., М. МИР 1983г
3 Основы микробиологии. М. “Медицина “ 1992 .
4 Хахина Л.Н. Концепция А.С. Фаминцына о значении симбиоза в эволюции Л. ; Наука, 1981 , с. 165 – 181 .
5 Альбертс Б. Молекулярная биология клетки; Пер. с англ. М. Мир 1986 т.1
Термин Prokaryotae (доядерные) объединяет микроорганизмы с примитивной организацией ядерных структур. Это название было предложено канадским ученым Мюрреем в 1968 г.
Во второй половине XX в. стало очевидным, что все живые организмы, имеющие клеточное строение, могут быть отнесены к одной из двух групп - прокариотам или эукариотам. Клетки прокариот устроены проще - они лишены окруженного оболочкой ядра. В течение некоторого времени считали, что термины «прокариоты» и «бактерии» - синонимы и что бактерии представляют собой самостоятельную и единую ветвь эволюции живых организмов. Однако постепенно накапливались данные о том, что различия между некоторыми бактериями могут быть существенными и свидетельствуют об отсутствии у них близкого эволюционного родства. Особенно большое значение придается данным о строении рибосомальных и транспортных рибонуклеиновых кислот (РНК). В настоящее время на основе результатов сравнения последовательностей оснований в молекулах рибосомальных РНК судят о родственных связях организмов. Такого рода данные, а также другие особенности прокариот показали, что на самом деле прокариоты должны быть разделены по крайней мере на две группы, имеющие различное происхождение и пути эволюции. Кроме того, эукариотические клетки не только более сложно организованы, но и обладают элементами, которые они не могли получить от предков современных прокариот. Это, прежде всего, цитоплазматические рибосомы, отличные от рибосом прокариот. Принципиальное сходство генетического кода, организации макромолекул и биохимического аппарата синтеза белка свидетельствует о единстве происхождения всех живых организмов. Предполагают, что существовал некий общий предок - «прогенот», но что он собой представлял - неизвестно. Этот прогенот мог дать начало трем самостоятельным ветвям эволюционного древа. С точки зрения иерархической систематики эти ветви предложено рассматривать в качестве доменов, имеющих ранг выше традиционных царств. Эти домены (империи) - Eubacteria, Archaebacteria, Eukaryota (эубактерии, архебактерии и эукариоты). Общие и дифференцирующие признаки архебактерий, эубактерий и эукариот даны в таблице.
Главная особенность прокариотной клетки - отсутствие ядра, ограниченного от цитоплазмы двойной мембраной. Наследственный материал сосредоточен в бактериальной хромосоме, обычно представленной в виде кольцевой молекулы двухцепочечной ДНК. У некоторых спирохет обнаружены хромосомы линейного типа, а у актиномицетов - «псевдокольцевые». Нитевидная молекула ДНК выполняет функцию ядра и располагается в центральной зоне клетки, называемой «нуклеоидом». В электронном микроскопе на срезе бактериальной клетки эта зона выглядит более светлой, чем остальная цитоплазма.
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации. Прежде всего, в клетке бактерий отсутствует мембранная сеть - эндоплазматический ретикулум. В клетках прокариот цитоплазматическая мембрана, лежащая под клеточной стенкой и отгораживающая снаружи протопласт, является единственной мембраной в клетке.
Цитоплазматическая мембрана, окружающая каждую клетку, определяет ее величину и обеспечивает сохранение существенных различий между клеточным содержимым и окружающей средой. Но мембрана - это не просто механическая перегородка. Она служит также высокоизбирательным фильтром, который поддерживает разницу концентраций ионов по обе стороны мембраны и позволяет питательным веществам проникать внутрь клетки, а продуктам выделения выходить наружу. Цитоплазматическая мембрана представляет собой ансамбль липидных и белковых молекул, удерживаемых с помощью нековалентных взаимодействий. В электронном микроскопе мембраны имеют вид листков толщиной около 7 нм с выраженной трехслойной структурой. Двойной слой липидных молекул определяет структурные особенности мембраны, белки же ответственны за большинство мембранных функций.
Основные признаки архебактерий, эубактерий и эукариот
| Признаки | Архебактерии | Эубактерии | Эукариоты |
| Типичные организмы | метаногены, экстремальные термофилы, галофилы | основные группы бактерий | протисты, грибы, растения, животные |
| Типичные размеры, мкм | 0,5-4 | 0,5-4 | более 5 |
| Геном | кольцевая хромосома | кольцевая хромосома | ядро со многими сложными хромосомами |
| Мембранные структуры: эндоплазматический ретикулум комплекс Гольджи лизосомы | нет нет нет | нет нет нет | есть есть есть |
| Митохондрии | нет | нет | есть |
| Хлоропласты | нет (иногда тилакоиды) | нет | нет (у грибов и животных), есть (у растений и водорослей) |
| Клеточные стенки | протеин, псевдомуреин | муреин, липополисахарид | различные |
| Рибосомы | 70S* | 70S | 80S (в цитоплазме)+705 рибосомы органелл, как у бактерий |
| РНК-полимераза | сложная | простая | сложная |
| Жгутики | простые | простые, состоят из одной или нескольких фибрилл | сложной структуры (2х9)+2 |
| Фиксация азота | есть | есть | нет |
| Анаэробное дыхание | есть | есть | нет |
| Хемосинтез | есть | есть | нет |
S* - единица осаждения Сведберга, характеризующая размеры рибосомы
В цитоплазматической мембране прокариот существуют множественные впячивания - инвагинации, которые выполняют разные функции. Они могут содержать ферменты и иметь вид пузырьков или являются мезосомами и участвуют в делении клетки. Цитоплазматическая мембрана прокариот является местом локализации ферментов энергетического метаболизма и, следовательно, выполняет функцию митохондрий, отсутствующих в клетках прокариот. Поэтому у бактерий с высоким уровнем дыхательного метаболизма цитоплазматическая мембрана особенно развита, т.е. она имеет много впячиваний, что увеличивает ее поверхность. Цитоплазматическая мембрана участвует также в делении нуклеоида. Кольцевая ДНК закрепляется на мембране и после репликации (удвоения) расходится по двум новым клеткам благодаря росту мембраны. Из цитоплазматической мембраны формируются покровы эндоспор. Она представляет собой также главный барьер для проникновения веществ внутрь клетки. Цитоплазматическая мембрана не пропускает крупные молекулы, она не способна к пиноцитозу и экзоцитозу.
Структуры, расположенные снаружи от цитоплазматической мембраны, - клеточная стенка, капсула, слизистый чехол, жгутики, фимбрии, пили, ворсинки - являются поверхностными структурами.
Клеточная стенка - важный и обязательный структурный элемент большинства прокариотных клеток (истинных бактерий, не считая архебактерий и микоплазм), располагающийся под капсулой или слизистым чехлом или в случае отсутствия последнего - контактирующая с окружающей средой. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клетке определенную присущую ей форму.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотичных организмов. Клеточная стенка прокариот состоит из нескольких слоев, из них основной - каркасный - слой, ответственный за прочность, формируется из особого вещества- пептидогликана муреина, который синтезируется только прокариотной клеткой и не встречается у других организмов. В группе истинных прокариот различают два главных типа строения клеточных стенок. Эти два типа соответствуют грамположительным и грамотрицательным бактериям. Названия этих групп происходят от способности или неспособности разных бактерий окрашиваться по Граму. Датским врачом К. Грамом, занимавшимся окрашиванием тканей, в 1884 г. был предложен метод окраски, который позднее использован для бактерий. Если фиксированные клетки эубактерий обработать сначала раствором кристаллического фиолетового, а затем йодом, то образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой и последние остаются окрашенными, у грамотрицательных видов окрашенный комплекс вымывается из клетки, и они обесцвечиваются. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки. Метод окраски по Граму нашел широкое применение в микробиологической практике для дифференциации бактерий.
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу, так и по ультраструктуре.
Основную массу клеточной стенки грамположительных бактерий составляет специфический гетерополимер - пептидогликан муреин. Полисахаридный остов молекулы построен из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой с помощью гликозидных связей. Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входят тейхоевые кислоты и полисахариды. Тейхоевые кислоты представляют собой полимеры, построенные на основе многоатомных спиртов (рибита и глицерина), соединенных между собой фосфодиэфирными связями.
У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных. В ее состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к цитоплазматической мембране. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки - наружная мембрана, которая состоит из фосфолипидов (типичных для элементарных мембран), белков, липопротеина и липополисахарида.
Эмпирически найденные различия между двумя группами бактерий в отношении окраски по Граму оказались коррелятивно связанными со многими другими важными признаками. Например, грамположительные бактерии размножаются бинарным делением за счет формирования поперечной перегородки в клетке, грамотрицательные - путем образования перетяжки. Первые чувствительны к пенициллину, вторые - нет. Грамотрицательные бактерии не образуют эндоспор, а некоторые грамположительные их образуют.
Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внешней формы, дает возможность клетке существовать в гипотонических растворах. В первую очередь это «заслуга» пептидогликана. С клеточной стенкой связана проницаемость и транспорт веществ в клетку.
Под цитоплазматической мембраной у бактерий находится цитоплазма. Это коллоидная система, состоящая из воды, белков, жиров, углеводов, минеральных соединений и других веществ, соотношение которых варьирует в зависимости от вида бактерий и их возраста. Цитоплазма прокариотной клетки не подвержена циклическому движению, как у растений, или сократительным явлениям, как у амеб. Бактериальные клетки осуществляют движение за счет других механизмов: либо при помощи просто устроенных жгутиков, отличающихся от сложных жгутиков эукариот, они активно плавают в жидкой среде либо скользят, выделяя слизь и перемещаясь по поверхности плотного субстрата.
Подвижные клетки бактерий активно перемещаются в направлении, определяемом теми или иными факторами. Такие направленные перемещения бактерий называются «таксисами». Различают хемотаксис, фототаксис, магнитотаксис, термотаксис, вискозитаксис (реагирование на изменение вязкости раствора).
Покоящиеся клетки бактерий представлены у грамположительных бактерий эндоспорами, у грамотрицательных - цистами. Споры бактерий обладают уникальной устойчивостью к неблагоприятным воздействиям. Особенно поразительна устойчивость бактериальных спор к нагреванию: некоторые споры переносят кипячение в течение 1 ч и более и при этом не теряют жизнеспособности. Очень стойки споры и к высушиванию. Устойчивость спор связана с наличием у них особого слоя в клеточной стенке - кортекса.
Клетки бактерий при спорообразовании иногда приобретают необычную для них форму веретена, лимона или барабанной палочки. Эндоспора, как правило, одна на клетку, но встречаются бактерии с числом спор более четырех. У некоторых анаэробных бактерий на спорах формируются ячеистые колпачки, составленные из газовых вакуолей, или разного рода выросты
Споры бактерий - это не единственные формы покоя у прокариот. Грамотридательные бактерии, которые не способны к образованию эндоспор, в ряде случаев превращаются в цисты - особые клетки с толстыми наружными покровами (например, Azotobacter chroococcum)
Снаружи клеточные стенки прокариот часто бывают окружены слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев, чехлов, или влагалищ (у нитчатых бактерий). Все они являются результатом биосинтеза прокариотами органических полимеров и отложения их вокруг клеток.
Капсулы, слизистые образования и чехлы могут содержать компоненты, одинаковые с клеточной стенкой, однако их химический состав не идентичен. Как правило, химический состав капсул, образуемых бактериями, родо- и видоспецифичен. Основные химические компоненты большинства капсул прокариот - полисахариды. Исключение составляют капсулы некоторых видов бацилл, построенные из полипептида, являющегося полимером глутаминовой кислоты. Чехлы - более сложные структуры, они имеют более сложный химический состав и содержат сахара, белки, липиды.
Капсулы имеют разную толщину и легко выявляются под
микроскопом при негативном контрастировании препарата черной тушью.
Капсулы и чехлы защищают клетку от повреждений, высыхания, создают дополнительный осмотический барьер, служат препятствием для проникновения фагов, они создают вокруг клеток особую среду, обеспечивающую определенный водный режим. Капсулы имеют специфические функции. У клубеньковых бактерий, например, они определяют избирательное прикрепление к корневым волоскам растений разных видов. Иногда капсулы могут использоваться в качестве запасных питательных веществ. С помощью слизи осуществляется связь между клетками в колонии, прикрепление клеток к различным поверхностям
Полисахариды капсул взаимодействуют с катионами почвенных растворов, они могут разрушать почвенные минералы, участвовать в процессах агрегации и образовании водопрочной почвенной структуры. Возможно включение микробных внеклеточных слизей в процесс образования гумусовых веществ.
Форма клетки прокарит очень разнообразна. Известны прокариоты сферической формы, палочковидной, спирально закрученной, в виде запятой, в виде кольца - замкнутого или разомкнутого, с выростами - простскгми. Клетки могут иметь квадратную или звездчатую форму. Многоклеточные прокариоты имеют форму нитей или червеобразную форму
Все прокариоты относятся к группе микроскопических организмов, размер их клеток измеряют микронами (микрометрами) (1мкм = 10 3 мм), а их детали- нанометрами (1 нм = 10 мм). Почти 40% бактерий в почвах имеют размеры, лежащие за пределами способности светооптического микроскопа, они видны только в электронном микроскопе. При таких малых размерах отношение поверхности к объему очень велико, следствием чего является высокая потенциальная интенсивность метаболизма
Все прокариоты делят на четыре группы по признаку строения наружных покровов клетки: грамотрицательные (Gracilicutes) эубактерии, грамположительные (Firmicutes) эубактерии, микоплазмы (Mollicutes), лишенные клеточных стенок, и архебактерии, или археи (Archaebacteria). Три первые группы объединяются как эубактерии (Eubacteria), архебактерии представляют собой обособленную группу организмов (Archaebacteria), отличающихся от других строением молекул рибосомальных и транспортных РНК, РНК-полимеразами, липидами, строением клеточной стенки
Грамотрицательные бактерии имеют клеточные стенки, где помимо основного каркасного слоя из муреина имеется еще наружная мембрана и перекрывающий ее сверху слой белка и/или гликопротеида. Между цитоплазматической мембраной и слоем муреина имеется пери плазматическое пространство. Эта группа бактерий наиболее обширна и разнообразна по числу видов и физиологическим функциям. Группа прокариот объединяет фототрофные и хемотрофные бактерии. Среди них есть обычные сапротрофы, копиотрофы, олиготрофы, внутриклеточные паразиты других бактерий, патогенные для человека, животных и растений формы со сложными потребностями в источниках пита ния. По морфологии это разнообразные бактерии от простых палочковидных псевдомонад до миксобактерий со сложным циклом развития.
Микоплазмы - бактерии, не имеющие клеточной стенки, функцию которой выполняет цитоплазматическая мембрана.
Архебактерии объединяют прокариоты с разной структурой наружных покровов, в составе которых отсутствует характерный для истинных бактерий муреин. У одних представителей архебактерий в клеточной стенке есть псевдомуреин, у других клеточная стенка состоит из слоя глобулярных белков, или гликопротеидов.
Разделение бактерий по группам на основании фенотипических признаков (морфологии, физиологии) удобно в практических целях для быстрой идентификации организмов. Организмы объединяются в искусственные группы, или «секции», не имеющие таксономического статуса и часто обозначаемые по наиболее характерному роду или названию. По такому принципу построен Определитель бактерий Берджи.
Изучая многообразие организмов, населявших и населяющих Землю, систематики стремятся создать филогенетическую систему, которая отражала бы генеологическое родство и общность происхождения организмов, объединяемых в ту или иную группу - таксон. Этой цели служит молекулярная систематика, основанная на сходстве строения биополимеров - ДНК, РНК, белков. За возможность выявить сходство генов ее часто называют геносистематикой. Одним из наиболее важных достижений геносистематики считается открытие новой группы микроорганизмов, выделенных в отдельное царство - архебактерий. Степень сходства бактериальных геномов определяют сравнением последовательности нуклеотидов в молекуле 16S рРНК. Использование этого метода в систематике бактерий послужили К. Везе основой для построения филогенетического древа бактерий.
Сравнивать филогенетическую и фенотипическую систематики очень трудно. В одни и те же филогенетические ветви попадают крайне разнообразные по физиологии организмы. При переходе от крупных ветвей грамположительных и грамотрицательных бактерий к низшим иерархическим уровням таких несовпадений становится все больше.
КУРСОВАЯ РАБОТА
по биологии
“Происхождение эукариотических
Выполнила: Сурова Вера
10-В класс,школа N55
Проверила: Струкова Н.А.
г.Рязань 1999 год
1 . Симбиотическая теория:
1.1 Фундаментальное разграничение между прокариотами и эукариотами;
Прокариотические организмы;
Эукариотические клетки;
Митохондрии и пластиды;
Способность эукариот к фотосинтезу;
2. Предшественники жизни;
3. Ферментирующие организмы:
Источники энергии ранних клеток;
Метаногенез как источник энергии;
Цианобактерии и хлорооксибактерии;
Фотосинтез и воздух;
Ассоциации и эукариоты:
Теории последовательных симбиозов;
Симбиотическое приобретение подвижности;
Эукариоты – фотосинтетики;
Взгляд на эволюцию клетки:
6.1 Гипотеза прямой филиации;
Гипотеза: от цианобактерий к водорослям;
Совместимость 2 –х гипотез: прямой филиации и
эндосимбиоза;
Литература.
ТЕОРИЯ ЭНДОСИМБИОЗА Все живые организмы на Земле обычно делят на прокари-от и эукариот. Главной особенностью прокариот в отличие от эукариот является отсутствие у них полноценного клеточного ядра, покрытого мембраной. Клетки прокариот имеют очень не-большие размеры порядка 1 мкм. Объем эукариотических клеток, содержащих полноценное ядро, в 800 – 1000 раз больше объема клеток прокариот. В связи с этим клеткам – эукариотам необходим для выполнения жизненных функций гораздо больший мембранный аппарат. Действительно, в эукариотической клетке выявлены субклеточные структуры, окруженные незави-симыми от плазмалеммы и друг от друга мембранами. Эти структуры называются органеллами. Каждой органелле: митохондрии, хлоропласту, ядру, лизосомам, аппарату Гольджи и т. д. присущи свои специфические функции в жизни клетки. С помощью молекулярно – биологических подходов получены убедительные доказательства того,что все живые существа должны быть разделены на 3 надцарства: бактерии, археи и эукариоты. Бактерии и археи возникли на Земле от общего предка - так называемого прогенота - около 4 млрд лет назад, а эукариоты по данным палеонтологов появились на Земле через 500 млн лет после прокариот. Наиболее популярной и достаточно обоснованной идеей происхождения эукариотических клеток, в настоящее время являктся идея многократного эндосимбиоза различных прокариот. По представлениям известных микробиологов А. Клюйвера и К. Ван Ниля, именно у древних прокариот произошел отбор клеток с наиболее экономичными и зачастую полифункциональными биохи-мическими реакциями, ставшими основой важнейших путей метаболизма.
Симбиотическая теория происхождения и эволюции клеток основана на двух концепциях, новых для биологии.Согласно первой из этих концепций, самое фундаментальное разграни-чение в живой природе - это разграничение между прокариотами и эукариотами,т.е. между бактериями и организмами, состоящими из клеток с истинными ядрами - протистами, животными, грибами и растениями. Вторая концепция состоит в том, что источником некоторых частей эукариотических клеток была эволюция симбиозов, т.е. формирование постоянных ассоциаций между организмами разных видов. Предполагается, что три класса органелл - митохондрии, реснички и фотосинтезирующие пластиды – произошли от свободно живущих бактерий, которые в результате симбиоза были в определенной последовательности включены в состав клеток прокариот - хозяев. Эта теория в большой мере опирается на неодарвинистские представления, развитые генетиками, экологами, цитологами и другими учеными, которые связали Менделевскую генетику с дар-виновской идеей естественного отбора.Она опирается также на совершенно новые или недавно возрожденные научные направ-ления: на молекулярную биологию,особенно на данные о структуре белков и последовательности аминокислот, на микро палеонтологию, изучающую наиболее ранние следы жизни на Земле, и даже на физику и химию атмосферы, поскольку эти науки имеют отношение к газам биологического происхсждения. Все организмы, состоящие из клеток, могут быть сгруппиро-ваны в пять царств: царство прокариот (Monera, куда относятся бактерии) и четыре царства эукариот (Protoctista, Animalia, Fungi , Plantae). Протоктисты- это эукариотические организмы, не относящиеся к животным, грибам или растениям. В царство протоктистов входят водоросли, протозои, слизевики и другие эукариотические организмы неясной принадлежности. Протисты определены более ограничительно- как однокле-точные эукариоты. Таким образом, царство Protoctista включает не только протистов – одноклеточных эукариот, но и их ближай-ших многоклеточных потомков, таких как красные и бурые водоросли,а также многие микроорганизмы,сходные с грибами,например хитридиевые. Согласно традиционному представ-лению о прямой филиации, такие клеточные органеллы, как митохондрии и пластиды, возникли путем компартментализации самой клетки.
НЕСКОЛЬКО ПРОКАРИОТ ОБРАЗУЮТ ОДНУ ЭУКАРИОТУ
Клетки - это ограниченные мембранами системы, наименьшие из биологических единиц, способных к саморепродукции. Даже мельчайшие клетки содержат гены в форме молекул ДНК (одной или нескольких) и белоксинтезирующий аппарат, состоящий из нескольких типов РНК и множества белков. Все клетки содержат рибосомы – тельца диаметром около 0,02 мкм, состоящие из РНК по меньшей мере трех типов, и примерно пятидесяти различных белков. Прокариотичесие организмы – наименьшие биологические единицы, которые удовлетворяют этому определению клетки. У них нет ядер. С другой стороны, одиночные эукариотические (имеющие ядро) клетки в известном смысле вообще не являются единицами. Это комплексы различных белоксинтезирующих единиц (ну-клеоцитоплазмы, митохондрий и пластид) , тип и число которых варьирует в зависимости от видовой принадлежности. Таким образом, согласно этой теории, эукариотические клетки возникли в результате кооперации первоначально независимых элементов, объединившихся в определенном порядке. Тэйлор назвал такое представление о раздельном происхождении и развитии частей эукариотической клетки и их последующем объединении - теорией последовательных эндосимбиозов. Условия высокой температуры и кислотности привели к выработке в нуклеоцитоплазме особого класса белков - гистонов - для защиты ее ДНК. Митохондрии были способны расщеплять трехуглеродные соединения до углекислоты и воды. Симбиотическая ассоци-ация нуклеоцитоплазмы и митохондрий начала изменяться в сторону усиления их взаимной зависимости. Этот комплекс включил в себя двигательные органеллы - реснички и жгутики. За этой трансформацией последовало морфогенетическое преобразование, которое состояло в развитии выраженных клеточных асимметрий. С интеграцией взаимозависимых нуклеоцитоплазмы, митохондрий, ундулиподий - завершилось создание полигеномной структуры эукариотической клетки. Первая эукариотическая клетка была гетеротрофной. Она питалась готовыми органическими соединениями. У этих эукариот возникли процессы митоза, а затем и мейоза. И наконец, приобретение эукариотами способности к фотосинтезу, имело место в процессе становления основной эукариотической организации и после его завершения- последнее событие в зтой эпопее. Симбиозы становились более и более интегрированными. В настоящее время зависимость каждой органеллы от продуктов метаболизма других органелл стала настолько полной, что только современные методы анализа позволяют проследить метаболические пути исходных партнеров. Эукариоты разнообразны по своему строению, но метаболически они остаются единообразными. Метаболические ухищрения эукариот были разработаны различными бактериями еще до того времени, когда они объединились, превратившись в хозяина и его органеллы. Прокариоты достигли большого разнообразия в способах получения энергии и в тонкостях биосинтеза. Они оставили след своего существования задолго до того, как на основе эукариотической клетки сформировались такие крупные организмы, как животные, обладающие скелетом.
Схема происхождения эукариотических клеток путем симбиоза
ПРЕДШЕСТВЕННИКИ ЖИЗНИ
Предки нынешних организмов возникли более трех милли-ардов лет назад из органических соединений, образовавшихся на поверхности Земли. Привлекательность этой концепции отчасти связана с ее доступностью для проверки.
С. Миллер и Х.. Юри показали, что при атмосферных усло-виях, имевших место на юной Земле, могут образовываться аминокислоты и другие органические молекулы. Различные типы органических молекул образовывались спонтанно в течение первого миллиарда лет существования Земли. Многие органические молекулы были обнаружены в межзвездном пространстве и в метеоритах. Значит, органические соединения возникают при взаимодействии легких элементов, в при-сутствии источников энергии. Универсальность химического механизма биорепродукции означает, что все живое на Земле происходит от общих предков: вся нынешняя жизнь произошла от клеток с информационными системами, осно-ванными на репликации ДНК, и на синтезе белков, направляемом информационной РНК.
ФЕРМЕНТИРУЮЩИЕ ОРГАНИЗМЫ
Возможно, что нуклеиновые кислоты с самого начала были отделены от окружающей среды липопротеидными мембранами. Информация стала кодироваться последовательностью нуклеотидов в молекулах ДНК. Древнейшие клетки использовали АТФ и аминокислоты, входящие в состав белков, как прямые источники энергии, и как структурные компоненты. Итак - древнейшая жизнь существовала в форме гетеротрофных бактерий, получавших пищу и энергию из органического материала абиотического происхождения. Для неодарвиновской органической эволюции необходимо три феномена: воспроизведение, мутации и селективное давление среды. Воспроизведение - это неприменное условие биологи-ческой эволюции. При достаточно точной репродукции - мутации и естественный отбор неизбежны. Объяснить проис-хождение такой репродукции - значит понять происхождение самой жизни. Мутирование - первичный источник изменчивости в эволюции, ведет к наследуемым изменениям в самовоспроизводяйщейся системе. Как только выработалось надежное воспроизведение, началась неодарвиновская эволю-ция. Организмы, у которых выработались пути синтеза нужных клетке компонентов, имели преимущество перед другими. Древнейшими автотрофами, то есть организмами способными удовлетворять свои потребности в энергии и органических веществах за счет неорганических источников, были анаэробные метанообразующие бактерии. Способность к фотосинтезу впервые выработалась у бактерий, чувствительных к кислороду. Поглощая солнечный свет с помощью бактериального хлорофилла, они превращали угле-кислоту атмосферы в органические вещества.Появление ана-эробного фотосинтеза необратимо изменило поверхность и атмосферу Земли.
Углекислота удалялась из атмосферы и превращалась в ор-ганическое вещество в соответствии с общим уравнением фотосинтеза: 2H2X+CO2=CH2O+2X+H2O . Фотосинтезирующие бактерии, которые использовали для восстановления СО2 сероводород, откладывали элементарную серу. Они дали начало бактериям, способным к фотосинтезу с выделением кислорода. Вероятно этот тип фотосинтеза зародился в результате накопления мутаций у фотосинтезирующих серных бактерий. Этот путь привел к возникновению сине-зеленых водорослей или цианобактерий. Недавно была открыта и изучена новая группа фотосинтезирующих прокариот, выделяющих О2 - хлороксибактерии. Можно предполагать, что именно циано- и хлорооксибактерии были предками пластид, находящихся в клетках водорослей и растений. По мере накопления О2 в результате микробного фотосинтеза земная атмосфера становилась окислительной, что вызвало кризис всей жизни – до этого времени мир был заселен анаэробными бактериями. Чтобы выжить микробы должны были избегать кислорода, укрываясь в болотах, или должны были выработать обмен веществ, способный предохранить их от возрастающего количества кислорода.
ФОТОСИНТЕЗ И ВОЗДУХ
Примерно 2 миллиарда лет назад один за другим происходили различные эволюционные процессы по созданию метаболи-ческого разнообразия. Способность к выработке специфических ферментов может передаваться в пределах одного вида с помощью вирусоподобных частиц, называемых плазмидами. Поразительны биосинтетические способности прокариот: из углекислоты, нескольких солей и восстановленных неорга-нических соединений, служащих источниками энергии, микро-организмы- хемоавтотрофы могут синтезировать в содержащей кислород неосвещенной воде все сложнейшие макромолекулы, необходимые для роста и размножения. Эти биохимические про-цессы приводили к системам круговорота элементов еще тогда, когда не было растений, грибов, животных.
Ассоциации и эукариоты.
Взгляд на эволюцию клетки.
Прямая филиация.
Традиционный взгляд на происхождение и эволюцию эукарио-тических клеток заключается в том, что все живые организмы произошли прямым путем от единственной предковой популя-ции в результате накопления мутаций под действием естест-венного отбора.
Точечные мутации, дупликации, делеции и другие наследствен-ные изменения, как известно игравшие роль в эволюции живот-ных и растений, ответственны и за дифференцировку эукарио-тических клеток из прокариотических- теория прямой филиации. т. е. представление о несимбиотическом происхождении клеток. Возможно, зеленые водоросли являются потомками родоначаль-ников всех растений. Многие особенности этих водорослей наводят на мыысль, что они и растения имеют общего предка. Филогения животных сложнее, но концепция прямой филиации вероятно приложима также и к их путям эволюции. Губки очевидно произошли от протистов, независимо от остальных групп животных. Итак, по крайней мере для животных и растений гипотеза, по которой сложные эукариотические орга-низмы развились из более просто устроенных эукариот путем накопления отдельных мутаций под действием естественного отбора, вполне правомерна. Эволюционные новшества опирают-ся на сохранение высокосовершенных благоприятных генов. Итак, новые организмы с повышенной способностью контроли-ровать свою среду обитания развивались больше благодаря сохранению дупликации и рекомбинации высокоадаптированных генов,чем путем приобретения новых свойств в результате простых, случайных точечных мутаций. Гипотеза прямой филиации не позволяет объяснить происхождение эукариот от прокариот. Например хиатус между неспособными к митозу цианобактериями, и такими формами с вполне развитым митозом, как красные водоросли, трудно понять с позиций прямой филиации, но он естественно вытекает из симбиотической теории.
Согласно теории прямой филиации и теории симбиоза, все организмы на Земле произошли от биохимически одно-типных бактериальных предков, а растениям и животным дали начало эукариотические микробы. Фотосинтезирующие бактерии дали начало водорослям, и в конце концов расте-ниям, а некоторые водоросли утратили свои пластиды и пре-вратились в предков грибов и животных. Теоретики прямой филиации согласны в том, что пластиды и другие органеллы эукариот, включая ядро, сформировались путем дифферен-циации внутри самих клеток и называют это ботаническим мифом.
Филогения типов растений
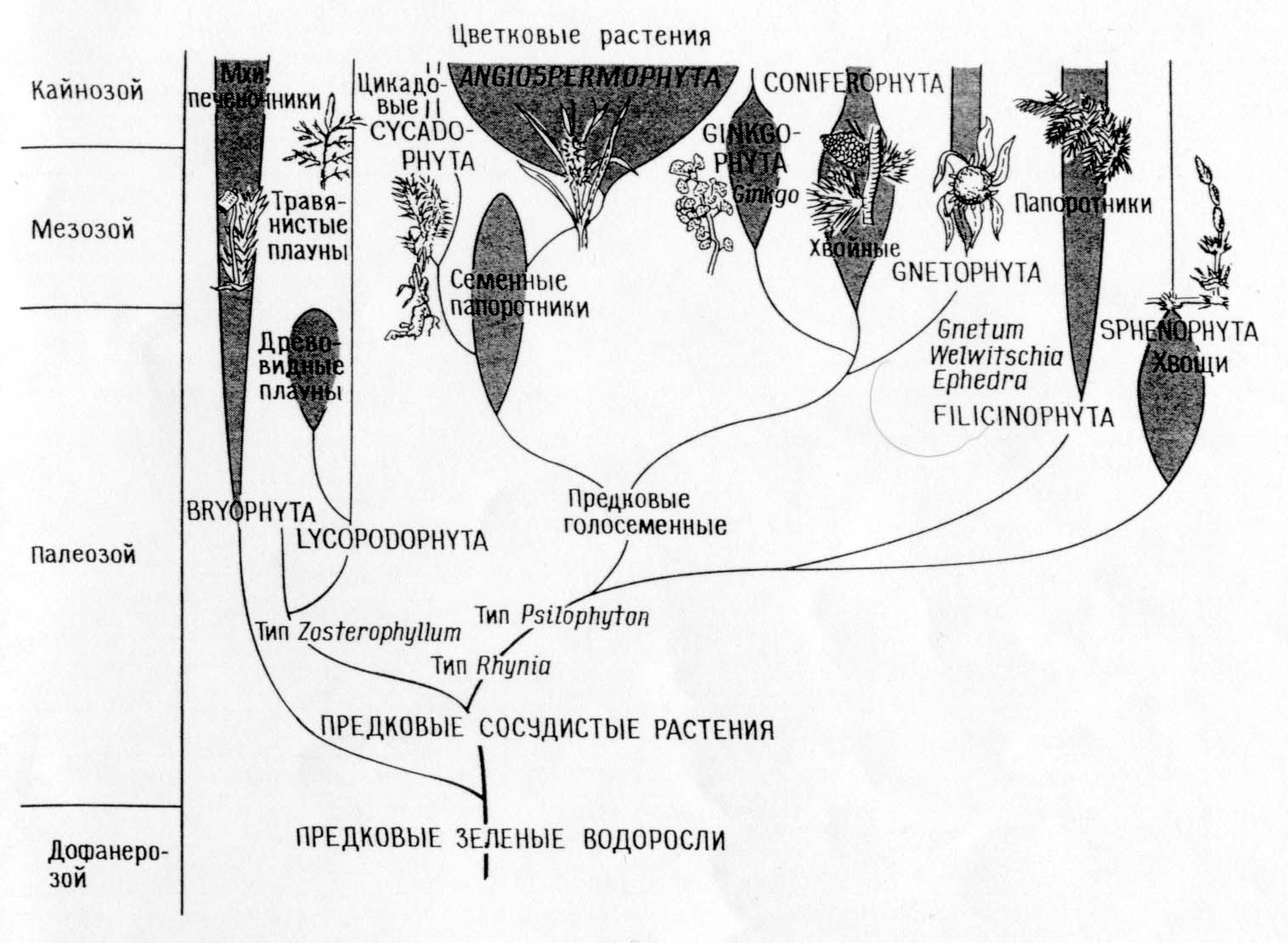
Сопоставление филогений, соответствующих теории прямой филиации (А)
и теории симбиоза (Б)
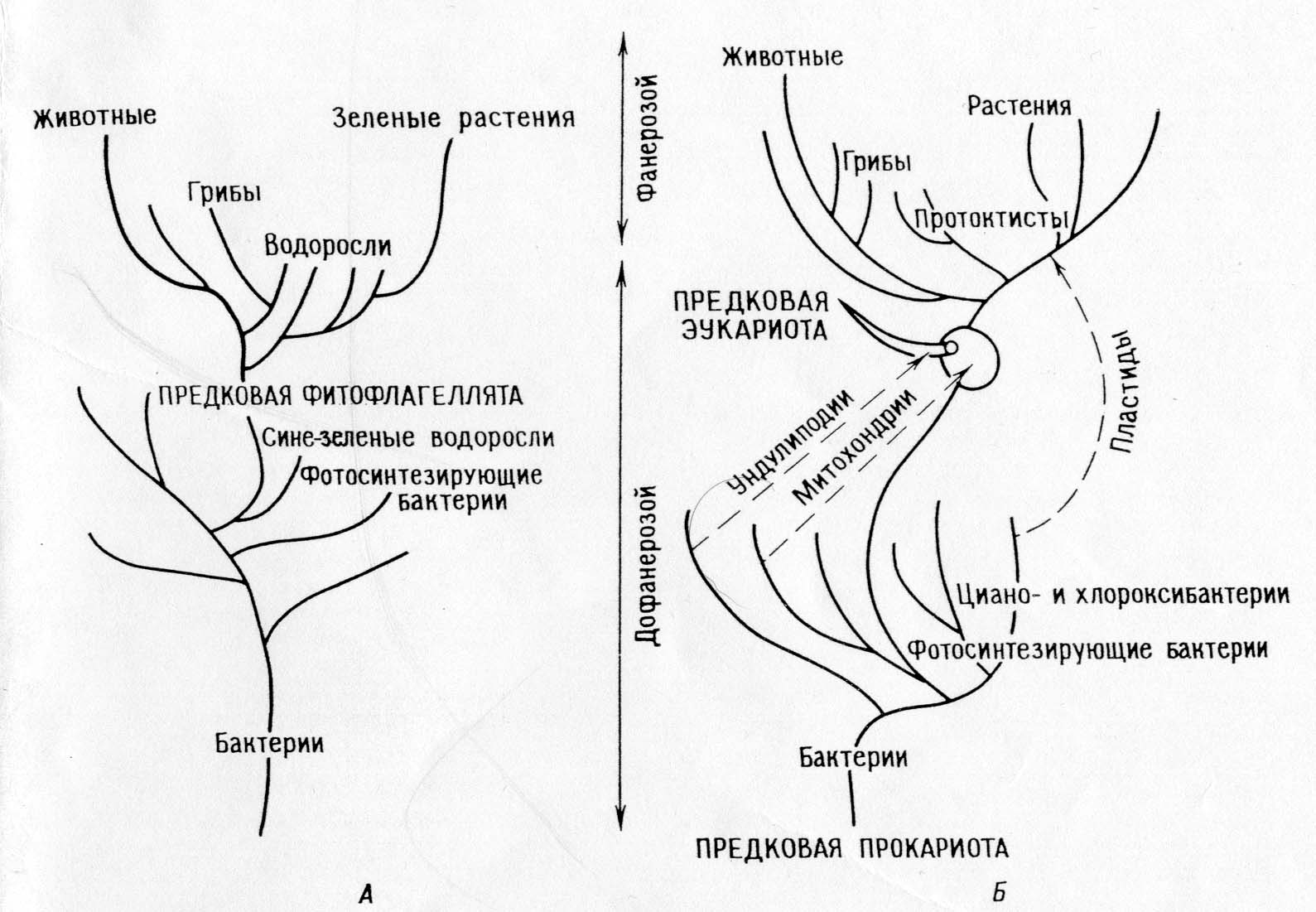
Различия между положениями теории прямой филиации и симбиотической теории:
Теория прямой филиации.
Главная дихотомия: животные – растения
Фотосинтезирующие эукариоты (водоросли и зеленые растения) произошли от фотосинтезирующих прокариот (сине-зеленых водорослей)
Растения и их фотосинтезируюшшие системы в докембрии эволюционировали монофилетически.
Животные и грибы произошли от автотрофов, утративщих пластиды.
Митохондрии дифференцировались внутриклеточно у фотосинтезирующих праводорослей.
У праводорослей внутриклеточно дифференцировались ундолиподии, митотический аппарат и все прочие эукариотические органеллы.
Все организмы произошли от предков – прокариот путем накопления одиночных мутаций.
СЛЕДСТВИЕ: согласно теории, должны существовать праводоросли; трудно связать эту теорию с ископаемой летописью и построить соответствующую филогению.
Теория последовательных эндосимбиозов:
Главная дихотомия: прокариоты – эукариоты.
Фотосинтезирующие эукариоты (ядросодержащие водоросли и растения) и нефотосинтезирующие эукариоты (животные, грибы, простейшие) произошли от общих гетеротрофных предков (амебофлагеллят).
Эволюция фотосинтеза в архее была монофилетической у бактерий (включая циано- и хлорооксибактерий), фотосин-тезирующие протисты возникли внезапно, когда их гетеро-трофные протисты – предки вступили в симбиоз с фото-синтезирующими прокариотами (протопластидами).
Животные и большинство водорослей и грибов произошли непосредственно от гетеротрофных протистов.
Митохондрии уже были у гетеротрофных протистов,когда последние в результате симбиоза приобрели пластиды. Промитохондрии были ранее приобретены симбиотическим путем.
Митоз выработался у гетеротрофных эукариотических протистов путем дифференциации ундолиподиальной системы.
Прокариоты произошли от ранних прокариотических предков путем ряда одиночных мутационных событий, эукариоты тоже произошли от эукариотических предков благодаря одиночным мутационным событиям, эукариотические же предки произошли от прокариот в результате серии симбиозов.
Следствия: теория объясняет биохимические пути и их избыточность у органелл. Она позволяет построить последо-вательную филогению, объясняющую “биологический разрыв “ между временами протерозоя и фанерозоя. Теория предска-зывает, что у органелл должны сохраняться остатки систем репликации.
Ботанический миф
Со времен Жюсье низшие растения называют таллофитами.а высшие - бриофитами и трахеофитами. Согласно ботаническому мифу,общие предки фотосинтезирующих бактерий и сине-зеленых водорослей путем прямой филиации дали начало эукариотическим водорослям в том числе морским. Группы организмов, филогению которых в наибольшшей мере затра-гивает концепция прямой филиации, оказались в области ботаники - это бактерии, водоросли, грибы и растения.
Гипотетические организмы, которые могли бы быть связующим звеном между цианобактериями и зелеными водорослями, были названы родоначальными фито-монадами (ancestral phytomonads) или праводорослями. Некоторые авторы утверждали, что подходящими кандитами на роль праводорослей могут быть эукариоты Cyanidium и Cyanophora, сходные по своим пигментным системам с цианобактериями. У таллофитов не было обнаружено промежуточных форм между немитотическим состоянием цианобактерий, и вполне развитым типичным митозом зеленых водорослей и растений. Хотя грибы и цветковые растения полностью лишены ундулиподий, им свойственны митоз и мейоз, для которых необходимы веретена, состоящие из микротрубочек. Имеется общее согласие в том,что все многоклеточные организмы,обладающие микротрубочками,происходят от эукариотических микроорганизмов с ундолиподиями, но никто не знает, от какой именно группы протистов. Подкреплением ботанического мифа служили древность и сложность фотосинтеза, а токже тот факт, что у многих организмов (у бактерий, евгленовых, пара-зититических растений) как в природе, так и в лаборатории, наблюдалась утрата фотосинтеза в результате мутаций. Поскольку фотосинтез - анаэробный процесс,обеспечивающий в конечном итоге существование всех организмов, то он должен был развиться очень рано в истории жизни на нашей планете, еще до того как возник митоз, для которого небходим кис-лород. в фотосинтезе участвует множество пигментов, липидов и ферментов,упорядоченно расположенных в очень сложных мембранах. При этом детали фотосинтетических механизмов у водорослей и растений, с одной стороны, и у цианобактерий, с другой - поразительно сходны. Все эти организмы освобождают газообразный кислород из воды, которая служит источником атомов водорода, используемых для восстановления СО2 до органических веществ клетки. Многие водоросли и простейщие не имеют ни митоза, ни генетической системы, сходной с таковой животных и растений. Инфузории имеют особую гене-тическую систему с ядрами 2 типов, причем клетки нередко многоядерные. Один тип яядер – макронуклеусы, а другой – намного меньшие микронуклеусы. Микронуклеусы физиологи-чески несущественны. однако они хранят копии генов, а при половом процессе - претерпевают мейоз. Физиологически необходимые макронуклеусы служат местом транскрипции РНК, и, таким образом, управляют физиологией клетки. Одна из трудностей при выяснении отношений между цианобактериями и эукариотическими водорослями связана с отсутствием данных о происхождении ундолиподий: не было найдено никаких промежуточных звеньев между цианобактериями, у которых ни-когда не бывает ундолиподий, и множеством водорослей, которые их имеют. Другая проблема – полное отсутствие полового процесса у цианобактерий: между бесполыми цианобактериями и водорослями, большинству которых свойственен половой процесс, не оказалось никаких промежуточных форм. Обнаружилось, что большинство водо-рослей имеют ундолиподии на какой – либо стадии жизненного цикла. Подвижные структуры у водорослей, простейших, жи-вотных и растений оказались одинаковыми: все они состоят из микротрубочек диаметром 0,024 мкм. Выяснилось,что мито-тическое веретено даже у грибов, у которых нет ундолипдий,состоит из белковых микротрубочек того же диаметра, что и в ундолиподиях. На протяжении полувека казалось чрезвычайно правдоподобным, что эукариотичкские водоросли произошли от цианобактерий. Этот ботанический миф привел к предположе-ниям, которые многие годы служили как стимулом для исследования, так и системой координат,в которой про-изводилась оценка получаемых результатов. Утверждение, что связующие звенья между цианобактериями и эукариотическими водорослями вымерли, не оставив ископаемых следов и живых реликтов, становилось все менее убедительным по мере поступ-ления новых данных о живых микробах и об окаменелостях. Главные промежуточные звенья между фотосинтезирующими прокариотами и эукариотами не вымерли бесследно, как того требует ботанический миф,- они никогда не существовали. Согласно теории последовательных эндосимбиозов, циано-бактерии – действительно родоначальники,но только пластид, а не остальных частей эукариотической клетки, и уж во всяком случае не ее ядра. И все же в вопросе о происхождении ядра гипотеза прямой филиации и теория симбиоза совместимы. Мыслимые модификации крайнего варианта теории симбиоза включают, например,идею о симбиотическом происхождении пластид, но не митохондрий, или о симбиотическом про-исхождении пластид и митохондрий, но не ундолиподий. Сторонники прямой филиации выдвинули также предположение о происхождении ядра путем симбиоза.
Решающие доказательства эндосимбиотического происхождения хлоропластов и митохондрий.
Наиболее существенные доказательства происхождения важней-ших органелл эукариотических клеток - хлоропластов и мито-хондрий из прокариот получены в последние годы при изуче-нии структуры рибосомальных РНК. Молекудярный биолог
К. Воз показал уже в 80-х годах, что структура РНК из рибосом хлоропластов растений почти идентична структуре РНК из некоторых цианобактерий. В то же время группа исследователей установила, что РНК рибосом, полученных из митохондрий разных эукариотов схожа с РНК некоторых бактерий, в частности бактерий рода Paracoccus . Набор дыхательных ферментов указанной бактерии очень похож на
“ дыхательный ансамбль “ митохондрий животных.
Эти данные достаточно ясно свидетельствуют в пользу большей вероятности эндосимбиотического происхождения эукариотических клеток. Явление эндосимбиоза и по сей день чрезвычайно широко распространено в природе. Известно, что на теле некоторых морских рыб часто поселяются светящиеся бактерии, которые не только сами получают большие преимущества для своей жизни, но и очень полезны для организмов хозяев. Все сказанное убедительно свидетельствует в пользу эндосимбиотического происхождения эукариотических клеток из гораздо меньщих по объему клеток прокариот. Однако главный вопрос заключается в установлении природы хозяйской клетки, ведь клетки эукариот обладают хорощо сформированным ядром, имеющим оболочку, а прокариоты такого ядра не имеют. Л. Маргелис, например, считает,что роль хозяйских клеток, возможно, сыграли прокариоты – предшественники современных бактерий, относящиеся к микоплазмам - очень примитивно построенным прокариотам, практически лишенным клеточных стенок. В последнее время появились и широко обсуждаются работы японского ученого
Т. Ошимы. На основании тщательного изучения структуры рибосомных РНК многих про- и эукариот, в том числе и многих представителей до сих пор малоизученного надцарства архей, этот автор приходит к выводу, что хозяйской клеткой являлся один из предков современных архей. Следует сказать, что у представителей именно этого надцарства, в отличие от бактерий, имеется очень много общих с эукариотами биохи-мических и молекулярно – биологических особенностей. В по-следних работах Т. Ошима приводятся доводы в пользу того, что хозяйской клеткой при возникновении эукариот являлся предок архей, подобный одному из современных представителей этих прокариот - термоплазме. Каково будет окончательное решение проблемы установления природы хозяйской клетки, давшей начало эукариотической клетке в процессе множествен-ного эндосимбиоза, покажут дальнейшие исследования.
Наиболее популярной и обоснованной гипотезой происхождения эукариотических клеток является в настоящее время представле-
ние об их образовании путем многократно происходившего в процессе эволюции в течении первых 1.5 – 2 млрд лет существования на Земле живых клеток эндосимбиоза различных прокариот, относящихся к бактериям и археям. Решающие доказательства эндосимбиотического происхождения хлоро-пластов и митохондрий были получены ведущими биологами мира при изучении структуры рибосомальных РНК. Эти данные достаточно ясно свидетельствуют в пользу большой вероятности эндосимбиотического происхождения эукарио-тических клеток из гораздо меньших по объему (на несколько порядков) клеток прокариот.
ЛИТЕРАТУРА
Соросовский образовательный журнал. Биология, Химия, Науки о земле, Физика, Математика. N 5 1998г
Маргелис Л. Роль симбиоза в эволюции клетки. Пер. с англ., М. МИР 1983г
Из основных разделов кинетики изучает кинетику биологических реакций; этот раздел принято называть биокинетикой. Биокинетика является пограничной наукой, возникшей на стыке биохимии и химической кинетики (см рис. 1). Выделение биокинетики в отдельную дисциплину неслучайно, оно логически оправдано и связано с исключительной значимостью кинетических процессов для всех живых организмов. Биокинетика...
Генов, но и создавала предпосылки для накопления в них изменений, дивергенции генов, увеличения разнообразия контролируемых ими продуктов. 2. Подвижные генетические элементы Определенная роль в эволюции геномов как про-, так и эукариотических клеток принадлежит так называемым подвижным генетическим элементам - транспозонам. Они представляют собой автономные единицы, несущие в нуклеотидной...
Представлены несколько возможных подходов к развитию будущей Общей Теории Динамических Систем. В главе 10 "Циркулярные процессы жизни" мы рассматриваем характерную циклическую организацию самоорганизующихся диссипативных систем. Обобщенная схема ставит характеристики генерирования трансформирующих циклов и циклов каталитических реакций, а также каталитических гиперциклов в соответствии с...