1. Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
2. Жгутики бактерий. Расположение жгутиков. Перитрихи. Монотрихи. Политрихи. Лофотрихи. Амфитрихи. Феномен роения. Диагностика подвижности бактерий.
3. Микроворсинки бактерий. Фимбрии бактерий. F-пили (секс-пили) бактерии. Клеточная оболочка бактерий. Гликокаликс.
4. Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
5. Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.
6. Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
7. Цитоплазматическая мембрана (ЦПМ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство.
9. Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
10. Усваиваемые бактериальной клеткой соединения. Пути поступления веществ в бактериальную клетку. Пассивный перенос. Диффузия.
Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Её основные составляющие - растворимые ферменты и растворимые РНК (мРНК и тРНК). Разнообразные органеллы, характерные для эукариотической клетки, у бактерий отсутствуют, а их функции выполняет бактериальная ЦПМ, отделяющая цитоплазму от клеточной стенки. У подавляющего числа бактерий цитоплазма относительно неподвижна, но у видов Streptococcus, Proteus, Clostridium имеются специальные трубочки - рапидосомы, аналогичные микротрубочкам простейших.
Бактериальный геном
В бактериальной клетке нет ядерной мембраны, ДНК сконцентрирована в цитоплазме в виде клубка. Поскольку в эукариотических клетках ДНК обычно находится в ядре , то по аналогии ДНК бактерий назвали нуклеоидом [от лат. nucleus, ядро + греч. eidos, сходство]. Её также называют генофором , или бактериальной хромосомой. Генофор бактерий представлен двойной спиральной, кольцевой, ковалентно замкнутой суперспирализованной молекулой ДНК. Она составляет 2-3% сухой массы клетки (более 10% по объёму). Генофор не содержит гистонов. Объём генетической информации, кодируемой в генофоре , различается в зависимости от вида бактерии (например, геном Escherichia coli кодирует примерно 4000 различных полипептидов). У бактерий может присутствовать дополнительная ДНК в виде включений. Эти включения, или плазмиды , несут ряд различных генов, кодирующих дополнительные свойства бактерий, но информация, содержащаяся в плазмидах, не является абсолютно необходимой для бактериальной клетки.
Бактериальные рибосомы
Бактериальные рибосомы - сложные глобулярные образования, состоящие из различных молекул РНК и связанных с ними белков. Всё образование функционирует как локус синтеза полипептидов. В зависимости от интенсивности роста бактериальная клетка может содержать от 5000 до 50 000 рибосом. Диаметр бактериальных рибосом около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70 S (единиц Свёдберга), тогда как у эукариотических клеток - 80 S. Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50 S и 30 S (у эукариотов 40 S и 60 S). Объединение субъединиц происходит перед началом трансляции. Рибосомы прокариот и эукариотов имеют сходную молекулярную структуру и механизмы функционирования, но различаются, помимо размеров, по составу белков и белковых факторов. Эти различия делают рибосомы эукариотов практически резистентными к действию антибиотиков, блокирующих синтез белка у бактерий.
Запасные гранулы бактерии
Запасные гранулы содержат временный избыток метаболитов; наличие и количество гранул изменяется в зависимости от вида бактерий и их метаболической активности. В виде гранул могут запасаться полисахариды (крахмал, гликоген, гранулёза), жиры (триглицериды, сходные с жирами высших животных, запасаются у дрожжей рода Candida), воска - у микобактерий и нокардий; полимеры р-оксимасляной кислоты (например, в клетках Bacillus megaterium), полифосфаты (волютин) у Spirillum volutans и Corynebacterium diphtheriae, сера (у бактерий, окисляющих сульфиды), кристаллизованные белки (например, токсичный для насекомых протоксин у Bacillus thuringiensis).
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как её ультраструктурной, так и химической организации.
Главная особенность прокариотной клетки - отсутствие ядра, ограниченного от цитоплазм двойной мембраной. Наследственный материал сосредоточен в бактериальной хромосоме, обычно представленной в виде кольцевой молекулы двухцепочечной ДНК. У некоторых спирохет, родококков, агробактерий обнаружены хромосомы линейного типа, а у актиномицетов - "псевдокольцевые". У некоторых видов в клетке обнаружены две или даже три неидентичные хромосомы. Нитевидная молекула ДНК выполняет функцию ядра и располагается в центральной зоне клетки, называемой "нуклеоидом ". В электронном микроскопе на срезе бактериальной клетки эта зона выглядит более светлой, чем остальная цитоплазма.
У бактерий может присутствовать дополнительная молекула ДНК в виде внехромосомных элементов либо интегрированных в генофор. подобные включения обозначаются термином "плазмиды" (соответственно эписомальные и интегрированные). Для ДНК эписом тоже характерна кольцевая форма, но по размеру эписомы меньше бактериальной хромосомы Плазмиды несут ряд различных генов и часто определяют вирулентность бактерий, но информация, содержащаяся в плазмидах, не абсолютно необходима для бактериальной клетки.
Цитоплазматическая мембрана , окружающая каждую клетку, определяет её величину и обеспечивает сохранение существенных различий между клеточным содержимым и окружающей средой. В клетке бактерий отсутствует мембранная сеть - эндоплазматический ретикулум, цитоплазматическая мембрана, лежащая под клеточной стенкой и отгораживающая снаружи протопласт, является единственной мембраной в клетке.
Но мембрана - это не просто механическая перегородка. Она служит также высокоизбирательным фильтром, который поддерживает разницу концентраций ионов по обе стороны мембраны и позволяет питательным веществам проникать внутрь клетки, а продуктам выделения выходить наружу. Цитоплазматическая мембрана представляет собой ансамбль липидных и белковых молекул. удерживаемых с помощью нековалентных взаимодействий. В электронном микроскопе мембраны имеют вид листков толщиной около 7 нм с выраженной трёхслойной структурой, двойной слой липидных молекул определяет структурные особенности мембраны, белки же ответственны за большинство мембранных функций.
В цитоплазматической мембране прокариот существуют многочисленные впячивания - инвагинации, которые выполняют разные функции. Они могут содержать ферменты энергетического метаболизма и соответственно выполнять функции митохондрий или являться мезосомами и участвовать в делении клетки. Из цитоплазматической мембраны формируются покровы эндоспор. Она представляет собой также главный барьер для проникновения веществ внутрь клетки.
Структуры, расположенные снаружи от цитоплазматической мембраны, - клеточная стенка, капсула, слизистый чехол, жгутики, фимбрии, пили, ворсинки - являются поверхностными структурами.
Клеточная стенка - важный и обязательный структурный компонент большинства прокариотных клеток (истинных бактерий, не считая архебактерий и микоплазм). Располагается под капсулой или слизистым чехлом или (в случае отсутствия последнего) - контактирует с окружающей средой. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придаёт клетке определённую присущую ей форму - ригидность. Состав и строение клеточной стенки – важный систематический признак, по которому все прокариоты подразделяются на следующие группы: грамположительные, грамотрицательные и не имеющие клеточной стенки.
Названия этих групп происходят от способности или неспособности разных бактерий окрашиваться по Граму. Датским врачом К. Грамом, занимавшимся окрашиванием тканей, в 1884 г. был предложен метод окраски, который позднее использован для бактерий. Его суть мы подробно рассмотрим на лабораторном занятии, а пока отметим, что клеточные стенки грамположительных и грамотрицательных бактерий резко различаются как по химическому составу, так и по ультраструктуре.
Г+ бактерии хорошо удерживают внутри клетки красящий комплекс генцианового фиолетового с иодом, а Г - - теряют окраску после обработки спиртом.
Эмпирически найденные различия между двумя группами бактерий в отношении окраски по Граму оказались коррелятивно связанными со многими другими важными признаками. Например, Г+ бактерии размножаются бинарным делением за счёт формирования поперечной перегородки в клетке, грамотрицательные - путём образования перетяжки. Первые чувствительны к пенициллину, вторые - нет. Г- бактерии не образуют эндоспор, а некоторые Г+ их образуют.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотичных организмов. Клеточная стенка прокариот состоит из нескольких слоёв, из них основной - каркасный - слой, ответственный за прочность, формируется из особого вещества - пептидогликана муреина, который синтезируется только прокариотной клеткой и не встречается у других организмов. Это поперечносшитый биополимер, гетерополисахарид, формирующий замкнутый мешок, полностью покрывающий клетку снаружи. Муреин составляет основную массу клеточной стенки Г+ бактерий (в 40 раз больше, чем Г -). Полисахаридный состав молекулы построен из чередующихся остатков N -ацетилглюкозамина и N - ацетилмурамовой кислоты , соединённых между собой с помощью гликозидных связей. Муреиновый мешок достаточно эластичен и под действием внешних факторов может растягиваться и сжиматься до 4-х раз. Интересно, что такая биологическая структура, как муреин, возникла в ходе эволюции дважды, поскольку некоторые археи содержат аналогичный по строению биополимер, состоящий из других исходных материалов.
В состав клеточной стенки входят также тейхоевые кислоты и полисахариды. Тейхоевые кислоты представляют собой полимеры, построенные на основе многоатомных спиртов (рибита и глицерина), соединённых между собой фосфодиэфирными связями. Считается, что тейхоевые кислоты придают муреиновому мешку определённую степень свободы при растяжении и сжатии и действуют наподобие пружин в матрасе.
У Г- эубактерий строение клеточной стенки намного сложнее. В её состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к цитоплазматической мембране. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки - наружная мембрана , которая состоит из фосфолипидов, белков, липопротеина и липополисахарида. Между цитоплазматической и внешней мембранами возникает уникальное периплазматическое пространство. Внешняя мембрана содержит белки-порины, формирующие поры. Кроме того, она обеспечивает контакт клеток между собой, с поверхностью субстрата и удерживает ряд внешних структурных образований, например пили.
Пили – нитеобразные полимерные органеллы белковой природы, локализованные на поверхности клеток. Обозначает все типы нежгутиковых образований на поверхности клетки (фимбрии). Пили выступают акцепторами бактериофагов. Помогают клеткам принимать и передовать ДНК при коньюгации и принимать участие в движении клетки. Основное назначение – поддержка специфических прикрепительных структур клеток.
Клеточная стенка прокариот механически защищает клетку от воздействия окружающей среды, обеспечивает поддержание её внешней формы, даёт возможность клетке существовать в гипотонических растворах. С клеточной стенкой связана проницаемость и транспорт веществ в клетку.
Под цитоплазматической мембраной у бактерий находится цитоплазма . Это коллоидная система, состоящая из воды, белков, жиров, углеводов, минеральных соединений и других веществ, соотношение которых варьирует в зависимости от вида бактерий и их возраста. В цитоплазме могут находиться включения (мезосомы, газовые вакуоли) и запасные вещества (β-гидроксибутират, гранулы гликогена и серы, карбоксисомы, пароспоральные тельца). Некоторые из включений просто лежат в цитоплазме, другие окружены тонкой мембраной (2-4 нм).
Цитоплазма прокариотной клетки не подвержена циклическому движению, как у растений или сократительным явлениям, как у амёб. Бактериальные клетки осуществляют движение за счёт других механизмов: либо при помощи просто устроенных жгутиков, отличающихся от сложных жгутиков эукариот, они активно плавают в жидкой среде либо скользят, выделяя слизь и перемещаясь по поверхности плотного субстрата. Несколько модифицированный тип движения наблюдается у спирохет, которые обладают «аксиальной нитью».
Бактериальные жгутики располагаются по полюсам или по разным сторонам клетки. В зависимости от числа и расположения жгутиков различают монотрихи (один полярный жгутик), амфитрихи (жгутики на двух полюсах), лофотрихи (пучок жгутиков) и перетрихи (жгутики по всей поверхности клетки). Жгутики построены иначе. Чем у эукариот (9+2). У бактерий жгутики правовращающие, у архей – левовращающие. Жгутик состоит из базального тела, включающего 4 или 2 кольца, стержень,белки мотора, а также из крючка и филломента (цилиндр состоит из белка флагеллина).Подвижные клетки бактерий активно перемещаются в направлении, определяемом теми или иными факторами. Такие направленные перемещения бактерий называют "таксисами".
Покоящиеся клетки бактерий представлены у Г+ бактерий эндоспорами, у Г- -цистами. Споры бактерий обладают уникальной устойчивостью к неблагоприятным воздействиям. Устойчивость спор связана с наличием у них особого слоя в клеточной стенке - кортекса .
Эндоспоры остаются жизнеспособными в течение 500 лет, споры актиномицетов до 7500 лет, бацилл -25-30 млн лет (в кишечнике пчелы в кусочке янтаря).
Тип спосрообразования (бациллярный, клостридиальный и плектридиальный) часто помогает идентифицировать культуру.
Циста – потерявшая подвижность клетка с утолщённой ослизненной оболочкой. Устойчивая к высушиванию и перепадам температур. Цисты характеризуются состоянием покоя со значительно сниженной метаболической активностью. Функции: защищают от вредных воздействий окружающей среды, служат способом сохранения ДНК, способ передачи информационного начала от хозяина к хозяину (у патогеннов). Прорастание стимулируется благоприятными внешними условиями.
Снаружи клеточные стенки прокариот часто бывают окружены слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоёв, чехлов или влагалищ (у нитчатых бактерий). Все они являются результатом биосинтеза прокариотами органических полимеров, в основном полисахаридной природы, и отложения их вокруг клеток. Как правило, химический состав капсул, образуемых бактериями, родо- и видоспецифичен. Капсулы и чехлы защищают клетку от повреждений, высыхания, создают дополнительный барьер, служат препятствием для проникновения фагов, создают вокруг клеток особую среду, обеспечивающую определённый водный режим.
Форма клетки прокариот очень разнообразна. Выделяют три основные формы бактерий - шаровидные или овальные - кокки, палочки (бациллы) и спиралевидные.
Кокки подразделяют на парные - диплококки, тетракокки , пакетообразные кокки или сарцины , располагающиеся этажами, стрептококки - в цепочках, стафилококки , образующие бесформенные скопления, напоминающие гроздья винограда.
Среди палочек выделяют одиночные , беспорядочно расположенные, диплобациллы , расположенные попарно и стрептобациллы - в цепочках
Спиралевидные бактерии разделяют на две группы - вибрионы и спирохеты и спириллы , имеющие изгибы, равные одному или нескольким оборотам спирали.
Многоклеточные прокариоты имеют форму нитей или червеобразную форму.
Таким образом, бактерии морфологически сравнительно слабо дифференцированы, поэтому среди них можно различить лишь ограниченное число форм. С таким внешним "единообразием" удивительно контрастирует чрезвычайное многообразие метаболических процессов.
Основы классификации бактерий
Классификация микроорганизмов, причисляемым к прокариотам, учитывает в первую очередь практические моменты и служит для распознавания описанных форм. Под классификацией понимают распределение единиц по группам более высокого порядка. Она осуществляется по иерархической системе. Основной единицей является чистая культура выделенной бактерии - "штамм". Штаммы объединяются в виды (species), виды в роды (genus, мн. числ. genera), а роды в семейства. Основой для классификации служит адекватное описание штаммов. в соответствии с которым и проводят сравнение и разграничение рассматриваемых единиц. Установлением соподчинения отдельных групп организмов занимается таксономия .
Различают два вида классификаций: филогенетические "естественные" и искусственные. Построение естественной классификации - конечная цель таксономии бактерий, которая состоит в том, чтобы объединить родственные формы, связанные общностью происхождения, и на этой основе создать филогенетическое древо бактерий. Несомненно, когда-нибудь это удастся сделать, исходя из химических признаков - таких, как последовательность аминокислот в функционально сходных белках или последовательность нуклеотидов в консервативных нуклеиновых кислотах, например в рибосомных РНК.
Искусственная классификация ставит перед собой более скромные задачи. Она довольствуется объединением организмов в отдельные группы на основе их сходства и используется для идентификации и определения организмов. Искусственная система рассчитана на использование её в качестве ключа для определения. Наиболее полный труд, в котором описаны бактерии, - это "Руководство по определению бактерий Берги". Первое издание определителя, подготовленное Американским обществом микробиологов, вышло в 1928, последнее - девятое - в 1994. Это признанный во всём мире, регулярно переиздаваемый многотомный труд, подготовленный международным коллективом из лучших специалистов по той или иной группе микроорганизмов (около 300 авторов). На русский 9-е американское издание переведено в 1997 г, благодаря бескорыстной работе русских микробиологов, с готовностью отдавших своё время и знания для перевода соответствующих глав. Это справочник для идентификации бактерий, необходимый в каждой лаборатории.
Разделение бактерий по группам на основании фенотипических признаков (морфологии, физиологии) удобно в практических целях для быстрой идентификации организмов. Организмы объединяются в искусственные группы, или "секции", не имеющие таксономического статуса и часто обозначаемые по наиболее характерному роду или названию. По такому принципу построен Определитель Берджи (9-е изд., 1997)
Краеугольное для каждой классификации понятие "вид" для бактерий до сих пор не имеет чёткого определения. Разнообразные механизмы изменчивости бактерий обусловливают определённую нестабильность признаков, совокупность которых определяет тот или иной вид. Поэтому в систематике бактерий широко применяют понятие "вариант". Различают морфологические, биологические, ферментативные, серологические и многие другие варианты
В микробиологии также применяют специализированные понятия и термины - "штамм" и "клон". Термин "штамм" определяет культуру микроорганизмов, выделенную из определённого конкретного источника. клоном называют культуру микроорганизмов, полученную из одной материнской клетки.
Главными отличиями прокариотической (бактериальной) клетки от эукариотической является: отсутствие оформленного ядра (т.е. ядерной мембраны), отсутствие внутриклеточных мембран,ядрышек, комплекса Гольджи, лизосом, митохондрий.
Основными структурами бактериальной клетки являются:
Нуклеоид – представляет собой наследственный (генетический) материал бактериальной клетки, представлен 1 молекулой ДНК, замкнутой в кольцо и суперспирализованной (скручена в рыхлый клубок). Длина ДНК около 1мм. Объем информации около 1000 генов (признаков). Нуклеоид не отделен от цитоплазмы мембраной.
Цитоплазма – коллоид, т.е. водный раствор белков, углеводов. Липидов, минеральных веществ, в котором находятся рибосомы, включения, плазмиды.
На рибосомах происходит биосинтез белка. Рибосомы прокариот отличаются от эукариотических более мелкими размерами (70 S).
Включения – запасные питательные вещества бактериальной клетки, а также скопления пигментов. К запасным питательным веществам относятся: гранулы волютина (неорганического полифосфата), гликоген, гранулеза, крахмал, капли жира, скопления пигмента, серы, кальция. Включения, как правило, образуется при выращивании бактерий на богатых питательных средах и исчезает при голодании.
Клеточная мембрана – ограничивает цитоплазму. Сосотоит из двойного слоя фосфолипидов и встроенных мембранных белков. КМ кроме барьерной и транспортной функций выполняют роль центра метаболической активности (в отличие от эукариотической клетки). Белки мембраны, ответственные за перенос необходимых веществ в клетку, называют пермеазами. На внутренней поверхности КМ находятся ферментные ансамбли, т.е.упорядоченные скопления молекул ферментов, ответственных за синтез энергоносителей – молекул АТФ. КМ может образовывать впячивания в цитоплазму, которые называют мезосомами.Существует два вида мезосом:
Септальные – образуют поперечные перегородки в процессе деления клетки.
Латеральные – служат для увеличения поверхности КМ и повышения скорости обменных процессов.
Нуклеоид, ЦП и КМ образуют протопласт.
Одним из отличительных свойств бактерий является очень высокое внутриклеточное осмотическое давление (от 5 до 20 атм), что является результатом интенсивного обмена веществ. Поэтому для защиты от осмотического шока бактериальная клетка окружена прочной клеточной стенкой.
По строению клеточной стенки все бактерии делятся на 2 группы: Имеющие однослойную клеточную стенку – Грам-положительные. Имеющие двухслойную клеточную стенку – Грам-отрицательные. Названия Грам+ и Грам- имеют свою предисторию. В 1884 датский микробиолог Ганс Христиан Грам разработал оригинальный метод окраски микробов, в результате которого одни бактерииокрашивались в синий цвет (грам+), а другие в красный (грам-). Химическая основа различной окраски бактерий по методу Грама была выяснена сравнительно недавно – около 35 лет назад. Оказалось, что Г- и Г+ бактерии имеют разное строение клеточной стенки. Строение клеточной стенки Г+ бактерий. Основу клеточной стенки Г+ бактерий составляют 2 полимера: пептидогликан и тейхоевые кислоты. Пептидогликан представляет собой линейный полимер, в котором чередуются остатки мурамовой кислоты и ацетилглюкозамина. С мурамовой кислоте ковалентно связан тетрапептид (белок). Нити пептидогликана связаны между собой через пептиды и образуют прочный каркас – основу клеточной стенки. Между нитями пептидогликана находится другой полимер – тейхоевые кислоты(глицерол ТК и рибитол ТК) - полимер полифосфатов. Тейхоевые кислоты выступают на поверхности клеточной стенки и являются главными АГ Г+ бактерий. Кроме этого, в состав клеточной стенки Г+ бактерий входит рибонуклеат Mg. Стенка Г- бактерий состоит из 2-х слоев: внутренний слой представлен моно- или бислоем пептидогликана (тонкий слой) . Наружный слой состоит из липополисахаридов, липопротеина, белков, фосфолипидов. ЛПС всех Г- бактерий обладают токсическими и порогенными свойствами и называются эндотоксинами.
При воздействии некоторых веществ, например пенициллина, нарушается синтез пептидогликанового слоя. При этом из Г+ бактерий образуется протопласт, а из Г- сферопласт (т.к. сохраняется наружный слой клеточной стенки).
При определенных условиях культивтрования клетки, лишенные клеточной стенки, сохраняют способность к росту и делению, и такие формы называют L- формами (по названи. Института Листера, где было открыто это явление). В некоторых случаях после устранения фактора, тормозящего синтез клеточной стенки L-формы могут превратиться в исходные формы.
Многие бактерии синтезируют слизистое вещество, состоящее из мукополисахаридов, которое откладывается с наружной стороны клеточной стенки, окружая бактериальную клетку слизистым чехлом. Это капсула. Функция капсулы – защита бактерий от фазоцитоза.
Поверхностные структуры бактериальной клетки.
Органы прикрепления к субстрату (адгезии) – пили (фимбрии) или реснички. Начинаются от мембраны клетки. Сосотоят из белка пилина. Число пилей может достигать 400 на 1 клетку.
Органы передачи наследственной информации – F-пили или sex-пили. F-пили образуются только в том случае, если клетка нечет плазмиду, т.к. белки F-пили кодирует ДНК плазмиды. Они представляют собой тонкую длинную трубочку, которая прикрепляется к другой бактериальной клетке. Через образовавшийся канал плазмида переходит в соседнюю бактериальную клетку.
Органы движения – жгутики – представляют собой спиральные нити. Их длина может превышать их диаметр в 10 и более раз. Жгутики состоят из белка флагеллина. Основание жгутика связано с клеточной мембраной посредством базального тельца. Базальное тельце состоит из системы колец, которые вращаясь передают вращательное движение жгутику. По расположению жгутиком бактерии делятся на моно-, лофо-, амфи-, перитрихи.
Конец работы -
Эта тема принадлежит разделу:
Медицинская микробиология, ее предмет, методы, связь с другими науками
Микроскопия люминесцентная свечение под воздействием энергии света эл лучей ионизир излуч Собственная без окраски наведенная окр... Микроскопич метод метод изучения морф и тинкториальных св в бактерий на... Химический состав бактериальной клетки Вода...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Медицинская микробиология, ее предмет, методы, связь с другими науками
Микробиология –наука о микроорганизмах, т.е. о живых существах, размеры которых меньше 0,1 мм. Микроорганизмы весьма разнообразны. К ним относятся некоторые многоклеточные организмы, простейшие, не
Основные этапы развития микробиологии
I. Эвристический – Гиппократ – причина болезней невидимые вещества – миазмы
II. Морфологический – Левенгук – обнаружил в микроскоп живые организмы. Ивановский – открыл вирусы.
III
Микроорганизмы и их положение в системе живого мира
Классификация микроорганизмов.
Представлены доклеточными (царство Vira – вирусы) и клеточными формами (бактерии, грибы, простейшие). Среди клеточных форм жизни различают 3 домена: Bacteria
Химический состав бактериальной клетки
Химический состав бактерий
Вода – 70%, Сухое вещество – 30%. Белки – 52, полисахариды – 16. Липиды – 9,4, РНК – 16, ДНК – 3.2, неорганические соединения – 0,4.
Потребность бактери
Спорообразование.
При неблагоприятных условиях некоторые бактерии способны образовывать эндоспоры - покоящиеся клетки. Одна клетка образует одну эндоспору, т.е. спорообразование не является формой размножения, а слу
Действие физических факторов на микроорганизмы
Стерилизация - полное освобождение какого-либо вещества или предмета от вегетативных форм и спор микроорганизмов путем воздействия на них физических и химических факторов. В хирургии стерилизуют
Действие химических факторов на микроорганизмы. Дезинфекция
Дезинфекция (обеззараживание) - (франц. "des" означающее удаление, уничтожение чего-либо; лат "inficere" -заражать, портить, отравлять) - удаление и уничтожение вегетативных фо
Питание бактерий
По типу питания бактерии подразделяются на аутотрофов и гетеротрофов. Аутотрофы способны усваивать углерод из СО2.
Гетеротрофы усваивают углерод только из органических соединений.
Питательные среды
В лабораторных условиях бактерии выращивают на питательных средах. Большое значение для роста и размножения бактерий имеют температурные условия. Все микроорганизмы по отношению к температурному р
Дыхание бактерий
Дыхание относится к реакциям катаболизма. В результате дыхания происходит расщепление сложных молекул до простых с выделением энергии, которая запасается в молекулах АТФ (КПД около 40%).
А
Ферменты бактерий, их биологическая роль
Ферменты - биологические катализаторы высокомолекулярной структуры, вырабатываемые живой клеткой. Они имеют белковую природу, строго специфичны и играют важнейшую роль в обмене веществ микроорганиз
Рост и размножение микроорганизмов
Под размножением бактерий подразумевают способность их к самовоспроизведению, увеличению количества особей на единицу объема. Рост означает увеличение массы бактерий в результате синтеза клеточного
Актиномицеты, их морфология
Актиномицеты (аей§ - луч, туЬм - гриб) - лучистые грибы, представляют собой многочисленную группу микроорганизмов, включенных в порядок АсйпотусеШез,
Большинство актиномицетов - свободнож
Риккетсии, их морфология и биологические свойства
Риккетсии представляют собой полиморфные микроорганизмы, живут и размножаются только в клетках (цитоплазме и ядре) тканей животных, человека и переносчиков. Кокковидные формы имеют вид очень мелких
Морфология и ультраструктура микоплазм
Микоплазмы принадлежат к классу Mollicutes, порядку Mycoplasmatales, семейству Мусор1аsmaсеае. Это мелкие бактерии размером 100-150 нм, иногда 200-700 нм, не образующие спор, неподвижные, грамотри
Хламидии, морфология и другие биологические свойства
К роду Chlamydia, семейству Chlamydiaceae, порядку Chlamydiales принадлежат возбудители трахомы, конъюнктивитов (бленнореи с включениями), пахового лимфогранулематоза (болезнь Никола - Фавра), орн
Грибы, их морфология и биологические свойства
Систематика и классификация грибов. Грибы отнесены к растительным гетеротрофным организмам-эукариотам, лишенным хлорофилла. Тип грибов (Fungi s. Mycetes) насчитывает свыше 100 000 видов, объедине
Дрожжеподобные грибы рода кандида
Возбудители кандидоза. К ним относят дрожжеподобные грибы из рода Candida. Они представляют собой одноклеточные организмы (рис. 134), размножающиеся почкованием; конидий и аскоспор не образуют, ист
Дейтеромицеты
Дейтеромицеты - несовершенные грибы (Fungi imperfecti), очень большая группа (25 000 видов) грибов, обладающих многоклеточным мицелием, но не имеющих ни сумчатого, ни базидиального спороношения, а
Взаимодействие вируса с клеткой
Репродукция вирусов.
Цикл репродукции вирусов состоит из ряда последовательных событий:
Специфическая адсорбция вируса на поверхности клетки
Проникновение внутрь клетки
Культуры клеток, их виды
Для культивирования вирусов используют культуры клеток, куриные эмбрионы и лабораторных животных. Наиболее широкое применение нашли однослойные культуры трипсинизированных клеток, а также перевивае
Изменчивость микроорганизмов
Исследователи, стоявшие у истоков бактериологии, столкнулись с разнообразием видов бактерий и кажущейся возможностью метаморфоз, взаимопревращений одних бактерий в другие создали учение о плеоморфи
Мутации
Изменения последовательности азотистых оснований в ДНК.
Виды:
Выпадение (делеция) или вставка (инсерция) одного или нескольких оснований со сдвигом рамки считывания.
Заме
Генетические рекомбинации
Рекомбинации - Наследственная изменчивость, обусловленная обменом участками ДНК между микроорганизмами, один из которых является донором, а другой – реципиентом. Виды рекомбинаций: Трансформация, к
Распространение микробов в природе
Объект
Характер загрязнения
Санитарно-показательные бактерии
Вода
Фекаль
Нормальная микрофлора тела человека
Макроорганизм и среда представляют единую экологическую систему, в которой важная физиологическая роль принадлежит микроорганизмам. Макроорганизм и его микрофлора (аутофлора) в нормальных условиях
Для изучения строения бактериальной клетки наряду со световым микроскопом применяют электронно-микроскопические и микрохимические исследования, позволяющие определить ультраструктуру бактериальной клетки.
Бактериальная клетка (рис. 5) состоит из следующих частей: трехслойной оболочки, цитоплазмы с различными включениями и ядерного вещества (нуклеоида). Дополнительными структурными образованиями являются капсулы, споры, жгутики, пили.
Рис. 5. Схематическое изображение строения бактериальной клетки. 1 - оболочка; 2 - слизистый слой; 3 - клеточная стенка; 4 - цитоплазматическая мембрана; 5 - цитоплазма; 6 - рибосома; 7 - полисома; 8 - включения; 9 - нуклеоид; 10 - жгутик; 11 - пили
Оболочка клетки состоит из наружного слизистого слоя, клеточной стенки и цитоплазматической мембраны.
Слизистый капсульный слой находится снаружи клетки и выполняет защитную функцию.
Клеточная стенка - один из основных структурных элементов клетки, сохраняющий ее форму и отделяющий клетку от окружающей среды. Важным свойством клеточной стенки является избирательная проницаемость, которая обеспечивает проникновение в клетку необходимых питательных веществ (аминокислот, углеводов и др.) и выведение из клетки продуктов обмена. Клеточная стенка сохраняет внутри клетки постоянное осмотическое давление. Прочность стенки обеспечивает муреин, вещество полисахаридной природы. Некоторые вещества разрушают клеточную стенку, например лизоцим.
Бактерии, полностью лишенные клеточной стенки, называются протопластами. Они сохраняют способность к дыханию, делению, синтезу ферментов; к воздействию внешних факторов: механическому повреждению, осмотическому давлению, аэрации и др. Сохранить протопласты можно только в гипертонических растворах.
Бактерии с частично разрушенной клеточной стенкой называются сферопластами. Если подавить процесс синтеза клеточной стенки с помощью пенициллина, то образуются L-формы, которые у всех видов бактерий представляют шаровидные крупные и мелкие клетки с вакуолями.
Цитоплазматическая мембрана плотно прилегает к клеточной стенке с внутренней стороны. Она очень тонкая (8-10 нм) и состоит из белков и фосфолипидов. Это пограничный полупроницаемый слой, через который осуществляется питание клетки. В мембране находятся ферменты пермеазы, осуществляющие активный перенос веществ, и ферменты дыхания. Цитоплазматическая мембрана образует мезосомы, принимающие участие в делении клетки. При помещении клетки в гипертонический раствор мембрана может отделиться от клеточной стенки.
Цитоплазма - внутреннее содержимое бактериальной клетки. Она представляет собой коллоидную систему, состоящую из воды, белков, углеводов, липидов, различных минеральных солей. Химический состав и консистенция цитоплазмы изменяются в зависимости от возраста клетки и условий окружающей среды. В цитоплазме находятся ядерное вещество, рибосомы и различные включения.
Нуклеоид, ядерное вещество клетки, ее наследственный аппарат. Ядерное вещество прокариотов в отличие от эукариотов не имеет собственной мембраны. Нуклеоид зрелой клетки представляет собой двойную нить ДНК, свернутую в кольцо. В молекуле ДНК закодирована генетическая информация клетки. По генетической терминологии ядерное вещество получило название генофор или геном.
Рибосомы находятся в цитоплазме клетки и выполняют функцию синтеза белка. В состав рибосомы входит 60% РНК и 40% белка. Количество рибосом в клетке достигает 10000. Соединяясь вместе, рибосомы образуют полисомы.
Включения - гранулы, содержащие различные запасные питательные вещества: крахмал, гликоген, жир, волютин. Они расположены в цитоплазме.
Клетки бактерий в процессе жизнедеятельности образуют защитные органеллы - капсулы и споры.
Капсула - внешний уплотненный слизистый слой, примыкающий к клеточной стенке. Это защитный орган, который появляется у некоторых бактерий при попадании их в организм человека и животных. Капсула предохраняет микроорганизм от защитных факторов организма (возбудители пневмонии и сибирской язвы). Некоторые микроорганизмы имеют постоянную капсулу (клебсиеллы).
Споры встречаются только у палочковидных бактерий. Они образуются при попадании микроорганизма в неблагоприятные условия внешней среды (действие высоких температур, высыхание, изменение рН, уменьшение количества питательных веществ в среде и т. д.). Споры находятся внутри бактериальной клетки и представляют уплотненный участок цитоплазмы с нуклеоидом, одетый собственной плотной оболочкой. По химическому составу они отличаются от вегетативных клеток малым количеством воды, увеличенным содержанием липидов и солей кальция, что способствует высокой устойчивости спор. Спорообразование происходит в течение 18-20 ч; при попадании микроорганизма в благоприятные условия спора в течение 4-5 ч прорастает в вегетативную форму. В бактериальной клетке образуется только одна спора, следовательно, споры не являются органами размножения, а служат для переживания неблагоприятных условий.
Спорообразующие аэробные бактерии называются бациллами, а анаэробные - клостридиями.
Споры отличаются по форме, размерам и расположению в клетке. Они могут располагаться центрально, субтерминально и терминально (рис. 6). У возбудителя сибирской язвы спора располагается центрально, ее размер не превышает поперечника клетки. Спора возбудителя ботулизма расположена ближе к концу клетки - субтерминально и превышает ширину клетки. У возбудителя столбняка округлая спора располагается на конце клетки - терминально и значительно превышает ширину клетки.
Жгутики - органы движения, характерны для палочковидных бактерий. Это тонкие нитевидные фибриллы, состоящие из белка - флагеллина. Длина их значительно превышает длину бактериальной клетки. Жгутики отходят от базального тельца, расположенного в цитоплазме, и выходят на поверхность клетки. Наличие их можно обнаружить по определению подвижности клеток под микроскопом, в полужидкой питательной среде или при окраске специальными методами. Ультраструктура жгутиков изучена в электронном микроскопе. По расположению жгутиков бактерии делят на группы (см. рис. 6): монотрихи - с одним жгутиком (возбудитель холеры); амфитрихи - с пучками или единичными жгутиками на обоих концах клетки (спириллы); лофотрихи - с пучком жгутиков на одном конце клетки (фекальный щелочеобразователь); перитрихи - жгутики расположены по всей поверхности клетки (кишечные бактерии). Скорость движения бактерий зависит от количества и расположения жгутиков (наиболее активны монотрихи), от возраста бактерий и влияния окружающих факторов.
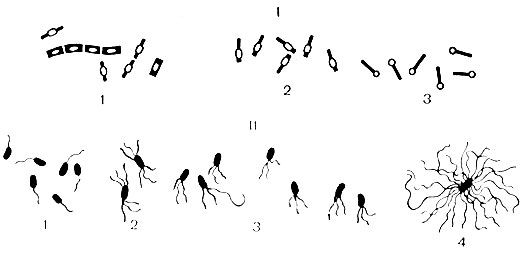
Рис. 6. Варианты расположения спор и жгутиков у бактерий. I - споры: 1 - центральное; 2 - субтерминальное; 3 - терминальное; II - жгутики: 1 - монотрихи; 2 - амфитрихи; 3 - лофотрихи; 4 - перитрихи
Пили или фимбрии - ворсинки, расположенные на поверхности бактериальных клеток. Они короче и тоньше жгутиков и также имеют спиральную структуру. Состоят пили из белка - пилина. Одни пили (их несколько сотен) служат для прикрепления бактерий к клеткам животных и человека, с другими (единичными) связана передача генетического материала из клетки в клетку.
Микоплазмы
Микоплазмы - клетки, не имеющие клеточной стенки, но окруженные трехслойной липопротеидной цитоплазматической мембраной. Микоплазмы могут быть сферической, овальной формы, в виде нитей и звезд. Микоплазмы по классификации Берги выделены в отдельную группу. В настоящее время этим микроорганизмам уделяется все большее внимание как возбудителям заболеваний воспалительного характера. Размеры их различны: от нескольких микрометров до 125-150 нм. Мелкие микоплазмы проходят через бактериальные фильтры и называются фильтрующимися формами.
Спирохеты
Спирохеты (см. рис. 52) (от лат. speira - изгиб, chaite - волосы) - тонкие, извитые, подвижные одноклеточные организмы, имеющие размеры от 5 до 500 мкм в длину и 0,3-0,75 мкм в ширину. С простейшими их роднит способ движения путем сокращения внутренней осевой нити, состоящей из пучка фибрилл. Характер движения спирохет различен: поступательное, вращательное, сгибательное, волнообразное. В остальном строение клетки типичное для бактерий. Некоторые спирохеты слабо окрашиваются анилиновыми красителями. Спирохеты разделяют на роды по количеству и форме завитков нити и ее окончанию. Кроме сапрофитных форм, распространенных в природе и организме человека, среди спирохет имеются болезнетворные - возбудители сифилиса и других заболеваний.
Риккетсии
Вирусы
Среди вирусов выделяют группу фагов (от лат. phagos - пожирающий), вызывающих лизис (разрушение) клеток микроорганизмов. Сохраняя присущие вирусам свойства и состав, фаги отличаются структурой вириона (см. главу 8). Они не вызывают заболеваний человека и животных.
Контрольные вопросы
1. Расскажите о классификации микроорганизмов.
2. Назовите основные свойства представителей царства прокариотов.
3. Перечислите и охарактеризуйте основные формы бактерий.
4. Назовите основные органеллы клетки и их назначение.
5. Дайте краткую характеристику основных групп бактерий и вирусов.
Один из основных признаков прокариотической клетки - отсутствие внутреннего разделения, обеспечиваемого элементарными мембранами. По сути, единственная мембранная система прокариот - ЦПМ, входящая в состав клеточной стенки, часто сложно устроенная и проникающая поперечными складками глубоко в цитоплазму.
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с рибосомами и неоформленного ядра, называемого нуклеоидом. Могут быть дополнительные структуры: капсула, микрокапсула, слизь, жгутики, включения. Некоторые бактерии способны образовывать споры для переживания неблагоприятных условий. Все структурные элементы бактериальной клетки условно можно разделить на обязательные (нуклеоид, ЦПМ, клеточная стенка, цитоплазма, 70S рибосомы) и необязательные, которые могут отсутствовать у некоторых видов (капсула, плазмиды, включения, жгутики, пили и др.)
Капсула. Клеточную оболочку многих бактерий окружает слой аморфного, гидрофильного вещества. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов; например, капсула В. anthracis состоит из полимеров D-глутаминовой кислоты. Основную роль в организации капсул играет ЦПМ. Выделяют микрокапсулы (выявляемые только при
электронной микроскопии в виде слоя мукополисахаридных фибрилл) и макрокапсулы (обнаруживают при световой микроскопии). У некоторых бактерий полимеры клеточной оболочки, выделяемые наружу, свободно располагаются вокруг нее, образуя слизистый слой. Слизь растворима в воде. Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных скистозным фиброзом. Капсула и слизь предохраняют бактерии от повреждений, высыхания, фагоцитов. воздействия бактерицидных факторов, обеспечивают адгезию на различных субстратах, могут содержать запасы питательных веществ. Капсула антигена (К-антиген). Для визуализации капсулы используют негативные методы окраски (по Бурри-Гинсу).
Клеточная стенка. Клеточная стенка является уникальной, по своему химическому составу, структурой прокариотической клетки. Она выполняет ряд важных функций: защищает бактерии от внешних воздействий, придает им характерную форму, поддерживает постоянство внутренней среды, участвует в делении, на поверхности клеточной стенки находятся рецепторы для бактериофагов, бактериоцинов и различных химических веществ, через клеточную стенку осуществляется транспорт питательных веществ и выделение метаболитов. Это прочная, упругая структура, имеющая сложное строение. Опорным каркасом клеточной стенки бактерий является пептидогликан (муреин, мукопептид) - гетерополимер, состоящий из повторяющихся дисахаридных групп, соединенных поперечными и боковыми цепочками. Дисахарид, являющийся структурной единицей пептидогликана, состоит из N - ацетилглюкозамина и N - ацетилмурамовой кислоты, соединенных β - гликозидными связями. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки. Связывание фрагментов пептидогликана заключается в образовании пептидной связи между терминальным остатком аминокислотного мостика (D-аланином) с предпоследним остатком примыкающего аминокислотного мостика (L-лизином или диаминопимелиновой кислотой). Боковые мостики образуют четыре аминокислоты, поперечные (вертикально связывающие слои пептидогликана) - пять аминокислот. В состав боковых и поперечных мостиков входят уникальные аминокислоты, присутствующие только у прокариот, что делает эти связи мишенью для ряда антибактериальных препаратов.
По строению и составу клеточной стенки микроорганизмы делятся на грамположительные и грамотрицательные, что является важным признаком, положенным в основу искусственной систематики микробов. Основным компонентом толстой клеточной стенки грамположительных бактерий является многослойный пептидогликан, составляющий 40-90 % массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков.
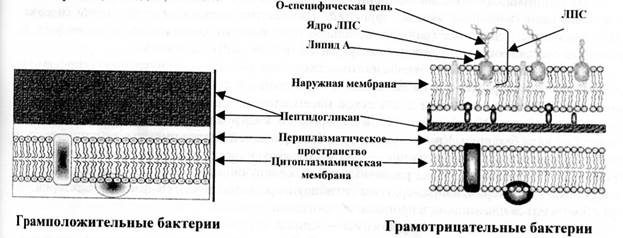
Рис. 2. Строение клеточной стенки бактерии
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Липополисахарид наружной мембраны состоит из 3 фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (от лат. соrе - ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием различных факторов образуются клетки с измененной формой (чаще шаровидной): протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана и способные размножаться, называются L-формами. Существуют нестабильные L-формы, которые могут реверсировать, «возвращаясь» в исходную бактериальную клетку, и стабильные L-формы, не способные к реверсии.
Цитоплазматическая мембрана. ЦПМ - это обязательная клеточная структура, являющаяся физическим, осмотическим, метаболическим барьером между внутренним содержимым бактериальной клетки и внешней средой. ЦПМ состоит из двух слоев фосфолипидов и встроенных в липидную мембрану белковых молекул (как большинство биологических мембран). Белки составляют 20-75%, липиды - 25-40% и в незначительных количествах в ЦПМ присутствуют углеводы и РНК, Белки ЦПМ подразделяют на структурные и функциональные. Первые образуют различные структуры ЦПМ, вторые представлены ферментами, участвующими в синтетических реакциях на поверхности мембраны и окислительно-восстановительных процессах, а также некоторыми специализированными энзимами (пермеазы участвуют в транспорте веществ). При избыточном росте (по сравнению с ростом клеточной стенки) цито-плазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Мезосомы образуют поперечные перегородки между делящимися клетками и являются местом прикрепления бактериальной хромосомы.
У некоторых бактерий между ЦПМ и клеточной стенкой располагается периплазматическое пространство - полость шириной около 10 нм. Снаружи в это пространство открываются поры клеточной стенки, изнутри выходят некоторые клеточные ферменты (рибонуклеазы, фосфатазы, |3-лактамазы).
Цитоплазма. Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Ее основные составляющие - растворимые ферменты и РНК (мРНК и тРНК).
Бактериальный геном. Эквивалентом ядра у бактерий является нуклеоид (генофор). Он расположен в центральной зоне бактерий в виде двойной суперспирализованной кольцевой молекулы ДНК. Она составляет 2-3% сухой массы клетки (более 10% по объему). Генофор не содержит гистонов. Объем генетической информации, кодируемой в бактериальной хромосоме зависит от вида бактерии. Кроме нуклеоида в бактериальной клетке имеются внехромосомные носители генетической информации - плазмиды, являющиеся ковалентно замкнутыми кольцами ДНК. Плазмиды несут ряд различных генов, кодирующих дополнительные (необязательные) признаки бактерий, например, гены антибиотикорезистентности (К-фактор). Бактерии могут обмениваться плазмидами в процессе конъюгации.
Рибосомы. Бактериальные рибосомы - сложные глобулярные образования, состоящие из молекул РНК и связанных с ними белками. Рибосомы необходимы для синтеза полипептидов. Количество рибосом в различных бактериальных клетках колеблется от 5 до 50 тысяч. Диаметр рибосом составляет около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70S (единиц Сведберга), тогда как у эукариотических клеток 80S. Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50S и 30S (у эукариот от 40S и 60S). Объединение субъединиц происходит перед началом трансляции. Рибосомальные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяющей оценить степень родства организмов. Различия в строении рибосом про- и эукариотических клеток делают рибосомы эукариот практически резистентными к действию антибиотиков, блокирующих синтез белка у бактерий.
Запасные гранулы. В цитоплазме бактерий содержатся различные включения, которые содержат временный избыток метаболитов. В виде гранул могут запасаться полисахариды (крахмал, гликоген), жиры (триглицериды, запасаются у дрожжеподобных грибов рода ^1 СапсНс1а), полимеры β - оксимасляной кислоты, полифосфаты (волютин) у C. diphtheriae, сера, кристаллизованные белки и др.
Жгутики. Жгутики бактерий являются органами движения (локомоции) бактерий. Расположение жгутиков - характерный признак, имеющий таксономическое значение. По количеству и расположению жгутиков различают монотрихи - один жгутик (V. cholerae), перитрихи (от греч. реri - вокруг и trichos - волос) - жгутики по всей поверхности бактериальной клетки (Е.сoli), лофотрихи (от греч. 1оphos - пучок и trichos - волос) - пучок жгутиков на одном конце клетки (Рseudomonas), амфитрихи (от греч. атрhi - двойной, двусторонний и trichos - волос) - единичные жгутики или пучки жгутиков на разных полюсах клетки (Spirillum). Жгутик - спирально изогнутая полая нить, образованная субъединицами белка флагеллина. Толщина жгутиков 12-20 нм, длина 3-15 мкм (больше длины клетки). Жгутики состоят из 3 частей: спиралевидной нити, крюка и базального тельца. Базальное тельце содержит стержень со специальными дисками: одна пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий. Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - мотором, вращающим жгутик. Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Жгутики являются Н-антигенами, что используется в серологической идентификации.
Подвижность бактерий определяют микроскопией препаратов «раздавленной» или «вися-
чей» капли. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные - по уколу) или посевом бактерий в водный конденсат скошенного столбика агара.
(подвижные виды переплывают из конденсата на поверхность среды и колонизируют ее).
Микроворсинки. Помимо жгутиков, поверхность многих бактерий покрыта цитоплазматическими выростами - микроворсинками, встречающимися у подвижных и неподвижных видов. Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, прикрепление бактерий к поражаемой клетке; пили, ответственные за питание, водно-солевой обмен и половые (F-пили), или коньюгационные, пили. Пили многочисленны - несколько сотен на клетку. Одна-
ко половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми «мужскими»
клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Со1- плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях. Споры. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Способностью к спорообразованию обладают только бактерии родов Bacillus и Clostridium, включающие патогенные для человека виды, и многие сапрофиты. У бактерий рода Васillus диаметр споры не превышает диаметр (поперечник) бактериальной клетки. У бактерий рода Clostridium (от лат. closter, веретено) диаметр споры превышает поперечник бактерии, что придает им форму веретена (С.perfringens), барабанной палочки (С. tetani) или теннисной ракетки (С. botulinum) при центральной, терминальной и субтерминальной локализации спор соответственно. Основное значение спор в выживаемости бактерий определяет их терморезистентность, и резистентность к другим факторам (дезинфектанты и др.), оказывающим губительное воздействие на бактерии. Высокая устойчивость спор связана с низким содержанием свободной воды, высокой концентрацией кальция, вялостью метаболических процессов, наличием дипиколиновой кислоты и белка, богатого цистеином (что делает его похожим на кератин), а также наличие нескольких оболочек, которые являются дополнительной защитой от неблагоприятных внешних воздействий. При попадании спор в благоприятные условия они прорастают, проходя три последовательные стадии: активацию, инициацию, вырастание.
Споры плохо окрашиваются и остаются бесцветными в окрашенных клетках при использовании обычных методов окраски (простые методы, метод Грама). Для окраски спор используют метод Ожешко (Ауэски).
5. Основные формы бактерий
Отдельным видам бактерий с достаточным постоянством присущи: определенная форма, размер и расположение. Длина бактериальных клеток варьирует от 0,1-0,2 мкм (виды Муcoplasma) до 10-15 мкм (виды Clostridium), толщина - от 0,1 до 2,5 мкм. Средние размеры бактерий 2-3x0.3-0.8 мкм. Выделяют три основные формы бактерий - шаровидные (кокки), палочковидные (цилиндрические) и извитые (спиральные).
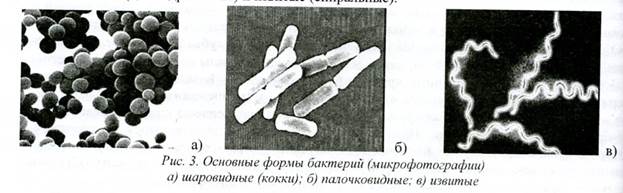
Рис. 3. Основные формы бактерии (микрофотографии) а) шаровидные (кокки); 6) палочковидные: в) извитые
Большинство кокков (от греч. kokkos - ягода, зерно) имеют шаровидную или овальную форму, клетки некоторых видов могут быть ланцетовидными (пневмококки) или бобовидными (нейссерии). По характеру расположения клеток в мазках выделяют диплококки (располагаются парами), стрептококки (располагаются цепочками), стафилококки (располагаются в виде гроздьев винограда), сарцины (располагаются пакетами по 8. 16. 32 и более клеток) и др.
Палочковидные бактерии различаются по форме концов клетки и взаимному расположению. Палочки могут быть правильной (кишечная палочка), неправильной (коринебактерии) формы, ветвящиеся (актиномицеты). Палочковидные бактерии могут располагаться в мачках одиночно и беспорядочно (монобактерии), попарно (диплобактерии) или цепочкой (стрепто-бактерии. стрептобациллы) (рис. 4).

Рис. 4. Основные формы бактерий
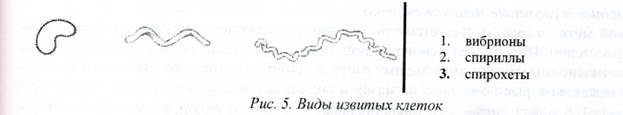
Извитые формы - представлены изогнутыми палочками. В зависимости от формы и количества завитков различают три типа клеток: вибрионы (от греч. vibrio- извиваюсь, изгибаюсь) имеют один завиток. не превышающий четверти оборота спирали (изогнутые клетки наподобие запятой): спириллы (от греч. speira- спираль) имеют 3-5 крупных завитков и спирохеты - большое количество мелких завитков (рис. 5).