STRUCTURE OF THE PLANT CELL
The plant cell is composed of more or less rigid cell wall and protoplast. The cell wall is the cell wall and cytoplasmic membrane. The term protoplast comes from the word protoplasm, which for a long time used to refer to all living things. The protoplast is the protoplasm of an individual cell.
The protoplast consists of the cytoplasm and the nucleus. The cytoplasm contains organelles (ribosomes, microtubules, plastids, mitochondria) and membrane systems (endoplasmic reticulum, dictyosomes). The cytoplasm also includes the cytoplasmic matrix (basic substance) in which organelles and membrane systems are immersed. The cytoplasm is separated from the cell wall plasma membrane, which is an elementary membrane. Unlike most animal cells, plant cells contain one or more vacuoles. These are vesicles filled with liquid and surrounded by an elementary membrane (tonoplast).
In a living plant cell, the main substance is located in in constant motion. Organelles are involved in the movement, called the flow of the cytoplasm or cyclosis. Cyclosis facilitates the movement of substances in the cell and their exchange between the cell and the environment.
Plasma membrane. It is a bilayer phospholipid structure. Plant cells are characterized by invaginations of the plasma membrane.
The plasma membrane performs the following functions:
Participates in the exchange of substances between the cell and the environment;
Coordinates the synthesis and assembly of cellulose microfibrils of the cell wall;
Transmits hormonal and external signals that control cell growth and differentiation.
Core. It is the most prominent structure in the cytoplasm of a eukaryotic cell. The kernel performs two important functions:
Controls the vital activity of the cell, determining which proteins, and at what time should be synthesized;
Stores genetic information and passes it on to daughter cells during cell division.
The nucleus of a eukaryotic cell is surrounded by two elementary membranes that form the nuclear envelope. It is permeated with numerous pores with a diameter of 30 to 100 nm, visible only in an electron microscope. The pores have a complex structure. outer membrane the nuclear membrane in some places is combined with the endoplasmic reticulum. The nuclear envelope can be viewed as a specialized, locally differentiated part of the endoplasmic reticulum (ER).
In the nucleus stained with special dyes, one can distinguish thin threads and lumps of chromatin and nucleoplasm (the main substance of the nucleus). Chromatin is made up of DNA bound to special proteins called histones. In the process of cell division, chromatin becomes more and more compacted and assembled into chromosomes. DNA encodes genetic information.
Organisms differ in the number of chromosomes in somatic cells. For example, cabbage has - 20 chromosomes; sunflower - 34; wheat - 42; humans - 46, and one of the species of the fern Ophioglossum - 1250. Sex cells (gametes) have only half the number of chromosomes characteristic of somatic cells of the body. The number of chromosomes in gametes is called haploid (single), in somatic cells - diploid (double). Cells that have more than two sets of chromosomes are called polyploid.
Under a light microscope, spherical structures - nucleoli - can be seen. Each nucleus has one or more nucleoli, which are visible in non-dividing nuclei. Ribosomal RNA is synthesized in the nucleolus. Usually in the nuclei of diploid organisms there are two nucleoli, one for each haploid set chromosomes. The nucleoli do not have their own membrane. Biochemically, the nucleoli are characterized high concentration RNA, which is here associated with phosphoproteins. The size of the nucleoli depends on functional state cells. it has been noticed that in a rapidly growing cell, in which intensive processes of protein synthesis are taking place, the nucleoli increase in size. In the nucleoli, mRNA and ribosomes are produced, which perform a synthetic function only in the nucleus.
Nucleoplasm (karyoplasm) is represented by a homogeneous liquid in which various proteins, including enzymes, are dissolved.
Plastids. Vacuoles, cellulose cell walls, and plastids are characteristic components of plant cells. Each plastid has its own shell, consisting of two elementary membranes. Inside the plastid, a membrane system is distinguished and varying degrees homogeneous substance - stroma. Mature plastids are classified based on the pigments they contain.
Chloroplasts, in which photosynthesis takes place, contain chlorophylls and carotenoids. Usually have the form of a disk with a diameter of 4 - 5 microns. In one cell of the mesophyll (middle of the leaf) there can be 40 - 50 chloroplasts; in mm 2 sheets - about 500,000. in the cytoplasm, chloroplasts are usually located parallel to the cell membrane.
The internal structure of the chloroplast is complex. The stroma is permeated with a developed system of membranes in the form of vesicles - thylakoids. Each thylakoid consists of two membranes. Thylakoids form single system. As a rule, they are collected in piles - grains, resembling columns of coins. The thylakoids of individual granae are interconnected by stroma thylakoids, or intergranal thylakoids. Chlorophyll and carotenoids are embedded in thylakoid membranes. Chloroplasts in green plants and algae often contain grains of starch and small lipid (fat) droplets. Starch grains are temporary stores of photosynthesis products. They can disappear from the chloroplasts of a plant that has been in the dark for only 24 hours, and reappear 3–4 hours after the plant is transferred to light.
Chloroplasts are semi-autonomous organelles and resemble bacteria. For example, the ribosomes of bacteria and chloroplasts have a fairly high similarity. They are smaller than eukaryotic ribosomes. Protein synthesis on the ribosomes of bacteria and chloroplasts is inhibited by chloramphenicol, which has no effect in eukaryotic cells. In addition, both bacteria and chloroplasts have a similar type of nucleoid, organized in a similar way. Despite the fact that the formation of chloroplasts and the synthesis of pigments contained in them is largely controlled by the chromosomal DNA of the cell, nevertheless, in the absence of its own DNA, chloroplasts do not form.
Chloroplasts can be considered the main cellular organelles, since they are the first in the chain of transformation of solar energy, as a result of which humanity receives both food and fuel. In chloroplasts, not only photosynthesis takes place. They are also involved in the synthesis of amino acids and fatty acids, serve as a storage of temporary reserves of starch.
Chromoplasts are pigmented plastids. Diverse in form, they do not have chlorophyll, but synthesize and accumulate carotenoids, which give yellow, orange, red color to flowers, old leaves, fruits and roots. chromoplasts can develop from chloroplasts, which in this case lose chlorophyll and internal membrane structures, accumulate carotenoids. This occurs during the ripening of many fruits. Chromoplasts attract insects and other animals with which they co-evolved.
Leukoplasts are non-pigmented plastids. Some of them synthesize starch (amyloplasts), others are capable of forming various substances, including lipids and proteins. In the light, leukoplasts turn into chloroplasts.
Proplastids are small, colorless or pale green, undifferentiated plastids found in the meristematic (dividing) cells of roots and shoots. They are the precursors of other, more differentiated plastids - chloroplasts, chromoplasts, and aminoplasts. If the development of protoplastids is delayed due to lack of light, one or more prolamellar bodies, which are semi-crystalline accumulations of tubular membranes, may appear in them. Plastids containing prolamellar bodies are called etioplasts. In the light, etioplasts turn into chloroplasts, while the membranes of the prolamellar bodies form thylakoids. Etioplasts are formed in the leaves of plants that are in the dark. protoplasts of seed embryos first turn into etioplasts, from which chloroplasts then develop in the light. Plastids are characterized by relatively easy transitions from one type to another. Plastids, like bacteria, reproduce by dividing in two. In meristematic cells, the time of division of protoplastids approximately coincides with the time of cell division. However, in mature cells most of The plastid is formed as a result of the division of mature plastids.
Mitochondria. Like chloroplasts, mitochondria are surrounded by two elementary membranes. The inner membrane forms many folds and protrusions - cristae, which significantly increase the inner surface of the mitochondria. They are significantly smaller than plastids, about 0.5 µm in diameter, and variable in length and shape.
In mitochondria, the process of respiration takes place, as a result of which organic molecules are broken down with the release of energy and its transfer to ATP molecules, the main energy reserve of all eukaryotic cells. Most plant cells contain hundreds or thousands of mitochondria. Their number in one cell is determined by the cell's need for ATP. Mitochondria are in constant motion, moving from one part of the cell to another, merging with each other and dividing. Mitochondria usually assemble where energy is needed. If the plasma membrane actively transfers substances from cell to cell, then mitochondria are located along the surface of the membrane. In motile unicellular algae, mitochondria accumulate at the base of the flagella, supplying the energy necessary for their movement.
Mitochondria, like plastids, are semi-autonomous organelles containing the components necessary for the synthesis of their own proteins. The inner membrane surrounds a fluid matrix that contains proteins, RNA, DNA, bacterial-like ribosomes, and various solutes. DNA exists in the form of circular molecules located in one or more nucleoids.
Based on the similarity of bacteria with the mitochondria and chloroplasts of eukaryotic cells, it can be assumed that mitochondria and chloroplasts originated from bacteria that found "refuge" in larger heterotrophic cells - the precursors of eukaryotes.
Microbodies. Unlike plastids and mitochondria, which are delimited by two membranes, microbodies are spherical organelles surrounded by a single membrane. Microbodies have a granular (granular) content, sometimes they also contain crystalline protein inclusions. Microbodies are associated with one or two regions of the endoplasmic reticulum.
Some microbodies, called proxisomes, play important role in the metabolism of glycolic acid, which is directly related to photorespiration. In green leaves, they are associated with mitochondria and chloroplasts. Other microbodies, called glyoxisomes, contain the enzymes needed to convert fats into carbohydrates. This happens in many seeds during germination.
Vacuoles are areas of the cell delimited by a membrane, filled with liquid - cell sap. They are surrounded by a tonoplast (vacuolar membrane).
A young plant cell contains numerous small vacuoles, which merge into one large vacuole as the cell ages. In a mature cell, up to 90% of its volume can be occupied by a vacuole. In this case, the cytoplasm is pressed in the form of a thin peripheral layer to the cell membrane. The increase in cell size is mainly due to the growth of the vacuole. As a result, turgor pressure arises and tissue elasticity is maintained. This is one of the main functions of the vacuole and tonoplast.
The main component of the juice is water, the rest vary depending on the type of plant and its physiological state. Vacuoles contain salts, sugars, less often proteins. The tonoplast plays an active role in the transport and accumulation of certain ions in the vacuole. The concentration of ions in the cell sap can significantly exceed its concentration in the environment. At high content Some substances form crystals in vacuoles. The most common calcium oxalate crystals are different shape.
Vacuoles are places of accumulation of metabolic products (metabolism). These can be proteins, acids, and even substances toxic to humans (alkaloids). Pigments are often deposited. Blue, violet, purple, dark red, crimson give plant cells pigments from the anthocyanin group. Unlike other pigments, they dissolve well in water and are found in cell sap. They determine the red and blue color of many vegetables (radishes, turnips, cabbage), fruits (grapes, plums, cherries), flowers (cornflowers, geraniums, delphiniums, roses, peonies). Sometimes these pigments mask chlorophyll in the leaves, for example, in the decorative red maple. Anthocyanins color autumn leaves bright red color. They are formed in cold sunny weather, when the synthesis of chlorophyll stops in the leaves. In leaves, when anthocyanins are not formed, after the destruction of chlorophyll, yellow-orange carotenoids of chloroplasts become noticeable. The leaves are most brightly colored in cold, clear autumn.
Vacuoles are involved in the destruction of macromolecules, in the circulation of their components in the cell. Ribosomes, mitochondria, plastids, getting into vacuoles, are destroyed. According to this digestive activity, they can be compared with lysosomes - organelles of animal cells.
Vacuoles are formed from the endoplasmic reticulum (reticulum)
Ribosomes. Small particles (17 - 23nm), consisting of approximately equal amounts of protein and RNA. In ribosomes, amino acids combine to form proteins. There are more of them in cells with an active metabolism. Ribosomes are located in the cytoplasm of the cell freely or attached to the endoplasmic reticulum (80S). They are also found in the nucleus (80S), mitochondria (70S), plastids (70S).
Ribosomes can form a complex on which there is a simultaneous synthesis of identical polypeptides, information about which is taken from one molecule and RNA. Such a complex is called polyribosomes (polysomes). Cells that synthesize proteins in large quantities have an extensive system of polysomes that are often attached to outer surface shells of the nucleus.
Endoplasmic reticulum. It is a complex three-dimensional membrane system of indefinite extent. In the context of the ER looks like two elementary membranes with a narrow transparent space between them. The shape and length of the ER depend on the cell type, its metabolic activity, and the stage of differentiation. In cells that secrete or store proteins, the ER is shaped like flat sacs or cisterns, with numerous ribosomes attached to its outer surface. This reticulum is called the rough endoplasmic reticulum. A smooth ER is usually tubular in shape. Rough and smooth endoplasmic reticulum can be present in the same cell. As a rule, there are numerous connections between them.
The endoplasmic reticulum functions as the communication system of the cell. He is associated with outer shell kernels. In fact, these two structures form a single membrane system. When the nuclear envelope ruptures during cell division, its fragments resemble ER fragments. The endoplasmic reticulum is a system for transporting substances: proteins, lipids, carbohydrates, to different parts of the cell. endoplasmic reticulum of neighboring cells are connected through cytoplasmic strands - plasmodesmata - that pass through the cell membranes.
The endoplasmic reticulum is the main site of synthesis cell membranes. In some plant cells, membranes of vacuoles and microbodies, cisterns of dictyosomes are formed here.
Golgi apparatus. This term is used to refer to all dictyosomes, or Golgi bodies, in a cell. Dictyosomes are groups of flat, disc-shaped vesicles or cisterns that branch into complex system tubules. Dictyosomes in higher plants consist of 4 - 8 cisterns, collected together.
Usually, in a pack of tanks, a developing and maturing side is distinguished. the membranes of the forming cisterns resemble the membranes of the ER in structure, and the membranes of the maturing cisterns resemble the plasma membrane.
Dictyosomes are involved in secretion and, in most higher plants, in the formation of cell membranes. Cell wall polysaccharides synthesized by dictyosomes accumulate in vesicles, which then separate from maturing cisterns. These secretory vesicles migrate and fuse with the CMP; while the polysaccharides contained in them are built into the cell membrane. Some substances that accumulate in dictyosomes are formed in other structures, for example, in the ER, and then transported to dictyosomes, where they are modified (modified) before secretion. For example, glycoproteins are an important construction material cell membrane. The protein part is synthesized by polysomes of the rough ER, the carbohydrate part is synthesized in dictyosomes, where both parts combine to form glycoproteins.
Membranes are dynamic, mobile structures that constantly change their shape and area. The concept of the endoplasmic system is based on the mobility of membranes. According to this concept, the inner membranes of the cytoplasm, except for the membranes of mitochondria and plastids, are a single whole and originate from the endoplasmic reticulum. New cisterns of dictyosomes form from the endoplasmic reticulum through the intermediate vesicle stage, and the secretory vesicles that separate from the dictyosomes eventually contribute to the formation of the plasma membrane. Thus, the endoplasmic reticulum and dictyosomes form a functional whole in which dictyosomes play a role intermediate structures in the process of converting membranes similar to the endoplasmic reticulum into membranes similar to the plasma. In tissues whose cells grow and divide weakly, membrane components are constantly being updated.
Microtubules are found in almost all eukaryotic cells. They are cylindrical structures with a diameter of about 24 nm. Their length varies. Each tubule is made up of subunits of a protein called tubulin. The subunits form 13 longitudinal filaments surrounding the central cavity. Microtubules are dynamic structures, they are regularly destroyed and formed at certain stages of the cell cycle. They are assembled in special places called microtubule organizing centers. In plant cells, they have a weakly expressed amorphous structure.
Functions of microtubules: participate in the formation of the cell membrane; direct bubbles of dictyosomes to the forming membrane, like spindle threads that form in a dividing cell; play a role in the formation of the cell plate (the initial boundary between daughter cells). In addition, microtubules important component flagella and cilia, in the movement of which they play an important role.
Microfilaments, like microtubules, are found in almost all eukaryotic cells. They are long filaments 5–7 nm thick, consisting of the contractile protein actin. Bundles of microfilaments are found in many cells of higher plants. Apparently, they play an important role in the currents of the cytoplasm. Microfilaments, together with microtubules, form a flexible network called the cytoskeleton.
The ground substance has long been considered a homogeneous (homogeneous) protein-rich solution with few or no structures at all. However, at present, using a high-voltage electron microscope, it has been established that the ground substance is a three-dimensional lattice built from thin (3–6 nm in diameter) strands that fill the entire cell. Other components of the cytoplasm, including microtubules and microfilaments, are suspended from this microtrabecular lattice.
The microtrabecular structure is a lattice of protein strands, the space between which is filled with water. Together with water, the lattice has the consistency of a gel; the gel looks like gelatinous bodies.
Organelles are attached to the microtrabecular lattice. The lattice provides a link between separate parts cells and directs intracellular transport.
Lipid droplets are spherical structures that give granularity to the cytoplasm plant cell under a light microscope. On electron micrographs, they look amorphous. Very similar but smaller drops are found in plastids.
Lipid drops, mistaking them for organelles, called them spherosomes and believed that they were surrounded by a single or double layer membrane. However, recent data show that lipid droplets do not have membranes, but may be coated with protein.
Ergastic substances are the “passive products” of the protoplast: reserve substances or waste products. They may appear and disappear in different periods cell cycle. Except starch grains, crystals, anthocyanin pigments and lipid droplets. These include resins, gums, tannins and proteins. Ergastic substances are part of the cell membrane, the main substance of the cytoplasm and organelles, including vacuoles.
Flagella and cilia are thin, hair-like structures that extend from the surface of many eukaryotic cells. They have a constant diameter, but the length varies from 2 to 150 microns. Conventionally, the longer and few of them are called flagella, and the shorter and more numerous are called cilia. There are no clear differences between these two types of structures, so the term flagellum is used to refer to both.
In some algae and fungi, flagella are locomotor organs, with the help of which they move in water. In plants (for example, mosses, liverworts, ferns, some gymnosperms), only germ cells (gametes) have flagella.
Each flagellum has a specific organization. An outer ring of 9 pairs of microtubules surrounds two additional microtubules located in the center of the flagellum. Enzyme-containing "handles" extend from one microtubule of each of the outer pairs. This is the basic 9+2 organization pattern found in all flagella in eukaryotic organisms. It is believed that the movement of flagella is based on the sliding of microtubules, while the outer pairs of microtubules move one along the other without contraction. Sliding of pairs of microtubules relative to each other causes local bending of the flagellum.
Flagella "grow" from cytoplasmic cylindrical structures called basal bodies, which also form the basal part of the flagellum. The basal bodies have internal structure, resembling the structure of a flagellum, except that the outer tubules are collected in triplets, and not in pairs, and the central tubules are absent.
Cell wall. The cell wall delimits the size of the protoplast and prevents its rupture due to the absorption of water by the vacuole.
The cell wall has specific functions that are important not only for the cell and the tissue in which the cell is located, but also for the whole plant. Cell walls play an essential role in the absorption, transport and excretion of substances, and, in addition, lysosomal, or digestive activity, can be concentrated in them.
Cell wall components. The most typical component of the cell wall is cellulose, which largely determines its architecture. Cellulose molecules are made up of repeating glucose molecules connected end to end. Long thin cellulose molecules are combined into microfibrils with a thickness of 10 - 25 nm. Microfibrils intertwine and form thin threads, which in turn can be wrapped one around the other, like strands in a rope. Each such "rope", or macrofibril, has a thickness of about 0.5 microns, reaching a length of 4 microns. Macrofibrils are as strong as steel wire of equal size.
The cellulose frame of the cell wall is filled with cellulose matrix molecules intertwined with it. It consists of polysaccharides called hemicelluloses, and pectins, or pectins, chemically very close to hemicelluloses.
Another component of the cell wall, lignin, is the most abundant polymer in plant cells after cellulose. Lignin increases the rigidity of the wall and is usually found in cells that perform a supporting or mechanical function.
Cutin, suberin, wax - usually deposited in the shells of the protective tissues of plants. Cutin, for example, is found in the cell membranes of the epidermis, and suberin is found in the secondary protective tissue, the cork. Both substances are found in combination with waxes and prevent excessive water loss from the plant.
Layers of the cell wall. The wall thickness of plant cells varies widely, depending on the role of cells in plant structure and the age of the cell itself. Under electron microscope two layers are visible in the plant cell wall: the median lamina (also called the intercellular substance), and the primary cell wall. Many cells lay down another layer - the secondary cell wall. The median plate is located between the primary walls of adjacent cells. The secondary wall, if any, is deposited by the cell protoplast on the inner surface of the primary cell wall.
Middle plate. The median lamina consists mainly of pectin. Where a cell wall should arise, between two newly formed cells, a dense plexus of tubules of the endoplasmic reticulum and the cistern of the Golgi apparatus (dictyosomes) is first noted. Then bubbles appear in this place, filled with pectin (from polysaccharides). These bubbles are separated from the tanks of the Golgi apparatus. The early cell wall contains various polysaccharides, the main ones being pectins and hemicellulose. Later, it consists of denser substances - cellulose and lignin.
Primary cell wall. This is a layer of cellulose sheath that is deposited before or during cell growth. In addition to cellulose, hemicellulose and pectin, the primary membranes contain a glycoprotein. Primary shells can lignify. The pectin component imparts plasticity, which allows the primary shell to expand as the root, stem or leaf elongates.
Actively dividing cells (most mature cells involved in the processes of photosynthesis, respiration and secretion) have primary membranes. Such cells with a primary membrane and a living protoplast are capable of losing characteristic shape, divide and differentiate into a new cell type. They are involved in wound healing and tissue regeneration in plants.
Primary cell membranes are not uniform in thickness throughout their entire length, but have thin sections, which are called primary pore fields. The strands of cytoplasm, or plasmodesmata, that connect the protoplasts of neighboring cells usually pass through the primary pore fields.
secondary cell wall. Despite the fact that many plant cells have only a primary wall, in some, the protoplast deposits a secondary wall towards the center of the cell. This usually occurs after the cessation of cell growth and the area of the primary membrane no longer increases. For this reason, the secondary shell differs from the primary. Secondary membranes are especially needed by specialized cells that strengthen the plant and conduct water. After the deposition of the secondary membrane, the protoplast of these cells, as a rule, dies. There is more cellulose in the secondary shells than in the primary ones, and pectin substances and glycoproteins are absent in them. The secondary membrane is difficult to stretch, its matrix consists of hemicellulose.
In the secondary shell, three layers can be distinguished - outer, middle and inner (S 1, S 2, S 3). The layered structure of the secondary shells significantly increases their strength. Cellulose microfibrils in the secondary membrane are deposited denser than in the primary. Lignin - ordinary component secondary shells of wood.
The pores in the membranes of contacting cells are located opposite each other. Two opposing pores and a pore membrane form a pair of pores. In cells with secondary membranes, there are two main types of pores: simple and bordered. In bordered pores, the secondary wall hangs over the pore cavity. In simple pores, this is not the case.
Cell wall growth. As the cell grows, the thickness and area of the cell membrane increases. Shell stretching is a complex process. It is controlled by the protoplast and regulated by the hormone auxin.
In cells growing uniformly in all directions, the deposition of myofibrils is random. These myofibrils form an irregular network. Such cells were found in the core of the stem, storage tissues, and during cell cultivation in vitro. In elongating cells, the myofibrils of the lateral sheaths are deposited at right angles to the axis of elongation.
Matrix substances - pectins, hemicelluloses and glycoproteins are transferred to the membrane in vesicles of dictyosomes. At the same time, pectins are more characteristic of growing cells, while hemicelluloses predominate in non-growing cells.
Cellulose microfibrils are synthesized on the cell surface with the help of an enzyme complex associated with the plasma membrane. The orientation of microfibrils is controlled by microtubules located at inner surface plasma membrane.
Plasmodesma. These are thin threads of the cytoplasm that connect the protoplasts of neighboring cells. Plasmodesmata either pass through the cell wall anywhere, or are concentrated in primary pore fields or in membranes between pairs of pores. Under an electron microscope, plasmodesmata appear as narrow channels lined with the plasma membrane. Along the axis of the canal, a smaller cylindrical tube, the desmotubule, extends from one cell to another, which communicates with the endoplasmic reticulum of both adjacent cells. Many plasmodesmata form during cell division, when the tubular endoplasmic reticulum is taken up by the developing cell plate. Plasmodesmata can also form in the membranes of non-dividing cells. These structures provide efficient transfer of certain substances from cell to cell.
Cell division. At multicellular organisms cell division along with an increase in their size is a way of growth of the whole organism. New cells formed during division are similar in structure and function, both to the parent cell and to each other. The division process in eukaryotes can be divided into two partially overlapping stages: mitosis and cytokinesis.
Mitosis is the formation of two daughter nuclei from one nucleus, morphologically and genetically equivalent to each other. Cytokinesis is the division of the cytoplasmic part of the cell with the formation of daughter cells.
Cell cycle. A living cell goes through a series of successive events that make up the cell cycle. The duration of the cycle itself varies depending on the type of cell and external factors, for example, on temperature or security nutrients. Usually the cycle is divided into interphase and four phases of mitosis.
Interphase. The period between successive mitotic divisions.
The interphase is divided into three periods, denoted as G 1 , S, G 2 .
During the G 1 period, which begins after mitosis. During this period, the amount of cytoplasm increases, including various organelles. In addition, according to the modern hypothesis, during the G 1 period, substances are synthesized that either stimulate or inhibit the S period and the rest of the cycle, thus determining the division process.
The S period follows the G 1 period, at which time the genetic material (DNA) is duplicated.
During the G2 period, which follows S, the structures directly involved in mitosis, such as spindle components, are formed.
Some cells go through an unlimited number of cell cycles. This unicellular organisms and some cells of active growth zones (meristems). Some specialized cells after maturation lose their ability to reproduce. The third group of cells, for example, those forming wound tissue (callus), retains the ability to divide only under special conditions.
Mitosis, or nuclear fission. It is a continuous process divided into four phases: prophase, metaphase, anaphase, telophase. As a result of mitosis, the genetic material that has doubled in interphase is divided equally between the two daughter nuclei.
One of the most early signs the transition of a cell to division is the appearance of a narrow, ring-shaped belt of microtubules directly under the plasma membrane. This relatively dense belt surrounds the nucleus in the equatorial plane of the future mitotic spindle. Since it appears before prophase, it is called the preprophase girdle. It disappears after the mitotic spindle, long before the appearance in the late telophase of the cell plate, which grows from the center to the periphery and merges with the mother cell membrane in the area previously occupied by the preprophase belt.
Prophase. At the beginning of prophase, the chromosomes resemble long threads scattered inside the nucleus. Then, as the strands shorten and thicken, one can see that each chromosome is made up of not one, but two intertwined strands called chromatids. In late prophase, the two shortened paired chromatids of each chromosome lie side by side in parallel, connected by a narrow section called the centromere. It has a specific position on each chromosome and divides the chromosome into two arms of different lengths.
Microtubules are located parallel to the surface of the nucleus along the axis of the spindle. This is the earliest manifestation of mitotic spindle assembly.
By the end of prophase, the nucleolus gradually loses its clear outline and finally disappears. Shortly thereafter, the nuclear envelope also disintegrates.
Metaphase. At the beginning of metaphase, the spindle, which is a three-dimensional structure, widest in the middle and tapering towards the poles, takes the place previously occupied by the nucleus. The spindle fibers are bundles of microtubules. During metaphase, the chromosomes, each consisting of two chromatids, are arranged so that their centromeres lie in the equatorial plane of the spindle. With its centromere, each chromosome is attached to the spindle threads. However, some strands pass from one pole to the other without being attached to chromosomes.
When all chromosomes are located in the equatorial plane, metaphase is completed. Chromosomes are ready to divide.
Anaphase. The chromatids of each chromosome separate. Now these are daughter chromosomes. First of all, the centromere divides, and the two daughter chromosomes are drawn to opposite poles. In this case, the centromeres move forward, and the arms of the chromosomes stretch behind. The spindle filaments attached to the chromosomes shorten, contributing to the separation of chromatids and the movement of daughter chromosomes in opposite directions.
Telophase. In telophase, the separation of two identical groups of chromosomes is completed, while a nuclear membrane forms around each of them. The rough reticulum takes an active part in this. The spindle apparatus disappears. During telophase, the chromosomes lose their clarity of outline, stretch out, turning back into thin threads. The nucleoli are restored. When the chromosomes become invisible, mitosis is completed. The two daughter nuclei enter interphase. They are genetically equivalent to each other and to the parent nucleus. This is very important, since the genetic program, and with it all the signs, must be transferred to the daughter organisms.
The duration of mitosis varies various organisms And it depends on the type of fabric. However, prophase is the longest and anaphase is the shortest. In the cells of the root tip, the duration of prophase is 1–2 hours; metaphase - 5 - 15 min; anaphase - 2 - 10 min; telophase - 10 - 30 min. The duration of the interphase is from 12 to 30 hours.
In many eukaryotic cells, the microtubule organizing centers responsible for the formation of the mitotic spindle are associated with centrioles.
Cytokinesis. This is the process of division of the cytoplasm. In most organisms, cells divide by retracting the cell wall and forming a fission furrow, which gradually deepens, compressing the remaining filaments of the mitotic spindle. In all plants (bryophytes and vascular plants) and in some algae, cells divide due to the formation of a cell plate.
In the early telophase, a barrel-shaped system of fibers called the phragmoplast forms between the two daughter nuclei. The phragmoplast fibers, like the fibers of the mitotic spindle, consist of microtubules. Small drops appear in the equatorial plane of the phragmoplast. They fuse to form a cell plate that grows until it reaches the wall of the dividing cell. This completes the separation of the two daughter cells. Merging droplets are bubbles breaking away from the Golgi apparatus. Basically, they contain pectin substances, from which the middle plate is formed. Vesicle membranes are involved in the construction of the plasma membrane on both sides of the plate. At the same time, plasmodesmata are formed from fragments of the tubular endoplasmic reticulum.
After the formation of the median plate, each protoplast deposits a primary membrane on it. In addition, each daughter cell deposits a new layer of membrane around the entire protoplast, which continues the membrane that arose from the cell plate. The original shell of the parent cell is destroyed as the daughter cells grow.
Different types of eukaryotic mitosis
The cell division described above in plants, and also in animals, is not the only form of indirect cell division. The simplest type of mitosis is pleuromitosis. It resembles binary division prokaryotic cells, in which the nucleoids after replication remain associated with the plasma membrane. The membrane begins to grow between the DNA binding points and thereby spreads the chromosomes into different areas cells. After that, during the formation of a cell constriction, each of the DNA molecules will be in a new separate cell.
Characteristic for the division of eukaryotic cells is the formation of a spindle built of microtubules. With closed pleuromitosis (it is called closed because the divergence of chromosomes occurs without breaking the nuclear envelope), not centrioles, but other structures located on inside nuclear membrane. These are the so-called polar bodies of indeterminate morphology, from which microtubules extend. There are two of these bodies. They diverge from each other without losing contact with nuclear envelope. As a result of this, two half-spindles are formed associated with chromosomes. The entire process of mitotic spindle formation and chromosome segregation in this case occurs under the nuclear envelope. This type of mitosis occurs among protozoa and is widely distributed in fungi (chytridia, zygomycetes, yeasts, oomycetes, ascomycetes, myxomycetes, etc.). there are forms of semi-closed pleuromitosis, when the nuclear envelope is destroyed at the poles of the formed spindle.
The next form of mitosis is orthomitosis. In this case, the COMTs are located in the cytoplasm; from the very beginning, not half-spindles are formed, but a bipolar spindle. There are three forms of orthomitosis (normal mitosis), semi-closed and closed. In semi-closed orthomitosis, a bisymmetric spindle is formed with the help of TsOMT located in the cytoplasm, the nuclear envelope is preserved throughout mitosis, with the exception of the polar zones. Masses of granular material, or even centrioles, may be detected as COMT. This form of mitosis is found in zoospores of green, brown, red algae, in some lower mushrooms and gregarine. With closed orthomitosis, the nuclear membrane is completely preserved, in which a real spindle is formed. Microtubules are formed in the karyoplasm, less often they grow from the internal TsOMT, which is not associated (unlike pleuromitosis) with the nuclear membrane. This type of mitosis is typical for the division of ciliate micronuclei, but can also occur in protozoa. In open orthomitosis, the nuclear envelope completely disintegrates. This type of cell division is characteristic of animal organisms, some protozoa, and cells of higher plants. This form of mitosis, in turn, is represented by astral and anastral types.
From the brief review, it is clear that main feature Mitosis in general is the emergence of structures of the fission spindle, which is formed in connection with the TsOMT, which is diverse in structure.
Morphology of the mitotic figure
The mitotic apparatus is especially well expressed at the metaphase stage of mitosis. In metaphase, in the equatorial plane of the cell, chromosomes are located, from which the so-called spindle threads extend in opposite directions, converging at two different poles of the mitotic figure. Thus, the mitotic spindle is a collection of chromosomes, poles and fibers. The spindle fibers are single microtubules or their bundles. Microtubules start from the spindle poles, and some of them go to the centromeres, where the chromosome kinetochores (kinetochore microtubules) are located, some go further towards the opposite pole, but do not reach it. They are called "interpolar microtubules". A group of radial microtubules departs from the poles, forming around them a structure resembling a "radiant glow" - these are astral microtubules.
By morphology, mitotic figures are divided into astral and anastral types.
The astral spindle type, or convergent, is characterized by the fact that its poles are represented by a small zone to which microtubules converge (converge). Usually, centrosomes containing centrioles are located at the poles of the astral spindles. However, cases of centriolar astral mitoses are known (during meiosis of some invertebrates). In addition, radial microtubules radiating from the poles are noted, which are not part of the spindle, but form stellate zones - citasters. This type of mitotic division resembles a dumbbell. The anastrial type of the mitotic figure does not have citasters at the poles. The polar regions of the spindle here are wide, they are called polar caps, they include centrioles. In this case, the spindle fibers do not depart from one point, but diverge in a wide front (diverge) from the entire zone of the polar caps. This type of spindle is characteristic of dividing cells of higher plants, but can also be found in higher animals. In the early embryogenesis of mammals, centriolar (divergent) mitoses are observed during division of oocyte maturation and at divisions I and II of the zygote. But already in the third cell division and in all subsequent cells, they divide with the participation of astral spindles, in the poles of which centrioles are always found. In general, for all forms of mitosis, chromosomes with their kinetophores, polar bodies (centrosomes), and spindle fibers remain common structures.
Centromeres and kinetochores
Centromeres may have different localization along the length of the chromosomes. Holocentric centromeres occur when microtubules are linked along the length of the entire chromosome (some insects, nematodes, some plants). Monocentric centromeres - when microtubules are associated with chromosomes in one site. monocentric centromeres can be punctate (for example, in some budding yeasts), when only one microtubule approaches the kinetochore, and zonal, where a bundle of microtubules approaches the complex kinetochore. Despite the diversity of centromere zones, they are all associated with complex structure kinetochore, which has a fundamental similarity in structure and function in all eukaryotes. Kinetochores are specialized protein structures for the most part located in the centromeres of chromosomes. These are complex complexes consisting of many proteins. morphologically, they are very similar, have the same structure, ranging from diatoms to humans. They are three-layer structures: the inner dense layer adjacent to the body of the chromosome, the middle loose layer and the outer dense layer. Many fibrils extend from the outer layer, forming the so-called fibrous crown of the kinetochore. IN general form kinetochores have the form of plates or disks lying in the zone of the primary constriction of the chromosome, in the centromere. There is usually one kinetochore per chromosome or chromatid. Before anaphase, kinetochores on each sister chromatid are located, each communicating with its own bundle of microtubules. In plants, the kinetochore does not look like plates, but hemispheres. The functional role of kinetochores is to bind sister chromatids to each other, to fix mitotic microtubules, to regulate chromosome separation, and to actually move chromosomes during mitosis with the participation of microtubules. In general, protein structures, kinetochores, double in the S-period, parallel to chromosome doubling. But their proteins are present on the chromosomes at all periods of the cell cycle.
PLANT TISSUE
Educational fabrics(meristems)
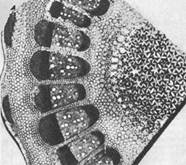
Educational tissues in the body of plants are located in different places, so they are divided into the following groups (Fig. 0; 1).
1. Apical (apical) meristems are located on the tops, or apexes, of the axial organs - stem, root. With the help of these meristems, the vegetative organs of plants carry out their growth in length.
2. Lateral meristems are characteristic of axial organs. There they are located concentrically, in the form of a clutch.
3. Intercalary, or intercalary, meristems originate from the apical meristems. These are groups of cells that are not yet able to multiply, but have embarked on the path of differentiation. There are no initial cells among them, but there are many specialized ones.
4. Wound meristems provide restoration of the damaged part of the body. Regeneration begins with dedifferentiation, that is, the reverse development from specialized cells to meristematic ones. They turn into phellogen, which forms a plug that covers the surface of the wound. Dedifferentiated cells, dividing, can form a loose parenchymal tissue - callus. From it, under certain conditions, plant organs are formed.
Integumentary tissues
They act as a boundary barrier, separating underlying tissues from environment. The primary integuments of a plant consist only of living cells. Secondary and tertiary integument - mostly from the dead with thick cell walls.
The main functions of integumentary tissues:
protection of the plant from drying out;
・Ingress protection harmful microorganisms;
· defence from sunburn;
· defence from mechanical damage;
regulation of metabolism between the plant and the environment;
perception of irritation.
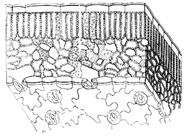
The primary integumentary tissue is the epidermis, the epidermis. Consists of living cells. Formed from apical meristems. Covers young growing stems and leaves.
The epidermis was formed in plants in connection with the exit from the aquatic habitat to land in order to prevent it from drying out. In addition to stomata, all cells of the epidermis are tightly interconnected. The outer walls of the main cells are thicker than the rest. The entire surface is covered with a layer of cutin and vegetable waxes. This layer is called the cuticle (skin). It is absent on the growing roots and underwater parts of plants. When dry, the permeability of the cuticle is significantly weakened.
In addition to the main cells, there are others in the epidermis, in particular hairs, or trichomes. They are unicellular and multicellular (Fig. 2). Functionally, they increase the surface of the epidermis, for example, in the root growth zone, serve as mechanical protection, cling to a support, and reduce water loss. A number of plants have glandular hairs, such as nettles.
Only higher plants have stomata in the epidermis, which regulate the exchange of water and gases. If there is no cuticle, then there is no need for stomata. Stomata are a group of cells that form the stomatal apparatus, which consists of two trailing cells and adjoining epidermal cells - side cells. They differ from the main epidermal cells (Figure 3). Guard cells differ from their surrounding cells in shape and the presence of a large number of chloroplasts and unevenly thickened walls. Those that face each other are thicker than the rest (Fig. 4). Between the guard cells, a stomatal gap is formed, which leads to a substomatal space called the substomatal cavity. Guard cells are highly photosynthetic. They contain a large number of spare starch and numerous mitochondria.
The number and distribution of stomata, types of stomatal apparatus varies widely in different plants. Stomata are absent in modern bryophytes. Photosynthesis in them is carried out by the gametophyte generation, and sporophytes are not capable of independent existence.
Usually stomata are located on the underside of the leaf. In plants floating on the water surface - on the upper surface. In the leaves of cereals, the stomata are often evenly spaced on both sides. Such leaves are illuminated relatively evenly. On 1 mm 2 of the surface, from 100 to 700 stomata can be located.
Secondary integumentary tissue (periderm). This tissue replaces the epidermis when green color annual shoots are replaced by brown. It is multilayered and consists of a central layer of cambial cells - phellogen. Phellogen cells, dividing, lay a layer of phellem outward, and phelloderm inward (Fig. 5).
Phelema, or cork. At first it consists of living thin-walled cells. Over time, their walls are impregnated with suberin and vegetable waxes and die off. The contents of the cell are filled with air.
Fellema functions:
Prevents moisture loss
protects the plant from mechanical damage;
Protects against pathogens
provides thermal insulation, as the cells are filled with air.
Cells of the phellogen, located in the epidermis itself, the underlying subepidermal layer, less often in the deep layers of the primary cortex, are the generating basis of the primary cortex.
The cork layer is not permanent. Breaks occur in it, which communicate with the intercellular spaces located nearby. At the same time, small tubercles are formed on the surface - lenticels, which communicate the spaces of the intercellular spaces with atmospheric air (Fig. 6.7).
In autumn, the phellogen under the lenticels lays a layer of corked cells, which greatly reduce transpiration, but do not completely exclude it. In the spring, this layer collapses from the inside. On light birch bark, lentils are clearly visible in the form of dark dashes.
The tertiary integumentary tissue (crust) is also characteristic only for woody forms of plants.
Phellogen is repeatedly laid down in the deeper layers of the cortex. The tissues that are outside of it eventually die off, forming a crust. Its cells are dead and incapable of stretching. However, living cells located deeper divide, which leads to an increase in the transverse size of the stem. Over time, the outer layer of the crust breaks. The time of occurrence of such a gap is a fairly constant value for individual plants. In an apple tree, this happens in the seventh year of life, in a hornbeam - in the fiftieth. In some species it does not occur at all. The main function of the peel is protection against mechanical and thermal damage.
Parenchyma
It is a group of specialized tissues that fill the spaces inside the plant body between conductive and mechanical tissues (Fig. 8). More often parenchyma cells have a rounded shape, less often an elongated shape. The presence of developed intercellular spaces is characteristic. The spaces between cells together form a transport system - the apoplast. In addition, the intercellular spaces form the "ventilation system" of the plant. Through stomata, or lenticels, they are connected with atmospheric air and provide optimal gas composition inside the plant. Developed intercellular spaces are especially necessary for plants growing on waterlogged soil, where normal gas exchange is difficult. This parenchyma is called aerenchyma (Fig. 9).
Elements of the parenchyma, filling the gaps between other tissues, also perform the function of a support. Parenchyma cells are alive, they do not have thick cell walls, like sclerenchyma. Therefore, mechanical properties are provided by turgor. If the water content drops, which leads to plasmolysis and wilting of the plant.
The assimilation parenchyma is formed by thin-walled cells with many intercellular spaces. The cells of this structure contain many chloroplasts, so it is called chlorenchyma. Chloroplasts are located along the wall without shading each other. In the assimilation parenchyma, photosynthesis reactions occur, which provide the plant with organic substances and energy. The result of photosynthetic processes is the possibility of the existence of all living organisms on the Earth.
Assimilation tissues are present only in the illuminated parts of the plant; they are separated from the environment by a transparent epidermis. If the epidermis is replaced by opaque secondary integumentary tissues, the assimilation parenchyma disappears.
The storage parenchyma serves as a receptacle for organic substances that are temporarily not used by the plant organism. Basically postponing organic matter as various kinds inclusions are capable of any cell with a living protoplast, but some cells specialize in this (Fig. 10, 11). Energy-rich compounds are deposited only during the growing season, consumed during the dormant period and in preparation for the next growing season. Therefore, reserve substances are deposited in the vegetative organs only in perennial plants.
The receptacle of stocks can be ordinary organs (escape, root), as well as specialized ones (rhizomes, tubers, bulbs). All seed plants store energetically valuable substances in seeds (cotyledons, endosperm). Many plants of an arid climate store not only organic matter, but also water (Fig. 12). For example, aloe stores water in fleshy leaves, cacti in shoots.
mechanical fabrics
The mechanical properties of plant cells are provided by:
The hard shell of the cell
turgidity, that is, the turgor state of cells.
Despite the fact that almost all tissue cells have mechanical properties, however, there are tissues in the plant for which mechanical properties are basic. These are collenchyma and sclerenchyma. They usually function in interaction with other tissues. Inside the body, the plants form a kind of frame. Therefore, they are called reinforcement.
Not all plants are equally well expressed mechanical fabrics. To a much lesser extent, plants living in aquatic environment than terrestrial ones. The reason is that aquatic plants need less internal support. Their body is largely supported by the surrounding water. Air on land does not create such support, since it has a lower density than water. It is for this reason that the presence of specialized mechanical tissues becomes relevant.
Improvement of internal support structures occurred in the process of evolution.
Collenchyma. It is formed only by living cells, elongated along the axis of the organ. This type of mechanical tissue is formed very early, during the period of primary growth. Therefore, it is important that the cells remain alive and retain the ability to stretch along with the stretching cells that are nearby.
Features of collenchyma cells:
Uneven thickening of the shell, as a result of which some parts of it remain thin, while others thicken;
shells do not lignify.
Collenchyma cells are located differently relative to each other. In adjacent cells, thickenings form on the corners facing each other (Fig. 13). Such a collenchyma is called a corner collenchyma. In another case, the cells are arranged in parallel layers. The cell membranes facing these layers are strongly thickened. This is lamellar collenchyma. Cells can be located loosely, with abundant intercellular spaces - this is a loose collenchyma. Such collenchyma is often found in plants on waterlogged soils.
Collenchyma is of particular importance in young plants, herbaceous forms, and also in plant parts where secondary growth does not occur, such as leaves. In this case, it is laid very close to the surface, sometimes just under the epidermis. If the organ has faces, then thick layers of collenchyma are found along their crests.
Collenchyma cells are functional only in the presence of turgor. Water deficiency reduces the effectiveness of collenchyma and the plant temporarily withers, for example, cucumber leaves sagging on a hot day. After filling the cells with water, the functions of collenchyma are restored.
Sclerenchyma. The second type of mechanical tissues. Unlike collenchyma, where all cells are alive, sclerenchyma cells are dead. Their walls are very thick. They perform a mechanical function. A strong thickening of the membrane leads to disruption of the transport of substances, as a result of which the protoplast dies (Fig. 14). The lignification of the cell membranes of sclerenchyma occurs when the plant organ has already completed its growth. Therefore, they no longer interfere with the stretching of surrounding tissues.
Depending on the shape, two types of sclerenchyma cells are distinguished - fibers and sclereids.
The fibers have a strongly elongated shape with very thick walls and a small cavity. They are somewhat smaller than wood fibers. Often under the epidermis form longitudinal layers and strands. In the phloem or xylem, they can be found singly or in groups. In the phloem they are called bast fibers (Fig. 15), and in the xylem they are called libriform fibers (Fig. 16).
Sclereids, or stony cells, are represented by rounded or branched cells with powerful membranes. In the body of a plant, they can be found singly (supporting cells) (Fig. 17) or in groups (Fig. 18). It should be noted that the mechanical properties strongly depend on the location of the sclereids. Part of the sclereids form continuous layers, as, for example, in the shell of nuts or in the stones of fruits (stone fruits).
excretory tissues
Any living cell initially has the function of removing substances, while transport can go both along the concentration gradient and against the gradient. Substances that have been synthesized in the cell can be removed and will subsequently affect the activity of other cells (phytohormones, enzymes). This process is called secretion. In the case when substances that are waste products of the cell are excreted, the process is called excretion. Despite the fact that the excretion of substances in plant and animal cells has a fundamental similarity, plants have a number of features that arise from fundamental differences in vital activity. The level of metabolic reactions in plants is much lower than in animals. Therefore, proportionately smaller amounts of waste are emitted. Another feature is that, independently synthesizing almost all the necessary organic compounds, plants never form excessive reserves. The released substances can serve as the starting material for other reactions (for example, CO 2 and H 2 O).
If in animals the process of excretion of toxins is associated with their removal from the body, plants may not do this, isolating unnecessary substances in the living protoplast (removal of various substances into the vacuole), in dead cells (most tissues perennial plant consists of such cells), in intercellular spaces.
The released substances can be divided into two large groups. The first group is organic substances synthesized directly by the cell (enzymes, polysaccharides, lignins, terpenes, the latter are constituent elements essential oils and resin). The second group - substances that initially enter the cell from the outside with the help of xylem or phloem transport (water, mineral salts, amino acids, monosugars, etc.). The excreted secret is rarely homogeneous and usually consists of a mixture in which one substance has the highest concentration.
Plants, unlike animals, lack a complete excretory system. There are only specialized structures scattered throughout the plant - idioblasts. The cells that form them are smaller than the parenchyma cells lying nearby. They have an electron-dense cytoplasm, with developed elements of the endoplasmic reticulum and the Golgi complex. Most often not expressed central vacuole. Between themselves and other living cells, these cells are connected by numerous plasmodesmata.
Depending on the location, excretory tissues can be external and internal.
External excretory tissues. Glandular hairs are derivatives of the epidermis. Morphologically, they are variable - they can have a multicellular head, be elongated, in the form of a shield on a leg, etc. (fig.19). These include stinging nettle hairs.
Hydathodes are structures that release excess water under conditions of low transpiration and high humidity - guttation. These structures are formed by groups of colorless living cells with thin walls - the epitheme. This tissue is adjacent to the conductive bundle (Fig. 20). Water is released through special water stomata, which differ from ordinary stomata by immobility and a constantly open gap. The composition of the guttation fluid varies widely from almost clean water to a very complex mixture of substances.
Nectaries. Many plants are characterized by the release of a liquid containing from 7 to 87% mono- and oligosaccharides. This process is carried out by special structures - nectaries. Depending on the location, flower, or floral, nectaries are distinguished (Fig. 21), as well as plants located on stems and leaves - extrafloral, or extrafloral nectaries. Nectar can be unaltered phloem juice, which is delivered to the surface through the intercellular spaces and excreted through the stomata. More complex nectars are formed by glandular parenchyma, covered with epidermis with glandular hairs. Nectar is excreted either by epidermal cells or glandular hairs. In this case, the secreted nectar differs from the phloem sap. Since it is dominated by glucose, fructose, sucrose, and in phloem juice - glucose. Ions are present in low concentrations. Ions are present in low concentrations. To attract pollinators, nectar may contain steroid hormones which are essential for insects.
Digestive glands are present in insectivorous plants (sundew, nepenthys, etc.). In response to the touch of small invertebrates, they secrete a secret containing hydrolytic enzymes, formic, butyric and propionic acids. As a result, biopolymers are broken down into monomers, which are used by the plant for its needs.
Salt glands develop in plants growing on saline soils. These glands are located on the leaves and, in a dissolved form, excess ions are brought to the surface. Salt is first deposited on the cuticle, then it is washed off by rain (Fig. 22). In addition, many plants have salt hairs on their leaves. Each hair consists of two cells: one forms the head, the other - the stem. Salts accumulate in the vacuole of the head cell, and when their concentration becomes high, the head falls off. In its place, a new cell grows, which also accumulates salts. During the growth of the leaf, the head may fall off and form several times. The advantage of salt hairs is that by releasing salts, they lose little water. This phenomenon is very important for plants growing on saline soils, where fresh water is scarce.
Internal excretory tissues. Internal excretory structures, scattered throughout the body of the plant in the form of idioblasts, do not remove substances outside the body, accumulating them in themselves. Idioblasts are plant cells that differ in shape, structure, or content from other cells in the same tissue. Depending on the origin, schizogenic (Greek schiso - split) and lysogenic (Greek lysis - dissolution) receptacles of secreted substances are distinguished (Fig. 23).
Schizogenic receptacles. These are more or less extensive intercellular spaces filled with excreted substances. For example, resin passages of conifers, aralia, umbrella and others. Resin is believed to have bactericidal properties, repels herbivores, making plants inedible for them.
Lysigenic receptacles are formed in place of living cells, which die and are destroyed after the accumulation of substances in them. Such formations can be observed in the peel of citrus fruits.
In addition to the main types, there are intermediate forms. Milkers are of two types: articulated and non-articulated. The former are formed from many living cells arranged in chains. At the point of contact, the shells are destroyed, the protoplasts merge, and as a result, a single network is formed. Such milky ones are found in Compositae. Non-segmented lactic cells are formed by a giant multinucleated cell. It arises in the early stages of development, lengthens and branches as the plant grows, forming a complex network, for example, in euphorbia plants (Fig. 25).
The cell walls of lactifers are heavily watered. They do not lignify and therefore are plastic. The central part of the lactis is occupied by a vacuole with latex, and the protoplast occupies a near-wall position. The boundary between the protoplast and the vacuole cannot always be determined. The lactifers penetrate the meristem and grow together with the plant organ.
Conductive tissues
This type refers to complex tissues, consists of differently differentiated cells. In addition to the actual conductive elements, the tissue contains mechanical, excretory and storage elements (Fig. 26). Conductive tissues unite all plant organs into a single system. There are two types of conductive tissues: xylem and phloem (Greek xylon - tree; phloios - bark, bast). They have both structural and functional differences.
The conducting elements of the xylem are formed by dead cells. They carry long-distance transport of water and substances dissolved in it from the root to the leaves. The conductive elements of the phloem keep the protoplast alive. They carry long-distance transport from photosynthetic leaves to the root.
Usually xylem and phloem are located in the plant body in certain order, forming layers or conductive bundles. Depending on the structure, several types of conductive bundles are distinguished, which are characteristic of certain groups plants. In the collateral open bundle between the xylem and the phloem, there is a cambium, which provides secondary growth (Fig. 27-A, 28). In the bicollateral open bundle, the phloem is located on both sides of the xylem (Fig. 27-B, 29). Closed bundles do not contain cambium, and hence are not capable of secondary thickening (Fig. 27-B, 27-D, 30.31). Two more types of concentric bundles can be found, where either the phloem surrounds the xylem (Fig. 27-E, 32), or the xylem surrounds the phloem (Fig. 27-E).
Xylem (wood). The development of xylem in higher plants is associated with the provision of water exchange. Since water is constantly excreted through the epidermis, the same amount of moisture must be absorbed by the plant and added to the organs that carry out transpiration. It should be taken into account that the presence of a living protoplast in water-conducting cells would greatly slow down transport, and dead cells are more functional here. However, a dead cell does not have turgidity, so the membrane must have mechanical properties. Note: turgescence is the state of plant cells, tissues and organs, in which they become elastic due to the pressure of the contents of the cells on their elastic membranes. Indeed, the conducting elements of the xylem consist of dead cells stretched along the axis of the organ with thick lignified shells.
Initially, xylem is formed from the primary meristem - procambium, located at the tops of the axial organs. The protoxylem differentiates first, followed by the metaxylem. Three types of xylem formation are known. In the exarch type, elements of the protoxylem first appear on the periphery of the procambial bundle, then elements of the metaxylem appear in the center. If the process goes in the opposite direction (i.e. from the center to the periphery), then this is an endarchic type. In the mesarchic type, xylem is laid in the center of the procambial bundle, after which it is deposited both towards the center and towards the periphery.
The root is characterized by an exarch type of xylem formation, while the stems have an endarch type. In low organized plants, the methods of xylem formation are very diverse and can serve as systematic characteristics.
In some plants (for example, monocots), all procambial cells differentiate into conductive tissues that are not capable of secondary thickening. In other forms (for example, woody), lateral meristems (cambium) remain between xylem and phloem. These cells are able to divide, renewing the xylem and phloem. This process is called secondary growth. Many plants growing in relatively stable climatic conditions grow constantly. In forms adapted to seasonal climate changes - periodically. As a result, well-defined annual growth rings are formed.
The main stages of differentiation of procambial cells. Its cells have thin membranes that do not prevent them from stretching during the growth of the organ. The protoplast then begins to lay down the secondary membrane. But this process has pronounced features. The secondary membrane is not deposited in a continuous layer, which would not allow the cell to stretch, but in the form of rings or in a spiral. Cell elongation is not difficult. In young cells, the rings or coils of the helix are located close to each other. In mature cells, they diverge as a result of cell stretching (Fig. 33). Ringed and spiral thickenings of the shell do not prevent growth, however, mechanically they are inferior to shells, where the secondary thickening forms a continuous layer. Therefore, after the cessation of growth in the xylem, elements with a continuous lignified shell (metaxylem) are formed. It should be noted that the secondary thickening here is not annular or spiral, but punctate, stair-like, reticulate (Fig. 34). Its cells are not able to stretch, and die within a few hours. This process in nearby cells occurs in a coordinated manner. A large number of lysosomes appear in the cytoplasm. Then they decompose, and the enzymes in them destroy the protoplast. When the transverse walls are destroyed, the cells located in a chain one above the other form a hollow vessel (Fig. 35). Majority angiosperms and some ferns have vessels.
A conducting cell that does not form through perforations in its wall is called a tracheid. The movement of water through the tracheids is slower than through the vessels. The fact is that in tracheids the primary membrane is not interrupted anywhere. The tracheids communicate with each other through pores. It should be clarified that in plants, the pore is only a depression in the secondary membrane up to the primary membrane, and there are no through perforations between the tracheids.
The most common are bordered pores (Fig. 35-1). Their channel, facing the cavity of the cell, forms an extension - the pore chamber. The pores of most coniferous plants on the primary shell have a thickening - a torus, which is a kind of valve and is able to regulate the intensity of water transport. By shifting, the torus blocks the flow of water through the pore, but after that it can no longer return to its previous position, performing a one-time action.
The pores are more or less rounded, elongated perpendicular to the elongated axis (a group of these pores resembles a ladder, so this porosity is called ladder porosity). Transport through the pores is carried out both in the longitudinal and transverse directions. Pores are present not only in tracheids, but also in individual vascular cells that form a vessel.
From the point of view of evolutionary theory, tracheids are the first and main structure that conducts water in the body of higher plants. It is believed that the vessels originated from tracheids due to the lysis of the transverse walls between them (Fig. 36). Most ferns and gymnosperms do not have. They move water through tracheids.
In the course of evolutionary development, vessels arose in different groups plants repeatedly, but they acquired the most important functional significance in angiosperms, which have them along with tracheids. It is believed that the possession of a more advanced mechanism of transport helped them not only to survive, but also to achieve a significant variety of forms.
Xylem is a complex tissue, in addition to water-conducting elements, it contains others. Mechanical functions are performed by libriform fibers (lat. liber - bast, forma - form). The presence of additional mechanical structures is important because, despite the thickening, the walls of the water-carrying elements are still too thin. They are unable to sustain themselves a large mass perennial plant. The fibers developed from tracheids. They are characterized by smaller sizes, lignified (lignified) shells and narrow cavities. On the wall, one can find pores devoid of edging. These fibers cannot conduct water, their main function is support.
The xylem also contains living cells. Their mass can reach 25% of the total volume of wood. Since these cells have a rounded shape, they are called wood parenchyma. In the body of the plant, the parenchyma is located in two ways. In the first case, the cells are arranged in the form of vertical cords - this is the cord parenchyma. In another case, the parenchyma forms horizontal rays. They are called core rays, as they connect the core and the cortex. The core performs a number of functions, including the storage of substances.
Phloem (bast). This is a complex tissue, as it is formed by heterogeneous cells. The main conducting cells are called sieve elements (Fig. 37). The conducting elements of the xylem are formed by dead cells, while in the phloem they retain a living, albeit greatly altered, protoplast during the period of functioning. The phloem carries out the outflow of plastic substances from the photosynthetic organs. All living cells of plants possess the ability to conduct organic substances. And hence, if xylem can be found only in higher plants, then the transport of organic substances between cells is also carried out in lower plants.
Xylem and phloem develop from apical meristems. At the first stage, protophloem is formed in the procambial cord. As the surrounding tissues grow, it stretches, and when growth is completed, metaphloem is formed instead of protophloem.
At various groups In higher plants, two types of sieve elements can be found. In ferns and gymnosperms, it is represented by sieve cells. The sieve fields in the cells are scattered along the side walls. The protoplast retains a slightly destructed nucleus.
In angiosperms, sieve elements are called sieve tubes. They communicate with each other through sieve plates. Mature cells lack nuclei. However, a companion cell is located next to the sieve tube, which is formed together with the sieve tube as a result of mitotic division of the common mother cell (Fig. 38). The companion cell has a denser cytoplasm with a large number of active mitochondria, as well as a fully functioning nucleus, great amount plasmodesmata (ten times more than other cells). Companion cells affect the functional activity of non-nuclear sieve tube cells.
The structure of mature sieve cells has some peculiarities. There is no vacuole, so the cytoplasm is highly liquefied. The nucleus may be absent (in angiosperms) or be in a wrinkled, functionally inactive state. Ribosomes and the Golgi complex are also absent, but the endoplasmic reticulum is well developed, which not only penetrates the cytoplasm, but also passes into neighboring cells through the pores of the sieve fields. Well-developed mitochondria and plastids are found in abundance.
Between cells, the transport of substances goes through holes located on the cell membranes. Such openings are called pores, but unlike the tracheid pores, they are through. It is assumed that they are highly expanded plasmodesmata, on the walls of which the callose polysaccharide is deposited. The pores are arranged in groups, forming sieve fields. In primitive forms, sieve fields are randomly scattered over the entire surface of the shell, in more advanced angiosperms they are located at the ends of neighboring cells adjacent to each other, forming a sieve plate (Fig. 39). If there is one sieve field on it, it is called simple, if there are several - complex.
The speed of movement of solutions through the sieve elements is up to 150 cm ∕ hour. This is a thousand times the rate of free diffusion. Probably, active transport takes place, and numerous mitochondria of sieve elements and satellite cells supply the necessary ATP for this.
The life of the sieve elements of the phloem depends on the presence of lateral meristems. If they are present, then the sieve elements work throughout the life of the plant.
In addition to sieve elements and satellite cells, the phloem contains bast fibers, sclereids, and parenchyma.
Questions for control and repetition
Signs of higher plants? What generation of higher plants is called a gametophyte? Sporophyte?
What are the reasons that contribute to the development of differentiated tissues in terrestrial plants.
List the types of plants.
What groups of meristems do you know? Where are they located in the plant body?
Integumentary tissues, their functions, significance.
Organization various types covering tissues.
The structure of the stomatal apparatus. List the types of stomatal apparatus known to you. How do stomata function?
Features of parenchymal tissues, types of parenchyma.
Signs of parenchyma and sclerenchyma.
List the types of excretory structures of a plant. What is their significance?
Methods for the formation of receptacles for excreted substances.
What are milkers? What types are they divided into?
How are conductive tissues organized?
What are conductive bundles? Their types.
Directions of movement of substances through the xylem? Phloem? Does the composition of transported substances differ in xylem and phloem?
The structure and functions of xylem, phloem.
The organization of the conducting elements of the phloem in plants from various systematic groups.
How are substances transported in the phloem?
Additional proteins associated with them, the so-called MAP proteins. These proteins, by stabilizing microtubules, accelerate the process of tubulin polymerization. Structure and function of microfilaments Microfilaments are very thin and long filamentous protein structures found throughout the cytoplasm. Under the plasma membrane, microfilaments form a continuous plexus, forming ...

As, for example, dill, carrots, turnips, etc., in most cases it has the same kind of tissue that I just indicated in the cork. Thus, the plant cell was discovered for the first time. But Hooke could not deal with his microscope for a long time: ideas of other inventions swarm in his head (spring clocks, improved compasses, etc.), and he willingly handed over the further maintenance of microscopic ...
At least some idea of the appropriate methods of cell research. Light microscopy The most ancient and, at the same time, the most common method of studying cells is microscopy. We can say that the beginning of the study of the cell was laid by the invention of the light optical microscope. Unarmed human eye has a resolution of about 1/10 mm. This...
With many cell organelles, it provides a connection between individual intracellular structures. Lysosomes Lysosomes (from lis- and Greek. soma - body), structures in the cells of animal and plant organisms containing enzymes that can break down (ie lyse - hence the name) proteins, polysaccharides, peptides, nucleic acids. This is a very variegated class of bubbles with a size of 0.1-0.4 microns, ...
The structure of a plant cell. Plant tissues
STRUCTURE OF THE PLANT CELL
A plant cell consists of a more or less rigid cell wall and a protoplast. The cell wall is the cell wall and the cytoplasmic membrane. The term protoplast comes from the word protoplasm, which has long been used to refer to all living things. The protoplast is the protoplasm of an individual cell.
The protoplast consists of the cytoplasm and the nucleus. The cytoplasm contains organelles (ribosomes, microtubules, plastids, mitochondria) and membrane systems (endoplasmic reticulum, dictyosomes). The cytoplasm also includes the cytoplasmic matrix (basic substance) in which organelles and membrane systems are immersed. The cytoplasm is separated from the cell wall by the plasma membrane, which is an elementary membrane. Unlike most animal cells, plant cells contain one or more vacuoles. These are vesicles filled with liquid and surrounded by an elementary membrane (tonoplast).
In a living plant cell, the ground substance is in constant motion. Organelles are involved in the movement, called the flow of the cytoplasm or cyclosis. Cyclosis facilitates the movement of substances in the cell and their exchange between the cell and the environment.
Plasma membrane. It is a bilayer phospholipid structure. Plant cells are characterized by invaginations of the plasma membrane.
The plasma membrane performs the following functions:
Participates in the exchange of substances between the cell and the environment;
Coordinates the synthesis and assembly of cellulose microfibrils of the cell wall;
Transmits hormonal and external signals that control cell growth and differentiation.
Core. It is the most prominent structure in the cytoplasm of a eukaryotic cell. The kernel performs two important functions:
Controls the vital activity of the cell, determining which proteins, and at what time should be synthesized;
Stores genetic information and passes it on to daughter cells during cell division.
The nucleus of a eukaryotic cell is surrounded by two elementary membranes that form the nuclear envelope. It is permeated with numerous pores with a diameter of 30 to 100 nm, visible only in an electron microscope. The pores have a complex structure. The outer membrane of the nuclear membrane in some places is combined with the endoplasmic reticulum. The nuclear envelope can be viewed as a specialized, locally differentiated part of the endoplasmic reticulum (ER).
In the nucleus stained with special dyes, thin filaments and clumps of chromatin and nucleoplasm (the main substance of the nucleus) can be distinguished. Chromatin is made up of DNA bound to special proteins called histones. In the process of cell division, chromatin becomes more and more compacted and assembled into chromosomes. DNA encodes genetic information.
Organisms differ in the number of chromosomes in somatic cells. For example, cabbage has - 20 chromosomes; sunflower - 34; wheat - 42; humans - 46, and one of the species of the fern Ophioglossum - 1250. Sex cells (gametes) have only half the number of chromosomes characteristic of somatic cells of the body. The number of chromosomes in gametes is called haploid (single), in somatic cells - diploid (double). Cells that have more than two sets of chromosomes are called polyploid.
Under a light microscope, spherical structures - nucleoli - can be seen. Each nucleus has one or more nucleoli, which are visible in non-dividing nuclei. Ribosomal RNA is synthesized in the nucleolus. Typically, the nuclei of diploid organisms have two nucleoli, one for each haploid set of chromosomes. The nucleoli do not have their own membrane. Biochemically, the nucleoli are characterized by a high concentration of RNA, which is associated here with phosphoproteins. The size of the nucleoli depends on the functional state of the cell. it has been noticed that in a rapidly growing cell, in which intensive processes of protein synthesis are taking place, the nucleoli increase in size. In the nucleoli, mRNA and ribosomes are produced, which perform a synthetic function only in the nucleus.
Nucleoplasm (karyoplasm) is represented by a homogeneous liquid in which various proteins, including enzymes, are dissolved.
Plastids. Vacuoles, cellulose cell walls, and plastids are characteristic components of plant cells. Each plastid has its own shell, consisting of two elementary membranes. Inside the plastid, a membrane system and a varying degree of a homogeneous substance, the stroma, are distinguished. Mature plastids are classified based on the pigments they contain.
Chloroplasts, in which photosynthesis takes place, contain chlorophylls and carotenoids. Usually have the form of a disk with a diameter of 4 - 5 microns. In one cell of the mesophyll (middle of the leaf) there can be 40 - 50 chloroplasts; in mm2 of a leaf - about 500,000. In the cytoplasm, chloroplasts are usually located parallel to the cell membrane.
The internal structure of the chloroplast is complex. The stroma is permeated with a developed system of membranes in the form of vesicles - thylakoids. Each thylakoid consists of two membranes. Thylakoids form a single system. As a rule, they are collected in piles - grains, resembling columns of coins. The thylakoids of individual granae are interconnected by stroma thylakoids, or intergranal thylakoids. Chlorophyll and carotenoids are embedded in thylakoid membranes. Chloroplasts in green plants and algae often contain grains of starch and small lipid (fat) droplets. Starch grains are temporary stores of photosynthesis products. They can disappear from the chloroplasts of a plant that has been in the dark for only 24 hours, and reappear 3–4 hours after the plant is transferred to light.
Chloroplasts are semi-autonomous organelles and resemble bacteria. For example, the ribosomes of bacteria and chloroplasts have a fairly high similarity. They are smaller than eukaryotic ribosomes. Protein synthesis on the ribosomes of bacteria and chloroplasts is inhibited by chloramphenicol, which has no effect in eukaryotic cells. In addition, both bacteria and chloroplasts have a similar type of nucleoid, organized in a similar way. Despite the fact that the formation of chloroplasts and the synthesis of pigments contained in them is largely controlled by the chromosomal DNA of the cell, nevertheless, in the absence of its own DNA, chloroplasts do not form.
Chloroplasts can be considered the main cellular organelles, since they are the first in the chain of transformation of solar energy, as a result of which humanity receives both food and fuel. In chloroplasts, not only photosynthesis takes place. They are also involved in the synthesis of amino acids and fatty acids, serve as a storage of temporary reserves of starch.
Chromoplasts are pigmented plastids. Diverse in form, they do not have chlorophyll, but synthesize and accumulate carotenoids, which give yellow, orange, red color to flowers, old leaves, fruits and roots. chromoplasts can develop from chloroplasts, which in this case lose chlorophyll and internal membrane structures, accumulate carotenoids. This occurs during the ripening of many fruits. Chromoplasts attract insects and other animals with which they co-evolved.
Leukoplasts are non-pigmented plastids. Some of them synthesize starch (amyloplasts), others are capable of forming various substances, including lipids and proteins. In the light, leukoplasts turn into chloroplasts.
Proplastids are small, colorless or pale green, undifferentiated plastids found in the meristematic (dividing) cells of roots and shoots. They are the precursors of other, more differentiated plastids - chloroplasts, chromoplasts, and aminoplasts. If the development of protoplastids is delayed due to lack of light, one or more prolamellar bodies, which are semi-crystalline accumulations of tubular membranes, may appear in them. Plastids containing prolamellar bodies are called etioplasts. In the light, etioplasts turn into chloroplasts, while the membranes of the prolamellar bodies form thylakoids. Etioplasts are formed in the leaves of plants that are in the dark. protoplasts of seed embryos first turn into etioplasts, from which chloroplasts then develop in the light. Plastids are characterized by relatively easy transitions from one type to another. Plastids, like bacteria, reproduce by dividing in two. In meristematic cells, the time of division of protoplastids approximately coincides with the time of cell division. However, in mature cells, most of the plastids are formed as a result of the division of mature plastids.
Mitochondria. Like chloroplasts, mitochondria are surrounded by two elementary membranes. The inner membrane forms many folds and protrusions - cristae, which significantly increase the inner surface of the mitochondria. They are significantly smaller than plastids, about 0.5 µm in diameter, and variable in length and shape.
In mitochondria, the process of respiration takes place, as a result of which organic molecules are broken down with the release of energy and its transfer to ATP molecules, the main energy reserve of all eukaryotic cells. Most plant cells contain hundreds or thousands of mitochondria. Their number in one cell is determined by the cell's need for ATP. Mitochondria are in constant motion, moving from one part of the cell to another, merging with each other and dividing. Mitochondria usually assemble where energy is needed. If the plasma membrane actively transfers substances from cell to cell, then mitochondria are located along the surface of the membrane. In motile unicellular algae, mitochondria accumulate at the base of the flagella, supplying the energy necessary for their movement.
Mitochondria, like plastids, are semi-autonomous organelles containing the components necessary for the synthesis of their own proteins. The inner membrane surrounds a fluid matrix that contains proteins, RNA, DNA, bacterial-like ribosomes, and various solutes. DNA exists in the form of circular molecules located in one or more nucleoids.
Based on the similarity of bacteria with the mitochondria and chloroplasts of eukaryotic cells, it can be assumed that mitochondria and chloroplasts originated from bacteria that found "refuge" in larger heterotrophic cells - the precursors of eukaryotes.
Microbodies. Unlike plastids and mitochondria, which are delimited by two membranes, microbodies are spherical organelles surrounded by a single membrane. Microbodies have a granular (granular) content, sometimes they also contain crystalline protein inclusions. Microbodies are associated with one or two regions of the endoplasmic reticulum.
Some microbodies, called proxisomes, play an important role in the metabolism of glycolic acid, which is directly related to photorespiration. In green leaves, they are associated with mitochondria and chloroplasts. Other microbodies, called glyoxisomes, contain the enzymes needed to convert fats into carbohydrates. This happens in many seeds during germination.
Vacuoles - These are areas of the cell delimited by the membrane, filled with liquid - cell sap. They are surrounded by a tonoplast (vacuolar membrane).
A young plant cell contains numerous small vacuoles, which merge into one large vacuole as the cell ages. In a mature cell, up to 90% of its volume can be occupied by a vacuole. In this case, the cytoplasm is pressed in the form of a thin peripheral layer to the cell membrane. The increase in cell size is mainly due to the growth of the vacuole. As a result, turgor pressure arises and tissue elasticity is maintained. This is one of the main functions of the vacuole and tonoplast.
The main component of the juice is water, the rest vary depending on the type of plant and its physiological state. Vacuoles contain salts, sugars, less often proteins. The tonoplast plays an active role in the transport and accumulation of certain ions in the vacuole. The concentration of ions in the cell sap can significantly exceed its concentration in the environment. With a high content of certain substances, crystals form in the vacuoles. The most common are calcium oxalate crystals, which have a different shape.
Vacuoles are places of accumulation of metabolic products (metabolism). These can be proteins, acids, and even substances toxic to humans (alkaloids). Pigments are often deposited. Blue, violet, purple, dark red, crimson give plant cells pigments from the anthocyanin group. Unlike other pigments, they dissolve well in water and are found in cell sap. They determine the red and blue color of many vegetables (radishes, turnips, cabbage), fruits (grapes, plums, cherries), flowers (cornflowers, geraniums, delphiniums, roses, peonies). Sometimes these pigments mask chlorophyll in the leaves, for example, in the decorative red maple. Anthocyanins color autumn leaves bright red. They are formed in cold sunny weather, when the synthesis of chlorophyll stops in the leaves. In leaves, when anthocyanins are not formed, after the destruction of chlorophyll, yellow-orange carotenoids of chloroplasts become noticeable. The leaves are most brightly colored in cold, clear autumn.
Vacuoles are involved in the destruction of macromolecules, in the circulation of their components in the cell. Ribosomes, mitochondria, plastids, getting into vacuoles, are destroyed. According to this digestive activity, they can be compared with lysosomes - organelles of animal cells.
Vacuoles are formed from the endoplasmic reticulum (reticulum)
Ribosomes. Small particles (17 - 23nm), consisting of approximately equal amounts of protein and RNA. In ribosomes, amino acids combine to form proteins. There are more of them in cells with an active metabolism. Ribosomes are located in the cytoplasm of the cell freely or attached to the endoplasmic reticulum (80S). They are also found in the nucleus (80S), mitochondria (70S), plastids (70S).
Ribosomes can form a complex on which there is a simultaneous synthesis of identical polypeptides, information about which is taken from one molecule and RNA. Such a complex is called polyribosomes (polysomes). Cells that synthesize proteins in large quantities have an extensive system of polysomes, which are often attached to the outer surface of the nuclear envelope.
Endoplasmic reticulum. It is a complex three-dimensional membrane system of indefinite extent. In the context of the ER looks like two elementary membranes with a narrow transparent space between them. The shape and length of the ER depend on the cell type, its metabolic activity, and the stage of differentiation. In cells that secrete or store proteins, the ER is shaped like flat sacs or cisterns, with numerous ribosomes attached to its outer surface. This reticulum is called the rough endoplasmic reticulum. A smooth ER is usually tubular in shape. Rough and smooth endoplasmic reticulum can be present in the same cell. As a rule, there are numerous connections between them.
The endoplasmic reticulum functions as the communication system of the cell. It is associated with the outer shell of the kernel. In fact, these two structures form a single membrane system. When the nuclear envelope ruptures during cell division, its fragments resemble ER fragments. The endoplasmic reticulum is a system for transporting substances: proteins, lipids, carbohydrates, to different parts of the cell. endoplasmic reticulum of neighboring cells are connected through cytoplasmic strands - plasmodesmata - that pass through the cell membranes.
The endoplasmic reticulum is the main site for the synthesis of cell membranes. In some plant cells, membranes of vacuoles and microbodies, cisterns of dictyosomes are formed here.
Golgi apparatus. This term is used to refer to all dictyosomes, or Golgi bodies, in a cell. Dictyosomes are groups of flat, disc-shaped vesicles, or cisterns, that branch along the edges into a complex system of tubules. Dictyosomes in higher plants consist of 4 - 8 cisterns, collected together.
Usually, in a pack of tanks, a developing and maturing side is distinguished. the membranes of the forming cisterns resemble the membranes of the ER in structure, and the membranes of the maturing cisterns resemble the plasma membrane.
Dictyosomes are involved in secretion and, in most higher plants, in the formation of cell membranes. Cell wall polysaccharides synthesized by dictyosomes accumulate in vesicles, which then separate from maturing cisterns. These secretory vesicles migrate and fuse with the CMP; while the polysaccharides contained in them are built into the cell membrane. Some substances that accumulate in dictyosomes are formed in other structures, for example, in the ER, and then transported to dictyosomes, where they are modified (modified) before secretion. For example, glycoproteins are an important building material of the cell membrane. The protein part is synthesized by polysomes of the rough ER, the carbohydrate part is synthesized in dictyosomes, where both parts combine to form glycoproteins.
Membranes are dynamic, mobile structures that constantly change their shape and area. The concept of the endoplasmic system is based on the mobility of membranes. According to this concept, the inner membranes of the cytoplasm, except for the membranes of mitochondria and plastids, are a single whole and originate from the endoplasmic reticulum. New cisterns of dictyosomes form from the endoplasmic reticulum through the intermediate vesicle stage, and the secretory vesicles that separate from the dictyosomes eventually contribute to the formation of the plasma membrane. Thus, the endoplasmic reticulum and dictyosomes form a functional unit in which dictyosomes play the role of intermediate structures in the process of converting membranes like the endoplasmic reticulum into membranes like the plasma. In tissues whose cells grow and divide weakly, membrane components are constantly being updated.
microtubules found in almost all eukaryotic cells. They are cylindrical structures with a diameter of about 24 nm. Their length varies. Each tubule is made up of subunits of a protein called tubulin. The subunits form 13 longitudinal filaments surrounding the central cavity. Microtubules are dynamic structures, they are regularly destroyed and formed at certain stages of the cell cycle. Their assembly occurs in special places, which are called microtubule organization centers. In plant cells, they have a weakly expressed amorphous structure.
Functions of microtubules: participate in the formation of the cell membrane; direct bubbles of dictyosomes to the forming membrane, like spindle threads that form in a dividing cell; play a role in the formation of the cell plate (the initial boundary between daughter cells). In addition, microtubules are an important component of flagella and cilia, in the movement of which they play an important role.
microfilaments, like microtubules, they are found in almost all eukaryotic cells. They are long filaments 5–7 nm thick, consisting of the contractile protein actin. Bundles of microfilaments are found in many cells of higher plants. Apparently, they play an important role in the currents of the cytoplasm. Microfilaments, together with microtubules, form a flexible network called the cytoskeleton.
Base substance for a long time was considered a homogeneous (homogeneous) protein-rich solution with a small number of structures or even structureless. However, at present, using a high-voltage electron microscope, it has been established that the ground substance is a three-dimensional lattice built from thin (3–6 nm in diameter) strands that fill the entire cell. Other components of the cytoplasm, including microtubules and microfilaments, are suspended from this microtrabecular lattice.
The microtrabecular structure is a lattice of protein strands, the space between which is filled with water. Together with water, the lattice has the consistency of a gel; the gel looks like gelatinous bodies.
Organelles are attached to the microtrabecular lattice. The lattice provides communication between individual parts of the cell and directs intracellular transport.
Lipid drops- structures of a spherical shape, giving granularity to the cytoplasm of a plant cell under a light microscope. On electron micrographs, they look amorphous. Very similar but smaller drops are found in plastids.
Lipid drops, mistaking them for organelles, called them spherosomes and believed that they were surrounded by a single or double layer membrane. However, recent data show that lipid droplets do not have membranes, but may be coated with protein.
Ergastic substances - these are the “passive products” of the protoplast: reserve substances or waste products. They can appear and disappear at different periods of the cell cycle. Except starch grains, crystals, anthocyanin pigments and lipid droplets. These include resins, gums, tannins and proteins. Ergastic substances are part of the cell membrane, the main substance of the cytoplasm and organelles, including vacuoles.
Flagella and cilia - they are thin, hair-like structures that extend from the surface of many eukaryotic cells. They have a constant diameter, but the length varies from 2 to 150 microns. Conventionally, the longer and few of them are called flagella, and the shorter and more numerous are called cilia. There are no clear differences between these two types of structures, so the term flagellum is used to refer to both.
In some algae and fungi, flagella are locomotor organs, with the help of which they move in water. In plants (for example, mosses, liverworts, ferns, some gymnosperms), only germ cells (gametes) have flagella.
Each flagellum has a specific organization. An outer ring of 9 pairs of microtubules surrounds two additional microtubules located in the center of the flagellum. Enzyme-containing "handles" extend from one microtubule of each of the outer pairs. This is the basic 9+2 organization pattern found in all flagella in eukaryotic organisms. It is believed that the movement of flagella is based on the sliding of microtubules, while the outer pairs of microtubules move one along the other without contraction. Sliding of pairs of microtubules relative to each other causes local bending of the flagellum.
Flagella "grow" from cytoplasmic cylindrical structures called basal bodies, which also form the basal part of the flagellum. The basal bodies have an internal structure resembling that of a flagellum, except that the outer tubules are arranged in triplets rather than pairs, and the central tubules are absent.
Cell wall. The cell wall delimits the size of the protoplast and prevents its rupture due to the absorption of water by the vacuole.
The cell wall has specific functions that are important not only for the cell and the tissue in which the cell is located, but also for the whole plant. Cell walls play an essential role in the absorption, transport and excretion of substances, and, in addition, lysosomal, or digestive activity, can be concentrated in them.
Cell wall components. The most typical component of the cell wall is cellulose, which largely determines its architecture. Cellulose molecules are made up of repeating glucose molecules connected end to end. Long thin cellulose molecules are combined into microfibrils with a thickness of 10 - 25 nm. Microfibrils intertwine and form thin threads, which in turn can be wrapped one around the other, like strands in a rope. Each such "rope", or macrofibril, has a thickness of about 0.5 microns, reaching a length of 4 microns. Macrofibrils are as strong as steel wire of equal size.
The cellulose frame of the cell wall is filled with cellulose matrix molecules intertwined with it. It consists of polysaccharides called hemicelluloses, and pectins, or pectins, chemically very close to hemicelluloses.
Another component of the cell wall, lignin, is the most abundant polymer in plant cells after cellulose. Lignin increases the rigidity of the wall and is usually found in cells that perform a supporting or mechanical function.
Cutin, suberin, wax - usually deposited in the shells of the protective tissues of plants. Cutin, for example, is found in the cell membranes of the epidermis, and suberin is found in the secondary protective tissue, the cork. Both substances are found in combination with waxes and prevent excessive water loss from the plant.
Layers of the cell wall. The wall thickness of plant cells varies widely, depending on the role of cells in plant structure and the age of the cell itself. Under an electron microscope, two layers are visible in the plant cell wall: the median lamina (also called the intercellular substance), and the primary cell wall. Many cells lay down another layer, the secondary cell wall. The median plate is located between the primary walls of adjacent cells. The secondary wall, if any, is deposited by the cell protoplast on the inner surface of the primary cell wall.
Middle plate. The median lamina consists mainly of pectin. Where a cell wall should arise, between two newly formed cells, a dense plexus of tubules of the endoplasmic reticulum and the cistern of the Golgi apparatus (dictyosomes) is first noted. Then bubbles appear in this place, filled with pectin (from polysaccharides). These bubbles are separated from the tanks of the Golgi apparatus. The early cell wall contains various polysaccharides, the main ones being pectins and hemicellulose. Later, it consists of denser substances - cellulose and lignin.
Primary cell wall. This is a layer of cellulose sheath that is deposited before or during cell growth. In addition to cellulose, hemicellulose and pectin, the primary membranes contain a glycoprotein. Primary shells can lignify. The pectin component imparts plasticity, which allows the primary shell to expand as the root, stem or leaf elongates.
Actively dividing cells (most mature cells involved in the processes of photosynthesis, respiration and secretion) have primary membranes. Such cells with a primary membrane and a living protoplast are able to lose their characteristic shape, divide and differentiate into a new cell type. They are involved in wound healing and tissue regeneration in plants.
Primary cell membranes are not uniform in thickness throughout their entire length, but have thin sections, which are called primary pore fields. The strands of cytoplasm, or plasmodesmata, that connect the protoplasts of neighboring cells usually pass through the primary pore fields.
secondary cell wall. Despite the fact that many plant cells have only a primary wall, in some, the protoplast deposits a secondary wall towards the center of the cell. This usually occurs after the cessation of cell growth and the area of the primary membrane no longer increases. For this reason, the secondary shell differs from the primary. Secondary membranes are especially needed by specialized cells that strengthen the plant and conduct water. After the deposition of the secondary membrane, the protoplast of these cells, as a rule, dies. There is more cellulose in the secondary shells than in the primary ones, and pectin substances and glycoproteins are absent in them. The secondary membrane is difficult to stretch, its matrix consists of hemicellulose.
In the secondary shell, three layers can be distinguished - outer, middle and inner (S1, S2, S3). The layered structure of the secondary shells significantly increases their strength. Cellulose microfibrils in the secondary membrane are deposited denser than in the primary. Lignin is a common component of the secondary skins of wood.
The pores in the membranes of contacting cells are located opposite each other. Two opposing pores and a pore membrane form a pair of pores. In cells with secondary membranes, there are two main types of pores: simple and bordered. In bordered pores, the secondary wall hangs over the pore cavity. In simple pores, this is not the case.
Cell wall growth. As the cell grows, the thickness and area of the cell membrane increases. Shell stretching is a complex process. It is controlled by the protoplast and regulated by the hormone auxin.
In cells growing uniformly in all directions, the deposition of myofibrils is random. These myofibrils form an irregular network. Such cells were found in the core of the stem, storage tissues, and during cell cultivation in vitro. In elongating cells, the myofibrils of the lateral sheaths are deposited at right angles to the axis of elongation.
Matrix substances - pectins, hemicelluloses and glycoproteins are transferred to the membrane in vesicles of dictyosomes. At the same time, pectins are more characteristic of growing cells, while hemicelluloses predominate in non-growing cells.
Cellulose microfibrils are synthesized on the cell surface with the help of an enzyme complex associated with the plasma membrane. The orientation of microfibrils is controlled by microtubules located near the inner surface of the plasma membrane.
Plasmodesma. These are thin threads of the cytoplasm that connect the protoplasts of neighboring cells. Plasmodesmata either pass through the cell wall anywhere, or are concentrated in primary pore fields or in membranes between pairs of pores. Under an electron microscope, plasmodesmata appear as narrow channels lined with the plasma membrane. Along the axis of the canal, a smaller cylindrical tube, the desmotubule, extends from one cell to another, which communicates with the endoplasmic reticulum of both adjacent cells. Many plasmodesmata form during cell division, when the tubular endoplasmic reticulum is taken up by the developing cell plate. Plasmodesmata can also form in the membranes of non-dividing cells. These structures provide efficient transfer of certain substances from cell to cell.
Cell division. In multicellular organisms, cell division, along with an increase in their size, is a way of growth of the whole organism. New cells formed during division are similar in structure and function, both to the parent cell and to each other. The division process in eukaryotes can be divided into two partially overlapping stages: mitosis and cytokinesis.
Mitosis is the formation of two daughter nuclei from one nucleus, morphologically and genetically equivalent to each other. Cytokinesis is the division of the cytoplasmic part of the cell with the formation of daughter cells.
Cell cycle. A living cell goes through a series of successive events that make up the cell cycle. The duration of the cycle itself varies depending on the type of cell and external factors such as temperature or nutrient availability. Usually the cycle is divided into interphase and four phases of mitosis.
Interphase. The period between successive mitotic divisions.
The interphase is divided into three periods, denoted as G1, S, G2.
During the G1 period, which begins after mitosis. During this period, the amount of cytoplasm increases, including various organelles. In addition, according to the current hypothesis, substances are synthesized during the G1 period that either stimulate or inhibit the S period and the rest of the cycle, thus determining the division process.
The S period follows the G1 period, at which time the genetic material (DNA) is duplicated.
During the G2 period, which follows S, structures that are directly involved in mitosis, such as spindle components, are formed.
Some cells go through an unlimited number of cell cycles. These are unicellular organisms and some cells of active growth zones (meristems). Some specialized cells after maturation lose their ability to reproduce. The third group of cells, for example, those forming wound tissue (callus), retains the ability to divide only under special conditions.
Mitosis, or nuclear fission. It is a continuous process divided into four phases: prophase, metaphase, anaphase, telophase. As a result of mitosis, the genetic material that has doubled in interphase is divided equally between the two daughter nuclei.
One of the earliest signs of a cell's transition to division is the appearance of a narrow, annular band of microtubules just below the plasma membrane. This relatively dense belt surrounds the nucleus in the equatorial plane of the future mitotic spindle. Since it appears before prophase, it is called the preprophase girdle. It disappears after the mitotic spindle, long before the appearance in the late telophase of the cell plate, which grows from the center to the periphery and merges with the mother cell membrane in the area previously occupied by the preprophase belt.
Prophase. At the beginning of prophase, the chromosomes resemble long threads scattered inside the nucleus. Then, as the strands shorten and thicken, one can see that each chromosome is made up of not one, but two intertwined strands called chromatids. In late prophase, the two shortened paired chromatids of each chromosome lie side by side in parallel, connected by a narrow section called the centromere. It has a specific position on each chromosome and divides the chromosome into two arms of different lengths.
Microtubules are located parallel to the surface of the nucleus along the axis of the spindle. This is the earliest manifestation of mitotic spindle assembly.
By the end of prophase, the nucleolus gradually loses its clear outline and finally disappears. Shortly thereafter, the nuclear envelope also disintegrates.
Metaphase. At the beginning of metaphase, the spindle, which is a three-dimensional structure, widest in the middle and tapering towards the poles, takes the place previously occupied by the nucleus. The spindle fibers are bundles of microtubules. During metaphase, the chromosomes, each consisting of two chromatids, are arranged so that their centromeres lie in the equatorial plane of the spindle. With its centromere, each chromosome is attached to the spindle threads. However, some strands pass from one pole to the other without being attached to chromosomes.
When all chromosomes are located in the equatorial plane, metaphase is completed. Chromosomes are ready to divide.
Anaphase. The chromatids of each chromosome separate. Now these are daughter chromosomes. First of all, the centromere divides, and the two daughter chromosomes are drawn to opposite poles. In this case, the centromeres move forward, and the arms of the chromosomes stretch behind. The spindle filaments attached to the chromosomes shorten, contributing to the separation of chromatids and the movement of daughter chromosomes in opposite directions.
STRUCTURE OF THE PLANT CELL
A plant cell consists of a more or less rigid cell wall and a protoplast. The cell wall is the cell wall and the cytoplasmic membrane. The term protoplast comes from the word protoplasm, which has long been used to refer to all living things. The protoplast is the protoplasm of an individual cell.
The protoplast consists of the cytoplasm and the nucleus. The cytoplasm contains organelles (ribosomes, microtubules, plastids, mitochondria) and membrane systems (endoplasmic reticulum, dictyosomes). The cytoplasm also includes the cytoplasmic matrix (basic substance) in which organelles and membrane systems are immersed. The cytoplasm is separated from the cell wall by the plasma membrane, which is an elementary membrane. Unlike most animal cells, plant cells contain one or more vacuoles. These are vesicles filled with liquid and surrounded by an elementary membrane (tonoplast).
In a living plant cell, the ground substance is in constant motion. Organelles are involved in the movement, called the flow of the cytoplasm or cyclosis. Cyclosis facilitates the movement of substances in the cell and their exchange between the cell and the environment.
Plasma membrane. It is a bilayer phospholipid structure. Plant cells are characterized by invaginations of the plasma membrane.
The plasma membrane performs the following functions:
Participates in the exchange of substances between the cell and the environment;
Coordinates the synthesis and assembly of cellulose microfibrils of the cell wall;
Transmits hormonal and external signals that control cell growth and differentiation.
Core. It is the most prominent structure in the cytoplasm of a eukaryotic cell. The kernel performs two important functions:
Controls the vital activity of the cell, determining which proteins, and at what time should be synthesized;
Stores genetic information and passes it on to daughter cells during cell division.
The nucleus of a eukaryotic cell is surrounded by two elementary membranes that form the nuclear envelope. It is permeated with numerous pores with a diameter of 30 to 100 nm, visible only in an electron microscope. The pores have a complex structure. The outer membrane of the nuclear membrane in some places is combined with the endoplasmic reticulum. The nuclear envelope can be viewed as a specialized, locally differentiated part of the endoplasmic reticulum (ER).
In the nucleus stained with special dyes, thin filaments and clumps of chromatin and nucleoplasm (the main substance of the nucleus) can be distinguished. Chromatin is made up of DNA bound to special proteins called histones. In the process of cell division, chromatin becomes more and more compacted and assembled into chromosomes. DNA encodes genetic information.
Organisms differ in the number of chromosomes in somatic cells. For example, cabbage has - 20 chromosomes; sunflower - 34; wheat - 42; humans - 46, and one of the species of the fern Ophioglossum - 1250. Sex cells (gametes) have only half the number of chromosomes characteristic of somatic cells of the body. The number of chromosomes in gametes is called haploid (single), in somatic cells - diploid (double). Cells that have more than two sets of chromosomes are called polyploid.
Under a light microscope, spherical structures - nucleoli - can be seen. Each nucleus has one or more nucleoli, which are visible in non-dividing nuclei. Ribosomal RNA is synthesized in the nucleolus. Typically, the nuclei of diploid organisms have two nucleoli, one for each haploid set of chromosomes. The nucleoli do not have their own membrane. Biochemically, the nucleoli are characterized by a high concentration of RNA, which is associated here with phosphoproteins. The size of the nucleoli depends on the functional state of the cell. it has been noticed that in a rapidly growing cell, in which intensive processes of protein synthesis are taking place, the nucleoli increase in size. In the nucleoli, mRNA and ribosomes are produced, which perform a synthetic function only in the nucleus.
Nucleoplasm (karyoplasm) is represented by a homogeneous liquid in which various proteins, including enzymes, are dissolved.
Plastids. Vacuoles, cellulose cell walls, and plastids are characteristic components of plant cells. Each plastid has its own shell, consisting of two elementary membranes. Inside the plastid, a membrane system and a varying degree of a homogeneous substance, the stroma, are distinguished. Mature plastids are classified based on the pigments they contain.
Chloroplasts, in which photosynthesis takes place, contain chlorophylls and carotenoids. Usually have the form of a disk with a diameter of 4 - 5 microns. In one cell of the mesophyll (middle of the leaf) there can be 40 - 50 chloroplasts; in mm2 of a leaf - about 500,000. In the cytoplasm, chloroplasts are usually located parallel to the cell membrane.
The internal structure of the chloroplast is complex. The stroma is permeated with a developed system of membranes in the form of vesicles - thylakoids. Each thylakoid consists of two membranes. Thylakoids form a single system. As a rule, they are collected in piles - grains, resembling columns of coins. The thylakoids of individual granae are interconnected by stroma thylakoids, or intergranal thylakoids. Chlorophyll and carotenoids are embedded in thylakoid membranes. Chloroplasts in green plants and algae often contain grains of starch and small lipid (fat) droplets. Starch grains are temporary stores of photosynthesis products. They can disappear from the chloroplasts of a plant that has been in the dark for only 24 hours, and reappear 3–4 hours after the plant is transferred to light.
Chloroplasts are semi-autonomous organelles and resemble bacteria. For example, the ribosomes of bacteria and chloroplasts have a fairly high similarity. They are smaller than eukaryotic ribosomes. Protein synthesis on the ribosomes of bacteria and chloroplasts is inhibited by chloramphenicol, which has no effect in eukaryotic cells. In addition, both bacteria and chloroplasts have a similar type of nucleoid, organized in a similar way. Despite the fact that the formation of chloroplasts and the synthesis of pigments contained in them is largely controlled by the chromosomal DNA of the cell, nevertheless, in the absence of its own DNA, chloroplasts do not form.
Chloroplasts can be considered the main cellular organelles, since they are the first in the chain of transformation of solar energy, as a result of which humanity receives both food and fuel. In chloroplasts, not only photosynthesis takes place. They are also involved in the synthesis of amino acids and fatty acids, serve as a storage of temporary reserves of starch.
Chromoplasts are pigmented plastids. Diverse in form, they do not have chlorophyll, but synthesize and accumulate carotenoids, which give yellow, orange, red color to flowers, old leaves, fruits and roots. chromoplasts can develop from chloroplasts, which in this case lose chlorophyll and internal membrane structures, accumulate carotenoids. This occurs during the ripening of many fruits. Chromoplasts attract insects and other animals with which they co-evolved.
Leukoplasts are non-pigmented plastids. Some of them synthesize starch (amyloplasts), others are capable of forming various substances, including lipids and proteins. In the light, leukoplasts turn into chloroplasts.
Proplastids are small, colorless or pale green, undifferentiated plastids found in the meristematic (dividing) cells of roots and shoots. They are the precursors of other, more differentiated plastids - chloroplasts, chromoplasts, and aminoplasts. If the development of protoplastids is delayed due to lack of light, one or more prolamellar bodies, which are semi-crystalline accumulations of tubular membranes, may appear in them. Plastids containing prolamellar bodies are called etioplasts. In the light, etioplasts turn into chloroplasts, while the membranes of the prolamellar bodies form thylakoids. Etioplasts are formed in the leaves of plants that are in the dark. protoplasts of seed embryos first turn into etioplasts, from which chloroplasts then develop in the light. Plastids are characterized by relatively easy transitions from one type to another. Plastids, like bacteria, reproduce by dividing in two. In meristematic cells, the time of division of protoplastids approximately coincides with the time of cell division. However, in mature cells, most of the plastids are formed as a result of the division of mature plastids.
Mitochondria. Like chloroplasts, mitochondria are surrounded by two elementary membranes. The inner membrane forms many folds and protrusions - cristae, which significantly increase the inner surface of the mitochondria. They are significantly smaller than plastids, about 0.5 µm in diameter, and variable in length and shape.
In mitochondria, the process of respiration takes place, as a result of which organic molecules are broken down with the release of energy and its transfer to ATP molecules, the main energy reserve of all eukaryotic cells. Most plant cells contain hundreds or thousands of mitochondria. Their number in one cell is determined by the cell's need for ATP. Mitochondria are in constant motion, moving from one part of the cell to another, merging with each other and dividing. Mitochondria usually assemble where energy is needed. If the plasma membrane actively transfers substances from cell to cell, then mitochondria are located along the surface of the membrane. In motile unicellular algae, mitochondria accumulate at the base of the flagella, supplying the energy necessary for their movement.
Mitochondria, like plastids, are semi-autonomous organelles containing the components necessary for the synthesis of their own proteins. The inner membrane surrounds a fluid matrix that contains proteins, RNA, DNA, bacterial-like ribosomes, and various solutes. DNA exists in the form of circular molecules located in one or more nucleoids.
Based on the similarity of bacteria with the mitochondria and chloroplasts of eukaryotic cells, it can be assumed that mitochondria and chloroplasts originated from bacteria that found "refuge" in larger heterotrophic cells - the precursors of eukaryotes.
Microbodies. Unlike plastids and mitochondria, which are delimited by two membranes, microbodies are spherical organelles surrounded by a single membrane. Microbodies have a granular (granular) content, sometimes they also contain crystalline protein inclusions. Microbodies are associated with one or two regions of the endoplasmic reticulum.
Some microbodies, called proxisomes, play an important role in the metabolism of glycolic acid, which is directly related to photorespiration. In green leaves, they are associated with mitochondria and chloroplasts. Other microbodies, called glyoxisomes, contain the enzymes needed to convert fats into carbohydrates. This happens in many seeds during germination.
Vacuoles - These are areas of the cell delimited by the membrane, filled with liquid - cell sap. They are surrounded by a tonoplast (vacuolar membrane).
A young plant cell contains numerous small vacuoles, which merge into one large vacuole as the cell ages. In a mature cell, up to 90% of its volume can be occupied by a vacuole. In this case, the cytoplasm is pressed in the form of a thin peripheral layer to the cell membrane. The increase in cell size is mainly due to the growth of the vacuole. As a result, turgor pressure arises and tissue elasticity is maintained. This is one of the main functions of the vacuole and tonoplast.
The main component of the juice is water, the rest vary depending on the type of plant and its physiological state. Vacuoles contain salts, sugars, less often proteins. The tonoplast plays an active role in the transport and accumulation of certain ions in the vacuole. The concentration of ions in the cell sap can significantly exceed its concentration in the environment. With a high content of certain substances, crystals form in the vacuoles. The most common are calcium oxalate crystals, which have a different shape.
Vacuoles are places of accumulation of metabolic products (metabolism). These can be proteins, acids, and even substances toxic to humans (alkaloids). Pigments are often deposited. Blue, violet, purple, dark red, crimson give plant cells pigments from the anthocyanin group. Unlike other pigments, they dissolve well in water and are found in cell sap. They determine the red and blue color of many vegetables (radishes, turnips, cabbage), fruits (grapes, plums, cherries), flowers (cornflowers, geraniums, delphiniums, roses, peonies). Sometimes these pigments mask chlorophyll in the leaves, for example, in the decorative red maple. Anthocyanins color autumn leaves bright red. They are formed in cold sunny weather, when the synthesis of chlorophyll stops in the leaves. In leaves, when anthocyanins are not formed, after the destruction of chlorophyll, yellow-orange carotenoids of chloroplasts become noticeable. The leaves are most brightly colored in cold, clear autumn.
Vacuoles are involved in the destruction of macromolecules, in the circulation of their components in the cell. Ribosomes, mitochondria, plastids, getting into vacuoles, are destroyed. According to this digestive activity, they can be compared with lysosomes - organelles of animal cells.
Vacuoles are formed from the endoplasmic reticulum (reticulum)
Ribosomes. Small particles (17 - 23nm), consisting of approximately equal amounts of protein and RNA. In ribosomes, amino acids combine to form proteins. There are more of them in cells with an active metabolism. Ribosomes are located in the cytoplasm of the cell freely or attached to the endoplasmic reticulum (80S). They are also found in the nucleus (80S), mitochondria (70S), plastids (70S).
Ribosomes can form a complex on which there is a simultaneous synthesis of identical polypeptides, information about which is taken from one molecule and RNA. Such a complex is called polyribosomes (polysomes). Cells that synthesize proteins in large quantities have an extensive system of polysomes, which are often attached to the outer surface of the nuclear envelope.
Endoplasmic reticulum. It is a complex three-dimensional membrane system of indefinite extent. In the context of the ER looks like two elementary membranes with a narrow transparent space between them. The shape and length of the ER depend on the cell type, its metabolic activity, and the stage of differentiation. In cells that secrete or store proteins, the ER is shaped like flat sacs or cisterns, with numerous ribosomes attached to its outer surface. This reticulum is called the rough endoplasmic reticulum. A smooth ER is usually tubular in shape. Rough and smooth endoplasmic reticulum can be present in the same cell. As a rule, there are numerous connections between them.
The endoplasmic reticulum functions as the communication system of the cell. It is associated with the outer shell of the kernel. In fact, these two structures form a single membrane system. When the nuclear envelope ruptures during cell division, its fragments resemble ER fragments. The endoplasmic reticulum is a system for transporting substances: proteins, lipids, carbohydrates, to different parts of the cell. endoplasmic reticulum of neighboring cells are connected through cytoplasmic strands - plasmodesmata - that pass through the cell membranes.
The endoplasmic reticulum is the main site for the synthesis of cell membranes. In some plant cells, membranes of vacuoles and microbodies, cisterns of dictyosomes are formed here.
Golgi apparatus. This term is used to refer to all dictyosomes, or Golgi bodies, in a cell. Dictyosomes are groups of flat, disc-shaped vesicles, or cisterns, that branch along the edges into a complex system of tubules. Dictyosomes in higher plants consist of 4 - 8 cisterns, collected together.
Usually, in a pack of tanks, a developing and maturing side is distinguished. the membranes of the forming cisterns resemble the membranes of the ER in structure, and the membranes of the maturing cisterns resemble the plasma membrane.
Dictyosomes are involved in secretion and, in most higher plants, in the formation of cell membranes. Cell wall polysaccharides synthesized by dictyosomes accumulate in vesicles, which then separate from maturing cisterns. These secretory vesicles migrate and fuse with the CMP; while the polysaccharides contained in them are built into the cell membrane. Some substances that accumulate in dictyosomes are formed in other structures, for example, in the ER, and then transported to dictyosomes, where they are modified (modified) before secretion. For example, glycoproteins are an important building material of the cell membrane. The protein part is synthesized by polysomes of the rough ER, the carbohydrate part is synthesized in dictyosomes, where both parts combine to form glycoproteins.
Membranes are dynamic, mobile structures that constantly change their shape and area. The concept of the endoplasmic system is based on the mobility of membranes. According to this concept, the inner membranes of the cytoplasm, except for the membranes of mitochondria and plastids, are a single whole and originate from the endoplasmic reticulum. New cisterns of dictyosomes form from the endoplasmic reticulum through the intermediate vesicle stage, and the secretory vesicles that separate from the dictyosomes eventually contribute to the formation of the plasma membrane. Thus, the endoplasmic reticulum and dictyosomes form a functional unit in which dictyosomes play the role of intermediate structures in the process of converting membranes like the endoplasmic reticulum into membranes like the plasma. In tissues whose cells grow and divide weakly, membrane components are constantly being updated.
microtubules found in almost all eukaryotic cells. They are cylindrical structures with a diameter of about 24 nm. Their length varies. Each tubule is made up of subunits of a protein called tubulin. The subunits form 13 longitudinal filaments surrounding the central cavity. Microtubules are dynamic structures, they are regularly destroyed and formed at certain stages of the cell cycle. Their assembly occurs in special places, which are called microtubule organization centers. In plant cells, they have a weakly expressed amorphous structure.
Functions of microtubules: participate in the formation of the cell membrane; direct bubbles of dictyosomes to the forming membrane, like spindle threads that form in a dividing cell; play a role in the formation of the cell plate (the initial boundary between daughter cells). In addition, microtubules are an important component of flagella and cilia, in the movement of which they play an important role.
microfilaments, like microtubules, they are found in almost all eukaryotic cells. They are long filaments 5–7 nm thick, consisting of the contractile protein actin. Bundles of microfilaments are found in many cells of higher plants. Apparently, they play an important role in the currents of the cytoplasm. Microfilaments, together with microtubules, form a flexible network called the cytoskeleton.
Base substance for a long time was considered a homogeneous (homogeneous) protein-rich solution with a small number of structures or even structureless. However, at present, using a high-voltage electron microscope, it has been established that the ground substance is a three-dimensional lattice built from thin (3–6 nm in diameter) strands that fill the entire cell. Other components of the cytoplasm, including microtubules and microfilaments, are suspended from this microtrabecular lattice.
The microtrabecular structure is a lattice of protein strands, the space between which is filled with water. Together with water, the lattice has the consistency of a gel; the gel looks like gelatinous bodies.
Organelles are attached to the microtrabecular lattice. The lattice provides communication between individual parts of the cell and directs intracellular transport.
Lipid drops- structures of a spherical shape, giving granularity to the cytoplasm of a plant cell under a light microscope. On electron micrographs, they look amorphous. Very similar but smaller drops are found in plastids.
Lipid drops, mistaking them for organelles, called them spherosomes and believed that they were surrounded by a single or double layer membrane. However, recent data show that lipid droplets do not have membranes, but may be coated with protein.
Ergastic substances - these are the “passive products” of the protoplast: reserve substances or waste products. They can appear and disappear at different periods of the cell cycle. Except starch grains, crystals, anthocyanin pigments and lipid droplets. These include resins, gums, tannins and proteins. Ergastic substances are part of the cell membrane, the main substance of the cytoplasm and organelles, including vacuoles.
Flagella and cilia - they are thin, hair-like structures that extend from the surface of many eukaryotic cells. They have a constant diameter, but the length varies from 2 to 150 microns. Conventionally, the longer and few of them are called flagella, and the shorter and more numerous are called cilia. There are no clear differences between these two types of structures, so the term flagellum is used to refer to both.
In some algae and fungi, flagella are locomotor organs, with the help of which they move in water. In plants (for example, mosses, liverworts, ferns, some gymnosperms), only germ cells (gametes) have flagella.
Each flagellum has a specific organization. An outer ring of 9 pairs of microtubules surrounds two additional microtubules located in the center of the flagellum. Enzyme-containing "handles" extend from one microtubule of each of the outer pairs. This is the basic 9+2 organization pattern found in all flagella in eukaryotic organisms. It is believed that the movement of flagella is based on the sliding of microtubules, while the outer pairs of microtubules move one along the other without contraction. Sliding of pairs of microtubules relative to each other causes local bending of the flagellum.
Flagella "grow" from cytoplasmic cylindrical structures called basal bodies, which also form the basal part of the flagellum. The basal bodies have an internal structure resembling that of a flagellum, except that the outer tubules are arranged in triplets rather than pairs, and the central tubules are absent.
Cell wall. The cell wall delimits the size of the protoplast and prevents its rupture due to the absorption of water by the vacuole.
The cell wall has specific functions that are important not only for the cell and the tissue in which the cell is located, but also for the whole plant. Cell walls play an essential role in the absorption, transport and excretion of substances, and, in addition, lysosomal, or digestive activity, can be concentrated in them.
Cell wall components. The most typical component of the cell wall is cellulose, which largely determines its architecture. Cellulose molecules are made up of repeating glucose molecules connected end to end. Long thin cellulose molecules are combined into microfibrils with a thickness of 10 - 25 nm. Microfibrils intertwine and form thin threads, which in turn can be wrapped one around the other, like strands in a rope. Each such "rope", or macrofibril, has a thickness of about 0.5 microns, reaching a length of 4 microns. Macrofibrils are as strong as steel wire of equal size.
The cellulose frame of the cell wall is filled with cellulose matrix molecules intertwined with it. It consists of polysaccharides called hemicelluloses, and pectins, or pectins, chemically very close to hemicelluloses.
Another component of the cell wall, lignin, is the most abundant polymer in plant cells after cellulose. Lignin increases the rigidity of the wall and is usually found in cells that perform a supporting or mechanical function.
Cutin, suberin, wax - usually deposited in the shells of the protective tissues of plants. Cutin, for example, is found in the cell membranes of the epidermis, and suberin is found in the secondary protective tissue, the cork. Both substances are found in combination with waxes and prevent excessive water loss from the plant.
Layers of the cell wall. The wall thickness of plant cells varies widely, depending on the role of cells in plant structure and the age of the cell itself. Under an electron microscope, two layers are visible in the plant cell wall: the median lamina (also called the intercellular substance), and the primary cell wall. Many cells lay down another layer, the secondary cell wall. The median plate is located between the primary walls of adjacent cells. The secondary wall, if any, is deposited by the cell protoplast on the inner surface of the primary cell wall.
Middle plate. The median lamina consists mainly of pectin. Where a cell wall should arise, between two newly formed cells, a dense plexus of tubules of the endoplasmic reticulum and the cistern of the Golgi apparatus (dictyosomes) is first noted. Then bubbles appear in this place, filled with pectin (from polysaccharides). These bubbles are separated from the tanks of the Golgi apparatus. The early cell wall contains various polysaccharides, the main ones being pectins and hemicellulose. Later, it consists of denser substances - cellulose and lignin.
Primary cell wall. This is a layer of cellulose sheath that is deposited before or during cell growth. In addition to cellulose, hemicellulose and pectin, the primary membranes contain a glycoprotein. Primary shells can lignify. The pectin component imparts plasticity, which allows the primary shell to expand as the root, stem or leaf elongates.
Actively dividing cells (most mature cells involved in the processes of photosynthesis, respiration and secretion) have primary membranes. Such cells with a primary membrane and a living protoplast are able to lose their characteristic shape, divide and differentiate into a new cell type. They are involved in wound healing and tissue regeneration in plants.
Primary cell membranes are not uniform in thickness throughout their entire length, but have thin sections, which are called primary pore fields. The strands of cytoplasm, or plasmodesmata, that connect the protoplasts of neighboring cells usually pass through the primary pore fields.
secondary cell wall. Despite the fact that many plant cells have only a primary wall, in some, the protoplast deposits a secondary wall towards the center of the cell. This usually occurs after the cessation of cell growth and the area of the primary membrane no longer increases. For this reason, the secondary shell differs from the primary. Secondary membranes are especially needed by specialized cells that strengthen the plant and conduct water. After the deposition of the secondary membrane, the protoplast of these cells, as a rule, dies. There is more cellulose in the secondary shells than in the primary ones, and pectin substances and glycoproteins are absent in them. The secondary membrane is difficult to stretch, its matrix consists of hemicellulose.
In the secondary shell, three layers can be distinguished - outer, middle and inner (S1, S2, S3). The layered structure of the secondary shells significantly increases their strength. Cellulose microfibrils in the secondary membrane are deposited denser than in the primary. Lignin is a common component of the secondary skins of wood.
The pores in the membranes of contacting cells are located opposite each other. Two opposing pores and a pore membrane form a pair of pores. In cells with secondary membranes, there are two main types of pores: simple and bordered. In bordered pores, the secondary wall hangs over the pore cavity. In simple pores, this is not the case.
Cell wall growth. As the cell grows, the thickness and area of the cell membrane increases. Shell stretching is a complex process. It is controlled by the protoplast and regulated by the hormone auxin.
In cells growing uniformly in all directions, the deposition of myofibrils is random. These myofibrils form an irregular network. Such cells were found in the core of the stem, storage tissues, and during cell cultivation in vitro. In elongating cells, the myofibrils of the lateral sheaths are deposited at right angles to the axis of elongation.
Matrix substances - pectins, hemicelluloses and glycoproteins are transferred to the membrane in vesicles of dictyosomes. At the same time, pectins are more characteristic of growing cells, while hemicelluloses predominate in non-growing cells.
Cellulose microfibrils are synthesized on the cell surface with the help of an enzyme complex associated with the plasma membrane. The orientation of microfibrils is controlled by microtubules located near the inner surface of the plasma membrane.
Plasmodesma. These are thin threads of the cytoplasm that connect the protoplasts of neighboring cells. Plasmodesmata either pass through the cell wall anywhere, or are concentrated in primary pore fields or in membranes between pairs of pores. Under an electron microscope, plasmodesmata appear as narrow channels lined with the plasma membrane. Along the axis of the canal, a smaller cylindrical tube, the desmotubule, extends from one cell to another, which communicates with the endoplasmic reticulum of both adjacent cells. Many plasmodesmata form during cell division, when the tubular endoplasmic reticulum is taken up by the developing cell plate. Plasmodesmata can also form in the membranes of non-dividing cells. These structures provide efficient transfer of certain substances from cell to cell.
Cell division. In multicellular organisms, cell division, along with an increase in their size, is a way of growth of the whole organism. New cells formed during division are similar in structure and function, both to the parent cell and to each other. The division process in eukaryotes can be divided into two partially overlapping stages: mitosis and cytokinesis.
Mitosis is the formation of two daughter nuclei from one nucleus, morphologically and genetically equivalent to each other. Cytokinesis is the division of the cytoplasmic part of the cell with the formation of daughter cells.
Cell cycle. A living cell goes through a series of successive events that make up the cell cycle. The duration of the cycle itself varies depending on the type of cell and external factors such as temperature or nutrient availability. Usually the cycle is divided into interphase and four phases of mitosis.
Interphase. The period between successive mitotic divisions.
The interphase is divided into three periods, denoted as G1, S, G2.
During the G1 period, which begins after mitosis. During this period, the amount of cytoplasm increases, including various organelles. In addition, according to the current hypothesis, substances are synthesized during the G1 period that either stimulate or inhibit the S period and the rest of the cycle, thus determining the division process.
The S period follows the G1 period, at which time the genetic material (DNA) is duplicated.
During the G2 period, which follows S, structures that are directly involved in mitosis, such as spindle components, are formed.
Some cells go through an unlimited number of cell cycles. These are unicellular organisms and some cells of active growth zones (meristems). Some specialized cells after maturation lose their ability to reproduce. The third group of cells, for example, those forming wound tissue (callus), retains the ability to divide only under special conditions.
Mitosis, or nuclear fission. It is a continuous process divided into four phases: prophase, metaphase, anaphase, telophase. As a result of mitosis, the genetic material that has doubled in interphase is divided equally between the two daughter nuclei.
One of the earliest signs of a cell's transition to division is the appearance of a narrow, annular band of microtubules just below the plasma membrane. This relatively dense belt surrounds the nucleus in the equatorial plane of the future mitotic spindle. Since it appears before prophase, it is called the preprophase girdle. It disappears after the mitotic spindle, long before the appearance in the late telophase of the cell plate, which grows from the center to the periphery and merges with the mother cell membrane in the area previously occupied by the preprophase belt.
Prophase. At the beginning of prophase, the chromosomes resemble long threads scattered inside the nucleus. Then, as the strands shorten and thicken, one can see that each chromosome is made up of not one, but two intertwined strands called chromatids. In late prophase, the two shortened paired chromatids of each chromosome lie side by side in parallel, connected by a narrow section called the centromere. It has a specific position on each chromosome and divides the chromosome into two arms of different lengths.
Microtubules are located parallel to the surface of the nucleus along the axis of the spindle. This is the earliest manifestation of mitotic spindle assembly.
By the end of prophase, the nucleolus gradually loses its clear outline and finally disappears. Shortly thereafter, the nuclear envelope also disintegrates.
Metaphase. At the beginning of metaphase, the spindle, which is a three-dimensional structure, widest in the middle and tapering towards the poles, takes the place previously occupied by the nucleus. The spindle fibers are bundles of microtubules. During metaphase, the chromosomes, each consisting of two chromatids, are arranged so that their centromeres lie in the equatorial plane of the spindle. With its centromere, each chromosome is attached to the spindle threads. However, some strands pass from one pole to the other without being attached to chromosomes.
When all chromosomes are located in the equatorial plane, metaphase is completed. Chromosomes are ready to divide.
Anaphase. The chromatids of each chromosome separate. Now these are daughter chromosomes. First of all, the centromere divides, and the two daughter chromosomes are drawn to opposite poles. In this case, the centromeres move forward, and the arms of the chromosomes stretch behind. The spindle filaments attached to the chromosomes shorten, contributing to the separation of chromatids and the movement of daughter chromosomes in opposite directions.
Telophase. In telophase, the separation of two identical groups of chromosomes is completed, while a nuclear membrane forms around each of them. The rough reticulum takes an active part in this. The spindle apparatus disappears. During telophase, the chromosomes lose their clarity of outline, stretch out, turning back into thin threads. The nucleoli are restored. When the chromosomes become invisible, mitosis is completed. The two daughter nuclei enter interphase. They are genetically equivalent to each other and to the parent nucleus. This is very important, since the genetic program, and with it all the signs, must be transferred to the daughter organisms.
The duration of mitosis varies in different organisms and depends on the type of tissue. However, prophase is the longest and anaphase is the shortest. In the cells of the root tip, the duration of prophase is 1–2 hours; metaphase - 5 - 15 min; anaphase - 2 - 10 min; telophase - 10 - 30 min. The duration of the interphase is from 12 to 30 hours.
In many eukaryotic cells, the microtubule organizing centers responsible for the formation of the mitotic spindle are associated with centrioles.
Cytokinesis. This is the process of division of the cytoplasm. In most organisms, cells divide by retracting the cell membrane and forming a fission furrow, which gradually deepens, compressing the remaining filaments of the mitotic spindle. In all plants (bryophytes and vascular) and in some algae, cells divide due to the formation of a cell plate.
In the early telophase, a barrel-shaped system of fibers called the phragmoplast forms between the two daughter nuclei. The phragmoplast fibers, like the fibers of the mitotic spindle, consist of microtubules. Small drops appear in the equatorial plane of the phragmoplast. They fuse to form a cell plate that grows until it reaches the wall of the dividing cell. This completes the separation of the two daughter cells. Merging droplets are bubbles breaking away from the Golgi apparatus. Basically, they contain pectin substances, from which the middle plate is formed. Vesicle membranes are involved in the construction of the plasma membrane on both sides of the plate. At the same time, plasmodesmata are formed from fragments of the tubular endoplasmic reticulum.
After the formation of the median plate, each protoplast deposits a primary membrane on it. In addition, each daughter cell deposits a new layer of membrane around the entire protoplast, which continues the membrane that arose from the cell plate. The original shell of the parent cell is destroyed as the daughter cells grow.
Different types of eukaryotic mitosis
The cell division described above in plants, and also in animals, is not the only form of indirect cell division. The simplest type of mitosis is pleuromitosis. It resembles the binary fission of prokaryotic cells, in which the nucleoids after replication remain associated with the plasma membrane. The membrane begins to grow between the DNA binding points and thereby spreads the chromosomes to different parts of the cell. After that, during the formation of a cell constriction, each of the DNA molecules will be in a new separate cell.
Characteristic for the division of eukaryotic cells is the formation of a spindle built of microtubules. At closed pleuromitosis(it is called closed because the divergence of chromosomes occurs without disrupting the nuclear membrane) as microtubule organizing centers (MCMT), not centrioles are involved, but other structures located on the inner side of the nuclear membrane. These are the so-called polar bodies of indeterminate morphology, from which microtubules extend. There are two of these bodies. They diverge from each other without losing contact with the nuclear envelope. As a result of this, two half-spindles are formed associated with chromosomes. The entire process of mitotic spindle formation and chromosome segregation in this case occurs under the nuclear envelope. This type of mitosis occurs among protozoa and is widely distributed in fungi (chytridia, zygomycetes, yeasts, oomycetes, ascomycetes, myxomycetes, etc.). meet forms semi-closed pleuromitosis when the nuclear envelope is destroyed at the poles of the formed spindle.
The next form of mitosis is orthomitosis. In this case, the COMTs are located in the cytoplasm; from the very beginning, not half-spindles are formed, but a bipolar spindle. There are three forms orthomitosis ( normal mitosis) semi-closed And closed. In semi-closed orthomitosis, a bisymmetric spindle is formed with the help of TsOMT located in the cytoplasm, the nuclear envelope is preserved throughout mitosis, with the exception of the polar zones. Masses of granular material, or even centrioles, may be detected as COMT. This form of mitosis is found in zoospores of green, brown, and red algae, in some lower fungi, and in gregarines. With closed orthomitosis, the nuclear membrane is completely preserved, in which a real spindle is formed. Microtubules are formed in the karyoplasm, less often they grow from the internal TsOMT, which is not associated (unlike pleuromitosis) with the nuclear membrane. This type of mitosis is typical for the division of ciliate micronuclei, but can also occur in protozoa. In open orthomitosis, the nuclear envelope completely disintegrates. This type of cell division is characteristic of animal organisms, some protozoa, and cells of higher plants. This form of mitosis, in turn, is represented by astral and anastral types.
From the briefly reviewed material, it can be seen that the main feature of mitosis in general is the appearance of fission spindle structures, which are formed in connection with the CMTs, which are diverse in their structure.
Morphology of the mitotic figure
The mitotic apparatus is especially well expressed at the metaphase stage of mitosis. In metaphase, in the equatorial plane of the cell, chromosomes are located, from which the so-called spindle threads extend in opposite directions, converging at two different poles of the mitotic figure. Thus, the mitotic spindle is a collection of chromosomes, poles and fibers. The spindle fibers are single microtubules or their bundles. Microtubules start from the spindle poles, and some of them go to centromeres where are located kinetochores chromosomes (kinetochore microtubules), some pass further towards the opposite pole, but do not reach it. They are called "interpolar microtubules". A group of radial microtubules departs from the poles, forming around them a structure resembling a "radiant glow" - these are astral microtubules.
By morphology, mitotic figures are divided into astral and anastral types.
The astral spindle type, or convergent, is characterized by the fact that its poles are represented by a small zone to which microtubules converge (converge). Usually, centrosomes containing centrioles are located at the poles of the astral spindles. However, cases of centriolar astral mitoses are known (during meiosis of some invertebrates). In addition, radial microtubules radiating from the poles are noted, which are not part of the spindle, but form stellate zones - citasters. This type of mitotic division resembles a dumbbell. The anastrial type of the mitotic figure does not have citasters at the poles. The polar regions of the spindle here are wide, they are called polar caps, they include centrioles. In this case, the spindle fibers do not depart from one point, but diverge in a wide front (diverge) from the entire zone of the polar caps. This type of spindle is characteristic of dividing cells of higher plants, but can also be found in higher animals. In the early embryogenesis of mammals, centriolar (divergent) mitoses are observed during division of oocyte maturation and at divisions I and II of the zygote. But already in the third cell division and in all subsequent cells, they divide with the participation of astral spindles, in the poles of which centrioles are always found. In general, for all forms of mitosis, chromosomes with their kinetophores, polar bodies (centrosomes), and spindle fibers remain common structures.
Centromeres and kinetochores
Centromeres can have different localization along the length of chromosomes. Holocentric centromeres occur when microtubules are linked along the length of the entire chromosome (some insects, nematodes, some plants). Monocentric centromeres - when microtubules are associated with chromosomes in one site. monocentric centromeres can be punctate (for example, in some budding yeasts), when only one microtubule approaches the kinetochore, and zonal, where a bundle of microtubules approaches the complex kinetochore. Despite the diversity of centromere zones, all of them are associated with the complex structure of the kinetochore, which has a fundamental similarity in structure and functions in all eukaryotes. Kinetochores are special protein structures, mostly located in the centromere zones of chromosomes. These are complex complexes consisting of many proteins. morphologically, they are very similar, have the same structure, ranging from diatoms to humans. They are three-layer structures: the inner dense layer adjacent to the body of the chromosome, the middle loose layer and the outer dense layer. Many fibrils extend from the outer layer, forming the so-called fibrous crown of the kinetochore. In the general form, kinetochores look like plates or disks lying in the zone of the primary constriction of the chromosome, in the centromere. There is usually one kinetochore per chromosome or chromatid. Before anaphase, kinetochores on each sister chromatid are located, each communicating with its own bundle of microtubules. In plants, the kinetochore does not look like plates, but hemispheres. The functional role of kinetochores is to bind sister chromatids to each other, to fix mitotic microtubules, to regulate chromosome separation, and to actually move chromosomes during mitosis with the participation of microtubules. In general, protein structures, kinetochores, double in the S-period, parallel to chromosome doubling. But their proteins are present on the chromosomes at all periods of the cell cycle.
PLANT TISSUE
Educational tissues (meristems)
Educational tissues in the body of plants are located in different places, so they are divided into the following groups (Fig. 0;1).
Apical (apical) meristems are located on the tops, or apexes, of the axial organs - stem, root. With the help of these meristems, the vegetative organs of plants carry out their growth in length.
Lateral meristems are characteristic of axial organs. There they are located concentrically, in the form of a clutch.
Intercalary, or intercalary, meristems originate from apical meristems. These are groups of cells that are not yet able to multiply, but have embarked on the path of differentiation. There are no initial cells among them, but there are many specialized ones.
Wound meristems provide restoration of the damaged part of the body. Regeneration begins with dedifferentiation, that is, the reverse development from specialized cells to meristematic ones. They turn into phellogen, which forms a plug that covers the surface of the wound. Dedifferentiated cells, dividing, can form a loose parenchymal tissue - callus. From it, under certain conditions, plant organs are formed.
Integumentary tissues
They act as a boundary barrier, separating the underlying tissues from the environment. The primary integuments of a plant consist only of living cells. Secondary and tertiary integument - mostly from the dead with thick cell walls.
The main functions of integumentary tissues:
protection of the plant from drying out;
protection against ingress of harmful microorganisms;
sunburn protection;
protection against mechanical damage;
regulation of metabolism between the plant and the environment;
perception of irritation.
The primary integumentary tissue is the epidermis, the epidermis. Consists of living cells. Formed from apical meristems. Covers young growing stems and leaves.
The epidermis was formed in plants in connection with the exit from the aquatic habitat to land in order to prevent it from drying out. In addition to stomata, all cells of the epidermis are tightly interconnected. The outer walls of the main cells are thicker than the rest. The entire surface is covered with a layer of cutin and vegetable waxes. This layer is called the cuticle (skin). It is absent on the growing roots and underwater parts of plants. When dry, the permeability of the cuticle is significantly weakened.
In addition to the main cells, there are others in the epidermis, in particular hairs, or trichomes. They are unicellular and multicellular (Fig. 2). Functionally, they increase the surface of the epidermis, for example, in the root growth zone, serve as mechanical protection, cling to a support, and reduce water loss. A number of plants have glandular hairs, such as nettles.
Only higher plants have stomata in the epidermis, which regulate the exchange of water and gases. If there is no cuticle, then there is no need for stomata. Stomata are a group of cells that form the stomatal apparatus, which consists of two trailing cells and adjoining epidermal cells - side cells. They are different from the main epidermal cells (fig.3). Guard cells differ from their surrounding cells in shape and the presence of a large number of chloroplasts and unevenly thickened walls. Those that face each other are thicker than the rest (fig.4). Between the guard cells, a stomatal gap is formed, which leads to a substomatal space called the substomatal cavity. Guard cells are highly photosynthetic. They contain a large amount of reserve starch and numerous mitochondria.
The number and distribution of stomata, types of stomatal apparatus varies widely in different plants. Stomata are absent in modern bryophytes. Photosynthesis in them is carried out by the gametophyte generation, and sporophytes are not capable of independent existence.
Usually stomata are located on the underside of the leaf. In plants floating on the water surface - on the upper surface. In the leaves of cereals, the stomata are often evenly spaced on both sides. Such leaves are illuminated relatively evenly. On 1 mm2 of the surface, from 100 to 700 stomata can be located.
Secondary integumentary tissue (periderm). This tissue replaces the epidermis when the green color of the annual shoots changes to brown. It is multilayered and consists of a central layer of cambial cells - phellogen. Phellogen cells, dividing, lay a layer of phellem on the outside, and phelloderm on the inside. (Fig.5).
Phelema, or cork. At first it consists of living thin-walled cells. Over time, their walls are impregnated with suberin and vegetable waxes and die off. The contents of the cell are filled with air.
Fellema functions:
prevents moisture loss;
protects the plant from mechanical damage;
protects against pathogens;
provides thermal insulation, as the cells are filled with air.
Cells of the phellogen, located in the epidermis itself, the underlying subepidermal layer, less often in the deep layers of the primary cortex, are the generating basis of the primary cortex.
The cork layer is not permanent. Breaks occur in it, which communicate with the intercellular spaces located nearby. At the same time, small tubercles are formed on the surface - lentils, which communicate the intercellular spaces with atmospheric air. (Fig.6,7).
In autumn, the phellogen under the lenticels lays a layer of corked cells, which greatly reduce transpiration, but do not completely exclude it. In the spring, this layer collapses from the inside. On light birch bark, lentils are clearly visible in the form of dark dashes.
The tertiary integumentary tissue (crust) is also characteristic only for woody forms of plants.
Phellogen is repeatedly laid down in the deeper layers of the cortex. The tissues that are outside of it eventually die off, forming a crust. Its cells are dead and incapable of stretching. However, living cells located deeper divide, which leads to an increase in the transverse size of the stem. Over time, the outer layer of the crust breaks. The time of occurrence of such a gap is a fairly constant value for individual plants. In an apple tree, this happens in the seventh year of life, in a hornbeam - in the fiftieth. In some species it does not occur at all. The main function of the peel is protection against mechanical and thermal damage.
Parenchyma
It is a group of specialized tissues that fill the spaces inside the plant body between conductive and mechanical tissues. (Fig. 8). More often parenchyma cells have a rounded shape, less often an elongated shape. The presence of developed intercellular spaces is characteristic. The spaces between cells together form a transport system - the apoplast. In addition, the intercellular spaces form the "ventilation system" of the plant. Through stomata, or lenticels, they are connected with atmospheric air and provide an optimal gas composition inside the plant. Developed intercellular spaces are especially necessary for plants growing on waterlogged soil, where normal gas exchange is difficult. This parenchyma is called aerenchyma. (Fig. 9).
Elements of the parenchyma, filling the gaps between other tissues, also perform the function of a support. Parenchyma cells are alive, they do not have thick cell walls, like sclerenchyma. Therefore, mechanical properties are provided by turgor. If the water content drops, which leads to plasmolysis and wilting of the plant.
The assimilation parenchyma is formed by thin-walled cells with many intercellular spaces. The cells of this structure contain many chloroplasts, so it is called chlorenchyma. Chloroplasts are located along the wall without shading each other. In the assimilation parenchyma, photosynthesis reactions occur, which provide the plant with organic substances and energy. The result of photosynthetic processes is the possibility of the existence of all living organisms on the Earth.
Assimilation tissues are present only in the illuminated parts of the plant; they are separated from the environment by a transparent epidermis. If the epidermis is replaced by opaque secondary integumentary tissues, the assimilation parenchyma disappears.
The storage parenchyma serves as a receptacle for organic substances that are temporarily not used by the plant organism. In principle, any cell with a living protoplast is capable of depositing organic matter in the form of various kinds of inclusions, but some cells specialize in this. (Figure 10, 11). Energy-rich compounds are deposited only during the growing season, consumed during the dormant period and in preparation for the next growing season. Therefore, reserve substances are deposited in the vegetative organs only in perennial plants.
The receptacle of stocks can be ordinary organs (escape, root), as well as specialized ones (rhizomes, tubers, bulbs). All seed plants store energetically valuable substances in seeds (cotyledons, endosperm). Many plants in arid climates store not only organic matter, but also water. (Fig. 12). For example, aloe stores water in fleshy leaves, cacti in shoots.
mechanical fabrics
The mechanical properties of plant cells are provided by:
hard shell of the cell;
turgidity, that is, the turgor state of cells.
Despite the fact that almost all tissue cells have mechanical properties, however, there are tissues in the plant for which mechanical properties are basic. These are collenchyma and sclerenchyma. They usually function in interaction with other tissues. Inside the body, the plants form a kind of frame. Therefore, they are called reinforcement.
Not all plants have equally well-defined mechanical tissues. To a much lesser extent, plants living in the aquatic environment need internal support than terrestrial ones. The reason is that aquatic plants need less internal support. Their body is largely supported by the surrounding water. Air on land does not create such support, since it has a lower density than water. It is for this reason that the presence of specialized mechanical tissues becomes relevant.
Improvement of internal support structures occurred in the process of evolution.
Collenchyma. It is formed only by living cells, elongated along the axis of the organ. This type of mechanical tissue is formed very early, during the period of primary growth. Therefore, it is important that the cells remain alive and retain the ability to stretch along with the stretching cells that are nearby.
Features of collenchyma cells:
uneven thickening of the shell, as a result of which some parts of it remain thin, while others thicken;
shells do not lignify.
Collenchyma cells are located differently relative to each other. In adjacent cells, thickenings form on the corners facing each other ( rice. 13). Such a collenchyma is called a corner collenchyma. In another case, the cells are arranged in parallel layers. The cell membranes facing these layers are strongly thickened. This is lamellar collenchyma. Cells can be located loosely, with abundant intercellular spaces - this is a loose collenchyma. Such collenchyma is often found in plants on waterlogged soils.
Collenchyma is of particular importance in young plants, herbaceous forms, and also in plant parts where secondary growth does not occur, such as leaves. In this case, it is laid very close to the surface, sometimes just under the epidermis. If the organ has faces, then thick layers of collenchyma are found along their crests.
Collenchyma cells are functional only in the presence of turgor. Water deficiency reduces the effectiveness of collenchyma and the plant temporarily withers, for example, cucumber leaves sagging on a hot day. After filling the cells with water, the functions of collenchyma are restored.
Sclerenchyma. The second type of mechanical tissues. Unlike collenchyma, where all cells are alive, sclerenchyma cells are dead. Their walls are very thick. They perform a mechanical function. A strong thickening of the membrane leads to a violation of the transport of substances, as a result of which the protoplast dies ( rice. 14). The lignification of the cell membranes of sclerenchyma occurs when the plant organ has already completed its growth. Therefore, they no longer interfere with the stretching of surrounding tissues.
Depending on the shape, two types of sclerenchyma cells are distinguished - fibers and sclereids.
The fibers have a strongly elongated shape with very thick walls and a small cavity. They are somewhat smaller than wood fibers. Often under the epidermis form longitudinal layers and strands. In the phloem or xylem, they can be found singly or in groups. In the phloem they are called bast fibers ( rice. 15), and in the xylem - libriform fibers ( rice. 16).
Sclereids, or stony cells, are represented by rounded or branched cells with powerful membranes. In the body of a plant, they can be found singly (supporting cells) ( fig.17) or groups (Fig. 18). It should be noted that the mechanical properties strongly depend on the location of the sclereids. Part of the sclereids form continuous layers, as, for example, in the shell of nuts or in the stones of fruits (stone fruits).
excretory tissues
Any living cell initially has the function of removing substances, while transport can go both along the concentration gradient and against the gradient. Substances that have been synthesized in the cell can be removed and will subsequently affect the activity of other cells (phytohormones, enzymes). This process is called secretion. In the case when substances that are waste products of the cell are excreted, the process is called excretion. Despite the fact that the excretion of substances in plant and animal cells has a fundamental similarity, plants have a number of features that arise from fundamental differences in vital activity. The level of metabolic reactions in plants is much lower than in animals. Therefore, proportionately smaller amounts of waste are emitted. Another feature is that, independently synthesizing almost all the necessary organic compounds, plants never form excessive reserves of them. The released substances can serve as starting material for other reactions (for example, CO2 and H2O).
If in animals the process of excretion of toxins is associated with their removal from the body, plants may not do this, isolating unnecessary substances in the living protoplast (removal of various substances into the vacuole), in dead cells (most tissues of a perennial plant consist of such cells), in intercellular spaces.
The emitted substances can be divided into two large groups. The first group is organic substances synthesized directly by the cell (enzymes, polysaccharides, lignins, terpenes, the latter are constituent elements of essential oils and resins). The second group - substances that initially enter the cell from the outside with the help of xylem or phloem transport (water, mineral salts, amino acids, monosaccharides, etc.). The excreted secret is rarely homogeneous and usually consists of a mixture in which one substance has the highest concentration.
Plants, unlike animals, do not have a complete excretory system. There are only specialized structures scattered throughout the plant - idioblasts. The cells that form them are smaller than the parenchyma cells lying nearby. They have an electron-dense cytoplasm, with developed elements of the endoplasmic reticulum and the Golgi complex. Most often not expressed, there is a central vacuole. Between themselves and other living cells, these cells are connected by numerous plasmodesmata.
Depending on the location, excretory tissues can be external and internal.
External excretory tissues. Glandular hairs are derivatives of the epidermis. Morphologically, they are variable - they can have a multicellular head, be elongated, in the form of a shield on a leg, etc. (fig.19). These include stinging nettle hairs.
Hydathodes are structures that release excess water under conditions of low transpiration and high humidity - guttation. These structures are formed by groups of colorless living cells with thin walls - the epitheme. This tissue is adjacent to the conductive bundle (Fig. 20). Water is released through special water stomata, which differ from ordinary stomata by immobility and a constantly open gap. The composition of the guttation fluid varies widely from almost pure water to a very complex mixture of substances.
Nectaries. Many plants are characterized by the release of a liquid containing from 7 to 87% mono- and oligosaccharides. This process is carried out by special structures - nectaries. Depending on the location, flower, or floral, nectaries are distinguished (Fig. 21), as well as plants located on stems and leaves - extrafloral, or extrafloral nectaries. Nectar can be unaltered phloem juice, which is delivered to the surface through the intercellular spaces and excreted through the stomata. More complex nectars are formed by glandular parenchyma, covered with epidermis with glandular hairs. Nectar is excreted either by epidermal cells or glandular hairs. In this case, the secreted nectar differs from the phloem sap. Since it is dominated by glucose, fructose, sucrose, and in phloem juice - glucose. Ions are present in low concentrations. Ions are present in low concentrations. To attract pollinators, the nectar may contain steroid hormones, which are necessary for insects.
Digestive glands are present in insectivorous plants (sundew, nepenthys, etc.). In response to the touch of small invertebrates, they secrete a secret containing hydrolytic enzymes, formic, butyric and propionic acids. As a result, biopolymers are broken down into monomers, which are used by the plant for its needs.
Salt glands develop in plants growing on saline soils. These glands are located on the leaves and, in a dissolved form, excess ions are brought to the surface. Salt is first deposited on the cuticle, then it is washed off by rain (Fig. 22). In addition, many plants have salt hairs on their leaves. Each hair consists of two cells: one forms the head, the other - the stem. Salts accumulate in the vacuole of the head cell, and when their concentration becomes high, the head falls off. In its place, a new cell grows, which also accumulates salts. During the growth of the leaf, the head may fall off and form several times. The advantage of salt hairs is that by releasing salts, they lose little water. This phenomenon is very important for plants growing on saline soils, where fresh water is scarce.
Internal excretory tissues. Internal excretory structures, scattered throughout the body of the plant in the form of idioblasts, do not remove substances outside the body, accumulating them in themselves. Idioblasts are plant cells that differ in shape, structure, or content from other cells in the same tissue. Depending on the origin, schizogenic (Greek schiso - split) and lysogenic (Greek lysis - dissolution) receptacles of secreted substances are distinguished (Fig. 23).
Schizogenic receptacles. These are more or less extensive intercellular spaces filled with excreted substances. For example, resin passages of conifers, aralia, umbrella and others. It is believed that the resin has bactericidal properties, repels herbivores, making plants inedible for them.
Lysigenic receptacles are formed in place of living cells, which die and are destroyed after the accumulation of substances in them. Such formations can be observed in the peel
STRUCTURE OF THE PLANT CELL
The plant cell is composed of more or less rigid cell wall And protoplast. The cell wall is the cell wall and the cytoplasmic membrane. The term protoplast comes from the word protoplasm, which has long been used to refer to all living things. The protoplast is the protoplasm of an individual cell.
The protoplast is made up of cytoplasm And kernels. The cytoplasm contains organelles (ribosomes, microtubules, plastids, mitochondria) and membrane systems (endoplasmic reticulum, dictyosomes). The cytoplasm also includes the cytoplasmic matrix ( ground substance) in which organelles and membrane systems are immersed. The cytoplasm is separated from the cell wall plasma membrane, which is an elementary membrane. Unlike most animal cells, plant cells contain one or more vacuoles. These are fluid-filled vesicles surrounded by an elementary membrane ( tonoplast).
In a living plant cell, the ground substance is in constant motion. In a movement called cytoplasmic current or cyclosis, organelles are involved. Cyclosis facilitates the movement of substances in the cell and their exchange between the cell and the environment.
Plasma membrane. It is a bilayer phospholipid structure. Plant cells are characterized by invaginations of the plasma membrane.
The plasma membrane performs the following functions:
Participates in the exchange of substances between the cell and the environment;
Coordinates the synthesis and assembly of cellulose microfibrils of the cell wall;
Transmits hormonal and external signals that control cell growth and differentiation.
Core. It is the most prominent structure in the cytoplasm of a eukaryotic cell. The kernel performs two important functions:
Controls the vital activity of the cell, determining which proteins, and at what time should be synthesized;
Stores genetic information and passes it on to daughter cells during cell division.
The nucleus of a eukaryotic cell is surrounded by two elementary membranes that form nuclear envelope. It is permeated with numerous pores with a diameter of 30 to 100 nm, visible only in an electron microscope. The pores have a complex structure. The outer membrane of the nuclear membrane in some places is combined with the endoplasmic reticulum. The nuclear envelope can be viewed as a specialized, locally differentiated part of the endoplasmic reticulum (ER).
In the core dyed with special dyes, thin threads and lumps can be distinguished chromatin And nucleoplasm(the main substance of the nucleus). Chromatin is made up of DNA bound to special proteins called histones. In the process of cell division, chromatin becomes more and more dense and gathers into chromosomes. DNA encodes genetic information.
Organisms differ in the number of chromosomes in somatic cells. For example, cabbage has - 20 chromosomes; sunflower - 34; wheat - 42; people - 46, and one of the species of fern Ophioglossum – 1250. Sex cells (gametes) have only half of the number of chromosomes characteristic of somatic cells of the body. The number of chromosomes in gametes is called haploid(single), in somatic cells - diploid(double). Cells with more than two sets of chromosomes are called polyploid.
Under a light microscope, spherical structures can be seen - nucleoli. Each nucleus has one or more nucleoli, which are visible in non-dividing nuclei. Ribosomal RNA is synthesized in the nucleolus. Typically, the nuclei of diploid organisms have two nucleoli, one for each haploid set of chromosomes. The nucleoli do not have their own membrane. Biochemically, the nucleoli are characterized by a high concentration of RNA, which is associated here with phosphoproteins. The size of the nucleoli depends on the functional state of the cell. it has been noticed that in a rapidly growing cell, in which intensive processes of protein synthesis are taking place, the nucleoli increase in size. In the nucleoli, mRNA and ribosomes are produced, which perform a synthetic function only in the nucleus.
Nucleoplasm (karyoplasm) is represented by a homogeneous liquid in which various proteins, including enzymes, are dissolved.
Plastids. Vacuoles, cellulose cell wall and plastids are characteristic components of plant cells. Each plastid has its own shell, consisting of two elementary membranes. Inside the plastid, a membrane system and a varying degree of homogeneous substance are distinguished - stroma. Mature plastids are classified based on the pigments they contain.
chloroplasts, in which photosynthesis takes place, contain chlorophylls and carotenoids. Usually have the form of a disk with a diameter of 4 - 5 microns. In one cell of the mesophyll (middle of the leaf) there can be 40 - 50 chloroplasts; in mm 2 sheets - about 500,000. in the cytoplasm, chloroplasts are usually located parallel to the cell membrane.
The internal structure of the chloroplast is complex. The stroma is permeated with a developed system of membranes in the form of bubbles - thylakoids. Each thylakoid consists of two membranes. Thylakoids form a single system. As a rule, they are collected in piles - grains resembling columns of coins. The thylakoids of individual granae are interconnected by stroma thylakoids, or intergranal thylakoids. Chlorophyll and carotenoids are embedded in thylakoid membranes. Chloroplasts in green plants and algae often contain grains of starch and small lipid (fat) droplets. Starch grains are temporary stores of photosynthesis products. They can disappear from the chloroplasts of a plant that has been in the dark for only 24 hours, and reappear 3–4 hours after the plant is transferred to light.
Chloroplasts are semi-autonomous organelles and resemble bacteria. For example, the ribosomes of bacteria and chloroplasts have a fairly high similarity. They are smaller than eukaryotic ribosomes. Protein synthesis on the ribosomes of bacteria and chloroplasts is inhibited by chloramphenicol, which has no effect in eukaryotic cells. In addition, both bacteria and chloroplasts have a similar type of nucleoid, organized in a similar way. Despite the fact that the formation of chloroplasts and the synthesis of pigments contained in them is largely controlled by the chromosomal DNA of the cell, nevertheless, in the absence of its own DNA, chloroplasts do not form.
Chloroplasts can be considered the main cellular organelles, since they are the first in the chain of transformation of solar energy, as a result of which humanity receives both food and fuel. In chloroplasts, not only photosynthesis takes place. They are also involved in the synthesis of amino acids and fatty acids, serve as a storage of temporary reserves of starch.
Chromoplasts - pigmented plastids. Diverse in form, they do not have chlorophyll, but synthesize and accumulate carotenoids, which give yellow, orange, red color to flowers, old leaves, fruits and roots. chromoplasts can develop from chloroplasts, which in this case lose chlorophyll and internal membrane structures, accumulate carotenoids. This occurs during the ripening of many fruits. Chromoplasts attract insects and other animals with which they co-evolved.
Leukoplasts - unpigmented plastids. Some of them synthesize starch ( amyloplasts), others are capable of forming various substances, including lipids and proteins. In the light, leukoplasts turn into chloroplasts.
proplastids - small colorless or pale green undifferentiated plastids that are found in meristematic (dividing) cells of roots and shoots. They are the precursors of other, more differentiated plastids - chloroplasts, chromoplasts, and aminoplasts. If the development of protoplastids is delayed due to lack of light, one or more prolamellar bodies, which are semi-crystalline accumulations of tubular membranes. Plastids containing prolamellar bodies are called etioplasts. In the light, etioplasts turn into chloroplasts, while the membranes of the prolamellar bodies form thylakoids. Etioplasts are formed in the leaves of plants that are in the dark. protoplasts of seed embryos first turn into etioplasts, from which chloroplasts then develop in the light. Plastids are characterized by relatively easy transitions from one type to another. Plastids, like bacteria, reproduce by dividing in two. In meristematic cells, the time of division of protoplastids approximately coincides with the time of cell division. However, in mature cells, most of the plastids are formed as a result of the division of mature plastids.
Mitochondria. Like chloroplasts, mitochondria are surrounded by two elementary membranes. The inner membrane forms many folds and protrusions - crist, which significantly increase the inner surface of the mitochondria. They are significantly smaller than plastids, about 0.5 µm in diameter, and variable in length and shape.
In mitochondria, the process of respiration takes place, as a result of which organic molecules are broken down with the release of energy and its transfer to ATP molecules, the main energy reserve of all eukaryotic cells. Most plant cells contain hundreds or thousands of mitochondria. Their number in one cell is determined by the cell's need for ATP. Mitochondria are in constant motion, moving from one part of the cell to another, merging with each other and dividing. Mitochondria usually assemble where energy is needed. If the plasma membrane actively transfers substances from cell to cell, then mitochondria are located along the surface of the membrane. In motile unicellular algae, mitochondria accumulate at the base of the flagella, supplying the energy necessary for their movement.
Mitochondria, like plastids, are semi-autonomous organelles containing the components necessary for the synthesis of their own proteins. The inner membrane surrounds a fluid matrix that contains proteins, RNA, DNA, bacterial-like ribosomes, and various solutes. DNA exists in the form of circular molecules located in one or more nucleoids.
Based on the similarity of bacteria with the mitochondria and chloroplasts of eukaryotic cells, it can be assumed that mitochondria and chloroplasts originated from bacteria that found "refuge" in larger heterotrophic cells - the precursors of eukaryotes.
Microbodies. Unlike plastids and mitochondria, which are delimited by two membranes, microbody are spherical organelles surrounded by a single membrane. Microbodies have a granular (granular) content, sometimes they also contain crystalline protein inclusions. Microbodies are associated with one or two regions of the endoplasmic reticulum.
Some microbodies called proxisomes, play an important role in the metabolism of glycolic acid, which is directly related to photorespiration. In green leaves, they are associated with mitochondria and chloroplasts. Other microbodies, called glyoxysomes, contain the enzymes needed to convert fats into carbohydrates. This happens in many seeds during germination.
Vacuoles - These are areas of the cell delimited by the membrane, filled with liquid - cell sap. They are surrounded tonoplast(vacuolar membrane).
A young plant cell contains numerous small vacuoles, which merge into one large vacuole as the cell ages. In a mature cell, up to 90% of its volume can be occupied by a vacuole. In this case, the cytoplasm is pressed in the form of a thin peripheral layer to the cell membrane. The increase in cell size is mainly due to the growth of the vacuole. As a result, turgor pressure arises and tissue elasticity is maintained. This is one of the main functions of the vacuole and tonoplast.
The main component of the juice is water, the rest vary depending on the type of plant and its physiological state. Vacuoles contain salts, sugars, less often proteins. The tonoplast plays an active role in the transport and accumulation of certain ions in the vacuole. The concentration of ions in the cell sap can significantly exceed its concentration in the environment. With a high content of certain substances, crystals form in the vacuoles. The most common are calcium oxalate crystals, which have a different shape.
Vacuoles are places of accumulation of metabolic products (metabolism). These can be proteins, acids, and even substances toxic to humans (alkaloids). Pigments are often deposited. Blue, violet, purple, dark red, crimson give plant cells pigments from the anthocyanin group. Unlike other pigments, they dissolve well in water and are found in cell sap. They determine the red and blue color of many vegetables (radishes, turnips, cabbage), fruits (grapes, plums, cherries), flowers (cornflowers, geraniums, delphiniums, roses, peonies). Sometimes these pigments mask chlorophyll in the leaves, for example, in the decorative red maple. Anthocyanins color autumn leaves bright red. They are formed in cold sunny weather, when the synthesis of chlorophyll stops in the leaves. In leaves, when anthocyanins are not formed, after the destruction of chlorophyll, yellow-orange carotenoids of chloroplasts become noticeable. The leaves are most brightly colored in cold, clear autumn.
Vacuoles are involved in the destruction of macromolecules, in the circulation of their components in the cell. Ribosomes, mitochondria, plastids, getting into vacuoles, are destroyed. By this digestive activity they can be compared with lysosomes- organelles of animal cells.
Vacuoles are formed from the endoplasmic reticulum (reticulum)
Ribosomes. Small particles (17 - 23nm), consisting of approximately equal amounts of protein and RNA. In ribosomes, amino acids combine to form proteins. There are more of them in cells with an active metabolism. Ribosomes are located in the cytoplasm of the cell freely or attached to the endoplasmic reticulum (80S). They are also found in the nucleus (80S), mitochondria (70S), plastids (70S).
Ribosomes can form a complex on which there is a simultaneous synthesis of identical polypeptides, information about which is taken from one molecule and RNA. Such a complex is called polyribosomes (polysomes). Cells that synthesize proteins in large quantities have an extensive system of polysomes, which are often attached to the outer surface of the nuclear envelope.
Endoplasmic reticulum. It is a complex three-dimensional membrane system of indefinite extent. In the context of the ER looks like two elementary membranes with a narrow transparent space between them. The shape and length of the ER depend on the cell type, its metabolic activity, and the stage of differentiation. In cells that secrete or store proteins, the ER is in the form of flat sacs or cisterns, with numerous ribosomes attached to its outer surface. This reticulum is called rough endoplasmic reticulum. A smooth ER is usually tubular in shape. Rough and smooth endoplasmic reticulum can be present in the same cell. As a rule, there are numerous connections between them.
The endoplasmic reticulum functions as the communication system of the cell. It is associated with the outer shell of the kernel. In fact, these two structures form a single membrane system. When the nuclear envelope ruptures during cell division, its fragments resemble ER fragments. The endoplasmic reticulum is a system for transporting substances: proteins, lipids, carbohydrates, to different parts of the cell. endoplasmic reticulum of neighboring cells are connected through cytoplasmic strands - plasmodesmata - that pass through the cell walls.
The endoplasmic reticulum is the main site for the synthesis of cell membranes. In some plant cells, membranes of vacuoles and microbodies, cisterns dictyosomes.
Golgi apparatus. This term is used to refer to all dictyosome, or golgi body, in a cage. Dictyosomes are groups of flat, disc-shaped vesicles, or cisterns, that branch along the edges into a complex system of tubules. Dictyosomes in higher plants consist of 4 - 8 cisterns, collected together.
Usually, in a pack of tanks, a developing and maturing side is distinguished. the membranes of the forming cisterns resemble the membranes of the ER in structure, and the membranes of the maturing cisterns resemble the plasma membrane.
Dictyosomes are involved in secretion and, in most higher plants, in the formation of cell membranes. Cell wall polysaccharides synthesized by dictyosomes accumulate in vesicles, which then separate from maturing cisterns. These secretory vesicles migrate and fuse with the CMP; while the polysaccharides contained in them are built into the cell membrane. Some substances that accumulate in dictyosomes are formed in other structures, for example, in the ER, and then transported to dictyosomes, where they are modified (modified) before secretion. For example, glycoproteins are an important building material of the cell membrane. The protein part is synthesized by polysomes of the rough ER, the carbohydrate part is synthesized in dictyosomes, where both parts combine to form glycoproteins.
Membranes are dynamic, mobile structures that constantly change their shape and area. The concept is based on membrane mobility endoplasmic system. According to this concept, the inner membranes of the cytoplasm, except for the membranes of mitochondria and plastids, are a single whole and originate from the endoplasmic reticulum. New cisterns of dictyosomes form from the endoplasmic reticulum through the intermediate vesicle stage, and the secretory vesicles that separate from the dictyosomes eventually contribute to the formation of the plasma membrane. Thus, the endoplasmic reticulum and dictyosomes form a functional unit in which dictyosomes play the role of intermediate structures in the process of converting membranes like the endoplasmic reticulum into membranes like the plasma. In tissues whose cells grow and divide weakly, membrane components are constantly being updated.
microtubules found in almost all eukaryotic cells. They are cylindrical structures with a diameter of about 24 nm. Their length varies. Each tubule is made up of subunits of a protein called tubulin. The subunits form 13 longitudinal filaments surrounding the central cavity. Microtubules are dynamic structures, they are regularly destroyed and formed at certain stages of the cell cycle. Their assembly occurs in special places, which are called microtubule organization centers. In plant cells, they have a weakly expressed amorphous structure.
Functions of microtubules: participate in the formation of the cell membrane; direct bubbles of dictyosomes to the forming membrane, like spindle threads that form in a dividing cell; play a role in the formation of the cell plate (the initial boundary between daughter cells). In addition, microtubules are an important component of flagella and cilia, in the movement of which they play an important role.
microfilaments, like microtubules, they are found in almost all eukaryotic cells. They are long filaments 5–7 nm thick, consisting of the contractile protein actin. Bundles of microfilaments are found in many cells of higher plants. Apparently, they play an important role in the currents of the cytoplasm. Microfilaments, together with microtubules, form a flexible network called cytoskeleton.
Base substance for a long time was considered a homogeneous (homogeneous) protein-rich solution with a small number of structures or even structureless. However, at present, using a high-voltage electron microscope, it has been established that the ground substance is a three-dimensional lattice built from thin (3–6 nm in diameter) strands that fill the entire cell. Other components of the cytoplasm, including microtubules and microfilaments, are suspended from this microtrabecular grid.
The microtrabecular structure is a lattice of protein strands, the space between which is filled with water. Together with water, the lattice has the consistency of a gel; the gel looks like gelatinous bodies.
Organelles are attached to the microtrabecular lattice. The lattice provides communication between individual parts of the cell and directs intracellular transport.
Lipid drops- structures of a spherical shape, giving granularity to the cytoplasm of a plant cell under a light microscope. On electron micrographs, they look amorphous. Very similar but smaller drops are found in plastids.
Lipid drops, mistaking them for organelles, called them spherosomes and believed that they were surrounded by a single or double layer membrane. However, recent data show that lipid droplets do not have membranes, but may be coated with protein.
Ergastic substances - these are the “passive products” of the protoplast: reserve substances or waste products. They can appear and disappear at different periods of the cell cycle. Except starch grains, crystals, anthocyanin pigments and lipid droplets. These include resins, gums, tannins and proteins. Ergastic substances are part of the cell membrane, the main substance of the cytoplasm and organelles, including vacuoles.
Flagella and cilia - they are thin, hair-like structures that extend from the surface of many eukaryotic cells. They have a constant diameter, but the length varies from 2 to 150 microns. Conventionally, the longer and few of them are called flagella, and the shorter and more numerous are called cilia. There are no clear differences between these two types of structures, so the term is used to refer to both. flagellum.
In some algae and fungi, flagella are locomotor organs, with the help of which they move in water. In plants (for example, mosses, liverworts, ferns, some gymnosperms), only germ cells (gametes) have flagella.
Each flagellum has a specific organization. An outer ring of 9 pairs of microtubules surrounds two additional microtubules located in the center of the flagellum. Enzyme-containing "handles" extend from one microtubule of each of the outer pairs. This is the basic 9+2 organization pattern found in all flagella in eukaryotic organisms. It is believed that the movement of flagella is based on the sliding of microtubules, while the outer pairs of microtubules move one along the other without contraction. Sliding of pairs of microtubules relative to each other causes local bending of the flagellum.
Flagella "grow" from cytoplasmic cylindrical structures called basal bodies, formed and the basal part of the flagellum. The basal bodies have an internal structure resembling that of a flagellum, except that the outer tubules are arranged in triplets rather than pairs, and the central tubules are absent.
Cell wall. The cell wall delimits the size of the protoplast and prevents its rupture due to the absorption of water by the vacuole.
The cell wall has specific functions that are important not only for the cell and the tissue in which the cell is located, but also for the whole plant. Cell walls play an essential role in the absorption, transport and excretion of substances, and, in addition, lysosomal, or digestive activity, can be concentrated in them.
Cell wall components. The most typical component of the cell wall is cellulose, which largely determines its architecture. Cellulose molecules are made up of repeating glucose molecules connected end to end. Long thin cellulose molecules are combined into microfibrils thickness 10 - 25 nm. Microfibrils intertwine and form thin threads, which in turn can be wrapped one around the other, like strands in a rope. Each such "rope", or macrofibril, has a thickness of about 0.5 microns, reaching a length of 4 microns. Macrofibrils are as strong as steel wire of equal size.
The cellulose frame of the cell wall is filled with cellulose matrix molecules intertwined with it. It contains polysaccharides called hemicelluloses, And pectins, or pectins, chemically very close to hemicelluloses.
Another component of the cell wall is lignin - is the most abundant polymer in plant cells after cellulose. Lignin increases the rigidity of the wall and is usually found in cells that perform a supporting or mechanical function.
Cutin, suberin, wax- are usually deposited in the shells of the protective tissues of plants. Cutin, for example, is found in the cell membranes of the epidermis, and suberin is found in the secondary protective tissue, the cork. Both substances are found in combination with waxes and prevent excessive water loss from the plant.
Layers of the cell wall. The wall thickness of plant cells varies widely, depending on the role of cells in plant structure and the age of the cell itself. Under an electron microscope, two layers are visible in the plant cell wall: middle plate(also called intercellular substance), and primary cell wall. Many cells lay another layer - secondary cell wall. The median plate is located between the primary walls of adjacent cells. The secondary wall, if any, is deposited by the cell protoplast on the inner surface of the primary cell wall.
Middle plate. The median lamina consists mainly of pectin. Where a cell wall should arise, between two newly formed cells, a dense plexus of tubules of the endoplasmic reticulum and the cistern of the Golgi apparatus (dictyosomes) is first noted. Then bubbles appear in this place, filled with pectin (from polysaccharides). These bubbles are separated from the tanks of the Golgi apparatus. The early cell wall contains various polysaccharides, the main ones being pectins and hemicellulose. Later, it consists of denser substances - cellulose and lignin.
Primary cell wall. This is a layer of cellulose sheath that is deposited before or during cell growth. In addition to cellulose, hemicellulose and pectin, the primary membranes contain a glycoprotein. Primary shells can lignify. The pectin component imparts plasticity, which allows the primary shell to expand as the root, stem or leaf elongates.
Actively dividing cells (most mature cells involved in the processes of photosynthesis, respiration and secretion) have primary membranes. Such cells with a primary membrane and a living protoplast are able to lose their characteristic shape, divide and differentiate into a new cell type. They are involved in wound healing and tissue regeneration in plants.
Primary cell membranes are not uniform in thickness throughout their entire length, but have thin sections called primary pore fields. The strands of cytoplasm, or plasmodesmata, that connect the protoplasts of neighboring cells usually pass through the primary pore fields.
secondary cell wall. Despite the fact that many plant cells have only a primary wall, in some, the protoplast deposits a secondary wall towards the center of the cell. This usually occurs after the cessation of cell growth and the area of the primary membrane no longer increases. For this reason, the secondary shell differs from the primary. Secondary membranes are especially needed by specialized cells that strengthen the plant and conduct water. After the deposition of the secondary membrane, the protoplast of these cells, as a rule, dies. There is more cellulose in the secondary shells than in the primary ones, and pectin substances and glycoproteins are absent in them. The secondary membrane is difficult to stretch, its matrix consists of hemicellulose.
In the secondary shell, three layers can be distinguished - outer, middle and inner (S 1, S 2, S 3). The layered structure of the secondary shells significantly increases their strength. Cellulose microfibrils in the secondary membrane are deposited denser than in the primary. Lignin is a common component of the secondary skins of wood.
The pores in the membranes of contacting cells are located opposite each other. Two opposing pores and a pore membrane form a couple of times. In cells with secondary membranes, there are two main types of pores: simple and bordered. In bordered pores, the secondary membrane hangs over pore cavity. In simple pores, this is not the case.
Cell wall growth. As the cell grows, the thickness and area of the cell membrane increases. Shell stretching is a complex process. It is controlled by the protoplast and regulated by a hormone auxin.
In cells growing uniformly in all directions, the deposition of myofibrils is random. These myofibrils form an irregular network. Such cells are found in the stem core, storage tissues, and during cell cultivation in vitro. In elongating cells, the myofibrils of the lateral sheaths are deposited at right angles to the axis of elongation.
Matrix substances - pectins, hemicelluloses and glycoproteins are transferred to the membrane in vesicles of dictyosomes. At the same time, pectins are more characteristic of growing cells, while hemicelluloses predominate in non-growing cells.
Cellulose microfibrils are synthesized on the cell surface with the help of an enzyme complex associated with the plasma membrane. The orientation of microfibrils is controlled by microtubules located near the inner surface of the plasma membrane.
Plasmodesma. These are thin threads of the cytoplasm that connect the protoplasts of neighboring cells. Plasmodesmata either pass through the cell wall anywhere, or are concentrated in primary pore fields or in membranes between pairs of pores. Under an electron microscope, plasmodesmata appear as narrow channels lined with the plasma membrane. A smaller cylindrical tube stretches along the axis of the canal from one cell to another - desmotube, which communicates with the endoplasmic reticulum of both adjacent cells. Many plasmodesmata form during cell division, when the tubular endoplasmic reticulum is taken up by the developing cell plate. Plasmodesmata can also form in the membranes of non-dividing cells. These structures provide efficient transfer of certain substances from cell to cell.
Cell division. In multicellular organisms, cell division, along with an increase in their size, is a way of growth of the whole organism. New cells formed during division are similar in structure and function, both to the parent cell and to each other. The division process in eukaryotes can be divided into two partially overlapping stages: mitosis And cytokinesis.
Mitosis- this is the formation of two daughter nuclei from one nucleus, morphologically and genetically equivalent to each other. Cytokinesis - This is the division of the cytoplasmic part of the cell with the formation of daughter cells.
Cell cycle. A living cell goes through a series of successive events that make up the cell cycle. The duration of the cycle itself varies depending on the type of cell and external factors such as temperature or nutrient availability. The cycle is usually divided into interphase and four phases mitosis.
Interphase. The period between successive mitotic divisions.
The interphase is divided into three periods, denoted as G 1 , S , G 2 .
During the G 1 period, which begins after mitosis. During this period, the amount of cytoplasm increases, including various organelles. In addition, according to the modern hypothesis, during the G 1 period, substances are synthesized that either stimulate or inhibit the S period and the rest of the cycle, thus determining the division process.
The S period follows the G 1 period, at which time the genetic material (DNA) is duplicated.
During the G2 period, which follows S, structures are formed that are directly involved in mitosis, for example, spindle components.
Some cells go through an unlimited number of cell cycles. These are unicellular organisms and some cells of active growth zones (meristems). Some specialized cells after maturation lose their ability to reproduce. The third group of cells, for example, those forming wound tissue (callus), retains the ability to divide only under special conditions.
Mitosis, or nuclear fission. It is an ongoing process divided into four phases: prophase, metaphase, anaphase, telophase. As a result of mitosis, the genetic material that has doubled in interphase is divided equally between the two daughter nuclei.
One of the earliest signs of a cell's transition to division is the appearance of a narrow, annular band of microtubules just below the plasma membrane. This relatively dense belt surrounds the nucleus in the equatorial plane of the future mitotic spindle. Since it appears before prophase, it is called preprophase belt. It disappears after the mitotic spindle, long before the appearance in the late telophase of the cell plate, which grows from the center to the periphery and merges with the mother cell membrane in the area previously occupied by the preprophase belt.
Prophase. At the beginning of prophase, the chromosomes resemble long threads scattered inside the nucleus. Then, as the strands shorten and thicken, it can be seen that each chromosome consists of not one, but two intertwined strands, called chromatids. In late prophase, the two shortened paired chromatids of each chromosome lie side by side in parallel, connected by a narrow section called centromere. It has a specific position on each chromosome and divides the chromosome into two arms of different lengths.
Microtubules are located parallel to the surface of the nucleus along the axis of the spindle. This is the earliest manifestation of mitotic spindle assembly.
By the end of prophase, the nucleolus gradually loses its clear outline and finally disappears. Shortly thereafter, the nuclear envelope also disintegrates.
Metaphase. At first metaphase spindle, which represents a three-dimensional structure, widest in the middle and tapering towards the poles, takes the place formerly occupied by the nucleus. The spindle fibers are bundles of microtubules. During metaphase, the chromosomes, each consisting of two chromatids, are arranged so that their centromeres lie in the equatorial plane of the spindle. With its centromere, each chromosome is attached to the spindle threads. However, some strands pass from one pole to the other without being attached to chromosomes.
When all chromosomes are located in the equatorial plane, metaphase is completed. Chromosomes are ready to divide.
Anaphase. The chromatids of each chromosome separate. now this child chromosomes. First of all, the centromere divides, and the two daughter chromosomes are drawn to opposite poles. In this case, the centromeres move forward, and the arms of the chromosomes stretch behind. The spindle filaments attached to the chromosomes shorten, contributing to the separation of chromatids and the movement of daughter chromosomes in opposite directions.
Telophase. In telophase, the separation of two identical groups of chromosomes is completed, while a nuclear membrane forms around each of them. The rough reticulum takes an active part in this. The spindle apparatus disappears. During telophase, the chromosomes lose their clarity of outline, stretch out, turning back into thin threads. The nucleoli are restored. When the chromosomes become invisible, mitosis is completed. The two daughter nuclei enter interphase. They are genetically equivalent to each other and to the parent nucleus. This is very important, since the genetic program, and with it all the signs, must be transferred to the daughter organisms.
The duration of mitosis varies in different organisms and depends on the type of tissue. However, prophase is the longest and anaphase is the shortest. In the cells of the root tip, the duration of prophase is 1–2 hours; metaphase - 5 - 15 min; anaphase - 2 - 10 min; telophase - 10 - 30 min. The duration of the interphase is from 12 to 30 hours.
In many eukaryotic cells, the microtubule organizing centers responsible for the formation of the mitotic spindle are associated with centrioles.
Cytokinesis. This is the process of division of the cytoplasm. In most organisms, cells divide by retracting the cell membrane and forming a fission furrow, which gradually deepens, compressing the remaining filaments of the mitotic spindle. In all plants (bryophytes and vascular) and in some algae, cells divide due to the formation cell plate.
In early telophase, a barrel-shaped system of fibers is formed between the two daughter nuclei, called phragmoplast. The phragmoplast fibers, like the fibers of the mitotic spindle, consist of microtubules. Small drops appear in the equatorial plane of the phragmoplast. They fuse to form a cell plate that grows until it reaches the wall of the dividing cell. This completes the separation of the two daughter cells. Merging droplets are bubbles breaking away from the Golgi apparatus. Basically, they contain pectin substances, from which the middle plate is formed. Vesicle membranes are involved in the construction of the plasma membrane on both sides of the plate. At the same time, plasmodesmata are formed from fragments of the tubular endoplasmic reticulum.
After the formation of the median plate, each protoplast deposits a primary membrane on it. In addition, each daughter cell deposits a new layer of membrane around the entire protoplast, which continues the membrane that arose from the cell plate. The original shell of the parent cell is destroyed as the daughter cells grow.
Different types of eukaryotic mitosis
The cell division described above in plants, and also in animals, is not the only form of indirect cell division. The simplest type of mitosis is pleuromitosis. It resembles binary division prokaryotic cells, in which the nucleoids after replication remain associated with the plasma membrane. The membrane begins to grow between the DNA binding points and thereby spreads the chromosomes to different parts of the cell. After that, during the formation of a cell constriction, each of the DNA molecules will be in a new separate cell.
Characteristic for division eukaryotic cells is the formation of a spindle built of microtubules. At closed pleuromitosis(it is called closed because the divergence of chromosomes occurs without disrupting the nuclear membrane) as microtubule organizing centers (MCMT), not centrioles are involved, but other structures located on the inner side of the nuclear membrane. These are the so-called polar bodies of indeterminate morphology, from which microtubules extend. There are two of these bodies. They diverge from each other without losing contact with the nuclear envelope. As a result of this, two half-spindles are formed associated with chromosomes. The entire process of mitotic spindle formation and chromosome segregation in this case occurs under the nuclear envelope. This type of mitosis occurs among protozoa and is widely distributed in fungi (chytridia, zygomycetes, yeasts, oomycetes, ascomycetes, myxomycetes, etc.). meet forms semi-closed pleuromitosis when the nuclear envelope is destroyed at the poles of the formed spindle.
The next form of mitosis is orthomitosis. In this case, the COMTs are located in the cytoplasm; from the very beginning, not half-spindles are formed, but a bipolar spindle. There are three forms orthomitosis ( normal mitosis) semi-closed And closed. In semi-closed orthomitosis, a bisymmetric spindle is formed with the help of TsOMT located in the cytoplasm, the nuclear envelope is preserved throughout mitosis, with the exception of the polar zones. Masses of granular material, or even centrioles, may be detected as COMT. This form of mitosis is found in zoospores of green, brown, and red algae, in some lower fungi, and in gregarines. With closed orthomitosis, the nuclear membrane is completely preserved, in which a real spindle is formed. Microtubules are formed in the karyoplasm, less often they grow from the internal TsOMT, which is not associated (unlike pleuromitosis) with the nuclear membrane. This type of mitosis is typical for the division of ciliate micronuclei, but can also occur in protozoa. In open orthomitosis, the nuclear envelope completely disintegrates. This type of cell division is characteristic of animal organisms, some protozoa, and cells of higher plants. This form of mitosis, in turn, is represented by astral and anastral types.
From the briefly reviewed material, it can be seen that the main feature of mitosis in general is the appearance of fission spindle structures, which are formed in connection with the CMTs, which are diverse in their structure.
Morphology of the mitotic figure
The mitotic apparatus is especially well expressed at the metaphase stage of mitosis. In metaphase, in the equatorial plane of the cell, chromosomes are located, from which the so-called spindle threads, converging at two different poles of the mitotic figure. Thus, the mitotic spindle is a collection of chromosomes, poles and fibers. The spindle fibers are single microtubules or their bundles. Microtubules start from the spindle poles, and some of them go to centromeres where are located kinetochores chromosomes (kinetochore microtubules), some pass further towards the opposite pole, but do not reach it. They are called "interpolar microtubules". A group of radial microtubules departs from the poles, forming around them a structure resembling a "radiant glow" - these are astral microtubules.
By morphology, mitotic figures are divided into astral and anastral types.
The astral spindle type, or convergent, is characterized by the fact that its poles are represented by a small zone to which microtubules converge (converge). Usually, centrosomes containing centrioles are located at the poles of the astral spindles. However, cases of centriolar astral mitoses are known (during meiosis of some invertebrates). In addition, radial microtubules radiating from the poles are noted, which are not part of the spindle, but form stellate zones - citasters. This type of mitotic division resembles a dumbbell. The anastrial type of the mitotic figure does not have citasters at the poles. The polar regions of the spindle here are wide, they are called polar caps, they include centrioles. In this case, the spindle fibers do not depart from one point, but diverge in a wide front (diverge) from the entire zone of the polar caps. This type of spindle is characteristic of dividing cells of higher plants, but can also be found in higher animals. In the early embryogenesis of mammals, centriolar (divergent) mitoses are observed during division of oocyte maturation and at divisions I and II of the zygote. But already in the third cell division and in all subsequent cells, they divide with the participation of astral spindles, in the poles of which centrioles are always found. In general, for all forms of mitosis, chromosomes with their kinetophores, polar bodies (centrosomes), and spindle fibers remain common structures.
Centromeres and kinetochores
Centromeres can have different localization along the length of chromosomes. holocentric centromeres occur when microtubules are linked along the length of the entire chromosome (some insects, nematodes, some plants). Monocentric centromeres - when microtubules are associated with chromosomes in one area. monocentric centromeres can be pinpoint(for example, in some budding yeasts), when only one microtubule approaches the kinetochore, and zonal, where a bundle of microtubules approaches the complex kinetochore. Despite the variety of centromere zones, they are all associated with a complex structure. kinetochore, which has a fundamental similarity in structure and function in all eukaryotes. Kinetochores are special protein structures, mostly located in the centromere zones of chromosomes. These are complex complexes consisting of many proteins. morphologically, they are very similar, have the same structure, ranging from diatoms to humans. They are three-layer structures: the inner dense layer adjacent to the body of the chromosome, the middle loose layer and the outer dense layer. Many fibrils extend from the outer layer, forming the so-called fibrous crown of the kinetochore. In the general form, kinetochores look like plates or disks lying in the zone of the primary constriction of the chromosome, in the centromere. There is usually one kinetochore per chromosome or chromatid. Before anaphase, kinetochores on each sister chromatid are located, each communicating with its own bundle of microtubules. In plants, the kinetochore does not look like plates, but hemispheres. The functional role of kinetochores is to bind sister chromatids to each other, to fix mitotic microtubules, to regulate chromosome separation, and to actually move chromosomes during mitosis with the participation of microtubules. In general, protein structures, kinetochores, double in the S-period, parallel to chromosome doubling. But their proteins are present on the chromosomes at all periods of the cell cycle.
PLANT TISSUE
Educational tissues (meristems)
Educational tissues in the body of plants are located in different places, so they are divided into the following groups (Fig. 0;1).
Apical (apical) meristems located on the tops, or apexes, of the axial organs - the stem, root. With the help of these meristems, the vegetative organs of plants carry out their growth in length.
Lateral meristems characteristic of axial organs. There they are located concentrically, in the form of a clutch.
Intercalary, or intercalary, meristems originate from the apical meristems. These are groups of cells that are not yet able to multiply, but have embarked on the path of differentiation. There are no initial cells among them, but there are many specialized ones.
wound meristems provide restoration of the damaged part of the body. Regeneration begins with dedifferentiation, that is, the reverse development from specialized cells to meristematic ones. They turn into fellogen, which forms cork covering the surface of the wound. Dedifferentiated cells, dividing, can form loose parenchymal tissue - callus. From it, under certain conditions, plant organs are formed.
Integumentary tissues
They act as a boundary barrier, separating the underlying tissues from the environment. The primary integuments of a plant consist only of living cells. Secondary and tertiary integument - mostly from the dead with thick cell walls.
The main functions of integumentary tissues:
protection of the plant from drying out;
protection against ingress of harmful microorganisms;
sunburn protection;
protection against mechanical damage;
regulation of metabolism between the plant and the environment;
perception of irritation.
Primary integumentary tissue - epidermis, epidermis . Consists of living cells. Formed from apical meristems. Covers young growing stems and leaves.
The epidermis was formed in plants in connection with the exit from the aquatic habitat to land in order to prevent it from drying out. In addition to stomata, all cells of the epidermis are tightly interconnected. The outer walls of the main cells are thicker than the rest. The entire surface is covered with a layer of cutin and vegetable waxes. This layer is called cuticle(skin). It is absent on the growing roots and underwater parts of plants. When dry, the permeability of the cuticle is significantly weakened.
Except basic cells, there are others in the epidermis, in particular hairs, or trichomes. They are unicellular and multicellular (Fig. 2). Functionally, they increase the surface of the epidermis, for example, in the root growth zone, serve as mechanical protection, cling to a support, and reduce water loss. A number of plants have glandular hairs eg nettle.
Only in higher plants in the epidermis are there stomata that regulate the exchange of water and gases. If there is no cuticle, then there is no need for stomata. Stomata are a group of cells that form stomatal apparatus, which consists of two guard cells and adjoining epidermal cells side cells. They are different from the main epidermal cells (fig.3 ). Guard cells differ from their surrounding cells in shape and the presence of a large number of chloroplasts and unevenly thickened walls. Those that face each other are thicker than the rest (fig.4) . Between the guard cells is formed stomatal opening, which leads to substomatal space called substomatal cavity. Guard cells are highly photosynthetic. They contain a large amount of reserve starch and numerous mitochondria.
The number and distribution of stomata, types of stomatal apparatus varies widely in different plants. Stomata are absent in modern bryophytes. Photosynthesis in them is carried out by the gametophyte generation, and sporophytes are not capable of independent existence.
Usually stomata are located on the underside of the leaf. In plants floating on the water surface - on the upper surface. In the leaves of cereals, the stomata are often evenly spaced on both sides. Such leaves are illuminated relatively evenly. On 1 mm 2 of the surface, from 100 to 700 stomata can be located.
Secondary integumentary tissue (periderm). This tissue replaces the epidermis when the green color of the annual shoots changes to brown. It is multilayered and consists of a central layer of cambial cells - phellogen. Phellogen cells, dividing, lay a layer outside fellems, and inside - phelloderma(Fig.5).
Fellema, or cork. At first it consists of living thin-walled cells. Over time, their walls are impregnated with suberin and vegetable waxes and die off. The contents of the cell are filled with air.
Fellema functions:
prevents moisture loss;
protects the plant from mechanical damage;
protects against pathogens;
provides thermal insulation, as the cells are filled with air.
Cells of the phellogen, located in the epidermis itself, the underlying subepidermal layer, less often in the deep layers of the primary cortex, are the generating basis of the primary cortex.
The cork layer is not permanent. Breaks occur in it, which communicate with the intercellular spaces located nearby. At the same time, small tubercles form on the surface - lentils, which communicate intercellular spaces with atmospheric air (Fig.6,7).
In autumn, the phellogen under the lenticels lays a layer of corked cells, which greatly reduce transpiration, but do not completely exclude it. In the spring, this layer collapses from the inside. On light birch bark, lentils are clearly visible in the form of dark dashes.
Tertiary integumentary tissue (crust), is also characteristic only for woody forms of plants.
Phellogen is repeatedly laid down in the deeper layers of the cortex. The tissues that are outside of it eventually die off, forming a crust. Its cells are dead and incapable of stretching. However, living cells located deeper divide, which leads to an increase in the transverse size of the stem. Over time, the outer layer of the crust breaks. The time of occurrence of such a gap is a fairly constant value for individual plants. In an apple tree, this happens in the seventh year of life, in a hornbeam - in the fiftieth. In some species it does not occur at all. The main function of the peel is protection against mechanical and thermal damage.
Parenchyma
It is a group of specialized tissues that fill the spaces inside the plant body between conductive and mechanical tissues. (Fig. 8). More often parenchyma cells have a rounded shape, less often an elongated shape. The presence of developed intercellular spaces is characteristic. The spaces between cells together form a transport system - apoplast In addition, the intercellular spaces form the "ventilation system" of the plant. Through stomata, or lenticels, they are connected with atmospheric air and provide an optimal gas composition inside the plant. Developed intercellular spaces are especially necessary for plants growing on waterlogged soil, where normal gas exchange is difficult. This parenchyma is called aerenchyma(Fig. 9).
Elements of the parenchyma, filling the gaps between other tissues, also perform the function of a support. Parenchyma cells are alive, they do not have thick cell walls, like sclerenchyma. Therefore, mechanical properties are provided by turgor. If the water content drops, which leads to plasmolysis and wilting of the plant.
Assimilation parenchyma formed by thin-walled cells with many intercellular spaces. The cells of this structure contain many chloroplasts, so it is called chlorenchyma. Chloroplasts are located along the wall without shading each other. In the assimilation parenchyma, photosynthesis reactions occur, which provide the plant with organic substances and energy. The result of photosynthetic processes is the possibility of the existence of all living organisms on the Earth.
Assimilation tissues are present only in the illuminated parts of the plant; they are separated from the environment by a transparent epidermis. If the epidermis is replaced by opaque secondary integumentary tissues, the assimilation parenchyma disappears.
Storage parenchyma serves as a reservoir of organic substances that are temporarily not used by the plant organism. In principle, any cell with a living protoplast is capable of depositing organic matter in the form of various kinds of inclusions, but some cells specialize in this. (Figure 10, 11). Energy-rich compounds are deposited only during the growing season, consumed during the dormant period and in preparation for the next growing season. Therefore, reserve substances are deposited in the vegetative organs only in perennial plants.
The receptacle of stocks can be ordinary organs (escape, root), as well as specialized ones (rhizomes, tubers, bulbs). All seed plants store energetically valuable substances in seeds (cotyledons, endosperm). Many plants in arid climates store not only organic matter, but also water. (Fig. 12). For example, aloe stores water in fleshy leaves, cacti in shoots.
mechanical fabrics
The mechanical properties of plant cells are provided by:
hard shell of the cell;
turgidity, that is, the turgor state of cells.
Despite the fact that almost all tissue cells have mechanical properties, however, there are tissues in the plant for which mechanical properties are basic. This collenchyma And sclerenchyma. They usually function in interaction with other tissues. Inside the body, the plants form a kind of frame. Therefore they are called reinforcing.
Not all plants have equally well-defined mechanical tissues. To a much lesser extent, plants living in the aquatic environment need internal support than terrestrial ones. The reason is that aquatic plants need less internal support. Their body is largely supported by the surrounding water. Air on land does not create such support, since it has a lower density than water. It is for this reason that the presence of specialized mechanical tissues becomes relevant.
Improvement of internal support structures occurred in the process of evolution.
Collenchyma. It is formed only by living cells, elongated along the axis of the organ. This type of mechanical tissue is formed very early, during the period of primary growth. Therefore, it is important that the cells remain alive and retain the ability to stretch along with the stretching cells that are nearby.
Features of collenchyma cells:
uneven thickening of the shell, as a result of which some parts of it remain thin, while others thicken;
shells do not lignify.
Collenchyma cells are located differently relative to each other. In adjacent cells, thickenings form on the corners facing each other ( rice. 13 ). This collenchyma is called corner. In another case, the cells are arranged in parallel layers. The cell membranes facing these layers are strongly thickened. This lamellar collenchyma. Cells can be arranged loosely, with abundant intercellular spaces is loose collenchyma. Such collenchyma is often found in plants on waterlogged soils.
Collenchyma is of particular importance in young plants, herbaceous forms, and also in plant parts where secondary growth does not occur, such as leaves. In this case, it is laid very close to the surface, sometimes just under the epidermis. If the organ has faces, then thick layers of collenchyma are found along their crests.
Collenchyma cells are functional only in the presence of turgor. Water deficiency reduces the effectiveness of collenchyma and the plant temporarily withers, for example, cucumber leaves sagging on a hot day. After filling the cells with water, the functions of collenchyma are restored.
Sclerenchyma. The second type of mechanical tissues. Unlike collenchyma, where all cells are alive, sclerenchyma cells are dead. Their walls are very thick. They perform a mechanical function. A strong thickening of the membrane leads to a violation of the transport of substances, as a result of which the protoplast dies ( rice. 14 ). The lignification of the cell membranes of sclerenchyma occurs when the plant organ has already completed its growth. Therefore, they no longer interfere with the stretching of surrounding tissues.
Depending on the shape, two types of sclerenchyma cells are distinguished - fibers and sclereids.
fibers have a strongly elongated shape with very thick walls and a small cavity. They are somewhat smaller than wood fibers. Often under the epidermis form longitudinal layers and strands. In the phloem or xylem, they can be found singly or in groups. In the phloem they are called bast fibers(rice. 15 ), and in the xylem libriform fibers(rice. 16 ).
sclereids, or stony cells, are represented by rounded or branched cells with powerful shells. In the body of a plant, they can be found singly (supporting cells) ( fig.17 ) or groups (Fig. 18). It should be noted that the mechanical properties strongly depend on the location of the sclereids. Part of the sclereids form continuous layers, as, for example, in the shell of nuts or in the stones of fruits (stone fruits).
excretory tissues
Any living cell initially has the function of removing substances, while transport can go both along the concentration gradient and against the gradient. Substances that have been synthesized in the cell can be removed and will subsequently affect the activity of other cells (phytohormones, enzymes). Such a process is called secretion. In the case when substances that are waste products of the cell are excreted, the process is called excretion. Despite the fact that the excretion of substances in plant and animal cells has a fundamental similarity, plants have a number of features that arise from fundamental differences in vital activity. The level of metabolic reactions in plants is much lower than in animals. Therefore, proportionately smaller amounts of waste are emitted. Another feature is that, independently synthesizing almost all the necessary organic compounds, plants never form excessive reserves of them. The released substances can serve as the starting material for other reactions (for example, CO 2 and H 2 O).
If in animals the process of excretion of toxins is associated with their removal from the body, plants may not do this, isolating unnecessary substances in the living protoplast (removal of various substances into the vacuole), in dead cells (most tissues of a perennial plant consist of such cells), in intercellular spaces.
The emitted substances can be divided into two large groups. The first group is organic substances synthesized directly by the cell (enzymes, polysaccharides, lignins, terpenes, the latter are constituent elements of essential oils and resins). The second group - substances that initially enter the cell from the outside with the help of xylem or phloem transport (water, mineral salts, amino acids, monosaccharides, etc.). The excreted secret is rarely homogeneous and usually consists of a mixture in which one substance has the highest concentration.
Plants, unlike animals, do not have a complete excretory system. There are only specialized structures scattered throughout the plant - idioblasts. The cells that form them are smaller than the parenchyma cells lying nearby. They have an electron-dense cytoplasm, with developed elements of the endoplasmic reticulum and the Golgi complex. Most often not expressed, there is a central vacuole. Between themselves and other living cells, these cells are connected by numerous plasmodesmata.
Depending on the location, excretory tissues can be external and internal.
External excretory tissues.glandular hairs are derivatives epidermis. Morphologically, they are variable - they can have a multicellular head, be elongated, in the form of a shield on a leg, etc. (fig.19). These include stinging nettle hairs.
Hydathodes- these are structures that release excess water under conditions of low transpiration and high humidity - guttation. These structures are formed by groups of colorless living cells with thin walls - epitheme. This tissue is adjacent to the conductive bundle (Fig. 20). Water is released through special water stomata, which differ from ordinary stomata by immobility and a constantly open gap. The composition of the guttation fluid varies widely from almost pure water to a very complex mixture of substances.
Nectaries. Many plants are characterized by the release of a liquid containing from 7 to 87% mono- and oligosaccharides. This process is carried out by special structures - nectaries. Depending on the location, there are flower, or floral, nectaries (Fig. 21), as well as plants located on stems, leaves - extraflora, or extrafloral nectaries. Nectar may represent It is an unchanged phloem juice, which is delivered to the surface through the intercellular spaces and excreted through the stomata. More complex nectars are formed by glandular parenchyma, covered with epidermis with glandular hairs. Nectar is excreted either by epidermal cells or glandular hairs. In this case, the secreted nectar differs from the phloem sap. Since it is dominated by glucose, fructose, sucrose, and in phloem juice - glucose. Ions are present in low concentrations. Ions are present in low concentrations. To attract pollinators, the nectar may contain steroid hormones, which are necessary for insects.
Digestive glands present in insectivorous plants (dew, nepentis, etc.). In response to the touch of small invertebrates, they secrete a secret containing hydrolytic enzymes, formic, butyric and propionic acids. As a result, biopolymers are broken down into monomers, which are used by the plant for its needs.
Salt glands develop in plants growing on saline soils. These glands are located on the leaves and, in a dissolved form, excess ions are brought to the surface. Salt is first deposited on the cuticle, then it is washed off by rain (Fig. 22). In addition, many plants have on the leaves salt hairs. Each hair consists of two cells: one forms the head, the other - the stem. Salts accumulate in the vacuole of the head cell, and when their concentration becomes high, the head falls off. In its place, a new cell grows, which also accumulates salts. During the growth of the leaf, the head may fall off and form several times. The advantage of salt hairs is that by releasing salts, they lose little water. This phenomenon is very important for plants growing on saline soils, where fresh water is scarce.
Internal excretory tissues. Internal excretory structures, scattered throughout the body of the plant in the form of idioblasts, do not remove substances outside the body, accumulating them in themselves. Idioblasts are plant cells that differ in shape, structure, or content from other cells in the same tissue. Depending on the origin, schizogenic (Greek schiso - split) and lysogenic (Greek lysis - dissolution) receptacles of secreted substances are distinguished (Fig. 23).
Schizogenic receptacles. These are more or less extensive intercellular spaces filled with excreted substances. For example, resin passages of conifers, aralia, umbrella and others. It is believed that the resin has bactericidal properties, repels herbivores, making plants inedible for them.
Lysigenic receptacles are formed in place of living cells that die and are destroyed after the accumulation of substances in them. Such formations can be observed in the peel of citrus fruits.
In addition to the main types, there are intermediate forms. milky- there are two types: jointed And non-segmented. The former are formed from many living cells arranged in chains. At the point of contact, the shells are destroyed, the protoplasts merge, and as a result, a single network is formed. Such milky ones are found in Compositae. non-segmented lactifers are formed by a giant multinucleated cell. It arises in the early stages of development, lengthens and branches as the plant grows, forming a complex network, for example, in euphorbia plants (Fig. 25).
The cell walls of lactifers are heavily watered. They do not lignify and therefore are plastic. The central part of the lactis is occupied by a vacuole with latex, and the protoplast occupies a near-wall position. The boundary between the protoplast and the vacuole cannot always be determined. The lactifers penetrate the meristem and grow together with the plant organ.
Conductive tissues
This type belongs to difficult tissues, consists of differently differentiated cells. In addition to the actual conductive elements, the tissue contains mechanical, excretory and storage elements (Fig. 26). Conductive tissues unite all plant organs into a single system. There are two types of conductive tissues: xylem And phloem(Greek xylon - tree; phloios - bark, bast). They have both structural and functional differences.
The conducting elements of the xylem are formed by dead cells. They carry long-distance transport of water and substances dissolved in it from the root to the leaves. The conductive elements of the phloem keep the protoplast alive. They carry long-distance transport from photosynthetic leaves to the root.
Usually xylem and phloem are located in the plant body in a certain order, forming layers or conductive bundles. Depending on the structure, several types of conducting bundles are distinguished, which are characteristic of certain groups of plants. In the collateral open bundle between the xylem and the phloem is the cambium, which provides secondary growth (Fig. 27-A, 28). In a bicollateral open bundle the phloem is located relative to the xylem on both sides (Fig. 27-B, 29). closed bundles do not contain cambium, and hence are not capable of secondary thickening (Fig. 27-B, 27-D, 30.31). There are two more types concentric bundles, where either the phloem surrounds the xylem (Fig. 27-E, 32), or the xylem surrounds the phloem (Fig. 27-E).
Xylem (wood). The development of xylem in higher plants is associated with the provision of water exchange. Since water is constantly excreted through the epidermis, the same amount of moisture must be absorbed by the plant and added to the organs that carry out transpiration. It should be taken into account that the presence of a living protoplast in water-conducting cells would greatly slow down transport, and dead cells are more functional here. However, a dead cell does not have turgidity , therefore, the shell must have mechanical properties. Note: turgescence - the state of plant cells, tissues and organs, in which they become elastic due to the pressure of the contents of the cells on their elastic membranes. Indeed, the conducting elements of the xylem consist of dead cells stretched along the axis of the organ with thick lignified shells.
Initially, xylem is formed from the primary meristem - procambium, located at the tops of the axial organs. First differentiates protoxylem, then metaxylem. Three types of xylem formation are known. At exarch type, protoxylem elements first appear on the periphery of the procambial bundle, then metaxylem elements appear in the center. If the process goes in the opposite direction (i.e. from the center to the periphery), then this endarchic type. At mesarch type xylem is laid down in the center of the procambial bundle, after which it is deposited both towards the center and towards the periphery.
The root is characterized by an exarch type of xylem formation, while the stems have an endarch type. In low organized plants, the methods of xylem formation are very diverse and can serve as systematic characteristics.
In some plants (for example, monocots), all procambial cells differentiate into conductive tissues that are not capable of secondary thickening. In other forms (for example, woody), lateral meristems (cambium) remain between xylem and phloem. These cells are able to divide, renewing the xylem and phloem. Such a process is called secondary growth. Many plants growing in relatively stable climatic conditions grow constantly. In forms adapted to seasonal climate changes - periodically. As a result, well-defined annual growth rings are formed.
The main stages of differentiation of procambial cells. Its cells have thin membranes that do not prevent them from stretching during the growth of the organ. The protoplast then begins to lay down the secondary membrane. But this process has pronounced features. The secondary membrane is not deposited in a continuous layer, which would not allow the cell to stretch, but in the form of rings or in a spiral. Cell elongation is not difficult. In young cells, the rings or coils of the helix are located close to each other. In mature cells, they diverge as a result of cell stretching (Fig. 33). Ringed and spiral thickenings of the shell do not prevent growth, however, mechanically they are inferior to shells, where the secondary thickening forms a continuous layer. Therefore, after the cessation of growth in the xylem, elements with a continuous lignified shell are formed ( metaxylem). It should be noted that the secondary thickening here is not annular or spiral, but punctate, stair-like, reticulate (Fig. 34). Its cells are not able to stretch, and die within a few hours. This process in nearby cells occurs in a coordinated manner. A large number of lysosomes appear in the cytoplasm. Then they decompose, and the enzymes in them destroy the protoplast. When the transverse walls are destroyed, the cells located in a chain one above the other form a hollow vessel (Fig. 35). Most angiosperms and some ferns have vessels.
A conductive cell that does not form through perforations in its wall is called tracheid. The movement of water through the tracheids is slower than through the vessels. The fact is that in tracheids the primary membrane is not interrupted anywhere. The tracheids communicate with each other through since. It should be clarified that in plants, the pore is only a depression in the secondary membrane up to the primary membrane, and there are no through perforations between the tracheids.
The most common are bordered pores (Fig. 35-1). Their channel, facing the cavity of the cell, forms an extension - pore chamber. The pores of most coniferous plants on the primary shell have a thickening - torus, which is a kind of valve and is able to regulate the intensity of water transport. By shifting, the torus blocks the flow of water through the pore, but after that it can no longer return to its previous position, performing a one-time action.
The pores are more or less rounded, elongated perpendicular to the elongated axis (a group of these pores resembles a ladder, so this porosity is called ladder porosity). Transport through the pores is carried out both in the longitudinal and transverse directions. Pores are present not only in tracheids, but also in individual vascular cells that form a vessel.
From the point of view of evolutionary theory, tracheids are the first and main structure that conducts water in the body of higher plants. It is believed that the vessels originated from tracheids due to the lysis of the transverse walls between them (Fig. 36). Most ferns and gymnosperms do not have. They move water through tracheids.
In the process of evolutionary development, vessels arose repeatedly in different groups of plants, but they acquired the most important functional significance in angiosperms, which have them along with tracheids. It is believed that the possession of a more advanced mechanism of transport helped them not only to survive, but also to achieve a significant variety of forms.
Xylem is a complex tissue, in addition to water-conducting elements, it contains others. Mechanical functions are performed by libriform fibers ( lat. liber - lub, forma - form). The presence of additional mechanical structures is important because, despite the thickening, the walls of the water-carrying elements are still too thin. They are not able to independently hold a large mass of a perennial plant. The fibers developed from tracheids. They are characterized by smaller sizes, lignified (lignified) shells and narrow cavities. On the wall, one can find pores devoid of edging. These fibers cannot conduct water, their main function is support.
The xylem also contains living cells. Their mass can reach 25% of the total volume of wood. Since these cells have a rounded shape, they are called wood parenchyma. In the body of the plant, the parenchyma is located in two ways. In the first case, the cells are arranged in the form of vertical strands - this is strand parenchyma. In another case, the parenchyma forms horizontal rays. They're called core rays, as they connect the core and bark. The core performs a number of functions, including the storage of substances.
Phloem (bast). This is a complex tissue, as it is formed by heterogeneous cells. The main conducting cells are called sieve elements(Fig. 37). The conducting elements of the xylem are formed by dead cells, while in the phloem they retain a living, albeit greatly altered, protoplast during the period of functioning. The phloem carries out the outflow of plastic substances from the photosynthetic organs. All living cells of plants possess the ability to conduct organic substances. And hence, if xylem can be found only in higher plants, then the transport of organic substances between cells is also carried out in lower plants.
Xylem and phloem develop from apical meristems. At the first stage, in the procambial cord, a protophloem. As the surrounding tissues grow, it stretches, and when growth is completed, instead of protophloem, it forms metaphloem.
In various groups of higher plants, two types of sieve elements can be found. In ferns and gymnosperms, it is represented sieve cells. The sieve fields in the cells are scattered along the side walls. The protoplast retains a slightly destructed nucleus.
In angiosperms, sieve elements are called sieve tubes. They communicate with each other through sieve plates. Mature cells lack nuclei. However, next to the sieve tube is located satellite cell, which is formed together with the sieve tube as a result of mitotic division of the common mother cell (Fig. 38). The companion cell has a denser cytoplasm with a large number of active mitochondria, as well as a fully functioning nucleus, a huge number of plasmodesmata (ten times more than other cells). Companion cells affect the functional activity of non-nuclear sieve tube cells.
The structure of mature sieve cells has some peculiarities. There is no vacuole, so the cytoplasm is highly liquefied. The nucleus may be absent (in angiosperms) or be in a wrinkled, functionally inactive state. Ribosomes and the Golgi complex are also absent, but the endoplasmic reticulum is well developed, which not only penetrates the cytoplasm, but also passes into neighboring cells through the pores of the sieve fields. Well-developed mitochondria and plastids are found in abundance.
Between cells, the transport of substances goes through holes located on the cell membranes. Such openings are called pores, but unlike the tracheid pores, they are through. It is assumed that they are highly expanded plasmodesmata, on the walls of which the callose polysaccharide is deposited. The pores are arranged in groups, forming sieve fields. In primitive forms, sieve fields are randomly scattered over the entire surface of the shell; in more advanced angiosperms, they are located at the ends of neighboring cells adjacent to each other, forming sieve plate(Fig. 39). If there is one sieve field on it, it is called simple, if there are several - complex.
The speed of movement of solutions through the sieve elements is up to 150 cm ∕ hour. This is a thousand times the rate of free diffusion. Probably, active transport takes place, and numerous mitochondria of sieve elements and satellite cells supply the necessary ATP for this.
The life of the sieve elements of the phloem depends on the presence of lateral meristems. If they are present, then the sieve elements work throughout the life of the plant.
In addition to sieve elements and satellite cells, the phloem contains bast fibers, sclereids, and parenchyma.
Questions for control and repetition
Signs of higher plants? What generation of higher plants is called a gametophyte? Sporophyte?
What are the reasons that contribute to the development of differentiated tissues in terrestrial plants.
List the types of plants.
What groups of meristems do you know? Where are they located in the plant body?
Integumentary tissues, their functions, significance.
Organization of various types of integumentary tissues.
The structure of the stomatal apparatus. List the types of stomatal apparatus known to you. How do stomata function?
Features of parenchymal tissues, types of parenchyma.
Signs of parenchyma and sclerenchyma.
List the types of excretory structures of a plant. What is their significance?
Methods for the formation of receptacles for excreted substances.
What are milkers? What types are they divided into?
How are conductive tissues organized?
What are conductive bundles? Their types.
Directions of movement of substances through the xylem? Phloem? Does the composition of transported substances differ in xylem and phloem?
The structure and functions of xylem, phloem.
Organization of phloem conducting elements in plants from various systematic groups.
Specialized and bacterial, have an outer cell wall. In higher cells Structure and functions of organelles cells. Laws of G. MendelControl work >>
Type fabrics. Our closest relatives, other mammals, structure and... , consisting of one cells. Plants also consist of various ... microtubules, centrioles cell center. IN vegetable cells chloroplasts are also secreted, in which ...
Red color. IN tissues plants anthracene derivatives can be found ... anthracene derivatives as in medicinal vegetable raw materials, and ... are surrounded by 2-3, less often 4 cells epidermis. Unicellular hairs, ... compounds, based buildings of which the structure of anthracene...