Ядро клетки — это обязательная структура каждого эукариотического организма. выполняет самые разнообразные функции, но основное ее предназначение — это хранение и передача наследственного генетического материала.
Практически каждая клетка человеческого организма имеет ядро. Исключение составляют лишь тромбоциты и эритроциты крови. Большинство клеток одноядерные, но, например, мышечные волокна и нейроны могут иметь и несколько этих органелл. Ядро в клетке может иметь разные размеры — самые большие ядерные структуры в женской яйцеклетке.
Ядро клетки: строение
Ядро имеет довольно сложную структуру и состоит их ядерной оболочки, хроматина, ядрышка и нуклеоплазмы. Давайте рассмотрим каждую его часть более подробно.
- Кариотека, или ядерная оболочка — это структура, которая отделяет внутреннюю среду ядра от цитоплазмы. Состоит эта оболочка из внешней и внутренней мембран, между которыми есть так называемое перинуклеарное пространство. Интересно, что внешняя мембрана оболочки переходит непосредственно в мембрану гранулярной эндоплазматической сети, поэтому полости цистерн ЭПС и ядра соединены между собой. А оболочке имеются ядерные поры, закрытые диафрагмой. Они предназначены для проникновения внутрь крупных молекул, а также для обмена веществ между кариоплазмой и цитоплазмой.
- Кариоплазма — гомогенное вещество, которое наполняет внутреннюю полость ядра. В ней содержится ядрышко, а также хроматин.
- Хроматин — генетический материал клетки. Его структурной единицей является нуклеосома, которая представляет собой нить ДНК, намотанную на специфический белок - гистон. В клетке различают два состояния генетического материала. Гетерохроматин —представляет собой небольшие, плотные осмофильные гранулы. Эухроматин, или разрыхленный хроматин — это участки, в которых активно протекают синтетические процессы. Во время хроматин конденсируется, формируя хромосомы.
- Ядрышко — небольшая, овальная структура, которая состоит из нитей РНК и белковых молекул. Именно здесь происходит образование субъединиц рибосом. В ядре может быть одно или несколько ядрышек, но заметить их можно только в неделящихся клетках.
Ядро клетки: функции
Функции можно определить, ознакомившись с его строением. Во-первых, ядро отвечает за передачу наследственного набора информации во время деления клетки, причем как митоза, так и мейоза. Во время митоза дочерние клетки получают геном, который идентичен с материнской клеткой. При мейозе (образовании человека) каждая клетка получает только половину хромосомного набора — полный набор хромосом формируется только после слияния с половой клеткой другого организма.
Кроме того, ядро клетки отвечает за один из самых важных этапов метаболизма — синтез белка. Дело в том, что именно в ядре образуется информационная, или матричная РНК. Затем она выходит в эндоплазматическую сеть, присоединяется к рибосоме и служит моделью для формирования аминокислотной последовательности пептидной молекулы.
И как уже было сказано, в ядре осуществляется синтез субъединиц рибосомы.
Ядро клетки: происхождение
На сегодняшний день существует несколько совершенно разных гипотез, с помощью которых ученые пытаются объяснить, как именно в клетке образовалось ядро. Но, к сожалению, ни одно из этих утверждений еще не нашло фактического подтверждения.
Существует теория, что ядро как клеточная структура образовалось в результате симбиоза и археи. Другие же ученые считают, что ядро — это результат заражения клетки специфическим вирусом.
Наиболее полное объяснение содержит в себе так называемая экзомембранная гипотеза. Согласно ей, в процессе эволюции у клетки возникла еще одна внешняя клеточная оболочка. При этом старая, внутренняя мембрана превратилась в оболочку ядра — со временем в ней возникла сложная система пор, а затем в ее полости начали концентрироваться молекулы хроматина.
Всем известно, что человек является эукариотом. Это значит, что все его клетки имеют органеллу, в которой заключена вся генетическая информация, - ядро. Однако существуют и исключения. Есть ли в организме человека безъядерные клетки и каково их значение для жизнедеятельности?
Безъядерные клетки человека
Их нельзя сравнивать с прокариотами, обладающими типичным строением. Что же это за безъядерные нет в - эритроцитах. Вместо данной органеллы они содержат сложный химический комплекс веществ, позволяющий им выполнять важнейшие для организма функции. Кровяные пластинки - тромбоциты и лимфоциты - также безъядерные клетки. Ядра нет и в клетках, которые называют стволовыми. Все перечисленные структуры объединяет еще один признак. Поскольку в них отсутствует ядро, они не способны к размножению. Это значит, что безъядерные клетки, примеры которых были приведены, после выполнения своей функции гибнут, а новые образуются в специализированных органах.
Эритроциты
Именно они определяют цвет нашей крови. Безъядерные клетки крови эритроциты имеют необычную форму - двояковогнутого диска, которая значительно увеличивает их поверхность при относительно малых размерах. Зато количество их просто поражает: в 1 кв. мм крови их находится до 5 млн! В среднем эритроцит живет до четырех месяцев, после чего погибает и нейтрализуется в селезенке и печени. Новые клетки формируются каждую секунду в красном костном мозге.
Функции эритроцитов
Что же вместо ядра содержат эти безъядерные клетки? Называются эти вещества гем и глобин. Первое является железосодержащим. Оно не только окрашивает кровь в красный цвет, но и образует нестойкие соединения с кислородом и углекислым газом. Глобин представляет собой вещество белковой природы. В его крупную молекулу погружен гем, содержащий заряженный ион железа. По механизму действия эти клетки можно сравнить с маршрутным такси. В легких они присоединяют кислород. С током крови он разносится ко всем клеткам и высвобождается там. При участии кислорода происходит процесс окисления органических веществ с выделением определенного количества энергии, которую человек использует для осуществления жизнедеятельности. Освободившееся место тут же занимает углекислый газ, который движется в обратном направлении - в легкие, где выдыхается. Этот процесс является необходимым условием жизни. Если кислород не поступает к клеткам, происходит их постепенное отмирание. Это может быть опасным для жизни организма в целом.
Эритроциты выполняют еще одну важную функцию. На их мембранах находится белковый маркер, который называется резус-фактором. Этот показатель, как и группа крови, очень важен во время переливания крови, при беременности, донорстве и хирургических операциях. Его обязательно устанавливают, поскольку при несовместимости может произойти так называемый резус-конфликт. Он является защитной реакцией, но может привести к отторжению плода или органов.
Нерациональное питание, вредные привычки, загрязненный воздух могут вызвать разрушение эритроцитов. Вследствие этого возникает тяжелое заболевание, которое называется анемией, или малокровием. При этом человек чувствует головокружение, слабость, одышку, шум в ушах. Кислородная недостаточность негативно сказывается на физической и умственной деятельности человека. Особенно опасна она в период беременности. Если через пуповину к плоду поступает недостаточно кислорода, это может привести к серьезным нарушениям в его развитии.
Строение тромбоцитов
Безъядерные клетки тромбоциты еще называют кровяными пластинками. В неактивном состоянии они действительно имеют плоскую форму, напоминающую линзу. А вот при повреждении сосудов они набухают, округляются, образуют непостоянные выросты наружного слоя - псевдоподии. Тромбоциты образуются в и живут недолго - до 10 дней, обезвреживаясь в селезенке.
Процесс образования тромба
Матрикс кровяных пластинок содержит фермент, который называется тромбопластином. При нарушении целостности сосудов он оказывается в плазме. Под его действием белок крови протромбин переходит в свою активную форму, в свою очередь, действуя на фибриноген. В результате это вещество переходит в нерастворимое состояние. Оно превращается в белок фибрин. Его нити тесно переплетаются и образуют тромб. Защитная реакция свертывания крови предотвращает кровопотери. Однако образование тромба внутри сосуда очень опасно. Это может привести к его разрыву и даже гибели организма. Нарушение процесса свертываемости называется гемофилией. Это наследственное заболевание характеризуется недостаточным количеством тромбоцитов и приводит к излишней потере крови.
Стволовые клетки
Эти безъядерные клетки называются стволовыми не зря. Они действительно являются основой для всех других. Их еще называют "генетически чистыми". Стволовые клетки находятся во всех тканях и органах, но больше всего их содержит костный мозг. Они способствуют восстановлению целостности там, где это необходимо. Стволовые превращаются в любые другие при их разрушении. Казалось бы, при наличии такого волшебного механизма человек должен жить вечно. Почему же этого не происходит? Все дело в том, что с возрастом интенсивность дифференциации стволовых клеток значительно уменьшается. Они уже неспособны восстановить разрушенные ткани. Но есть и еще одна опасность. Существует большая вероятность превращения стволовых клеток в раковые, что неминуемо приведет к гибели любой живой организм.
Безъядерные клетки: примеры и черты отличия
В природе безъядерные клетки встречаются достаточно часто. Например, прокариотическими являются сине-зеленые водоросли и бактерии. Но, в отличие от безъядерных клеток человека, они не гибнут после выполнения своей биологической роли. Дело в том, что прокариоты имеют генетический материал. Поэтому они способны к делению, которое происходит путем две генетические копии материнской клетки. Наследственная информация прокариот представлена кольцевой молекулой ДНК, которая удваивается перед делением. Этот аналог ядра еще называют нуклеоидом. У растений безъядерными являются живые клетки - ситовидные трубки.
Итак, безъядерные клетки человека неспособны к делению, поэтому они существуют непродолжительный промежуток времени до выполнения своей функции. После этого происходит их разрушение и внутриклеточное переваривание. К ним относятся форменные элементы (эритроциты), кровяные пластинки (тромбоциты) и стволовые клетки.
Броун не делал предположений относительно функций ядра. В 1838 году Маттиас Шлейден предположил, что ядро участвует в образовании новых клеток, поэтому он ввёл для обозначения ядер термин «цитобласт» (клеточный строитель). Он был уверен, что наблюдал сборку новых клеток вокруг «цитобластов». Убеждённым оппонентом этого взгляда был Франц Мейен , открывший, что клетки размножаются посредством деления , и считавший, что у многих клеток может не быть ядра. Идея об образовании клеток de novo , то есть с нуля, посредством цитобластов или иначе, противоречила работам Роберта Ремака (1852) и Рудольфа Вирхова (1855), которые окончательно утвердили новую парадигму, утверждающую, что клетки могут образовываться только из клеток («Omnis cellula e cellula»). Функции ядра оставались неясными .
Структуры
Ядро - крупнейшая органелла животных клеток . У млекопитающих диаметр ядра составляет примерно 6 мкм , а само ядро составляет около 10 % объёма клетки . Вязкая жидкость, заполняющая ядро, называется нуклеоплазмой и по химическому составу близка к цитозолю , окружающему ядро .
Ядерная оболочка и ядерные поры
Ядерная оболочка состоит из двух мембран (наружной и внутренней), которые расположены параллельно на расстоянии от 10 до 50 нм . Ядерная оболочка полностью окружает ядро, отделяя генетический материал клетки от цитоплазмы и служа барьером, предотвращающим свободную диффузию макромолекул между нуклеоплазмой и цитоплазмой. Наружная ядерная мембрана продолжается в мембрану шероховатого эндоплазматического ретикулума (ЭПР) и покрыта рибосомами . Промежуток между ядерными мембранами называется перинуклеарным пространством и продолжается в люмен ЭПР .
Ядерные поры, представляющие собой заполненные водой каналы в ядерной оболочке , состоят из множества белков, называемых нуклеопоринами . У человека масса пор составляет около 120 000 кДа ,что в 40 раз больше массы рибосомы ; при этом у дрожжей в состав ядерных пор входит около 50 белков, а у позвоночных - несколько сотен . Хотя диаметр пор составляет 100 нм , ширина щели, через которую могут проходить молекулы, из-за наличия внутри пор регуляторных систем составляет всего 9 нм . В такую щель могут проходить водорастворимые малые молекулы, но не крупные молекулы - такие, как нуклеиновые кислоты и большие белки; для переноса этих молекул в ядро необходим активный (то есть энергозатратный) транспорт. На оболочке ядра типичной клетки млекопитающего располагается от 3000 до 4000 пор , и у каждой в месте слияния двух ядерных мембран находится кольцевая структура, имеющая 8 осей симметрии . К кольцу прикрепляется особая структура, известная как ядерная корзина, которая выдаётся в нуклеоплазму, а несколько её филаментов выдаются в цитоплазму. Обе структуры необходимы для опосредования связывания транспортных ядерных белков .
Большинство белков, субъединицы рибосом и некоторые ДНК переносятся через ядерные поры посредством семейства транспортных факторов, известных как кариоферины . Кариоферины, опосредующие транспорт в ядро, также называются импортинами , а опосредующие транспорт из ядра - экспортинами. Большинство кариоферинов непосредственно взаимодействуют со своим грузом, но некоторые используют для этого адаптерные белки . Стероидные гормоны (такие, как кортизол и альдостерон), а также другие жирорастворимые малые молекулы могут диффундировать в цитоплазму внутрь клетки через клеточную мембрану; в цитоплазме они связываются с белковыми ядерными рецепторами, которые доставляют их в ядро. Здесь ядерные рецепторы , связанные со своими лигандами, функционируют как транскрипционные факторы, а в отсутствие лиганда многие рецепторы функционируют как гистондеацетилазы , подавляющие экспрессию некоторых генов .
Ядерная ламина
В клетках животных механическую поддержку ядра обеспечивают две сети из промежуточных филаментов : ядерная ламина, представляющая собой сеть промежуточных филаментов на внутренней поверхности ядра, а также менее организованные филаменты на цитозольной поверхности ядра. Обе системы филаментов обеспечивают поддержку ядра и служат для закрепления хромосом и ядерных пор .
Ядерная ламина состоит в основном из белков, известных как ламины . Как и все белки, ламины синтезируются в цитоплазме и далее транспортируются внутрь ядра, где они вставляются в ядерную ламину . Расположенные на наружной стороне ядерной оболочки ламины (такие, как эмерин и несприн ), связываются с элементами цитоскелета, что обеспечивает структурную поддержку ядру. Ламины также обнаруживаются в нуклеоплазме, где они образуют другую регулярную структуру, известную как нуклеоплазматическая вуаль (англ. nucleoplasmic veil ) ; последнюю можно визуализировать с использованием флуоресцентной микроскопии . Функция вуали неизвестна, но известно, что её нет в ядрышке и она присутствует в интерфазе клеточного цикла . Входящие в состав вуали ламины (такие, как LEM3) связываются с хроматином , и нарушения в их структуре подавляют транскрипцию белоккодирующих генов .
Как и другие белки промежуточных филаментов, мономеры ламинов содержат α-спиральный домен , используемый двумя мономерами, чтобы обвиться вокруг друг друга, образуя димер , имеющий структуру биспирали . Два димера далее связываются своими боковыми сторонами в антипараллельной ориентации, образуя тетрамер, известный как протофиламент. Восемь тетрамеров объединяются в скрученный, похожий на верёвку филамент. Филаменты могут собираться и разбираться динамическим образом, то есть длина филамента зависит от относительных скоростей его сборки и разборки .
Хромосомы
В ядре находится большая часть генетического материала клетки, представленного множеством линейных молекул ДНК, которые организованы в структуры, известные как хромосомы . Суммарная длина молекул ДНК клетки человека составляет около 2 м . В течение большей части клеточного цикла данные молекулы в комплексе с белками формируют так называемый хроматин , а при клеточном делении хромосомы предстают в виде отдельных хорошо различимых хромосом, составляющих кариотип . Небольшое количество клеточного генетического материала располагается в митохондриях и, в случае растительной клетки , в хлоропластах .
Известно два вида хроматина. В эухроматине ДНК наименее плотно организована; он содержит гены, которые транскрибируются наиболее часто. Другой вид хроматина, гетерохроматин , более компактен и содержит ДНК, транскрибируемую редко или никогда. Гетерохроматин подразделяется на факультативный, который присутствует только в клетках определённого типа и на определённой стадии клеточного цикла, и конститутивный, представленный такими структурами хромосом, как теломеры и центромеры . В течение интерфазы хроматин каждой хромосомы занимает строго определённую область ядра - хромосомную территорию . Активные гены, которые, как правило, располагаются в эухроматине, обычно располагаются на границе хромосомной территории .
Ядерные тельца
В ядре клеток млекопитающих содержится ряд дискретных субкомпартментов , которые называются ядерными тельцами. Они осуществляют компартментализацию ядра, создавая внутри него отдельные пространства, которые обладают определёнными свойствами. Многие ядерные тельца осуществляют специфические функции - например, синтез и процессинг пре-рибосомных РНК в ядрышке, накопление и сборку компоментов сплайсосом в спеклах (см. ниже) или накопление молекул РНК в параспеклах . Механизмы, которые обеспечивают выполнение ядрышковыми тельцами этих функций, очень разнообразны. В некоторых случаях ядерное тельце может служить местом протекания определённых процессов, например, транскрипции. В других случаях ядерные тельца, по-видимому, опосредованно регулируют локальные концентрации своих компонентов в нуклеоплазме. Подобно цитоплазматическим органеллам, ядерные тельца содержат специфический набор белков, которые определяют их структуру на молекулярном уровне. Однако, в отличие от органелл цитоплазмы, ядерные тельца не окружены липидными мембранами, и их структурная целостность целиком обеспечивается белок-белковыми и РНК-белковыми взаимодействиями. Ниже в таблице перечислены основные характеристики ядерных телец .
| Ядерное тельце | Функции | Характерные компоненты | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|---|
| Ядрышко | Биогенез рибосом | Машинерия РНК-полимеразы I , факторы процессинга рРНК и сборки рибосомных субъединиц | 3-8 | 1-4 |
| Спеклы | Накопление и сборка факторов сплайсинга | Факторы сплайсинга пре-мРНК | 2-3 | 20-50 |
| Стрессовые ядерные тельца | Регуляция транскрипции и сплайсинга в условиях стресса | HSF1 , HAP | 1-2 | 3-6 |
| Тельце гистоновых локусов | Процессинг пре-мРНК гистонов | NPAT , FLASH, U7 мяРНП | 0,2-1,2 | 2-4 |
| Тельце Кахаля | Биогенез, созревание и кругооборот малых РНК | Коилин , SMN | 0,2-1,5 | 1-10 |
| PML-тельце | Регуляция стабильности генома, репарация ДНК , контроль транскрипции, защита от вирусов | PML | 0,1-1 | 10-30 |
| Параспеклы | Регуляция мРНК, редактирование РНК | Некодирующие РНК NEAT1/MENε/β, белки PSP1, p54 nrb /NONO | 0,2-1 | 2-20 |
| Околоядрышковый компартмент | Посттранскрипционная регуляция набора РНК, синтезированных РНК-полимеразой III | PTB | 0,2-1 | 1-2 |
Ядрышко
Ядрышко - это отдельная плотная структура в ядре. Она не окружена мембраной и формируется в области расположения рДНК - тандемных повторов генов рибосомной РНК (рРНК) , называемых ядрышковыми организаторами . Главная функция ядрышка - синтез рРНК и образование рибосом. Структурная целостность ядрышка зависит от его активности, и инактивация генов рРНК приводит к смешению ядрышковых структур .
На первой стадии образования рибосом фермент РНК-полимераза I транскрибирует рДНК и образует пре-рРНК, которая далее разрезается на 5,8S, 18S и 28S рРНК . Транскрипция и посттранскрипционный процессинг рРНК происходят в ядрышке при участии малых ядрышковых РНК (snoРНК), некоторые из которых происходят из сплайсированных интронов мРНК генов, кодирующих белки, связанные с работой рибосом. Собранные рибосомные субъединицы - это самые крупные структуры, проходящие через ядерные поры .
При рассматривании под электронным микроскопом в ядрышке можно выделить три компонента: фибриллярные центры (ФЦ), окружающий их плотный фибриллярный компонент (ПФК) и гранулярный компонент (ГК), который, в свою очередь, окружает ПФК. Транскрипция рРНК происходит в ФЦ и на границе ФЦ и ПФК, поэтому при активации образования рибосом ФЦ становятся хорошо различимы. Разрезание и модификации рРНК происходят в ПФК, а последующие этапы образования рибосомных субъединиц, включающие загрузку рибосомных белков, происходят в ГК .
Тельце Кахаля
Тельце Кахаля (ТК) - ядерное тельце, имеющееся у всех эукариот. Оно идентифицируется по наличию сигнатурного белка коилина и специфических РНК (scaРНК). В ТК также содержится белок SMN (англ. survival of motor neurons ). В ТК наблюдается высокая концентрация сплайсирующих малых ядерных рибонуклеопротеинов (мяРНП) и других факторов процессинга РНК, поэтому считается, что ТК служат местами сборки и/или посттранскрипционной модификации факторов сплайсинга. ТК присутствует в ядре во время интерфазы, но исчезает в митозе. В биогенезе ТК прослеживаются свойства самоорганизующейся структуры .
Когда внутриклеточная локализация SMN впервые изучалась методом иммунофлуоресценции , то белок обнаруживался во всей цитоплазме, а также в ядрышковом тельце, сходном по размеру с ТК и часто расположенном рядом с ТК. По этой причине данное тельце было названо «близнецом ТК» (англ. gemini of CB ) или просто gem. Однако оказалось, что линия клеток HeLa , в которой было открыто новое тельце, была необычной: в других линиях клеток человека, а также у плодовой мушки Drosophila melanogaster SMN колокализовался с коилином в ТК. Поэтому в общем случае SMN можно рассматривать как важный компонент ТК, а не как маркер отдельного ядерного тельца .
Тельце гистоновых локусов
PML-тельца
Спеклы
Параспеклы
Параспеклы - это ядерные тельца неправильной формы, располагающиеся в интерхроматиновом пространстве ядра . Впервые они были описаны у клеток HeLa, у которых имеется 10-30 параспеклов на ядро, но сейчас параспеклы обнаружены во всех первичных клетках человека, в клетках трансформированных линий и на срезах тканей . Своё название они получили из-за своего расположения в ядре - вблизи спеклов .
Параспеклы - динамические структуры, которые изменяются в ответ на изменения в метаболической активности клетки. Они зависят от транскрипции , и в отсутствие транскрипции, проводимой РНК-полимеразой II , параспеклы исчезают, а все входящие в их состав белки (PSP1, p54nrb, PSP2, CFI(m)68 и PSF) формируют серповидный околоядрышковый кэп. Этот феномен наблюдается в ходе клеточного цикла: параспеклы присутствуют в интерфазе и всех фазах митоза, за исключением телофазы . В ходе телофазы формируются дочерние ядра, и РНК-полимераза II ничего не транскрибирует, поэтому белки параспеклов и формируют околоядрышковый кэп . Параспеклы участвуют в регуляции экспрессии генов, накапливая те РНК, где есть двухцепочечные участки, которые подвергаются редактированию, а именно превращению аденозина в инозин . Благодаря этому механизму параспеклы задействованы в контроле экспрессии генов при дифференцировке , вирусной инфекции и стрессе .
Околоядрышковый компартмент
Околоядрышковый компартмент (ОК) - ядерное тельце неправильной формы, которое характеризуется тем, что располагается на периферии ядрышка. Несмотря на физическую связь, эти два компартмента структурно различны. Обычно ОК обнаруживают в клетках злокачественных опухолей . ОК - динамическая структура, и содержит очень много РНК-связывающих белков и РНК-полимеразу III. Структурная стабильность ОК обеспечивается транскрипцией, осуществляемой РНК-полимеразой III, и наличием ключевых белков. Поскольку присутствие ОК обычно связано со злокачественностью и со способностью к метастазированию , их рассматривают как потенциальные маркеры рака и других злокачественных опухолей. Показана ассоциация ОК со специфическими локусами ДНК .
Стрессовые ядерные тельца
Стрессовые ядерные тельца формируются в ядре при тепловом шоке. Они образуются при непосредственном взаимодействии транскрипционного фактора теплового шока 1 (HSF1 ) и перицентрических тандемных повторов в последовательности сателлита III, что соответствует сайтам активной траснкрипции некодирующих транскриптов сателлита III. Распространено мнение, что такие тельца соответствуют очень плотно упакованным формам рибонуклеопротеиновых комплексов. Считается, что в клетках, подвергающихся стрессу, они участвуют в быстрых, временных и глобальных изменениях в экспрессии генов посредством различных механизмов - например, ремоделирования хроматина и захватывания факторов транскрипции и сплайсинга. В клетках, находящихся в нормальных (не стрессовых) условиях, стрессовые ядерные тельца обнаруживаются редко, однако их количество резко увеличивается под действием теплового шока. Стрессовые ядерные тельца найдены только в клетках человека и других приматов .
Ядерные тельца-сироты
| Ядерное тельце | Описание | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|
| Кластосома | Концентрирует протеасомные комплексы 20S и 19S и белки, связанные с убиквитином. Обнаруживается, главным образом, тогда, когда стимулируется активность протеасом, и разбирается при ингибировании активности протеасом. | 0,2-1,2 | 0-3 |
| Тельце деления (англ. cleavage body ) | Обогащено факторами деления CstF и CPSF , а также белком DDX1 , содержащим DEAD-бокс . Обнаруживается в основном в S-фазе , ингибирование транскрипции на него не влияет. | 0,2-1,0 | 1-4 |
| Домен OPT | Обогащён факторами транскрипции Oct1 и PTF. Частично колокализуется с сайтами транскрипции. Обнаруживается в основном в поздней G1-фазе , разбирается при ингибировании транскрипции. | 1,0-1,5 | 1-3 |
| Тельце Polycomb | Обнаруживается в клетках человека и дрозофилы, обогащено белком PcG . У человека накапливает белки RING1 , BMI1 , HPC, может быть связано с околоцентромерным гетерохроматином. | 0,3-1,0 | 12-16 |
| Тельце Sam68 | Накапливает белок Sam68 и схожие с ним белки SLM-1 и SLM-2. Разбирается при ингибировании транскрипции. Вероятно, обогащено РНК. | 0,6-1,0 | 2-5 |
| Тельце SUMO | Обогащено белками SUMO и SUMO-конъюгирующим ферментом Ubc9 . Концентрирует транскрипционные факторы pCREB , CBP , c-Jun . | 1-3 | 1-3 |
Функции
Ядерная оболочка защищает ДНК клетки и участвует в гораздо более сложной регуляции экспрессии генов по сравнению с прокариотической клеткой. У прокариот транскрипция и трансляция являются сопряжёнными процессами, и трансляция мРНК в белок начинается ещё до того, как она будет полностью синтезирована. В клетках эукариот цитоплазма, в которой проходит трансляция, и транскрипция, протекающая в ядре, пространственно разобщены, поэтому возникает необходимость в обеспечении транспорта молекул между ядром и цитоплазмой .
Ядерная оболочка даёт ядру возможность контролировать своё содержимое и отделяет его от остальной цитоплазмы. Это имеет важное значение для регуляции процессов, протекающих по обе стороны ядерной оболочки. Когда цитоплазматический процесс должен быть как-то ограничен, то обычно его ключевой участник переносится в ядро, где он взаимодействует с факторами транскрипции и таким образом запускает подавление образования некоторых ферментов, задействованных в цитоплазматическом процессе. Например, такой регуляторный механизм имеется у гликолиза - процесса, в ходе которого клетка извлекает энергию из молекулы глюкозы . Первую реакцию гликолиза осуществляет фермент гексокиназа , преобразуя молекулу глюкозы в глюкозо-6-фосфат . Когда концентрация фруктозо-6-фосфата (вещества, в ходе гликолиза образующегося из глюкозо-6-фосфата) возрастает, регуляторный белок отправляет гексокиназу в ядро , где она формирует транскрипционный репрессирующий комплекс, который подавляет экспрессию генов, кодирующих ферменты гликолиза .
Чтобы контролировать, какие именно гены транскрибируются, в клетке транскрипционные факторы не имеют физического доступа к ДНК, пока они не будут активированы в ходе определённого сигнального пути . Это предотвращает даже низкую экспрессию неправильных генов. В частности, в случае контролируемых NF-κB генов, которые принимают участие в воспалительном процессе , транскрипция индуцируется под действием сигнального пути, например, начинающегося со связывании сигнальной молекулы TNF-α со своим рецептором на клеточной мембране и в конце концов приводящего к активации фактора транскрипции NF-κB. Сигнал ядерной локализации , имеющийся у NF-κB, позволяет ему проходить в ядро и из него через ядерные поры; в ядре он стимулирует транскрипцию генов-мишеней .
Компартментализация предотвращает транскрипцию клеткой несплайсированной мРНК. Эукариотические мРНК содержат интроны, которые должны быть удалены до того, как начнётся трансляция мРНК. Сплайсинг, то есть удаление интронов, протекает в ядре, что предотвращает доступ к пре-мРНК рибосом, находящихся вне ядра. Если бы ядра не было, то рибосомы начинали бы транслировать незрелые мРНК, что привело бы к образованию неправильных белковых продуктов .
Поскольку транскрипция протекает в ядре, ядро содержит множество белков, непосредственно участвующих в транскрипции или регулирующих этот процесс. К этим белкам относятся хеликазы , которые расплетают двойную спираль ДНК, облегчая доступ к ней других белков, РНК-полимеразы , которые синтезируют РНК, топоизомеразы , влияющие на топологию ДНК, а также разнообразные факторы транскрипции .
Ядерный транспорт
Выход из ядра и вход в ядро крупных молекул контролируется ядерными порами. Хотя малые молекулы могут проникать в ядро без всякой регуляции, макромолекулы - такие, как белки и РНК - должны связаться с кариоферинами для транспорта в ядро (импортинами) и из ядра (экспортинами). Белки, которые должны быть транспортированы из цитоплазмы в ядро, содержат особую аминокислотную последовательность, известную как сигнал ядерной локализации, с которой связываются импортины. Аналогичным образом белки, которые должны выйти из ядра, содержат сигнал ядерного экспорта , распознаваемый экспортинами. Способность импортинов и экспортинов переносить свой груз регулируется ГТФазами - ферментами, которые гидролизуют ГТФ с высвобождением энергии . Ключевая ГТФаза ядерного транспорта - Ran , которая может связываться с ГТФ или ГДФ , в зависимости от своего местонахождения (в ядре или в цитоплазме). В ядре взаимодействие Ran-ГТФ с импортином вызывает конформационные изменения в последнем, так что он отделяется от переносимого груза. Образованный комплекс Ran-ГТФ и импортина транспортируется в цитоплазму, где белок RanBP отделяет Ran-ГТФ от импортина. Отделение от импортина позволяет белку GAP связаться с Ran-ГТФ и катализировать гидролиз ГТФ до ГДФ. Далее комплекс Ran-GDP распознаётся белком NUTF2 , который возвращает его в нуклеоплазму. В ядре белок GEF заменяет ГДФ на ГТФ, образуя Ran-ГТФ и замыкая цикл .
Сборка и разборка
В течение жизни клетки ядро может быть разобрано (при делении клетки или при апоптозе). В ходе этих процессов структурные компоненты ядра - ядерная оболочка и ядерная ламина - разрушаются. В большинстве клеток разборка ядра наблюдается в профазе митоза. Однако разборка ядра не приурочена строго к митозу и происходит не во всех клетках. Некоторые одноклеточные эукариоты (например, дрожжи) подвергаются так называемому закрытому митозу, при котором ядерная оболочка остаётся целой. При закрытом митозе хромосомы перемещаются к разным сторонам ядра, которое потом делится надвое. Клетки высших эукариот, напротив, обычно подвергаются открытому митозу, в ходе которого ядерная оболочка распадается. Хромосомы мигрируют к разным полюсам веретена деления , и вокруг них заново формируются два ядра. Ядерная ламина тоже подвергается разборке из-за фосфорилирования ламинов такими киназами , как циклинзависимая протеинкиназа 1 . Сборка ядерной ламины в дочерних ядрах начинается после дефосфорилирования ламинов .
Апоптоз - это контролируемый процесс разрушения клеточных компонентов, приводящего к гибели клетки. Перемены, связанные с апоптозом, происходят непосредственно с ядром и его содержимым. К их числу относится конденсация хроматина, а также дезинтеграция ядерной оболочки и ядерной ламины. Разрушение сети ламинов происходит с участием апоптотических протеаз , известных как каспазы , которые разрушают ламины и, таким образом, влияют на структурную целостность ядра. Разрушение ламинов иногда используется в качестве индикатора активности каспаз в исследованиях, посвящённых апоптозу. Клетки, в которых экспрессируются мутантные ламины, устойчивые к действию каспаз, при апоптозе не утрачивают целостность ядра, поэтому ламины играют ключевую роль в начале изменений, которое претерпевает ядро при апоптозе . Кроме того, ингибирование сборки ламинов в сеть запускает апоптоз .
Особенности ядер у различных эукариот
Размеры, формы и морфология ядер эукариот изменяются в самых широких пределах. Если у пироплазмид и лейшманий диаметр ядра составляет 1-3 мкм , то у некоторых радиолярий ядра в диаметре достигают 400 мкм и даже 1 мм . Как правило, форма ядра у большинства эукариот близка к сферической, но иногда она способна принимать довольно причудливые очертания (это, в частности, относится к макронуклеусам инфузорий). Хотя у всех эукариот оболочка ядра состоит из двух мембран, число пор в ней у различных видов сильно варьирует, причём иногда к ней (как снаружи, так и изнутри) могут примыкать дополнительные слои; например, у многих свободноживущих амёб к внутренней стороне оболочки прилегает фиброзный слой с ячеистым строением, который значительно превосходит ядерную оболочку по толщине, а у радиолярий с внешней стороны оболочки располагаются дополнительные фибриллярные слои .
Значительным своеобразием отличается организация ядра у протистов из типа динофлагеллят (Dinoflagellata). Большинство их представителей имеет ядро, в котором хромосомы конденсированы на протяжении всего клеточного цикла (в том числе и в интерфазе) и практически лишены гистонов . Такой тип ядра получил название динокарион . При этом количество ДНК в динокарионе в десятки и сотни раз превосходит количество ДНК, приходящееся на клетку у представителей других групп эукариот . Впрочем, некоторые динофлагелляты (Noctiluca , Oodinium ) имеют обычные эукариотические ядра ; у других представителей типа в вегетативных клетках ядра обычные, а динокарион присутствует на других стадиях клеточного цикла (например, в гаметах) .
Клетки протистов обладают по крайней мере одним ядром . В то же время в организмах Metazoa встречаются и безъядерные клетки, которые, не имея ядра, утратили способность делиться с образованием двух дочерних клеток. Наиболее известным примером безъядерных клеток являются эритроциты млекопитающих, в которых отсутствуют и другие органеллы - такие, как митохондрии . Эритроциты созревают в костном мозге в процессе эритропоэза , в ходе которого они утрачивают ядра, другие органеллы и рибосомы. Ядро выталкивается из клетки при процессе дифференцировки эритробласта в ретикулоцит , который выступает непосредственным предшественником эритроцита . Под действием некоторых мутагенов в кровь могут выпускаться незрелые эритроциты, содержащие микроядра.
Большинство протистов имеет только одно ядро; у протистов, для которых характерен сложный жизненный цикл (например, у представителей типа апикомплексы (Apicomplexa) встречаются одноядерные и многоядерные стадии .
Многоядерные клетки протистов
В составе хлоропластов у криптофитовых и хлорарахниофитовых водорослей присутствует нуклеоморф - редуцированное ядро фототрофного эндосимбионта , инкорпорированного предками данных водорослей в ходе вторичного эндосимбиоза (у Cryptophyta происходила инкорпорация красной , а у Chlorarachnea - зелёной водоросли)
КЛЕТОЧНОЕ ЯДРО
Ядро (nucleus, s. kаryon) имеется во всех клетках человека, кроме эритроцитов и тромбоцитов. Функции ядра - хранение и передача новым (дочерним) клеткам наследственной информации. Эти функции связаны с наличием в ядре ДНК. В ядре происходит также синтез белков - рибонуклеиновой кислоты РНК и рибосомных материалов.
У большинства клеток ядро шаровидное или овоидное, однако встречаются и другие формы ядра (кольцевидное, палочковидное, веретеновидное, четковидное, бобовидное, сегментированное, грушевидное, полиморфное). Размеры ядра колеблются в широких пределах - от 3 до 25 мкм. Наиболее крупное ядро имеет яйцеклетка. Большинство клеток человека одноядерные, однако имеются двухъядерные (некоторые нейроны, гепатоциты, кардиомиоциты). Некоторые структуры являются многоядерными (мышечные волокна). У ядра различают ядер- ную оболочку, хроматин, ядрышко и нуклеоплазму (рис. 4).
Ядерная оболочка, или кариотека (caryothйca), отделяющая содержимое ядра от цитоплазмы, состоит из внутренней и наружной ядерных мембран толщиной 8 нм каждая. Мембраны разделены перинуклеарным пространством (цистерна кариотеки) шириной 20-50 нм, которое содержит мелкозернистый материал умеренной электронной плотности. Наружная ядерная мембрана переходит в зернистую эндоплазматическую сеть. Поэтому перинуклеарное пространство составляет единую полость с эндоплазматической сетью. Внутренняя ядерная мембрана изнутри соединена с разветвленной сетью белковых фибрилл, состоящих из отдельных субъединиц.
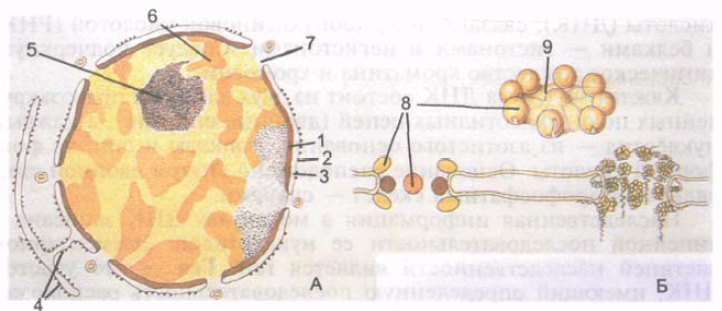
Рис. 4. Клеточное ядро (А) и комплекс ядерной поры (Б). 1 - внутренняя ядерная мембрана; 2 - наружная ядерная мембрана; 3 - пе- ринуклеарное пространство; 4 - мембраны эндоплазматической сети; 5 - ядрышко; 6 - деконденсированный хроматин; 7 - ядерная пора; 8 - гранулы ядерной поры; 9 - диафрагма ядерной поры.
В ядерной оболочке имеется множество округлых ядерных пор диаметром 50-70 нм каждая. Ядерные поры в обшей сложности занимают до 25 % от поверхности ядра. Количество пор у одного ядра достигает 3000-4000. По краям пор наружная и внутренняя мембраны соединяются одна с другой и образуют так называемое кольцо поры. Каждая пора закрыта диафрагмой, которую называют также комплексом поры. Диафрагмы пор имеют сложное строение, они образованы соединенными между собой белковыми гранулами. Через ядерные поры осуществляется избирательный транспорт крупных частиц, а также обмен веществ между ядром и цитозолем клетки.
Под ядерной оболочкой находятся нуклеоплазма (кариоплазма) (nucleoplаsma, s. karyoplаsma), имеющая гомогенное строение, и ядрышко. В нуклеоплазме неделящегося ядра, в его ядерном белковом матриксе, расположены осмиофильные гранулы (глыбки) так называемого гетерохроматина. Участки более разрыхленного хроматина, расположенные между гранулами, называют эухроматином. Разрыхленный хроматин называют также деконденсированным хроматином, в нем наиболее интенсивно протекают синтетические процессы. Во время деления клетки хроматин уплотняется, конденсируется, образует хромосомы.
Хроматин (chromatinum) неделящегося ядра и хромосомы делящегося образованы молекулами дезоксирибонуклеиновой кислоты (ДНК), связанной с рибонуклеиновой кислотой (РНК) и белками - гистонами и негистонами. Следует подчеркнуть химическое тождество хроматина и хромосом.
Каждая молекула ДНК состоит из двух длинных правозакрученных полинуклеотидных цепей (двойные спирали), а каждый нуклеотид - из азотистого основания, глюкозы и остатка фосфорной кислоты. Основание расположено внутри двойной спирали, а сахарофосфатный скелет - снаружи.
Наследственная информация в молекулах ДНК записана в линейной последовательности ее нуклеотидов. Элементарной частицей наследственности является ген. Ген - это участок ДНК, имеющий определенную последовательность расположения нуклеотидов, ответственных за синтез одного определенного специфического белка.
Молекула ДНК в ядре упакована компактно. Так, одна молекула ДНК, содержащая 1 млн нуклеотидов, при их линейном расположении заняла бы отрезок длиной всего 0,34 мм. Длина одной хромосомы человека в растянутом виде составляет около 5 см, однако в уплотненном состоянии хромосома имеет объем около 10 -15 см 3 .
Молекулы ДНК, связанные с белками-гистонами, образуют нуклеосомы, являющиеся структурными единицами хроматина. Нуклеосома имеет вид бусинки диаметром 10 нм. Каждая нуклеосома состоит из гистонов, вокруг которых закручен участок ДНК, включающий 146 пар нуклеотидов. Между нуклеосомами располагаются линейные участки ДНК, состоящие из 60 пар нуклеотидов.
Хроматин представлен фибриллами, которые образуют петли длиной около 0,4 мкм, содержащие от 20 000 до 30 000 пар нуклеотидов.
В результате уплотнения (конденсация) и закручивания (суперспециализация) дезоксирибонуклеопротеидов (ДНП) в делящемся ядре становятся видными хромосомы. Эти структуры - хромосомы (chromasоmae, от греч. chrуma - краска, soma - тело) - представляют собой удлиненные палочковидные образования, имеющие два плеча, разделенные так называемой перетяжкой - центромерой. В зависимости от расположения центромеры и взаимного расположения и длины плеч (ножек) выделяют три типа хромосом: метацентрические, имеющие примерно одинаковые плечи; субметацентрические, у которых длина плеч различная; акроцентрические, у которых одно плечо длинное, а другое - очень короткое, еле заметное. В хромосоме имеются эу- и гетерохроматиновые участки. Последние в неделящемся ядре и в ранней профазе митоза остаются компактными. Чередование эу- и гетерохроматиновых участков используют для идентификации хромосом.
Поверхность хромосом покрыта различными молекулами, главным образом рибонуклеопротеидами (РНП). В соматических клетках имеется по 2 копии каждой хромосомы, их называют гомологичными. Они одинаковые по длине, форме, строению, несут одни и те же гены, которые расположены одинаково. Особенности строения, количество и размеры хромосом называют кариотипом. Нормальный кариотип человека включает 22 пары аутосом и одну пару половых хромосом (XX или XY). Соматические клетки человека (диплоидные) имеют удвоенное число хромосом - 46. Половые клетки содержат гаплоидный (одинарный) набор - 23 хромосомы. Поэтому в половых клетках ДНК в 2 раза меньше, чем в диплоидных соматических клетках.
Ядрышко (nucleоlus), одно или несколько, выявляется во всех неделящихся клетках. Оно имеет вид интенсивно окрашивающегося округлого тельца, величина которого пропорциональна интенсивности белкового синтеза. Ядрышко состоит из электронно-плотной нуклеолонемы (от греч. пета - нить), в которой различают нитчатую (фибриллярную) часть, состоящую из множества переплетающихся нитей РНК толщиной около 5 нм, и гранулярную часть. Гранулярная (зернистая) часть образована зернами диаметром около 15 нм, представляющими собой частицы РНП - предшественников рибосомных субъединиц. Околоядрышковый хроматин внедряется в углубления нуклеолонемы. В ядрышке образуются рибосомы.
ДЕЛЕНИЕ КЛЕТОК. КЛЕТОЧНЫЙ ЦИКЛ
Рост организма происходит за счет увеличения числа клеток путем деления. Основными способами деления клеток в человеческом организме являются митоз и мейоз. Процессы, происходящие при этих способах деления клеток, протекают одинаково, однако приводят к разным результатам.
Митотическое деление клеток (митоз) приводит к увеличению числа клеток, росту организма. Таким способом обеспечивается обновление клеток при их износе, гибели. В настоящее время известно, что клетки эпидермиса живут 10-30 дней, эритроциты - до 4-5 мес. Нервные и мышечные клетки (волокна) живут в течение всей жизни человека.
У всех клеток при размножении (делении) наблюдаются изменения, укладывающиеся в рамки клеточного цикла. Клеточным циклом называют процессы, которые происходят в клетке от деления до деления или от деления до смерти (гибели) клетки. В клеточном цикле выделяют подготовку клетки к делению (интерфаза) и митоз (процесс деления клетки).
В интерфазе, которая длится примерно 20-30 ч, скорость биосинтетических процессов возрастает, увеличивается количество органелл. В это время удваивается масса клетки и всех ее структурных компонентов, в том числе центриолей.
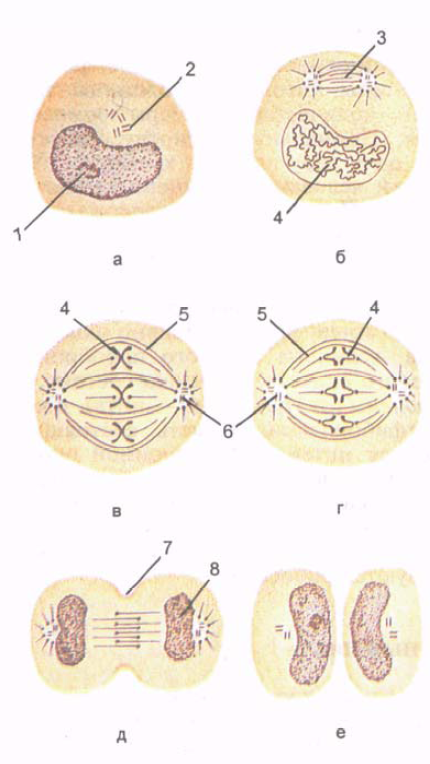
Рис. 5. Деление клетки. Стадии митоза. Показаны образование хромосом, формирование веретена деления и равномерное распределение хромосом и центриолей по двум дочерним клеткам.
А - интерфаза; Б - профаза; В - метафаза; Г - анафаза; Д - телофаза; Е - поздняя те- лофаза. 1 - ядро; 2 - клеточный центр (центриоль); 3 - веретено деления клетки; 4 - хромосомы; 5 - непрерывные микротрубочки; 6 - клеточный центр; 7 - борозда деления клетки; 8 - формирование ядра.
Происходит репликация (повторение, удвоение) молекул нуклеиновых кислот. Этот процесс передачи генетической информации, хранящейся в родительской ДНК, путем точного ее воспроизведения в дочерних клетках. Родительская цепь ДНК служит матрицей для синтеза дочерних ДНК. В итоге репликации каждая из двух дочерних молекул ДНК состоит из одной старой и одной новой цепей. В период подготовки к митозу в клетке синтезируются белки, необходимые для деления клетки. К концу интерфазы хроматин в ядре конденсирован.
Митоз (mitоsis; от греч. mitоs - нить) представляет собой период, когда материнская клетка делится на две дочерние (рис. 5). Митотическое деление клеток обеспечивает равномерное распределение структур клетки, ее ядерного вещества - хроматина - между двумя дочерними клетками. Длительность митоза - от 30 мин до 3 ч. Митоз подразделяют на профазу, метафазу, анафазу, телофазу.
В профазе постепенно распадается ядрышко, центрио- ли расходятся к полюсам клетки. Микротрубочки центриолей направлены к экватору, а в области экватора они перекрывают друг друга.
В метафазе разрушается ядерная оболочка, хромосомные нити направляются к полюсам, сохраняя связь с экватори
альной областью клетки. Структуры эндоплазматической сети и комплекса Гольджи распадаются на мелкие пузырьки (везикулы), которые вместе с митохондриями распределяются в обе половины делящейся клетки. В конце метафазы каждая хромосома начинает расщепляться продольной щелью на две новые дочерние хромосомы.
В анафазе хромосомы отделяются друг от друга и расходятся к полюсам клетки со скоростью до 0,5 мкм/мин. В конце анафазы плазматическая мембрана впячивается по экватору клетки перпендикулярно ее продольной оси, образуя борозду деления.
В телофазе хромосомы, разошедшиеся к полюсам клетки, деконденсируются, переходят в хроматин и начинается транскрипция (продукция) РНК. Образуются ядерная оболочка, ядрышко, быстро формируются мембранные структуры будущих дочерних клеток. На поверхности клетки, по ее экватору, перетяжка углубляется, клетка разделяется на две дочерние клетки.
Благодаря митотическому делению дочерние клетки получают набор хромосом, идентичный материнскому. Митоз обеспечивает генетическую стабильность, увеличение числа клеток и, следовательно, рост организма, а также процессы регенерации.
Мейоз (от греч. meiоsis - уменьшение) наблюдается у половых клеток. В результате деления этих клеток образуются новые клетки с одинарным (гаплоидным) набором хромосом, что важно для передачи генетической информации. При слиянии одной половой клетки с клеткой противоположного пола (при оплодотворении) набор хромосом удваивается, становится полным, двойным (диплоидным). В образовавшейся после слияния половых клеток диплоидной (двуядерной) зиготе оказывается два набора одинаковых (гомологичных) хромосом. Каждая пара гомологичных хромосом диплоидного организма (зиготы) происходит из ядра яйцеклетки и из ядра сперматозоида.
В результате мейоза половых клеток в зрелом организме в каждой дочерней клетке оказывается лишь по одной из всех пар гомологичных хромосом исходных клеток. Это становится возможным потому, что при мейозе происходят лишь репликация ДНК и два последовательных деления ядер. В результате из одной диплоидной клетки образуются две гаплоидные клетки. В каждой из таких дочерних клеток содержится вдвое меньше хромосом (23), чем в ядре материнской клетки (46). В результате мейоза гаплоидные половые клетки имеют не только уменьшенное вдвое число хромосом, но иное расположение генов в хромосомах. Поэтому новый организм несет не просто сумму признаков своих родителей, но и собственные (индивидуальные) черты.
Вопросы для повторения и самоконтроля
1. Из каких элементов состоит ядро клетки? Какие функции оно выполняет?
2. Расскажите о строении молекул ДНК.
3. Дайте морфологическую характеристику хромосом, приведите их классификацию.
4. Что собой представляет клеточный цикл, какие периоды (фазы) выделяют в этом цикле?
5. Что такое мейоз, чем он отличается от митоза?