La evidencia paleontológica sugiere que algunas plantas pertenecientes al grupo Eucaesalpinioideae fueron las leguminosas más antiguas que tenían nódulos.
En especies modernas de plantas leguminosas, se han encontrado nódulos en las raíces de muchos representantes de la familia Papilijonaceae.
Los representantes filogenéticamente más primitivos de familias como Caesalpiniaceae Mimosaceae no forman nódulos en la mayoría de los casos.
De las 13.000 especies (550 géneros) de plantas leguminosas, la presencia de nódulos hasta ahora se ha identificado sólo en aproximadamente 1300 especies (243 géneros). Esto incluye principalmente especies de plantas utilizadas en la agricultura (más de 200).
Habiendo formado nódulos, las leguminosas adquieren la capacidad de absorber nitrógeno atmosférico. Sin embargo, pueden alimentarse de formas unidas de nitrógeno: sales de amonio y ácido nítrico. Solo una planta, Hedysarum coronarium, asimila solo nitrógeno molecular. Por lo tanto, sin nódulos en la naturaleza, esta planta no se produce.
Las bacterias del nódulo suministran nitrógeno a la planta leguminosa, que se fija del aire. Las plantas, a su vez, suministran a las bacterias los productos del metabolismo de los carbohidratos y las sales minerales que necesitan para su crecimiento y desarrollo.
En 1866, el famoso botánico y científico del suelo M.S. Voronin vio los "terneros" más pequeños en los nódulos de las raíces de las plantas leguminosas. Voronin presentó suposiciones audaces para ese momento: vinculó la formación de nódulos con la actividad de las bacterias y el aumento de la división de las células del tejido de la raíz con la reacción de la planta a las bacterias que penetraron en la raíz.
20 años después, el científico holandés Beijerinck aisló bacterias de los nódulos de guisantes, arvejas, chiny, frijoles, seradella y piruletas y estudió sus propiedades, probando la capacidad de infectar plantas y provocar la formación de nódulos. Llamó a estos microorganismos Bacillus radicicola. Dado que las bacterias que forman esporas pertenecen al género Bacillus, y las bacterias de los nódulos carecen de esta capacidad, A. Prazhmovsky las rebautizó como Bacterium radicicola. B. Frank propuso un nombre genérico más exitoso para la bacteria del nódulo: Rhizobium (del griego rhizo - raíz, bio - vida; vida en las raíces). Este nombre ha echado raíces y todavía se usa en la literatura.
Para designar una especie de bacteria nódulo se acostumbra añadir al nombre genérico Rhizobium un término correspondiente al nombre en latín de la especie vegetal de cuyos nódulos se aíslan y sobre la que pueden formar nódulos. Por ejemplo, Rhizobium trifolii - bacterias del nódulo del trébol, Rhizobium lupini - bacterias del nódulo del lupino, etc. En los casos en que las bacterias del nódulo pueden formar nódulos en las raíces de diferentes tipos de plantas leguminosas, es decir, causar la llamada infección cruzada, el nombre de la especie es, por así decirlo, colectivo: refleja esta "infección cruzada". capacidad. Por ejemplo, Rhizobium leguminosarum - bacterias nodulares de guisantes (Pisum), lentejas (Lens), filas (Lathyrus).
Morfología y fisiología de las bacterias del nódulo: las bacterias del nódulo se caracterizan por una sorprendente variedad de formas: polimorfismo. Muchos investigadores llamaron la atención sobre esto al estudiar bacterias de nódulos en cultivo puro en condiciones de laboratorio y suelo. Las bacterias del nódulo pueden tener forma de bastón y forma ovalada. Entre estas bacterias también hay formas filtrables, formas L, cocoides inmóviles y organismos móviles.
Las bacterias de nódulos jóvenes en cultivo puro en medios nutritivos generalmente tienen forma de varilla, el tamaño de las varillas es de aproximadamente 0.5-0.9 X 1.2-3.0 micrones, móviles, se multiplican por división. En las células en forma de varilla de las bacterias del nódulo del trébol, se observa división por ligamiento. Con la edad, las células en forma de bastón pueden pasar a brotar. Según Gram, las células se tiñen negativamente, su estructura ultrafina es típica de las bacterias Gram-negativas.
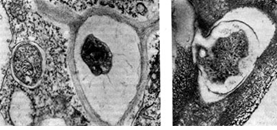
Con el envejecimiento, las bacterias de los nódulos pierden su movilidad y pasan al estado de los llamados bastones ceñidos. Obtuvieron este nombre debido a la alternancia de secciones densas y sueltas de protoplasma en las células. La estriación de las células se detecta bien cuando se observa bajo un microscopio óptico después del tratamiento de las células con tintes de anilina. Las secciones densas de protoplasma (faja) se tiñen peor que los espacios entre ellas. En un microscopio fluorescente, las bandas son de color verde claro, los espacios entre ellas no brillan y se ven oscuros Las bandas pueden estar ubicadas en el medio de la celda o en los extremos. El anillado de células también es visible en patrones de difracción de electrones, si la preparación no se trata con agentes de contraste antes de verla. Probablemente, con la edad, la célula bacteriana se va llenando de inclusiones grasas que no perciben el color y, como consecuencia, hacen que la célula se estrie. La etapa de "bastones en cinturón" precede a la etapa de formación de bacteroides, células de forma irregular: engrosadas, ramificadas, esféricas, en forma de pera y en forma de matraz que las células bacterianas en forma de bastón que se encuentran en los tejidos de los nódulos.
Los bacteroides contienen más gránulos de volutina y se caracterizan por un mayor contenido de glucógeno y grasa que las células en forma de bastón. Los bacteroides cultivados en medios nutritivos artificiales y formados en tejidos de nódulos son fisiológicamente del mismo tipo. Se cree que los bacteroides son formas de bacterias con un proceso de división incompleto. Con la división celular incompleta de las bacterias del nódulo, surgen formas de bacteroides que se ramifican dicotómicamente. El número de bacteroides aumenta con el envejecimiento del cultivo; su aparición se ve facilitada por el agotamiento del medio nutritivo, la acumulación de productos metabólicos y la introducción de alcaloides en el medio.
Arroz. 3
En cultivos antiguos (de dos meses) de bacterias de nódulos, utilizando un microscopio electrónico, es posible identificar formaciones esféricas claramente definidas en muchas células: artrosporas. Su número en las celdas varía de 1 a 5.
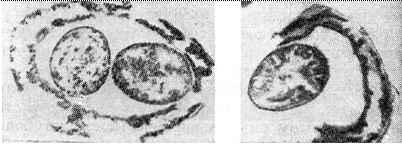
Arroz. 4
Las bacterias del nódulo de varios tipos de plantas leguminosas crecen a diferentes velocidades en medios nutritivos. Las bacterias de crecimiento rápido incluyen rizobios de guisantes, tréboles, alfalfa, frijoles forrajeros, arvejas, lentejas, chiny, trébol de olor, fenogreco, frijoles, garbanzos y patas de pájaro; a las de crecimiento lento: bacterias de nódulos de lupino, soja, maní, seradella, frijol mungo, caupí, esparceta, aulaga. Se pueden obtener colonias completamente formadas de cultivos de crecimiento rápido en el 3er - 4to día de incubación, colonias de cultivos de crecimiento lento - en el 7 - 8.
Las bacterias de los nódulos de rápido crecimiento se caracterizan por una disposición peritrica de los flagelos, mientras que las bacterias de crecimiento lento son monotriquiales.
Además de los flagelos, en las células de bacterias del nódulo se forman excrecencias filamentosas y similares a perlas cuando se cultivan en medios líquidos.Su longitud alcanza las 8-10 micras. Suelen estar situados en la superficie de la célula peritricialmente, contienen de 4 a 10 o más por célula.
Las colonias de bacterias de nódulos de rápido crecimiento tienen el color de la leche horneada, a menudo translúcidas, viscosas, con bordes lisos, moderadamente convexas y eventualmente crecen en la superficie del medio de agar. Las colonias de bacterias de crecimiento lento son más convexas, pequeñas, secas, densas y, por regla general, no crecen en la superficie del medio. El moco producido por las bacterias del nódulo es un compuesto complejo del tipo polisacárido, que incluye hexosas, pentosas y ácidos urónicos.
Las bacterias del nódulo son microaerófilas (se desarrollan con pequeñas cantidades de oxígeno en el ambiente), sin embargo, prefieren condiciones aeróbicas.
Las bacterias de los nódulos utilizan carbohidratos y ácidos orgánicos como fuente de carbono en medios nutritivos, y varios compuestos minerales y orgánicos que contienen nitrógeno como fuente de nitrógeno. Cuando se cultiva en medios con un alto contenido de sustancias que contienen nitrógeno, las bacterias de los nódulos pueden perder su capacidad de penetrar en la planta y formar nódulos. Por lo tanto, las bacterias del nódulo generalmente se cultivan en extractos de plantas (caldo de frijoles, guisantes) o extractos de suelo. El fósforo necesario para el desarrollo puede ser obtenido por bacterias de nódulos a partir de compuestos minerales y orgánicos que contienen fósforo; Los compuestos minerales pueden servir como fuente de calcio, potasio y otros elementos minerales.
Para suprimir la microflora saprofita extraña cuando se aíslan bacterias de nódulos de nódulos o directamente del suelo, se recomiendan medios nutritivos con la adición de cristal violeta, tanino o antibióticos.
Para el desarrollo de la mayoría de los cultivos de bacterias de nódulos, se requiere una temperatura óptima en el rango de 24-26 °. A 0° y 37°C se detiene el crecimiento. Por lo general, los cultivos de bacterias de nódulos se almacenan en condiciones de laboratorio a bajas temperaturas (2-4 °C).
Muchos tipos de bacterias del nódulo son capaces de sintetizar vitaminas B, así como sustancias de crecimiento como la heteroauxina (ácido beta-indolacético).
Todas las bacterias del nódulo son aproximadamente igualmente resistentes a una reacción alcalina del medio (pH = 8,0), pero desigualmente sensibles a una reacción ácida.
Especificidad, virulencia, competitividad y actividad de las bacterias del nódulo: el concepto de especificidad de las bacterias del nódulo es colectivo. Caracteriza la capacidad de las bacterias para formar nódulos en las plantas. Si hablamos de bacterias de nódulos en general, entonces para ellos la formación de nódulos solo en un grupo de plantas leguminosas ya es específica en sí misma: tienen selectividad por las plantas leguminosas.
Sin embargo, si consideramos cultivos individuales de bacterias de nódulos, resulta que entre ellos hay aquellos que son capaces de infectar solo un determinado grupo de plantas leguminosas, a veces más grande, a veces más pequeño, y en este sentido, la especificidad de las bacterias de nódulos es una capacidad selectiva en relación con la planta huésped. La especificidad de las bacterias del nódulo puede ser limitada (las bacterias del nódulo del trébol infectan solo a un grupo de tréboles, especificidad de especie, y las bacterias del nódulo de lupino pueden incluso caracterizarse por especificidad de variedad, infectan solo variedades de lupino con alcaloides o libres de alcaloides). Con una amplia especificidad, las bacterias del nódulo del guisante pueden infectar plantas de guisante, barbilla y frijol, y las bacterias del nódulo del guisante y frijol pueden infectar plantas de guisante, es decir, todas se caracterizan por la capacidad de "infectarse de forma cruzada". La especificidad de las bacterias del nódulo es la base de su clasificación.
La especificidad de las bacterias del nódulo surgió como resultado de su adaptación a largo plazo a una planta oa un grupo de ellas y la transmisión genética de esta propiedad. En este sentido, también existe una adaptabilidad diferente de las bacterias del nódulo a las plantas dentro del grupo de infecciones cruzadas. Por lo tanto, las bacterias del nódulo de la alfalfa pueden formar nódulos en el trébol de olor. Sin embargo, están más adaptadas a la alfalfa, y las bacterias del trébol de olor están más adaptadas al trébol de olor.
En el proceso de infección del sistema radicular de las plantas leguminosas con bacterias del nódulo, la virulencia de los microorganismos es de gran importancia. Si la especificidad determina el espectro de acción de las bacterias, entonces la virulencia de las bacterias del nódulo caracteriza la actividad de su acción dentro de este espectro. La virulencia se refiere a la capacidad de las bacterias del nódulo para penetrar en el tejido de la raíz, multiplicarse allí y causar la formación de nódulos.
Un papel importante lo juega no solo la capacidad misma de penetrar en las raíces de la planta, sino también la velocidad de esta penetración.
Para determinar la virulencia de una cepa de bacterias de nódulos, es necesario establecer su capacidad para provocar la formación de nódulos. El criterio de virulencia de cualquier cepa puede ser el mínimo número de bacterias que proporcione una infección más vigorosa de las raíces en comparación con otras cepas, culminando en la formación de nódulos.

Arroz. 5
En el suelo, en presencia de otras cepas, la cepa más virulenta no siempre infectará primero a la planta. En este caso, se debe tener en cuenta su capacidad competitiva, que muchas veces enmascara la propiedad de virulencia en condiciones naturales.
Es necesario que las cepas virulentas también tengan competitividad, es decir, puedan competir con éxito no solo con representantes de la microflora saprofita local, sino también con otras cepas de bacterias nodulares. Un indicador de la competitividad de una cepa es el número de nódulos formados por ella como porcentaje del número total de nódulos en las raíces de las plantas.
Una propiedad importante de las bacterias del nódulo es su actividad (eficiencia), es decir, la capacidad en simbiosis con leguminosas para asimilar nitrógeno molecular y satisfacer en él las necesidades de la planta huésped. Dependiendo de la medida en que las bacterias del nódulo contribuyan al aumento del rendimiento de las leguminosas, generalmente se dividen en activas (eficaces), inactivas (ineficaces) e inactivas (ineficaces).
Una cepa de bacterias inactivas para una planta huésped en simbiosis con otra especie de planta leguminosa puede ser muy eficaz. Por tanto, a la hora de caracterizar una cepa en cuanto a su eficacia, siempre se debe indicar en relación a qué especie vegetal huésped se manifiesta su efecto.
La actividad de las bacterias del nódulo no es su propiedad permanente. A menudo, en la práctica de laboratorio, hay una pérdida de actividad en cultivos de bacterias de nódulos. En este caso, se pierde la actividad de todo el cultivo o aparecen células individuales con baja actividad. Se produce una disminución en la actividad de las bacterias del nódulo en presencia de ciertos antibióticos, aminoácidos. Una de las razones de la pérdida de actividad de las bacterias del nódulo puede ser la influencia del fago. Al pasar, es decir, pasar repetidamente bacterias a través de la planta huésped (adaptación a una especie de planta en particular), es posible obtener cepas efectivas de las ineficaces.
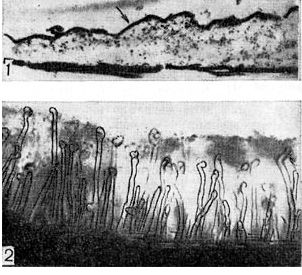
Arroz. 6 - Una capa de sustancia granular en la superficie del pelo de la raíz (1) durante el examen con microscopio electrónico (según P. Dart, F. Mercer, aumento X 30.000) y una capa viscosa de zoogley (2), en la que se encuentran pelos de la raíz del trébol. sumergido, bajo un microscopio de luz (zoom x 80)
La exposición a los rayos gamma permite obtener cepas con mayor eficiencia. Hay casos conocidos de aparición de radiomutantes altamente activos de bacterias del nódulo de alfalfa a partir de una cepa inactiva. El uso de radiación ionizante, que tiene un efecto directo sobre el cambio en las características genéticas de la célula, con toda probabilidad, puede ser una técnica prometedora en la selección de cepas altamente activas de bacterias nodulares.
Infección de una planta leguminosa con bacterias de nódulos: para garantizar el proceso normal de infección del sistema radicular con bacterias de nódulos, es necesario tener una cantidad bastante grande de células bacterianas viables en la zona de la raíz.
Durante el desarrollo del sistema de raíces de una planta leguminosa, las secreciones de las raíces estimulan la multiplicación de bacterias nodulares en la superficie de la raíz. Los productos de destrucción de las cofias radiculares y los pelos también juegan un papel importante en proporcionar un sustrato adecuado a las bacterias del nódulo.
En la superficie de la raíz hay una capa de sustancia mucosa (matriz), que se forma independientemente de la presencia de bacterias en la rizosfera. Esta capa es claramente visible cuando se examina bajo un microscopio óptico de luz. Las bacterias del nódulo después de la inoculación generalmente se precipitan hacia esta capa y se acumulan debido al efecto estimulante de la raíz, que se manifiesta incluso a una distancia de hasta 30 mm.
Hay una serie de hipótesis sobre el mecanismo de penetración de las bacterias del nódulo en la raíz de la planta. Los más interesantes de ellos son los siguientes. Los autores de una de las hipótesis afirman que las bacterias del nódulo penetran en la raíz a través del daño al tejido epidérmico y cortical (especialmente en los lugares donde se ramifican las raíces laterales). Esta hipótesis se planteó sobre la base de la investigación de Bril (1888), quien provocó la formación de nódulos en plantas leguminosas perforando las raíces con una aguja previamente sumergida en una suspensión de bacterias nodulares. Como caso especial, tal ruta de implementación es bastante real. Por ejemplo, en el maní, los nódulos se ubican predominantemente en las axilas de las ramas de la raíz, lo que sugiere la penetración de bacterias del nódulo en la raíz a través de espacios durante la germinación de las raíces laterales.
Una hipótesis interesante y no infundada es la penetración de las bacterias del nódulo en el tejido de la raíz a través de los pelos de la raíz. La mayoría de los investigadores reconocen el paso de las bacterias del nódulo a través de los pelos radiculares.
Es posible que las bacterias del nódulo puedan penetrar en la raíz a través de las células epidérmicas de las puntas de las raíces jóvenes. Según Prazhmovsky (1889), las bacterias pueden penetrar en la raíz solo a través de la pared celular joven (de los pelos de la raíz o de las células epidérmicas) y son completamente incapaces de superar la capa corchosa o alterada químicamente de la corteza. Esto puede explicar que los nódulos se desarrollen generalmente en las secciones jóvenes de la raíz principal y en las raíces laterales emergentes.
Recientemente, la hipótesis de las auxinas ha recibido una gran popularidad. Los autores de esta hipótesis creen que las bacterias del nódulo penetran en la raíz estimulando la síntesis de ácido p-indolacético (heteroauxina) a partir del triptófano, que siempre está presente en las secreciones de las raíces de las plantas. La presencia de heteroauxina está asociada con la curvatura de los pelos radiculares, lo que generalmente se observa cuando el sistema radicular está infectado con bacterias del nódulo.
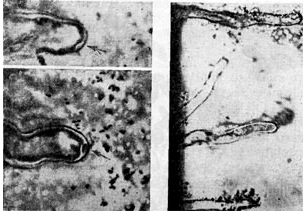
Arroz. 7
La fuente de ácido beta-indolilacético en el momento de la infección de la planta, obviamente, no son solo las plantas que secretan triptófano a través del sistema radicular, que muchos tipos de bacterias, incluidos los nódulos de la raíz, pueden convertir en ácido beta-indolilacético. Las mismas bacterias del nódulo, y posiblemente otros tipos de microorganismos del suelo que viven en la zona de la raíz, también pueden participar en la síntesis de heteroauxina.
La acción de la heteroauxina no es específica y provoca la curvatura de los pelos de la raíz en varias especies de plantas, no solo en las leguminosas. Al mismo tiempo, las bacterias de los nódulos provocan la curvatura de los pelos radiculares solo en las plantas leguminosas, mientras que exhiben una selectividad bastante significativa. Si el efecto en consideración fuera determinado solo por el ácido β-indolacético, entonces tal especificidad no existiría.Además, la naturaleza de los cambios en los pelos radiculares bajo la influencia de las bacterias del nódulo es algo diferente que bajo la influencia de la heteroauxina.
También se debe tener en cuenta que, en algunos casos, los pelos de raíz no curvados están expuestos a infecciones. Las observaciones muestran que en la alfalfa y los guisantes, el 60-70% de los pelos de la raíz están torcidos y torcidos, y en el trébol, alrededor del 50%. En algunas especies de trébol, esta reacción se observa en no más de 1/4 de los cabellos infectados. En la reacción de curvatura, obviamente, el estado del cabello radicular es de gran importancia. Los pelos de raíz en crecimiento son más sensibles a la acción de sustancias producidas por bacterias.
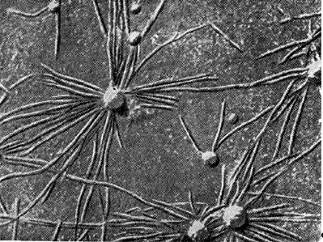
Arroz. 8
leguminosa bacteria nitrogenada nódulo
Cabe señalar otra hipótesis, según la cual las bacterias del nódulo ingresan a la raíz durante la formación de una protuberancia en forma de dedo en la superficie del cabello de la raíz. Un patrón de difracción de electrones de una sección de cabello de raíz que confirma esta hipótesis muestra un cabello de raíz doblado en forma de mango de paraguas, en cuya curva hay una acumulación de bacterias nodulares. Las bacterias del nódulo son como si fueran atraídas (tragadas) por el vello de la raíz (similar a la pinocitosis).
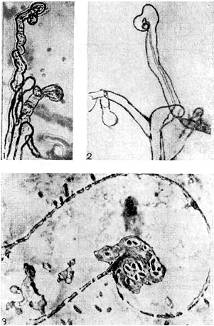
Arroz. 9 - Curvatura de los pelos radiculares de plantas leguminosas en presencia de bacterias nodulares: 1,2 - estola, x 120; 3 - Sección ultrafina a través de la raíz del cabello. Aumentó X 10.000 (según K. Salman y G. Fareus)
El proceso de introducción de las bacterias del nódulo en el tejido radicular es el mismo en todos los tipos de plantas leguminosas y consta de dos fases. En la primera fase se produce la infección de los pelos radiculares. En la segunda fase, el proceso de formación de nódulos avanza intensamente. La duración de las fases es diferente en diferentes especies de plantas: en Trifolium fragiferum la primera fase dura 6 días, en Trifolium nigrescens - 3 días. En algunos casos es muy difícil detectar los límites entre fases. La introducción más intensa de bacterias del nódulo en los pelos radiculares ocurre en las primeras etapas del desarrollo de la planta. La segunda fase termina durante la formación masiva de nódulos. A menudo, la penetración de las bacterias de los nódulos en los pelos de la raíz continúa incluso después de que se hayan formado los nódulos en las raíces. Este llamado exceso o infección adicional se produce porque la infección de los pelos no se detiene durante mucho tiempo. En las últimas etapas de la infección, los nódulos generalmente se ubican más abajo a lo largo de la raíz.
Habiendo penetrado en la raíz (a través del cabello de la raíz, células epidérmicas, lugares dañados por la raíz), las bacterias del nódulo se mueven hacia los tejidos de la raíz de la planta. Más fácilmente, las bacterias pasan a través de los espacios intercelulares.
Ya sea una sola célula o un grupo de células bacterianas pueden penetrar en el tejido de la raíz. Si se ha introducido una célula separada, puede continuar moviéndose a través del tejido como una sola célula. La forma de infección de la raíz por células individuales es característica de las plantas de lupino.
Sin embargo, en la mayoría de los casos, la célula invasora, que se multiplica activamente, forma los llamados hilos infecciosos (o tji infecciosos) y, ya en forma de tales hilos, se mueve hacia los tejidos vegetales.
El término "hilo de infección" surgió a partir del estudio del proceso de infección en un microscopio óptico. A partir del trabajo de Beijerinck, el hilo de infección comenzó a ser considerado como una masa viscosa parecida a una hifa con
De acuerdo con el método de formación, los nódulos de las plantas leguminosas se dividen en dos tipos: 1er tipo: los nódulos surgen durante la división de las células del periciclo (capa de la raíz), generalmente ubicado contra el protoxilema (primero en el tiempo para la formación de vasos ) - tipo endógeno de formación de nódulos; Tipo 2: los nódulos se originan en la corteza de la raíz como resultado de la introducción del patógeno en las células parenquimatosas de la corteza y el endodermo (la capa interna de la corteza primaria), un tipo exógeno de formación de nódulos.
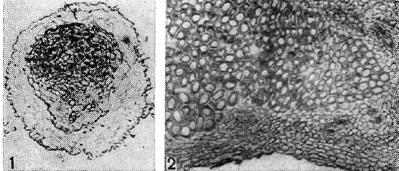
Arroz. 10 - Secciones a través del tejido del nódulo de las plantas leguminosas: 1 - sección transversal a través del nódulo de los guisantes. El comienzo de la diferenciación del tejido vascular en el nódulo (según G. Bond); d - sección longitudinal a través del nódulo del rango. Aumentó x90
En la naturaleza, predomina este último tipo. Los tejidos del cilindro central de la raíz participan solo en la formación del sistema vascular de nódulos de tipo endógeno y exógeno.
La diferencia en la naturaleza de la formación de nódulos y raíces laterales se observa especialmente claramente en Seradella, ya que el tejido cortical de la raíz principal de esta planta, el sitio de los primeros nódulos, consiste en una capa relativamente pequeña de células y los nódulos se vuelven muy visibles. rápidamente después de la infección de la raíz con bacterias. Primero forman protuberancias aplanadas en la raíz, lo que permite distinguirlas de las protuberancias cónicas de las raíces laterales. Los nódulos se diferencian de las raíces laterales en una serie de características anatómicas: la ausencia de un cilindro central, cubiertas de raíces y epidermis, y la presencia de una capa significativa de corteza que cubre el nódulo.
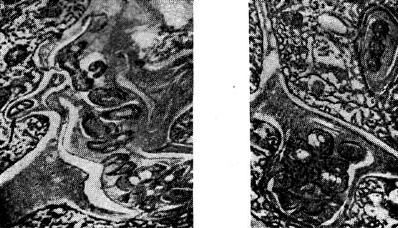
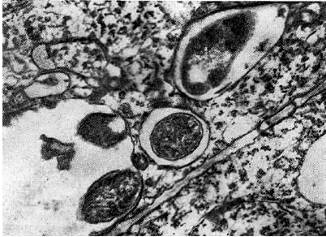
Arroz. once - Hilos de infección intercelular en forma de bolsillo (derecha) y antorcha (izquierda) con nódulo bacteriano. La flecha muestra una célula bacteriana preparándose para ingresar al citoplasma de una célula vegetal. Aumentó 20.000x20.000
La formación de nódulos de las leguminosas ocurre durante el período en que la raíz todavía tiene una estructura primaria. Comienza con la división de células corticales ubicadas a una distancia de 2-3 capas de los extremos de los hilos infecciosos. Las capas de la corteza, penetradas por hilos infecciosos, permanecen sin cambios. Al mismo tiempo, en seradella, la división de las células corticales ocurre directamente debajo del cabello de la raíz infectada, mientras que en los guisantes, la división celular se observa solo en la penúltima capa de la corteza.
La división con la formación de una estructura de tejido radial continúa hacia las células del núcleo interno. Ocurre sin una dirección específica, al azar, y como resultado de esto, surge un meristema (un sistema de tejidos educativos) del nódulo, formado por pequeñas células granulares.
Las células divididas de la corteza cambian: los núcleos se redondean y aumentan de tamaño, especialmente los nucléolos. Después de la mitosis, los núcleos se dispersan y, sin tomar su forma original, comienzan a dividirse nuevamente.
Aparece un meristema secundario. Pronto, en el endodermo y el periciclo, aparecen signos de división incipiente, que en las primeras células externas se produce principalmente por tabiques tangenciales. Esta división finalmente se extiende al complejo meristemático común, cuyas pequeñas células se alargan, las vacuolas desaparecen y el núcleo llena la mayor parte de la célula. Se forma el llamado nódulo primario, en el plasma de las células de las cuales las bacterias del nódulo están ausentes, ya que en esta etapa todavía se encuentran dentro de los hilos de infección. Mientras se forma el nódulo primario, los hilos de infección se ramifican muchas veces y pueden pasar entre las células (intercelularmente) oa través de las células (intracelularmente) e introducir bacterias.
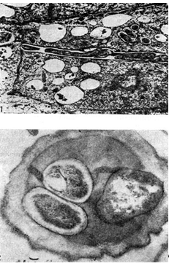
Arroz. 12 - Hilos infecciosos intracelulares con bacterias nodulares: 1 - sección longitudinal (según D. Goodchild y F. Bergersen), 2 - sección transversal. Llevado, x 35000
Los hilos infecciosos intercelulares debido a la reproducción activa de bacterias nodulares en ellos a menudo adquieren una forma extraña: se forman en forma de bolsillos (divertículos) o antorchas.
El proceso de movimiento de los hilos de infección de una célula a otra no está del todo claro. Aparentemente, los hilos infecciosos, como cree el microbiólogo canadiense D. Jordan, vagan en forma de hilos mucosos desnudos en los espacios intercelulares del tejido vegetal hasta que, por algunas razones aún inexplicables, comienzan a invaginarse en el citoplasma de las células adyacentes.
En algunos casos, la invaginación del hilo de infección ocurre en uno, en algunos casos, en cada celda vecina. A través de estas cavidades tubulares invaginadas (divertículos), fluye el contenido del hilo encerrado en moco. El crecimiento más activo de hilos infecciosos generalmente ocurre cerca del núcleo de la célula vegetal. La penetración del hilo va acompañada del movimiento del núcleo, que se desplaza hacia el sitio de la infección, aumenta, cambia de forma y degenera. Un cuadro similar se observa en una infección fúngica, cuando el núcleo a menudo se precipita hacia las hifas invasoras, es atraído por el daño hacia el lugar de mayor actividad fisiológica, se acerca al hilo, se hincha y colapsa. Aparentemente, esto es característico de la respuesta de la planta a la infección.
En las plantas anuales, los hilos infecciosos suelen aparecer durante el primer período de infección de la raíz, en las plantas perennes, durante un largo período de desarrollo.
Arroz. 13
Las bacterias pueden liberarse del hilo de infección en diferentes momentos y de diferentes maneras. La salida de bacterias, por regla general, es un proceso muy largo, especialmente en plantas perennes. Por lo general, la liberación de bacterias desde el hilo de infección al citoplasma de la planta huésped está asociada con la presión interna resultante de la multiplicación intensiva de bacterias en el hilo y su excreción de moco. A veces, las bacterias se escapan del hilo en grupos rodeados por la mucosidad del hilo de infección. en forma de vesículas (formaciones vesiculares). Como las vesículas no tienen membranas, la salida de bacterias de ellas es muy sencilla. Las bacterias del nódulo también pueden ingresar a las células vegetales individualmente desde los espacios intercelulares.
Las bacterias del nódulo que han surgido del hilo de infección continúan multiplicándose en el tejido huésped. Su reproducción durante este período se produce por división de constricción. La mayor parte de las bacterias se multiplican en el citoplasma de la célula y no en el hilo de infección. Las células infectadas dan lugar al futuro tejido bacteroide.
Llenas de células de bacterias de nódulos que se multiplican rápidamente, las células vegetales comienzan a dividirse intensamente. En el momento de la división mitótica de las células infectadas, las bacterias del nódulo pueden acumularse en dos polos opuestos de la célula madre y entrar pasivamente en las células hijas. Cada una de las células no infectadas está bajo un fuerte efecto estimulante de las bacterias del nódulo y, como resultado, también se divide. Gracias a esta vigorosa división mitótica de las células meristemáticas, las bacterias del nódulo se propagan a los tejidos del nódulo y aumenta el volumen de la región bacteroide.
El tejido infectado, que consiste en células densamente tendidas y en división activa, primero tiene la forma de un cono truncado. Posteriormente, debido al crecimiento paulatino de este cono ya la simultánea división y desarrollo de las células meristemáticas, el tejido del nódulo crece perdiendo su forma de cono.
Así, el nódulo aumenta primero como resultado de la división radial y tangencial de las células centrales, y luego debido a un aumento en su tamaño y división simultánea. Una vez que las células vegetales están completamente llenas de bacterias, la mitosis se detiene. Sin embargo, las células continúan aumentando de tamaño y, a menudo, están muy alargadas. Su tamaño es varias veces mayor que el de las células vegetales no infectadas, que se ubican entre ellas en la zona bacteroide del nódulo.
La conexión de un nódulo joven con la raíz de una planta leguminosa se lleva a cabo gracias a haces vasculares y fibrosos. Por primera vez, los haces fibrosos vasculares fueron observados por M.S. Voronin (1866). El tiempo de aparición del sistema vascular en los nódulos de varios tipos de plantas leguminosas es diferente. Así, en los nódulos de soja, el comienzo del desarrollo de los haces vasculares coincide con el momento de penetración de las bacterias del nódulo en dos capas del parénquima de la vaca. Con el crecimiento del nódulo, el sistema conductor crece, se ramifica y rodea la región del bacteroide.
Paralelamente al proceso de diferenciación del sistema vascular, se forma el endodermo del nódulo a partir de la capa exterior del nódulo primario. Luego, el nódulo se redondea, su capa de células periféricas está rodeada por la corteza del nódulo.
La epidermis de la raíz se rompe y el nódulo continúa desarrollándose y aumentando de tamaño.
Utilizando un microscopio óptico en secciones longitudinales de nódulos maduros, se suelen distinguir claramente 4 zonas características de diferenciación tisular: corteza, meristemo, zona bacteroide y sistema vascular. Todos los tejidos de los nódulos se diferencian en una secuencia acrópeta a medida que el meristemo inicia nuevas células.
Arroz. 14
Corteza de nódulo: la cubierta del nódulo, que realiza una función protectora. La corteza consta de varias filas de células parenquimatosas no infectadas, cuyo tamaño y características son diferentes en las diferentes leguminosas. Muy a menudo, las células de la corteza tienen una forma alargada y son más grandes en comparación con otras células del nódulo.
Arroz. 15
En la corteza de los nódulos de especies leñosas perennes, a menudo se encuentran células con membranas corchosas que contienen resinas, tanino y taninos.
El meristema del nódulo se encuentra debajo de las células de la corteza y es una zona de células no infectadas que se dividen intensamente. El meristema del nódulo se caracteriza por células pequeñas de paredes delgadas y de forma irregular densamente espaciadas, sin espacios intercelulares. Las células meristemáticas del nódulo son similares a las células de otros tipos de tejido meristemático (parte superior de la raíz, parte superior del tallo). Las células del meristemo de los nódulos contienen un citoplasma denso y finamente granular con ribosomas, cuerpos de Golgi, protoplástidos, mitocondrias y otras estructuras. Hay pequeñas vacuolas. En el centro del citoplasma hay un gran núcleo con una membrana nuclear, poros y un nucléolo claramente definido. Las funciones de las células meristemáticas son formar células de la corteza del nódulo, la región bacteroide y el sistema vascular. Dependiendo de la ubicación del meristemo, los nódulos tienen una variedad de formas: esféricas (guisantes, frijoles, seradella, cacahuetes) o cilíndricas (alfalfa, arveja, rancio, acacia, trébol). El meristemo, ubicado en áreas separadas a lo largo de la periferia del nódulo, conduce a la formación de nódulos en forma de manguito en lupino.
El meristema del nódulo funciona durante mucho tiempo, incluso durante la necrosis del nódulo, cuando ya están llenos de masa bacterioide lisante y células vegetales destruidas.
La zona bacteroide de un nódulo ocupa su parte central y constituye del 16 al 50% de la masa seca total de los nódulos. En el primer período de formación de nódulos, es esencialmente una zona bacteriana, ya que está llena de células bacterianas que se encuentran en la etapa de desarrollo bacteriana en lugar de bacterioide. Sin embargo, cuando se trata de la zona de tejido del nódulo que contiene bacterias, se acostumbra llamarlo bacteroide.
La región bacteroide del nódulo se compone principalmente de células infectadas con bacterias del nódulo y en parte de células no infectadas adyacentes a ellas, llenas de pigmentos, taninos y, en otoño, de almidón.
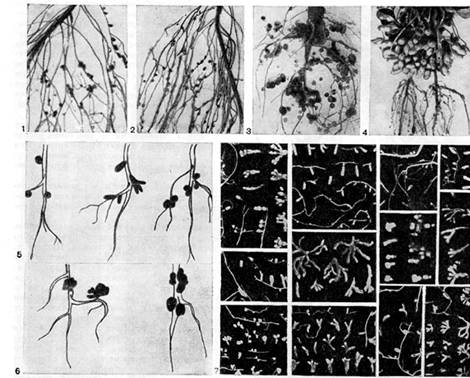
Arroz. dieciséis - Forma de nódulos en plantas leguminosas: 1, 2 - guisantes; 3, 6 (derecha) - lupino; 4 - cacahuetes (según E. Schwinghamer); 5 - filas, trébol, wiki, 6 (izquierda) - alfalfa (según J. Date); 7 - diferentes tipos de plantas leguminosas silvestres (según A.R. Werner, N.Ya. Gordienko)
En los nódulos formados por cepas eficaces de bacterias nodulares, el volumen relativo medio de la zona bacteroide es mayor que en los nódulos formados por la introducción de cepas ineficaces.
En algunos casos, el volumen de la región bacteroide alcanza un máximo en el período inicial de la vida del nódulo y, posteriormente, permanece relativamente constante. La zona bacteroide es penetrada por una densa red de hilos infecciosos y está rodeada por haces fibrosos vasculares a lo largo de la periferia.
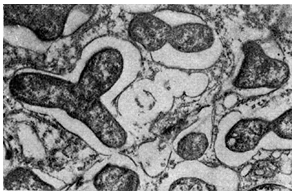
La forma de los bacteroides en los nódulos de diferentes tipos de leguminosas puede variar. Entonces, en wiki, rank y pea, son bifurcados o bifurcados. Para el trébol y la esparceta, la forma predominante de bacteroides es esférica, en forma de pera, hinchada, ovoide y redonda para los garbanzos. La forma de los bacteroides del frijol, seradella, pata de pájaro y lupino es casi en forma de varilla.
Los bacteroides llenan la mayor parte de la célula vegetal, con excepción de la zona central del núcleo y las vacuolas. Así, el porcentaje de bacteroides en la zona bacteroide de un nódulo de color rosa es 94,2 del número total de bacterias del nódulo. Las células bacterioides son de 3 a 5 veces más grandes que las células bacterianas.
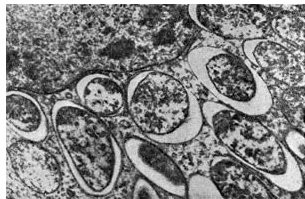
Arroz. 17
Los bacteroides de las bacterias de los nódulos son de particular interés debido al hecho de que son casi los únicos habitantes de los nódulos de las plantas leguminosas durante el período de fijación intensiva del nitrógeno atmosférico por parte de ellas. Algunos investigadores consideran que los bacteroides son formas degenerativas patológicas y no asocian el proceso de fijación de nitrógeno con la forma bacteroide de las bacterias del nódulo. La mayoría de los investigadores encuentran que los bacteroides son las formas más viables y activas de bacterias de nódulos y que las leguminosas fijan nitrógeno atmosférico solo con su participación.
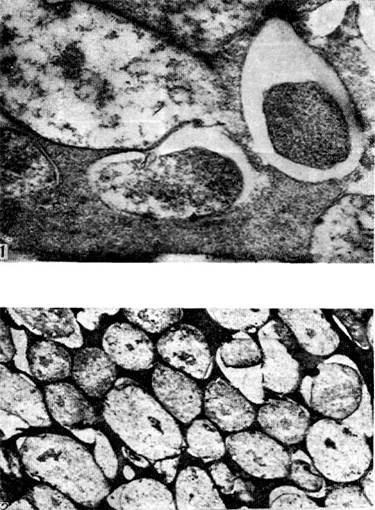
Arroz. 18
El sistema vascular del nódulo proporciona un vínculo entre la bacteria y la planta huésped. Los nutrientes y los productos metabólicos se transportan a través de los haces vasculares. El sistema vascular se desarrolla temprano y funciona durante mucho tiempo.
Los vasos completamente formados tienen una estructura específica: consisten en traqueidas de xilema, fibras de floema, tubos cribosos y células acompañantes.
Según el tipo de leguminosa, la conexión del glomérulo se realiza a través de uno o más haces vasculares. Por ejemplo, en los guisantes hay dos nódulos vasculares diferenciados en la base del nódulo. Cada uno de ellos generalmente se ramifica dicotómicamente dos veces y, como resultado, 8 paquetes pasan a través del nódulo desde el lugar de la segunda ramificación dicotómica. Muchas plantas tienen solo un racimo, mientras que al mismo tiempo, en un nódulo de Sesbania grandiflora a la edad de un año, pudieron contar hasta 126. Muy a menudo, el sistema vascular del nódulo está separado del lado exterior de su corteza por una capa de células parcial o totalmente suberizadas, denominada endodermo nodular, adherida a la endodermis de la raíz. El endodermo del nódulo es la capa externa del parénquima bovino no infectado, ubicado entre el tejido del nódulo y la corteza de la raíz.
En la mayoría de las especies de plantas, los nódulos se forman según el tipo descrito. Por lo tanto, la formación de nódulos es el resultado de fenómenos complejos que comienzan fuera de la raíz. Después de las fases iniciales de infección, se induce la formación de un nódulo, luego se produce la propagación de bacterias en la zona del tejido del nódulo y la fijación de nitrógeno.
Todas las etapas de desarrollo de las bacterias de los nódulos, según el microbiólogo checo V. Kas (1928), se pueden rastrear en las secciones de los nódulos. Entonces, en la parte superior del nódulo, por ejemplo, la alfalfa contiene principalmente pequeñas células en división en forma de bastón, una pequeña cantidad de bacteroides jóvenes, cuyo número aumenta gradualmente a medida que se desarrolla el nódulo. En la parte central del nódulo, de color rosado, se encuentran predominantemente células bacteroides y, con menos frecuencia, células pequeñas en forma de bastón. En la base del nódulo en las primeras etapas de vegetación de la planta huésped, los bacteroides son los mismos que en su parte media, pero al final de la temporada de crecimiento están más hinchados y degeneran antes.
El momento de aparición de los primeros nódulos visibles en las raíces de varios tipos de plantas leguminosas es diferente. Su aparición en la mayoría de las leguminosas ocurre con mayor frecuencia durante el desarrollo de las primeras hojas verdaderas. Así, la formación de los primeros nódulos de la alfalfa se observa entre el 4° y 5° día después de la germinación, ya los 7° - 8° día este proceso ocurre en todas las plantas. Los nódulos de la alfalfa falciforme aparecen a los 10 días.
Durante el período de funcionamiento, los nódulos suelen ser densos. Los nódulos formados por cultivos activos de bacterias son de color blanquecino a una edad temprana. En el momento de la manifestación de la actividad óptima, se vuelven rosados. Los nódulos que han surgido durante la infección con cultivos bacterianos inactivos son de color verdoso. A menudo, su estructura prácticamente no difiere de la estructura de los nódulos formados con la participación de cepas activas de bacterias de nódulos, pero se destruyen prematuramente.
En algunos casos, la estructura de los nódulos formados por bacterias inactivas se desvía de la norma. Esto se expresa en la desorganización del tejido del nódulo, que suele perder su diferenciación zonal claramente definida.
El color rosa está determinado por la presencia de un pigmento en los nódulos, que es similar en composición química a la hemoglobina sanguínea. En este sentido, el pigmento se llama leghemoglobina (legoglobina) - Leguminosae hemoglobina. La legoglobina se encuentra solo en las células de los nódulos que contienen bacteroides. Se localiza en el espacio entre los bacteroides y la membrana que los rodea.
Su cantidad oscila entre 1 y 3 mg por 1 g de nódulo, según el tipo de leguminosa.
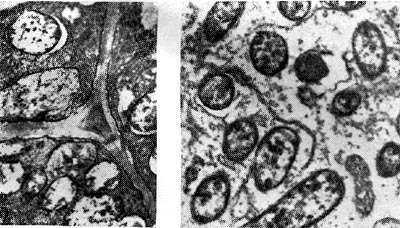
Arroz. 19
En las leguminosas anuales, al final de la temporada de crecimiento, cuando termina el proceso de fijación de nitrógeno, el pigmento rojo se vuelve verde. El cambio de color comienza en la base del nódulo, luego su parte superior se vuelve verde. En las leguminosas perennes no se produce el enverdecimiento de los nódulos o se observa sólo en la base del nódulo. En diferentes tipos de plantas leguminosas, la transición del pigmento rojo al verde se produce con diversos grados de intensidad ya diferentes velocidades.
Los nódulos de las plantas anuales funcionan por un tiempo relativamente corto. En la mayoría de las leguminosas, la necrosis de los nódulos comienza durante el período de floración de la planta huésped y, por lo general, procede del centro hacia la periferia del nódulo. Uno de los primeros signos de destrucción es la formación de una capa de células con fuertes paredes en la base del nódulo. Esta capa de células, situada perpendicularmente al vaso principal de la raíz, lo separa del nódulo y retrasa el intercambio de nutrientes entre la planta huésped y los tejidos del nódulo.
Numerosas vacuolas aparecen en las células del tejido en degeneración del nódulo, los núcleos pierden su capacidad de teñirse, algunas de las células bacterianas del nódulo se lisan y algunas migran al medio ambiente en forma de pequeñas células de artrosporas cocoides.
Los nódulos viejos son oscuros, flácidos, blandos. Cuando se cortan, una mucosidad acuosa sobresale de ellos. El proceso de destrucción de los nódulos, a partir del taponamiento de las células del sistema vascular, se ve facilitado por la disminución de la actividad fotosintética de la planta, la sequedad o la excesiva humedad del ambiente.
En un nódulo mucilaginoso destruido, a menudo se encuentran protozoos, hongos, bacilos y pequeñas bacterias de nódulos en forma de varilla.
El estado de la planta huésped afecta la duración del funcionamiento del nódulo. Así, según F. F. Yukhimchuk (1957), al castrar o quitar las flores de lupino, es posible prolongar el período de su vegetación y, al mismo tiempo, el tiempo de actividad activa de las bacterias del nódulo.
Los nódulos de las plantas perennes, a diferencia de los nódulos anuales, pueden funcionar durante muchos años. Entonces, por ejemplo, caragana tiene nódulos perennes, en los que el proceso de envejecimiento celular ocurre simultáneamente con la formación de otros nuevos. En la glicinia (glicinia china), también funcionan nódulos perennes que forman hinchazones esféricas en las raíces del huésped. Al final de la temporada de crecimiento, el tejido bacteroide de los nódulos perennes se degrada, pero el nódulo completo no muere. Al año siguiente vuelve a funcionar.
Factores que determinan la relación simbiótica de las bacterias del nódulo con las leguminosas - Para una simbiosis que asegure un buen desarrollo de las plantas, es necesario un determinado conjunto de condiciones ambientales. Si las condiciones ambientales son desfavorables, incluso a pesar de la alta virulencia, capacidad competitiva y actividad del microsimbionte, la eficacia de la simbiosis será baja.
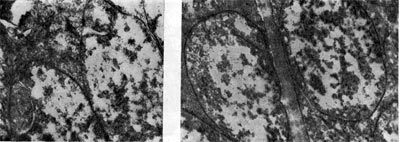
Arroz. 20
Para el desarrollo de nódulos, el contenido de humedad óptimo es 60-70% de la capacidad total de humedad del suelo. La humedad mínima del suelo en la que todavía es posible el desarrollo de bacterias nodulares en el suelo es aproximadamente igual al 16% de la capacidad de humedad total. Cuando la humedad está por debajo de este límite, las bacterias del nódulo ya no suelen multiplicarse, pero sin embargo no mueren y pueden permanecer inactivas durante mucho tiempo. La falta de humedad también conduce a la muerte de los nódulos ya formados.
El exceso de humedad, así como su falta, también es desfavorable para la simbiosis: debido a una disminución en el grado de aireación en la zona de la raíz, empeora el suministro de oxígeno al sistema de raíces de la planta. La aireación insuficiente también afecta negativamente a las bacterias del nódulo que viven en el suelo, las cuales, como saben, se multiplican mejor cuando hay oxígeno disponible. Sin embargo, la alta aireación en la zona de la raíz conduce al hecho de que los reductores de nitrógeno molecular comienzan a fijar el oxígeno, reduciendo el grado de fijación de nitrógeno de los nódulos.
El factor temperatura juega un papel importante en la relación entre las bacterias del nódulo y las leguminosas. Las características de temperatura de los diferentes tipos de plantas leguminosas son diferentes. Además, las diferentes cepas de bacterias del nódulo tienen sus propios valores óptimos de temperatura específicos para el desarrollo y la fijación de nitrógeno activo. Cabe señalar que las temperaturas óptimas para el desarrollo de las leguminosas, la formación de nódulos y la fijación de nitrógeno no coinciden. Así, en condiciones naturales, la formación de nódulos se puede observar a temperaturas ligeramente superiores a 0°C, la fijación de nitrógeno prácticamente no se produce en tales condiciones. Quizás solo las leguminosas simbióticas del Ártico fijan nitrógeno a temperaturas muy bajas. Por lo general, este proceso ocurre solo a 10°C y más. La máxima fijación de nitrógeno de varias leguminosas se observa a 20-25°C. Las temperaturas superiores a 30°C afectan negativamente al proceso de acumulación de nitrógeno.
La reacción del suelo tiene una gran influencia en la actividad vital de las bacterias del nódulo y la formación de nódulos. Para diferentes especies e incluso cepas de bacterias de nódulos, el valor de pH del hábitat es algo diferente. Por ejemplo, las bacterias del nódulo del trébol son más resistentes a los valores bajos de pH que las bacterias del nódulo de la alfalfa. Evidentemente, aquí también incide la adaptación de los microorganismos al medio. El trébol crece en suelos más ácidos que la alfalfa. La reacción del suelo como factor ecológico afecta la actividad y virulencia de las bacterias del nódulo. Las cepas más activas tienden a ser más fáciles de aislar de suelos con valores de pH neutros. En suelos ácidos, las cepas inactivas y débilmente virulentas son más comunes. Un ambiente ácido (pH 4.0 - 4.5) tiene un efecto directo sobre las plantas, en particular, interrumpiendo los procesos de síntesis del metabolismo de las plantas y el desarrollo normal de los pelos radiculares. En un ambiente ácido en plantas inoculadas, el período de funcionamiento del tejido bacteroide se reduce drásticamente, lo que conduce a una disminución en el grado de fijación de nitrógeno.
En suelos ácidos, como señaló A.V. San Petersburgo, las sales de aluminio y manganeso pasan a la solución del suelo, afectando negativamente el desarrollo del sistema de raíces de las plantas y el proceso de asimilación de nitrógeno, y también disminuye el contenido de formas asimilables de fósforo, calcio, molibdeno y dióxido de carbono. La reacción desfavorable del suelo se elimina mejor mediante el encalado.
Para las bacterias del nódulo, el tejido huésped es un medio nutritivo que puede satisfacer incluso a la cepa más exigente debido al contenido de todo tipo de nutrientes en el tejido. Sin embargo, después de la introducción de bacterias nodulares en el tejido de la planta huésped, su desarrollo no solo está determinado por procesos internos, sino que también depende en gran medida de la acción de factores externos que afectan todo el curso del proceso infeccioso. El contenido o ausencia de uno u otro nutriente en el ambiente puede ser un momento determinante para la manifestación de la fijación simbiótica de nitrógeno.
El grado de suministro de las plantas leguminosas con formas disponibles de compuestos de nitrógeno mineral determina la efectividad de la simbiosis. Sobre la base de numerosos experimentos de laboratorio y vegetativos, se sabe que cuantos más compuestos que contienen nitrógeno en el medio ambiente, más difícil es que las bacterias penetren en la raíz.
De gran importancia para activar la absorción de nitrógeno por parte de las leguminosas es la nutrición con fósforo. Con un bajo contenido de fósforo en el medio, las bacterias penetran en la raíz, pero no se forman nódulos. Las leguminosas tienen algunas peculiaridades en el intercambio de compuestos que contienen fósforo. Las semillas de leguminosas son ricas en fósforo. El fósforo de reserva durante la germinación de las semillas no se usa de la misma manera que en otros cultivos, de manera relativamente uniforme para la formación de todos los órganos, pero se concentra en mayor medida en las raíces. Por tanto, en las primeras etapas de desarrollo, las leguminosas, a diferencia de los cereales, satisfacen en mayor medida sus necesidades de fósforo a expensas de los cotiledones, y no de las reservas del suelo. Cuanto más grandes son las semillas, menos leguminosas dependen del fósforo del suelo. Sin embargo, con un modo de existencia simbiótico, la necesidad de fósforo en las leguminosas es mayor que con una autótrofa. Por lo tanto, con una falta de fósforo en el medio de las plantas inoculadas, empeora el suministro de nitrógeno a las plantas.
La fijación simbiótica de nitrógeno también requiere magnesio, azufre y hierro. Con la falta de magnesio, se inhibe la reproducción de las bacterias del nódulo, disminuye su actividad vital y se suprime la fijación simbiótica de nitrógeno. El azufre y el hierro también tienen un efecto beneficioso sobre la formación de nódulos y el proceso de fijación de nitrógeno, en particular, desempeñando un papel indudable en la síntesis de leghemoglobina.
De los oligoelementos, destacamos especialmente el papel del molibdeno y el boro. Con la falta de molibdeno, los nódulos se forman mal, se altera la síntesis de aminoácidos libres y se suprime la síntesis de leghemoglobina. El molibdeno, junto con otros elementos de valencia variable (Fe, Co, Cu) sirve como intermediario en la transferencia de electrones en reacciones enzimáticas redox. Con una deficiencia de boro, los haces vasculares no se forman en los nódulos y, como resultado, se interrumpe el desarrollo del tejido bacteroide.
La formación de nódulos en las leguminosas está muy influenciada por el metabolismo de los carbohidratos de las plantas, que está determinado por una serie de factores: la fotosíntesis, la presencia de dióxido de carbono en el medio ambiente y las características fisiológicas de las plantas. Mejorar la nutrición de carbohidratos tiene un efecto positivo en el proceso de inoculación y la acumulación de nitrógeno. Desde un punto de vista práctico, el uso de paja y estiércol fresco de paja para la fertilización de plantas leguminosas como fuente de carbohidratos es de gran interés. Pero en el primer año después de la introducción de paja en el suelo, las sustancias tóxicas se acumulan durante su descomposición. Cabe señalar que no todos los tipos de plantas leguminosas son sensibles a los productos tóxicos de descomposición de la paja; los guisantes, por ejemplo, no reaccionan a ellos.