עדויות פליאונטולוגיות מצביעות על כך שכמה צמחים השייכים לקבוצת Eucaesalpinioideae היו הקטניות העתיקות ביותר שהיו להן גושים.
במינים מודרניים של צמחי קטניות, נמצאו גושים על השורשים של נציגים רבים ממשפחת Papilijonaceae.
מבחינה פילוגנטית נציגים פרימיטיביים יותר של משפחות כמו Caesalpiniaceae Mimosaceae אינם יוצרים גושים ברוב המקרים.
מתוך 13,000 המינים (550 סוגים) של צמחי קטניות, נוכחותם של גושים זוהתה עד כה רק בכ-1300 מינים (243 סוגים). זה כולל בעיקר מיני צמחים המשמשים בחקלאות (יותר מ-200).
לאחר שנוצרו גושים, קטניות רוכשות את היכולת לספוג חנקן אטמוספרי. עם זאת, הם מסוגלים להאכיל מצורות קשורות של חנקן - מלחי אמוניום וחומצה חנקתית. רק צמח אחד, Hedysarum coronarium, מטמיע חנקן מולקולרי בלבד. לכן, ללא גושים בטבע, צמח זה אינו מתרחש.
חיידקי הגושים מספקים לצמח הקטניות חנקן, הנקבע מהאוויר. צמחים, בתורם, מספקים לחיידקים תוצרי חילוף חומרים של פחמימות ומלחים מינרליים להם הם זקוקים לצמיחה והתפתחות.
בשנת 1866, הבוטנאי ומדען הקרקע המפורסם מ.ס. וורונין ראה את ה"עגלים" הקטנים ביותר בגושים על שורשי צמחי הקטניות. וורונין הציג הנחות נועזות לאותה תקופה: הוא קישר בין היווצרות גושים לפעילות החיידקים, והחלוקה המוגברת של תאי רקמת השורש עם התגובה של הצמח לחיידקים שחדרו לשורש.
20 שנה מאוחר יותר, המדען ההולנדי ביירינק בודד חיידקים מגושים של אפונה, בקיה, צ'יני, שעועית, סרדלה וסוכרייה וחקר את תכונותיהם, תוך בדיקת היכולת להדביק צמחים ולגרום להיווצרות גושים. הוא קרא למיקרואורגניזמים הללו Bacillus radicicola. מכיוון שחיידקים היוצרים נבגים שייכים לסוג Bacillus, ולחיידקי גושים חסרים יכולת זו, א.פרז'מובסקי שינה את שמם לחיידק Bacterium radicicola. B. Frank הציע שם גנרי מוצלח יותר לחיידקי גושים - Rhizobium (מיוונית rhizo - שורש, ביו - חיים; חיים על השורשים). שם זה השתרש ועדיין נמצא בשימוש בספרות.
כדי לייעד זן של חיידקי גושים, נהוג להוסיף לשם הגנרי Rhizobium מונח המקביל לשם הלטיני של מיני הצמחים שמהם הם מבודדים ועליהם הם יכולים ליצור גושים. לדוגמא, Rhizobium trifolii - חיידקי גוש תלתן, Rhizobium lupini - חיידקי תורמוס וכו'. במקרים בהם חיידקי גושים מסוגלים ליצור גושים על שורשיהם של סוגים שונים של צמחי קטניות, כלומר לגרום למה שנקרא זיהום צולב, שם המין הוא, כביכול, קולקטיבי - הוא משקף את ה"הדבקה ההולכת" הזו. יְכוֹלֶת. לדוגמה, Rhizobium leguminosarum - חיידקי גושים של אפונה (Pisum), עדשים (עדשה), דרגות (Lathyrus).
מורפולוגיה ופיזיולוגיה של חיידקי הגושים - חיידקי הגושים מאופיינים במגוון מדהים של צורות - פולימורפיזם. חוקרים רבים הפנו לכך תשומת לב כאשר חקרו חיידקי גושים בתרבית טהורה בתנאי מעבדה ובאדמה. חיידקי גושים יכולים להיות בצורת מוט וסגלגל. בין החיידקים הללו ישנן גם צורות הניתנות לסינון, צורות L, אורגניזמים ניידים של coccoid.
חיידקי גושים צעירים בתרבית טהורה על חומרי הזנה הם בדרך כלל בעלי צורה בצורת מוט, גודל המוטות הוא כ-0.5-0.9 X 1.2-3.0 מיקרון, ניידים, מתרבים בחלוקה. בתאים בצורת מוט של חיידקי גושים של תלתן, נצפית חלוקה על ידי קשירה. עם הגיל, תאים בצורת מוט יכולים לעבור לניצנים. לפי גראם, התאים צובעים בצורה שלילית, המבנה האולטרה-דק שלהם אופייני לחיידקים גראם שליליים.
עם ההזדקנות, חיידקי גושים מאבדים את הניידות שלהם ועוברים למצב של מה שנקרא מוטות חגורה. הם קיבלו את השם הזה בשל התחלופה של חלקים צפופים ורופפים של פרוטופלזמה בתאים. הפסים של התאים מזוהים היטב כאשר צופים במיקרוסקופ אור לאחר טיפול בתאים בצבעי אנילין. חלקים צפופים של פרוטופלזמה (חגורה) מוכתמים גרוע יותר מהרווחים ביניהם. במיקרוסקופ פלואורסצנטי הרצועות בצבע ירוק בהיר, המרווחים ביניהן אינם זוהרים ונראים כהים. ניתן למקם רצועות באמצע התא או בקצוות. חגורת התאים נראית גם על דפוסי עקיפה של אלקטרונים, אם התכשיר אינו מטופל בחומרי ניגוד לפני הצפייה. ככל הנראה, עם הגיל, תא החיידק מתמלא בתכלילים שומניים שאינם קולטים צבע וכתוצאה מכך גורמים לתא להיות מפוספס. שלב ה"מוטות החגורים" מקדים את שלב היווצרותם של בקטרואידים - תאים בעלי צורה לא סדירה: מעובה, מסועפת, כדורית, בצורת אגס וצורת צלוחית. מאשר תאי חיידקים בצורת מוט המצויים ברקמות גושים.
בקטרואידים מכילים יותר גרגירי וולוטין ומאופיינים בתכולה גבוהה יותר של גליקוגן ושומן מאשר תאים בצורת מוט. בקטרואידים הגדלים באמצעי תזונה מלאכותיים ונוצרים ברקמות גושים הם מבחינה פיזיולוגית מאותו סוג. הוא האמין כי בקטרואידים הם צורות של חיידקים עם תהליך חלוקה לא שלם. עם חלוקת תאים לא מלאה של חיידקי גושים, נוצרות צורות מסתעפות דיכוטומיות של בקטרואידים. מספר הבקטרואידים עולה עם הזדקנות התרבית; הופעתם מוקל על ידי דלדול המדיום התזונתי, הצטברות של מוצרים מטבוליים והחדרת אלקלואידים למדיום.
אורז. 3
בתרביות ישנות (בני חודשיים) של חיידקי גושים, באמצעות מיקרוסקופ אלקטרונים, ניתן לזהות תצורות כדוריות מוגדרות בבירור בתאים רבים - ארתרוספורות. מספרם בתאים משתנה בין 1 ל-5.
אורז. 4
חיידקי גושים מסוגים שונים של צמחי קטניות גדלים בקצבים שונים על חומרי הזנה. חיידקים הגדלים במהירות כוללים rhizobia של אפונה, תלתן, אספסת, שעועית מספוא, בקיה, עדשים, צ'יני, תלתן מתוק, חילבה, שעועית, חומוס ורגל ציפור; לגדילה איטית - חיידקי גושים של תורמוס, סויה, בוטנים, סרדלה, שעועית מונג, חומוס, סיפיניון, דלעת. ניתן להשיג מושבות מלאות של תרבויות הגדלות במהירות ביום ה-3-4 של הדגירה, מושבות של תרבויות הגדלות לאט - ב-7-8.
חיידקי גושים הגדלים במהירות מאופיינים בסידור פריטריכי של דגלים, בעוד שחיידקים הגדלים לאט הם מונוטריכיאליים.
בנוסף לפגללה, נוצרים צמחים חוטים ודמויי חרוזים בתאי חיידקי גושים כאשר הם גדלים על גבי מדיה נוזלית.אורכם מגיע ל-8-10 מיקרון. הם ממוקמים בדרך כלל על פני התא באופן פריטריכיאלי, הם מכילים בין 4 ל-10 או יותר לתא.
למושבות של חיידקי גושים הגדלים במהירות יש צבע של חלב אפוי, לעתים קרובות שקוף, רירי, עם קצוות חלקים, קמור בינוני, ולבסוף צומח על פני השטח של המדיום אגר. מושבות של חיידקים הגדלים לאט הן קמורות יותר, קטנות, יבשות, צפופות וככלל אינן צומחות על פני המדיום. הריר המיוצר על ידי חיידקי הגושים הוא תרכובת מורכבת מסוג פוליסכריד, הכוללת הקסוזות, פנטוזות וחומצות אורוניות.
חיידקי הגושים הם מיקרו-אירופילים (הם מתפתחים עם כמויות קטנות של חמצן בסביבה), עם זאת, הם מעדיפים תנאים אירוביים.
חיידקי הגושים משתמשים בפחמימות ובחומצות אורגניות כמקור פחמן במדיות תזונתיות, ובתרכובות שונות המכילות חנקן וחנקן כמקור חנקן. כאשר מגדלים אותם על מדיה עם תכולה גבוהה של חומרים המכילים חנקן, חיידקי הגושים עלולים לאבד את יכולתם לחדור לצמח וליצור גושים. לכן, חיידקי גושים גדלים בדרך כלל על תמציות צמחים (שעועית, מרק אפונה) או תמציות אדמה. את הזרחן הדרוש להתפתחות ניתן להשיג על ידי חיידקי גושים מתרכובות המכילות זרחן אורגניות; תרכובות מינרלים יכולות לשמש כמקור לסידן, אשלגן ואלמנטים מינרלים אחרים.
כדי לדכא מיקרופלורה ספרופיטית חיצונית בעת בידוד חיידקי גושים מגושים או ישירות מהאדמה, מומלצים מדיה תזונתית בתוספת של סגול קריסטל, טאנין או אנטיביוטיקה.
לפיתוח רוב התרבויות של חיידקי גושים, נדרשת טמפרטורה אופטימלית בטווח של 24-26 מעלות. ב-0° ו-37°C הצמיחה נעצרת. בדרך כלל תרביות של חיידקי גושים מאוחסנות בתנאי מעבדה בטמפרטורות נמוכות (2-4 מעלות צלזיוס).
סוגים רבים של חיידקי גושים מסוגלים לסנתז ויטמינים מקבוצת B, כמו גם חומרי גדילה כגון הטרואוסין (חומצה בטא אינדולאצטית).
כל חיידקי הגושים עמידים בערך באותה מידה לתגובה בסיסית של המדיום (pH = 8.0), אך רגישים באופן לא שווה לתגובה חומצית.
ספציפיות, ארסיות, תחרותיות ופעילות של חיידקי גושים - מושג הספציפיות של חיידקי גושים הוא מושג קולקטיבי. הוא מאפיין את יכולתם של חיידקים ליצור גושים בצמחים. אם מדברים על חיידקי גושים באופן כללי, אז מבחינתם היווצרות גושים רק בקבוצת צמחי קטניות היא כבר ספציפית בפני עצמה - יש להם סלקטיביות לצמחי קטניות.
אולם אם נתייחס לתרביות בודדות של חיידקי גושים, מסתבר שביניהם יש כאלה המסוגלים להדביק רק קבוצה מסוימת, לפעמים גדולה יותר, לפעמים קטנה יותר, של צמחי קטניות, ובמובן זה, הספציפיות של חיידקי הגושים. היא יכולת סלקטיבית ביחס לצמח המארח. הספציפיות של חיידקי הגוש יכולה להיות צרה (חיידקי גוש תלתן מדביקים רק קבוצה של תלתן - ספציפיות מינים, וחיידקי גוש התורמוס יכולים אפילו להתאפיין בספציפיות זנית - מדביקים רק זנים נטולי אלקלואיד או אלקלואידים של תורמוס). עם ספציפיות רחבה, חיידקי אפונה יכולים להדביק צמחי אפונה, סנטר ושעועית, וחיידקי גושי אפונה ושעועית יכולים להדביק צמחי אפונה, כלומר כולם מתאפיינים ביכולת "להצליב". הספציפיות של חיידקי הגושים עומדת בבסיס הסיווג שלהם.
הספציפיות של חיידקי הגושים נוצרה כתוצאה מהסתגלותם ארוכת הטווח לצמח אחד או לקבוצה מהם ומההעברה הגנטית של תכונה זו. בהקשר זה, קיימת גם הסתגלות שונה של חיידקי הגושים לצמחים בקבוצת ההדבקה הצולבת. לפיכך, חיידקי אספסת יכולים ליצור גושים בתלתן מתוק. אף על פי כן, הם מותאמים יותר לאספסת, וחיידקי תלתן מתוק מותאמים יותר לתלתן מתוק.
בתהליך ההדבקה של מערכת השורשים של צמחי קטניות בחיידקי גושים, יש חשיבות רבה לארסיות של מיקרואורגניזמים. אם הספציפיות קובעת את ספקטרום הפעולה של חיידקים, אז הארסיות של חיידקי הגושים מאפיינת את פעילות פעולתם בתוך ספקטרום זה. ארסיות מתייחסת ליכולת של חיידקי גושים לחדור לרקמת השורש, להתרבות שם ולגרום להיווצרות גושים.
תפקיד חשוב הוא לא רק מעצם היכולת לחדור לשורשי הצמח, אלא גם ממהירות החדירה הזו.
כדי לקבוע את הארסיות של זן של חיידקי גושים, יש צורך לבסס את יכולתו לגרום להיווצרות גושים. הקריטריון לארסיות של כל זן יכול להיות המספר המינימלי של חיידקים המספק זיהום נמרץ יותר של השורשים בהשוואה לזנים אחרים, ששיאו ביצירת גושים.

אורז. 5
באדמה, בנוכחות זנים אחרים, הזן הארס יותר לא תמיד ידביק את הצמח ראשון. במקרה זה, יש לקחת בחשבון את היכולת התחרותית שלו, אשר לעתים קרובות מסווה את תכונת הארסיות בתנאים טבעיים.
זה הכרחי שלזנים ארסיים יש גם תחרותיות, כלומר, הם יכולים להתחרות בהצלחה לא רק עם נציגי המיקרופלורה הספרופיטית המקומית, אלא גם עם זנים אחרים של חיידקי גושים. אינדיקטור לתחרותיות של זן הוא מספר הגושים הנוצרים על ידו כאחוז ממספר הגושים הכולל על שורשי הצמח.
תכונה חשובה של חיידקי הגושים היא פעילותם (יעילותם), כלומר. היכולת בסימביוזה עם צמחי קטניות להטמיע חנקן מולקולרי ולספק את צורכי הצמח המארח בו. בהתאם למידת התרומה של חיידקי הגושים לעלייה בתפוקת הקטניות, לרוב הם מחולקים לפעילים (יעילים), לא פעילים (לא יעילים) ולא פעילים (לא יעילים).
זן של חיידקים שאינו פעיל עבור צמח מארח אחד בסימביוזה עם זן אחר של צמח קטניות יכול להיות יעיל למדי. לכן, כאשר מאפיינים זן מבחינת יעילותו, יש לציין תמיד ביחס לאיזה מין צמח מארח באה לידי ביטוי השפעתו.
פעילותם של חיידקי הגושים אינה רכושם הקבוע. לעתים קרובות במעבדה, יש אובדן פעילות בתרביות של חיידקי גושים. במקרה זה, או שהפעילות של התרבית כולה אובדת, או שתאים בודדים עם פעילות נמוכה מופיעים. ירידה בפעילות של חיידקי הגושים מתרחשת בנוכחות אנטיביוטיקה מסויימת, חומצות אמינו. אחת הסיבות לאובדן הפעילות של חיידקי הגושים עשויה להיות השפעת הפאג'. על ידי מעבר, כלומר, העברת חיידקים שוב ושוב דרך הצמח המארח (הסתגלות לזן צמחי מסוים), ניתן להשיג זנים יעילים מאלה שאינם יעילים.
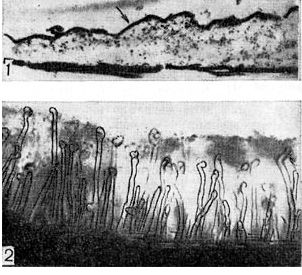
אורז. 6 - שכבת חומר גרגירי על פני השטח של שער השורש (1) במהלך בדיקה מיקרוסקופית אלקטרונית (לפי P. Dart, F. Mercer, הגדלה X 30,000) ושכבת זוגלי רירית (2), שבה נמצאות שערות שורש תלתן. שקוע, תחת מיקרוסקופ אור (זום x 80)
חשיפה לקרני גמא מאפשרת להשיג זנים ביעילות משופרת. ידועים מקרים של הופעת רדיו-מוטנטים פעילים ביותר של חיידקי אספסת מזן לא פעיל. השימוש בקרינה מייננת, שיש לה השפעה ישירה על השינוי במאפיינים הגנטיים של התא, ככל הנראה, יכול להיות טכניקה מבטיחה בבחירת זנים פעילים ביותר של חיידקי גושים.
הדבקה של צמח קטניות בחיידקי גושים - כדי להבטיח תהליך תקין של הדבקה של מערכת השורשים בחיידקי גושים, יש צורך בכמות די גדולה של תאי חיידקים ברי קיימא באזור השורשים.
במהלך התפתחות מערכת השורשים של צמח קטניות, ריבוי חיידקי הגושים על פני השורש מעורר על ידי הפרשות שורשים. גם תוצרי ההרס של כובעי השורשים והשערות ממלאים תפקיד חשוב במתן מצע מתאים לחיידקי הגושים.
על פני השורש יש שכבה של חומר רירי (מטריקס), שנוצרת ללא קשר להימצאות חיידקים בריזוספרה. שכבה זו נראית בבירור כאשר בוחנים אותה תחת מיקרוסקופ אור-אופטי. חיידקי גושים לאחר חיסון בדרך כלל ממהרים לשכבה זו ומצטברים בה בשל ההשפעה המעוררת של השורש, המתבטאת גם במרחק של עד 30 מ"מ.
קיימות מספר השערות לגבי מנגנון החדירה של חיידקי הגושים לשורש הצמח. המעניינים שבהם הם הבאים. מחברי אחת ההשערות קובעים כי חיידקי הגושים חודרים לשורש באמצעות פגיעה ברקמת האפידרמיס וקליפת המוח (במיוחד במקומות בהם מסתעפים שורשים לרוחב). השערה זו הועלתה על בסיס מחקר של בריל (1888), שגרם להיווצרות גושים בצמחי קטניות על ידי ניקוב השורשים במחט שהוטבעה קודם לכן בתרחיף של חיידקי גושים. כמקרה מיוחד, נתיב יישום כזה הוא די אמיתי. לדוגמה, בבוטנים ממוקמים בעיקר בציר של ענפי שורש, מה שמרמז על חדירת חיידקי גושים לשורש דרך מרווחים במהלך הנביטה של שורשים לרוחב.
השערה מעניינת ולא מופרכת היא חדירת חיידקי גושים לרקמת השורש דרך שערות השורש. המעבר של חיידקי גושים דרך שערות השורש מוכר על ידי רוב החוקרים.
ייתכן שחיידקי גושים יכולים לחדור לשורש דרך תאי האפידרמיס של קצות שורשים צעירים. לפי פרז'מובסקי (1889), חיידקים יכולים לחדור לשורש רק דרך דופן התא הצעיר (של שערות שורש או תאי אפידרמיס) ואינם מסוגלים לחלוטין להתגבר על השכבה שהשתנתה כימית או הפקקת של הקורטקס. זה עשוי להסביר שגבשושיות בדרך כלל מתפתחות בחלקים צעירים של השורש הראשי ובשורשים הצדדיים העולים.
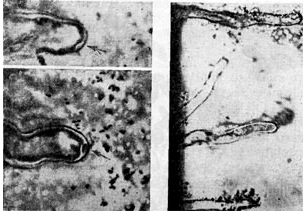
לאחרונה, השערת האוקסין זכתה לפופולריות רבה. מחברי השערה זו מאמינים שחיידקי גושים חודרים לשורש על ידי גירוי הסינתזה של חומצה p-indoleacetic (הטרואוקסין) מטריפטופן, הקיימת תמיד בהפרשות שורש צמחים. הנוכחות של heteroauxin קשורה לעקמומיות של שערות שורש, אשר נצפתה בדרך כלל כאשר מערכת השורשים נגועה בחיידקי גושים.
אורז. 7
המקור לחומצה בטא אינדולילאצטית בזמן ההדבקה של הצמח, מן הסתם, אינו רק צמחים המפרישים טריפטופן דרך מערכת השורשים, שסוגים רבים של חיידקים, כולל גושים שורשיים, יכולים להמיר לחומצה בטא אינדולילאצטית. חיידקי גושים עצמם, ואולי סוגים אחרים של מיקרואורגניזמים בקרקע החיים באזור השורש, יכולים גם הם להשתתף בסינתזה של הטרואוסין.
פעולת ההטרואוסין אינה ספציפית וגורמת לעקמומיות של שערות שורש במיני צמחים שונים, לא רק בקטניות. יחד עם זאת, חיידקי הגושים גורמים לעקמומיות של שערות השורשים רק בצמחי קטניות, תוך שהם מציגים סלקטיביות משמעותית למדי. אם ההשפעה הנידונה הייתה נקבעת רק ע"י חומצה β-indoleacetic, אז ספציפיות כזו לא הייתה קיימת. בנוסף, אופי השינויים בשערות השורש בהשפעת חיידקי הגושים שונה במקצת מאשר בהשפעת ההטרואוסין.
כמו כן, יש לציין שבמקרים מסוימים, שערות שורש לא מעוקלות חשופות לזיהום. תצפיות מראות כי באספסת ובאפונה, 60-70% משערות השורש מעוותות ומפותלות, ובתלתן - כ-50%. במינים מסוימים של תלתן, תגובה זו מופיעה בלא יותר מ-1/4 מהשערות הנגועות. בתגובה של עקמומיות, כמובן, יש חשיבות רבה למצב של שיער השורש. שערות שורש גדלות רגישות ביותר לפעולת חומרים המיוצרים על ידי חיידקים.
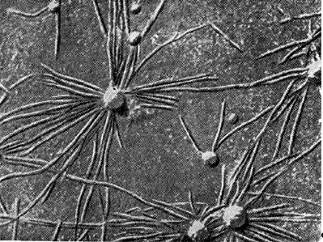
אורז. 8
גושים קטניות חיידקים חנקן
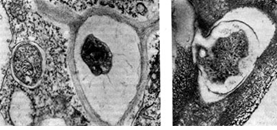
יש לציין השערה נוספת, לפיה חיידקי גושים חודרים לשורש במהלך היווצרות בליטה דמוית אצבע על פני שער השורש. תבנית עקיפה אלקטרונית של חתך שיער שורש המאששת השערה זו מציגה שערת שורש כפופה בצורת ידית מטרייה, בעיקול שלה יש הצטברות של חיידקי גושים. חיידקי הגושים כאילו נמשכים פנימה (בולעים) על ידי שיער השורש (בדומה לפינוציטוזיס).
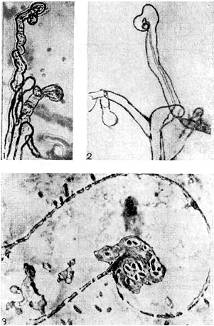
אורז. 9 - עקמומיות של שערות השורש של צמחים קטניות בנוכחות חיידקי גושים: 1.2 - גנב, x 120; 3 - חתך דק במיוחד דרך שיער השורש. מוּגדָל X 10,000 (לפי ק' סלמאן וג'י פאראוס)
תהליך החדרת חיידקי הגושים לרקמת השורש זהה בכל סוגי הצמחים הקטניות ומורכב משני שלבים. בשלב הראשון מתרחשת זיהום של שערות השורש. בשלב השני, תהליך היווצרות הגושים מתקדם באינטנסיביות. משך השלבים שונה במיני צמחים שונים: ב-Trifolium fragiferum השלב הראשון נמשך 6 ימים, ב-Trifolium nigrescens - 3 ימים. במקרים מסוימים קשה מאוד לזהות את הגבולות בין השלבים. ההחדרה האינטנסיבית ביותר של חיידקי גושים לשערות השורש מתרחשת בשלבים המוקדמים של התפתחות הצמח. השלב השני מסתיים במהלך היווצרות המונית של גושים. לעתים קרובות, חדירת חיידקי הגושים לשערות השורשים נמשכת גם לאחר היווצרות הגושים על השורשים. מה שנקרא עודף או זיהום נוסף זה מתרחש מכיוון שהזיהום של השערות אינו מפסיק במשך זמן רב. בשלבים המאוחרים יותר של ההדבקה, הגושים ממוקמים בדרך כלל נמוך יותר לאורך השורש.
לאחר שחדרו לתוך השורש (דרך שיער השורש, תא האפידרמיס, מקומות של נזק לשורש), חיידקי גושים עוברים לרקמות שורש הצמח. הכי קל, חיידקים עוברים דרך החללים הבין-תאיים.
תא בודד או קבוצה של תאים חיידקיים יכולים לחדור לתוך רקמת השורש. אם הוכנס תא נפרד, הוא יכול להמשיך לנוע ברקמה כתא בודד. דרך ההדבקה בשורש על ידי תאים בודדים אופיינית לצמחי תורמוס.
עם זאת, ברוב המקרים, התא הפולש, מתרבה באופן פעיל, יוצר את מה שנקרא חוטים זיהומיות (או tji זיהומיות) וכבר בצורה של חוטים כאלה, עובר לתוך רקמות הצמח.
המונח "חוט זיהום" עלה על בסיס חקר תהליך ההדבקה במיקרוסקופ אור. החל מעבודתו של ביירינק, חוט ההדבקה החל להיחשב כגוש דמוי היפוף רזה עם סגור
על פי שיטת היווצרות, הגושים של צמחי הקטניות מתחלקים לשני סוגים: סוג 1 - נודולים נוצרים במהלך חלוקת תאי הפריציקל (שכבת השורש), הממוקמים לרוב כנגד הפרוטוקסילם (ראשון בזמן ליצירת כלי דם). ) - סוג אנדוגני של היווצרות של גושים; סוג 2 – נודולות מקורן בקורטקס השורש כתוצאה מהחדרת הפתוגן לתאי הפרנכימליים של הקורטקס והאנדודרם (השכבה הפנימית של הקורטקס הראשוני) – סוג אקסוגני של היווצרות גושים.
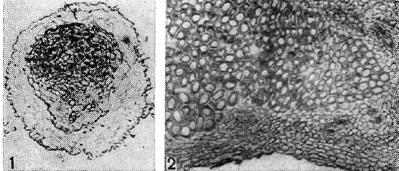
אורז. 10 - חתכים דרך רקמת הגוש של צמחים קטניות: 1 - חתך דרך גוש האפונה. תחילת ההתמיינות של רקמת כלי הדם בגוש (לפי G. Bond); d - חתך אורך דרך הגוש של הדרגה. מוּגדָל x90
בטבע, הסוג האחרון שולט. הרקמות של הגליל המרכזי של השורש לוקחות חלק רק ביצירת מערכת כלי הדם של גושים מסוג אנדוגני ואקסוגני כאחד.
ההבדל באופי היווצרות הגוש והשורש לרוחב נצפה במיוחד בסרדלה, שכן רקמת קליפת המוח של השורש הראשי של צמח זה - המקום של הגושים הראשונים - מורכבת משכבה קטנה יחסית של תאים וגושים הופכים לנראים מאוד. מהר לאחר הדבקה של השורש בחיידקים. תחילה הם יוצרים בליטות פחוסות על השורש, מה שמאפשר להבדיל בינם לבין הבליטות החרוטיות של השורשים הצדדיים. גושים נבדלים משורשים לרוחב במספר מאפיינים אנטומיים: היעדר גליל מרכזי, כובעי שורש ואפידרמיס, ונוכחות של שכבה משמעותית של קליפה המכסה את הגוש.
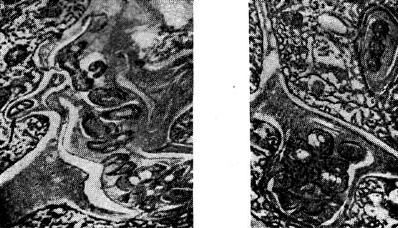
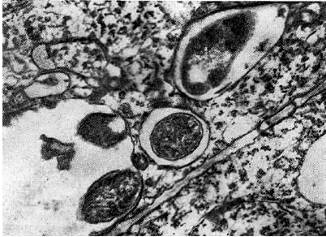
אורז. אחד עשר - חוטי זיהום בין תאי בצורת כיס (מימין) ולפיד (משמאל) עם חיידקי גושים. החץ מציג תא חיידקי שמתכונן להיכנס לציטופלזמה של תא צמחי. מוּגדָל X 20,000
היווצרות גושים של צמחי קטניות מתרחשת בתקופה שבה לשורש עדיין יש מבנה ראשוני. זה מתחיל בחלוקה של תאי קליפת המוח הממוקמים במרחק של 2-3 שכבות מקצות החוטים הזיהומיים. שכבות הקורטקס, שחודרות על ידי חוטים זיהומיים, נותרות ללא שינוי. יחד עם זאת, בסרדלה, חלוקת תאי קליפת המוח מתרחשת ישירות מתחת לשיער השורש הנגוע, בעוד שבאפונה, חלוקת התאים מצוינת רק בשכבה הלפני אחרונה של הקורטקס.
חלוקה עם היווצרות של מבנה רקמה רדיאלי ממשיכה לתאי הליבה הפנימיים. זה מתרחש ללא כיוון ספציפי, באופן אקראי, וכתוצאה מכך, נוצרת מריסטם (מערכת של רקמות חינוכיות) של הגוש, המורכב מתאי גרגירים קטנים.
תאים מחולקים של הקורטקס משתנים: הגרעינים מעוגלים וגדלים בגודלם, במיוחד הגרעינים. לאחר המיטוזה, הגרעינים מתפזרים ומבלי לקבל את צורתם המקורית, מתחילים להתחלק שוב.
מופיע מריסטם משני. עד מהרה, באנדודרם ובפריציקל, מופיעים סימני חלוקה מתחילה, אשר בתאים החיצוניים לשעבר מתרחשת בעיקר על ידי מחיצות משיקות. חלוקה זו מתרחבת לבסוף לקומפלקס המריסטמטי המשותף, שתאיו הקטנים מתארכים, הוואקוולים נעלמים, הגרעין ממלא את רוב התא. נוצר מה שנקרא הגוש הראשוני, בפלזמה של תאים שחיידקי הגוש נעדרים מהם, שכן בשלב זה הם עדיין נמצאים בתוך חוטי הזיהום. בזמן שהנודולה הראשונית נוצרת, חוטי זיהום מסתעפים פעמים רבות ויכולים לעבור בין תאים - בין תאי, או דרך תאים - תוך תאי - ולהחדיר חיידקים.
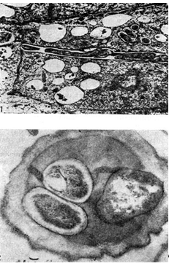
אורז. 12 - חוטים זיהומיים תוך תאיים עם חיידקי גושים: 1 - חתך אורך (לפי ד' גודצ'יילד ופ. ברגרסן), 2 - חתך רוחב. הוביל משם, x 35000
חוטים זיהומיים בין-תאיים עקב רבייה פעילה של חיידקי גושים בהם מקבלים לעתים קרובות צורה מוזרה - הם נוצרים בצורה של כיסים (דיברטיקולה) או לפידים.
תהליך התנועה של חוטי זיהום מתא לתא אינו ברור לחלוטין. ככל הנראה, חוטים מדבקים, כפי שסבור המיקרוביולוג הקנדי ד' ג'ורדן, מסתובבים בצורה של גדילים ריריים עירומים במרחבים הבין-תאיים של רקמת הצמח עד, בגלל כמה סיבות שעדיין בלתי מוסברות, הם מתחילים לחדור לתוך הציטופלזמה של תאים סמוכים.
בחלק מהמקרים, פלישת חוט הזיהום מתרחשת באחד, במקרים מסוימים - בכל תא שכן. מבעד לחללים הצינוריים המופלטים הללו (דיברטיקולה), תוכן החוט המוקף בריר זורם. הצמיחה הפעילה ביותר של חוטים זיהומיים מתרחשת בדרך כלל ליד הגרעין של תא הצמח. חדירת החוט מלווה בתנועת הגרעין, שנע לעבר מקום הזיהום, מתגבר, משנה צורה ומתנוון. תמונה דומה נצפית בזיהום פטרייתי, כאשר הגרעין שועט לעיתים קרובות לעבר ההיפיות הפולשות, נמשך לנזק באשר למקום הפעילות הפיזיולוגית הגדולה ביותר, מתקרב לחוט, מתנפח ומתמוטט. ככל הנראה, הדבר מאפיין את תגובת הצמח לזיהום.
בצמחים חד-שנתיים מופיעים בדרך כלל חוטים מדבקים בתקופת ההדבקה הראשונה של השורש, בצמחים רב-שנתיים - במהלך תקופה ארוכה של התפתחות.
אורז. 13
חיידקים יכולים להשתחרר מחוט הזיהום בזמנים שונים ובדרכים שונות. יציאת חיידקים, ככלל, היא תהליך ארוך מאוד, במיוחד בצמחים רב-שנתיים. בדרך כלל, שחרור חיידקים מחוט הזיהום לציטופלזמה של הצמח המארח קשור ללחץ פנימי הנובע מהכפלה אינטנסיבית של חיידקים בחוט והפרשת הריר שלהם. לפעמים חיידקים חומקים מהחוט בקבוצות המוקפות בריר של חוט הזיהום. בצורה של שלפוחיות (תצורות שלפוחיות). מכיוון שלשלפוחיות אין ממברנות, יציאת החיידקים מהן פשוטה מאוד. חיידקי גושים יכולים גם לחדור לתאי צמחים בנפרד מהחללים הבין-תאיים.
חיידקי גושים שיצאו מחוט הזיהום ממשיכים להתרבות ברקמת המארח. רבייתם בתקופה זו מתרחשת על ידי חלוקת היצרות. עיקר החיידקים מתרבים בציטופלזמה של התא, ולא בחוט הזיהום. תאים נגועים יוצרים רקמת בקטרואידית עתידית.
מלאים בתאים מתרבים במהירות של חיידקי גושים, תאי צמחים מתחילים להתחלק באופן אינטנסיבי. ברגע של חלוקה מיטוטית של תאים נגועים, חיידקי גושים יכולים להצטבר בשני קטבים מנוגדים של תא האם ולהיכנס באופן פסיבי לתאי הבת. כל אחד מהתאים הלא נגועים נמצא תחת השפעה מגרה חזקה של חיידקי גושים וכתוצאה מכך גם מתחלק. הודות לחלוקה מיטוטית נמרצת זו של תאים מריסטמטיים, חיידקי גושים מתפשטים לרקמות גושים ונפח אזור הבקטרואיד גדל.
לרקמה הנגועה, המורכבת מתאי שוכב צפוף ומתחלקים באופן פעיל, יש תחילה צורה של חרוט קטום. לאחר מכן, עקב הצמיחה ההדרגתית של חרוט זה וחלוקה והתפתחות בו-זמנית של תאים מריסטמטיים, רקמת הגושים גדלה ומאבדת את צורת החרוט שלה.
כך, הגוש גדל תחילה כתוצאה מחלוקה רדיאלית ומשיקית של תאי הליבה, ולאחר מכן עקב עלייה בגודלם וחלוקה בו זמנית. לאחר שתאי הצמח מתמלאים לחלוטין בחיידקים, המיטוזה נעצרת. עם זאת, התאים ממשיכים להגדיל את גודלם ולעתים קרובות הם מוארכים מאוד. גודלם גדול פי כמה מזה של תאי צמחים לא נגועים, הממוקמים ביניהם באזור הבקטירואיד של הגוש.
החיבור של גוש צעיר עם שורש של צמח קטניות מתבצע הודות לצרורות כלי דם-סיביים. בפעם הראשונה, צרורות סיבי כלי דם נצפו על ידי M.S. וורונין (1866). זמן התרחשות מערכת כלי הדם בגושים של סוגים שונים של צמחים קטניות שונה. אז, בגושים של סויה, תחילת ההתפתחות של צרורות כלי דם עולה בקנה אחד עם רגע החדירה של חיידקי הגושים לשתי שכבות של הפרנכימה של הפרה. עם צמיחת הגוש, מערכת המוליכה גדלה, מסתעפת ומקיפה את אזור החיידקים.
במקביל לתהליך ההתמיינות של מערכת כלי הדם, האנדודרם הגוש נוצר מהשכבה החיצונית של הגוש הראשוני. לאחר מכן הגוש מעוגל, שכבת התאים ההיקפית שלו מוקפת בקליפת הגוש.
האפידרמיס השורש נשבר, והגוש ממשיך להתפתח ולהגדיל את גודלו.
באמצעות מיקרוסקופ אור על חתכים אורכיים של גושים בוגרים, 4 אזורים אופייניים של התמיינות רקמות מובחנים בדרך כלל בבירור: קורטקס, מריסטם, אזור בקטרואיד ומערכת כלי הדם. כל רקמות הגושים מתמיינות ברצף אקרופטלי כאשר תאים חדשים יוזמים על ידי המריסטם.
אורז. 14
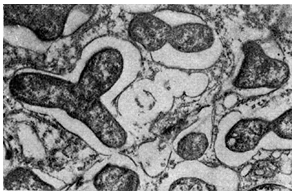
קליפת הגוש - המעטפת של הגוש, המבצעת תפקיד מגן. הקליפה מורכבת ממספר שורות של תאים פרנכימים לא נגועים, שגודלם ומאפייניהם שונים בקטניות שונות. לרוב, לתאי הקורטקס יש צורה מוארכת והם גדולים יותר בהשוואה לתאים אחרים של הגוש.
אורז. 15
בקליפת הגושים של מינים עציים רב-שנתיים, נמצאים לעתים קרובות תאים בעלי ממברנות פקקות המכילות שרפים, טאנינים וטאנינים.
מריסטם הגוש ממוקם מתחת לתאי הקורטקס והוא אזור של חלוקה אינטנסיבית של תאים לא נגועים. המריסטם של הגוש מאופיין במרווחים בצפיפות, ללא מרווחים בין-תאיים, תאים קטנים בעלי דופן דקה וצורה לא סדירה. תאי מריסטם גושים דומים לתאים מסוגים אחרים של רקמה מריסטמטית (השורש העליון, הגבעול העליון). תאי מריסטם גושים מכילים ציטופלזמה צפופה וגרגירית עדינה עם ריבוזומים, גופי גולגי, פרוטופלסטידים, מיטוכונדריה ומבנים אחרים. יש ואקוולים קטנים. במרכז הציטופלזמה נמצא גרעין גדול עם קרום גרעיני, נקבוביות וגרעין מוגדר בבירור. התפקידים של תאים מריסטמטיים הם ליצור תאים של קליפת הגוש, אזור הבקטרואיד ומערכת כלי הדם. בהתאם למיקום המריסטם, לגושים יש מגוון צורות: כדורית (אפונה, שעועית, סרדלה, בוטנים) או גלילית (אספסת, בקיה, דרגה, שיטה, תלתן). המריסטם, הממוקם באזורים נפרדים לאורך הפריפריה של הגוש, מוביל להיווצרות גושים דמויי גוש בתורמוס.
מריסטם הגושים מתפקד במשך זמן רב, אפילו בזמן נמק של גושים, כאשר הם כבר מלאים במסת בקטרואיד מתמזגת ותאי צמחים הרוסים.
אזור החיידקים של הגוש תופס את החלק המרכזי שלו ומהווה בין 16 ל-50% מסך המסה היבשה של הגושים. בתקופה הראשונה של היווצרות הגושים, זהו בעצם אזור חיידקי, שכן הוא מלא בתאי חיידקים שנמצאים בשלב ההתפתחות החיידקי ולא בקטרואידי. למרות זאת, כאשר מדובר באזור רקמת הגוש המכיל חיידקים, נהוג לקרוא לזה בקטרואיד.
אזור הבקטרואיד של הגוש מורכב בעיקר מתאים נגועים בחיידקי גוש וחלקו מתאים לא נגועים הסמוכים אליהם, מלאים בפיגמנטים, טאנינים ועד הסתיו - בעמילן.
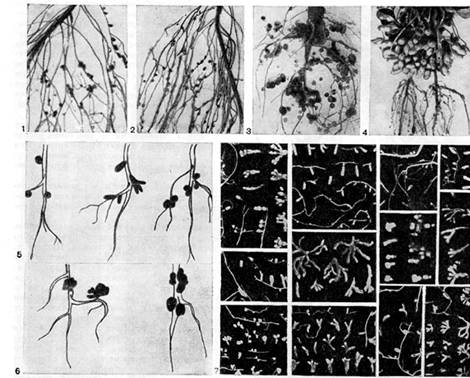
אורז. 16 - צורת גושים בצמחי קטניות: 1, 2 - אפונה; 3, 6 (מימין) - תורמוס; 4 - בוטנים (לפי E. Schwinghamer); 5 - דרגות, תלתן, ויקי, 6 (משמאל) - אספסת (לפי J. Date); 7 - סוגים שונים של צמחי קטניות בר (לפי א.ר. ורנר, נ.י. גורדיאנקו)
בגושים הנוצרים על ידי זנים יעילים של חיידקי גושים, הנפח היחסי הממוצע של אזור הבקטרואיד גבוה יותר מאשר בגושים הנוצרים עם החדרת זנים לא יעילים.
במקרים מסוימים, נפח אזור הבקטרואיד מגיע למקסימום בתקופה המוקדמת של חיי הגושים ולאחר מכן נשאר קבוע יחסית. אזור החיידקים חודר על ידי רשת צפופה של חוטים זיהומיים, והוא מוקף בצרורות סיביים כלי דם לאורך הפריפריה.
ניתן לגוון את צורת הבקטרואידים בגושים של סוגים שונים של קטניות. אז בוויקי, דרגה ואפונה, הם דו-ענפים או מפוצלים. עבור תלתן וסיפיניון, הצורה השלטת של בקטרואידים היא כדורית, בצורת אגס, נפוחה, ביצית ועגולה עבור חומוס. צורת החיידקים של השעועית, הסרדלה, רגל הציפור והתורמוס היא כמעט בצורת מוט.
בקטרואידים ממלאים את רוב תא הצמח, למעט האזור המרכזי של הגרעין וה-vacuoles. לפיכך, אחוז הבקטרואידים באזור הבקטרואיד של גוש בצבע ורוד הוא 94.2 מסך חיידקי הגוש. תאים בקטרואידים גדולים פי 3-5 מתאי חיידקים.
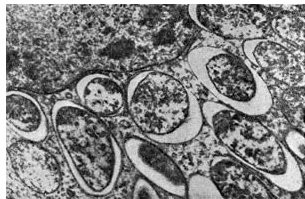
אורז. 17
בקטרואידים של חיידקי גושים מעניינים במיוחד בשל העובדה שהם כמעט התושבים היחידים של הגושיות של צמחי קטניות במהלך תקופת הקישור האינטנסיבי של חנקן אטמוספרי על ידם. יש חוקרים הרואים בבקטרואידים צורות ניווניות פתולוגיות ואינם מקשרים את תהליך קיבוע החנקן עם הצורה הבקטרואידית של חיידקי גושים. רוב החוקרים מגלים כי בקטרואידים הם הצורות הקיימות והפעילות ביותר של חיידקי גושים וכי קטניות מקבעות חנקן אטמוספרי רק בהשתתפותן.
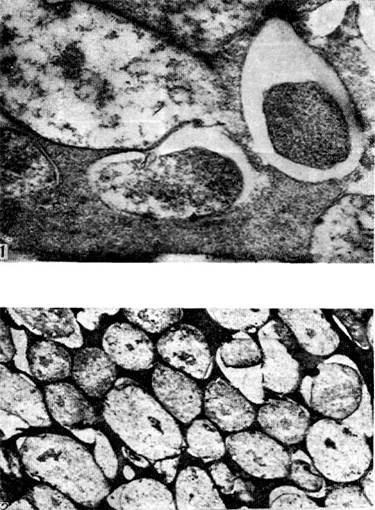
אורז. 18
מערכת כלי הדם של הגוש מספקת קישור בין החיידקים לצמח המארח. חומרים מזינים ומוצרים מטבוליים מועברים דרך צרורות כלי הדם. מערכת כלי הדם מתפתחת מוקדם ומתפקדת לאורך זמן.
לכלים המעוצבים במלואם יש מבנה ספציפי: הם מורכבים מטרכאידים של קסילם, סיבי פלואם, צינורות מסננת ותאים נלווים.
בהתאם לסוג הקטניות, חיבור הגלומרולוס מתבצע דרך צרור כלי דם אחד או יותר. לדוגמה, באפונה, יש שני צמתים מובחנים בכלי דם בבסיס הגוש. כל אחד מהם בדרך כלל מסתעף באופן דיכוטומי פעמיים, וכתוצאה מכך עוברים 8 צרורות דרך הגוש ממקום ההסתעפות הדיכוטומית השנייה. לצמחים רבים יש רק חבורה אחת, בעוד שבמקביל, בגוש אחד של Sesbania grandiflora בגיל שנה, הם הצליחו לספור עד 126. לעתים קרובות למדי, מערכת כלי הדם של הגוש מופרדת מהצד החיצוני של הגוש. קליפתו על ידי שכבה של תאים חלקיים או מלאים, הנקראים אנדורם נודולים, המחוברים לאנדודרמיס השורש. האנדודרם הגוש הוא השכבה החיצונית של פרנכימה בקר לא נגועה, הממוקמת בין רקמת הגוש לקליפת השורש.
ברוב מיני הצמחים נוצרים גושים לפי הסוג המתואר. לכן, היווצרות גושים היא תוצאה של תופעות מורכבות שמתחילות מחוץ לשורש. לאחר השלבים הראשוניים של הזיהום, נגרמת היווצרות של גושים, ואז מתרחשים התפשטות של חיידקים באזור רקמת הגוש וקיבוע חנקן.
על פי המיקרוביולוג הצ'כי V. Kas (1928), ניתן לאתר את כל שלבי ההתפתחות של חיידקי גושים על קטעים של גושים. אז בחלק העליון של הגוש, למשל, אספסת מכילה בעיקר תאים קטנים בצורת מוט מתחלק, כמות קטנה של בקטרואידים צעירים, שמספרם גדל בהדרגה עם התפתחות הגוש. בחלק האמצעי, בצבע ורוד של הגוש, נמצאים בעיקר תאים בקטרואידים ולעתים רחוקות יותר תאים קטנים בצורת מוט. בבסיס הגוש בשלבי הצמחייה המוקדמים של הצמח המארח, הבקטרואידים זהים לחלקו האמצעי, אך בסוף עונת הגידול הם נפוחים יותר ומתנוונים מוקדם יותר.
העיתוי של הופעת הגושים הראשונים הנראים לעין על השורשים של סוגים שונים של צמחי קטניות שונה. הופעתם ברוב הקטניות מתרחשת לרוב במהלך התפתחות העלים האמיתיים הראשונים. לפיכך, היווצרות הגושים הראשונים של אספסת נצפית בין הימים הרביעי והחמישי לאחר הנביטה, וביום השביעי - השמיני תהליך זה מתרחש בכל הצמחים. הגושים של אספסת מגל מופיעים לאחר 10 ימים.
במהלך תקופת התפקוד, הגושים בדרך כלל צפופים. גושים הנוצרים על ידי תרבויות פעילות של חיידקים בצבע לבנבן בגיל צעיר. עד לביטוי של פעילות אופטימלית, הם הופכים ורודים. גושים שהתעוררו במהלך הדבקה בתרביות חיידקים לא פעילות הם בצבע ירקרק. לעתים קרובות, המבנה שלהם למעשה אינו שונה ממבנה הגושים הנוצרים בהשתתפות זנים פעילים של חיידקי גושים, אך הם נהרסים בטרם עת.
במקרים מסוימים, מבנה הגושים הנוצרים על ידי חיידקים לא פעילים חורג מהנורמה. זה מתבטא בחוסר ארגון של רקמת הגוש, שלרוב מאבדת את ההתמיינות האזורית המוגדרת בבירור שלה.
הצבע הוורוד נקבע על ידי נוכחות של פיגמנט בגושים, הדומה בהרכבו הכימי להמוגלובין בדם. בהקשר זה, הפיגמנט נקרא leghemoglobin (לגגלובין) - Leguminosae hemoglobin. לגוגלובין נמצא רק באותם תאי גושים המכילים בקטרואידים. הוא ממוקם ברווח שבין הבקטרואידים והממברנה המקיפה אותם.
הכמות שלו נעה בין 1 ל-3 מ"ג ל-1 גרם של גוש, תלוי בסוג הצמח הקטנית.
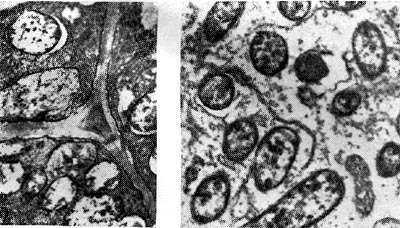
אורז. 19
בצמחי קטניות חד-שנתיים, עד סוף עונת הגידול, כאשר מסתיים תהליך קיבוע החנקן, הפיגמנט האדום הופך לירוק. שינוי הצבע מתחיל בבסיס הגוש, מאוחר יותר החלק העליון שלו הופך לירוק. בצמחי קטניות רב-שנתיים לא מתרחשת ירוקת של גושים או שהיא נצפית רק בבסיס הגוש. בסוגים שונים של צמחי קטניות, המעבר של פיגמנט אדום לירוק מתרחש בדרגות עוצמה שונות ובקצבים שונים.
גושים של צמחים חד-שנתיים מתפקדים לזמן קצר יחסית. ברוב הקטניות, נמק הגושים מתחיל בתקופת הפריחה של הצמח המארח וממשיך בדרך כלל מהמרכז לפריפריה של הגוש. אחד הסימנים הראשונים להרס הוא היווצרות של שכבת תאים עם קירות חזקים בבסיס הגוש. שכבת תאים זו, הממוקמת בניצב לכלי הראשי של השורש, מפרידה בינו לבין הגוש ומעכבת את חילופי החומרים המזינים בין הצמח המארח לרקמות הגוש.
בתאי הרקמה המתנוונת של הגוש מופיעים מספר רב של ואקוולים, הגרעינים מאבדים את יכולתם להכתים, חלק מתאי חיידקי הגוש מתמזגים, וחלקם נודדים לסביבה בצורה של תאי ארתרוספור קוקואידים קטנים.
גושים ישנים כהים, רופפים, רכים. כשחותכים, מבצבץ מהם ריר מימי. תהליך הרס הגוש, החל בפקק של תאי מערכת כלי הדם, מוקל על ידי ירידה בפעילות הפוטוסינתטית של הצמח, יובש או לחות יתר של הסביבה.
בגוש נהרס, רירי, פרוטוזואה, פטריות, חיידקים וחיידקים קטנים בצורת מוטות נמצאים לעתים קרובות.
מצב הצמח המארח משפיע על משך תפקודו של הגוש. לפיכך, לפי F. F. Yukhimchuk (1957), על ידי סירוס או הסרה של פרחי התורמוס, ניתן להאריך את תקופת הצמחייה שלו ובמקביל, את זמן הפעילות הפעילה של חיידקי הגוש.
גושים של צמחים רב-שנתיים, בניגוד לגושים חד-שנתיים, יכולים לתפקד במשך שנים רבות. כך, למשל, לקראגנה יש גושים רב-שנתיים, שבהם תהליך הזדקנות התאים מתרחש במקביל להיווצרותם של חדשים. בוויסטריה (וויסטריה סינית), מתפקדים גם גושים רב-שנתיים, היוצרים נפיחות כדוריות על שורשי המארח. עד סוף עונת הגידול, רקמת החיידקים של גושים רב-שנתיים מתפוררת, אך כל הגוש אינו מת. בשנה הבאה הוא מתחיל לתפקד שוב.
גורמים הקובעים את הקשר הסימביוטי של חיידקי הגושים עם צמחי קטניות - לסימביוזה המבטיחה התפתחות טובה של צמחים, יש צורך בסט מסוים של תנאים סביבתיים. אם התנאים הסביבתיים אינם נוחים, הרי שגם למרות הארסיות הגבוהה, יכולת התחרותיות והפעילות של המיקרוסימביונט, יעילות הסימביוזה תהיה נמוכה.
אורז. 20
לפיתוח גושים, תכולת הלחות האופטימלית היא 60-70% מיכולת הלחות הכוללת של הקרקע. לחות הקרקע המינימלית שבה עדיין מתאפשרת התפתחות של חיידקי גושים בקרקע שווה בקירוב ל-16% מיכולת הלחות הכוללת. כאשר הלחות מתחת לגבול זה, חיידקי הגושים בדרך כלל אינם מתרבים יותר, אך למרות זאת הם אינם מתים ויכולים להישאר במצב לא פעיל למשך זמן רב. חוסר הלחות מוביל גם למוות של גושים שכבר נוצרו.
עודף לחות, כמו גם חסרונה, אף הוא שלילי לסימביוזה - עקב ירידה במידת האוורור באזור השורשים, אספקת החמצן למערכת השורשים של הצמח מתדרדרת. אוורור לא מספיק משפיע לרעה גם על חיידקי הגושים החיים באדמה, שכידוע, מתרבים טוב יותר כאשר חמצן זמין. עם זאת, אוורור גבוה באזור השורש מוביל לעובדה שמפחיתי חנקן מולקולריים מתחילים לקשור חמצן, ומפחיתים את מידת קיבוע החנקן של גושים.
גורם הטמפרטורה ממלא תפקיד חשוב ביחסים בין חיידקי הגושים לצמחי קטניות. מאפייני הטמפרטורה של סוגים שונים של צמחי קטניות שונים. כמו כן, לזנים שונים של חיידקי גושים יש אופטימום טמפרטורה ספציפי משלהם להתפתחות וקיבוע חנקן פעיל. יש לציין שהטמפרטורות האופטימליות להתפתחות צמחי קטניות, היווצרות גושים וקיבוע חנקן אינן חופפות. אז, בתנאים טבעיים, ניתן לראות היווצרות של גושים בטמפרטורות מעט מעל 0 מעלות צלזיוס, קיבוע חנקן כמעט אינו מתרחש בתנאים כאלה. אולי רק קטניות סימביוטיות ארקטיות מקבעות חנקן בטמפרטורות נמוכות מאוד. בדרך כלל, תהליך זה מתרחש רק ב-10 מעלות צלזיוס ומעלה. קיבוע החנקן המרבי של מספר צמחי קטניות נצפה ב-20-25 מעלות צלזיוס. טמפרטורות מעל 30 מעלות צלזיוס משפיעות לרעה על תהליך הצטברות החנקן.
לתגובת הקרקע השפעה רבה על הפעילות החיונית של חיידקי הגושים ויצירת גושים. עבור מינים שונים ואפילו זנים של חיידקי גושים, ערך ה-pH של בית הגידול שונה במקצת. לדוגמה, חיידקי קשרי תלתן עמידים יותר לערכי pH נמוכים מאשר חיידקי גושי אספסת. ברור שגם התאמת המיקרואורגניזמים לסביבה משפיעה כאן. תלתן גדל בקרקעות חומציות יותר מאשר אספסת. תגובת הקרקע כגורם אקולוגי משפיעה על הפעילות והארסיות של חיידקי הגושים. הזנים הפעילים ביותר נוטים להיות קלים יותר לבידוד מקרקעות עם ערכי pH ניטרליים. בקרקעות חומציות, זנים לא פעילים וחולשים ארסיים שכיחים יותר. לסביבה חומצית (pH 4.0 - 4.5) יש השפעה ישירה על צמחים, בפרט, משבשת את התהליכים הסינתטיים של חילוף החומרים בצמחים והתפתחות תקינה של שערות שורש. בסביבה חומצית בצמחים מחוסנים, תקופת התפקוד של רקמת הבקטרואיד מצטמצמת בחדות, מה שמוביל לירידה במידת קיבוע החנקן.
בקרקעות חומציות, כפי שצוין על ידי A.V. פטרבורג, מלחי אלומיניום ומנגן עוברים לתמיסת הקרקע, ומשפיעים לרעה על התפתחות מערכת השורשים של הצמחים ועל תהליך הטמעת החנקן, ותכולת הצורות הניתנות להטמעה של זרחן, סידן, מוליבדן ופחמן דו חמצני פוחתת אף היא. תגובת אדמה לא חיובית בוטלה בצורה הטובה ביותר על ידי סיד.
עבור חיידקי גושים, הרקמה המארחת היא מדיום תזונתי כזה שיכול לספק אפילו את הזנים התובעניים ביותר בשל תכולת כל סוגי החומרים התזונתיים ברקמה. עם זאת, לאחר החדרת חיידקי גושים לרקמת הצמח המארח, התפתחותם נקבעת לא רק על ידי תהליכים פנימיים, אלא גם תלויה במידה רבה בפעולה של גורמים חיצוניים המשפיעים על כל מהלך התהליך הזיהומי. התוכן או היעדר של חומר תזונתי כזה או אחר בסביבה יכולים להיות רגע מכריע לביטוי של קיבוע חנקן סימביוטי.
מידת האספקה של צמחי קטניות עם צורות זמינות של תרכובות חנקן מינרליות קובעת את יעילות הסימביוזה. בהתבסס על ניסויים מעבדתיים וצמחיים רבים, ידוע שככל שיש יותר תרכובות המכילות חנקן בסביבה, כך קשה יותר לחיידקים לחדור לשורש.
חשיבות רבה בהפעלת ספיגת החנקן על ידי קטניות היא תזונת הזרחן. בתכולת זרחן נמוכה במדיום, חיידקים חודרים לשורש, אך לא נוצרים גושים. לצמחי קטניות יש כמה מוזרויות בחילופי תרכובות המכילות זרחן. זרעי קטניות עשירים בזרחן. זרחן רזרב במהלך נביטת הזרעים אינו משמש באותו אופן כמו בגידולים אחרים - באופן שווה יחסית להיווצרות כל האיברים, אלא מרוכז במידה רבה יותר בשורשים. לכן, בשלבי התפתחות מוקדמים, צמחי קטניות, בניגוד לדגנים, מספקים את צורכיהם לזרחן במידה רבה יותר על חשבון קוטלידונים, ולא עתודות קרקע. ככל שהזרעים גדולים יותר, כך פחות קטניות תלויות בזרחן הקרקע. עם זאת, עם אופן קיום סימביוטי, הצורך בזרחן בצמחי קטניות גבוה יותר מאשר באוטוטרופי. לכן, עם מחסור בזרחן בתווך של צמחים מחוסנים, אספקת הצמחים עם חנקן מחמירה.
קיבוע חנקן סימביוטי דורש גם מגנזיום, גופרית וברזל. עם חוסר במגנזיום מעכבת רביית חיידקי הגושים, פעילותם החיונית פוחתת וקיבוע חנקן סימביוטי מדוכא. לגופרית ולברזל יש גם השפעה מועילה על היווצרות גושים ועל תהליך קיבוע החנקן, בפרט, ממלאים תפקיד ללא ספק בסינתזה של להמוגלובין.
מבין יסודות הקורט, אנו מציינים במיוחד את התפקיד של מוליבדן ובור. עם חוסר מוליבדן, גושים נוצרים בצורה גרועה, הסינתזה של חומצות אמינו חופשיות מופרעת בהם, וסינתזה של להמוגלובין מדוכאת. מוליבדן, יחד עם יסודות אחרים בעלי ערכיות משתנה (Fe, Co, Cu) משמש כמתווך בהעברת אלקטרונים בתגובות אנזימטיות חיזור. עם מחסור בורון, צרורות כלי דם אינם נוצרים בגושים, וכתוצאה מכך, התפתחות רקמת בקטרואיד מופרעת.
היווצרות גושים בקטניות מושפעת רבות מחילוף החומרים של הפחמימות בצמחים, הנקבע על ידי מספר גורמים: פוטוסינתזה, נוכחות פחמן דו חמצני בסביבה ומאפיינים פיזיולוגיים של צמחים. לשיפור תזונת הפחמימות יש השפעה חיובית על תהליך החיסון והצטברות חנקן. מבחינה מעשית יש עניין רב בשימוש בזבל טרי וקש לדישון צמחי קטניות כמקור לפחמימות. אבל בשנה הראשונה לאחר הכנסת הקש לאדמה, מצטברים חומרים רעילים במהלך הפירוק שלו. יש לציין שלא כל סוגי הצמחים הקטניים רגישים לתוצרי פירוק רעילים של קש; אפונה, למשל, לא מגיבה אליה.