רבייה של מיקרואורגניזמים - עלייה בריכוז המיקרואורגניזמים ביחידת נפח של המדיום, שמטרתה שימור המין.
מיקרואורגניזמים מאופיינים ב:
מגוון שיטות רבייה;
מעבר משיטת רבייה אחת לאחרת;
האפשרות להשתמש במספר שיטות בו זמנית;
שיעור רבייה גבוה.
שיטות רבייה של מיקרואורגניזמים
אני. מיני עםשיטת רבייהנראה רק באוקריוטים.
II. שיטות רבייה א-מיניות.
ביקוע רוחבי בינארי בשטח שווה (חלוקה פשוטה, חלוקה איזומורפית, מיטוזה) נצפתה ברוב המיקרואורגניזמים החד-תאיים (חיידקים, ריקטזיה, פרוטוזואה, שמרים), כתוצאה מכך, נוצרות שתי יחידות בנות חדשות, שניחנו במידע הגנטי של תא האם, סימטריים ביחס לצירי האורך והרוחב, תא האם עצמו נעלם.
יחד עם זאת, ברוב חיידקי Gram+, החלוקה מתרחשת על ידי סינתזה של מחיצה רוחבית המשתרעת מהפריפריה למרכז (איור 63A). התאים של רוב חיידקי הגראם מתחלקים על ידי התכווצות התא (התא הופך דק יותר באמצע) (איור 63B).
ניצנים (ביקוע בינארי לא שווה) נראה אצל בני הסוג פרנסיסלהו מיקופלזמהופטריות דמויי שמרים. בעת ניצנים, תא האם מוליד תא בת: באחד הקטבים של תא האם נוצרת צמח קטן (ניצן) שמתגבר בתהליך הגדילה. בהדרגה מגיעה הכליה לגודל תא האם ולאחר מכן היא נפרדת. CS הכליות מסונתז לחלוטין מחדש (איור 63B). בתהליך הניצנים נצפית סימטריה ביחס לציר האורך בלבד. ישנם הבדלים מורפולוגיים ופיזיולוגיים בין תאי האם והבת. תא הבת החדש מסתגל טוב יותר לתנאים משתנים.
פרגמנטציה של צורות חוטיות מאפיין את הסוג Actinomycesו מיקופלזמה.
היווצרות אקסוספור מאפיין של Streptomycetes, פטריות דמויי שמרים ועובש.
מחזור פיתוח מיוחד tiyaנראה ב כלמידיה. רק צורות וגטטיביות של כלמידיה (גופים רשתיים או ראשוניים) מסוגלות להתחלק בתאי המקרואורגניזם. המחזור שלהם, המורכב מכמה חלוקות, מסתיים בהיווצרות צורות ביניים, שמהן נוצרים גופים יסודיים, המביאים לצורות וגטטיביות. לאחר הרס של דופן ה-vacuole והתא המארח, הגופים היסודיים משתחררים והמחזור חוזר על עצמו. המחזור נמשך 40-48 שעות.
חלוקה מרובה מתואר עבור קבוצה אחת של ציאנובקטריות חד-תאיות. ביקוע מרובה מבוסס על העיקרון של ביקוע בינארי שווה. ההבדל טמון בעובדה שבמקרה זה, לאחר ביקוע בינארי, תאי הבת המתקבלים אינם גדלים, אלא הם שוב עוברים חלוקה (איור 63D).
חלוקה מרובה (סכיזוגוניה)מתואר גם בפרוטוזואה (מלריה פלסמודיה): החומר הגרעיני מחולק להרבה נוקלאולים, מוקפים בקטעים של הציטופלזמה, וכתוצאה מכך נוצרים תאי בת רבים.
מנגנון ושלבים של חלוקה פשוטה
א. צמיחה לדרגה מסוימת של בשלות. גידול התאים אינו בלתי מוגבל, ולאחר שהגיע לגודל מסוים, תא החיידק מתחיל להתחלק. צמיחת התאים מואטת במהלך החלוקה ומתחילה שוב לאחר החלוקה.
ב. קריוקינזיס ( שכפול הדנ"א ו-ד חלוקת נוקלואיד). מהציטופלזמה הבוגרת מגיע אות המפעיל את הגן היוזם ב-DNA. מיקרואורגניזמים בפעולת הגן היוזם מסנתזים חלבון יוזם הפועל על גן המשכפל - קטע מיוחד של DNA שממנו מתחילה שכפול וחלוקה של DNA לשני גדילים.
החלוקה של מולקולת ה-DNA (שכפול) מתרחשת על פי מנגנון חצי שמרני ובדרך כלל תמיד קודמת לחלוקת התא. שכפול ה-DNA מתחיל בנקודת ההתקשרות של הכרומוזום המעגלי ל-CMP, שם המנגנון האנזימטי האחראי לשכפול ממוקם מקומי.
מנגנון שכפול ה-DNA מתבטא בשבירת קשרי מימן בין שתי שרשראות הפולינוקלאוטידים שלו, התפרקותם וסינתזה בעזרת DNA פולימראז לאורך כל שרשרת ישנה של שרשראות חדשות עם רצף בסיסים משלים. לאחר התפצלות לתאי בת לאורך שרשרת פולינוקלאוטיד אחת ישנה ואחת חדשה, קשרי מימן משוחזרים ביניהם ונוצר DNA דו-גדילי למחצה.
בדרך כלל, קיים קשר זמני מסוים בין שכפול הכרומוזומים וחלוקת תאי חיידקים. גם חשיפה לכימיקלים ולגורמים פיזיקליים שונים, המובילה לדיכוי שכפול ה-DNA, עוצרת את חלוקת התא. עם זאת, בתנאים מסוימים, הקשר בין שני התהליכים יכול להישבר, ותאים מסוגלים להתחלק בהיעדר סינתזת DNA.
B. ציטוקינזיס (חלוקת תאים). במקביל לשכפול של מולקולות DNA, הממברנה מסונתזת ליד המזוזום, באזור המגע של ה-DNA עם ה-CPM. היווצרות מחיצה מובילה לחלוקת תאים. הרגע שבו מתחילים את חלוקת התא הוא סוף שכפול ה-DNA. זה מוביל להפרדה של מולקולות DNA של בת וליצירת כרומוזומים נפרדים. תאי הבת החדשים שנוצרו נפרדים זה מזה.
עיכוב סינתזת הממברנה לפני סיום השכפול מוביל לשיבוש תהליך החלוקה: התא מפסיק להתחלק וגדל לאורכו. בחיידקים מסוימים, היווצרות מחיצה אינה מובילה לחלוקת תאים: נוצרים תאים רב-חדריים.
ד. ההתפצלות של תאי הבת שנוצרו מתרחשת כתוצאה מתמוגגת של השכבה האמצעית של ה-COP. אם לאחר חלוקה חוזרת ונשנית באותו מישור, התאים אינם מתפצלים, הם נוצרים שרשראות בצורת מוט (בצילוס) אוֹ כַּדוּרִי(סטרפטוקוקוס) תאים או תאים מזווגים(נייסריה) . הפרדת תאים אפשרית עם בידוד של אחד התאים על ידי תנועה לאורך פני השטח של השני, כתוצאה מכך, החיידקים נמצאים פָּרוּעַ (Escherichia). אם, במהלך ההפרדה, אחד מתאי הבת, מבלי להתנתק מנקודת החלוקה, נע לאורך קשת, V-מְעוּצָבטופס (Corynebacterium, Bifidobacterium). לאחר ביקוע בינארי והתפצלות תאים במספר מישורים, אשכולות תאים בצורות שונות:צרורות (סטפילוקוקוס), חבילות (סרצ'ינה) (איור 65). אם חלוקת נוקלואיד קודמת לחלוקת התא, מולטינוקלואידמיקרואורגניזמים. בהשפעת גורמים חיצוניים שליליים (מלחי מרה, קרני UV, פעילי שטח, אנטיביוטיקה), חלוקת התא יכולה להיפסק תוך שמירה על גדילתו. במקרה זה, היווצרות של מוארך פיליפורםתאים.
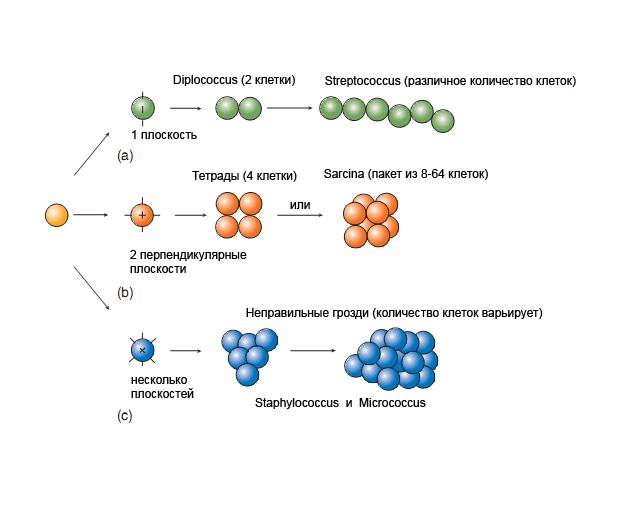
אורז. 65.חלוקה של קוקי
תקופת דור- מרווח הזמן שבמהלכו מכפיל את מספר החיידקים קצב רביית המיקרואורגניזמים ותקופת היצירה תלויים בסוג המיקרואורגניזם, גודל ותכונות החיסון, הרכב המדיום התזונתי, ה-pH שלו, אוורור, טמפרטורת הדגירה וגורמים נוספים. בתנאים נוחים, מיקרואורגניזמים רבים מתחלקים תוך 15-30 דקות. (ה. coli, ס. טיפי). במיקרואורגניזמים גחמניים, החלוקה מתבצעת לאחר 45-90 דקות (סטרפטוקוקוס, Corynebacterium) ואפילו לאחר 18 שעות (M. שַׁחֶפֶת).