Celično jedro je bistvena struktura vsakega evkariontskega organizma. opravlja najrazličnejše funkcije, vendar je njegov glavni namen shranjevanje in prenos dednega genskega materiala.
Skoraj vsaka celica v človeškem telesu ima jedro. Edina izjema so trombociti in rdeče krvničke. Večina celic je mononuklearnih, vendar imajo lahko na primer mišična vlakna in nevroni več teh organelov. Jedro v celici je lahko različnih velikosti - največje jedrske strukture v jajčecu.
Celično jedro: struktura
Jedro ima precej zapleteno zgradbo in je sestavljeno iz jedrske membrane, kromatina, nukleola in nukleoplazme. Oglejmo si vsak del podrobneje.
- Karioteka ali jedrska ovojnica je struktura, ki ločuje notranje okolje jedra od citoplazme. Ta lupina je sestavljena iz zunanje in notranje membrane, med katerimi je tako imenovani perinuklearni prostor. Zanimivo je, da zunanja membrana lupine prehaja neposredno v membrano zrnatega endoplazmatskega retikuluma, zato sta votlini rezervoarjev EPS in jedra med seboj povezani. In lupina ima jedrske pore, zaprte z diafragmo. Namenjeni so prodiranju v velike molekule, pa tudi za izmenjavo snovi med karioplazmo in citoplazmo.
- Karioplazma je homogena snov, ki zapolnjuje notranjo votlino jedra. Vsebuje nukleolus in kromatin.
- Kromatin je genetski material celice. Njegova strukturna enota je nukleosom, ki je veriga DNK, ovita okoli določenega proteina, histona. Genski material v celici ima dve stanji. Heterokromatin je majhna, gosta osmofilna zrnca. Evkromatin ali razrahljan kromatin so področja, v katerih aktivno potekajo sintetični procesi. Sčasoma se kromatin kondenzira in tvori kromosome.
- Jedro je majhna ovalna struktura, ki je sestavljena iz verig RNK in beljakovinskih molekul. Tu pride do tvorbe ribosomskih podenot. V jedru je lahko eno ali več nukleolov, vendar jih je mogoče videti le v celicah, ki se ne delijo.
Celično jedro: funkcije
Funkcije lahko določite tako, da se seznanite z njegovo strukturo. Prvič, jedro je odgovorno za prenos dednega niza informacij med celično delitvijo, tako mitozo kot mejozo. Med mitozo hčerinske celice prejmejo genom, ki je enak matični celici. Med mejozo (nastankom človeka) vsaka celica prejme le polovico kromosomskega nabora – celoten nabor kromosomov nastane šele po zlitju z zarodno celico drugega organizma.
Poleg tega je celično jedro odgovorno za eno najpomembnejših stopenj metabolizma - sintezo beljakovin. Dejstvo je, da se v jedru tvori informacija ali matrična RNA. Nato vstopi v endoplazmatski retikulum, se pritrdi na ribosom in služi kot model za nastanek aminokislinskega zaporedja peptidne molekule.
In kot že omenjeno, se sinteza ribosomskih podenot izvaja v jedru.
Celično jedro: izvor
Do danes obstaja več popolnoma različnih hipotez, s pomočjo katerih znanstveniki poskušajo pojasniti, kako natančno je nastalo jedro v celici. Toda na žalost nobena od teh izjav še ni našla dejanske potrditve.
Obstaja teorija, da je jedro kot celična struktura nastala kot posledica simbioze in arhej. Drugi znanstveniki verjamejo, da je jedro posledica okužbe celice z določenim virusom.
Najbolj popolna razlaga vsebuje tako imenovano eksomembransko hipotezo. Po njenih besedah je imela celica v procesu evolucije še eno zunanjo celično membrano. Istočasno se je stara, notranja membrana spremenila v lupino jedra - sčasoma je v njej nastal kompleksen sistem por, nato pa so se molekule kromatina začele koncentrirati v njeni votlini.
Vsi vemo, da smo ljudje evkarionti. To pomeni, da imajo vse njene celice organel, ki vsebuje vse genetske informacije – jedro. Vendar obstajajo izjeme. Ali v človeškem telesu obstajajo celice brez jedra in kakšen je njihov pomen za življenje?
Človeške celice brez jedra
Ne morejo se primerjati s prokarionti, ki imajo tipično strukturo. Kakšnih brezjedrnih ni v – eritrocitih. Namesto tega organela vsebujejo zapleten kemični kompleks snovi, ki jim omogoča opravljanje najpomembnejših funkcij v telesu. Tudi trombociti – trombociti in limfociti – so brezjedrne celice. V celicah ni jedra, imenujemo jih matične celice. Vse te strukture združuje še ena lastnost. Ker nimajo jedra, se ne morejo razmnoževati. To pomeni, da nejedrne celice, katerih primeri so bili navedeni, po opravljeni funkciji odmrejo, nove pa nastanejo v specializiranih organih.
rdeče krvne celice
Določajo barvo naše krvi. Krvne celice brez jedra, eritrociti, imajo nenavadno obliko - bikonkavni disk, ki znatno poveča njihovo površino pri relativno majhni velikosti. Toda njihovo število je preprosto neverjetno: na 1 kvadrat. mm njihove krvi je do 5 milijonov! V povprečju živi eritrocit do štiri mesece, nato pa odmre in se nevtralizira v vranici in jetrih. Vsako sekundo se v rdečem kostnem mozgu tvorijo nove celice.
Funkcije rdečih krvnih celic
Kaj vsebujejo te nejedrne celice namesto jedra? Te snovi se imenujejo hem in globin. Prvi vsebuje železo. Ne le obarva krvi rdeče, ampak tudi tvori nestabilne spojine s kisikom in ogljikovim dioksidom. Globin je beljakovinska snov. Hem, ki vsebuje nabit železov ion, je potopljen v svojo veliko molekulo. Po mehanizmu delovanja lahko te celice primerjamo s taksijem na fiksni poti. V pljučih dodajajo kisik. S krvnim tokom se prenaša v vse celice in se tam sprosti. S sodelovanjem kisika poteka proces oksidacije organskih snovi s sproščanjem določene količine energije, ki jo človek porabi za življenje. Izpraznjen prostor takoj zasede ogljikov dioksid, ki se pomakne v nasprotno smer – v pljuča, kjer se izdihne. Ta proces je nujen pogoj za življenje. Če celice ne dobijo kisika, pride do njihove postopne smrti. To je lahko življenjsko nevarno za organizem kot celoto.
Eritrociti opravljajo še eno pomembno funkcijo. Na njihovih membranah je beljakovinski marker, imenovan Rh faktor. Ta kazalnik je tako kot krvna skupina zelo pomemben pri transfuziji krvi, med nosečnostjo, darovanjem in kirurškimi posegi. Mora biti nameščen, saj lahko v primeru nezdružljivosti pride do tako imenovanega Rh konflikta. Je zaščitna reakcija, vendar lahko povzroči zavrnitev ploda ali organov.
Slaba prehrana, slabe navade, onesnažen zrak lahko povzročijo uničenje rdečih krvnih celic. Posledično se pojavi resna bolezen, ki se imenuje anemija ali slabokrvnost. V tem primeru oseba čuti omotico, šibkost, težko dihanje, tinitus. Pomanjkanje kisika negativno vpliva na telesno in duševno aktivnost osebe. Še posebej nevarno je med nosečnostjo. Če do ploda skozi popkovino ne pride dovolj kisika, lahko pride do resnih motenj v njegovem razvoju.
Struktura trombocitov
Nejedrne celice, trombocite, imenujemo tudi trombociti. V neaktivnem stanju imajo res ravno obliko, ki spominja na lečo. Ko pa so žile poškodovane, nabreknejo, okrogle, tvorijo nestabilne izrastke zunanje plasti - psevdopodije. Trombociti nastanejo v vranici in ne živijo dolgo - do 10 dni.
Proces nastajanja trombov
Trombocitni matriks vsebuje encim, imenovan tromboplastin. Če je celovitost posod kršena, se pojavi v plazmi. Pod njegovim delovanjem krvni protein protrombin preide v svojo aktivno obliko, nato pa deluje na fibrinogen. Posledično ta snov preide v netopno stanje. Pretvori se v beljakovino fibrin. Njegove niti so tesno prepletene in tvorijo trombo. Zaščitna reakcija koagulacije krvi preprečuje izgubo krvi. Vendar pa je nastanek krvnega strdka v žili zelo nevaren. To lahko privede do njegovega zloma in celo smrti telesa. Kršitev procesa strjevanja krvi se imenuje hemofilija. Za to dedno bolezen je značilno nezadostno število trombocitov in vodi do prekomerne izgube krvi.
stebelna celica
Te celice brez jedra z razlogom imenujemo matične celice. Pravzaprav so osnova za vse druge. Imenujejo jih tudi "gensko čiste". Matične celice najdemo v vseh tkivih in organih, največ pa jih vsebuje kostni mozeg. Po potrebi prispevajo k ponovni vzpostavitvi integritete. Stebelne se spremenijo v vse druge, ko so uničene. Zdi se, da bi moral človek ob prisotnosti takšnega čarobnega mehanizma živeti večno. Zakaj se to ne zgodi? Dejstvo je, da se s starostjo intenzivnost diferenciacije matičnih celic bistveno zmanjša. Niso več sposobni obnoviti uničenega tkiva. Obstaja pa tudi druga nevarnost. Obstaja velika verjetnost, da se matične celice spremenijo v rakave celice, kar bo neizogibno povzročilo smrt katerega koli živega organizma.
Celice brez jedra: primeri in razlike
V naravi so celice brez jedra precej pogoste. Na primer, modrozelene alge in bakterije so prokarionti. Toda v nasprotju s človeškimi celicami brez jedra ne umrejo, ko izpolnijo svojo biološko vlogo. Dejstvo je, da imajo prokarionti genetski material. Zato so sposobni delitve, ki se zgodi z izdelavo dveh genetskih kopij matične celice. Dedno informacijo prokariontov predstavlja krožna molekula DNK, ki se pred delitvijo podvoji. Ta analog jedra se imenuje tudi nukleoid. Pri rastlinah so žive celice brez jedra – sitaste cevi.
Človeške celice brez jedra se torej ne morejo deliti, zato obstajajo kratek čas, preden opravijo svojo funkcijo. Po tem pride do njihovega uničenja in znotrajcelične prebave. Sem spadajo oblikovani elementi (eritrociti), trombociti (trombociti) in izvorne celice.
Brown ni podal nobenih domnev o funkcijah jedra. Leta 1838 je Matthias Schleiden predlagal, da jedro sodeluje pri tvorbi novih celic, zato je uvedel izraz "citoblast" (graditelj celic), ki se nanaša na jedra. Prepričan je bil, da opazuje sestavljanje novih celic okoli »citoblastov«. Odločen nasprotnik tega stališča je bil Franz Meyen, ki je odkril, da se celice razmnožujejo z delitvijo, in verjel, da mnoge celice morda nimajo jedra. Ideja o tvorbi celic de novo , torej iz nič, prek citoblastov ali kako drugače, je bilo v nasprotju z delom Roberta Remacka (1852) in Rudolfa Virchowa (1855), ki sta končno potrdila novo paradigmo, da lahko celice nastanejo le iz celic ("Omnis cellula e cellula") . Funkcije jedra so ostale nejasne.
strukture
Jedro je največji organel v živalskih celicah. Pri sesalcih je premer jedra približno 6 µm, samo jedro pa predstavlja približno 10 % prostornine celice. Viskozna tekočina, ki napolnjuje jedro, se imenuje nukleoplazma in je kemično podobna citosolu, ki obdaja jedro.
Jedrska ovojnica in jedrske pore
Jedrska ovojnica je sestavljena iz dveh membran (zunanje in notranje), ki se nahajata vzporedno na razdalji 10 do 50 nm. Jedrska ovojnica popolnoma obdaja jedro, ločuje genetski material celice od citoplazme in služi kot ovira, ki preprečuje prosto difuzijo makromolekul med nukleoplazmo in citoplazmo. Zunanja jedrska membrana se nadaljuje v membrano grobega endoplazmatskega retikuluma (ER) in je obložena z ribosomi. Prostor med jedrnimi membranami se imenuje perinuklearni prostor in se nadaljuje v lumen ER.
Jedrske pore, ki so z vodo napolnjeni kanali v jedrski ovojnici, so sestavljene iz različnih beljakovin, imenovanih nukleoporini. Pri ljudeh je masa por približno 120.000 kDa, ki je 40-krat večja od mase ribosoma; hkrati je v jedrske pore pri kvasovkah vključenih približno 50 beljakovin, pri vretenčarjih pa nekaj sto. Čeprav je premer por 100 nm, je širina reže, skozi katero lahko prehajajo molekule, zaradi prisotnosti regulatornih sistemov znotraj por le 9 nm. V vodi topne majhne molekule lahko preidejo skozi takšno vrzel, ne pa tudi velike molekule, kot so nukleinske kisline in veliki proteini; za prenos teh molekul v jedro je potreben aktivni (to je energetsko potraten) transport. Na lupini jedra tipične sesalske celice je od 3000 do 4000 por, vsaka na stičišču dveh jedrskih membran pa ima obročasto strukturo z 8 simetričnimi osemi. Na obroč je pritrjena posebna struktura, znana kot jedrska košarica, ki štrli v nukleoplazmo, več njenih filamentov pa štrli v citoplazmo. Obe strukturi sta potrebni za posredovanje vezave transportnih jedrnih proteinov.
Večino beljakovin, ribosomskih podenot in nekaj DNK prenaša skozi jedrske pore družina transportnih faktorjev, znanih kot karioferini. Karioferini, ki posredujejo pri transportu v jedro, se imenujejo tudi importins, in posredovanje transporta iz jedra - eksportini. Večina karioferinov neposredno sodeluje s svojim tovorom, nekateri pa uporabljajo adapter veverice. Steroidni hormoni (kot sta kortizol in aldosteron) kot tudi druge majhne molekule, topne v maščobi, lahko difundirajo v citoplazmo v notranjost celice preko celične membrane; v citoplazmi se vežejo na proteinske jedrne receptorje, ki jih dostavijo v jedro. Tukaj jedrske receptorje vezani na svoje ligande delujejo kot transkripcijski faktorji, v odsotnosti liganda pa številni receptorji delujejo kot histonske deacetilaze, ki zavirajo izražanje določenih genov.
jedrska lamina
V živalskih celicah mehansko podporo jedru zagotavljata dve mreži vmesnih filamentov: jedrska lamina, ki je mreža vmesnih filamentov na notranji površini jedra, in manj organizirani filamenti na citosolni površini jedra. Oba sistema filamentov nudita podporo jedru in služita za sidranje kromosomov in jedrnih por.
Jedrska lamina je sestavljena predvsem iz beljakovin, znanih kot lamini. Kot vsi proteini se tudi lamini sintetizirajo v citoplazmi in se naprej transportirajo v notranjost jedra, kjer se vstavijo v jedrno lamino. Lamine, ki se nahajajo na zunanji strani jedrske ovojnice (kot npr emerin in nesprin), se vežejo na elemente citoskeleta, ki zagotavlja strukturno podporo jedru. Lamine najdemo tudi v nukleoplazmi, kjer tvorijo drugo pravilno strukturo, znano kot nukleoplazmatična tančica. nukleoplazmatsko tančico) ; slednje je mogoče prikazati s fluorescenčno mikroskopijo. Funkcija tančice ni znana, vendar je znano, da ne obstaja v nukleolu in je prisotna v interfazi celičnega cikla. Lamini, ki tvorijo tančico (kot je LEM3), se vežejo na kromatin in motnje v njihovi strukturi zavirajo prepisovanje genov, ki kodirajo beljakovine.
Tako kot drugi vmesni filamentni proteini tudi laminski monomeri vsebujejo α-vijačno domeno, ki jo dva monomera uporabljata za zvijanje drug okoli drugega, da tvorita dimer, ki ima strukturo bispirale. Dva dimera sta nadalje povezana s svojimi stranskimi ploskvami v antiparalelni orientaciji in tvorita tetramer, znan kot protofilament. Osem tetramerov je združenih v zvit, vrvi podoben filament. Filamente lahko sestavljamo in razstavljamo dinamično, kar pomeni, da je dolžina filamenta odvisna od relativne hitrosti njegovega sestavljanja in razstavljanja.
kromosomi
Jedro vsebuje večino genetskega materiala celice, ki je sestavljen iz številnih linearnih molekul DNK, ki so organizirane v strukture, znane kot kromosomi. Skupna dolžina molekul DNK v človeški celici je približno 2 m. Med večino celičnega cikla te molekule v kombinaciji z beljakovinami tvorijo tako imenovani kromatin, med celično delitvijo pa se kromosomi pojavljajo kot ločeni, dobro ločljivi kromosomi, ki sestavljajo kariotip. Majhna količina celičnega genskega materiala se nahaja v mitohondrijih in, v primeru rastlinske celice, v kloroplastih.
Obstajata dve vrsti kromatina. V evhromatinu je DNK najmanj gosto organizirana; vsebuje gene, ki se najpogosteje prepisujejo. Druga vrsta kromatina, heterokromatin, je bolj kompakten in vsebuje DNK, ki se redko ali nikoli ne prepisuje. Heterokromatin delimo na fakultativni, ki je prisoten samo v celicah določene vrste in na določeni stopnji celičnega cikla, in konstitutivni, ki ga predstavljajo kromosomske strukture, kot so telomeri in centromeri. Med interfazo kromatin vsakega kromosoma zaseda strogo določeno območje jedra - kromosomsko ozemlje. Aktivni geni, ki se ponavadi nahajajo v evhromatinu, se običajno nahajajo na meji kromosomskega ozemlja.
jedrska telesa
Jedro celic sesalcev vsebuje številne ločene podkompartmente, imenovane jedrna telesa. Izvajajo razdelitev jedra in v njem ustvarjajo ločene prostore, ki imajo določene lastnosti. Številna jedrska telesa opravljajo posebne funkcije, kot so sinteza in obdelava predribosomske RNA v nukleolusu, kopičenje in sestavljanje komponent spliceosoma v pegah (glej spodaj) ali kopičenje molekul RNA v paraspeckles. Mehanizmi, ki zagotavljajo izvajanje teh funkcij nukleolarnih teles, so zelo raznoliki. V nekaterih primerih lahko jedrsko telo služi kot mesto za določene procese, kot je transkripcija. V drugih primerih jedrska telesa očitno posredno uravnavajo lokalne koncentracije svojih komponent v nukleoplazmi. Tako kot citoplazemski organeli tudi jedrna telesa vsebujejo specifičen niz beljakovin, ki določajo njihovo strukturo na molekularni ravni. Vendar za razliko od citoplazemskih organelov jedrska telesa niso obdana z lipidnimi membranami, njihova strukturna celovitost pa je v celoti zagotovljena z interakcijami protein-protein in RNA-protein. V spodnji tabeli so navedene glavne značilnosti jedrskih teles.
| jedrsko telo | Funkcije | Značilne komponente | Tipična velikost (v µm) | Količina na jedro |
|---|---|---|---|---|
| nukleolus | Ribosomska biogeneza | Stroji RNA polimeraza I, procesni faktorji rRNA in sestavljanje ribosomske podenote | 3-8 | 1-4 |
| Pege | Kopičenje in sestavljanje spojnih faktorjev | Pre-mRNA faktorji spajanja | 2-3 | 20-50 |
| Stresna jedrska telesa | Regulacija transkripcije in spajanja pod stresom | HSF1, HAP | 1-2 | 3-6 |
| Telo histonskih lokusov | Predelava histonske mRNA | NPAT, FLASH, U7 snRNP | 0,2-1,2 | 2-4 |
| Telo Cajal | Biogeneza, zorenje in kroženje malih RNA | coilin, SMN | 0,2-1,5 | 1-10 |
| telo PML | Regulacija stabilnosti genoma, popravljanje DNA, nadzor transkripcije, zaščita pred virusi | PML | 0,1-1 | 10-30 |
| Paraspeckles | Regulacija mRNA, urejanje RNA | Nekodirajoče RNA NEAT1/MENε/β, proteini PSP1, p54 nrb /NONO | 0,2-1 | 2-20 |
| Perinukleolarni oddelek | Posttranskripcijska regulacija nabora sintetiziranih RNA RNA polimeraza III | PTB | 0,2-1 | 1-2 |
nukleolus
Nukleolus je ločena gosta struktura v jedru. Ni obdan z membrano in nastane na območju, kjer se nahaja rDNA – tandemske ponovitve genov ribosomske RNA (rRNA), imenovane nukleolarni organizatorji. Glavna funkcija nukleolusa je sinteza rRNA in tvorba ribosomov. Strukturna celovitost nukleolusa je odvisna od njegove aktivnosti, inaktivacija genov rRNA pa vodi do mešanice nukleolarnih struktur.
V prvi fazi tvorbe ribosoma encim RNA polimeraza I prepisuje rDNA in tvori pre-rRNA, ki se nadalje razreže na 5,8S, 18S in 28S rRNA. Transkripcija in post-transkripcijska obdelava rRNA potekata v nukleolusu s sodelovanjem majhnih nukleolarnih RNA (snoRNA), od katerih nekatere izvirajo iz spojenih intronov mRNA genov, ki kodirajo proteine, povezane s funkcijo ribosoma. Sestavljene ribosomske podenote so največje strukture, ki potekajo skozi jedrne pore.
Ko gledamo pod elektronskim mikroskopom, lahko v nukleolusu ločimo tri komponente: fibrilarne centre (FC), gosto fibrilarno komponento (CFC), ki jih obdaja, in zrnato komponento (GC), ki pa obdaja CFC. Transkripcija rRNA se pojavi v FC in na meji FC in PFC; zato postanejo FC, ko se aktivira tvorba ribosomov, jasno razločljivi. Rezanje in modifikacija rRNA se pojavita v PFC, naslednji koraki v tvorbi ribosomskih podenot, vključno z nalaganjem ribosomskih proteinov, pa se pojavijo v HA.
Telo Cajal
Cajalovo telo (TC) je jedrsko telo, ki ga najdemo pri vseh evkariontih. Prepoznamo ga po prisotnosti značilnega proteina coilin in specifičnih RNA (scaRNA). TK vsebuje tudi protein SMN. preživetje motoričnih nevronov). MA imajo visoko koncentracijo majhnih jedrskih ribonukleoproteinov (snRNP) in drugih procesnih faktorjev RNA, zato se domneva, da MA služijo kot mesta za sestavljanje in/ali post-transkripcijsko modifikacijo faktorjev spajanja. TK je prisoten v jedru med interfazo, vendar izgine med mitozo. V biogenezi TC se zasledijo lastnosti samoorganizirajoče strukture.
Ko so znotrajcelično lokalizacijo SMN prvič proučevali z imunofluorescenco, so protein našli v celotni citoplazmi, pa tudi v nukleolarnem telesu, ki je po velikosti podobno velikosti MC in pogosto ob MC. Zaradi tega so to telo poimenovali "TK dvojček" (angl. dvojčka CB) ali samo dragulj. Vendar se je izkazalo, da je celična linija HeLa, v kateri so odkrili novo telo, nenavadna: v drugih človeških celičnih linijah, pa tudi v vinski mušici Drosophila melanogaster SMN kolokaliziran s coilinom v TC. Zato lahko v splošnem primeru SMN obravnavamo kot pomembno komponento TC in ne kot marker posameznega jedrskega telesa.
Telo histonskih lokusov
telesa PML
Pege
Paraspeckles
Parapekle so nepravilno oblikovana jedrska telesa, ki se nahajajo v interkromatskem prostoru jedra. Najprej so jih opisali v celicah HeLa, ki imajo 10-30 parapekel na jedro, zdaj pa parapekle najdemo v vseh primarnih človeških celicah, v celicah transformiranih linij in na tkivnih odsekih. Ime so dobili zaradi svoje lokacije v jedru - blizu peg.
Parapekle so dinamične strukture, ki se spreminjajo kot odgovor na spremembe presnovne aktivnosti celice. Odvisni so od transkripcije in v odsotnosti transkripcije z RNA polimerazo II parapeke izginejo in vsi njihovi proteini (PSP1, p54nrb, PSP2, CFI(m)68 in PSF) tvorijo srpasto perinukleolarno kapico. Ta pojav opazimo med celičnim ciklom: paraspekle so prisotne v interfazi in vseh fazah mitoze razen telofaze. Med telofazo se oblikujejo hčerinska jedra, RNA polimeraza II pa ne prepisuje ničesar, zato beljakovine parapeke tvorijo perinukleolarni pokrovček. Parapeke sodelujejo pri uravnavanju izražanja genov s kopičenjem tistih RNA, kjer so dvoverižne regije, ki so predmet urejanja, namreč pretvorbe adenozina v inozin. Preko tega mehanizma so paraspeckles vključeni v nadzor izražanja genov med diferenciacijo, virusno okužbo in stresom.
Perinukleolarni oddelek
Perinukleolarni kompartment (OK) je nepravilno oblikovano jedrno telo, za katerega je značilno, da se nahaja na obodu nukleolusa. Čeprav sta fizično povezana, sta dva oddelka strukturno različna. TC se običajno nahajajo v celicah malignih tumorjev. OK je dinamična struktura in vsebuje veliko RNA-vezavnih proteinov in RNA polimeraze III. Strukturno stabilnost OK zagotavlja transkripcija, ki jo izvaja RNA polimeraza III, in prisotnost ključnih proteinov. Ker je prisotnost TC običajno povezana z malignostjo in sposobnostjo metastaziranja, se obravnavajo kot potencialni označevalci raka in drugih malignih tumorjev. Prikazana je bila povezanost OK s specifičnimi lokusi DNA.
Stresna jedrska telesa
Med toplotnim udarom v jedru nastanejo stresna jedrska telesa. Nastanejo z neposredno interakcijo transkripcijskega faktorja toplotnega šoka 1 ( HSF1) in pericentrične tandemske ponovitve v zaporedju satelita III, ki ustrezajo mestom aktivne transkripcije nekodirajočih transkriptov satelita III. Splošno prepričanje je, da takšna telesa ustrezajo zelo gosto zapakiranim oblikam ribonukleoproteinskih kompleksov. V celicah pod stresom se domneva, da so vključeni v hitre, prehodne in globalne spremembe izražanja genov prek različnih mehanizmov, kot je preoblikovanje kromatina in privzem faktorjev transkripcije in spajanja. V celicah v normalnih (ne stresnih) pogojih jedrska telesa pod stresom redko najdemo, vendar se njihovo število pod vplivom toplotnega šoka močno poveča. Stresna jedrska telesa najdemo le v celicah človeka in drugih primatov.
Jedrska telesa sirote
| jedrsko telo | Opis | Tipična velikost (v µm) | Količina na jedro |
|---|---|---|---|
| Klastosom | Koncentrira proteasomske komplekse 20S in 19S ter proteine, povezane z ubikvitinom. Najdemo ga predvsem, ko je aktivnost proteasoma stimulirana, in izgine, ko je aktivnost proteasoma zavrta. | 0,2-1,2 | 0-3 |
| Delitveni organ jasno telo) | Obogatena s faktorji cepitve CstF in CPSF, kot tudi beljakovine DDX1 ki vsebuje MRTVA škatla. Najdemo ga predvsem v S-fazi in nanj ne vpliva inhibicija transkripcije. | 0,2-1,0 | 1-4 |
| OPT domena | Obogatena s transkripcijskimi faktorji 1. okt in PTF. Delno se kolokalizira s transkripcijskimi mesti. Najdeno predvsem v pozni fazi G1, razstavljeno z inhibicijo transkripcije. | 1,0-1,5 | 1-3 |
| Polycomb telo | Najdemo ga v človeških celicah in celicah Drosophile, obogatene s proteinom PcG. Kopiči beljakovine RING1 pri ljudeh, BMI1, HPC, je lahko povezan s pericentromernim heterokromatinom. | 0,3-1,0 | 12-16 |
| Bik Sam68 | Kopiči beljakovino Sam68 in podobne beljakovine SLM-1 in SLM-2. Razstavljen z inhibicijo transkripcije. Verjetno bogat z RNA. | 0,6-1,0 | 2-5 |
| Telo SUMO | Obogaten s SUMO proteini in SUMO konjugacijskim encimom Ubc9. Koncentrira transkripcijske faktorje pCREB, CBP, c-junij. | 1-3 | 1-3 |
Funkcije
Jedrska ovojnica ščiti DNK celice in je v primerjavi s prokariontsko celico vključena v veliko bolj zapleteno regulacijo izražanja genov. Pri prokariontih sta transkripcija in translacija povezana procesa in translacija mRNA v beljakovino se začne, še preden je ta v celoti sintetizirana. V evkariontskih celicah sta citoplazma, v kateri poteka translacija, in transkripcija, ki poteka v jedru, prostorsko ločeni, zato je treba zagotoviti transport molekul med jedrom in citoplazmo.
Jedrska ovojnica daje jedru nadzor nad svojo vsebino in jo ločuje od ostale citoplazme. To je pomembno za uravnavanje procesov, ki potekajo na obeh straneh jedrske ovojnice. Kadar je treba citoplazmatski proces nekako omejiti, se običajno njegov ključni udeleženec prenese v jedro, kjer sodeluje s transkripcijskimi faktorji in tako sproži zatiranje tvorbe nekaterih encimov, ki sodelujejo v citoplazmatskem procesu. Tak regulacijski mehanizem ima na primer glikoliza, proces, pri katerem celica črpa energijo iz molekule glukoze. Prvo reakcijo glikolize izvede encim heksokinaza, ki pretvori molekulo glukoze v glukoza-6-fosfat. Ko se koncentracija fruktozo-6-fosfata (snov, ki nastane iz glukoza-6-fosfata med glikolizo) poveča, regulatorni protein pošlje heksokinazo v jedro, kjer tvori transkripcijski represivni kompleks, ki zavira izražanje genov, ki kodirajo glikolitične encime.
Za nadzor nad tem, kateri geni se prepisujejo, transkripcijski faktorji v celici nimajo fizičnega dostopa do DNK, dokler se ne aktivirajo na določeni signalni poti. To prepreči celo nizko izraženost napačnih genov. Zlasti v primeru genov, nadzorovanih z NF-κB, ki so vključeni v vnetni proces, je transkripcija inducirana s signalno potjo, na primer, ki se začne z vezavo signalne molekule TNF-α na njen receptor na celični membrani. in končno vodi do aktivacije transkripcijskega faktorja NF-κB. Jedrski lokalizacijski signal, ki je prisoten v NF-κB, omogoča prehod v jedro in iz njega skozi jedrske pore; v jedru spodbuja prepisovanje ciljnih genov.
Kompartmentalizacija preprečuje, da bi celica prepisala nerazcepljeno mRNA. Evkariontske mRNA vsebujejo introne, ki jih je treba odstraniti, preden se lahko začne prevajanje mRNA. Spajanje, to je odstranitev intronov, poteka v jedru, kar ribosomom izven jedra onemogoči dostop do pre-mRNA. Če ne bi bilo jedra, bi ribosomi začeli prevajati nezrelo mRNA, kar bi povzročilo nastanek nepravilnih beljakovinskih produktov.
Ker transkripcija poteka v jedru, jedro vsebuje veliko beljakovin, ki neposredno sodelujejo pri transkripciji ali jo uravnavajo. Ti proteini vključujejo helikaze, ki odvijajo dvojno vijačnico DNK in olajšajo dostop drugih beljakovin do nje, polimeraze RNK, ki sintetizirajo RNK, topoizomeraze, ki vplivajo na topologijo DNK, in različne transkripcijske faktorje.
jedrski transport
Izhod iz jedra in vstop v jedro velikih molekul nadzorujejo jedrske pore. Čeprav lahko majhne molekule vstopijo v jedro brez kakršne koli regulacije, se morajo makromolekule, kot so beljakovine in RNA, vezati na karioferine za transport v jedro (importini) in iz jedra (eksportini). Proteini, ki jih je treba prenesti iz citoplazme v jedro, vsebujejo specifično aminokislinsko zaporedje, znano kot jedrski lokalizacijski signal, na katerega se vežejo importini. Podobno vsebujejo beljakovine, ki morajo zapustiti jedro jedrski izvozni signal, ki ga priznava exportins. Sposobnost uvoznikov in izvoznikov, da prenašajo svoj tovor, uravnavajo GTPaze, encimi, ki hidrolizirajo GTP, da sprostijo energijo. Ključna GTPaza jedrskega transporta je tekel, ki se lahko veže na GTP ali GDP, odvisno od lokacije (v jedru ali v citoplazmi). V bistvu interakcija Ran-GTP z importinom povzroči konformacijsko spremembo slednjega, tako da se loči od prepeljanega tovora. Nastali kompleks Ran-GTP in importina se transportira v citoplazmo, kjer protein RanBP loči Ran-GTP od importina. Ločitev od importin omogoča beljakovine vrzel vzpostavi stik z Ran-GTP in katalizira hidrolizo GTP v BDP. Poleg tega beljakovina prepozna kompleks Ran-GDP NUTF2, ki ga vrne v nukleoplazmo. beljakovine v jedru GEF nadomesti BDP z GTP, tvori Ran-GTP in zapre cikel.
Montaža in demontaža
Med življenjem celice se lahko jedro razgradi (med celično delitvijo ali apoptozo). Med temi procesi se uničijo strukturne komponente jedra – jedrna ovojnica in jedrska lamina. V večini celic pride do razgradnje jedra med profazo mitoze. Vendar pa razstavljanje jedra ni strogo omejeno na mitozo in se ne pojavi v vseh celicah. Nekateri enocelični evkarionti (kot so kvasovke) so podvrženi tako imenovani zaprti mitozi, pri kateri ostane jedrska ovojnica nedotaknjena. Pri zaprti mitozi se kromosomi premaknejo na različne strani jedra, ki se nato razdeli na dvoje. V nasprotju s tem so celice višjih evkariontov običajno podvržene odprti mitozi, med katero razpade jedrska ovojnica. Kromosomi migrirajo na različne pole vretena in okoli njih se ponovno oblikujeta dve jedri. Jedrska lamina je tudi podvržena razgradnji zaradi fosforilacije laminov s kinazami, kot je npr. od ciklina odvisna protein kinaza 1. Sestavljanje jedrske lamine v hčerinskih jedrih se začne po defosforilaciji laminov.
Apoptoza je nadzorovan proces uničenja celičnih komponent, ki vodi v celično smrt. Spremembe, povezane z apoptozo, se pojavijo neposredno v jedru in njegovi vsebini. Ti vključujejo kondenzacijo kromatina, pa tudi razpad jedrske ovojnice in jedrske lamine. Razpad laminske mreže posredujejo apoptotične proteaze, znane kot kaspaze, ki razgrajujejo lamine in tako vplivajo na strukturno celovitost jedra. Uničenje lamina se včasih uporablja kot indikator aktivnosti kaspaze v študijah apoptoze. Celice, ki izražajo mutirane lamine, odporne na kaspazo, med apoptozo ne izgubijo jedrske celovitosti, zato imajo lamini ključno vlogo pri uvajanju sprememb, ki jim je jedro podvrženo med apoptozo. Poleg tega zaviranje sestavljanja laminov v mrežo sproži apoptozo.
Značilnosti jeder pri različnih evkariontih
Velikosti, oblike in morfologija evkariontskih jeder se zelo razlikujejo. če piroplazmid in Leishmania je premer jedra 1-3 mikronov, nato pa pri nekaterih radiolarijah premer jedra doseže 400 mikronov in celo 1 mm. Praviloma je oblika jedra pri večini evkariontov blizu sferične, včasih pa lahko prevzame precej bizarne oblike (to še posebej velja za makronukleuse ciliatov). Čeprav je pri vseh evkariontih lupina jedra sestavljena iz dveh membran, se število por v njem zelo razlikuje pri različnih vrstah, včasih pa se lahko pridružijo dodatni plasti (tako zunaj kot znotraj); na primer, pri mnogih prostoživečih amebah se vlaknasta plast s celično strukturo meji na notranjo stran lupine, ki po debelini znatno presega jedrsko lupino, pri radiolarijah pa se na zunanji strani lupine nahajajo dodatne fibrilarne plasti. .
Organizacijo jedra v protistih vrste Dinoflagellate (Dinoflagellata) odlikuje pomembna izvirnost. Večina njihovih predstavnikov ima jedro, v katerem so kromosomi zgoščeno v celotnem celičnem ciklu (tudi v interfazi) in so praktično brez histonov. Ta vrsta jedra se imenuje dinokarion. Hkrati je količina DNK v dinokarionu več deset in stokrat večja od količine DNK na celico pri predstavnikih drugih skupin evkariontov. Vendar pa nekateri dinoflagelati ( Noctiluca, Oodinium ) imajo pravilna evkariontska jedra; v drugih predstavnikih vrste v vegetativnih celicah so jedra navadna, dinokarion pa je prisoten na drugih stopnjah celičnega cikla (na primer v gametah).
Protistične celice imajo vsaj eno jedro. Hkrati se v organizmih Metazoa nahajajo tudi nejedrne celice, ki so brez jedra izgubile sposobnost delitve s tvorbo dveh hčerinskih celic. Najbolj znan primer celic brez jedra so eritrociti sesalcev, ki nimajo tudi drugih organelov, na primer mitohondrijev. Rdeče krvne celice zorijo v kostnem mozgu skozi proces eritropoeze, med katerim izgubijo jedra, druge organele in ribosome. Jedro se v procesu diferenciacije eritroblasta potisne iz celice v retikulocit, ki deluje kot neposredni predhodnik eritrocita. Pod vplivom nekaterih mutagenov se lahko v kri sprostijo nezreli eritrociti, ki vsebujejo mikronukleuse.
Večina protistov ima samo eno jedro; pri protistih, za katere je značilen kompleksen življenjski cikel (na primer, predstavniki vrste apikompleksa (Apicomplexa) imajo enojedrne in večjedrne stopnje.
Večjedrne protistične celice
Sestava kloroplastov v kriptofitih in kloarahniofitih vsebuje nukleomorf- reducirano jedro fototrofnega endosimbionta, ki so ga vgradili predniki teh alg med sekundarno endosimbiozo (pri Cryptophyta so bile vgrajene rdeče alge, pri Chlorarachnea pa zelene alge)
CELIČNO JEDRO
Jedro(nucleus, s. karyon) se nahaja v vseh človeških celicah, razen v eritrocitih in trombocitih. Naloge jedra so shranjevanje in prenos dednih informacij novim (hčerinskim) celicam. Te funkcije so povezane s prisotnostjo DNK v jedru. Sinteza beljakovin poteka tudi v jedru - ribonukleinska kislina RNA in ribosomski materiali.
V večini celic je jedro sferično ali jajčasto, obstajajo pa tudi druge oblike jedra (obročasto, paličasto, fuziformno, kroglasto, fižolasto, segmentirano, hruškasto, polimorfno). Velikost jedra je zelo različna - od 3 do 25 mikronov. Jajčece ima največje jedro. Večina človeških celic je mononuklearnih, vendar obstajajo dvojedrne (nekateri nevroni, hepatociti, kardiomiociti). Nekatere strukture so večjedrne (mišična vlakna). Jedro ima jedrno ovojnico, kromatin, nukleol in nukleoplazmo (slika 4).
Jedrska membrana ali karioteka (caryothéca), ki ločuje vsebino jedra od citoplazme, je sestavljena iz notranje in zunanje jedrske membrane, debeline vsaka po 8 nm. Membrani sta med seboj ločeni s perinuklearnim prostorom (kariotečna cisterna) širine 20-50 nm, ki vsebuje drobnozrnat material zmerne elektronske gostote. Zunanja jedrska membrana prehaja v granularni endoplazmatski retikulum. Poeto Mu perinuklearni prostor je ena sama votlina z endoplazmatskim retikulumom. Notranja jedrska membrana je znotraj povezana z obsežno mrežo beljakovinskih vlaken, ki jih sestavljajo posamezne podenote.
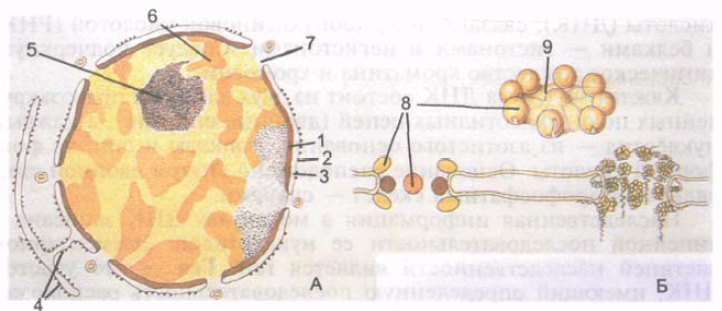
riž. 4. Celično jedro (A) in kompleks jedrnih por (B). 1 - notranja jedrska membrana; 2 - zunanja jedrska membrana; 3 - perinuklearni prostor; 4 - membrane endoplazmatskega retikuluma; 5 - nukleolus; 6 - dekondenzirani kromatin; 7 - jedrski čas; 8 - zrnca jedrske pore; 9 - diafragma jedrskih por.
Jedrska ovojnica vsebuje veliko zaobljenih jedrnih por s premerom 50–70 nm. Jedrne pore skupaj zasedajo do 25% površine jedra. Število por v enem jedru doseže 3000-4000. Ob robovih por sta zunanja in notranja membrana med seboj povezani in tvorita tako imenovani porni obroč. Vsaka pora je zaprta z diafragmo, imenovano tudi kompleks por. Diafragme por imajo zapleteno strukturo, tvorijo jih medsebojno povezane beljakovinske granule. Skozi jedrske pore se izvaja selektivni transport velikih delcev, pa tudi izmenjava snovi med jedrom in citosolom celice.
Pod jedrsko ovojnico so nukleoplazma (karioplazma)(nukleoplazma, s. karyoplasma), ki ima homogeno strukturo, in nukleolus. V nukleoplazmi nedelljivega jedra, v njegovem jedrskem proteinskem matriksu, so osmiofilna zrnca (grude) tako imenovanega heterokromatina. Področja bolj ohlapnega kromatina, ki se nahajajo med zrnci, se imenujejo evhromatin. Ohlapni kromatin se imenuje tudi dekondenzirani kromatin, sintetični procesi v njem potekajo najbolj intenzivno. Med delitvijo celic se kromatin zgosti, kondenzira in tvori kromosome.
Kromatin(kromatinu) nedelljivega jedra in kromosome delitvenega tvorijo molekule deoksiribonukleinske kisline (DNA), povezane z ribonukleinsko kislino (RNA) in proteini - histoni in nehistoni. Poudariti je treba kemijsko identiteto kromatina in kromosomov.
Vsaka molekula DNA je sestavljena iz dveh dolgih desnosučnih polinukleotidnih verig (dvojne vijačnice), vsak nukleotid pa je sestavljen iz dušikove baze, glukoze in ostanka fosforne kisline. Osnova se nahaja znotraj dvojne vijačnice, sladkorno-fosfatni skelet pa zunaj.
Dedna informacija v molekulah DNK je zapisana v linearnem zaporedju njenih nukleotidov. Osnovni delec dednosti je gen. Gen je del DNA, ki ima specifično zaporedje nukleotidov, odgovornih za sintezo enega določenega specifičnega proteina.
Molekula DNK v jedru je kompaktno zapakirana. Tako bi ena molekula DNK, ki vsebuje 1 milijon nukleotidov, z njihovo linearno razporeditvijo zasedla le 0,34 mm dolg segment. Dolžina enega človeškega kromosoma v raztegnjeni obliki je približno 5 cm, v stisnjenem stanju pa ima kromosom približno 10 -15 cm3.
Molekule DNA, povezane s histonskimi proteini, tvorijo nukleosome, ki so strukturne enote kromatina. Nukleosom ima obliko kroglice s premerom 10 nm. Vsak nukleosom je sestavljen iz histonov, okoli katerih je zvita 146 bp veriga DNK. Med nukleosomi so linearni odseki DNK, sestavljeni iz 60 parov nukleotidov.
Kromatin predstavljajo fibrile, ki tvorijo približno 0,4 μm dolge zanke, ki vsebujejo od 20.000 do 30.000 baznih parov.
Zaradi zbijanja (kondenzacije) in zvijanja (superspecializacije) deoksiribonukleoproteinov (DNP) v delitvenem jedru postanejo kromosomi vidni. Te strukture so kromosomi(kromasomi, iz grščine chruma - barva, soma - telo) - so podolgovate paličaste tvorbe z dvema krakoma, ločenima s tako imenovano zožitvijo - centromero. Glede na lokacijo centromera ter relativni položaj in dolžino krakov (nog) ločimo tri vrste kromosomov: metacentrične, ki imajo približno enake krake; submetacentrični, pri katerem je dolžina ramen drugačna; akrocentrično, pri katerem je eno rame dolgo, drugo pa zelo kratko, komaj opazno. Kromosom vsebuje eu- in heterokromatske regije. Slednji v nedelljivem jedru in v zgodnji profazi mitoze ostanejo kompaktni. Za identifikacijo kromosomov se uporablja menjavanje eu- in heterokromatskih regij.
Površina kromosomov je prekrita z različnimi molekulami, predvsem ribonukleoproteini (RNP). Somatske celice imajo 2 kopiji vsakega kromosoma, imenujemo jih homologne. Enaki so po dolžini, obliki, strukturi, nosijo iste gene, ki se nahajajo na enak način. Strukturne značilnosti, število in velikost kromosomov imenujemo kariotip. Normalni človeški kariotip vključuje 22 parov avtosomov in en par spolnih kromosomov (XX ali XY). Somatske človeške celice (diploidne) imajo dvojno število kromosomov - 46. Spolne celice vsebujejo haploiden (enojni) niz - 23 kromosomov. Zato je DNK v zarodnih celicah 2-krat manj kot v diploidnih somatskih celicah.
nukleolus(nucleolus), enega ali več, najdemo v vseh celicah, ki se ne delijo. Ima obliko intenzivno obarvanega okroglega telesa, katerega velikost je sorazmerna z intenzivnostjo sinteze beljakovin. Jedro je sestavljeno iz elektronsko gostega nukleolonema (iz grške pete - nit), v katerem se razlikuje filamentni (fibrilarni) del, sestavljen iz številnih prepletajočih se verig RNK debeline približno 5 nm, in zrnat del. Zrnati (granularni) del tvorijo zrna s premerom okoli 15 nm, ki so delci RNP – prekurzorji ribosomskih podenot. Perinukleolarni kromatin je vgrajen v vdolbine nukleolonema. Ribosomi nastanejo v nukleolu.
DELITEV CELICE. CELIČNI CIKLUS
Organizem raste tako, da z delitvijo povečuje število celic. Glavni metodi delitve celic v človeškem telesu sta mitoza in mejoza. Procesi, ki se pojavljajo pri teh metodah delitve celic, potekajo na enak način, vendar vodijo do različnih rezultatov.
Mitotična celična delitev(mitoza) vodi do povečanja števila celic, rasti organizma. Na ta način je zagotovljena obnova celic, ko se iztrošijo in odmrejo. Zdaj je znano, da epidermalne celice živijo 10-30 dni, eritrociti - do 4-5 mesecev. Živčne in mišične celice (vlakna) živijo vse človekovo življenje.
V vseh celicah med razmnoževanjem (delitvijo) opazimo spremembe, ki se umeščajo v okvir celičnega cikla. celični cikel imenujemo procesi, ki potekajo v celici od delitve do delitve ali od delitve do smrti (odmiranja) celice. V celičnem ciklu ločimo pripravo celice na delitev (interfaza) in mitozo (proces celične delitve).
V interfazi, ki traja približno 20-30 ur, se poveča hitrost biosintetskih procesov in poveča se število organelov. V tem času se masa celice in vseh njenih strukturnih komponent, vključno s centrioli, podvoji.
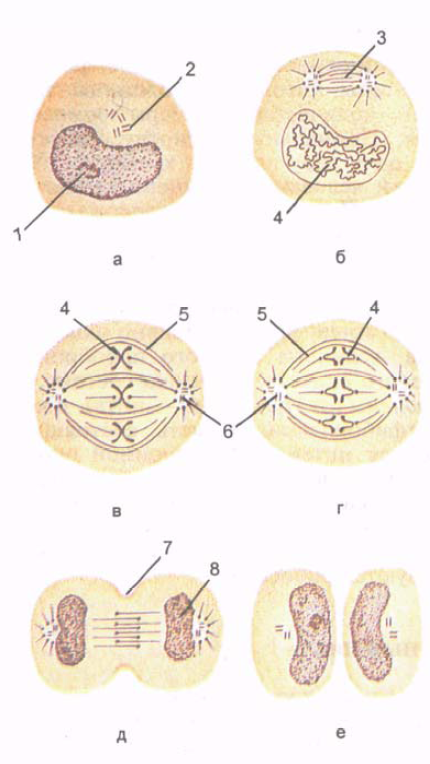
riž. 5. Celična delitev. faze mitoze. Prikazani so nastanek kromosomov, nastanek delitvenega vretena in enakomerna porazdelitev kromosomov in centriolov v dveh hčerinskih celicah.
A - medfaza; B - profaza; B - metafaza; G - anafaza; D - telofaza; E - pozna telofaza. 1 - jedro; 2 - celični center (centriole); 3 - vreteno delitve celic; 4 - kromosomi; 5 - neprekinjene mikrotubule; 6 - celični center; 7 - brazda celične delitve; 8 - tvorba jedra.
Pride do replikacije (ponovitve, podvojitve) molekul nukleinske kisline. To je proces prenosa genetskih informacij, shranjenih v DNK staršev, z natančno reprodukcijo v hčerinskih celicah. Starševska veriga DNK služi kot predloga za sintezo hčerinske DNK. Zaradi replikacije je vsaka od dveh hčerinskih molekul DNK sestavljena iz ene stare in ene nove verige. V obdobju priprave na mitozo se v celici sintetizirajo beljakovine, potrebne za celično delitev. Do konca interfaze se kromatin v jedru kondenzira.
Mitoza(mitoza; iz grščine mitos - nit) je obdobje, ko se matična celica razdeli na dve hčerinski celici (slika 5). Mitotična celična delitev zagotavlja enakomerno porazdelitev celičnih struktur, njene jedrske snovi - kromatina - med dvema hčerinskima celicama. Trajanje mitoze je od 30 minut do 3 ur.Mitozo delimo na profazo, metafazo, anafazo, telofazo.
V profazi nukleolus postopoma razpade, centrioli se razhajajo proti polom celice. Mikrotubuli centriolov so usmerjeni proti ekvatorju, v območju ekvatorja pa se med seboj prekrivajo.
V metafazi se jedrska ovojnica uniči, kromosomske niti se pošljejo na poli in ohranjajo povezavo z ekvatorjem.
celotnem območju celice. Strukture endoplazemskega retikuluma in Golgijevega kompleksa razpadejo na majhne vezikle (mehurčke), ki se skupaj z mitohondriji porazdelijo v obe polovici deleče se celice. Na koncu metafaze se začne vsak kromosom z vzdolžno razcepom cepiti na dva nova hčerinska kromosoma.
V anafazi se kromosomi ločijo drug od drugega in razhajajo proti polom celice s hitrostjo do 0,5 µm/min. Na koncu anafaze se plazemska membrana invaginira vzdolž ekvatorja celice pravokotno na njeno vzdolžno os in tvori cepitveno brazdo.
V telofazi se kromosomi, ki so se oddaljili do polov celice, dekondenzirajo, preidejo v kromatin in začne se transkripcija (proizvodnja) RNK. Oblikuje se jedrska ovojnica, nukleolus in hitro nastanejo membranske strukture bodočih hčerinskih celic. Na površini celice, vzdolž njenega ekvatorja, se zožitev poglobi, celica se razdeli na dve hčerinski celici.
Zaradi mitotske delitve hčerinske celice prejmejo nabor kromosomov, enak matičnemu. Mitoza zagotavlja genetsko stabilnost, povečanje števila celic in posledično rast organizma ter procese regeneracije.
Mejoza(iz grške mejoze - zmanjšanje) opazimo v zarodnih celicah. Zaradi delitve teh celic nastanejo nove celice z enim samim (haploidnim) nizom kromosomov, ki je pomemben za prenos genetske informacije. Ko se ena spolna celica združi s celico nasprotnega spola (med oploditvijo), se nabor kromosomov podvoji, postane popoln, dvojni (diploiden). V diploidni (dvojedrni) zigoti, ki nastane po zlitju zarodnih celic, sta dva niza enakih (homolognih) kromosomov. Vsak par homolognih kromosomov diploidnega organizma (zigote) izhaja iz jedra jajčeca in iz jedra semenčice.
Zaradi mejoze zarodnih celic v zrelem organizmu vsaka hčerinska celica vsebuje samo enega od vseh parov homolognih kromosomov prvotnih celic. To postane mogoče, ker se med mejozo zgodi samo replikacija DNA in dve zaporedni delitvi jedra. Posledično iz ene diploidne celice nastaneta dve haploidni celici. Vsaka od teh hčerinskih celic vsebuje polovico manj kromosomov (23) kot v jedru matične celice (46). Zaradi mejoze imajo haploidne spolne celice ne le prepolovljeno število kromosomov, ampak tudi drugačno razporeditev genov na kromosomih. Zato novi organizem ne nosi le vsote značilnosti svojih staršev, temveč tudi lastne (individualne) značilnosti.
Vprašanja za ponavljanje in samokontrolo
1. Iz katerih elementov je sestavljeno celično jedro? Katere funkcije opravlja?
2. Povejte nam o strukturi molekul DNA.
3. Podajte morfološke značilnosti kromosomov, podajte njihovo razvrstitev.
4. Kaj je celični cikel, katera obdobja (faze) se razlikujejo v tem ciklu?
5. Kaj je mejoza, kako se razlikuje od mitoze?