גרעין התא הוא המבנה החיוני של כל אורגניזם אוקריוטי. מבצע מגוון רחב של פונקציות, אך מטרתו העיקרית היא אחסון והעברה של חומר גנטי תורשתי.
כמעט לכל תא בגוף האדם יש גרעין. החריגים היחידים הם טסיות דם ותאי דם אדומים. רוב התאים הם חד-גרעיניים, אבל, למשל, לסיבי שריר ונוירונים עשויים להיות כמה מהאברונים הללו. לגרעין בתא יכולים להיות גדלים שונים - המבנים הגרעיניים הגדולים ביותר בביצית הנקבה.
גרעין התא: מבנה
לגרעין מבנה מורכב למדי והוא מורכב מהממברנה הגרעינית, כרומטין, נוקלאולוס ונוקלאופלזמה. בואו נסתכל על כל חלק שלו ביתר פירוט.
- הקאריוטקה, או המעטפת הגרעינית, היא המבנה המפריד בין הסביבה הפנימית של הגרעין לציטופלזמה. קליפה זו מורכבת ממברנות חיצוניות ופנימיות, שביניהם יש מה שנקרא חלל פרי-גרעיני. באופן מעניין, הממברנה החיצונית של הקליפה עוברת ישירות לתוך הממברנה של הרשת האנדופלזמית הגרנורית, כך שהחללים של מיכלי ה-EPS והגרעין מחוברים זה לזה. ולקליפה יש נקבוביות גרעיניות סגורות על ידי דיאפרגמה. הם מיועדים לחדירה למולקולות גדולות, כמו גם לחילופי חומרים בין הקריופלזמה לציטופלזמה.
- קריופלזמה היא חומר הומוגני הממלא את החלל הפנימי של הגרעין. הוא מכיל את הגרעין והכרומטין.
- כרומטין הוא החומר הגנטי של התא. היחידה המבנית שלו היא הנוקלאוזום, שהוא גדיל DNA הכרוך סביב חלבון ספציפי, ההיסטון. ישנם שני מצבים של חומר גנטי בתא. הטרוכרומטין הוא גרגירים אוסמופיליים קטנים וצפופים. Euchromatin, או כרומטין משוחרר, הם אזורים שבהם מתרחשים באופן פעיל תהליכים סינתטיים. עם הזמן, הכרומטין מתעבה ויוצר כרומוזומים.
- הגרעין הוא מבנה סגלגל קטן המורכב מגדילים של מולקולות RNA וחלבון. כאן מתרחשת היווצרות תת-יחידות ריבוזום. ייתכן שיש גרעין אחד או יותר בגרעין, אך ניתן לראות אותם רק בתאים שאינם מתחלקים.
גרעין התא: פונקציות
ניתן לקבוע פונקציות על ידי היכרות עם המבנה שלה. ראשית, הגרעין אחראי להעברת מערך המידע התורשתי במהלך חלוקת התא, הן מיטוזה והן מיוזה. במהלך מיטוזה, תאי הבת מקבלים גנום זהה לתא האב. במהלך המיוזה (היווצרות אנושית), כל תא מקבל רק מחצית ממערך הכרומוזומים - קבוצה שלמה של כרומוזומים נוצרת רק לאחר איחוי עם תא הנבט של אורגניזם אחר.
בנוסף, גרעין התא אחראי לאחד השלבים החשובים ביותר של חילוף החומרים – סינתזת חלבון. העובדה היא שבגרעין נוצר מידע, או RNA מטריקס. לאחר מכן הוא נכנס לרטיקולום האנדופלזמי, נצמד לריבוזום ומשמש מודל ליצירת רצף חומצות האמינו של מולקולת הפפטידים.
וכפי שכבר הוזכר, הסינתזה של תת-יחידות ריבוזום מתבצעת בגרעין.
גרעין התא: מקור
נכון להיום ישנן כמה השערות שונות לחלוטין, בעזרתן מנסים מדענים להסביר כיצד בדיוק נוצר הגרעין בתא. אבל, למרבה הצער, אף אחת מההצהרות הללו עדיין לא מצאה אישור ממשי.
קיימת תיאוריה שהגרעין כמבנה תאי נוצר כתוצאה מסימביוזה וארכיאה. מדענים אחרים מאמינים שהגרעין הוא תוצאה של הדבקה של התא בנגיף ספציפי.
ההסבר השלם ביותר מכיל את מה שנקרא השערת אקסוממברנה. לדבריה, בתהליך האבולוציה היה לתא קרום תא חיצוני נוסף. במקביל, הקרום הישן והפנימי הפך לקליפה של הגרעין - עם הזמן נוצרה בו מערכת מורכבת של נקבוביות, ואז החלו להתרכז מולקולות כרומטין בחלל שלה.
כולנו יודעים שבני אדם הם אוקריוטים. זה אומר שלכל התאים שלו יש אברון שמכיל את כל המידע הגנטי - הגרעין. עם זאת, ישנם יוצאי דופן. האם ישנם תאים נטולי גרעין בגוף האדם ומה משמעותם לחיים?
תאים אנושיים נטולי גרעין
לא ניתן להשוות אותם עם פרוקריוטים, בעלי מבנה אופייני. איזה סוג של נטולי גרעין אין - אריתרוציטים. במקום אברון זה, הם מכילים קומפלקס כימי מורכב של חומרים המאפשר להם לבצע את הפונקציות החשובות ביותר עבור הגוף. טסיות דם - טסיות ולימפוציטים - הן גם תאים לא גרעיניים. אין גרעין בתאים, הנקראים תאי גזע. כל המבנים הללו מאוחדים על ידי תכונה אחת נוספת. מכיוון שחסר להם גרעין, הם אינם מסוגלים להתרבות. המשמעות היא שתאים שאינם גרעיניים, שדוגמאות לכך ניתנו, מתים לאחר ביצוע תפקידם, ותאים חדשים נוצרים באיברים מיוחדים.
תאי דם אדומים
הם קובעים את צבע הדם שלנו. לתאי דם נטולי גרעין, אריתרוציטים, יש צורה יוצאת דופן - דיסק דו קעורה, אשר מגדיל באופן משמעותי את פני השטח שלהם בגודל קטן יחסית. אבל המספר שלהם פשוט מדהים: בריבוע אחד. מ"מ מהדם שלהם הוא עד 5 מיליון! בממוצע, אריתרוציט חי עד ארבעה חודשים, ולאחר מכן הוא מת ומנוטרל בטחול ובכבד. תאים חדשים נוצרים בכל שנייה במח העצם האדום.
פונקציות של תאי דם אדומים
מה מכילים התאים הלא-גרעיניים האלה במקום גרעין? חומרים אלו נקראים hem וגלובין. הראשון מכיל ברזל. זה לא רק מכתים את הדם באדום, אלא גם יוצר תרכובות לא יציבות עם חמצן ופחמן דו חמצני. גלובין הוא חומר חלבוני. Hem המכיל יון ברזל טעון שקוע במולקולה הגדולה שלו. על פי מנגנון הפעולה, ניתן להשוות תאים אלו למונית במסלול קבוע. לריאות הם מוסיפים חמצן. עם זרימת הדם, הוא נישא לכל התאים ומשוחרר שם. בהשתתפות חמצן, תהליך החמצון של חומרים אורגניים מתרחש עם שחרור של כמות מסוימת של אנרגיה שאדם משתמש בה כדי לבצע חיים. את החלל המתפנה תופס פחמן דו חמצני מיד, שנע בכיוון ההפוך - אל הריאות, שם הוא נשוף. תהליך זה הוא תנאי הכרחי לחיים. אם לא מסופק חמצן לתאים, מותם ההדרגתי מתרחש. זה יכול להיות סכנת חיים עבור האורגניזם בכללותו.
אריתרוציטים ממלאים תפקיד חשוב נוסף. על הממברנות שלהם סמן חלבון הנקרא גורם Rh. מדד זה, כמו סוג הדם, חשוב מאוד במהלך עירוי דם, במהלך הריון, תרומה ופעולות כירורגיות. זה חייב להיות מותקן, כי במקרה של חוסר תאימות, מה שנקרא קונפליקט Rh יכול להתרחש. זוהי תגובה מגנה, אך עלולה להוביל לדחייה של העובר או האיברים.
תזונה לקויה, הרגלים רעים, אוויר מזוהם עלולים לגרום להרס של תאי דם אדומים. כתוצאה מכך, מתרחשת מחלה קשה, הנקראת אנמיה, או אנמיה. במקרה זה, האדם מרגיש סחרחורת, חולשה, קוצר נשימה, טינטון. מחסור בחמצן משפיע לרעה על הפעילות הגופנית והנפשית של האדם. זה מסוכן במיוחד במהלך ההריון. אם לא מספקים מספיק חמצן לעובר דרך חבל הטבור, הדבר עלול להוביל להפרעות חמורות בהתפתחותו.
מבנה הטסיות
תאים לא גרעיניים, טסיות דם, נקראים גם טסיות. במצב לא פעיל, באמת יש להם צורה שטוחה, המזכירה עדשה. אבל כשהכלים נפגעים, הם מתנפחים, עגולים, יוצרים יציאות לא יציבות של השכבה החיצונית - פסאודופודיה. טסיות הדם נוצרות ואינן חיות זמן רב - עד 10 ימים, כשהן מנוטרלות בטחול.
תהליך היווצרות פקקת
מטריצת הטסיות מכילה אנזים הנקרא טרומבופלסטין. אם שלמות הכלים מופרת, היא מופיעה בפלזמה. תחת פעולתו, חלבון הדם פרוטרומבין עובר לצורתו הפעילה, בתורו, פועל על פיברינוגן. כתוצאה מכך, חומר זה עובר למצב בלתי מסיס. זה הופך לחלבון פיברין. החוטים שלו שלובים זה בזה ויוצרים פקקת. התגובה המגנה של קרישת דם מונעת איבוד דם. עם זאת, היווצרות קריש דם בתוך הכלי מסוכנת מאוד. זה יכול להוביל לקרע שלו ואפילו למוות של הגוף. הפרה של תהליך הקרישה נקראת המופיליה. מחלה תורשתית זו מאופיינת במספר לא מספיק של טסיות דם ומובילה לאיבוד דם מוגזם.
תאי גזע
תאים נטולי גרעין אלה נקראים תאי גזע מסיבה כלשהי. הם אכן הבסיס לכל השאר. הם נקראים גם "טהורים מבחינה גנטית". תאי גזע נמצאים בכל הרקמות והאיברים, אך מח העצם מכיל את המרב. הם תורמים לשיקום שלמות במידת הצורך. הגזעים הופכים לכל השאר כשהם מושמדים. נראה כי בנוכחות מנגנון קסום כזה, אדם צריך לחיות לנצח. למה זה לא קורה? העניין הוא שעם הגיל, עוצמת ההתמיינות של תאי הגזע יורדת משמעותית. הם אינם מסוגלים עוד לשחזר את הרקמה ההרוסה. אבל יש גם סכנה אחרת. ישנה סבירות גבוהה שתאי גזע יהפכו לתאים סרטניים, מה שיוביל בהכרח למוות של כל אורגניזם חי.
תאים נטולי גרעין: דוגמאות והבדלים
בטבע, תאים נטולי גרעין נפוצים למדי. לדוגמה, אצות כחולות ירוקות וחיידקים הם פרוקריוטים. אבל, בניגוד לתאים אנושיים נטולי גרעין, הם אינם מתים לאחר מילוי תפקידם הביולוגי. העובדה היא שלפרוקריוטים יש חומר גנטי. לכן, הם מסוגלים להתחלק, מה שמתרחש על ידי יצירת שני עותקים גנטיים של תא האם. המידע התורשתי של פרוקריוטים מיוצג על ידי מולקולת DNA מעגלית, המוכפלת לפני חלוקתה. אנלוגי זה של הגרעין נקרא גם נוקלואיד. בצמחים, תאים חיים הם נטולי גרעין - צינורות מסננת.
אז, תאים אנושיים נטולי גרעין אינם מסוגלים להתחלק, ולכן הם קיימים לפרק זמן קצר לפני ביצוע תפקידם. לאחר מכן, הרס שלהם ועיכול תוך תאי מתרחשים. אלה כוללים אלמנטים שנוצרו (אריתרוציטים), טסיות דם (טסיות) ותאי גזע.
בראון לא הניח הנחות לגבי תפקידי הגרעין. בשנת 1838, מתיאס שליידן הציע שהגרעין מעורב ביצירת תאים חדשים, אז הוא הציג את המונח "ציטובבלסט" (בונה תאים) כדי להתייחס לגרעינים. הוא היה בטוח שהוא צופה בהרכבה של תאים חדשים סביב "ציטובלסטים". מתנגד נחרץ לדעה זו היה פרנץ מיין, שגילה שתאים מתרבים בחלוקה, והאמין שלתאים רבים אולי אין גרעין. הרעיון של היווצרות תאים דה נובו , כלומר מאפס, דרך ציטובלסטים או אחר, סתרה את עבודתם של רוברט רמאק (1852) ורודולף וירצ'וב (1855), שאישרו לבסוף את הפרדיגמה החדשה לפיה תאים יכולים להיווצר רק מתאי ("Omnis cellula e cellula") . תפקידי הליבה נותרו לא ברורים.
מבנים
הגרעין הוא האברון הגדול ביותר בתאי בעלי חיים. אצל יונקים, קוטר הגרעין הוא בערך 6 מיקרומטר, והגרעין עצמו מהווה כ-10% מנפח התא. הנוזל הצמיג הממלא את הגרעין נקרא נוקלאופלזמה והוא דומה מבחינה כימית לציטוזול המקיף את הגרעין.
מעטפת גרעינית ונקבוביות גרעיניות
המעטפת הגרעינית מורכבת משני ממברנות (חיצוניות ופנימיות), הממוקמות במקביל במרחק של 10 ל 50 ננומטר. המעטפת הגרעינית מקיפה לחלוטין את הגרעין, מפרידה את החומר הגנטי של התא מהציטופלזמה ומשמשת מחסום המונע דיפוזיה חופשית של מקרומולקולות בין הנוקלאופלזמה לציטופלזמה. הממברנה הגרעינית החיצונית ממשיכה לתוך קרום הרשת האנדופלזמית המחוספסת (ER) ומרופדת בריבוזומים. החלל בין הממברנות הגרעיניות נקרא החלל הפרי-גרעיני וממשיך לתוך לומן ה-ER.
נקבוביות גרעיניות, שהן תעלות מלאות מים במעטפת הגרעינית, מורכבות ממגוון חלבונים הנקראים נוקלאופורינים. בבני אדם, מסת הנקבוביות היא בערך 120,000 kDa, שהיא פי 40 מהמסה של הריבוזום; במקביל, כ-50 חלבונים כלולים בנקבוביות הגרעיניות בשמרים, וכמה מאות בבעלי חוליות. למרות שקוטר הנקבוביות הוא 100 ננומטר, רוחב הרווח שדרכו מולקולות יכולות לעבור, עקב נוכחותן של מערכות רגולטוריות בתוך הנקבוביות, הוא 9 ננומטר בלבד. מולקולות קטנות מסיסות במים יכולות לעבור דרך פער כזה, אבל לא מולקולות גדולות כמו חומצות גרעין וחלבונים גדולים; נדרשת הובלה פעילה (כלומר, גוזלת אנרגיה) כדי להעביר את המולקולות הללו לגרעין. על קליפת הגרעין של תא יונק טיפוסי, יש בין 3,000 ל-4,000 נקבוביות, ולכל אחת מהן במפגש של שני ממברנות גרעיניות יש מבנה טבעת עם 8 צירי סימטריה. לטבעת מחובר מבנה מיוחד המכונה הסל הגרעיני, הבולט לתוך הנוקלאופלזמה, וכמה מחוטי החוטים שלו בולטים לתוך הציטופלזמה. שני המבנים נדרשים כדי לתווך את הקישור של חלבוני תחבורה גרעיניים.
רוב החלבונים, יחידות המשנה של הריבוזומים וחלק מה-DNA מועברים דרך נקבוביות גרעיניות על ידי משפחה של גורמי הובלה הידועים בשם קריופרינים. קריופרינים המתווכים הובלה לגרעין נקראים גם יבואנים, ותיווך הובלה מהגרעין - יצואנים. רוב הקאריופרינים מקיימים אינטראקציה ישירה עם המטען שלהם, אך חלקם משתמשים מַתאֵםסנאים. הורמונים סטרואידים (כגון קורטיזול ואלדוסטרון) כמו גם מולקולות קטנות מסיסות בשומן יכולים להתפזר לתוך הציטופלזמה אל פנים התא על פני קרום התא; בציטופלזמה, הם נקשרים לקולטנים גרעיניים של חלבון, המעבירים אותם לגרעין. כאן קולטנים גרעינייםהקשורים לליגנדים שלהם מתפקדים כגורמי שעתוק, ובהיעדר ליגנד, קולטנים רבים מתפקדים כ-Histone deacetylases המדכאים את הביטוי של גנים מסוימים.
למינה גרעינית
בתאי בעלי חיים, התמיכה המכנית של הגרעין מסופקת על ידי שתי רשתות של חוטי ביניים: הלמינה הגרעינית, שהיא רשת של חוטי ביניים על פני השטח הפנימיים של הגרעין, וחוטים פחות מאורגנים על פני השטח הציטוסוליים של הגרעין. שתי מערכות החוטים מספקות תמיכה לגרעין ומשמשות לעיגון כרומוזומים ונקבוביות גרעיניות.
הלמינה הגרעינית מורכבת בעיקר מחלבונים המכונים למינים. כמו כל החלבונים, הלמינים מסונתזים בציטופלזמה ומועברים הלאה אל פנים הגרעין, שם הם מוכנסים אל הלמינה הגרעינית. למינים הממוקמים בצד החיצוני של המעטפת הגרעינית (כגון אמריןו נספרין), נקשרים לאלמנטים של שלד הציטו, המספק תמיכה מבנית לגרעין. למינים נמצאים גם בנוקלאופלזמה, שם הם יוצרים מבנה רגיל נוסף המכונה צעיף הנוקלאופלזמה. צעיף נוקלאופלזמי) ; את האחרון ניתן להמחיש באמצעות מיקרוסקופ פלואורסצנטי. תפקידו של הצעיף אינו ידוע, אך ידוע שהוא אינו קיים בגרעין ונמצא ב-interphase של מחזור התא. למינים המרכיבים את הצעיף (כגון LEM3) נקשרים לכרומטין, והפרעות במבנה שלהם מדכאות את השעתוק של גנים מקודדי חלבון.
כמו חלבוני נימה ביניים אחרים, מונומרים למינים מכילים תחום α-סלילי, המשמש את שני המונומרים כדי להתפתל זה סביב זה ליצירת דימר, בעל המבנה ביספירלים. שני הדימרים מחוברים עוד יותר על ידי פני הצד שלהם בכיוון אנטי-מקביל, ויוצרים טטרמר המכונה פרוטופילמנט. שמונה טטרמרים משולבים לתוך חוט מעוות דמוי חבל. ניתן להרכיב ולפרק חוטים באופן דינמי, כלומר אורכו של חוט נימה תלוי במהירויות היחסיות של ההרכבה והפירוק שלו.
כרומוזומים
הגרעין מכיל את רוב החומר הגנטי של התא, המורכב ממולקולות DNA ליניאריות רבות המאורגנות למבנים המכונים כרומוזומים. האורך הכולל של מולקולות ה-DNA בתא אנושי הוא כ-2 מ'. במהלך רוב מחזור התא, מולקולות אלו בשילוב עם חלבונים יוצרות את מה שנקרא כרומטין, ובמהלך חלוקת התא, הכרומוזומים מופיעים ככרומוזומים נפרדים ומובחנים היטב המרכיבים את הקריוטיפ. כמות קטנה של חומר גנטי תאי שוכנת במיטוכונדריה, ובמקרה של תא צמחי, בכלורופלסטים.
ישנם שני סוגים של כרומטין. באוכרומטין, ה-DNA הוא המאורגן הכי פחות בצפיפות; הוא מכיל את הגנים המתועתקים בתדירות הגבוהה ביותר. סוג אחר של כרומטין, הטרוכרומטין, הוא קומפקטי יותר ומכיל DNA שלעיתים רחוקות או אף פעם לא מתועתק. הטרוכרומטין מחולק לפקולטטיבי, הקיים רק בתאים מסוג מסוים ובשלב מסוים של מחזור התא, ומכונן, המיוצג על ידי מבני כרומוזומים כגון טלומרים וצנטרומרים. במהלך interphase, הכרומטין של כל כרומוזום תופס אזור מוגדר בקפדנות של הגרעין - טריטוריה של כרומוזומים. גנים פעילים, הנוטים להיות ממוקמים באוכרומטין, ממוקמים בדרך כלל בגבול טריטוריית הכרומוזומים.
גופים גרעיניים
הגרעין של תאי יונקים מכיל מספר תת-תאים נפרדים הנקראים גופים גרעיניים. הם מבצעים את המידור של הגרעין, ויוצרים בתוכו חללים נפרדים בעלי תכונות מסוימות. גופים גרעיניים רבים מבצעים פונקציות ספציפיות, כגון סינתזה ועיבוד של RNA פרה-ריבוזומלי בגרעין, הצטברות והרכבה של רכיבי spliceosome בכתמים (ראה להלן), או הצטברות של מולקולות RNA ב משקפיים. המנגנונים המבטיחים את ביצוע הפונקציות הללו על ידי גופים גרעיניים מגוונים מאוד. במקרים מסוימים, הגוף הגרעיני יכול לשמש כאתר לתהליכים מסוימים, כגון שעתוק. במקרים אחרים, גופים גרעיניים מווסתים כנראה בעקיפין את הריכוזים המקומיים של מרכיביהם בנוקלאופלזמה. כמו אברונים ציטופלזמיים, גופים גרעיניים מכילים קבוצה מסוימת של חלבונים הקובעים את המבנה שלהם ברמה המולקולרית. עם זאת, בניגוד לאברונים ציטופלזמיים, גופים גרעיניים אינם מוקפים בממברנות שומניות, ושלמותם המבנית מובטחת לחלוטין על ידי אינטראקציות חלבון וחלבון RNA-חלבון. הטבלה שלהלן מפרטת את המאפיינים העיקריים של גופים גרעיניים.
| גוף גרעיני | פונקציות | רכיבים אופייניים | גודל אופייני (במיקרומטר) | כמות לכל ליבה |
|---|---|---|---|---|
| גרעין | ביוגנזה של ריבוזום | מְכוֹנוֹת RNA פולימראז I, גורמי עיבוד rRNA והרכבת תת-יחידות ריבוזומליות | 3-8 | 1-4 |
| כתמים | הצטברות והרכבה של גורמי שחבור | גורמי שחבור טרום-mRNA | 2-3 | 20-50 |
| לחץ על גופים גרעיניים | ויסות של תמלול וחבור תחת לחץ | HSF1, HAP | 1-2 | 3-6 |
| גוף של לוקוסים היסטונים | עיבוד היסטון טרום-mRNA | NPAT, פלאש, U7 snRNP | 0,2-1,2 | 2-4 |
| גוף קג'ל | ביוגנזה, התבגרות ומחזור של RNAs קטנים | סליל, SMN | 0,2-1,5 | 1-10 |
| גוף PML | ויסות יציבות הגנום, תיקון DNA, בקרת שעתוק, הגנה מפני וירוסים | PML | 0,1-1 | 10-30 |
| משקפיים | ויסות mRNA, עריכת RNA | RNA לא מקודדים NEAT1/MENε/β, חלבוני PSP1, p54 nrb /NONO | 0,2-1 | 2-20 |
| תא פרינוקלאולרי | ויסות פוסט-תעתיק של קבוצה של RNA שסונתז RNA פולימראז III | PTB | 0,2-1 | 1-2 |
גרעין
הגרעין הוא מבנה צפוף נפרד בגרעין. הוא אינו מוקף בממברנה ונוצר באזור בו נמצא rDNA - חזרות טנדם של גנים ריבוזומיים RNA (rRNA) הנקראים מארגנים גרעיניים. תפקידו העיקרי של הגרעין הוא סינתזה של rRNA ויצירת ריבוזומים. השלמות המבנית של הגרעין תלויה בפעילותו, והשבתת גנים של rRNA מובילה לתערובת של מבנים גרעיניים.
בשלב הראשון של יצירת הריבוזום, האנזים RNA polymerase I מתמלל rDNA ויוצר קדם-rRNA, אשר נחתך עוד יותר ל-5.8S, 18S ו-28S rRNA. שעתוק ועיבוד פוסט-תעתיק של rRNA מתרחשים בגרעין בהשתתפות RNAs גרעיניים קטנים (snoRNAs), שחלקם מקורם באינטרונים של mRNA שחוברים של גנים המקודדים לחלבונים הקשורים לתפקוד הריבוזום. תת-היחידות הריבוזומליות המורכבות הן המבנים הגדולים ביותר העוברים דרך הנקבובית הגרעינית.
כאשר צופים במיקרוסקופ אלקטרונים, ניתן להבחין בשלושה מרכיבים בגרעין: מרכזים פיברילרים (FC), הרכיב הפיברילרי הצפוף (CFC) המקיף אותם, והרכיב הגרנורי (GC), אשר, בתורו, מקיף את ה-CFC. שעתוק rRNA מתרחש ב-FC ובגבול של FC ו- PFC; לכן, כאשר היווצרות של ריבוזומים מופעלת, ה-FC הופך להיות ברור. חיתוך ושינוי של rRNA מתרחשים ב-PFC, ושלבים הבאים ביצירת תת-יחידות ריבוזומליות, כולל העמסת חלבונים ריבוזומליים, מתרחשים ב-HA.
גוף קג'ל
גוף הקאג'ל (TC) הוא הגוף הגרעיני שנמצא בכל האיקריוטים. זה מזוהה על ידי נוכחות של חלבון הסליל החתימה ו-RNAs ספציפיים (scaRNAs). TK מכיל גם את החלבון SMN. הישרדות של נוירונים מוטוריים). ל-MAs יש ריכוז גבוה של שחבור ריבונוקלאופרוטאין גרעיני קטן (snRNPs) וגורמי עיבוד RNA אחרים, ולכן מאמינים ש-MAs משמשים כאתרים להרכבה ו/או שינוי פוסט-תעתיק של גורמי שחבור. TK קיים בגרעין במהלך אינטרפאזה אך נעלם במהלך מיטוזה. בביוגנזה של TC, מאפיינים של מבנה מארגן עצמי מתחקות אחר.
כאשר הלוקליזציה התוך תאית של SMN נחקרה לראשונה על ידי אימונופלואורסצנטי, החלבון נמצא בכל הציטופלזמה, כמו גם בגוף גרעיני דומה בגודלו ל-MC ולעיתים קרובות סמוך ל-MC. מסיבה זו, הגוף הזה נקרא "תאום TK" (אנג'. מזל תאומים של CB) או סתם פנינה. אולם, התברר שקו תאי ה-HeLa שבו התגלה הגוף החדש היה יוצא דופן: בשורות תאים אנושיות אחרות, כמו גם בזבוב הפירות. תסיסנית מלנוגסטר SMN colocalized עם coilin ב-TC. לכן, במקרה הכללי, SMN יכול להיחשב כמרכיב חשוב של ה-TC, ולא כסמן של גוף גרעיני אינדיבידואלי.
גוף של לוקוסים היסטונים
גופי PML
כתמים
משקפיים
כתמים הם גופים גרעיניים בעלי צורה לא סדירה הממוקמים במרחב הבין-כרומטי של הגרעין. הם תוארו לראשונה בתאי HeLa, שיש להם 10-30 כתמים לכל גרעין, אך כתמים נמצאים כעת בכל התאים האנושיים הראשוניים, בתאים של קווים שעברו טרנספורמציה ובחתכי רקמה. הם קיבלו את שמם בגלל מיקומם בליבה - ליד הכתמים.
כתמים הם מבנים דינמיים המשתנים בתגובה לשינויים בפעילות המטבולית של התא. הם תלויי שעתוק, ובהיעדר שעתוק על ידי RNA פולימראז II, הכתמים נעלמים וכל החלבונים שלהם (PSP1, p54nrb, PSP2, CFI(m)68 ו-PSF) יוצרים מכסה פרינוקלאולרי בצורת מגל. תופעה זו נצפית במהלך מחזור התא: כתמים נמצאים באינטרפאזה ובכל שלבי המיטוזה למעט טלופאז. במהלך הטלופאז נוצרים גרעיני בת, ו-RNA פולימראז II אינו מתמלל דבר, ולכן חלבוני ה-paraspectle יוצרים את המכסה הפרי-נוקלאולרי. כתמים מעורבים בוויסות ביטוי הגנים על ידי צבירת אותם RNAs שבהם יש אזורים דו-גדיליים הכפופים לעריכה, כלומר ההמרה של אדנוזין לאינוזין. באמצעות מנגנון זה, כתמים מעורבים בשליטה על ביטוי גנים במהלך התמיינות, זיהום ויראלי ומתח.
תא פרינוקלאולרי
המדור הפרינוקלאולרי (OK) הוא גוף גרעיני בעל צורה לא סדירה, המתאפיין בכך שהוא ממוקם בפריפריה של הגרעין. למרות היותם קשורים פיזית, שני התאים נבדלים מבחינה מבנית. TCs נמצאים בדרך כלל בתאי גידול ממאירים. OK הוא מבנה דינמי, ומכיל הרבה חלבונים קושרי RNA ו-RNA פולימראז III. היציבות המבנית של OK מובטחת על ידי שעתוק המבוצע על ידי RNA פולימראז III ונוכחות של חלבוני מפתח. מכיוון שנוכחות TC קשורה בדרך כלל לממאירות וליכולת לגרורות, הם נחשבים כסמנים פוטנציאליים לסרטן וגידולים ממאירים אחרים. הוכח הקשר של OK עם לוקוסי DNA ספציפיים.
לחץ על גופים גרעיניים
גופים גרעיניים מתח נוצרים בגרעין במהלך הלם חום. הם נוצרים על ידי אינטראקציה ישירה של גורם שעתוק הלם חום 1 ( HSF1) וחזרות טנדם פריצנטריות ברצף של לוויין III, התואמים לאתרים של תעתיק פעיל של תמלילים לא מקודדים של לוויין III. הדעה הרווחת היא שגופים כאלה תואמים לצורות ארוזות בצפיפות רבה של קומפלקסים של ריבונוקלאופרוטאין. בתאים לחוצים, הם חושבים שהם מעורבים בשינויים מהירים, חולפים וגלובליים בביטוי גנים באמצעות מנגנונים שונים, כגון עיצוב מחדש של כרומטין וקליטת גורמי שעתוק ושחבור. בתאים בתנאים רגילים (לא מלחיצים), גופים גרעיניים לחוצים נמצאים לעתים רחוקות, אך מספרם גדל בחדות בהשפעת הלם חום. גופים גרעיניים של מתח נמצאים רק בתאי אדם ותאי פרימטים אחרים.
גופים גרעיניים יתומים
| גוף גרעיני | תיאור | גודל אופייני (במיקרומטר) | כמות לכל ליבה |
|---|---|---|---|
| קלסטוזום | מרכז קומפלקסים של פרוטאזום 20S ו-19S וחלבונים הקשורים לאוביקווטין. זה נמצא בעיקר כאשר פעילות פרוטאזום מגורה ומתנקה כאשר פעילות פרוטאזום מעוכבת. | 0,2-1,2 | 0-3 |
| גוף החטיבה גוף פינוי) | מועשר בגורמי ביקוע CstFו CPSF, כמו גם חלבון DDX1מֵכִיל קופסא מתה. הוא נמצא בעיקר ב-S-phase ואינו מושפע מעיכוב שעתוק. | 0,2-1,0 | 1-4 |
| דומיין OPT | מועשר בגורמי תמלול 1 באוקטוברו-PTF. מתמקם חלקית עם אתרי תמלול. נמצא בעיקר בשלב ה-G1 המאוחר, מפורק על ידי עיכוב שעתוק. | 1,0-1,5 | 1-3 |
| גוף פוליקומב | נמצא בתאי אדם ותאי תסיסנית, מועשר בחלבון PcG. צובר חלבוני RING1 בבני אדם, BMI1, HPC, עשוי להיות קשור להטרוכרומטין pericentromeric. | 0,3-1,0 | 12-16 |
| מזל שור Sam68 | צובר את חלבון Sam68 וחלבונים דומים SLM-1 ו-SLM-2. מפורק על ידי עיכוב שעתוק. כנראה עשיר ב-RNA. | 0,6-1,0 | 2-5 |
| גוף SUMO | מועשר בחלבוני SUMO ואנזים מצומד SUMO Ubc9. מרכז את גורמי השעתוק pCREB, CBP, c-יוני. | 1-3 | 1-3 |
פונקציות
המעטפת הגרעינית מגנה על ה-DNA של התא ומעורבת בוויסות הרבה יותר מורכב של ביטוי גנים בהשוואה לתא הפרוקריוטי. בפרוקריוטים, שעתוק ותרגום הם תהליכים משולבים, ותרגום של mRNA לחלבון מתחיל עוד לפני שהוא מסונתז במלואו. בתאים איקריוטים, הציטופלזמה שבה מתרחש התרגום והתעתוק המתרחש בגרעין מופרדות במרחב, ולכן יש צורך להבטיח את הובלת מולקולות בין הגרעין לציטופלזמה.
המעטפת הגרעינית מעניקה לגרעין שליטה על תכולתו ומפרידה בינו לבין שאר הציטופלזמה. זה חשוב לוויסות תהליכים המתרחשים משני צידי המעטפת הגרעינית. כאשר התהליך הציטופלזמי צריך להיות מוגבל איכשהו, אז בדרך כלל משתתף המפתח שלו מועבר לגרעין, שם הוא יוצר אינטראקציה עם גורמי שעתוק ובכך גורם לדיכוי היווצרות של כמה אנזימים המעורבים בתהליך הציטופלזמי. למשל, לגליקוליזה, תהליך שבו תא שואב אנרגיה ממולקולת גלוקוז, יש מנגנון ויסות כזה. התגובה הראשונה של הגליקוליזה מתבצעת על ידי האנזים הקסוקינאז, הממיר את מולקולת הגלוקוז לגלוקוז-6-פוספט. כאשר ריכוז הפרוקטוז-6-פוספט (חומר שנוצר מגלוקוז-6-פוספט במהלך הגליקוליזה) עולה, החלבון הרגולטורי שולח הקסוקינאז לגרעין, שם הוא יוצר קומפלקס מדכא שעתוק המדכא את הביטוי של גנים המקודדים לאנזימים גליקוליטים.
על מנת לשלוט אילו גנים מתועתקים, לגורמי שעתוק בתא אין גישה פיזית ל-DNA עד שהם מופעלים במסלול איתות מסוים. זה מונע אפילו ביטוי נמוך של הגנים הלא נכונים. בפרט, במקרה של גנים הנשלטים על ידי NF-κB המעורבים בתהליך הדלקתי, השעתוק מושרה על ידי מסלול איתות, למשל, המתחיל בקשירה של מולקולת האיתות TNF-α לקולטן שלה על קרום התא. ובסופו של דבר מוביל להפעלה של גורם השעתוק NF-κB. אות הלוקליזציה הגרעיני הקיים ב-NF-κB מאפשר לו לעבור אל תוך הגרעין והחוצה ממנו דרך הנקבוביות הגרעיניות; בגרעין, הוא ממריץ את התעתוק של גנים מטרה.
מידור מונע מהתא לתמלל את ה-mRNA הלא-חובר. mRNA אוקריוטיים מכילים אינטרונים שיש להסיר לפני שניתן להתחיל בתרגום של ה-mRNA. שחבור, כלומר הסרת אינטרונים, מתרחש בגרעין, מה שמונע גישה לפרה-mRNA על ידי ריבוזומים מחוץ לגרעין. אם לא היה גרעין, אז הריבוזומים היו מתחילים לתרגם mRNA לא בוגר, מה שיוביל ליצירת מוצרי חלבון שגויים.
מכיוון שהתעתוק מתרחש בגרעין, הגרעין מכיל חלבונים רבים המעורבים ישירות בתעתוק או מווסתים אותו. חלבונים אלו כוללים הליקאזות, המשחררות את הסליל הכפול של ה-DNA, ומקלים על הגישה של חלבונים אחרים אליו, פולימראזות RNA, המסנתזות RNA, טופואיזומראזות, המשפיעות על טופולוגיית ה-DNA, ומגוון גורמי שעתוק.
הובלה גרעינית
היציאה מהגרעין והכניסה לגרעין של מולקולות גדולות נשלטת על ידי נקבוביות גרעיניות. למרות שמולקולות קטנות יכולות להיכנס לגרעין ללא כל ויסות, מקרומולקולות כגון חלבונים ו-RNA חייבות להיקשר לקריופרינים לצורך הובלה אל הגרעין (אימפורטינים) ומחוץ לגרעין (אקספורטינים). חלבונים שחייבים להיות מועברים מהציטופלזמה לגרעין מכילים רצף חומצות אמינו ספציפי המכונה אות לוקליזציה גרעיני, שאליו נקשרים ה-importins. באופן דומה, חלבונים שחייבים לצאת מהגרעין מכילים אות ייצוא גרעיני, מוכר על ידי יצואנים. היכולת של יבואנים וייצואינים לשאת את המטען שלהם מווסתת על ידי GTPases, אנזימים המעבירים הידרוליזה של GTP לשחרור אנרגיה. המפתח GTPase של הובלה גרעינית הוא רץ, שיכול להיקשר ל-GTP או ל-GDP, בהתאם למיקומו (בגרעין או בציטופלזמה). בבסיסו, האינטראקציה של Ran-GTP עם importin גורמת לשינוי קונפורמטיבי אצל האחרון כך שהוא נפרד מהמטען הנישא. הקומפלקס שנוצר של Ran-GTP ו-importin מועבר לציטופלזמה, שם חלבון RanBP מפריד בין Ran-GTP ל-importin. ההפרדה מ-importin מאפשרת את החלבון פערצור קשר עם Ran-GTP וזרז את ההידרוליזה של GTP ל-GDP. יתר על כן, תסביך Ran-GDP מזוהה על ידי החלבון NUTF2, מה שמחזיר אותו לנוקלאופלזמה. חלבון בגרעין GEFמחליף GDP ב-GTP, יוצר Ran-GTP וסגירת המחזור.
הרכבה ופירוק
במהלך חיי התא ניתן לפרק את הגרעין (במהלך חלוקת התא או אפופטוזיס). במהלך תהליכים אלו מושמדים המרכיבים המבניים של הגרעין - המעטפת הגרעינית והלמינה הגרעינית. ברוב התאים, פירוק של הגרעין מתרחש במהלך פרוזה של מיטוזה. עם זאת, פירוק של הגרעין אינו מוגבל אך ורק למיטוזה ואינו מתרחש בכל התאים. כמה אוקריוטים חד-תאיים (כגון שמרים) עוברים מה שמכונה מיטוזה סגורה, שבה המעטפת הגרעינית נשארת שלמה. במיטוזה סגורה, הכרומוזומים עוברים לצדדים שונים של הגרעין, אשר לאחר מכן מתחלק לשניים. לעומת זאת, תאים של אוקריוטים גבוהים יותר עוברים בדרך כלל מיטוזה פתוחה, שבמהלכה מתפרקת המעטפת הגרעינית. כרומוזומים נודדים לקטבים שונים של הציר, ושני גרעינים נוצרים מחדש סביבם. הלמינה הגרעינית עוברת גם פירוק עקב זרחון של למינים על ידי קינאזות כגון חלבון קינאז תלוי ציקלין 1. הרכבה של הלמינה הגרעינית בגרעיני הבת מתחילה לאחר דה-פוספורילציה של הלמינים.
אפופטוזיס הוא תהליך מבוקר של הרס של רכיבים תאיים המוביל למוות תאי. שינויים הקשורים לאפופטוזיס מתרחשים ישירות בגרעין ובתכולתו. אלה כוללים עיבוי כרומטין, כמו גם התפוררות של המעטפת הגרעינית והלמינה הגרעינית. פירוק רשת הלמינים מתווכת על ידי פרוטאזות אפופטוטיות, המכונות קספסות, אשר מפרקות למינים ובכך משפיעות על השלמות המבנית של הגרעין. הרס למין משמש לעתים כאינדיקטור לפעילות קספאז במחקרי אפופטוזיס. תאים המבטאים למינים מוטנטיים עמידים לקספאז אינם מאבדים שלמות גרעינית במהלך אפופטוזיס, ולכן הלמינים ממלאים תפקיד מפתח בהתחלת שינויים שהגרעין עובר במהלך האפופטוזיס. בנוסף, עיכוב של הרכבה של למינים לרשת גורם לאפופטוזיס.
תכונות של גרעינים באוקריוטים שונים
הגדלים, הצורות והמורפולוגיה של גרעינים אוקריוטיים משתנים מאוד. אם פירופלסמידולישמניה, קוטר הגרעין הוא 1-3 מיקרון, ואז בחלק מהרדיולארים, הקוטר של הגרעינים מגיעים ל-400 מיקרון ואפילו 1 מ"מ. ככלל, צורת הגרעין ברוב האאוקריוטים קרובה לכדורית, אך לפעמים היא יכולה ללבוש צורות מוזרות למדי (זה, במיוחד, חל על מקרו-גרעיני ריסים). למרות שבכל האיקריוטים קליפת הגרעין מורכבת משני ממברנות, מספר הנקבוביות בה משתנה מאוד במינים שונים, ולעתים יכולות לצמוד אליו שכבות נוספות (הן מבחוץ והן מבפנים); למשל, באמבות רבות חיות חופשיות, צמודה לצד הפנימי של הקונכייה שכבה סיבית בעלת מבנה תאי, העולה באופן משמעותי על הקונכייה הגרעינית בעובי, וברדיולארים שכבות פיברילריות נוספות ממוקמות בצד החיצוני של הקונכייה. .
ארגון הגרעין בפרוטיסטים מסוג Dinoflagellate (Dinoflagellata) נבדל במקוריות משמעותית. לרוב הנציגים שלהם יש גרעין שבו הכרומוזומים מְעוּבֶּהלאורך כל מחזור התא (כולל ב-interphase) והם כמעט נטולי היסטונים. סוג זה של גרעין נקרא דינוקריון. יחד עם זאת, כמות ה-DNA בדינוקריון גדולה בעשרות ומאות מונים מכמות ה-DNA לתא בנציגי קבוצות אחרות של איקריוטים. עם זאת, כמה דינופלגלטים ( נוקטילוקה, אודיניום ) בעלי גרעינים אוקריוטיים תקינים; בנציגים אחרים מהסוג בתאים וגטטיביים, הגרעינים הם רגילים, והדינוקריון קיים בשלבים אחרים של מחזור התא (למשל, בגמטות).
לתאים פרוטיסטים יש לפחות גרעין אחד. במקביל, תאים שאינם גרעיניים נמצאים גם באורגניזמים של Metazoa, אשר, ללא גרעין, איבדו את היכולת להתחלק עם היווצרותם של שני תאי בת. הדוגמה המוכרת ביותר לתאים שאינם בעלי גרעין הם אריתרוציטים של יונקים, שגם חסרים להם אברונים אחרים, כמו מיטוכונדריה. תאי דם אדומים מבשילים במח העצם בתהליך של אריתרופואזיס, שבמהלכו הם מאבדים גרעינים, אברונים אחרים וריבוזומים. הגרעין נדחק אל מחוץ לתא במהלך תהליך ההתמיינות של האריתרובלסט אל הרטיקולוציט, המשמש כמבשר המיידי של האריתרוציט. בהשפעת חלק מהמוטגנים, אריתרוציטים לא בשלים המכילים מיקרוגרעינים יכולים להשתחרר לדם.
לרוב הפרוטיסטים יש רק גרעין אחד; בפרוטיסטים, המאופיינים במחזור חיים מורכב (לדוגמה, לנציגים מסוג apicomplexa (Apicomplexa) יש שלבים חד-ליבים ורב-ליבים.
תאים פרוטיסטים מרובי גרעינים
הרכב הכלורופלסטים בקריפטופיטים וכלורכניופיטים מכיל נוקלאומורף- גרעין מצומצם של אנדוסימביונט פוטוטרופי, ששולבו על ידי אבותיהן של אצות אלו במהלך אנדוסימביוזה משנית (ב-Cryptophyta שולבו אצות אדומות, וב-Chlorarachnea, אצות ירוקות)
גרעין התא
הליבה(גרעין, s. karyon) נמצא בכל תאי האדם, למעט אריתרוציטים וטסיות דם. תפקידיו של הגרעין הם אחסון והעברת מידע תורשתי לתאים חדשים (בת). פונקציות אלו קשורות לנוכחות של DNA בגרעין. סינתזת חלבון מתרחשת גם בגרעין - RNA חומצה ריבונוקלאית וחומרים ריבוזומים.
ברוב התאים, הגרעין הוא כדורי או ביצי, אך קיימות צורות נוספות של הגרעין (טבעתי, מוט, פתיל, חרוז, שעועית, מפולח, אגס, פולימורפי). גודלו של הגרעין משתנה מאוד - בין 3 ל-25 מיקרון. לביצית יש את הגרעין הגדול ביותר. רוב התאים האנושיים הם חד-גרעיניים, אך ישנם דו-גרעיניים (כמה נוירונים, הפטוציטים, קרדיומיוציטים). מבנים מסוימים הם מרובי גרעינים (סיבי שריר). לגרעין יש מעטפת גרעינית, כרומטין, גרעין ונוקלאופלזמה (איור 4).
הממברנה הגרעינית, או caryotheca (caryotheca), המפרידה בין תוכן הגרעין לציטופלזמה, מורכבת מממברנה גרעינית פנימית וחיצונית בעובי 8 ננומטר כל אחד. הממברנות מופרדות על ידי חלל פרי-גרעיני (בור קריוטקה) ברוחב 20-50 ננומטר, המכיל חומר עדין גרגר בעל צפיפות אלקטרונים בינונית. הממברנה הגרעינית החיצונית עוברת לתוך הרשת האנדופלזמית הגרנורית. פוטו החלל הפרי-גרעיני של Mu הוא חלל בודד עם הרשת האנדופלזמית. הממברנה הגרעינית הפנימית מחוברת פנימית לרשת נרחבת של סיבים של חלבון, המורכבת מתת-יחידות בודדות.
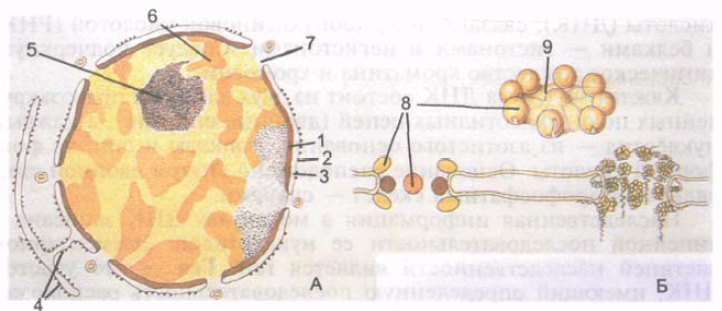
אורז. 4. גרעין התא (A) וקומפלקס נקבוביות גרעיניות (B). 1 - קרום גרעיני פנימי; 2 - קרום גרעיני חיצוני; 3 - חלל perinuclear; 4 - ממברנות של הרשת האנדופלזמית; 5 - גרעין; 6 - כרומטין מעובה; 7 - זמן גרעיני; 8 - גרגירים של הנקבובית הגרעינית; 9 - דיאפרגמה של נקבוביות גרעיניות.
המעטפת הגרעינית מכילה נקבוביות גרעיניות מעוגלות רבות בקוטר של 50-70 ננומטר כל אחת. נקבוביות גרעיניות בסך הכל תופסות עד 25% משטח הגרעין. מספר הנקבוביות בגרעין אחד מגיע ל-3000-4000. לאורך קצוות הנקבוביות, הממברנות החיצוניות והפנימיות מחוברות זו לזו ויוצרות את מה שנקרא טבעת הנקבוביות. כל נקבובית סגורה על ידי דיאפרגמה, הנקראת גם קומפלקס נקבוביות. לסרעפות נקבוביות יש מבנה מורכב; הן נוצרות על ידי גרגירי חלבון מחוברים זה לזה. דרך הנקבוביות הגרעיניות מתבצעת הובלה סלקטיבית של חלקיקים גדולים, כמו גם חילופי חומרים בין הגרעין לציטוזול של התא.
מתחת למעטפת הגרעין נמצאים נוקלאופלזמה (קריופלזמה)(נוקלאופלזמה, s. קריופלזמה), בעל מבנה הומוגני, והגרעין. בנוקלאופלזמה של הגרעין הלא מתחלק, במטריצת החלבון הגרעיני שלו, יש גרגירים אוסמיופיליים (גושים) של מה שנקרא הטרוכרומטין. האזורים של כרומטין משוחרר יותר הממוקמים בין הגרגירים נקראים אוכרומטין. כרומטין רופף נקרא גם כרומטין מעובה; תהליכים סינתטיים מתקיימים בו בצורה האינטנסיבית ביותר. במהלך חלוקת התא, הכרומטין מתעבה, מתעבה ויוצר כרומוזומים.
כרומטין(כרומטינום) של הגרעין הלא מתחלק והכרומוזומים של זה המתחלק נוצרים על ידי מולקולות של חומצה דאוקסיריבונוקלאית (DNA) הקשורה לחומצה ריבונוקלאית (RNA) ולחלבונים - היסטונים ולא היסטונים. יש להדגיש את הזהות הכימית של הכרומטין והכרומוזומים.
כל מולקולת DNA מורכבת משתי שרשראות פולינוקלאוטידים ארוכות ביד ימין (סלילים כפולים), וכל נוקלאוטיד מורכב מבסיס חנקני, גלוקוז ושארית חומצה זרחתית. הבסיס ממוקם בתוך הסליל הכפול, ושלד הסוכר-פוספט נמצא בחוץ.
מידע תורשתי במולקולות DNA כתוב ברצף הליניארי של הנוקלאוטידים שלו. החלקיק היסודי של התורשה הוא הגן. גן הוא קטע של DNA שיש לו רצף ספציפי של נוקלאוטידים האחראי לסינתזה של חלבון ספציפי אחד.
מולקולת ה-DNA בגרעין ארוזה בצורה קומפקטית. לפיכך, מולקולת DNA אחת המכילה מיליון נוקלאוטידים, עם הסידור הליניארי שלהם, תתפוס קטע באורך 0.34 מ"מ בלבד. אורכו של כרומוזום אנושי אחד בצורה מתוחה הוא כ-5 ס"מ, אולם במצב דחוס, לכרומוזום נפח של כ-10 -15 ס"מ 3.
מולקולות DNA הקשורות לחלבוני היסטון יוצרות נוקלאוזומים, שהם היחידות המבניות של הכרומטין. נוקלאוזוםיש צורה של חרוז בקוטר של 10 ננומטר. כל נוקלאוזום מורכב מהיסטונים, שסביבם מעוות גדיל DNA של 146 bp. בין הנוקלאוזומים יש קטעים ליניאריים של DNA, המורכבים מ-60 זוגות של נוקלאוטידים.
הכרומטין מיוצג על ידי סיבים, היוצרים לולאות באורך של כ-0.4 מיקרומטר, המכילות בין 20,000 ל-30,000 זוגות בסיסים.
כתוצאה מדחיסה (עיבוי) ופיתול (התמחות-על) של דאוקסיריבונוקלאופרוטאין (DNP) בגרעין המתחלק, הופכים כרומוזומים לגלויים. מבנים אלה הם כרומוזומים(chromasomae, מיוונית chruma - צבע, סומה - גוף) - הם תצורות מוארכות בצורת מוט עם שתי זרועות מופרדות על ידי מה שנקרא כיווץ - centromere. בהתאם למיקום הצנטרומר ולמיקום היחסי ואורך הזרועות (הרגליים), מבחינים בשלושה סוגים של כרומוזומים: מטאצנטרי, בעל אותן זרועות בערך; submetacentric, שבו אורך הכתפיים שונה; אקרוצנטרי, שבו כתף אחת ארוכה, והשנייה קצרה מאוד, בקושי מורגשת. הכרומוזום מכיל אזורים eu- והטרוכרומטיים. האחרון בגרעין הלא מתחלק ובתחזית המוקדמת של מיטוזה נשארים קומפקטיים. החלפה של אזורים איו והטרוכרומטיים משמשת לזיהוי כרומוזומים.
פני השטח של הכרומוזומים מכוסים במולקולות שונות, בעיקר ריבונוקלאופרוטאין (RNP). לתאים סומטיים יש 2 עותקים של כל כרומוזום, הם נקראים הומולוגיים. הם זהים באורך, צורה, מבנה, נושאים את אותם גנים הממוקמים באותו אופן. המאפיינים המבניים, מספרם וגודלם של הכרומוזומים נקראים קריוטיפ. הקריוטיפ האנושי הרגיל כולל 22 זוגות של אוטוזומים וזוג אחד של כרומוזומי מין (XX או XY). לתאים אנושיים סומטיים (דיפלואידים) יש מספר כפול של כרומוזומים - 46. תאי מין מכילים סט הפלואידי (יחיד) - 23 כרומוזומים. לכן, ה-DNA בתאי נבט קטן פי 2 מאשר בתאים סומטיים דיפלואידים.
גרעין(נוקלאולוס), אחד או יותר, נמצא בכל התאים שאינם מתחלקים. יש לו צורה של גוף מעוגל מוכתם בעוצמה, שגודלו פרופורציונלי לעוצמת סינתזת החלבון. הגרעין מורכב מנוקלאולונמה צפופה באלקטרון (מיוונית peta - חוט), שבה מובחן חלק חוטי (פיברילרי), המורכב מגדילים רבים של RNA שזורים זה בזה בעובי של כ-5 ננומטר, וחלק גרגירי. החלק הגרגירי (הגרגירי) נוצר על ידי גרגרים בקוטר של כ-15 ננומטר, שהם חלקיקי RNP - מבשרי תת-יחידות ריבוזומליות. כרומטין Perinucleolar מוטבע בשקעים של הנוקלאולונמה. ריבוזומים נוצרים בגרעין.
חלוקת תא. מחזור התא
אורגניזם גדל על ידי הגדלת מספר התאים על ידי חלוקה. השיטות העיקריות לחלוקת תאים בגוף האדם הן מיטוזה ומיוזה. התהליכים המתרחשים בשיטות אלו של חלוקת תאים מתנהלים באותו אופן, אך מביאים לתוצאות שונות.
חלוקת תאים מיטוטיים(מיטוזה) מוביל לעלייה במספר התאים, הצמיחה של הגוף. בדרך זו מובטחת חידוש התאים כשהם נשחקים ומתים. כיום ידוע שתאי האפידרמיס חיים 10-30 ימים, אריתרוציטים - עד 4-5 חודשים. תאי עצב ושריר (סיבים) חיים לאורך כל חייו של האדם.
בכל התאים, במהלך הרבייה (חלוקה), נצפים שינויים המשתלבים במסגרת מחזור התא. מחזור התאנקראים התהליכים המתרחשים בתא מחלוקה לחלוקה או מחלוקה למוות (מוות) של התא. במחזור התא מבחינים בהכנת התא לחלוקה (אינטרפאזה) ומיטוזה (תהליך חלוקת התא).
ב-interphase, שנמשך כ-20-30 שעות, קצב התהליכים הביוסינתטיים עולה, ומספר האברונים עולה. בשלב זה, מסת התא וכל מרכיביו המבניים, כולל צנטריולים, מוכפלת.
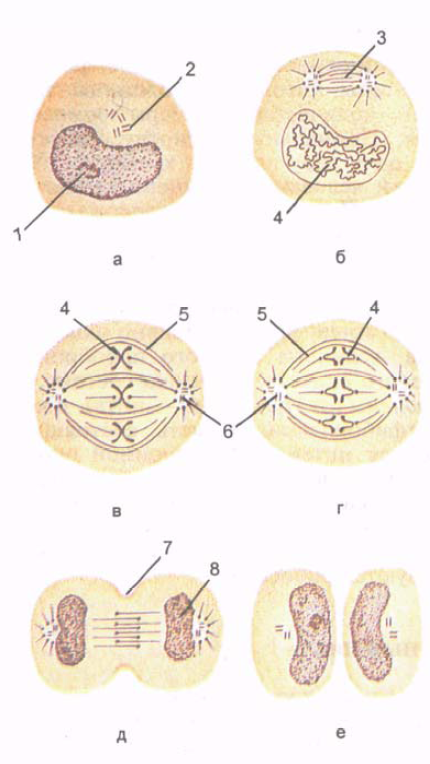
אורז. 5. חלוקת תאים. שלבים של מיטוזה. מוצגים היווצרות הכרומוזומים, היווצרות ציר החלוקה והתפלגות האחידה של כרומוזומים וצנטריולים בשני תאי בת.
A - interphase; ב' - פרופסה; B - מטאפאזה; G - אנפאזה; D - טלופאז; E - טלופאז מאוחר. 1 - ליבה; 2 - מרכז תא (צנטריול); 3 - ציר של חלוקת תאים; 4 - כרומוזומים; 5 - microtubules רציף; 6 - מרכז תאים; 7 - תלם של חלוקת תאים; 8 - היווצרות הליבה.
מתרחשת שכפול (חזרה, הכפלה) של מולקולות חומצת גרעין. זהו תהליך העברת מידע גנטי המאוחסן ב-DNA של ההורים על ידי שחזור מדויק שלו בתאי בת. גדיל ה-DNA האב משמש כתבנית לסינתזה של ה-DNA הבת. כתוצאה משכפול, כל אחת משתי מולקולות ה-DNA הבת מורכבת מגדיל אחד ישן ואחד חדש. במהלך תקופת ההכנה למיטוזה, החלבונים הדרושים לחלוקת התא מסונתזים בתא. בסוף האינטרפאזה, הכרומטין בגרעין מתעבה.
מיטוזה(מיטוזה; מיוונית מיטוס - חוט) היא התקופה שבה תא האם מתחלק לשני תאי בת (איור 5). חלוקת תאים מיטוטיים מספקת חלוקה אחידה של מבני התא, החומר הגרעיני שלו - כרומטין - בין שני תאי בת. משך המיטוזה הוא בין 30 דקות ל-3 שעות.המיטוזה מחולקת לפרופזה, מטפאזה, אנפאזה, טלופזה.
בפרופאזה, הגרעין מתפרק בהדרגה, הצנטרולים מתפצלים לכיוון הקטבים של התא. מיקרוטובולים של צנטריולים מכוונים לכיוון קו המשווה, ובאזור קו המשווה הם חופפים זה לזה.
במטאפאזה, המעטפת הגרעינית נהרסת, חוטי כרומוזומים נשלחים לקטבים, שומרים על קשר עם קו המשווה
כל שטח התא. המבנים של הרשת האנדופלזמית והקומפלקס של גולגי מתפרקים לשלפוחיות קטנות (שלפוחיות), אשר יחד עם המיטוכונדריה מפוזרות לשני חצאי התא המתחלק. בסוף המטאפאזה, כל כרומוזום מתחיל להתפצל עם שסע אורכי לשני כרומוזומי בת חדשים.
באנפאזה, הכרומוזומים נפרדים זה מזה ומתפצלים לכיוון הקטבים של התא בקצב של עד 0.5 מיקרומטר/דקה. בסוף האנפאזה, קרום הפלזמה חודר לאורך קו המשווה של התא בניצב לציר האורך שלו, ויוצר תלם ביקוע.
בטלופאזה, הכרומוזומים שהתפצלו לקטבים של התא מתפרקים, עוברים לכרומטין ומתחיל שעתוק (ייצור) של RNA. המעטפת הגרעינית, הגרעין נוצרים, ומבני הממברנה של תאי הבת העתידיים נוצרים במהירות. על פני התא, לאורך קו המשווה שלו, ההיצרות מעמיקה, התא מחולק לשני תאי בת.
עקב החלוקה המיטוטית, תאי הבת מקבלים קבוצה של כרומוזומים זהים להורה. מיטוזיס מספקת יציבות גנטית, עלייה במספר התאים, וכתוצאה מכך, את הצמיחה של האורגניזם, כמו גם תהליכי התחדשות.
מיוזיס(מהמיוזה מיוזה - הפחתה) נצפית בתאי נבט. כתוצאה מחלוקת התאים הללו נוצרים תאים חדשים עם קבוצה אחת (הפלואידית) של כרומוזומים, החשובה להעברת מידע גנטי. כאשר תא מין אחד מתמזג עם תא מהמין השני (במהלך ההפריה), מערך הכרומוזומים מכפיל את עצמו, הופך שלם, כפול (דיפלואידי). בזיגוטה הדיפלואידית (דו-גרעינית) שנוצרה לאחר איחוי תאי נבט, קיימות שתי קבוצות של כרומוזומים זהים (הומולוגיים). כל זוג כרומוזומים הומולוגיים של אורגניזם דיפלואידי (זיגוטה) מגיע מגרעין הביצית ומגרעין הזרע.
כתוצאה ממיוזה של תאי נבט באורגניזם בוגר, כל תא בת מכיל רק אחד מכל זוגות הכרומוזומים ההומולוגיים של התאים המקוריים. זה מתאפשר מכיוון שבמהלך המיוזה מתרחשות רק שכפול DNA ושתי חלוקות גרעיניות עוקבות. כתוצאה מכך, שני תאים הפלואידים נוצרים מתא דיפלואידי אחד. כל אחד מתאי הבת הללו מכיל חצי כרומוזומים (23) מאשר בגרעין תא האם (46). כתוצאה ממיוזה, לתאי מין הפלואידיים יש לא רק מספר חצי של כרומוזומים, אלא סידור שונה של גנים בכרומוזומים. לכן, האורגניזם החדש נושא לא רק את סך המאפיינים של הוריו, אלא גם את התכונות שלו (הפרטיות).
שאלות לחזרה ושליטה עצמית
1. מאילו יסודות מורכב גרעין התא? אילו פונקציות הוא מבצע?
2. ספר לנו על המבנה של מולקולות DNA.
3. תנו מאפיין מורפולוגי של כרומוזומים, תנו את הסיווג שלהם.
4. מהו מחזור התא, באילו תקופות (שלבים) מבחינים במחזור זה?
5. מהי מיוזה, במה היא שונה ממיטוזה?