Paleontološki dokazi kažejo, da so bile nekatere rastline iz skupine Eucaesalpinioideae najstarejše stročnice, ki so imele nodule.
Pri sodobnih vrstah stročnic so na koreninah številnih predstavnikov družine Papilijonaceae našli vozliče.
Filogenetsko primitivnejši predstavniki takih družin, kot so Caesalpiniaceae Mimosaceae, v večini primerov ne tvorijo nodulov.
Od 13.000 vrst (550 rodov) stročnic je bila prisotnost gomoljev doslej ugotovljena le pri približno 1300 vrstah (243 rodov). Sem sodijo predvsem rastlinske vrste, ki se uporabljajo v kmetijstvu (več kot 200).
Po oblikovanju nodulov stročnice pridobijo sposobnost absorbiranja atmosferskega dušika. Lahko pa se hranijo z vezanimi oblikami dušika - amonijeve soli in dušikove kisline. Samo ena rastlina, Hedysarum coronarium, asimilira samo molekularni dušik. Zato brez nodulov v naravi ta rastlina ne obstaja.
Nodulne bakterije oskrbujejo stročnico z dušikom, ki se veže iz zraka. Rastline pa oskrbujejo bakterije s produkti presnove ogljikovih hidratov in mineralnimi solmi, ki jih potrebujejo za rast in razvoj.
Leta 1866 je slavni botanik in znanstvenik M.S. Voronin je videl najmanjša "teleta" v vozličih na koreninah stročnic. Voronin je za tisti čas postavil drzne predpostavke: nastanek vozličev je povezal z aktivnostjo bakterij, povečano delitev celic koreninskega tkiva pa z reakcijo rastline na bakterije, ki so prodrle v korenino.
20 let kasneje je nizozemski znanstvenik Beijerinck izločil bakterije iz gomoljev graha, grašice, lubada, fižola, seradele in lizike ter preučeval njihove lastnosti in preizkusil sposobnost okužbe rastlin in povzročitve nastanka gomoljev. Te mikroorganizme je poimenoval Bacillus radicicola. Ker bakterije, ki tvorijo spore, pripadajo rodu Bacillus, nodulne bakterije pa te sposobnosti nimajo, jih je A. Prazhmovsky preimenoval v Bacterium radicicola. B. Frank je predlagal bolj uspešno generično ime za nodulne bakterije - Rhizobium (iz grškega rhizo - koren, bio - življenje; življenje na koreninah). To ime se je uveljavilo in se še vedno uporablja v literaturi.
Za označevanje vrste nodulnih bakterij je običajno generičnemu imenu Rhizobium dodati izraz, ki ustreza latinskemu imenu rastlinske vrste, iz katerih gomoljev so izolirani in na katerih lahko tvorijo nodule. Na primer Rhizobium trifolii - gomoljne bakterije detelje, Rhizobium lupini - gomoljne bakterije volčjega boba itd. V primerih, ko so nodulne bakterije sposobne tvoriti vozliče na koreninah različnih vrst stročnic, tj. povzročijo tako imenovano navzkrižno okužbo, je ime vrste tako rekoč skupno - odraža to "navzkrižno okužbo" sposobnost. Na primer, Rhizobium leguminosarum - gomoljne bakterije graha (Pisum), leče (Lens), vrst (Lathyrus).
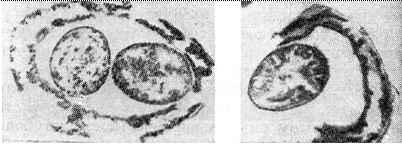
Morfologija in fiziologija nodulnih bakterij - za nodulne bakterije je značilna neverjetna raznolikost oblik - polimorfizem. Mnogi raziskovalci so na to opozorili pri proučevanju nodulnih bakterij v čisti kulturi v laboratorijskih pogojih in zemlji. Nodulne bakterije so lahko paličaste in ovalne. Med temi bakterijami so tudi filtrabilne oblike, L-oblike, kokoidno nepremični in mobilni organizmi.
Mlade nodulne bakterije v čisti kulturi na hranilnih medijih imajo običajno paličasto obliko, velikost palic je približno 0,5-0,9 X 1,2-3,0 mikronov, mobilne, razmnožujejo se z delitvijo. V paličastih celicah nodulnih bakterij detelje opazimo delitev z ligacijo. S starostjo se lahko paličaste celice premaknejo v brstenje. Po Gramu se celice obarvajo negativno, njihova ultrafina struktura je značilna za gramnegativne bakterije.
Nodulne bakterije s staranjem izgubijo svojo mobilnost in preidejo v stanje tako imenovanih pasov. To ime so dobili zaradi izmenjave gostih in ohlapnih delov protoplazme v celicah. Progastost celic je dobro zaznavna, če jo gledamo pod svetlobnim mikroskopom po obdelavi celic z anilinskimi barvili. Gosti odseki protoplazme (pas) so obarvani slabše kot prostori med njimi. V fluorescenčnem mikroskopu so pasovi svetlo zeleni, prostori med njimi se ne svetijo in so videti temni.Trakovi se lahko nahajajo na sredini celice ali na koncih. Opasnost celic je vidna tudi na elektronskih difraktogramih, če preparata pred ogledom ne obdelamo s kontrastnimi sredstvi. Verjetno je s starostjo bakterijska celica napolnjena z maščobnimi vključki, ki ne zaznavajo barve in posledično povzročijo progasto celico. Faza "pasastih palic" je pred fazo nastajanja bakteroidov - celic nepravilne oblike: zadebeljene, razvejane, sferične, hruškaste in bučke v obliki paličastih bakterijskih celic, ki jih najdemo v tkivih nodulov.
Bakteroidi vsebujejo več volutinskih zrnc in zanje je značilna večja vsebnost glikogena in maščob kot paličaste celice. Bakteroidi, ki rastejo v umetnih hranilnih medijih in nastajajo v tkivih nodulov, so fiziološko istega tipa. Menijo, da so bakteroidi oblike bakterij z nepopolnim procesom delitve. Z nepopolno delitvijo celic nodulnih bakterij nastanejo dihotomno razvejane oblike bakteroidov. Število bakteroidov narašča s staranjem kulture; njihov pojav olajšajo izčrpavanje hranilnega medija, kopičenje presnovnih produktov in vnos alkaloidov v medij.
riž. 3
V starih (dva meseca) kulturah nodulnih bakterij je mogoče z elektronskim mikroskopom prepoznati jasno definirane sferične tvorbe v številnih celicah – artrospore. Njihovo število v celicah se giblje od 1 do 5.
riž. 4
Nodulne bakterije različnih vrst stročnic rastejo na hranilnih gojiščih z različno hitrostjo. Hitro rastoče bakterije vključujejo rizobije graha, detelje, lucerne, krmnega fižola, grašice, leče, lubadarja, sladke detelje, triplata, fižola, čičerike in ptičje noge; do počasi rastočih nodulnih bakterij volčjega boba, soje, arašidov, seradele, mung fižola, kravjega fižola, esparzete, grebena. Popolnoma oblikovane kolonije hitro rastočih kultur lahko dobite 3.-4. dan inkubacije, kolonije počasi rastočih kultur - 7.-8.
Za hitro rastoče gomoljične bakterije je značilna peritrična razporeditev bičkov, medtem ko so počasi rastoče bakterije monotrihialne.
Pri gojenju na tekočem mediju se v celicah nodulnih bakterij poleg bičkov oblikujejo nitasti in kroglični izrastki, katerih dolžina doseže 8-10 mikronov. Običajno se nahajajo na površini celice peritrihialno, vsebujejo jih od 4 do 10 ali več na celico.
Kolonije hitro rastočih nodulnih bakterij imajo barvo pečenega mleka, pogosto prosojne, sluzaste, z gladkimi robovi, zmerno konveksne in sčasoma rastejo na površini agarnega medija. Kolonije počasi rastočih bakterij so bolj konveksne, majhne, suhe, goste in praviloma ne rastejo na površini gojišča. Sluz, ki jo proizvajajo nodulne bakterije, je kompleksna spojina polisaharidnega tipa, ki vključuje heksoze, pentoze in uronske kisline.
Nodulne bakterije so mikroaerofili (razvijajo se ob majhnih količinah kisika v okolju), vendar imajo raje aerobne pogoje.
Nodulne bakterije kot vir ogljika v hranilnih medijih uporabljajo ogljikove hidrate in organske kisline, kot vir dušika pa različne mineralne in organske spojine, ki vsebujejo dušik. Pri gojenju na gojiščih z visoko vsebnostjo snovi, ki vsebujejo dušik, lahko nodulne bakterije izgubijo sposobnost prodiranja v rastlino in tvorjenja nodulov. Zato nodulne bakterije običajno gojimo na rastlinskih izvlečkih (fižolova, grahova juha) ali izvlečkih iz zemlje. Fosfor, potreben za razvoj, lahko pridobijo nodulne bakterije iz mineralnih in organskih spojin, ki vsebujejo fosfor; mineralne spojine lahko služijo kot vir kalcija, kalija in drugih mineralnih elementov.
Za zatiranje tuje saprofitne mikroflore pri izolaciji nodulnih bakterij iz nodulov ali neposredno iz tal priporočamo hranilne medije z dodatkom kristalne vijolice, tanina ali antibiotikov.
Za razvoj večine kultur nodulnih bakterij je potrebna optimalna temperatura v območju 24-26 °. Pri 0° in 37°C se rast ustavi. Običajno se kulture nodulnih bakterij hranijo v laboratorijskih pogojih pri nizkih temperaturah (2-4 °C).
Številne vrste nodulnih bakterij lahko sintetizirajo vitamine B, pa tudi rastne snovi, kot je heteroavksin (beta-indolocetna kislina).
Vse gomoljne bakterije so približno enako odporne na alkalno reakcijo gojišča (pH = 8,0), vendar neenakomerno občutljive na kislo.
Specifičnost, virulentnost, kompetitivnost in aktivnost nodulnih bakterij - pojem specifičnosti nodulnih bakterij je skupen. Označuje sposobnost bakterij, da tvorijo gomolje v rastlinah. Če govorimo o nodulnih bakterijah na splošno, potem je zanje nastajanje nodulov samo v skupini stročnic že samo po sebi specifično - imajo selektivnost za stročnice.
Če pa upoštevamo posamezne kulture gomoljnih bakterij, se izkaže, da so med njimi tudi takšne, ki so sposobne okužiti le določeno, včasih večjo, včasih manjšo skupino stročnic, in v tem smislu je specifičnost nodialnih bakterij. je selektivna sposobnost glede na gostiteljsko rastlino. Specifičnost gomoljnih bakterij je lahko ozka (deteljne gomoljne bakterije okužijo le skupino detelj – vrstna specifičnost, gomoljne bakterije volčjega boba pa so lahko označene celo s sortno specifičnostjo – okužijo le alkaloidne ali brezalkaloidne sorte volčjega boba). S široko specifičnostjo lahko grahove nodulne bakterije okužijo rastline graha, brade in fižola, grahove in fižolove nodulne bakterije pa lahko okužijo rastline graha, kar pomeni, da je za vse značilna sposobnost "navzkrižne okužbe". Specifičnost nodulnih bakterij je osnova za njihovo razvrstitev.
Posebnost nodulnih bakterij je nastala kot posledica njihove dolgotrajne prilagoditve na eno rastlino ali skupino le-teh in genetskega prenosa te lastnosti. V zvezi s tem obstaja tudi različna prilagodljivost nodulnih bakterij rastlinam znotraj skupine navzkrižne okužbe. Tako lahko gomoljne bakterije lucerne tvorijo gomolje v sladki detelji. Kljub temu so bolj prilagojene na lucerno, na sladko deteljo pa so bolj prilagojene bakterije sladke detelje.
V procesu okužbe koreninskega sistema stročnic z nodulnimi bakterijami je virulentnost mikroorganizmov velikega pomena. Če specifičnost določa spekter delovanja bakterij, potem virulentnost nodulnih bakterij označuje aktivnost njihovega delovanja znotraj tega spektra. Virulenca se nanaša na sposobnost nodulnih bakterij, da prodrejo v koreninsko tkivo, se tam razmnožijo in povzročijo nastanek nodulov.
Pomembno vlogo igra ne le sama sposobnost prodiranja v korenine rastline, ampak tudi hitrost tega prodiranja.
Za določitev virulence seva nodulnih bakterij je treba ugotoviti njegovo sposobnost, da povzroči nastanek nodulov. Merilo za virulentnost katerega koli seva je lahko najmanjše število bakterij, ki zagotavlja močnejšo okužbo korenin v primerjavi z drugimi sevi, ki se zaključi z nastankom nodulov.

riž. 5
V prsti v prisotnosti drugih sevov bolj virulentni sev ne bo vedno prvi okužil rastline. V tem primeru je treba upoštevati njegovo konkurenčno sposobnost, ki pogosto prikrije lastnost virulence v naravnih razmerah.
Nujno je, da imajo virulentni sevi tudi konkurenčnost, tj. da lahko uspešno tekmujejo ne le s predstavniki lokalne saprofitske mikroflore, temveč tudi z drugimi sevi nodulnih bakterij. Indikator konkurenčnosti seva je število nodulov, ki jih tvori kot odstotek skupnega števila nodulov na rastlinskih koreninah.
Pomembna lastnost nodulnih bakterij je njihova aktivnost (učinkovitost), t.j. sposobnost asimilacije molekularnega dušika v sožitju s stročnicami in zadovoljevanja potreb gostiteljske rastline v njem. Glede na to, v kolikšni meri gomoljne bakterije prispevajo k povečanju pridelka metuljnic, jih običajno delimo na aktivne (učinkovite), neaktivne (neučinkovite) in neaktivne (neučinkovite).
Sev bakterij, neaktiven za eno gostiteljsko rastlino v simbiozi z drugo vrsto stročnic, je lahko zelo učinkovit. Zato je treba pri karakterizaciji seva glede na učinkovitost vedno navesti, v zvezi s katero gostiteljsko rastlinsko vrsto se njegov učinek kaže.
Dejavnost nodulnih bakterij ni njihova trajna last. V laboratorijski praksi pogosto pride do izgube aktivnosti v kulturah nodulnih bakterij. V tem primeru se bodisi izgubi aktivnost celotne kulture bodisi se pojavijo posamezne celice z nizko aktivnostjo. Zmanjšanje aktivnosti nodulnih bakterij se pojavi v prisotnosti nekaterih antibiotikov, aminokislin. Eden od razlogov za izgubo aktivnosti nodulnih bakterij je lahko vpliv faga. S pasažo, to je večkratnim prehajanjem bakterij skozi gostiteljsko rastlino (prilagajanje na posamezno rastlinsko vrsto), je mogoče iz neučinkovitih pridobiti učinkovite seve.
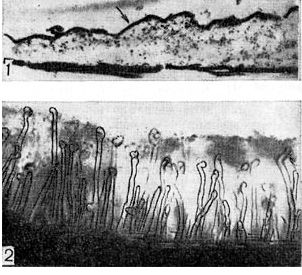
riž. 6 - Plast zrnate snovi na površini koreninskega dlaka (1) pri elektronskem mikroskopskem pregledu (po P. Dartu, F. Mercerju, povečava X 30.000) in sluzasta plast zoogleja (2), v kateri so koreninski dlaki detelje. potopljeno, pod svetlobnim mikroskopom (zoom x 80)
Izpostavljenost gama žarkom omogoča pridobivanje sevov z večjo učinkovitostjo. Znani so primeri nastanka visoko aktivnih radiomutantov nodulnih bakterij lucerne iz neaktivnega seva. Uporaba ionizirajočega sevanja, ki neposredno vpliva na spremembo genetskih lastnosti celice, je po vsej verjetnosti lahko obetavna tehnika pri selekciji visoko aktivnih sevov gomoljičnih bakterij.
Okužba rastline metuljnice z nodulnimi bakterijami - za zagotovitev normalnega procesa okužbe koreninskega sistema z nodulnimi bakterijami je potrebno imeti v območju korenin precej veliko število živih bakterijskih celic.
Med razvojem koreninskega sistema metuljnice spodbujajo razmnoževanje gomoljnih bakterij na površini korenin s koreninskimi izločki. Produkti uničenja koreninskih klobukov in laskov imajo tudi pomembno vlogo pri zagotavljanju gomoličnih bakterij primernega substrata.
Na površini korenine je plast sluzaste snovi (matriks), ki nastane ne glede na prisotnost bakterij v rizosferi. Ta plast je jasno vidna pri pregledu pod svetlobno-optičnim mikroskopom. Nodulne bakterije po inokulaciji običajno hitijo v to plast in se v njej kopičijo zaradi stimulativnega učinka korenine, ki se kaže že na razdalji do 30 mm.
Obstajajo številne hipoteze o mehanizmu prodiranja nodulnih bakterij v koren rastline. Najbolj zanimivi med njimi so naslednji. Avtorji ene izmed hipotez navajajo, da gomoljne bakterije prodrejo v korenino s poškodbo epidermalnega in kortikalnega tkiva (zlasti na mestih, kjer se stranske korenine odcepijo). To hipotezo je postavil na podlagi raziskav Bril (1888), ki je povzročil nastanek gomoljev pri metuljnicah tako, da je korenine prebodel z iglo, ki je bila predhodno potopljena v suspenzijo gomoljnih bakterij. Kot poseben primer je takšna pot izvajanja povsem realna. Na primer, pri arašidu se noduli nahajajo pretežno v pazduhah koreninskih vej, kar kaže na prodiranje nodulnih bakterij v korenino skozi vrzeli med kalitvijo stranskih korenin.
Zanimiva in ne neutemeljena hipoteza je prodiranje nodulnih bakterij v koreninsko tkivo skozi koreninske laske. Večina raziskovalcev priznava prehod nodulnih bakterij skozi koreninske laske.
Možno je, da gomoljne bakterije prodrejo v korenino skozi epidermalne celice mladih koreninskih vršičkov. Prazhmovsky (1889) lahko bakterije prodrejo v korenino le skozi mlado celično steno (koreninskih laskov ali epidermalnih celic) in so popolnoma nesposobne premagati kemično spremenjeno ali plutasto plast skorje. To lahko pojasni, da se noduli običajno razvijejo na mladih delih glavne korenine in nastajajočih stranskih koreninah.
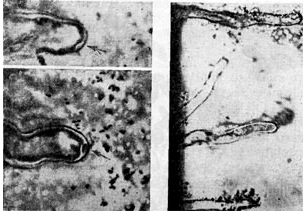
V zadnjem času je hipoteza o avksinu postala zelo priljubljena. Avtorji te hipoteze verjamejo, da gomoljne bakterije prodrejo v korenino tako, da spodbudijo sintezo p-indolocetne kisline (heteroavksina) iz triptofana, ki je vedno prisoten v rastlinskih koreninskih izločkih. Prisotnost heteroauksina je povezana z ukrivljenostjo koreninskih dlak, kar običajno opazimo, ko je koreninski sistem okužen z nodulnimi bakterijami.
riž. 7
Vir beta-indolilocetne kisline v času okužbe rastline očitno niso le rastline, ki skozi koreninski sistem izločajo triptofan, ki ga številne vrste bakterij, vključno s koreninskimi gomolji, lahko pretvorijo v beta-indolilocetno kislino. Pri sintezi heteroauksina lahko sodelujejo tudi same nodulne bakterije in morda druge vrste talnih mikroorganizmov, ki živijo v območju korenin.
Delovanje heteroavksina je nespecifično in povzroča ukrivljenost koreninskih dlačic pri različnih vrstah rastlin, ne le pri metuljnicah. Hkrati nodulne bakterije povzročajo ukrivljenost koreninskih dlačic le pri stročnicah, medtem ko kažejo precejšnjo selektivnost. Če bi obravnavani učinek določal samo β-indolocetna kislina, potem takšne specifičnosti ne bi bilo.Poleg tega je narava sprememb koreninskih laskov pod vplivom nodulnih bakterij nekoliko drugačna kot pod vplivom heteroavksina.
Prav tako je treba opozoriti, da so v nekaterih primerih neukrivljene koreninske dlake izpostavljene okužbi. Opazovanja kažejo, da je pri lucerni in grahu 60-70% koreninskih las zvitih in zvitih, pri detelji pa približno 50%. Pri nekaterih vrstah detelje je ta reakcija opažena pri največ 1/4 okuženih las. Pri reakciji ukrivljenosti je seveda zelo pomembno stanje koreninskega lasu. Rastoče koreninske dlake so najbolj občutljive na delovanje snovi, ki jih proizvajajo bakterije.
riž. 8
nodulna dušikova bakterija metuljnica
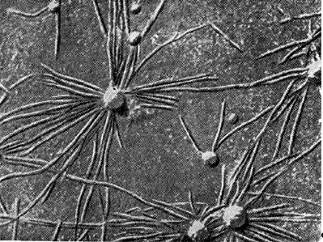
Opozoriti je treba še na eno hipotezo, po kateri nodulne bakterije vstopijo v korenino med tvorbo prstaste štrline na površini koreninskega dlaka. Elektronski difraktogram odseka koreninskega dlaka, ki potrjuje to hipotezo, kaže koreninski dlak, upognjen v obliki ročaja dežnika, v pregibu katerega se kopičijo gomoljne bakterije. Nodulne bakterije so kot da jih koreninski lasek vleče (pogoltne) (podobno kot pinocitoza).
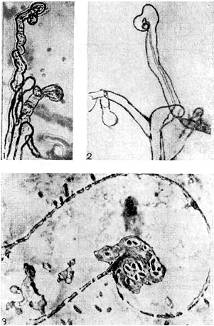
riž. 9 - Ukrivljenost koreninskih laskov stročnic v prisotnosti nodulnih bakterij: 1,2 - stola, x 120; 3 - ultratanek rez skozi koreninski las. Povečana X 10.000 (po K. Salmanu in G. Fareusu)
Proces vnosa gomoljnih bakterij v koreninsko tkivo je pri vseh vrstah stročnic enak in je sestavljen iz dveh faz. V prvi fazi pride do okužbe koreninskih laskov. V drugi fazi se intenzivno nadaljuje proces nastajanja vozličev. Trajanje faz je različno pri različnih rastlinskih vrstah: pri Trifolium fragiferum prva faza traja 6 dni, pri Trifolium nigrescens - 3 dni. V nekaterih primerih je zelo težko zaznati meje med fazami. Najintenzivnejši vnos nodulnih bakterij v koreninske dlake se zgodi v zgodnjih fazah razvoja rastline. Druga faza se konča med množično tvorbo nodul. Pogosto se prodiranje noduličnih bakterij v koreninske dlake nadaljuje tudi po tem, ko so na koreninah že nastali vozliči. Do te tako imenovane ekscesne ali dodatne okužbe pride, ker se okužba dlak ne ustavi dolgo časa. V kasnejših fazah okužbe so gomolji običajno nameščeni nižje ob korenini.
Ko prodrejo v koren (skozi koreninske dlake, epidermalne celice, mesta poškodb korenin), se nodulne bakterije nato premaknejo v tkiva korenine rastline. Najlažje bakterije prehajajo skozi medcelične prostore.
V koreninsko tkivo lahko prodre ena sama celica ali skupina bakterijskih celic. Če je bila vnesena ločena celica, se lahko še naprej premika skozi tkivo kot ena sama celica. Za rastline volčjega boba je značilen način okužbe korenin s posameznimi celicami.
Vendar pa v večini primerov invazivna celica, ki se aktivno razmnožuje, tvori tako imenovane nalezljive niti (ali nalezljive tji) in se že v obliki takih niti premakne v rastlinska tkiva.
Izraz "infekcijska nit" je nastal na podlagi preučevanja procesa okužbe v svetlobnem mikroskopu. Z delom Beijerincka so infekcijsko nit začeli obravnavati kot sluzasto hifam podobno maso z zaprto
Glede na način nastanka so noduli stročnic razdeljeni na dve vrsti: 1. vrsta - noduli nastanejo med delitvijo celic pericikla (koreninski sloj), ki se običajno nahajajo proti protoksilemu (prvi v času za nastanek žil ) - endogena vrsta tvorbe vozličev; Tip 2 - vozliči izvirajo iz koreninske skorje kot posledica vnosa patogena v parenhimske celice skorje in endoderme (notranja plast primarne skorje) - eksogeni tip tvorbe vozličev.
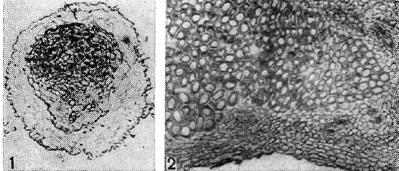
riž. 10 - Prerezi skozi nodulno tkivo stročnic: 1 - prerez skozi nodulo graha. Začetek diferenciacije žilnega tkiva v nodulu (po G. Bondu); d - vzdolžni prerez skozi nodul ranga. Povečana x90
V naravi prevladuje slednji tip. Tkiva osrednjega cilindra korenine sodelujejo samo pri tvorbi vaskularnega sistema vozličev tako endogenih kot eksogenih vrst.
Razlika v naravi tvorbe vozličev in stranskih korenin je še posebej jasno opazna pri Seradelli, saj je kortikalno tkivo glavne korenine te rastline - mesto prvih vozličev - sestavljeno iz relativno majhne plasti celic in vozliči postanejo vidni zelo hitro po okužbi korenine z bakterijami. Na korenini najprej tvorijo sploščene izrastke, po katerih jih je mogoče razlikovati od stožčastih izrastkov stranskih korenin. Nodule se od stranskih korenin razlikujejo po številnih anatomskih značilnostih: odsotnosti osrednjega valja, koreninskih pokrovčkov in povrhnjice ter prisotnosti znatne plasti lubja, ki pokriva vozlič.
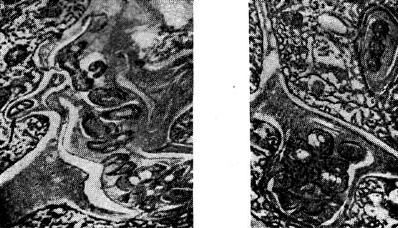
riž. enajst - Medcelične infekcijske niti v obliki žepka (desno) in bakle (levo) z nodulnimi bakterijami. Puščica prikazuje bakterijsko celico, ki se pripravlja na vstop v citoplazmo rastlinske celice. Povečana X 20.000
Nastajanje nodulov stročnic se pojavi v obdobju, ko ima korenina še primarno strukturo. Začne se z delitvijo kortikalnih celic, ki se nahajajo na razdalji 2-3 plasti od koncev nalezljivih niti. Plasti skorje, ki jih prodrejo nalezljive niti, ostanejo nespremenjene. Hkrati se pri seradeli delitev kortikalnih celic pojavi neposredno pod okuženo koreninsko dlako, medtem ko pri grahu opazimo delitev celic le v predzadnji plasti skorje.
Delitev s tvorbo radialne strukture tkiva se nadaljuje do celic notranjega jedra. Pojavi se brez določene smeri, naključno, zaradi česar nastane meristem (sistem izobraževalnih tkiv) nodula, sestavljen iz majhnih zrnatih celic.
Razdeljene celice korteksa se spremenijo: jedra se zaokrožijo in povečajo, zlasti nukleoli. Po mitozi se jedra razpršijo in se začnejo znova deliti, ne da bi prevzela prvotno obliko.
Pojavi se sekundarni meristem. Kmalu se v endodermu in periciklu pojavijo znaki začetne delitve, ki v nekdanjih zunanjih celicah poteka predvsem s tangencialnimi septumi. Ta delitev se končno razširi na skupni meristematski kompleks, katerega majhne celice se podaljšajo, vakuole izginejo, jedro zapolni večino celice. Nastane tako imenovani primarni nodul, v plazmi celic katerega ni nodulnih bakterij, saj so na tej stopnji še vedno znotraj infekcijskih niti. Medtem ko nastaja primarni vozlič, se niti okužbe večkrat razvejajo in lahko prehajajo med celicami – medcelično ali skozi celice – znotrajcelično – in vnašajo bakterije.
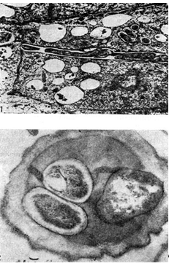
riž. 12 - Intracelularne infekcijske niti z nodulnimi bakterijami: 1 - vzdolžni prerez (po D. Goodchildu in F. Bergersenu), 2 - prečni prerez. Odpeljan, x 35000
Medcelične nalezljive niti zaradi aktivnega razmnoževanja nodulnih bakterij v njih pogosto pridobijo bizarno obliko - nastanejo v obliki žepov (divertikulov) ali bakel.
Proces premikanja okužbenih niti od celice do celice ni povsem jasen. Očitno kužne niti, kot meni kanadski mikrobiolog D. Jordan, tavajo v obliki golih sluzastih pramenov v medceličnih prostorih rastlinskega tkiva, dokler se zaradi še vedno nerazložljivih razlogov ne začnejo vriniti v citoplazmo sosednjih celic.
V nekaterih primerih pride do invaginacije okužbe v eni, v nekaterih primerih v vsaki sosednji celici. Skozi te invaginirane cevaste votline (divertikule) teče vsebina nitke, obdane s sluzjo. Najbolj aktivna rast kužnih niti se običajno pojavi v bližini jedra rastlinske celice. Prodiranje nitke spremlja gibanje jedra, ki se premika proti mestu okužbe, se povečuje, spreminja obliko in degenerira. Podobno sliko opazimo pri glivični okužbi, ko jedro pogosto hiti proti vdornim hifam, ga pritegne poškodba kot mesto največje fiziološke aktivnosti, se približa niti, nabrekne in propade. Očitno je to značilno za odziv rastline na okužbo.
V letnih rastlinah se nalezljive niti običajno pojavijo v prvem obdobju okužbe korenine, v trajnicah - v daljšem obdobju razvoja.
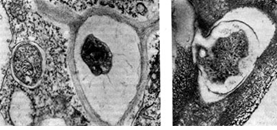
riž. 13
Bakterije se lahko sprostijo iz okužbe ob različnih časih in na različne načine. Izhod bakterij je praviloma zelo dolg proces, zlasti pri trajnicah. Običajno je sproščanje bakterij iz infekcijske niti v citoplazmo gostiteljske rastline povezano z notranjim pritiskom, ki je posledica intenzivnega razmnoževanja bakterij v niti in njihovega izločanja sluzi. Včasih bakterije zdrsnejo iz niti v skupinah, obdanih s sluzjo okužbe. v obliki veziklov (mehurjaste tvorbe). Ker vezikli nimajo membran, je izhod bakterij iz njih zelo preprost. Nodulne bakterije lahko vstopajo v rastlinske celice tudi posamično iz medceličnih prostorov.
Nodulne bakterije, ki so izšle iz okužbe, se še naprej razmnožujejo v tkivu gostitelja. Njihovo razmnoževanje v tem obdobju poteka s konstrikcijsko delitvijo. Glavnina bakterij se namnoži v citoplazmi celice in ne v niti okužbe. Okužene celice povzročijo prihodnje bakteroidno tkivo.
Napolnjene s hitro razmnoževalnimi celicami nodulnih bakterij se rastlinske celice začnejo intenzivno deliti. V trenutku mitotske delitve okuženih celic se lahko nodulne bakterije kopičijo na dveh nasprotnih polih matične celice in pasivno prehajajo v hčerinske celice. Vsaka od neokuženih celic je pod močnim stimulativnim učinkom nodulnih bakterij in se posledično tudi deli. Zahvaljujoč tej močni mitotični delitvi meristematskih celic se nodulne bakterije razširijo v nodulna tkiva in volumen bakteroidne regije se poveča.
Okuženo tkivo, sestavljeno iz gosto ležečih in aktivno delečih se celic, ima najprej obliko prisekanega stožca. Kasneje se zaradi postopne rasti tega stožca in hkratne delitve ter razvoja meristematskih celic gomoljno tkivo poveča in izgubi svojo stožčasto obliko.
Tako se nodul poveča najprej zaradi radialne in tangencialne delitve jedrnih celic, nato pa zaradi povečanja njihove velikosti in sočasne delitve. Ko so rastlinske celice popolnoma napolnjene z bakterijami, se mitoza ustavi. Vendar se celice še naprej povečujejo in so pogosto močno podaljšane. Njihova velikost je nekajkrat večja od velikosti neokuženih rastlinskih celic, ki se nahajajo med njimi v bakteroidnem območju nodula.
Povezava mladega nodula s korenino stročnic se izvaja zahvaljujoč vaskularno-vlaknastim snopom. Vaskularne fibrozne snope je prvič opazil M.S. Voronin (1866). Čas nastanka žilnega sistema v nodulih različnih vrst stročnic je različen. Torej, v sojinih nodulah začetek razvoja žilnih snopov sovpada s trenutkom prodiranja nodulnih bakterij v dve plasti kravjega parenhima. Z rastjo vozliča se prevodni sistem poveča, razveji in obda bakteroidni predel.
Vzporedno s procesom diferenciacije vaskularnega sistema se iz zunanje plasti primarnega vozliča oblikuje endoderm nodula. Nato je vozlič zaokrožen, njegova periferna celična plast je obdana s skorjo vozliča.
Koreninska povrhnjica se zlomi, vozlič pa se še naprej razvija in povečuje.
Z uporabo svetlobnega mikroskopa na vzdolžnih rezih zrelih nodulov se običajno jasno ločijo 4 značilne cone diferenciacije tkiv: skorja, meristem, bakteroidna cona in vaskularni sistem. Vsa tkiva nodulov se diferencirajo v akropetalnem zaporedju, ko meristem sproži nove celice.
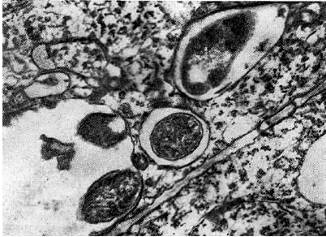
riž. 14
Lubje nodule - lupina nodule, ki opravlja zaščitno funkcijo. Lubje je sestavljeno iz več vrst neokuženih parenhimskih celic, katerih velikost in lastnosti so pri različnih metuljnicah različne. Najpogosteje imajo celice skorje podolgovato obliko in so večje v primerjavi z drugimi celicami vozlišča.
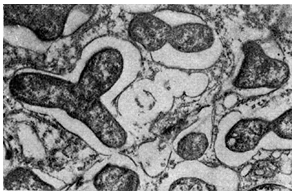
riž. 15
V lubju gomoljev trajnih lesnih vrst pogosto najdemo celice s plutastimi membranami, ki vsebujejo smole, tanin in tanine.
Nodulni meristem se nahaja pod celicami korteksa in je cona intenzivne delitve tudi neokuženih celic. Za meristem nodule so značilne gosto razporejene, brez medceličnih prostorov, majhne tankostenske celice nepravilne oblike. Nodulne meristemske celice so podobne celicam drugih vrst meristematskega tkiva (vrh korenine, vrh stebla). Nodulne meristemske celice vsebujejo gosto, fino zrnato citoplazmo z ribosomi, Golgijevimi telesi, protoplastidi, mitohondriji in drugimi strukturami. Obstajajo majhne vakuole. V središču citoplazme je veliko jedro z jedrno membrano, porami in jasno definiranim nukleolom. Funkcije meristematskih celic so tvorba celic skorje vozličkov, bakteroidne regije in vaskularnega sistema. Glede na lokacijo meristema imajo vozliči različne oblike: sferične (grah, fižol, seradella, arašidi) ali valjaste (lucerna, grašica, čin, akacija, detelja). Meristem, ki se nahaja v ločenih območjih vzdolž oboda nodula, vodi do tvorbe gomoljev podobnih gomoljev pri volčjem bobu.
Meristem vozličkov deluje dolgo časa, tudi med nekrozo nodulov, ko so že napolnjeni z lizirajočo bakteroidno maso in uničenimi rastlinskimi celicami.
Bakteroidna cona nodula zavzema njegov osrednji del in predstavlja od 16 do 50% celotne suhe mase nodulov. V prvem obdobju nastanka nodula je to v bistvu bakterijsko območje, saj je napolnjeno z bakterijskimi celicami, ki so v bakterijski in ne v bakteroidni fazi razvoja. Kljub temu, ko gre za območje nodulnega tkiva, ki vsebuje bakterije, je običajno, da ga imenujemo bakteroid.
Bakteroidno območje nodula je sestavljeno predvsem iz celic, okuženih z nodulnimi bakterijami, in delno neokuženih celic, ki mejijo na njih, napolnjene s pigmenti, tanini in do jeseni - s škrobom.
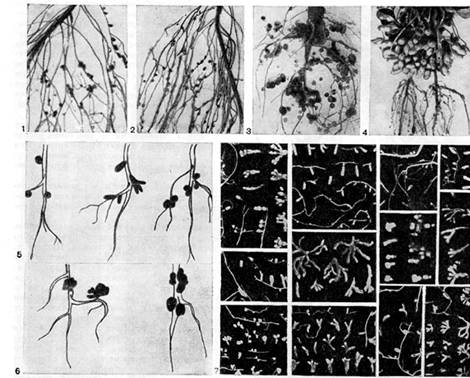
riž. 16 - Oblika nodul v stročnicah: 1, 2 - grah; 3, 6 (desno) - volčji bob; 4 - arašidi (po E. Schwinghamerju); 5 - vrste, detelja, wiki, 6 (levo) - lucerna (po J. Datumu); 7 - različne vrste divjih stročnic (po A.R. Werner, N.Ya. Gordienko)
V nodulih, ki jih tvorijo učinkoviti sevi nodulnih bakterij, je povprečna relativna prostornina bakteroidne cone večja kot v nodulih, ki nastanejo ob vnosu neučinkovitih sevov.
V nekaterih primerih obseg bakteroidne regije doseže največjo vrednost v zgodnjem obdobju življenja nodula in nato ostane relativno konstanten. Bakteroidno območje je prepredeno z gosto mrežo infekcijskih niti, po obodu pa je obdano z vaskularnimi fibroznimi snopi.
Oblika bakteroidov v nodulih različnih vrst stročnic je lahko različna. Torej, v wiki, čin in grah sta dvovejni ali viličasti. Pri detelji in esparzeti je prevladujoča oblika bakteroidov kroglasta, hruškasta, nabrekla, jajčasta in pri čičeriki okrogla oblika. Oblika bakteroidov fižola, seradele, ptičje noge in volčjega boba je skoraj paličasta.
Bakteroidi zapolnjujejo večino rastlinske celice, z izjemo osrednjega območja jedra in vakuol. Tako je odstotek bakteroidov v bakteroidnem območju rožnatega nodula 94,2 celotnega števila nodulnih bakterij. Bakteroidne celice so 3-5-krat večje od bakterijskih celic.
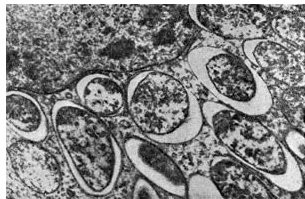
riž. 17
Bakteroidi nodulnih bakterij so še posebej zanimivi zaradi dejstva, da so skoraj edini prebivalci nodulov stročnic v obdobju intenzivne vezave atmosferskega dušika. Nekateri raziskovalci menijo, da so bakteroidi patološko degenerativne oblike in ne povezujejo procesa fiksacije dušika z bakteroidno obliko nodulnih bakterij. Večina raziskovalcev ugotavlja, da so bakteroidi najsposobnejše in najaktivnejše oblike nodulnih bakterij in da stročnice vežejo atmosferski dušik le z njihovo udeležbo.
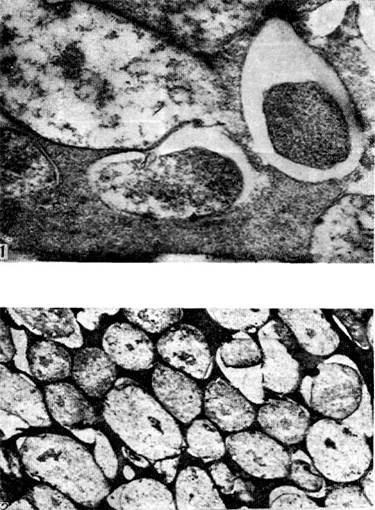
riž. 18
Žilni sistem vozliča zagotavlja povezavo med bakterijo in gostiteljsko rastlino. Hranila in presnovni produkti se prenašajo skozi žilne snope. Žilni sistem se razvije zgodaj in deluje dolgo časa.
Popolnoma oblikovane žile imajo specifično strukturo: sestavljene so iz ksilemskih traheidov, floemskih vlaken, sitastih cevk in spremljajočih celic.
Odvisno od vrste stročnic se povezava glomerula izvaja preko enega ali več žilnih snopov. Na primer, pri grahu sta na dnu nodula dva diferencirana žilna vozla. Vsak od njih se navadno dvakrat dihotomno razveji, zaradi česar skozi nodulo iz mesta druge dihotomne veje preide 8 snopov. Številne rastline imajo samo en šop, hkrati pa so lahko v enem gomolju Sesbania grandiflora pri starosti enega leta našteli do 126. Nemalokrat je žilni sistem gomolja ločen od zunanje strani gomolja. njegovo lubje s plastjo delno ali popolnoma suberiziranih celic, imenovanih nodulni endoderm, pritrjenih na koreninski endodermis. Nodulni endoderm je zunanja plast neokuženega govejega parenhima, ki se nahaja med tkivom nodula in koreninsko skorjo.
Pri večini rastlinskih vrst se vozliči oblikujejo po opisani vrsti. Zato je nastajanje vozličkov posledica kompleksnih pojavov, ki se začnejo zunaj korenine. Po začetnih fazah okužbe se sproži nastanek vozliča, nato pride do širjenja bakterij v območju tkiva vozliča in fiksacije dušika.
Vse stopnje razvoja nodulnih bakterij, po češkem mikrobiologu V. Kasu (1928), je mogoče izslediti na odsekih gomoljev. Torej, v zgornjem delu nodula, na primer, lucerna vsebuje predvsem majhne ločljive paličaste celice, majhno količino mladih bakteroidov, katerih število se postopoma povečuje z razvojem nodula. V srednjem, rožnato obarvanem delu vozliča se nahajajo pretežno bakteroidne celice, redkeje majhne paličaste celice. Na dnu gomolja v zgodnjih fazah vegetacije gostiteljske rastline so bakteroidi enaki kot v njegovem srednjem delu, vendar so do konca rastne sezone bolj nabrekli in prej degenerirajo.
Čas pojava prvih vidnih vozličev na koreninah različnih vrst stročnic je različen. Njihov pojav se pri večini metuljnic najpogosteje pojavi v času razvoja prvih pravih listov. Tako nastajanje prvih nodulov lucerne opazimo med 4. in 5. dnevom po kalitvi, 7. - 8. dan pa se ta proces pojavi pri vseh rastlinah. Nodule srpaste lucerne se pojavijo po 10 dneh.
V obdobju delovanja so vozlišča običajno gosta. Noduli, ki jih tvorijo aktivne kulture bakterij, so v mladosti belkaste barve. V času manifestacije optimalne aktivnosti postanejo rožnate. Noduli, ki so nastali med okužbo z neaktivnimi bakterijskimi kulturami, so zelenkaste barve. Pogosto se njihova struktura praktično ne razlikuje od strukture vozličev, ki nastanejo s sodelovanjem aktivnih sevov nodulnih bakterij, vendar se prezgodaj uničijo.
V nekaterih primerih struktura vozlov, ki jih tvorijo neaktivne bakterije, odstopa od norme. To se izraža v dezorganizaciji nodulnega tkiva, ki običajno izgubi jasno opredeljeno consko diferenciacijo.
Rožnato barvo določa prisotnost pigmenta v vozličih, ki je po kemični sestavi podoben krvnemu hemoglobinu. V zvezi s tem se pigment imenuje leghemoglobin (legoglobin) - Leguminosae hemoglobin. Legoglobin se nahaja le v tistih celicah vozličkov, ki vsebujejo bakteroide. Lokaliziran je v prostoru med bakteroidi in membrano, ki jih obdaja.
Njegova količina se giblje od 1 do 3 mg na 1 g gomolja, odvisno od vrste stročnic.
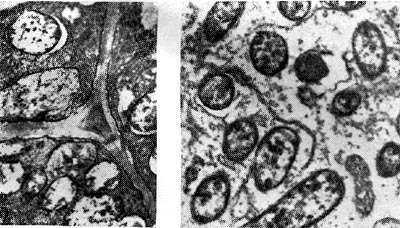
riž. 19
Pri enoletnih stročnicah se do konca rastne sezone, ko se konča proces fiksacije dušika, rdeči pigment spremeni v zelen. Spreminjanje barve se začne na dnu vozliča, kasneje njegov vrh postane zelen. Pri trajnih metuljnicah do ozelenitve gomoljev ne pride ali pa ga opazimo le na dnu gomolja. Pri različnih vrstah stročnic se prehod rdečega pigmenta v zeleni pojavi z različnimi stopnjami intenzivnosti in različno hitro.
Nodule enoletnih rastlin delujejo relativno kratek čas. Pri večini stročnic se nekroza vozličev začne v času cvetenja gostiteljske rastline in se običajno nadaljuje od središča do obrobja vozličev. Eden od prvih znakov uničenja je nastanek plasti celic z močnimi stenami na dnu vozliča. Ta plast celic, ki se nahaja pravokotno na glavno žilo korenine, jo ločuje od gomolja in upočasni izmenjavo hranil med gostiteljsko rastlino in tkivi nodula.
V celicah propadajočega tkiva nodusa se pojavijo številne vakuole, jedra izgubijo sposobnost obarvanja, nekatere celice nodusnih bakterij lizirajo, nekatere pa migrirajo v okolje v obliki majhnih kokoidnih artrospornih celic.
Stari noduli so temni, ohlapni, mehki. Pri prerezu iz njih štrli vodena sluz. Proces uničenja vozličkov, ki se začne z zamašitvijo celic vaskularnega sistema, olajša zmanjšanje fotosintetske aktivnosti rastline, suhost ali prekomerna vlažnost okolja.
V uničenem sluzastem gomolju pogosto najdemo protozoe, glive, bacile in majhne paličaste gomoljne bakterije.
Stanje gostiteljske rastline vpliva na trajanje delovanja nodule. Tako je po F. F. Yukhimchuk (1957) s kastracijo ali odstranjevanjem cvetov volčjega boba mogoče podaljšati obdobje njegove vegetacije in hkrati čas aktivnega delovanja nodulnih bakterij.
Nodule trajnih rastlin, za razliko od letnih vozličev, lahko delujejo več let. Tako ima na primer karagana večletne gomolje, v katerih se proces staranja celic pojavi sočasno z nastajanjem novih. V gliciniji (kitajski gliciniji) delujejo tudi trajnice, ki tvorijo sferične otekline na koreninah gostitelja. Do konca rastne sezone se bakteroidno tkivo trajnih gomoljev razgradi, vendar celoten nodus ne odmre. Naslednje leto ponovno začne delovati.
Dejavniki, ki določajo simbiotski odnos nodulnih bakterij s stročnicami - za simbiozo, ki zagotavlja dober razvoj rastlin, je potreben določen nabor okoljskih pogojev. Če so okoljske razmere neugodne, bo učinkovitost simbioze kljub visoki virulentnosti, tekmovalni sposobnosti in aktivnosti mikrosimbionta nizka.
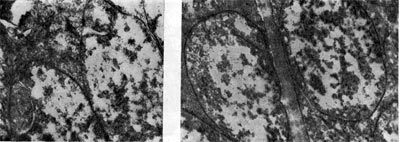
riž. 20
Za razvoj nodulov je optimalna vsebnost vlage 60-70% celotne vlažnosti tal. Minimalna vlažnost tal, pri kateri je še možen razvoj gomoljnih bakterij v tleh, je približno enaka 16 % celotne kapacitete vlage. Ko je vlažnost pod to mejo, se gomoljne bakterije navadno ne razmnožujejo več, kljub temu pa ne odmrejo in lahko ostanejo dolgo časa v neaktivnem stanju. Pomanjkanje vlage vodi tudi do smrti že oblikovanih vozličev.
Presežek vlage, pa tudi njeno pomanjkanje, je tudi neugoden za simbiozo - zaradi zmanjšanja stopnje prezračevanja v koreninskem območju se poslabša oskrba s kisikom koreninskega sistema rastline. Nezadostno prezračevanje negativno vpliva tudi na gomoljne bakterije, živeče v tleh, ki se, kot veste, bolje razmnožujejo, če je na voljo kisik. Kljub temu visoko prezračevanje v koreninskem območju vodi do dejstva, da reduktorji molekularnega dušika začnejo vezati kisik, kar zmanjša stopnjo fiksacije dušika gomoljev.
Temperaturni dejavnik igra pomembno vlogo v razmerju med nodulnimi bakterijami in stročnicami. Temperaturne značilnosti različnih vrst stročnic so različne. Poleg tega imajo različni sevi nodulnih bakterij svoje specifične temperaturne optimume za razvoj in aktivno fiksacijo dušika. Treba je opozoriti, da optimalne temperature za razvoj stročnic, nastanek nodulov in fiksacijo dušika ne sovpadajo. Torej, v naravnih razmerah lahko opazimo nastanek vozličev pri temperaturah nekoliko nad 0 ° C, fiksacija dušika v takih pogojih praktično ne poteka. Morda samo arktične simbiotske stročnice vežejo dušik pri zelo nizkih temperaturah. Običajno se ta proces zgodi le pri 10 °C in več. Največjo fiksacijo dušika pri številnih rastlinah stročnic opazimo pri 20-25 °C. Temperature nad 30°C negativno vplivajo na proces kopičenja dušika.
Reakcija tal ima velik vpliv na vitalno aktivnost gomoljnih bakterij in nastanek gomoljev. Za različne vrste in celo seve nodulnih bakterij je pH vrednost habitata nekoliko drugačna. Na primer, bakterije iz gomoljev detelje so bolj odporne na nizke vrednosti pH kot bakterije iz nodul lucerne. Očitno tu vpliva tudi prilagoditev mikroorganizmov na okolje. Detelja uspeva v bolj kislih tleh kot lucerna. Reakcija tal kot ekološki dejavnik vpliva na aktivnost in virulenco nodulnih bakterij. Najbolj aktivne seve je lažje izolirati iz tal z nevtralnimi pH vrednostmi. V kislih tleh so pogostejši neaktivni in šibko virulentni sevi. Kislo okolje (pH 4,0 - 4,5) ima neposreden učinek na rastline, zlasti moti sintetične procese rastlinskega metabolizma in normalen razvoj koreninskih laskov. V kislem okolju v inokuliranih rastlinah se obdobje delovanja bakteroidnega tkiva močno zmanjša, kar vodi do zmanjšanja stopnje fiksacije dušika.
V kislih tleh, kot ugotavlja A.V. Petersburgu soli aluminija in mangana prehajajo v raztopino tal, kar negativno vpliva na razvoj koreninskega sistema rastlin in proces asimilacije dušika, zmanjša pa se tudi vsebnost asimiliranih oblik fosforja, kalcija, molibdena in ogljikovega dioksida. Neugodno reakcijo tal najbolje odpravimo z apnenjem.
Gostiteljsko tkivo je za nodulne bakterije tako hranilno gojišče, ki lahko zaradi vsebnosti vseh vrst hranil v tkivu zadovolji tudi najzahtevnejši sev. Kljub temu pa po vnosu nodulnih bakterij v tkivo gostiteljske rastline njihov razvoj ne določajo le notranji procesi, ampak je v veliki meri odvisen tudi od delovanja zunanjih dejavnikov, ki vplivajo na celoten potek infekcijskega procesa. Vsebnost ali odsotnost enega ali drugega hranila v okolju je lahko odločilni trenutek za manifestacijo simbiotske fiksacije dušika.
Stopnja oskrbe rastlin stročnic z razpoložljivimi oblikami mineralnih dušikovih spojin določa učinkovitost simbioze. Na podlagi številnih laboratorijskih in vegetativnih poskusov je znano, da več kot je dušikovih spojin v okolju, težje bakterije prodrejo v korenino.
Velik pomen pri aktiviranju absorpcije dušika s stročnicami je prehrana s fosforjem. Pri nizki vsebnosti fosforja v mediju bakterije prodrejo v korenino, vendar se vozliči ne tvorijo. Stročnice imajo nekatere posebnosti pri izmenjavi spojin, ki vsebujejo fosfor. Semena stročnic vsebujejo veliko fosforja. Rezervni fosfor med kalitvijo semen se ne porabi tako kot pri drugih pridelkih - razmeroma enakomerno za tvorbo vseh organov, v večji meri pa koncentriran v koreninah. Zato stročnice v zgodnjih fazah razvoja, za razliko od žit, v večji meri zadovoljujejo svoje potrebe po fosforju na račun kličnih listov in ne zalog v tleh. Večja kot so semena, manj so stročnice odvisne od fosforja v tleh. Vendar pa je pri simbiotskem načinu obstoja potreba po fosforju pri stročnicah večja kot pri avtotrofnem. Zato se ob pomanjkanju fosforja v gojišču inokuliranih rastlin poslabša oskrba rastlin z dušikom.
Simbiotska fiksacija dušika zahteva tudi magnezij, žveplo in železo. S pomanjkanjem magnezija je razmnoževanje nodulnih bakterij zavirano, njihova vitalna aktivnost se zmanjša, simbiotska fiksacija dušika je zatrta. Žveplo in železo ugodno vplivata tudi na nastanek nodulov in proces fiksacije dušika, še posebej pa imata nedvomno vlogo pri sintezi leghemoglobina.
Od elementov v sledovih izpostavljamo predvsem vlogo molibdena in bora. S pomanjkanjem molibdena se vozliči slabo oblikujejo, v njih je motena sinteza prostih aminokislin, sinteza leghemoglobina pa je zatrta. Molibden skupaj z drugimi elementi s spremenljivo valenco (Fe, Co, Cu) služi kot posrednik pri prenosu elektronov v redoks encimskih reakcijah. Pri pomanjkanju bora se v vozličih ne tvorijo žilni snopi, zaradi česar je razvoj bakteroidnega tkiva moten.
Na nastanek nodulov pri metuljnicah velik vpliv ima presnova ogljikovih hidratov rastlin, ki jo določajo številni dejavniki: fotosinteza, prisotnost ogljikovega dioksida v okolju in fiziološke značilnosti rastlin. Izboljšanje prehrane z ogljikovimi hidrati pozitivno vpliva na proces inokulacije in kopičenje dušika. S praktičnega vidika je zelo zanimiva uporaba slame in slamnatega svežega gnoja za gnojenje stročnic kot vira ogljikovih hidratov. Toda v prvem letu po vnosu slame v tla se med njeno razgradnjo kopičijo strupene snovi. Treba je opozoriti, da niso vse vrste stročnic občutljive na strupene produkte razgradnje slame; grah na primer ne reagira nanje.