В ядрах простейших, так же как и в ядрах многоклеточных, имеется оболочка, ядерный сок (кариолимфа), хроматин (хромосомы) и ядрышки. Однако по размерам и строению ядра разные простейшие весьма разнообразны (рис.). Эти различия обусловлены соотношением структурных компонентов ядра: количеством ядерного сока, количеством и размерами ядрышек (нуклеол), степенью сохранения строения хромосом в интерфазном ядре и т. п.
Рис. Ядра простейших.
А - микронуклеус инфузории Paramecium aurelia;
Б - жгутиконосец Trypanosoma brucei,
В - амеба Amoeba sphaeronuclei;
Г - панцирный жгутиконосец Geratium fuscus;
Д - радиолярия Aulacantha scolymantha;
E - макрогамета кокцидий Aggregata eberthi;
Ж - макронуклеус инфузории Epidinium ecaudatum
У большинства простейших имеется одно ядро. Однако встречаются и многоядерные виды простейших.
У некоторых простейших, а именно у инфузорий и немногих корненожек - фораминифер, наблюдается интересное явление дуализма (двойственности) ядерного аппарата. Оно сводится к тому, что в теле простейшего имеются два ядра двух категорий, различающиеся как по своему строению, так и по физиологической роли в клетке. У инфузорий, например, имеется два типа ядер: большое, богатое хроматином ядро - макронуклеус и маленькое ядро - микронуклеус. Первое связано с выполнением вегетативных функций в клетке, второе-с половым процессом.
Простейшим, как и всем организмам, свойственно размножение. Существуют две основные формы размножения простейших: бесполое и половое. В основе того и другого лежит процесс деления клетки.
При бесполом размножении число особей возрастает в результате деления. Например, амеба при бесполом размножении делится на две амебы путем перетяжки тела. Процесс этот начинается с ядра, а затем захватывает цитоплазму. Иногда бесполое размножение приобретает характер множественного деления. При этом ядро предварительно делится несколько раз и простейшее становится многоядерным. Вслед за этим цитоплазма распадается на число отдельностей, соответствующих количеству ядер.
В результате организм простейшего сразу дает начало значительному количеству мелких особей. Так происходит, например, бесполое размножение малярийного плазмодия - возбудителя малярии человека.
Половое размножение простейших характеризуется тем, что собственно размножению (увеличению числа особей) предшествует половой процесс, характерным признаком которого является слияние двух половых клеток (гамет) или двух половых ядер, ведущее к образованию одной клетки - зиготы, дающей начало новому поколению.
Формы полового процесса и полового размножения у простейших в высшей степени разнообразны.
Для размножения простейших характерны два типа процессов. Один из них - агамное или бесполое размножение - сводится к увеличению числа особей данного вида и может происходить различными способами. Особи, образованные в результате такого размножения также могут иметь разную судьбу. Они могут непосредственно вырастать в организм, подобный материнскому, проходить через метаморфоз (как бродяжки сукторий или перитрих), целиком превращаться в гамету, или гаметоцит (как спорозоиты грегарин).
Кроме того, в жизненный цикл многих протист включается половой процесс. Он связан с образованием специальных стадий - гамет, которые могут формироваться разными способами. Половой процесс может вовсе не сопровождаться размножением (конъюгация, автогамия, партеногенез). Наиболее существенным моментом полового процесса является перестройка ядра и других компонентов клетки, протекающая через ряд фаз и приводящая к образованию нового поколения, отличающегося по своей жизнеспособности и наследственным свойствам от предыдущих поколений. Как уже отмечалось, не все типы полового процесса приводят к увеличению числа особей, поэтому в отношении простейших обычно говорят о наличии у них бесполого размножения с одной стороны и полового процесса с другой.
^
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Простое деление надвое или монотомия. Этот тип размножения наиболее распространен у простейших и клеток тканей многоклеточных. В типичном случае оно представляет собой правильное равномерное разделение тела простейшего на две одинаковые дочерние особи, этому предшествует период питания и роста. Классический пример - размножение амеб. У амеб деление не имеет определенной ориентации. Тело амеб вытягивается, на экваторе намечается поперечная перетяжка, она постепенно углубляется и, наконец, клетка разделяется на две половины, которые немедленно начинают самостоятельно двигаться и питаться. Деление цитоплазмы сопровождается митотическим делением ядра. Остальные клеточные органоиды либо сами делятся, либо просто распределяются между дочерними особями, а недостающие образуются заново. Такое же неориентированное деление наблюдается у части солнечников и радиолярий.
Наличие скелета, естественно, несколько усложняет этот процесс. У раковинных амеб - тестацей (Arcella) и громиид (Euglypha) перед делением в специальных вакуолях закладываются зачатки кремнеземных пластинок раковины. Затем начинается деление, плоскость которого расположена перпендикулярно продольной оси раковины. Ядро делится и примерно половина цитоплазмы вместе с дочерним ядром и кремнеземными пластинками выходит из устья наружу и окружается новой раковиной. У радиолярий, имеющих скелет из двух створок, положение плоскости деления определяется положением створок. При делении створки расходятся вместе с половиной цитоплазмы, и достраивается недостающая створка.
У жгутиконосцев и цилиат положение плоскости деления более постоянно и стабильно. У жгутиконосцев оно, как правило, происходит в продольном направлении, у инфузорий - в поперечном. У жгутиконосцев деление начинается с переднего конца тела и продолжается к заднему концу. При этом бывает, что отделяющиеся половины клетки отгибаются в противоположные стороны, что создает впечатление поперечного деления. У динофлагеллят плоскость деления отклонена от продольной - деление происходит в косом направлении. Также в косом направлении проходит плоскость деления у опалин.
У сложно организованных мастигофор деление связано с явлениями реорганизации. При этом некоторые органеллы резорбируются и формируются заново у дочерних особей. Подобные явления могут происходить с базальными зернами жгутика, аксостилем и жгутиками полимастигин.
У инфузорий процесс деления приводит к еще более сложной реорганизации, чем у жгутиковых. Так у гипотрих у передней дочерней особи старые адоральные мембранеллы постепенно уменьшаются в размерах, и, в то же время, у их основания появляются зачатки новых мембранелл, которые растут по мере резорбции старых. Точно так же заменяются брюшные цирри, так что обе дочерние особи получают полностью новый набор локомоторных органелл.
Особый случай реорганизации инфузорий - стоматогенез. Стоматогенез - морфогенетический процесс, который можно наблюдать при делении, регенерации инфузорий, а также при трансформации микростомных форм в макростомные. Первый этап стоматогенеза - образование новых кинетосом, составляющих зачаток околоротовой цилиатуры. Этот этап у разных групп инфузорий протекает по-разному, что служит основанием для филогенетических и систематических построений.
Исходной формой стоматогенеза считается телокинетальный - образование зачатков цилиатуры, при котором кинетосомы происходят от передних отрезков соматических кинет или же соседних с ними фрагментов кинет. Он характерен для представителей класса Kinetofragminophorea.
Многие гименостоматы, спиротрихи и перитрихи имеют буккокинетальный стоматогенез - новые кинетосомы возникают рядом с кинетосомами старого ротового аппарата. При этом базальные тела специализированных отделов перистома дают начало разным компонентам нового рта. Разделение двух новообразованных комплексов ресничек происходит путем серии сложных перемещений, типа сдвигов и поворотов.
Паракинетальный стоматогенез встречается у некоторых гименостомат и спиротрих. При нем старые ротовые структуры рассасываются, новые возникают за счет (или вблизи) соматических ресничных рядов на вентральной стороне организма.
Если не удается установить никакой связи между новыми кинетосомами и прежними ротовыми структурами, говорят об апокинетном стоматогенезе.
Непосредственно после образования базальных телец на них уже обнаруживаются различные системы микротрубочек, как, например, нематодесмы, постцилиарные и трансверсальные микротрубочки. Затем кинетосомы мигрируют к плазмалемме, где начинается рост ресничек.
На следующем этапе стоматогенеза кинетосомы выстраиваются в пределах зачатка в определенном порядке. Этот процесс происходит в определенном направлении и в строго определенной последовательности. В конце стоматогенеза развивается аппарат захват пищи - зона, от которой отшнуровываются пищевые вакуоли.
Палинтомия . Палинтомией называется деление надвое, повторяющееся многократно, без промежуточной стадии питания и роста. Характерными чертами палинтомии являются: 1. Предшествующий гипертрофический рост материнской особи, что дает ей возможность делиться многократно. 2. Палинтомия очень часто наступает в периоды жизненного цикла, которым предшествует усиленное питание организма. 3. Палинтомия обычно характеризуется временной дедифференцировкой продуктов деления, так как продукты деления не успевают пройти процесс реорганизации. Поэтому палинтомия часто проходит на стадии цисты. Палинтомию подразделяют на линейную, когда дочерние клетки располагаются в виде цепочки (динофлагелляты, инфузории-апостоматы) и табличную, которая возникает в результате чередования продольных делений в двух взаимно перпендикулярных плоскостях, вследствие чего продукты деления располагаются параллельно друг другу в плоскую табличку. Такая палинтомия характерна для фитомонад, некоторых динофлагеллят и эвгленид. Часто палинтомия наблюдается при образовании гаметоцитов и гамет.
Почкование . Почкование выводят из неравномерного деления. При этом морфологически различают материнскую особь (трофонт) и дочернюю (бродяжку, телотрох, почку, томит). Различают монотомическое почкование, когда образуется одна бродяжка, и множественное почкование. Кроме того, у разных групп инфузорий имеется внутреннее почкование, наружное и некоторые переходные формы почкования. Почкование характерно для сидячих инфузорий - сукторий, перитрих, хонотрих, ринходид, встречается у солнечников. Характерной особенностью почкования является отсутствие реорганизаций у материнских особей - они, как правило, сохраняют неизменными все, в том числе локомоторные или ротовые органеллы. Среди мастигофор почкование встречается крайне редко, в частности известно простое наружное почкование у Haplozoon и некоторых хризомонад. У солнечников известно множественное наружное почкование. У некоторых миксоспоридий отмечается обособление и отделение участков тела плазмодиальных стадий.
^ Синтомия или множественное деление . При этом типе размножения животное на определенном этапе жизненного цикла (обычно после периода питания и роста) испытывает серию повторных делений ядра и становится временно многоядерным, а затем сразу распадается на соответствующее количество клеток. В зависимости от того, образуются ли в результате процесса вегетативные особи или половые клетки, размножающаяся посредством синтомии стадия называется шизонтом или гамонтом, а когда множественное деление следует непосредственно за половым процессом - споронтом. Продукты шизогонии, соответственно, называются мерозоитами, гамогонии - гаметами, а спорогонии - спорозоитами для споровиков и зооспорами для других групп. Синтомия широко распространена среди споровиков, реже встречается у мастигофор (динофлагелляты - Noctiluca) и отсутствует у инфузорий.
Плазматомия. Плазматомией называют тип бесполого размножения, при котором тело многоядерных простейших делится на две или большее число многоядерных же дочерних особей. При этом деление клетки происходит несогласованно с ядерными делениями и независимо от них. К плазматомии относят деление плазмодиев слизневиков, миксоспоридий, амеб - пеломиксин и др. Некоторые авторы ставят под сомнение выделение этого способа размножения в особый тип.
^ Размножение колоний. Еще один тип бесполого размножения. Выделяют три способа размножения колоний. Первый заключается в делении целой колонии надвое. Таким способом размножаются колониальные радиолярии и хризомонады. Второй способ сводится к отделению от колонии особей (бродяжек), которые закладывают новые колонии. Этот способ встречается у сидячих жгутиконосцев и перитрих (Zoothamnium arbuscula). Третий способ сходен со вторым, но здесь бродяжки не образуются, а особи-основательницы остаются в составе старой колонии и дают начало дочерней колонии. Он характерен для колониальных вольвоцин.
Следует отметить, что колониальность также рассматривают как результат не доведенного до конца бесполого размножения и, поскольку колониальность считается эволюционной основой многоклеточности, то последняя тоже в какой-то мере может выводится из бесполого размножения.
Основной формой размножения простейших является монотомия –простое деление надвое. Между двумя делениями животное приобретает все утраченные в процессе деления структуры, активно питается, растет и к началу нового деления успевает достичь размеров материнской особи (амеба, эвглена, цератиум и др.).
Особую форму размножения простейших представляет палинтомия или повторное деление. Деление повторяется много раз без последующего периода питания и роста, а промежуточные продукты палинтомии остаются недифференцированными, и размеры их с каждым делением уменьшаются. Только последнее поколение достигает полного развития, питается и растет. Палинтомическое деление обычно протекает под покровом защитных оболочек (деление ооцисты у малярийного плазмодия, зиготы у вольвокса и т.п.).
У простейших также часто встречается множественное деление –синтомия (шизогония). При этом сначала происходит многократное деление ядра без разделения цитоплазмы, вследствие чего клетка временно становится полиэнергидной, затем цитоплазма распадается сразу, на много (соответственно числу ядер) дочерних клеток (трипанозомы, малярийный плазмодий, фораминиферы).
Половой процесс у простейших
Половой процесс может проходить у простейших в виде копуляции и конъюгации . При копуляции происходит полное слияние двух половых особей – гамет. Процесс, когда гаметы ничем не отличаются друг от друга, называется изогамией. В других случаях образуются крупные макрогаметы (женские) и мелкие микрогаметы (мужские); возникает анизогамия. Процесс, когда макрогаметы становятся неподвижными и накапливают в себе запасные питательные вещества, называется оогамией.
Конъюгация представляет собой более сложную форму полового процесса и встречается только у инфузорий. При этом происходит лишь временное соединение двух особей и обмен частями микронуклеусов. Настоящие гаметы при этом не образуются, но мигрирующее ядро одето тонким слоем цитоплазмы и соответствует микрогамете, а остальное тело инфузории со стационарным ядром представляет собой макрогамету. Мигрирующее (мужское) ядро каждого конъюганта переходит в тело партнера и копулирует с его стационарным (женским) ядром – происходит перекрестное оплодотворение. Макронуклеусы в этом процессе не участвуют.
Ядерные циклы
С половым процессом связан ядерный цикл. Общим для всех ядерных циклов является чередование гаплоидного и диплоидного поколений. Гаметы всегда гаплоидны, при их слиянии получается диплоидная зигота, на какой-то стадии жизненного цикла происходит редукционное деление (мейоз), которое возвращает ядра к гаплоидному состоянию.
Существуют три типа ядерных циклов, каждый из которых характеризуется тем, какое место в этом цикле занимает редукционное деление (рис. 3).
1) Циклы с зиготической редукцией. Большая часть жизненного цикла приходится на гаплоидное поколение. Диплоидное поколение представлено одной зиготой, которая делится редукционно, при этом восстанавливается гаплоидность. Такие циклы свойственны Phytomonadida, Sporozoa, части Polymastigida, части Hypermastlgida.
2) Циклы с гаметической редукцией. Большая часть цикла приходится на диплоидное поколение. Гаплоидное поколение представлено только гаметами, редукционное деление происходит прямо перед их образованием. Такие циклы характерны для остальных Polymastigida и Hypermaatlgida, для Heliozoa, Ciliophora, Opalinida, Cnidosporidia и почти всех Metazoa.
3) Циклы с промежуточной редукцией. Гаплоидное и диплоидное поколение занимают примерно равные части цикла: зигота делится ми-тотически. Возникают более развитые стадии диплоидного поколения (агамонт). В какой-то момент происходит редукционное деление с образованием одноядерных гаплоидных агамет – агамное (бесполое) размножение. Агаметы превращаются в более развитые стадии гаплоидного поколения. Затем это взрослое гаплоидное поколение (гамонт) распадается на гаметы, которые копулируют – происходит половой процесс. С образованием зиготы цикл замыкается. Такой цикл встречается только у фораминифер и в виде редкого исключения – у некоторых коловраток.
Жизненные циклы
Жизненные циклы простейших очень разнообразны. Есть простейшие, у которых имеется цикл без полового процесса, т.е. без ядерного цикла (например, у трипанозом, у которых чередуются разные вегетативные стадии).
В наиболее дифференцированном жизненном цикле различают четыре периода:
1) прогамный, завершающийся формированием гамет;
2) сингамный, включающий в себя копуляцию гамет и образование зиготы;
3) метагамный, охватывающий несколько поколений спорозоитов;
4) вегетативный, в течение которого происходит рост, увеличение массы особей.
В какой-то части жизненного цикла происходит редукция числа хромосом.
Для удобства сравнения жизненных циклов составим таблицу, которую разделим по вертикали на 4 части, соответственно указанным четырем периодам. В эту таблицу поместим несколько циклов простейших и Цикл многоклеточного организма, разместив периоды их циклов в соответствующих столбиках (рис. 4). Выберем простейших с наиболее простым и с наиболее расчлененным циклами.
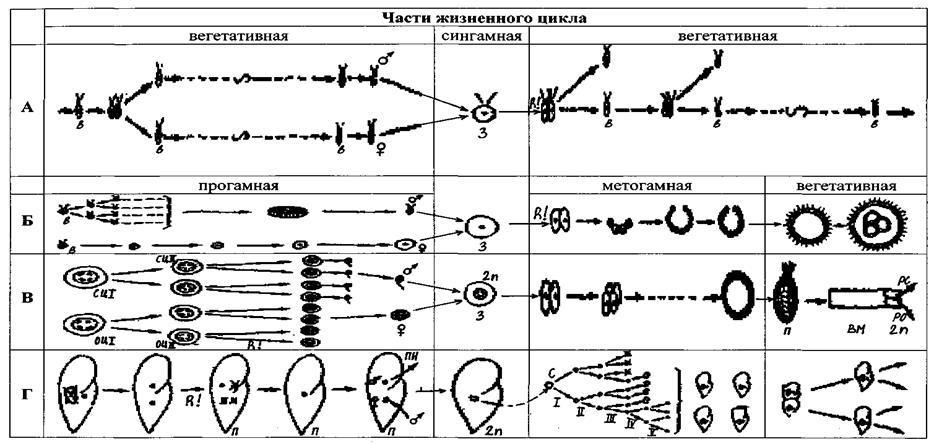
Рис 4. Сравнение жизненных циклов различных простейших и многоклеточных (из К.В. Беклемишева, 1979):
А – неопределенно многозвенный цикл низших фитомонад с факультативным половым процессом; Б - цикл вольвокса с разделением цикла на вегетативную, прогамную и метагамную части; В – цикл многоклеточных с усилением различий между частями цикла; Г – цикл инфузорий (в метагамной части цикла показано только деление ядер, контуры клеток не изображены). 3 – зигота, С – синкарион, R! – редукционное деление, ПН – пронуклеусы, П – паренхимула, рс – размножение и рост спорогониев, ро – размножение и рост оогониев, ВМ – взрослое многоклеточное, сцI и сцП – сперматоциты I и П порядка, оцI и оцП – ооциты I и П порядка
Из всего многообразия жизненных циклов простейших с половым процессом наиболее простым циклом обладают моноэнергидные жгутиконосцы, например, Chlamydomonas. У некоторых видов этого рода наблюдается множество бесполых поколений, которые образуются путем монотомии, половой процесс возникает при ухудшении условий существования. Гаметы морфологически мало отличаются от особей бесполых поколений. После копуляции зигота одевается оболочкой, под покровом которой путем палинтомического деления образуется 4–32 особи, которые выходят из оболочки и приступают к бесполому размножению. Редукция числа хромосом происходит при первых делениях зиготы. В этом цикле все стадии гаплоидны, единственная диплоидная стадия – зигота. Это цикл с зиготической редукцией. У других видов рода Chlamydomonas бесполое размножение происходит путем палинтомии, гаметы также возникают палинтомически.
Наиболее расчлененный жизненный цикл у «многоклеточного» простейшего – вольвокса. В курсе зоологии этому объекту уделяется мало внимания, т.к. он обладает аутотрофным типом питания. Однако вольвокс занимает важное место в сравнительной анатомии беспозвоночных, так как его жизненный цикл наиболее близок к циклу многоклеточных. Жизненный цикл вольвокса характеризуется строго фиксированным местом полового процесса, которому предшествует прогамная часть цикла, в течение которой происходит гамогония, т.е. превращение особых генеративных клеток в гаметы: макрогамонт растет, накапливает в себе питательные вещества и превращается в крупную неподвижную макрогамету, микрогамонт палинтомически делится, давая начало большому количеству микрогамет. Затем следует слияние гамет (сингамная часть цикла) и редукционное деление. Далее – метагамная часть – зигота, делится палинтомически, приводя к образованию бластулоподобной колонии, очень напоминающей стадию бластулы многоклеточных организмов; эта колония растет, внутри нее образуются новые дочерние колонии – после метагамной появляется еще вегетативная часть цикла. В жизненном цикле Volvox следует обратить внимание на то, что развитие дочерней колонии из зиготы происходит сначала в состоянии инкурвации (т.е. клетки, образующиеся в результате палинтомического деления зиготы, обращены жгутиковыми концами внутрь полого шара), и лишь перед выходом сформировавшейся дочерней колонии из материнской происходит экскурвация – выворачивание, в результате которого клетки принимают окончательное расположение жгутиками наружу. Редукционное деление у Volvox,как и у споровиков, происходит после образования зиготы – цикл с зиготической редукцией. Очень своеобразен цикл у инфузорий. В учебниках зоологии дается описание цикла инфузорий без учета ядерного цикла. Поэтому следует обратить внимание на то, что у них в прогамной части участвует только микронуклеус, претерпевший редукционное деление, из четырех образовавшихся гаплоидных ядер три редуцируются, остается одно, которое делится с образованием двух пронуклеусов. Женский пронуклеус превращается в стационарное ядро, остается в инфузории, и таким образом вся она соответствует макрогамете, а мужской – превращается в мигрирующее ядро, соответствующее микрогамете, и переходит в партнера, сливаясь с его стационарным ядром (сингамная часть цикла). В метагамной части цикла инфузорий палинтомия не наблюдается; происходит деление синкариона, приводящее после ряда перестроек к восстановлению двухядерности каждой особи и к восстановлению полиплоидности макронуклеуса. В вегетативной части цикла инфузории растут и размножаются бесполым путем. Это цикл с гаметической редукцией.
В жизненном цикле любого многоклеточного организма так же, как и у Volvox, выделяются прогамная, сингамная, метагамная и вегетативная части. Но прогамная часть усложняется за счет того, что у многоклеточных в этот период наблюдается сложная перестройка ядерного аппарата: образование тетраплоидных оо- и сперматоцитов I порядка, из них – диплоидных оо- и сперматоцитов П порядка и, наконец, редукционное деление, дающее начало гаплоидным гаметам.
Метагамная палинтомическая часть цикла многоклеточных очень сходна с таковой у фитомонад. Однако вегетативная часть цикла отлична: она характеризуется высокой степенью интеграции организма и большой разнокачественностыо клеток.
Из таблицы видно, что из рассмотренных простейших наиболее близок к жизненному циклу многоклеточных цикл вольвокса. Его метагамная часть завершается образованием шаровидной бластулообразной стадии, в нем так же, как и у Metazoa, есть вегетативная часть, в течение которой организм (или колония) растет, клетки его дифференцируются, и лишь небольшая часть их специализируется в генеративные. Основным отличием жизненного цикла вольвокса от цикла многоклеточных является место редукционного деления. Такое подробное сравнение циклов различных простейших и многоклеточных необходимо для понимания происхождения многоклеточных животных. Следует, однако, четко представлять, что вольвокс ни в коем случае не следует рассматривать в качестве прямого предка многоклеточных. Этот объект привлекается лишь как модель, удобная для понимания тех процессов, которые могли происходить в эволюции колониальных гетеротрофных жгутиконосцев, скорее всего близких к современным Choanoflagellata, которых в настоящее время считают предками многоклеточных. Среди современных воротничковых жгутиконосцев нет видов с половым процессом, но считают, что именно такие колониальные Choanoflagellata, имевшие форму шара и жизненный цикл типа Volvox, но с гаметической редукцией, и были предками многоклеточных.
Бесполое - митозом ядра и делением клетки надвое (у амебы, эвглены, инфузории), а также путем шизогонии - многократного деления (у споровиков).
Половое - копуляция . Клетка простейшего становится функциональной гаметой; в результате слияния гамет образуется зигота.
Для инфузорий характерен половой процесс - конъюгация . Он заключается в том, что клетки обмениваются генетической информацией, но увеличения числа особей не происходит.
Многие простейшие способны существовать в двух формах - трофозоита (вегетативной формы, способной к активному питанию и передвижению) и цисты , которая образуется при неблагоприятных условиях. Клетка обездвиживается, обезвоживается, покрывается плотной оболочкой, обмен веществ резко замедляется. В такой форме простейшие легко переносятся на большие расстояния животными, ветром и расселяются. При попадании в благоприятные условия обитания происходит эксцистирование, клетка начинает функционировать в состоянии трофозоита. Таким образом, инцистирование не является способом размножения, но помогает клетке переживать неблагоприятные условия среды.
Для многих представителей типа Protozoa характерно наличие жизненного цикла , состоящего в закономерном чередовании жизненных форм. Как правило, происходит смена поколений с бесполым и половым размножением. Образование цисты не является частью закономерного жизненного цикла.
Время генерации для простейших составляет 6-24 ч. Это означает, что, попав в организм хозяина, клетки начинают размножаться по экспоненте и теоретически могут привести его к гибели. Однако этого не происходит, так как вступают в силу защитные механизмы организма хозяина.
Медицинское значение имеют представители простейших, относящиеся к классам саркодовые, жгутиковые, инфузории и споровики.
К типу Простейшие относят организмы, тело которых состоит из одной клетки, функционирующей, однако, как целый организм. Клетки простейших способны к самостоятельному питанию, передвижению, защите от врагов и к переживанию неблагоприятных условий.
Питание простейших происходит с помощью пищеварительных вакуолей, осуществляется за счет фаго - или пиноцитоза. Остатки непереваренной пищи выбрасываются наружу. Некоторые простейшие содержат хлоропласты и способны питаться за счет фотосинтеза.
Большинство простейших имеют органеллы передвижения: жгутики, реснички и псевдоподии (временные подвижные выросты цитоплазмы). Формы органелл движения лежат в основе систематики простейших.
Размножение простейших осуществляется обычно разными формами деления - разновидностями митоза. Характерен также половой процесс: в виде слияния клеток - копуляция-или обмен частью наследственного материала - конъюгация.
Большинство простейших имеют одно ядро, но встречаются и многоядерные формы.
Медицинское значение имеют простейшие, относящиеся к классам Саркодовые, Жгутиковые, Инфузории и Споровики.