Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Введение
1.Структура и функции компонентов клетки
2.Митотический цикл. Факторы, влияющие на митотическую активность
3.Критика представлений о фатальности наследственных заболеваний
Литература
Введение
Клетка была открыта во второй половине 17 века. Особенно сильно изучение клетки развернулось во второй половине 19 века в связи с созданием клеточной теории. Клеточный уровень исследования сделался ведущим принципом важнейших биологических дисциплин. В биологии оформился новый раздел - цитология. Объектом изучения цитологии являются клетки многоклеточных организмов, а также организмы, тело которых представлено одной клеткой. Цитология изучает строение, химический состав, пути их размножения, адаптивные свойства. В данной работе будет рассмотрена структура и функции компонентов клетки.
Митоз - это непрямое деление клетки, в результате которого исходная клетка дает начало двум новым, имеющим совершенно одинаковый набор генов. Митотический цикл - совокупность процессов, в результате которых из одной клетки образуются две новые, он охватывает период митоза и часть интерфазы. Целью нашей работы по этому вопросу будет также анализ факторов, влияющих на митотическую активность.
Наследственность - это фундаментальное свойство всего живого, которое заключается в том, что все живые организмы способны хранить информацию о своем строении и передавать эту информацию другим поколениям. Человечество проделало долгий и нелегкий путь, чтобы правильно понять причины и законы наследственности. Целью нашей работы в аспекте наследственности будет рассмотрение критики представлений о фатальности наследственных заболеваний, что особенно актуально для сегодняшних медицинских исследований.
1.Структура и функции компонентов клетки
Теоретической базой цитологии является клеточная теория. Клеточная теория была сформулирована в 1838 году Т. Шванном, хотя первые два положения клеточной теории принадлежат М. Шлейдену, который занимался изучением клеток растений. Т. Шванн - известный специалист по строению клеток животных в 1838 году, опираясь на данные работ М. Шлейдена и результаты своих собственных исследований, сделал следующие выводы:
Клетка это наименьшая структурная единица живых организмов.
Клетки образуются в результате деятельности живых организмов.
Клетки животных и растений имеют больше сходств, чем различий.
Клетки многоклеточных организмов связаны между собой структурно и функционально.
Дальнейшее изучение строения и жизнедеятельности позволило узнать о ней много нового. Этому способствовало совершенство микроскопической техники, методов исследования и приход в цитологию многих талантливых исследователей. Было детально изучено строение ядра, проведен цитологический анализ таких важнейших биологических процессов как митоз, мейоз, оплодотворение. Стало известной микроструктура самой клетки. Были открыты и описаны органоиды клетки. Программа цитологических исследований 20 века поставила задачу выяснить и точнее разграничить свойства клетки. Отсюда особое внимание стало уделяться изучению химического состава клетки и механизма поглощения клеткой веществ окружающей средой.
Все эти исследования позволили умножить и расширить положения клеточной теории, основные постулаты которой в настоящее время выглядят следующим образом:
Клетка - основная и структурная единица всех живых организмов
Клетки образуются только из клеток в результате деления.
Клетки всех организмов сходны по своему строению, химическому составу, основным физиологическим функциям.
Клетки многоклеточных организмов образуют единый функциональный комплекс.
Клетки всех живых существ на земле можно поделить на два принципиально разных типа: ядерные (эукариотические) и безъядерные (прокариотические). Прокариотические клетки - самые древние на нашей планете, это клетки бактерий и синезеленых водорослей. Для них характерны следующие черты:
Отсутствие ядра.
Наличие ДНК кольцевого вида.
Многократное повторение одинаковых генов в ДНК.
Отсутствие самоделящихся органелл клетки: центриолей, митохондрий, пластид.
Деление клетки путем амитоза (прямого деления).
Из эукариотических клеток образованы организмы растений, грибов и животных. Они появились позднее прокариот. Для них характерны такие признаки как:
Наличие ядра, где всегда находятся молекулы ДНК. Некоторые клетки вторично утрачивают ядро (эритроциты млекопитающих и тромбоциты).
ДНК всегда в виде одной или нескольких нитей, незамкнутых на концах.
Гены в каждой молекуле ДНК, как правило, не повторяются.
В клетках всегда имеются самоделящиеся органеллы, обладающие собственными молекулами ДНК: центриоли, митохондрии, пластиды. Последние встречаются только в растительных клетках.
Деление клетки путем митоза (непрямого деления), в результате которого все гены равномерно распределяются между новыми клетками.
Эукариотические клетки в десятки и сотни раз крупнее прокариотических.
Рассмотрим более подробно строение эукариотической клетки.
Клетка имеет мембрану, цитоплазму и ядро.
Мембрана - органелла клетки, имеющая четырехслойное строение. Наружный и внутренний слои белковые. Между ними лежат два слоя из жироподобных веществ - липоидов. Один из концов молекулы липоида имеет хорошо выраженные гидрофобные свойства. В мембране все липоиды расположены так, что своими гидрофобными концами каждый слой сориентирован в противоположную сторону от другого. В разных местах клеточной мембраны встроены особые крупные молекулы белков, которые занимают всю ее толщину. Мембраны многих клеток снаружи покрываются дополнительными защитными оболочками, состоящими либо из углеводов (например, из целлюлозы в растительных клетках), либо из сложных веществ - глюкопротеидов (пелликула инфузорий и жгутиконосцев). Здоровье клетки, длительность ее жизни во многом зависят от состояния мембраны.
Полная проницаемость для воды. Мембрана всегда пропускает воду внутрь клетки или наружу, в зависимости от того, где концентрация воды больше. Такое движение вещества из области высокой его концентрации в область более низкой называется диффузией. Диффузия вещества не требует затрат энергии.
Избирательная проводимость растворенных веществ:
Отрицательно заряженные частицы быстрее и легче проникают через мембрану.
Вещества растворимые в жирах легче проникают через мембрану, чем вещества растворимые в воде.
Мелкие молекулы легче проникают через мембрану, чем крупные.
Активный транспорт веществ. Некоторые вещества способны проникать через мембрану в направлении обратном их диффузии, то есть из места низкой в место с более высокой концентрацией. Путем активного транспорта из клетки постоянно выводится избыток ионов натрия, водорода и хлора. А фосфаты, глюкоза, аминокислоты, наоборот активно проникают в цитоплазму. Активный транспорт всегда сопряжен с затратой энергии.
Мембрана регулярно восстанавливается в результате работы специальных органелл, синтезирующих мембранные вакуоли. Многие мембраны, не покрытые плотными оболочками, способны образовывать временные выросты, называемые ложноножками (псевдоподиями).
Функции мембран:
Фагоцитоз - захват ложноножками твердых частичек пищи. В результате образуется пищеварительная вакуоль, плавающая в цитоплазме.
Пиноцитоз - поглощение растворенных веществ.
Защитная. Мембрана защищает клетку от проникновения в нее чужеродных, опасных веществ.
Дыхательная. Через мембрану в клетку поступает кислород, а выделяется углекислый газ.
Гомеостатическая. Гомеостаз - это способность поддерживать относительно постоянным свой состав. Благодаря своим свойствам (избирательному поглощению веществ и активному транспорту) мембрана обеспечивает клетке постоянство своего состава.
Интегративная. Клетки контактируют между собой при помощи мембран. Через мембрану одна клетка может передавать различную информацию другой клетке. Эта информация может передаваться как при помощи электрических импульсов, так и при помощи химических веществ (гормонов, медиаторов).
Цитоплазма - клеточный сок, клеточная жидкость. Содержит воду, растворенные в ней неорганические и органические вещества, а также различные обособленные структуры, называемые органеллами:
Рибосомы - органеллы клетки, состоящие из двух частиц крупной и мелкой. Каждая частица образована белками и рибосомальной РНК. Рибосомы осуществляют синтез белка. Синтезируются в ядре.
Эндоплазматическая сеть (ЭПС) - мембранная органелла клетки, представляющая многочисленные каналы и полости из мембран, по структуре сходной с мембраной клетки. По строению и функциям делится на два типа: шероховатая ЭПС - содержит на поверхности рибосомы и является местом синтеза белков; гладкая ЭПС - не содержит рибосом, является местом синтеза углеводов, липоидов и жиров. Снаружи ЭПС контактирует с мембраной клетки, внутри - с мембраной ядра.
Аппарат Гольджи - по расположению является участком эндоплазматической сети. Имеет мембранную структуру. Выглядит как скопление многочисленных мешочков, полостей, вакуолей. Выполняет множество функций:
Доводит белки до окончательной рабочей формы, некоторые белки в крупные белковые комплексы, присоединяет к некоторым белкам необходимые ионы металлов.
Образует мембранные пузырьки, которые, покидая комплекс Гольджи либо реставрируют клеточную мембрану, либо превращаются в лизосомы.
Лизосомы - мембранные органеллы клетки, представляющие микроскопические пузырьки, наполненные пищеварительными ферментами. Выполняют пищеварительную и защитную функции. Могут слипаться с пищеварительной вакуолью, изливая в нее пищеварительные ферменты. При контакте клетки с чужеродным веществом или с чужой клеткой лизосомы слипаются с клеточной мембраной, выделяя свои ферменты во вне клетки. Ферменты лизосом могут также принимать участие в запрограммированной смерти собственной клетки.
Митохондрии - мембранные самоделящиеся органеллы. Образованы двумя слоями мембран: наружной гладкой и внутренней, имеющей многочисленные выросты внутрь митохондрии. Такие выросты внутренней мембраны называются кристами. В них протекает процесс окисления молочной кислоты, в результате которого выделяется энергия, запасаемая в виде АТФ (окислительное фосфорилирование). Следовательно, важнейшая функция митохондрий - энергетическая. Митохондрии имеют собственные молекулы ДНК, которые по строению не отличаются от ДНК бактерий. Размножаются митохондрии, как и бактерии, прямым делением.
Пластиды - мембранные самоделящиеся органеллы клеток. В отличие от всех органелл, рассмотренных выше, пластиды встречаются только в растительных клетках. По строению напоминают митохондрии: образованы двумя мембранами наружной гладкой и внутренней, образующей многочисленные плоские выросты - тилакоиды. Все тилакоиды расположены стопками наподобие стопок монет. Каждая стопка называется граной. Между гранами находится внутренняя жидкость пластида называемая стромой. В ней находится собственная ДНК, строением напоминающая бактериальную. Размножаются пластиды подобно бактериям прямым делением.
Центриоли - самоделящиеся органеллы клеток животных и некоторых низших растений. Каждая центриоль состоит из короткого полого цилиндра, стенки которого образованы микротрубочками, расположенными вдоль оси цилиндра. Центриоли содержат белки и небольшое количество РНК. В клетке имеется две пары центриолей.
2.Митотический цикл. Факторы, влияющие на митотическую активность
Митотический цикл, совокупность процессов, в результате которых из одной клетки образуются две новые. Митотический цикл охватывает период митоза и часть интерфазы. -- периода между делениями, когда происходит подготовка к следующему митозу. Митотический цикл -- часть жизненного цикла клетки; в быстро делящихся клеточных популяциях (например, у бластомеров дробящегося яйца) митотический цикл почти совпадает с жизненным циклом клетки.
Митоз - это непрямое деление клетки, в результате которого исходная клетка дает начало двум новым, имеющим совершенно одинаковый набор генов.
Митоз длится 1-2 часа и протекает в четыре фазы, из которых самыми продолжительными являются первая и последняя.
Фазы митоза.
Профаза. Наблюдается конденсация хроматиновых нитей, то есть их упаковка. Образуются хорошо заметные в световой микроскоп (при специальном подкрашивании) утолщенные хромосомы. Синтез РНК и белков заканчивается. Разрушается оболочка ядра. Образуется веретено деления.
Метафаза. Все хромосомы перемещаются в центр клетки, располагаясь по ее экватору. Каждая хромосома состоит из двух хорошо различимых дочерних хроматид, образованных дочерними ДНК, появившихся в результате редупликации материнской. Любая пара дочерних хроматид связывается между собой тонким перехватом, называемым центромерой. Это участок материнской ДНК, в котором редупликация еще не прошла. К каждой центромере присоединена своя нить веретена деления.
Анафаза. Дочерние хроматиды отделяются друг от друга в результате редупликации центромер и быстро расходятся к противоположным полюсам клетки. Теперь на каждом полюсе находится свой набор хроматид. Оба этих набора содержат одинаковые гены, так как все дочерние хроматиды, образованные в ходе редупликации материнской ДНК являются копиями друг друга.
Телофаза. На полюсах клетки хроматиды раскручиваются в хроматиновые нити. Возобновляется синтез РНК и белков. Вокруг каждого набора из дочерних хроматид формируются свои ядерные оболочки. Клетка перешнуровывается по экватору. Образуются две новые клетки.
В результате митотического деления появляются две клетки генетически абсолютно одинаковые. Это возможно только благодаря двум процессам:
редупликации ДНК, в основе которой лежит принцип комплиментарности.
расхождении каждой пары дочерних хроматид в новые клетки.
Митотическое деление клеток встречается:
при бесполом размножении растений грибов и животных,
при эмбриональном и постэмбриональном развитии всех многоклеточных организмов из оплодотворенной яйцеклетки,
при заживлении ран, образовании клеток крови, нарастании клеток кожного и кишечного эпителия и др. процессах.
В результате облучения очень большого количества однотипных клеток установлено, что при воздействии разных видов излучений длительность обратимого угнетения клеточного деления и процент клеток, у которых деление полностью прекратилось, возрастают по мере увеличения дозы излучения. С увеличением дозы излучений все большее число клеток теряет способность к размножению или по крайней мере у них временно прекращается процесс деления. Одним из показателей нарушения этой способности клеток к размножению как у одноклеточных, так и у клеток тканей высших организмов является возникновение гигантских форм клеток.
Некоторые радиационно-биохимические изменения появляются уже после воздействия относительно малых доз, другие изменения наступают лишь в результате воздействия средних или высоких доз излучений. Среди нарушений обмена веществ, возникающих при воздействии ионизирующих излучений, на первое место следует поставить нарушение самого радиочувствительного субстрата--нуклеиновых кислот. Лучевые поражения в виде угнетения синтеза нуклеиновых кислот нельзя рассматривать как непосредственную причину угнетения клеточного деления или разрыва хромосом, которые могут привести к их грубым морфологическим нарушениям, определяемым при митозах после облучения. Нарушения других видов обмена, например углеводного, дают право говорить об его очень низкой радио чувствительности. Изменения углеводного обмена после облучения, в частности угнетение анаэробного гликолиза, становятся заметными, как правило, лишь после воздействия в дозах порядка 5000--20000 р.; нарушение клеточного дыхания обычно наблюдается в результате воздействия еще больших доз--от 20000 до 100000 р.
При воздействии малых доз излучении наблюдается угнетение клеточного деления. При больших дозах клетки окончательно теряют способность к размножению. Временное угнетение митозов и полная стерильность не могут быть обусловлены единым механизмом, несмотря на то, что оба эти явления на первый взгляд могут показаться вполне родственными.
От качества излучений, кроме функциональных изменений, зависят также определенные виды лучевых хромосомных аберраций. В клеточных популяциях с митотическим делением клеток после облучения сначала отмечается кратковременное увеличение частоты митозов, а затем падение до определенной минимальной величины.
Для первичного и вторичного эффекта излучений характерны определенные типы хромосомных изменений.
Механизм хромосомных изменений при первичном и вторичном эффекте различен. Хромосомные изменения, типичные для первичного эффекта, возникают главным образом в тех клетках, которые во время облучения имели митотическую активность и находились в стадии метафаза. У определенного числа этих клеток наблюдаются митозы, частота которых снижается в результате облучения. У других митотически делящихся клеток, достигших или прошедших стадию метафазы, митозы продолжаются, но в более замедленном темпе.
С делением клеток (митозом) связано определённое излучение, которое обнаружил и измерял А.Г. Гурвич. Он назвал его “митогенетическим”. Было установлено, что если под это излучение попадают другие клетки, то и их митоз увеличивается, то есть стимулируется их рост.
3.Критика представлений о фатальности наследственных заболеваний
Еще недавно даже среди врачей господствовало мнение о фатальности наследственных заболеваний, невозможности их профилактики и лечения. Сегодня для некоторых заболеваний методы лечения уже найдены. Фенилкетонурия отмечается в среднем у каждого из 10 тыс. новорожденных. В результате отсутствия в организме фермента, контролирующего превращение аминокислоты фенилаланина в тирозин, концентрация фенилаланина возрастает в десятки раз. Часть его выводится с мочой, а остальное количество превращается в фенилпировиноградную, фенилуксусную, фенилмолочную кислоты и другие вещества. Это вызывает ряд вторичных биохимических изменений, в результате которых нарушается созревание мозга. Отклонения в психическом развитии ребенка становятся заметными лишь после 6-месячного возраста. Большинство детей, страдающих фенилкетонурией, вырастали умственно неполноценными. Сейчас такое нарушение обмена устраняется безбелковой диетой, на которой ребенок находится до 6-8 лет. Белковые вещества дают только в виде специальных препаратов, из которых удален фенилаланин. Диагностика заболевания довольно проста: она основана на положительной качественной реакции мочи с фенилпировиноградной кислотой.
Другое наследственное заболевание - галактоземия - нередко проявляется уже в первые дни жизни ребенка рвотой, резкой вялостью, гипотонией, желтухой, судорогами. Если заболевание развивается постепенно, то основные симптомы выявляются несколько позднее. К ним относятся катаракта, умственная отсталость и хроническое поражение печени - гепатит. У больных повышено содержание галактозы (молочного сахара), а уровень глюкозы снижен. Если диагноз поставлен своевременно и из рациона ребенка исключены молочные продукты, развитие ребенка осуществляется нормально.
Мы привели только два примера. Наследственных заболеваний значительно больше. По данным Всемирной организации здравоохранения, около 4% новорожденных страдают теми или иными генетическими дефектами. А ведь к ним надо прибавить тех детей, у которых заболевание проявляется не сразу после рождения, а в более позднем возрасте. Поэтому дальнейшее развитие медицинской генетики, распространение генетических знаний не только среди врачей, но и среди населения является важной задачей. Не божественное предопределение, а реальные причины лежат в основе каждого вида наследственной патологии. Борьба с этими заболеваниями осуществляется двумя путями. Первый путь - целенаправленное изменение условий среды, делающее невозможным развитие болезни. Второй - профилактика с помощью медико-генетического консультирования населения.
Наследственные нарушения обмена веществ корректируются специальной диетой: устранением из пищи неусваиваемых организмом веществ или, напротив, введением недостающих. Многие дефекты органов речи, слуха, зрения исправляют хирургическим путем. Не зная истинных причин появления на свет детей с врожденными уродствами, люди нередко расценивали это как "божью кару за грехи" или предвестник тяжелых напастей. Дж. У. Баллантайн в книге "Тератологические записи халдеев" (1894) приводит примеры различных толкований и предсказаний, связанных с рождением уродов: "Если какая-нибудь женщина родит ребенка, у которого отсутствуют ноздри, стране будет угрожать несчастье, и дом ее мужа будет разрушен. Если какая-нибудь женщина родит ребенка, у которого нет носа, беда постигнет страну, и хозяин дома умрет. Если какая-нибудь женщина родит ребенка, у которого нет полового члена, хозяин дома снимет богатый урожай с полей. Если женщина родит ребенка, пол которого не будет ясно обозначен, бедствия и несчастия постигнут страну, а мужа ее будут сопровождать несчастья".
В происхождении неврозов и реактивных психозов основную роль играют психические травмы, которые иногда только провоцируют наследственную предрасположенность к болезни. В происхождении психических заболеваний определенную роль играет комбинация причинных факторов с индивидуальными особенностями человека. Например, не у всех лиц, страдающих сифилисом, развивается сифилитический психоз, и только у небольшого числа больных атеросклерозом сосудов головного мозга возникает слабоумие или галлюцинаторно-бредовой психоз.
Сегодня многие наследственно обусловленные болезни будущего ребенка диагностируются в начале беременности с помощью исследования амниотической околоплодной жидкости. Это позволяет начать своевременное лечение в тех случаях, когда оно возможно и целесообразно, или прервать беременность, чтобы предотвратить появление на свет неполноценного ребенка. Родители должны быть информированы о степени грозящего им несчастья для того, чтобы принять правильное решение.
Выводы
Таким образом, все организмы состоят из одинаковых частей -- клеток; они образуются и растут по одним и тем же законам. Общий принцип развития для элементарных частей организма -- образование клеток. Каждая клетка в определенных границах есть индивидуум, некое самостоятельное целое. Но эти индивдуумы действуют совместно, так, что возникает гармоничное целое ткань. Все ткани состоят из клеток. Процессы, происходящие в клетках растений, сводятся к следующим: возникновение новых клеток, увеличение размеров клеток, изменение клеточного содержимого и утолщение клеточной стенки.
Благодаря созданию клеточной теории стало понятно, что клетка -- это важнейшая составляющая часть всех живых организмов. Из клеток состоят ткани и органы. Развитие всегда начинается с одной клетки, и поэтому можно сказать, что она представляет собой предшественник многоклеточного организма. Структура и функции клеточных компонентов были рассмотрены в настоящей работе.
Как видим, способность к делению -- важнейшее свойство клеток. Без деления невозможно представить себе увеличение числа одноклеточных существ, развитие сложного многоклеточного организма из одной оплодотворенной яйцеклетки, возобновление клеток, тканей и даже органов, утраченных в процессе жизнедеятельности организма.
Деление клеток осуществляется поэтапно. На каждом этапе деления происходят определенные процессы. Они приводят к удвоению генетического материала (синтезу ДНК) и его распределению между дочерними клетками. Период жизни клетки от одного деления до следующего называется клеточным циклом. Во время митоза клетка проходит ряд последовательных фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, какой был в материнской клетке.
Различают 4 фазы митоза: профаза, метафаза, анафаза и телофаза. Среди факторов, влияющих на митотическую активность особое внимание нами уделено радиационно-биохимическим влияниям.
Мнение о фатальности наследственных заболеваний, невозможности их профилактики и лечения, господствовавшее даже среди врачей до недавнего времени, все больше подвергается критике. И сегодня для некоторых заболеваний методы лечения уже найдены.
К тому же, настоящее время многие наследственно обусловленные болезни будущего ребенка можно диагностировать в начале беременности с помощью исследования амниотической околоплодной жидкости, что позволяет начать своевременное лечение в случае его целесообразности или прервать беременность во избежание появления на свет неполноценного ребенка.
Литература
1.Адамчик М.В.- Всемирная энциклопедия. Биология / Ред. Аникеев В.И. - М.: Современный литератор, 2006.
2.Калюжный В.Г. Справочник по биологии. (Серия "Учебники и учебные пособия") - Ростов-на-Дону, Феникс, 2004.
3.Гиляров М.С. Биология. Большой энциклопедический словарь. - 3-е изд. - (Золотой фонд). - М.: Большая Российская энциклопедия, 2005.
4.В. Смелова. Строение клетки. Газета "Биология". - №36/2001.
5.П. Кошель. Учение о растительной клетке.- http://bio.1september.ru/article.php?ID=200204304
6.И. Бологова. Клетка - структурная и функциональная единица живого. - http://bio.1september.ru/article.php?ID=200200603
7. Грекова Т.И. Атеизм и медицина. - http://lib.metromir.ru/author5836
8. Н.Грін, У.Стаут, Д.Тейлор. Биология. М.: Мир, 1996.
Подобные документы
История и основные этапы исследования клетки, ее структуры и компонентов. Содержание и значение клеточной теории, выдающиеся ученые, внесшие свой вклад в ее разработку. Симбиотическая теория (хлоропласты и митохондрии). Зарождения эукариотической клетки.
презентация , добавлен 20.04.2016
Методы изучения клетки, их зависимость от типа объектива микроскопа. Положения клеточной теории. Клетки животного и растительного происхождения. Фагоцитоз - поглощение клеткой из окружающей среды плотных частиц. Подходы к лечению наследственных болезней.
презентация , добавлен 12.09.2014
Ядро эукариотической клетки. Клетки, имеющие более двух наборов хромосом. Процесс деления у эукариот. Объединенные пары гомологичных хромосом. Онтогенез растительной клетки. Процесс разъединения клеток в результате разрушения срединной пластинки.
реферат , добавлен 28.01.2011
Периоды и фазы клеточного цикла. Последовательное прохождение клеткой периодов цикла без пропуска или возврата к предыдущим стадиям. Деление исходной клетки на две дочерние клетки. Циклины и циклин-зависимые киназы; деление эукариотической клетки; митоз.
контрольная работа , добавлен 21.11.2009
Авторы создания клеточной теории. Особенности архей и цианобактерий. Филогения живых организмов. Строение эукариотической клетки. Подвижность и текучесть мембран. Функции аппарата Гольджи. Симбиотическая теория происхождения полуавтономных органелл.
презентация , добавлен 14.04.2014
Элементы строения клетки и их характеристика. Функции мембраны, ядра, цитоплазмы, клеточного центра, рибосомы, эндоплазматической сети, комплекса Гольджи, лизосом, митохондрий и пластид. Отличия в строении клетки представителей разных царств организмов.
презентация , добавлен 26.11.2013
Виды повреждения клетки. Стадии хронического повреждения клетки. Виды гибели клетки. Некроз и апоптоз. Патогенез повреждения клеточных мембран. Высокоспециализированные клетки с высоким уровнем внутриклеточной регенерации. Состояния соединительной ткани.
презентация , добавлен 03.11.2013
Элементарная генетическая и структурно-функциональная биологическая система. Клеточная теория. Типы клеточной организации. Особенности строения прокариотической клетки. Принципы организации эукариотической клетки. Наследственный аппарат клеток.
контрольная работа , добавлен 22.12.2014
Строение животной клетки. Основные положения клеточной теории, понятие про прокариоты и эукариоты. Структура цитоплазмы и эндоплазматический ретикулум. Хромосомный набор человека. Способы деления клетки (амитоз, митоз и мейоз) и ее химический состав.
презентация , добавлен 09.10.2013
Компартментация в организации эукариотической клетки. Линейные размеры эукариотической клетки. Ядерно-цитоплазматическое соотношение. Различные формы хондриома. Митохондриальная система кардиомиоцитов. Признаки митохондриальных болезней у человека.
Клетка. Строение и взаимодействие клеточных элементов. Элементы клеточной патологии.
(В составлении лекции принимали участие проф. Семёнов В.В., асс. Кошпаева Е.С., асс. Колочкова Е.В.)
РАССМАТРИВАЕМЫЕ ВОПРОСЫ.
1. ВВЕДЕНИЕ
2. Структурные компоненты клетки.
2.1. Плазматическая мембрана и её роль в жизнедеятельности клетки.
2.1.1. Строение плазматической мембраны.
2.1.2. Функции плазматической мембраны.
а. Транспортная функция.
б. Перенос информации через плазматическую мембрану.
2.1.3. Участие компонентов мембран (липидов, белков и гликопротеинов) в патологических процессах.
а. Перекисное окисление липидов.
б. Фосфолипазная активность.
в. Гликопротеины и опухолевой процесс
2.2. Эндоплазматическая сеть.
2.3. Комплекс Гольджи.
2.4. Лизосомы.
2.5. Рибосомы.
2.6. Митохондрии.
2.7. Клеточный центр.
2.8. Ядро-система управления клеткой.
2.8.1. Ядерная оболочка.
2.8.2. Нуклеоплазма (кариолимфа, ядерный сок).
2.8.3. Морфо-функциональная характеристика и классификация хромосом.
а. Хроматин – эухроматин и гетерохроматин.
б. Нуклеосома.
2.8.4. Ядрышко.
2.8.5. Ядерный матрикс.
2.9. Цитоплазматические включения.
а. Трофические включения.
б. Секреторные.
в. Пигментные.
г. Экскреторные.
В этой лекции мы отойдём от традиционного изложения морфо-функциональных (структуры и функции) особенностей клетки, которые были характерны для школьной методики преподавания. Основной упор мы сделаем на функциональную сторону поведения клетки и её элементов. При этом мы исходим из трёх важных в медицинской биологии принципов. Во-первых, клетка, как целостная система находится не во внешней природной среде, а внутри целостного организма, в своеобразной жидкой межклеточной среде . И это не случайно. Эволюция живого началась в водной среде. Практически все основные морфологические образования клетки эволюционировали в водной среде. К ней они адаптированы. После перехода на сушу, в газовую среду, необходимо было существенно менять всю приспособленную к водной среде организацию живого. Сейчас трудно предположить в силу каких причин эволюция в газовой среде у вышедших на сушу организмов не смогла полностью исключить все приспособления, которые организмы получили находясь в жидкой среде. Возможно, эволюция в газовой среде потребовала таких коренных перестроек в живых системах, адаптированных к водной среде, что они были просто не совместимы с жизнью. По другому варианту эволюция организмов, приспособленных к водной среде была невозможна в газовой. То-ли были какие-то ещё причины, но в любом случае Природа пошла на компромисс – вышедшие на сушу организмы заключили в себе среду первичного океана, к которому они адаптированы. Фактически кровь, лимфа, межклеточная жидкость по своему составу напоминает колыбель нашего развития, первичный океан в котором проходил первый этап нашей эволюции. Клетка, находясь внутри организма, практически не соприкасается с внешней средой; вся её деятельность проходит в межклеточной жидкости, которая обеспечивает не только существование клетки, но и является инициатором перестройки её метаболизма. Эта перестройка переводит клетку на новый режим жизнедеятельности, в иное функциональное состоянии. Однако такой переход возможен в результате второго важного принципа – строение и функции субклеточных органоидов клетки не строго детерминированы, они пластичны, способны изменяться в определённых пределах. А поскольку внутриклеточные элементы участвуют в различных биохимических процессах, любые изменения структуры клетки непременно приведут к изменению выполняемых этой структурой функций. Для живого характерным является то, что один и тот же клеточный элемент может выполнять несколько функций. Например, это хорошо иллюстрируется ниже при перечислении функций, например, органоидов. И, наконец, необходимо помнить третий принцип:- все внутриклеточные элементы и процессы представляют собой единую взаимосвязанную систему, эту совокупность элементов и процессов можно представить как своеобразную сеть, в которой изменение одной ячейки или узла приводит к изменению всей внутриклеточной организации и её функции. Этот принцип имеет большое значение в медицине, поскольку иногда возникающие изменения настолько сильны, что слаженная внутренняя организация клетки нарушается. В этом случае процессы ответственные за саморегуляцию, адаптацию и другие клеточные функции могут стать ареной для развития патологии, вначале на уровне внутриклеточных элементарных структур и процессов, затем на уровнепатологии всей клетки, как элементарной саморегулирующейся живой системы, а затем на уровне клеточных образований, объединённых конечной функцией.
2. Структурные компоненты эукариотической клетки
На внешней стороне клетки (рис. 1) располагается наружная плазматическая мембрана, которая отделяет клетку от внешней среды. Под ней располагаются цитоплазма и ядро. Ядро не всегда находится в центре клетки. В тех случаях, когда в одной части клетки осуществляется интенсивная работа, например. идёт активный процесс всасывания пищевых веществ с затратой энергии, ядро сдвинуто к противоположному «нерабочему» отделу клетки, а в «рабочем» отделе концентрируются митохондрии. Цитоплазма и ядро, в свою очередь, состоят из нескольких компонентов, которые представлены на рисунке 1.
В основе структурной организации клетки лежит мембранный принцип. Это означает, что мембраны составляют существенную часть в строении клетки. Они отделяют клетку от окружающей межклеточной среды, делят клетку на отдельные отсеки – компартменты. В этих изолированных участках размещены специфические метаболические процессы. Такое разграничение в эволюции повысило организацию клетки, но этим не закончилось. С течением времени некоторые биохимические процессы в компартментах стали настолько специфическими, что появилась необходимость в создании более изолированных, почти автономных структур - органоидов. К органоидам, имеющим мембрану, относятся митохондрии, пластиды, лизосомы, эндоплазматический ретикулум и комплекс Гольджи. На этих плацдармах разместились жизненно важные для организма процессы. Одновременно создавались структуры, не имеющие мембранной оболочки, но так же выполняющих определённые клеточные функции – рибосомы, клеточный центр и др. В то же время цитоплазма клетки остаётся изолированной мембраной эндоплазматической сети, хотя и не полностью, на отдельные отсеки, в которых так же локализуются определённые звенья клеточного метаболизма, например, гликолиз. Все биомембраны построены примерно одинаково, поэтому рассмотрим строение только плазматической мембраны.

http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image004.gif" border="0" width="104" height="55 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image006.gif" border="0" alt="Подпись:" width="126" height="64 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image008.gif" border="0" width="204" height="48 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image010.gif" border="0" width="12" height="87 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image012.gif" border="0" width="12" height="99 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image014.gif" border="0" width="12" height="13 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image016.gif" border="0" width="12" height="81 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image011.gif" border="0" width="12" height="27 src=">
Липиды Ядерный сок
Белки Гиалоплазма
Органоиды Хромосомы
Полисахариды Включения
Ядерная мембрана
Рис. 1. Общий план строения эукариотической клетки.
2.1. Плазматическая мембрана, её роль в жизнедеятельности клетки
2.1.1. Строение плазматической мембраны.
Мембрана состоит из двух слоев липидных молекул (бислой), в который встроены белки. К некоторым липидным и белковым молекулам присоединены углеводы. Их немного. Толщина мембраны около 10 нм (0,00001 мм). Основную часть мембраны составляет непрерывный слой фосфолипидных молекул. В этот жидкий слой погружены молекулы различных по строению и функций белковых молекул. Белки не полностью покрывают липидный бислой, а располагаются в нём по отдельности или группами. В целом это напоминает мозаику (рис. 2. Б. В). В связи с этим принятая в настоящее время модель мембраны носит название жидкостно-мозаичная . Белки способны перемещаться по липидному слою. Перемещаются и молекулы липидного слоя. Понятно, что движение молекул мембраны меняет физико-химические характеристики последней, а это, в свою очередь, меняет функциональные характеристики мембраны. Необходимо отметить, что плазматическая мембрана большинства клеток не имеет форму идеального шара. Наоборот, она имеет множество выступов, углублений, которые постоянно меняют свою форму и величину. Полученные в последнее время результаты внесли некоторые коррективы в теорию строение мембран. Было показано, что не все мембранные белки способны к движению, а некоторые участки мембран отличаются по своей структуре от классического липидного бислоя.
Молекула фосфолипида имеет вид головки с двумя хвостиками (рис. 2 А). Головка (глицерин) растворима в воде, гидрофильна, хвостики (жирные кислоты) нерастворимы в воде, гидрофобны. Поэтому, находясь в воде, молекулы самопроизвольно занимают определённое положение по отношению к водной фазе. Отталкиваясь от молекул воды, хвостики располагаются в глубине липидного слоя, а водорастворимые головки обращены к внешней и внутренней водной среде (рис. 2. Б). Бислой липидов носит название матрикс. Особо следует отметить наличие в мембранах клеток липидов, хвостик которых содержит жирные кислоты, имеющие в своей структуре двойные связи, расположенные через CH2-группу (– СН = СН – СН = СН – СН –) . Такие жирные кислоты называют ненасыщенными. Эти кислоты наиболее всего подвержены воздействию активных форм кислорода (АФК), которые постоянно присутствуют в организме всех живых существ. Их количество особенно возрастает при различных заболеваниях. Это может привести к неблагоприятным последствиям, о чём мы скажем ниже.
Вкрапленные в матрикс белки (рис. 2 В) располагаются по-разному. Одни на внутренней и наружной поверхности липидного слоя – примембранные или поверхностные белки, другие полупогружены в мембрану – полуинтегральные белки, третьи пронизывают всю мембрану – интегральные белки. Обычно полуинтегральные и интегральные белки объединяют одним термином – внутренние белки, поскольку их трудно отличить друг от друга. Чаще всего в мембранах встречаются интегральные белки. Они могут быть представлены, как одной молекулой и выполнять какую-либо одну функцию, так и группой (ансамблем) белков. Каждый участник ансамбля выполняет строго определённую роль. Эти комплексы также выполняют одну или несколько конечных функций.
А Б ВMsoNormalTable" style="border-collapse: collapse; border: none; margin-left: 6.75pt; margin-right: 6.75pt;" align="left ">
Фосфолипид
1-головка,
2- хвостик
Липидный бислой
(матрикс)
Расположение молекул белка относительно липидного слоя: 1- примембранные белки, 2 – полупогружённый белок, 3 - интегральный белок
Рис. 2. Схематическое строение плазматической мембраны.
Следует отметить, что интегральные и поверхностные белки-ферменты, функционирующие в мембране, достаточно часто меняют своё положение. В некоторых случаях сложно определить к какому типу (поверхностные или интегральные) относится тот или иной белок мембраны. Например, фермент фосфолипаза А, осаждаясь на мембрану, является поверхностным белком, но затем она активируется, становится интегральным белком и, взаимодействуя с липидами бислоя, образует из них арахидоновую кислоту (рис.3). Последняя покидает мембрану и превращается в другие активные соединения, которые участвуют в развитии различных патологических процессов.
Na+, K+- АТФ-аза Адренорецептор Аденилатциклаза
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image020.gif" border="0" width="12" height="79 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image022.jpg" border="0" width="576" height="193 src="> Ca2+-АТФ-аза
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image030.gif" border="0" width="12" height="39 src="> цАМФ
Na+, K+ Сa2+ Арахидоновая
кислота G-белок Гликоген
Рис. 3. Гипотетическая схема локализации некоторых мембранных белков
Напротив, белки, участвующие в перемещении веществ через мембрану –например, белки, принимающие участие в облегчённой диффузии Na+, K+- АТФ-аза или Ca2+-АТФ-аза, как правило, не меняют своё положение, функционируя как интегральные белки (рис. 3). И, наконец, как мы уже говорили, в мембране могут находиться сложные комплексы из нескольких белков, связанных в единый ансамбль одной задачей. К таким комплексам относятся белки, принимающие участие в проведении информационного сигнала через мембрану (рис.3). К последним относится комплекс, содержащий три белка - адренорецептор, G-белок и аденилатциклазу. Все эти белки имеют существенное значение в нормальной жизнедеятельности клетки и при патологии. Об этом мы расскажем ниже.
Кроме липидов и белков в мембране имеются углеводы, но они располагаются не как самостоятельные компоненты, а как составные части липидов (гликолипиды) или белков (гликопротеинов). Углеводы располагаются на наружной поверхности плазмалеммы.
2.1.2. Функции плазматической мембраны.
Плазматическая мембрана выполняет множество функций. Перечислим наиболее важные.
· Перенос веществ через мембрану. Через мембрану осуществляется транспорт веществ в обе стороны мембраны.
· Перенос информации через мембрану. На мембране информация из вне воспринимается преобразуется и передаётся в клетку или из клетки. Существенную роль при это м играют рецепторы мембран.
· Защитная роль. а) защищает содержимое клетки от механических повреждений, химических реагентов и биологической агрессии, например от проникновения вирусов и др.;
б) в многоклеточном организме рецепторы плазматической мембраны формируют иммунный статус организма;
в) в многоклеточном организме мембрана обеспечивает протекание реакции фагоцитоза.
- Ферментативная - в мембранах находятся различные ферменты (например, фосфолипаза А и др.), которые осуществляют целый ряд ферментативных реакций. Гликопротеины и гликолипиды на цитоплазматической мембране осуществляют контакт с мембранами других клеток.
Некоторые из перечисленных функций рассмотрим более детально.
а. Транспортная функция. Через мембрану внутрь клетки и наружу происходит перемещение различных веществ, в том числе и лекарственных препаратов. В зависимости от размера переносимых через мембрану молекул различают два вида транспорта: без нарушения целостности мембраны и с нарушением целостности мембраны. Первый тип транспорта может осуществляется двумя путями – без затрата энергии (пассивный транспорт) и с затратой энергии (активный транспорт) (см. рис. 4). Пассивный перенос происходит за счёт диффузии по электрохимическому градиенту в результате броуновского движения атомов и молекул. Этот вид транспорта может осуществляться непосредственно через липидный слой, без какого-либо участия белков и углеводов или при помощи специальных белков – транслоказ. Через липидный слой в основном транспортируются молекулы веществ, которые растворимы в жирах, и малые незаряженные или слабозаряженные молекулы, такие как вода, кислород, углекислый газ, азот, мочевина, жирные кислоты, а также многие органические соединения (например, наркотики) хорошо растворимые в жирах . Транслоказы, могут переносить вещество через мембраны в сторону его меньшей концентрации, не затрачивая энергии, при помощи двух различных механизмов – через канал, который проходит внутри белка, или путём соединения выступающей из мембраны части белка с веществом, поворотом комплекса на 1800 и отсоединением вещества от белка. Диффузия веществ через мембрану с участием белков важна тем, что она идёт значительно быстрее простой диффузии, через липидный слой без участия белков. Поэтому диффузия, в которой принимают участие транслоказы, называют облегчённой диффузией. По такому принципу в клетку транспортируются некоторые ионы (например, ион хлора) и полярные молекулы, а также глюкоза.
Для активного переноса веществ через мембрану характерны три свойства:
1. Активный перенос осуществляется против градиента концентрации.
2. Осуществляется белком переносчиком.
3. Идёт с затратой энергии.
Энергия при активном переносе веществ необходима для того, чтобы перенести вещество против градиента его концентрации. Системы активного переноса часто называют мембранными насосами. Энергия в этих системах может быть получена из различных источников, чаще всего таким источником служит АТФ. Расщепление фосфатных связей в АТФ осуществляет интегральный белок-фермент АТФ-аза. Поэтому этот фермент и находится в мембране многих клеток в виде интегрального белка. Важно то, что этот фермент не только освобождает энергию из АТФ, но и осуществляет перемещение вещества. Поэтому система активного переноса состоит чаще всего из одного белка - АТФ-азы, который получает энергию и перемещает вещество. Иными словами, процесс перемещения и энергообеспечения в АТФ-азе сопряжены. В зависимости от того, какие вещества перекачивает АТФ-аза насосы называют или Na+, K+- АТФ-аза или Ca2+-АТФ-аза . Первые регулируют содержание в клетке натрия и калия, вторые кальция (этот тип насосов чаще всего размещён на каналах ЭПС). Сразу же отметим важный для медицинских работников факт: для успешной работы калий-натриевого насоса, клетка затрачивает около 30% энергии основного обмена. Это очень большой объём. Эта энергия тратится на поддержку определённых концентраций натрия и калия в клетке и межклеточном пространстве;- в клетке содержится калия больше, чем в межклеточном пространстве, натрия, наоборот, больше в межклеточном пространстве, чем в клетке. Такое распределение, далёкое от осмотического равновесия, обеспечивает наиболее оптимальный режим работы клетки.
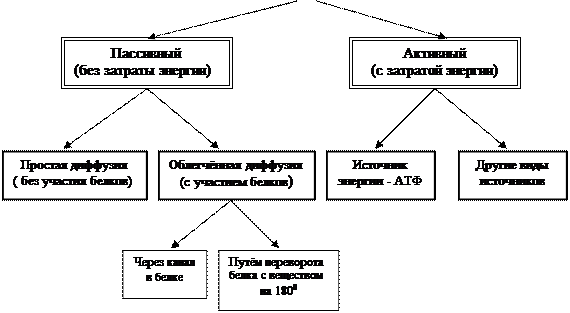 |
Рис. 4. Классификация типов транспорта веществ через мембрану.
Путём активного переноса происходит перемещение через мембрану неорганических ионов, аминокислот и сахаров, практически всех лекарственных веществ, имеющих полярные молекулы – парааминобензойная кислота, сульфаниламиды, йод, сердечные гликозиды, витамины группы В, кортикостероидные гормоны и др.
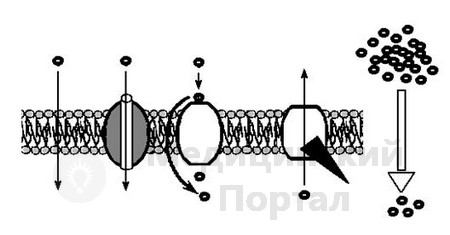
Для наглядной иллюстрации процесса переноса веществ через мембрану мы приводим (с небольшими изменениями) рисунок 5 взятый из книги «Молекулярная биология клетки» (1983) Б. Альбертса и др. учёных, считающихся лидерами в разработке теории
 http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image036.gif" border="0" width="38" height="19 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image038.gif" border="0" width="33" height="16 src="> белок переносчик
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image036.gif" border="0" width="38" height="19 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image038.gif" border="0" width="33" height="16 src="> белок переносчик
Пассивный транспорт Активный транспорт
Рис 5. Многие мелкие незаряженные молекулы свободно проходят через липидный бислой. Заряженные молекулы, крупные незаряженные молекулы и некоторые мелкие незаряженные молекулы проходят через мембраны по каналам или порам либо с помощью специфических белков переносчиков. Пассивный транспорт всегда направлен против электрохимического градиента в сторону установления равновесия. Активный же транспорт осуществляется против электрохимического градиента и требует энергетических затрат.
трансмембранного переноса, отражены основные типы переноса веществ через мембрану. Следует отметить что белки, участвующие в трансмембранном переносе, относятся к интегральным белкам и чаще всего представлены одним сложноорганизованным белком.
Перенос высокомолекулярных молекул белка и др. больших молекул через мембрану в клетку осуществляется эндоцитозом (пиноцитоз, фагоцитоз и эндоцитоз), а из клетки – экзоцитозом. Во всех случаях эти процессы отличаются от вышеизложенных тем, что переносимое вещество (частица, вода, микроорганизмы или др.) вначале упаковывается в мембрану и в таком виде переносится в клетку или выделяется из клетки. Процесс упаковки может происходить как на поверхности плазматической мембраны, так и внутри клетки
б. Перенос информации через плазматическую мембрану.
Кроме белков, участвующих в переносе веществ через мембрану, в ней выявлены сложные комплексы из нескольких белков. Пространственно разделённые, они объединены одной конечной функцией. К сложно устроенным белковым ансамблям относится комплекс белков, отвечающих за производство в клетке очень мощного биологически активного вещества – цАМФ (циклический аденозинмонофосфат). В этом ансамбле белков имеются как поверхностные, так и интегральные белки. Например, на внутренней поверхности мембраны расположен поверхностный белок, который носит название G – белок. Этот белок поддерживает взаимоотношения между двумя рядом расположенными интегральным белками – белком, который называется адреналиновый рецептор и белком - ферментом – аденилатциклазой. Адренорецептор способен соединятся с адреналином, который попадает из крови в межклеточное пространство и возбуждаться. Это возбуждение G – белок передаёт на аденилатциклазу – фермент, способный производить активное вещество – цАМФ. Последний, поступает в цитоплазму клетки и активирует в ней самые различные ферменты. Например, активируется фермент, расщепляющий гликоген до глюкозы. Образование глюкозы приводит к повышению активности митохондрий и повышению синтеза АТФ, которая поступает в качестве носителя энергии во все клеточные отсеки, усиливая работу лизосомы, натрий-калиевых и кальциевых насосов мембраны, рибосом и т.д. повышая в конечном итоге жизнедеятельность практически всех органов, особенно мышц. На этом примере, хотя и очень упрощенном, видно как связана деятельность мембраны с работой других элементов клетки. На бытовом уровне эта сложная схема выглядит достаточно просто. Представьте, что на человека неожиданно набросилась собака. Возникшее чувство страха приводит к выбросу в кровь адреналина. Последний, связывается с адренорецепторами на плазматической мембране изменяя при этом химическую структуру рецептора. Это, в свою очередь, приводит к изменению структуры G – белка. Изменённый G – белок становиться способным активировать аденилатциклазу, которая усиливает производство цАМФ. Последний стимулирует образование глюкозы из гликогена. В результате усиливается синтез энергоёмкой молекулы АТФ. Повышенное образование энергии у человека в мышцах приводит к быстрой и сильной реакции на нападение собаки (бегство, защита, борьба и т.д.).
2.1.3. Участие компонентов мембран (липидов, белков и гликопротеинов) в патологических процессах.
Прежде всего следует отметить, что практически при любом воздействии неблагоприятных факторов на мембрану в ней возникают два процесса
1. Происходит активация перекисного окисления липидов.
2. Происходит активация фосфолипазной активности.
Первый процесс касается липидного слоя, второй белкового.
а. Перекисное окисление липидов.
Мы уже говорили, что основу фосфолипидной молекулы мембраны составляет глицерол и 2 жирные кислоты. Эти кислоты могут быть насыщенными и ненасыщенными. Ненасыщенные кислоты содержат в своей молекуле двойные связи между атомами углерода (с – С = С – С = С – С = С –). Отсутствие таких связей характеризует насыщенную жирную кислоту (– С – С – С – С – С –). Ненасыщенные жирные кислоты легко вступают во взаимодействие с активными формами кислорода (АФК), которые могут попасть в мембрану из клетки или межклеточного пространства. АФК являются сильными реагентами и реагируют практически со всеми известными органическими соединениями. (белками, эфирами, аминокислотами, ДНК, РНК и т.д.). АФК опасны тем, что их реакции неуправляемые, они реагируют со всеми без исключения органическими молекулами, с которыми встречаются. Эти реакции ведут к разрушению органических веществ, потери специфической активности. Внутри клетки АФК образуются в самых различных местах – в органоидах, цитоплазме, в количествах которые не представляют опасность для клетки. Это связано с тем, что в клетке имеется мощная система защиты от АФК – витамины, некоторые продукты метаболизма клетки, белки и др. соединения. Если клетка попадает в неблагоприятные условия (травмы, вирусные или инфекционные заболевания, аутоиммунные конфликты и др.), то генерация АФК внутри клетки возрастает и их уровень начинает превышать физиологические возможности защитного барьера. В этом случае АФК распространяются по всему объёму клетки. Попав в мембрану они прежде всего взаимодействуют с ненасыщенными жирными кислотами. Это приводит к образованию различных не свойственных мембране соединений. Некоторые из них обладают сильными реакционными свойствами, их называют свободные радикалы. Они чрезвычайно опасны поскольку способны вступать во взаимодействие практически с любыми органическими соединениями – белками, жирами, углеводами, ДНК, РНК и т.д. с которыми они контактируют. Находясь в мембране, свободные радикалы взаимодействуют со всеми органическими молекулами мембраны, нарушая её целостность. Свободные радикалы могут выходить из мембраны внутрь клетки – тогда они взаимодействуют с различными компонентами цитоплазмы, органоидов, ядра. Если же они попадают в межклеточное пространство, то они могут при помощи переносчиков попасть в кровь и разноситься по всему организму. Отметим, что свободные радикалы, как правило, являются сильными мутагенами, что представляет угрозу для генетического материала половых клеток (возможно появление наследственной патологии) и соматических (возможно преобразование соматической клетки в результате мутации в злокачественную). Есть и другая опасность. В результате взаимодействия свободных радикалов с ненасыщенными жирными кислотами могут образовываться водорастворимые соединения, которые могут стать причиной неконтролируемого проникновения через мембрану самых различных соединений, например ионов. Иными словами в сплошном бислое липидов образуется канал, по которому могут проходить в клетку и выходить из неё самые различные ионы, в частности – натрий и калий. Как правило, при физиологических условиях в клетке концентрация калия выше, чем в межклеточном пространстве, а концентрация натрия, наоборот, в клетке ниже, чем в межклеточном пространстве. Такое состояние необходимо клетке для нормальной жизнедеятельности и поддерживается за счёт постоянной работы специального калий-натриевого насоса расположенного в толще мембраны. Через него, строго дозировано проходят ионы натрия и калия. В случае появления неестественного канала, через него начнётся неуправляемое движение натрия и калия. Натрий проникнув в клетку будет соединяться с хлором, образуя соль. Внутри клетки повысится осмотическое давление и в клетку будет поступать вода. Клетка будет набухать и перекроет узкие капиллярные русла. Это приведёт к уменьшению снабжением кислородом и питательными веществами других клеток. В условиях недостатка кислорода и питания в клетках автоматически усилится генерация АФК и они вновь будут попадать из цитоплазмы в мембрану, где будут вновь взаимодействовать с ненасыщенными жирными кислотами. Как мы говорили ранее это приведёт к образованию свободных радикалов и появлению в мембране неестественных каналов. Это усилит переход натрия внутрь клетки и образование там NaCl, сразу же начнётся перемещение воды внутрь клетки, её разбухание и вслед за этим сужение ближайших капилляр. и В соседних клетках вновь возникнут дефицит кислорода и питательных веществ. Сценарий повторится. Мы коснулись только небольшого фрагмента событий, которые разыгрываются в мембране при патологии. Но и из него отчётливо виден важный принцип характерный для любой патологии – процесс возникший в одном месте клетки, по одной какой-то причине, затем распространяется, захватывая другие объекты, усиливается и в конце концов может привести к непоправимым последствиям. Понятно, как важно найти в этой цепочки развёртывающихся процессов слабое звено, воздействуя на которое можно было бы блокировать развитие патологии.
б. Фосфолипазная активность .
Этот процесс связан с активацией находящегося на мембране фермента – фосфолипазы А. Об этом ферменте мы уже писали выше - он преобразует некоторые липиды матрикса в арахидоновую кислоту. Последняя выходит из липидного бислоя в цитоплазму клетки и превращается в целый ряд активных соединений, часть из которых остаётся в клетке, изменяя её метаболизм, часть выходит из клетки в межклеточное пространство воздействуя на соседние клетки, переводя их на новый режим работы. Другая часть активных веществ попадает в кровь, разносится по организму и воздействует на отдалённые клетки, также модифицируя их функцию. Приведём пример (рис.6). В бронхах, вокруг кровеносных сосудов, располагается большое количество тучных клеток . Во время
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image045.gif" border="0" width="464" height="137 src="> Рибосома иРНК
Рис. 7. Эндоплазматическая сеть:
А – фрагменты гладкой ЭПС; Б – фрагменты шероховатой ЭПС. В – функционирующая рибосома на шероховатой ЭПС.
В мембране гладкой ЭПС имеется набор ферментов, синтезирующих жиры и простые углеводы, а также стероидные гормоны, необходимые для организма. Особо следует отметить, что в мембране гладкой ЭПС клеток печени располагается система ферментов, осуществляющих расщепление чужеродных веществ (ксенобиотиков), попавших в клетку, в том числе и лекарственных соединений. Система состоит из разнообразных белков-ферментов (окислителей, восстановителей, ацетиляторов и др.).
Ксенобиотик или лекарственное вещество (ЛВ), взаимодействуя последовательно с определёнными ферментами, изменяет свою химическую структуру. В результате конечный продукт может сохранить свою специфическую активность, может стать неактивным или, наоборот, приобрести новое свойство – стать токсичным для организма. Система ферментов, расположенная в ЭПС и осуществляющая химическое преобразование ксенобиотиков (или ЛВ), носит название система биотрансформации. В настоящее время этой системе придают большое значение, т.к. от интенсивности её работы и количественного содержания в ней тех или иных ферментов зависит специфическая активность ЛВ (бактерицидность и т.п.) в организме и их токсичность. http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image047.gif" border="0" width="216" height="105 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image049.jpg" border="0" width="287" height="252 src="> Элементы цитоскелета
 |
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image052.gif" border="0" width="107" height="50 src="> Рибосома
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image054.gif" border="0" width="31" height="53 src=">
Ядро Клетка
Рис. 8. Схематическое изображение внутренней части клетки (масштабы не соблюдены).
Необходимо отметить важную роль ЭПС в строительстве всех внутриклеточных мембран. Здесь начинается самый первый этап такого строительства.
Существенную роль играет ЭПС и в обмене ионов кальция. Этот ион имеет большое значение в регуляции клеточного метаболизма, изменяя проницаемость мембранных каналов, активируя различные соединения в цитоплазме и т.д. Гладкая ЭПС является депо ионов кальция. При необходимости кальций высвобождается и принимает участие в жизнедеятельности клетки. Эта функция более всего свойственна ЭПС мышц. Освобождение ионов кальция из ЭПС является звеном в сложном процессе сокращения мышцы.
Необходимо отметить тесную связь ЭПС с митохондриями - энергетическими станциями клетки. При заболеваниях, связанных с энергодифецитом, рибосомы отсоединяются от мембраны шероховатой ЭПС. Последствия не трудно предсказать – нарушается синтез белков на экспорт. А поскольку к таким белкам относятся пищеварительные ферменты, то при заболеваниях, связанных с энергодифецитом, будет нарушена работа пищеварительных желёз и, как следствие, пострадает одна из основных функций организма – пищеварительная. Исходя из этого, должна вырабатываться и фармакологическая тактика врача.
2.3. Комплекс Гольджи
В железах внутренней секреции, например, в поджелудочной железе, некоторые пузырьки, отделяясь от ЭПС, уплощаются, сливаются с другими пузырьками, накладываются друг на друга, как блины в стопке, образуя комплекс Гольджи (КГ). Состоит он из нескольких структурных элементов – цистерн, пузырьков и трубочек (рис. 9). Все эти элементы образованы однослойной мембраной жидкостно-мозаичного типа. В цистернах происходит «созревание» содержимого пузырьков. Последние отшнуровываются от комплекса и передвигаются в цитозоле по микротрубочкам, фибриллам и филаментам. Однако основной путь пузырьков – движение к плазматической мембране. Сливаясь с ней, пузырьки опорожняют своё содержимое с пищеварительными ферментами в межклеточное пространство (рис. 10). Из него ферменты попадают в проток и изливаются в кишечник. Процесс выведения при помощи пузырьков секрета КГ носит название экзоцитоз.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image056.gif" border="0" width="150" height="18 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image058.gif" border="0" width="34" height="12 src="> ЭПС
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image060.gif" border="0" width="12" height="39 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image062.jpg" border="0" width="336" height="226 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image064.gif" border="0" width="47" height="12 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image066.gif" border="0" width="27" height="12 src="> Мембрана
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image072.gif" border="0" width="27" height="12 src=">мембраны клетки из
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image074.gif" border="0" width="51" height="18 src=">мембраны пузырька.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image076.gif" border="0" width="26" height="67 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image078.gif" border="0" width="155" height="14 src=">из КГ
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image080.jpg" border="0" width="456" height="259 src=">
Микроорганизмы
Растворённые
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image084.gif" border="0" width="17" height="25 src=">вещества
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image086.gif" border="0" width="19" height="29 src=">4
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image088.gif" border="0" width="34" height="25 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image090.gif" border="0" width="39" height="11 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image092.gif" border="0" width="31" height="48 src="> 1а 4а
Белки, жиры Лизосома Фрагменты
углеводы митохондрий
Рис. 12. Функции лизосом:
1, 1а – утилизация органических веществ гиалоплазмы; 2, 2а – утилизация содержимого пиноцитозных пузырьков; 3, 3а – утилизация содержимого фагоцитозных пузырьков; 4, 4а – ферментативное расщепление поврежденных митохондрий. 3а – фагосомы.
ных органических соединений, которые после выхода в цитоплазму становятся участниками клеточного метаболизма. Переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полости лизосомы накапливаются непереваренные продукты. Такая лизосома называется остаточным тельцем. Там же откладываются пигментные вещества. У человека при старении организма в остаточных тельцах клеток мозга, печени и в мышечных волокнах накапливается пигмент старения - липофусцин.
Если вышесказанное можно условно охарактеризовать как действие лизосом на уровне клетки, то другая сторона деятельности этих органоидов проявляется на уровне целого организма, его систем и органов. Прежде всего это касается удаления отмирающих в процессе эмбриогенеза органов (например хвост у головастика), при дифференцировке клеток некоторых тканей (замена хряща костью) и т.д.
Учитывая большое значение ферментов лизосом в жизнедеятельности клетки, можно предположить, что любые нарушения их работы могут привести к тяжёлым последствиям. При повреждении гена, контролирующего синтез какого-либо фермента лизосом, у последнего произойдёт нарушение структуры. Это приведёт к тому, что в лизосомах будут накапливаться «непереваренные» продукты. Если в клетке таких лизосом становится слишком много, клетка повреждается и как результат нарушается работа соответствующих органов. Наследственные болезни, развивающиеся по такому сценарию, носят название «лизосомные болезни накопления».
Следует обратить внимание также на участие лизосом в формировании иммунного статуса организма (рис 13). Попадая в организм, антиген (например, токсин микроорганизма) в основном (около 90%) разрушается, что предохраняет клетки от его повреждающего действия. Оставшиеся в крови молекулы антигена поглощаются (пиноцитозом или фагоцитозом) макрофагами или специальными клетками с развитой лизосомальной сис
Бактерия
 |
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image099.gif" border="0" width="444" height="244 src="> Антиген
Макрофаг
пиноцитоза
Пиноцитозный
Лизосома
Пептидные фрагменты антигена
Рис. 13. Формирование в макрофаге пептидных фрагментов антигена
(масштабы не соблюдены).
темой. Пиноцитозный пузырёк или фагосома с антигеном соединяется с лизосомой и ферменты последней, расщепляют антиген на фрагменты, которые обладают большей антигенной активностью и меньшей токсичностью, чем первоначальный микробный антиген. Эти фрагменты в большом количестве выносятся на поверхность клеток, и происходит мощная активация иммунных систем организма. Понятно, что усиление антигенных свойств (на фоне отсутствия токсического эффекта), в результате лизосомальной обработки, значительно ускорит процесс развития защитных иммунных реакций на этот микроорганизм. Процесс расщепления лизосомами антигена на пептидные фрагменты носит название процессинг антигена . Необходимо отметить, что непосредственное участие в этом явлении принимают ЭПС и комплекс Гольджи.
И, наконец, в последнее время широко рассматривается вопрос взаимоотношения лизосом и микроорганизмов, фагоцитированных клеткой. Как мы излагали ранее, слияние фагосомы и лизосомы приводит к перевариванию микроорганизмов в фаголизосоме. Это наиболее благоприятный исход. Однако возможны и другие варианты взаимоотношений. Так, некоторые патогенные (болезнетворные) микроорганизмы при проникновении в клетку внутри фагосомы выделяют вещества, блокирующие слияние лизосом с фагосомой. Это даёт возможность сохраниться им в фагосомах. Однако срок жизни клеток (фагоцитов) с поглощёнными микроорганизмами невелик, они распадаются, выбрасывая в кровь фагосомы с микробами. Вышедшие в кровеносное русло микроорганизмы способны вновь спровоцировать рецидив (возврат) заболевания. Возможен и другой вариант, когда части разрушенного фагоцита, в том числе и фагосомы с микробами, вновь поглощаются другими фагоцитами, снова оставаясь в живом состоянии и в новой клетке. Цикл может повторяться достаточно длительное время. Описан случай заболевания сыпным тифом у пожилого пациента, который ещё юношей-красноармейцем перенёс сыпной тиф, сражаясь в Первой конной армии. Через пятьдесят с лишним лет повторились не только симптомы заболевания – даже бредовые видения возвращали старика в эпоху гражданской войны. Всё дело в том, что возбудители сыпного тифа обладают способностью блокировать процесс соединения фагосом и лизосом.
Функция лизосом:
· пищеварительная (переваривая части цитоплазмы и микроорганизмы, поставляет элементарные органические соединения для нужд клетки),
· утилизационная (очищает цитоплазму от распавшихся частей),
· участвуют в удалении отмирающих клеток и органов,
· защитная (переваривание микроорганизмов, участие в иммунных реакциях организма).
2.5. Рибосомы.
Это аппарат синтеза белка в клетке. В рибосому входят две субъединицы – большая и малая. Субъединицы имеют сложную конфигурацию (см. рис. 14) и состоят из белков и рибосомальной РНК (рРНК). Рибосомальная РНК служит своеобразным каркасом, на который крепятся молекулы белка.
Образование рибосом происходит в ядрышке ядра клетки (этот процесс будет рассмотрен ниже). Сформированные большая и малая субъединица выходят через ядерные поры в цитоплазму.
В цитоплазме рибосомы находятся в диссоциированном или диспергированном состоянии, это диссоциированные рибосомы . В таком состоянии они не способны прикрепиться к мембране. Это не рабочее состояние рибосомы. В рабочем состоянии рибосома представляет собой органоид, состоящий из двух скрепленных между собой субъединиц, между которыми проходит нить иРНК. Такие рибосомы могут свободно «плавать» в цитозоле, они называются свободные рибосомы , или прикрепляться к различным мембранам,
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image106.gif" border="0" width="444" height="240 src=">зывается полисомой (рис. 15).
Окончание синтеза белка Начало синтеза белка
Рис. 15. Схема синтеза белка полисомой.
На рисунке полисома состоит из пяти разных рибосом.
Обычно на мембранах шероховатой ЭПС синтезируются белки на экспорт, а в гиалоплазме – на нужды клетки. Если при заболевании обнаруживается отсоединение рибосом от мембран и переход их в гиалоплазму, то это можно рассматривать как защитную реакцию – с одной стороны, клетки сокращают экспорт белка и увеличивает синтез белка на внутренние нужды. С другой стороны, такое отсоединение рибосом свидетельствует о наступающем энергодифеците клетки, так как прикрепление и удержание рибосом на мембранах требует затраты энергии, основным поставщиком которой в клетке является АТФ. Недостаток АТФ закономерно приводит не только к отсоединению рибосом от мембраны, но и неспособности свободных рибосом прикрепиться к мембране. Это приводит к выключению из молекулярного хозяйства клетки эффективного генератора белка – шероховатой ЭПС. Считается, что энергодефицит – это серьёзное нарушение клеточного метаболизма, чаще всего связанное с нарушением в деятельности энергозависимых процессов (например в митохондриях).
В рибосоме имеются три различных участка, с которыми связывается РНК - один для матричной, или информационной РНК (мРНК, или иРНК), и два для транспортной РНК. Первый располагается в месте контакта большой и малой субъединицы. Из двух последних - один участок удерживает молекулу тРНК и формирует связи между аминокислотами (пептидные связи), поэтому его называют Р-центр. Он располагается в малой субъединице. А второй служит для удержания только что прибывшей молекулы тРНК, нагруженной аминокислотой. Его называют А-центром.
Следует подчеркнуть, что при синтезе белка некоторые антибиотики могут блокировать этот процесс (подробнее на этом мы остановимся, когда будем описывать трансляцию).
2.6. Митохондрии.
Их называют «энергетическими станциями клетки». У эукариот в процессе гликолиза, цикла Кребса и других биохимических реакций формируется большое количество электронов и протонов. Часть из них участвует в разнообразных биохимических реакциях, другая часть аккумулируется в специальных соединениях. Их несколько. Наиболее важные из них НАДН и НАДФН (никотинамидадениндинуклеотид и никатинамидадениндинуклеотид-фосфат). Эти соединения в форме НАД и НАДФ являются акцепторами – своеобразными «ловушками» электронов и протонов. После присоединения к ним электронов и протонов они превращаются в НАДН и НАДФН и являются уже донорами элементарных частиц. «Отлавливая» их в самых различных частях клетки, они переносят частицы в различные отделы цитоплазмы и, отдавая их на нужды биохимических реакций, обеспечивают бесперебойное течение метаболизма. Эти же соединения поставляют электроны и протоны в митохондрии из цитоплазмы и из матрикса митохондрий, где располагается мощный генератор элементарных частиц – цикл Кребса. НАДН и НАДФН, встраиваясь в цепь переноса электронов (см. далее), передают частицы на синтез АТФ. Из АТФ энергия черпается на все процессы, идущие в клетке с затратой энергии.
Митохондрии имеют две мембраны жидкостно-мозаичного типа. Между ними располагается межмембранное пространство. Внутренняя мембрана имеет складки – кристы (рис. 16). Внутренняя поверхность крист усеяна грибовидными тельцами, имеющими ножку и головку.
В грибовидных тельцах происходит синтез АТФ. В самой толще внутренней мембраны митохондрий располагаются ферментные комплексы, переносящие электроны с НАДН2 на кислород. Эти комплексы называются дыхательной цепью или цепью пере-
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image108.gif" border="0" width="51" height="12 src="> Рибосома
text-decoration: underline;">В неделящихся клетках этот комплекс образован двумя одинаково устроенными центриолями – материнской и дочерней. Одна центриоль представляет собой цилиндр, стенку которого формирует девять групп, в группе - три микротрубочки. Всего 9 триплетов. Центриоли располагаются перпендикулярно друг к другу (рис. 17). Органоид принимает участие в формировании микротрубочек цитоскелета. В двигающихся клетках микротрубочки непрерывно распадаются и образуются вновь. В специализированных клетках (нейроны) микротрубочки относительно стабильны.
В делящихся клетках обе центриоли вначале расходятся по полюсам, и между ними формируется система из микротрубочек. Самосборка микротрубочек происходит из белковых блоков. Один из белков, входящий в блок, носит название тубулин . Центриоли и
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image117.jpg" border="0" width="312" height="215 src=">
Рис. 17. Центриоли: клеточный центр (А), состоящий из двух перпендикулярно расположенных центриолей (Б, С).
система микротрубочек между центриолями напоминают веретено. Отсюда и название всей фигуры – веретено деления. Микротрубочки под микроскопом напоминают нити, поэтому иногда их так и называют – нити веретена. Микротрубочки прикрепляются к центромерам хромосом и принимают участие в «растаскивании» хроматид по полюсам делящейся клетки.
После завершения деления клетки в обеих дочерних клетках около каждой центриоли образуется новая дочерняя копия, которая сразу же ориентируется перпендикулярно по отношению к материнской. Воспроизведение новых центриолей осуществляется путём самосборки из белковых субъединиц, находящихся в цитоплазме.
Функции клеточного центра многообразны. Основные – участие в делении клетки и транспортировка клеточных органоидов, пино-и фагосом по цитоплазме. В последнее время появились данные, указывающие на то, что центриоли являются главными пространственно-временными координаторами клетки.
Необходимо отметить, что процесс, приводящий к формированию веретена деления, очень чувствителен к различным препаратам. Один из алкалоидов безвременника осеннего, колхицин, соединяясь с тубулином, препятствует формированию митотического веретена и блокирует деление клетки. Такие вещества называют антимитотическими агентами. К ним относят лекарственные препараты винбластин и винкристин, которые широко используются в терапии рака.
При этом заболевании клетки злокачественной опухоли интенсивно и бесконтрольно размножаются; повреждение веретена деления останавливает их деление.
2.8. Ядро - система управления клетки
Состоит из следующих частей: ядерной мембраны, кариоплазмы (кариолимфы), хромосом (во время деления клетки) или хроматина (в интерфазе), ядрышка и ядерного скелета (матрикса).
Почти вся ДНК эукариотической клетки в период интерфазы сосредоточена в ядре, объем которого составляет приблизительно 10% от общего объема клетки. Как правило, клетки содержат только одно ядро, но некоторые клетки, в зрелом состоянии, могут не иметь ядра. К ним относятся эритроциты периферической крови человека (млекопитающих). Встречаются клетки, имеющие два (клетки печени, инфузории) или много ядер (поперечно-полосатая мышца).
Ядро окружено ядерной оболочкой и содержит хроматин, а также одно или несколько ядрышек (рис. 18).
2.8.1. Ядерная оболочка
Состоит из двух мембран - наружной и внутренней. Пространство между ними называется перинуклеарным. Внешняя мембрана может переходить в мембраны ЭПС. Это позволяет постоянно обмениваться содержимым перинуклеарного пространства и каналов ЭПС. Кроме того, наружная мембрана способна формировать пузырьки, которые встраиваются в КГ. Возможен и обратный процесс – пузырьки, сформированные КГ, включаются в ядерную мембрану, изливая содержимое в перинуклеарное пространство.
Ядерная мембрана пронизана порами, через которые из ядра в цитоплазму транспортируются молекула иРНК, сложные образования из рРНК и белков (предшественники рибосом) и некоторые вещества. Пора представляет собой сложный комплекс, состоящий из нескольких белковых глобул.
Изнутри поверхность внутренней ядерной мембраны подстилает белковая сеть – ядерная ламина, которая участвует в организации хроматина и порового комплекса. При делении клетки ламина участвует в разрушении ядерной оболочки.
2.8.2. Нуклеоплазма (кариолимфа, ядерный сок) - внутренняя среда клетки
В виде бесструктурной массы окружает хромосомы и ядрышко. Вязкость кариоплазмы примерно такая же, как и гиалоплазмы, а кислотность выше. Она состоит из воды (основная часть) и растворённых в ней органических и неорганических соединений. Предполагается, что в кариоплазме происходят специфические биохимические процессы.
Шероховатая ЭПС
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image122.gif" border="0" width="98" height="45 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image124.gif" border="0" width="38" height="41 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image126.gif" border="0" width="125" height="12 src=">left">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image115.gif" border="0" width="63" height="12 src=">left">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image134.gif" border="0" height="12 src=">мембрана
Комплекс
text-decoration: underline;">Во время интерфазы экспрессия (функция, работа) генов максимальна и хромосомы имеют вид тонких нитей. Те участки нити, в которых происходит синтез РНК – деконденсированы, а те участки, где синтез не происходит, – наоборот, конденсированы (рис. 19).
Во время деления, когда ДНК в хромосомах практически не функционирует, хромосомы представляют собой плотные тельца, похожие на «Х» или «У». Это связано с сильной конденсацией ДНК в хромосомах.
Особо необходимо уяснить, что наследственный материал по-разному представлен в клетках, находящихся в интерфазе и в момент деления. В интерфазе в клетке отчётливо просматривается ядро, наследственный материал, в котором представлен хроматином. Хроматин, в свою очередь, состоит из частично конденсированных нитей хромосом. Если же рассматривать клетку во время деления, когда ядра уже нет, то весь наследственный материал концентрируется в хромосомах, которые максимально конденсированы (рис. 20).
Совокупность всех нитей хромосом, состоящих из ДНК и различных белков, в ядрах эукариотических клеток носит название хроматин (см. рис. 19. В). Хроматин в свою очередь делится на эухроматин и гетерохроматин . Первый слабо окрашивается красителями, т.к. содержит тонкие неконденсированные нити хромосом. Гетерохроматин, напротив, – содержит конденсированную, а следовательно, хорошо прокрашиваемую нить хромосомы. Неконденсированные участки хроматина содержат ДНК, в которой функционируют гены (т.е. происходит синтез РНК).
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image137.gif" border="0" width="43" height="54 src="> 1 Ядро
А Б В
Рис. 19. Хромосомы в интерфазе.
А – выделенная нить хромосомы из ядра клетки, находящейся в интерфазе. 1- конденсированный участок; 2 – неконденсированный участок.
Б – выделенные несколько нитей хромосом из ядра клетки, находящейся в интерфазе. 1 – конденсированный участок; 2 – неконденсированный участок. В – ядро клетки с нитями хромосом, находящейся в интерфазе. 1 – конденсированный участок; 2 – неконденсированный участок; 1 и 2 – хроматин ядра.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image150.gif" border="0" width="39" height="12 src=">Ядро Хромосомы
Рис. 20. Два состояния наследственного материала в клетках в клеточном цикле: А – в интерфазе наследственный материал расположен в хромосомах, которые частично деконденсированы и расположены в ядре; Б – при делении клетки наследственный материал выходит из ядра, хромосомы располагаются в цитоплазме.
Необходимо помнить, что если ген функционирует, то ДНК в этом участке деконденсирована. И наоборот, конденсация ДНК гена свидетельствует о блокаде активности гена. Феномен конденсации и деконденсации участков ДНК достаточно часто можно обнаружить, когда в клетке регулируется активность (включения или выключения) генов.
Субмолекулярное строение хроматина (в дальнейшем мы будем их называть интерфазные хромосомы) и хромосом делящейся клетки (в дальнейшем мы будем их называть метафазные хромосомы) до настоящего времени полностью не выяснено. Однако ясно, что при различных состояниях клетки (интерфаза и деление) организация наследственного материала различна. В основе интерфазных (ИХ) и метафазных хромосом (МХ) лежит нуклеосома . Нуклеосома состоит из центральной белковой части, вокруг которой обёрнута нить ДНК. Центральную часть образуют восемь молекул белка-гистона – Н2А, Н2В, Н3, Н4 (каждый гистон представлен двумя молекулами). В связи с этим сердцевина нуклеосомы носит название тетрамер, октамер или кор . Молекула ДНК в форме спирали обвивает кор 1,75 раз и переходит на соседний кор, обвивает его и переходит на следующую. Таким образом создаётся своеобразная фигура, напоминающая нитку (ДНК) с нанизанными на ней бусами (нуклеосомами).
Между нуклеосомами лежит ДНК, которая называется линкерной . С ней может связываться ещё один гистон – Н1. Если он связывается с линкерным участком, то ДНК изгибается и сворачивается в спираль (рис. 21. Б). Гистон Н1 принимает участие в сложном процессе конденсации ДНК, при котором нитка бус сворачивается в спираль толщиной 30 нм. Эта спираль носит название соленоид . Нити хромосом интерфазных клеток состоят из нитей бус и соленоидов. В метафазных хромосомах соленоид сворачивается в суперспираль, которая соединяется с сетчатой структурой (из белков), формируя петли, которые укладываются уже в виде хромосомы. Такая упаковка приводит к почти 5000-кратному уплотнению ДНК в метафазной хромосоме. На рисунке 23 представлена схема последовательной укладки хроматина. Понятно, что процесс спирализации ДНК в ИХ и МХ намного сложнее, но сказанное даёт возможность уяснить наиболее общие принципы упаковки хромосом.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image156.gif" border="0" width="64" height="19 src="> Н1
Рис. 21. Строение нуклеосом:
А – в неконденсированной хромосоме. Гистон Н1 не связан с линкерной ДНК. Б – в конденсированной хромосоме. Гистон Н1 связан с линкерной ДНК.
Необходимо отметить, что, каждая хромосома в метафазе состоит из двух хроматид, удерживаемых с помощью центромеры (первичной перетяжки). В основе каждой из этих хроматид лежат упакованные порознь дочерние молекулы ДНК. После процесса компактизации они становятся хорошо различимыми в световой микроскоп хроматидами одной хромосомы. В конце митоза они расходятся по дочерним клеткам. С момента отделения хроматид одной хромосомы друг от друга, их уже называют хромосомами, то есть хромосома содержит либо две хроматиды, перед делением, либо – одну (но она называется уже хромосомой) после деления.
Некоторые хромосомы, кроме первичной перетяжки, имеют вторичную. Её ещё называют ядрышковый организатор . Это тонкая нить хромосомы, на конце которой помещается спутник. Вторичная перетяжка, как и основная хромосома, состоит из ДНК, на которой располагаются гены, ответственные за синтез рибосомальных РНК. На концах хромосомы располагается участок, называемый теломерой . Он как бы «запечатывает» хромосому. Если теломера случайно отрывается, образуется «липкий» конец, который может соединиться с таким же концом другой хромосомы.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image158.gif" border="0" width="105" height="33 src="> Нить хромосомы
Хромосома
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image161.gif" border="0" width="87" height="12 src="> Нитка бус
Совокупность всех метафазных хромосом клетки, их форма и морфология называется кариотипом . По форме различают три типа хромосом – метацентрические, субметацентрические и акроцентрические (рис. 23). В метацентрических хромосомах центромера
2.8.4. Ядрышко
Это плотное, хорошо прокрашиваемое тельце, расположенное внутри ядра. В нем обнаружены ДНК, РНК и белки. Основу ядрышка составляют ядрышковые организаторы – участки ДНК, несущие множественные копии генов рРНК. На ДНК ядрышковых организаторов происходит синтез рибосомальных РНК. К ним присоединяются белки и формируется сложное образование - рибонуклеопротеидные (РНП) частицы. Это предшественники (или полуфабрикаты) малой и большой субъединиц рибосом. Процесс образования РНП в основном происходит в периферической части ядрышек. Предшественники ри-
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image170.jpg" border="0" width="288" height="99 src="> Спутник
Рибосомы
Предшественники рибосом
Рис. 24. Формирование рибосом в ядрышке ядра.
Размер ядрышка отражает степень его функциональной активности, которая широко варьирует в различных клетках и может изменяться в индивидуальной клетке. Чем интенсивнее происходит процесс формирования рибосом в цитоплазме, тем активнее осуществляется синтез специфических белков на рибосомах. В этом отношении примечательно действие стероидных гормонов (СГ) на клетки-мишени. СГ попадают в ядро и активируют синтез рРНК. В результате возрастает количество РНП и, как следствие, увеличивается число рибосом в цитоплазме. Это приводит к значительному возрастанию уровня синтеза специальных белков, которые через ряд биохимических и физиологических реакций обеспечивают определённый фармакологический эффект (например, разрастается железистый эпителий в матке).
В зависимости от фазы клеточного цикла внешний вид ядрышка заметно меняется. С началом митоза ядрышко уменьшается, а затем и вовсе исчезает. В конце митоза, когда возобновляется синтез рРНК, миниатюрные ядрышки вновь возникают на участках хромосом, содержащих гены рРНК.
2.8.5. Ядерный матрикс
Хромосомы в трёхмерном пространстве ядра располагаются не хаотично, а строго упорядоченно. Этому способствует каркасная внутриядерная структура, называемая ядерным матриксом или скелетом. В основе этой структуры - ядерная ламина (см. рис. 19). К ней прикрепляется внутренний белковый каркас, занимающий весь объём ядра. Хромосомы в интерфазе прикрепляются и к ламине и к участкам внутреннего белкового матрикса.
Все перечисленные компоненты – не застывшие жёсткие структуры, а подвижные образования, архитектура которых меняется в зависимости от функциональной особенности клетки.
Ядерный матрикс играет важную роль в организации хромосом, репликации ДНК и транскрипции генов. Ферменты репликации и транскрипции закреплены на ядерном матриксе, а нить ДНК «протаскивается» через этот фиксированный комплекс.
В последнее время ламина ядерного матрикса привлекает внимание исследователей, работающих над проблемой долгожительства. Исследования показали, что ламина состоит из нескольких различных белков, которые кодируются генами. Нарушение структуры этих генов (а следовательно, и белков ламины) резко сокращает продолжительность жизни экспериментальных животных.
2.9. Цитоплазматические включения
Включения – это непостоянные компоненты клетки, представляющие собой отложения веществ, не участвующие в данный момент в клеточном метаболизме. Большинство включений видимы в световой микроскоп в виде гранул и располагаются в гиалоплазме, органоидах, либо в вакуолях. Существуют твердые и жидкие включения. В зависимости от нужд клетки, они могут присутствовать в больших количествах или исчезать.
Любые твёрдые включения в клетке, как правило, максимально обезвожены. Этот феномен не только снижает объём включений, но и препятствует развитию в них бактерий. Кроме того, в процессе образования твёрдые включения становятся химически инертными, что не позволяет им вступать в химические реакции с клеточными соединениями.
По химическому составу различают углеводные, белковые, липидные и минеральные включения. По функции - трофические, секреторные, специальные и т.д.
а. Трофические (питательные) включения
К ним относятся белковые, жировые и полисахаридные включения.
Белковые включения . В клетке есть соединения, важность которых определяется тем, что в случае нужды они могут стать предшественниками целого ряда других, жизненно необходимых, для клетки веществ. К таким соединениям относятся аминокислоты. Они могут использоваться в клетке в качестве источников энергии для синтеза углеводов, жиров, гормонов и других метаболитов. Поэтому белковые включения фактически представляют собой своеобразное клеточное сырьё для производства аминокислот.
Судьба белковых включений во всех клетках примерно одинакова. Прежде всего они сливаются с лизосомой, где специальные ферменты расщепляют белки до аминокислот. Последние выходят из лизосом в цитоплазму. Часть из них в цитоплазме взаимодействует с тРНК и в таком виде транспортируется к рибосомам на синтез белка. Другая часть вступает в специальные биохимические циклы, где из них синтезируются жиры, углеводы, гормоны и другие метаболиты. И наконец, аминокислоты принимают участие в энергетическом обмене клетки.
Полисахаридные включения . Для животных клеток и клеток грибов - основным запасным питательным включением является гликоген. Для растений таким включением является крахмал.
Гликоген у человека в основном депонируется в клетках печени и используется не только для нужд самой клетки, но и в качестве энергетических ресурсов для всего организма. В последнем случае гликоген расщепляется в клетке до глюкозы, которая выходит из клетки в кровь и разносится по организму.
Гликоген – это большая ветвистая молекула, состоящая из глюкозных остатков. Специальные внутриклеточные процессы при необходимости отщепляют от молекулы гликогена глюкозные остатки и синтезируют глюкозу. Последняя поступает в кровь и расходуется на нужды клетки. Казалось бы, проще запасти в клетке саму глюкозу, не превращая её в гликоген, тем более что молекула глюкозы растворима и достаточно быстро проходит в клетку через плазматическую мембрану. Однако этому мешает то, что глюкоза также быстро, не задерживаясь, выходит из клетки. Задержать её в клетке в чистом виде практически невозможно. Кроме того, депонирование глюкозы в больших количествах опасно, т.к. это может привести к созданию такого градиента концентраций, что вначале произойдёт набухание клетки из-за поступления воды, а затем и её гибель. Поэтому специальная система ферментов, слегка видоизменив молекулу глюкозы, связывает её с такой же молекулой. Создаётся гигантская ветвистая молекула, состоящая из глюкозных остатков – гликоген. Эта молекула уже нерастворима, как глюкоза, и не способна изменить осмотические свойства клетки.
Жировые включения . Эти включения в гиалоплазме могут находиться в виде капель. Многие растения содержат масла, например подсолнечник, арахис и т.д. Богата жировыми включениями жировая ткань человека, служащая для предохранения организма от потерь тепла, энергетическим депо и как амортизатор при механических воздействиях.
Необходимо отметить, что запасов гликогена в организме среднего взрослого человека достаточно на один день нормальной активности, тогда как запаса жиров хватит на месяц. Если бы главным энергетическим резервом в нашем организме был гликоген, а не жиры, вес тела увеличился бы в среднем на 25 кг.
В некоторых случаях появление жировых включений в клетке является тревожным сигналом неблагополучия. Так, при заболевании дифтерией токсин микроорганизма блокирует утилизацию жирных кислот и они накапливаются в больших количествах в цитоплазме. При этом нарушается метаболизм и клетка гибнет. Чаще всего такие нарушения происходят в клетках сердечной мышцы. Заболевание называется дифтерийный миокардит.
Все питательные включения используются клеткой в моменты интенсивной жизнедеятельности. В эмбриогенезе существует потребность в большом количестве питательных веществ. Поэтому ещё на стадии овогенеза яйцеклетка интенсивно запасает различные питательные вещества (желток и др.) в виде включений, которые обеспечивают прохождение первых этапов эмбрионального развития.
б. Секреторные включения
Различные секреторные гранулы, образующиеся в железистых клетках животных, разнообразны по химической природе и могут быть представлены ионами, ферментами, гормонами, гликопротеинами и т.д., например, пищеварительные ферменты, синтезируемые клетками поджелудочной железы. Сигналом к формированию и опорожнению секреторных включений в поджелудочной железе является приём пищи. До приёма пищи происходит накопление включений в цитоплазме. Определяя количество включений в клетках поджелудочной железы, можно примерно предположить, чьи это клетки – голодного или сытого человека.
в. Пигментные включения
Обеспечивают окраску тканей и органов. В клетках животных часто запасается пигмент меланин. У растений - каротиноиды, флавоноиды.
г. Экскреторные включения
У растений, не имеющих специальных выделительных органов, нередко в вакуолях накапливаются конечные продукты жизнедеятельности в виде солей оксалата или карбоната кальция. В дальнейшем растение сбрасывает листья или кору вместе с такими накопленными кристаллическими включениями.
Ядро - обязательная часть клеток эукариот. Это основной регуляторный компонент клетки. Оно отвечает за хранение и передачу наследственной информации, управляет всеми обменными процессами в клетке. Не органоид, а компонент клетки.
Ядро состоит из:
1) ядерную оболочку (ядерную мембрану), через поры которой осуществляется обмен между ядром клетки и цитоплазмой.
2) ядерный сок, или кариоплазму,- полужидкую, слабо окрашиваемую плазматическую массу, заполняющую все ядра клетки и содержащую в себе остальные компоненты ядра;
3) хромосомы, которые в неделящемся ядре видны только с помощью специальных методов микроскопии. Совокупность хромосом клетки называется кариотипом. Хроматин на окрашенных препаратах клетки представляет собой сеть тонких тяжей (фибрилл), мелких гранул или глыбок.
4) одно или несколько сферических телец - ядрышек, являющихся специализированной частью ядра клетки и связанных с синтезом рибонуклеиновой кислоты и белков.
два состояния ядра:
1. интерфазное ядро - имеет ядер. оболочку- кариолемму.
2. ядро при делений клетки. присутствует только хроматин в разном состоянии.
ядрышки включают две зоны:
1. внутренняя- фибриллярная- молекул белка и пре РНК
2. наружняя- гранулярная- формируют субъединицы рибосом.
Оболочка ядра состоит из двух мембран, разделенных перинуклеарным пространством. Обе они пронизаны многочисленными порами, благодаря которым возможен обмен веществами между ядром и цитоплазмой.
Основные компоненты ядра - хромосомы, образованные из молекулы ДНК и различных белков. В световом микроскопе они хорошо различимы лишь в период клеточного деления (митоза, мейоза). В неделящейся клетке хромосомы имеют вид длинных тонких нитей, распределенных по всему объему ядра.
Главные функции клеточного ядра следующие:
- хранение информации;
- передача информации в цитоплазму с помощью транскрипции, т. е. синтеза переносящей информацию и-РНК;
- передача информации дочерним клеткам при репликации - делении клеток и ядер.
- регулирует биохимические, физиологические и морфологические процессы в клетке.
В ядре происходит репликация - удвоение молекул ДНК, а также транскрипция - синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Компартмент для ядра - кариотека - образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется - люменом или перинуклеарным пространством . Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной - жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
12. Двумембранные органоиды (митохондрии, пластиды). Их строение и функции.
Митохондрии - это структуры округлой или палочковидной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5-10 мкм.
Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана
образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс
органеллы
. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20-40 нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген.
В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2-6 копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. Главная функция
митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата -АТФ). В целом этот процесс называется окислительным фосфорилированием
.
Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая).
Пластиды – это полуавтономные (могут существовать относительно автономно от ядерной ДНК клетки) двумембранные органоиды, характерные для фотосинтезирующих эукариотных организмов. Различают три основных типа пластид: хлоропласты, хромопласты и лейкопласты. Совокупность пластид в клетке называют пластидомом . Каждый их этих типов при определенных условиях может переходить один в другой. Как и митохондрии, пластиды содержат собственные молекулы ДНК. Поэтому они также способны размножаться независимо от деления клетки. Пластиды характерны только для растительных клеток.
Хлоропласты. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр - от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом . Группа тилакоидов, уложенных наподобие стопки монет, называется граной . Граны связываются друг с другом уплощенными каналами - ламеллами. В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой . В строме имеются кольцевая «голая» ДНК, рибосомы, ферменты цикла Кальвина, зерна крахмала. Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н+. Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Хлоропласты низших растений называют хроматофорами.
Лейкопласты . Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Амилопласты -синтезируют и накапливают крахмал, элайопласты - масла, протеинопласты - белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты - каротиноиды , придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях и др. Хромопласты считаются конечной стадией развития пластид.
Пластиды могут взаимно превращаться друг в друга: лейкопласты - хлоропласты - хромопласты.
Одномембранные органоиды (ЭПС, аппарат Гольджи, лизосомы). Их строение и функции.
Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети . Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой.
Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.
Комплекс Гольджи -представляет собой стопку плоских мембранных мешочков, которые называются цистернами . Цистерны полностью изолированы друг от друга и не соединяются между собой. По краям от цистерн ответвляются многочисленные трубочки и пузырьки. От ЭПС время от времени отшнуровываются вакуоли (пузырьки) с синтезированными веществами, которые перемещаются к комплексу Гольджи и соединяются с ним. Вещества, синтезированные в ЭПС, усложняются и накапливаются в комплексе Гольджи. Функции комплекса Гольджи :1- В цистернах комплекса Гольджи происходит дальнейшее химическое преобразование и усложнение веществ, поступивших в него из ЭПС. Например, формируются вещества, необходимые для обновления мембраны клетки (гликопротеиды, гликолипиды),полисахариды.
2- В комплексе Гольджи происходит накопление веществ и их временное «хранение»
3- Образованные вещества «упаковываются» в пузырьки (в вакуоли) и в таком виде перемещаются по клетке.
4- В комплексе Гольджи образуются лизосомы (сферические органоиды с расщепляющими ферментами).
Лизосомы - мелкие сферические органоиды, стенки которых образованы одинарной мембраной; содержат литические (расщепляющие) ферменты. Сначала лизосомы, отшнуровавшиеся от комплекса Гольджи, содержат неактивные ферменты. При определенных условиях их ферменты активизируются. При слиянии лизосомы с фагоцитозной или пиноцитозной вакуолью образуется пищеварительная вакуоль, в которой происходит внутриклеточное переваривание различных веществ.
Функции лизосом :1- Осуществляют расщепление веществ, поглощенных в результате фагоцитоза и пиноцитоза. Биополимеры расщепляются до мономеров, которые поступают в клетку и используются на ее нужды. Например, они могут быть использованы для синтеза новых органических веществ или могут подвергаться дальнейшему расщеплению для получения энергии.
2- Разрушают старые, поврежденные, избыточные органоиды. Ращепление органоидов может происходить и во время голодания клетки.
Вакуоли - сферические одномембранные органоиды, представляющие собой резервуары воды и растворенных в ней веществ. К вакуолям относятся: фагоцитозные и пиноцитозные вакуоли , пищеварительные вакуоли, пузырьки, отшнуровывающиеся от ЭПС и комплекса Гольджи. Вакуоли животной клетки - мелкие, многочисленные, но их объем не превышает 5% от всего объема клетки. Их основная функция - транспорт веществ по клетке, осуществление взаимосвязи между органоидами.
В клетке растений на долю вакуолей приходится до 90% объема. В зрелой растительной клетки вакуоль одна, занимает центральное положение. Мембрана вакуоли растительной клетки - тонопласт, ее содержимое - клеточный сок. Функции вакуолей в растительной клетке: поддержание клеточной оболочки в напряжении, накопление различных веществ, в том числе отходов жизнедеятельности клетки. Вакуоли поставляют воду для процессов фотосинтеза. Могут входить:
Запасные вещества, которые могут использоваться самой клеткой (органические кислоты, аминокислоты, сахара, белки). - вещества, которые выводятся из обмена веществ клетки и накапливаются в вакуоли (фенолы, дубильные вещества, алкалоиды и др.) - фитогормоны, фитонциды,
Пигменты (красящие вещества), которые придают клеточному соку пурпурный, красный, синий, фиолетовый цвет, а иногда желтый или кремовый. Именно пигменты клеточного сока окрашивают лепестки цветков, плоды, корнеплоды
14.Немембранные органоиды (микротрубочки, клеточный центр, рибосомы). Их строение и функции. Рибосома - немембранный органоид клетки, осуществляющий биосинтез белка. Состоит из двух субъединиц - малой и большой. Рибосома состоит из 3-4 молекул р-РНК, образующих ее каркас, и нескольких десятков молекул различных белков. Рибосомы синтезируются в ядрышке. В клетке рибосомы могут располагаться на поверхности гранулярной ЭПС или в гиалоплазме клетки в виде полисом. Полисома - это комплекс и-РНК и нескольких рибосом, считывающих с нее информацию. Функция рибосом - биосинтез белка. Если рибосомы располагаются на ЭПС, то синтезируемые ими белки используются на нужды всего организма, рибосомы гиалоплазмы синтезируют белки на нужды самой клетки. Рибосомы прокариотических клеток мельче, чем рибосомы эукариот. Такие же мелкие рибосомы находятся в митохондриях и пластидах.
Микротрубочки - полые цилиндрические структуры клетки, состоящие из несократимогобелка тубулина. Микротрубочки не способны к сокращению. Стенки микротрубочки образованы 13 нитями белка тубулина. Микротрубочки располагаются в толще гиалоплазмы клеток.
Реснички и жгутики - органоиды движения. Главная функция - передвижение клеток или перемещение вдоль клеток окружающей их жидкости или частиц. В многоклеточном организме реснички характерны для эпителия дыхательных путей, маточных труб, а жгутики - для сперматозоидов. Реснички и жгутики отличаются только размерами - жгутики более длинные. В их основе - микротрубочки, расположенные по системе 9(2) + 2. Это значит, что 9 двойных микротрубочек (дуплетов) образуют стенку цилиндра, в центре которого располагаются 2 одиночные микротрубочки. Опорой ресничек и жгутиков являются базальные тельца. Базальное тельце имееет цилиндрическую форму, образовано 9 тройками (триплетами) микротрубочек, в центре базального тельца микротрубочек нет.
Кле точный центр - митотический центр, постоянная структура почти всех животных и некоторых растительных клеток, определяет полюса делящейся клетки (см. Митоз). Клеточный центр обычно состоит из двух центриолей - плотных гранул размером 0,2-0,8 мкм, расположенных под прямым углом друг к другу. При образовании митотического аппарата центриоли расходятся к полюсам клетки, определяя ориентировку веретена деления клетки. Поэтому правильнее К. ц. называть митотическим центром , отражая этим его функциональное значение, тем более что лишь в некоторых клетках К. ц. расположен в ее центре. В ходе развития организма изменяются как положение К. ц. в клетках, так и форма его. При делении клетки каждая из дочерних клеток получает пару центриолей. Процесс их удвоения происходит чаще в конце предыдущего клеточного деления. Возникновение ряда патологических форм деления клетки связано с ненормальным делением К. ц.