Уникальная по своему обеспечивает их разнообразие и особенности жизнедеятельности.
Современная микробиология для изучения и классификации использует следующие отличительные особенности бактерий:
- Морфологическое клеточное строение:
- шаровидные «кокки»;
- палочковидные «бациллы»;
- спиралевидные «спирохеты»;
- извитой формы «вибрионы».
- Дыхание:
- дышат кислородом «аэробные»;
- бескислородные «анаэробные».
- Способ питания:
- автотрофные «самостоятельные» – способны синтезировать органические вещества из неорганических с помощью энергии химических реакций или фотосинтеза (к примеру, сине-зеленые водоросли);
- гетеротрофные – преобразовывают и перерабатывают готовые органические вещества, полученные в результате жизнедеятельности других организмов, сюда относятся патогенные (болезнетворные), и симбиотические (полезные) микроорганизмы, а также бактерии, живущие за счет процессов брожения, гниения и т.д.
- Среда обитания:
- внешняя среда (почва, вода и т.д.);
- внутри живых организмов;
- мертвая органика (трупы, отложения, продукты жизнедеятельности).
- Способ распространения и выживания (образовывают ли клеточные споры, чехлы, слизь).
- Реакция на тест по Граму (разделение по особенностям состава и строения стенок, ключевая классификация в разрезе темы строения клеточной стенки):
- грамположительные – окрашиваются во время проведения процедуры окрашивания по Граму (оболочка проницаема для анилинового красителя (кристаллвиолет, метиловый фиолетовый и др.));
- грамотрицательные – не окрашиваются во время проведения процедуры окрашивания по Граму (клеточная оболочка непроницаема для красителя).
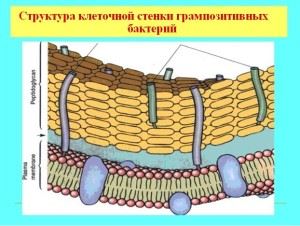
Особенности клеточной оболочки грамположительных бактерий
Суть проведения теста (предложен Г.К.Грамом в 1884 году) состоит в процедуре обработки образца бактериальной культуры анилиновым красителем (кристаллвиолет, кристаллический фиолетовый). При последующей промывке спиртом, Грам (+) виды микроорганизмов сохраняют синюю окраску, другие же обесцвечиваются. Способность клеточной оболочки фиксировать краситель определяется составом и строением клеточной стенки. Классификация одноклеточных по типу структуры клеточной стенки, поддающейся окраске или нет с помощью метода Грама, является одной из ключевых. На практике результаты грам-теста помогают диагностировать инфекционные заболевания изучать биохимические свойства микроорганизмов.
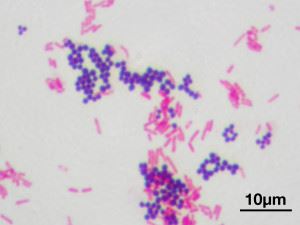
Окрашивание по Граму. Кокки (шаровидные) — грамположительные и бациллы (палочки) — грамотрицательные
Большинство грамположительных бактерий имеют массивную однослойную оболочку, проницаемую для анилинового красителя. Состав стенки включает в себя сложные органические молекулы, основу механической жесткости придает муреин (гетерополимер), гликопептиды, мукопептиды. Строение стенки определяет форму микроорганизма и является чем-то вроде внешнего скелета, с пористой структурой, толщиной порядка 40 молекул гликопептида. Кроме пептидогликановой основы, в состав оболочки входят тейхоевые кислоты и полисахариды.
Несмотря на общую реакцию на краситель, среди грамположительных можно встретить организмы, принадлежащие к самым разным подвидам, разных форм, размеров, среды обитания и способа питания.
Большинство патогенных для человека бактерий являются граположительными, среди которых, к примеру, возбудители таких инфекционных заболеваний:
- столбняк (Clostridium tetani),
- ботулизм (Clostridium botulinum),
- сибирская язва (),
- стрептококки,
- стафилококки (Staphylococcus aureus), обладающие высокой устойчивостью к температуре и действию лекарственных препаратов.
Кислотоустойчивые , благодаря особому многослойному строению оболочки (в нее входят воски, полисахариды и белки), как правило, не окрашиваются по методу Грама. Для них применяют специальный метод Циля-Нельсена. Но некоторые дают положительный результат по методу Грама в случае очень высокой концентрации красителя или повышенной температуры во время проведения процедуры.
Кислотоустойчивые одноклеточные сохраняют жизнеспособность в кислых и щелочных средах, хотя нормальная для их развития и роста среда – нейтральная. обычно классифицируются как грамположительные. Опасными для людей и животных представителями этой группы являются возбудители таких болезней, как туберкулез и проказа. Также кислотоустойчивыми являются некоторые грунтовые бактерии, способные к фиксации атмосферного азота и являющиеся симбионтами растений.
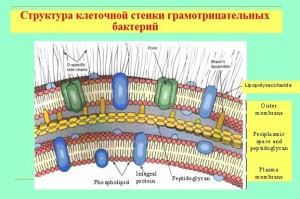
Особенности оболочки грамотрицательных бактерий
Несмотря на то, что толщина мукопептидной стенки у них намного меньше, стенка является непроницаемой для кристаллического фиолетового красителя. Все дело в принципиально ином строении и химическом составе клеточной . Оболочка имеет многослойную структуру и состоит из:
- внешней оболочки (мембаны), основной ее состав – липосахариды и белки;
- периплазматического пространства;
- внутреннего слоя (муреин пептидогликановый), его состав аналогичен, как и у стенок грамположительных бактерий, но как правило, более чем в два раза тоньше.
Благодаря более сложной структуре некоторые представители этой группы имеют повышенную устойчивость к антителам и антибиотикам.
С усложнением клеточной стенки появились новые функции и возможности. Некоторые виды бактерий, живущие на твердых поверхностях, приобрели возможность менять форму клетки (стенки «скелета» стали пластичными). Это важно при скольжении и перемещении по неровностям твердой среды обитания.
Периплазматическое пространство приобрело функцию изолированного места хранения некоторых гидролизующих ферментов, необходимых для жизнедеятельности клетки, но в то же время способных расщепить и полимерные молекулы самой бактерии. Благодаря гидролизу попадающих извне полимерных молекул живая клетка расширяет круг пригодных для питания веществ, в то же время непроницаемая для ферментов внутренняя оболочка препятствует «самоперевариванию» бактериальной клетки.
В структуру внешней мембраны входят белки, способные образовывать гидрофильные поры, через которые внутрь клетки могут проходить некоторые макромолекулы (сахара и аминокислоты), необходимые для питания бактерии.
К данному классу бактерий относятся многие бактерии, способные менять химический состав среды обитания (бактерии уксусного и спиртового брожения); симбиотические – кишечная палочка (E coli), многие патогенные (болезнетворные) организмы, такие как менингококк (Neisseria meningitidis); легионелла (Legionella pneumophila), и знаменитая Helicobacter pylori вызывающая язвенную болезнь и обладающая высокой кислотоустойчивостью.

Использование человеком особенностей строения бактерий
Первыми микроорганизмами, которые были использованы человеком для своих нужд, были молочнокислые и бактерии спиртового брожения. Именно они готовили для нас и готовят до сих пор сыр, хлеб и вино. Причем пользоваться продуктом их работы люди начали задолго до открытия бактерий и начала изучения строения и состава продуктов их жизнедеятельности.
В настоящее время биологические методы очистки сточных и фекальных вод стали доступны не только муниципальным предприятиям, но и частным домовладельцам. Спящие культуры входят в состав современных препаратов для выгребных ям и локальных канализаций. Фермерские хозяйства часто используют кислотоустойчивые для скоростной переработки компоста и животноводческих отходов на удобрения и для многих других целей.
Также многие генетически модифицированные бактерии участвуют в процессах производства лекарственных препаратов, синтеза новых видов полимеров и других материалов с уникальными химическими свойствами и сложным строением.
Химические компоненты клеточной стенки обычно разделяют на основные, инкрустирующие и откладывающиеся на ее поверхности (адкрустирующие). К основным химическим компонентам относятся целлюлоза, гемицеллюлоза, пектиновые вещества, белки. Компонентами, инкрустирующими клеточную стенку, являются лигнин и суберин. На поверхности клеточной стенки могут откладываться кутин, воск. Кроме того, клеточные стенки содержат силикаты и карбонаты кальция.
Самыми распространенными компонентами возобновляемого растительного сырья являются целлюлоза, лигнин и гемицеллюлозы (полиозы), которые составляют, соответственно, 40; 30 и 26% его общей массы. Целлюлоза является армирующим материалом клеточных стенок, образующим упорядоченные нитевидные структуры (фибриллы). Лигнин и гемицеллюлозы представляют аморфные биополимеры сетчатого или разветвленного строения и играют роль связующих (наполнителей).
Строение этих веществ, их межмолекулярное взаимодействие определяют свойства растительных субстратов в целом и особенности поведения их при переработке.
Целлюлоза
Целлюлоза является самым распространенным веществом на Земле, это скелет клеточной стенки растений. Лучше всего изучена структура целлюлозы хлопкового волокна и волокон, синтезируемых некоторыми бактериями, водорослями, а также моллюсками, так как в них целлюлоза встречается в наиболее чистой форме. В древесине она встречается только в виде композита с другими биополимерами, откуда ее сложно извлечь без частичного разрушения.
В химическом отношении целлюлоза представляет собой полисахарид, макромолекулы которого построены из остатков ß-D-глюкозы и представляют собой линейную цепь ангидроглюкопиранозных звеньев, соединенных 1,4-глюкозидными связями (рис.9).
Рис.9. Химическое строение целлюлозы
Это стереорегулярный полимер, в цепи которого стереоповторяющимся звеном служит остаток целлобиозы. Суммарная формула целлюлозы имеет вид – (С 6 Н 10 О 5)n или [С 6 Н 7 О 2 (ОН) 3 ]n. В мономерном звене целлюлозы содержатся три спиртовых гидроксильных группы, из которых одна первичная – СН 2 ОН, а две у С 2 и С 3 вторичные – СНОН. Концевые звенья отличаются от остальных звеньев цепи. Одно концевое звено (у С 4) имеет дополнительный, свободный вторичный спиртовый гидроксил. Концевое звено (у С 1) содержит свободный гликозидный (полуацетальный) гидроксил и может существовать в двух таутомерных формах – циклической (полуацетальной) и открытой (альдегидной). Концевая альдегидная группа придает целлюлозе редуцирующую (восстанвливающую) способность. По способности целлюлозы к реакции восстановления можно охарактеризовать длину ее цепей, свойственную данному виду растений.
В глюкозном остатке принято выделять гидрофильную и гидрофобную плоскости, в зависимости от того, сколько ОН- и СН-групп ей соотвествуют. Это важно при изучении взаимодействия глюкозного остатка с остатками ароматических аминокислот при ферментативном расщеплении молекулы целлюлозы в активном центре фермента.
Благодаря тому, что гидроксильные группы у первого и четвертого атомов глюкозы отличаются расположением относительно плоскости кольца, соседние остатки глюкозы в полимерной цепи оказываются повернутыми по отношению друг к другу на 180 0 . Полимерные цепи стабилизируются за счет водородных связей, образующихся между остатками глюкозы при взаимодействии гидроксильных групп при 6,2,3 углеродных атомов и кислорода кольца. Линейные полимерные цепи соединяются между собой с помощью межмолекулярных водородных связей, которые возникают между атомами кислорода одной цепи и гидроксильными группами при С 6 глюкозы другой цепи (рис. 10). В результате таких взаимодействий формируются прочные волокна. Гибкость молекулы целлюлозы определяется возможностью вращения ангидроглюкозных остатков в целлобиозном звене относительно гликозидной связи. Устойчивость линейной конформации целлюлозы определяется, в частности, минимальным отталкиванием С1-H и C4-H.
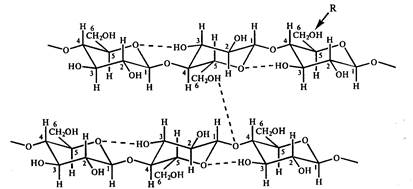
Рис. 10. Схематическое изображение системы водородных связей между остатками глюкозы в целлюлозе. R- центр возможной химической модификации целлюлозы. Так, в метилцеллюлозе, ацетилцеллюлозе и карбоксиметилцеллюлозе R= CH 3 , СОСН 3 , СН 2 СООNa соответственно
Молекула глюкозы представлена в конформации кресла, при которой атомы С2, С3, С5 и О5 лежат в одной плоскости, а атомы С1 и С4 - по разные стороны от нее. Остаток глюкозы может принимать также и энергетически менее выгодные конформации, которые по степени устойчивости располагаются в следующем порядке: скошенная конформация, конформация лодки (или ванны) и наименее выгодная конформация полукресла, при которой в одну плоскость попадают сразу 5 атомов 6-членного пиранозного цикла. Способность ангидроглюкозного звена принимать в молекуле целлюлозы помимо наиболее устойчивой менее устойчивые конформации (например, конформацию лодкиили конформациюполукресла) имеет важное значение при нарушении упорядоченной надмолекулярной структуры целлюлозы, а также при ферментативном гидролизе гликозидных связей в ее молекуле.
Элементарные волокна (мицеллы) целлюлозы состоят из 100-200 полимерных цепей, соединенных водородными мостиками. Эти волокна в клеточной стенке располагаются параллельно друг другу и группируются таким образом, что образуются пучки овальной формы – микрофибриллы. Около 10 мицелл образуют микрофибриллу диаметром 0,025 нм (рис.11).
Каждый пучок имеет центральную кристаллическую часть – ядро, в которой цепи упакованы настолько плотно, что в них с большим трудом проникают даже молекулы воды. Вокруг ядра располагается паракристаллический кортекс с менее упорядоченной структурой. Эти менее упорядоченные участки, называемые также аморфными,обычно составляют около 15% микроструктуры целлюлозы. Микрофибриллы могут объединяться в отдельные группы или слои – макрофибриллы – толщиной 0,4-0,5 мкм, видимые в световой микроскоп. Диаметр и степень кристалличности микрофибрилл довольно сильно варьируют в зависимости от вида растения и физиологического состояния клетки.
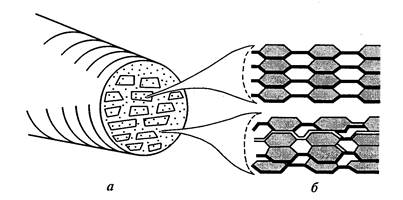
Рис.11. Модель строения микрофибриллы:а) поперечный разрез микрофибриллы с мицеллами и межмицелярными промежутками; б) схема расположения целлюлозы
Для древесной целлюлозы с индексом упорядоченности 50-70% поперечные размеры кристаллита не превышают 4х6 нм. По данным электронной микроскопии средние размеры в поперечнике могут составлять 1,5-2х2,5-3,5 нм. От 40 до 80% молекул при таких размерах расположено на поверхности кристаллита, где возможны их большая гидратация, нарушение дальнего порядка в структуре водородных связей, а также более сильное влияние химических воздействий.
Природная целлюлоза является полидисперсной, число глюкозных остатков в молекуле целлюлозы и, следовательно, длина самих молекул могут резко различаться в клеточных оболочках различных растений. Так, для целлюлозы волосков хлопчатника степень полимеризации первичной клеточной стенки равна 2-6 тысяч, вторичной – 13-14 тысяч, для целлюлозы Acetobacter xylinum- 2-6 тысяч, для древесины- 8-10 тысяч. Максимальная экспериментально установленная молекулярная масса целлюлозы льна составляет 6 000 000. В связи с тем, что степень полимеризации является усредненным показателем, наиболее точной характеристикой является кривая распределения молекул целлюлозы по длине (молекулярно-массовое распределение). Однако эту характеристику получить трудно и поэтому применяют усредненные характеристики: среднечисловую (DP n) и средневесовую (DP w) степени полимеризации. Среднечисловая степень полимеризации равна общему числу ангидроглюкозных остатков в образце целлюлозы, деленному на общее число молекул полимера. Для ее определения в образце полисахарида оттитровывают восстанавливающие концы. Средневесовую степень полимеризации целлюлозы определяют растворением образца целлюлозы в щелочном комплексе меди, цинка, кобальта или кадмия с этилендиамином, либо комплексе LiCl с N,N’-диметилацетамидом. Отношение DPw /DPn равно единице только у монодисперсных полимеров.
Для целлюлозы древесины ели, сосны, березы, и осины средневесовая степень полимеризации составляет 12000, при этом длина такой молекулы в вытянутом состоянии достигает почти 6 мкм, что в 6000 раз больше чем ее диаметр. Столь длинная и регулярно построенная молекула уложена определенным образом. Способ укладки определяет надмолекулярную структуру биополимера, основой которой являются внутри- и межмолекулярные водородные связи, образуемые целлобиозными звеньями. Исследование способа укладки и типов водородных связей в макромолекуле целлюлозы осуществляют с помощью конформационного анализа, дифракции (малоугловое рентгеновское рассеяние, рассеяние электронов), ИК-спектроскопии и ЯМР. Несмотря на значительные достижения в этой области вопрос о способах укладки молекул целлюлозы окончательно не решен.
Целлюлоза нерастворима в воде и органических растворителях, в химическом отношении инертна, механически очень прочна. Вода между близко расположенными, но не взаимодействующими непосредственно поверхностями кристаллитов, структурируется. Удаление гидратной воды, например в результате сушки способствует сближению изначально не контактировавших целлюлозных молекул и образованию между ними разнообразных типов водородных связей, которые не разрушаются при регидратации. Повышение степени кристалличности в процессе увлажнения исследователи связывают с проникновением воды из менее упорядоченных в более упорядоченные области. В результате наблюдается уменьшение абсолютных размеров кристаллитов и увеличение степени общей упорядоченности. Полагают, что вода при этом действует по принципу «молекулярной смазки», упрощая неупорядоченным фрагментам молекул достройку упорядоченной структуры путем кооперативного взаимодействия с кристаллической «затравкой». Замещение полярных групп целлюлозы, например, карбоксиметилом, ведет к нарушению молекулярной регулярности, способствует возникновению множества водородных связей с водой и делает целлюлозу растворимой.
Устойчивость целлюлозы к расщеплению, как в природных, так и в лабораторных условиях обусловлена не столько особенностями b-1,4- глюкозидной связи, сколько кристаллической структурой целлюлозы и особой «упаковкой» ее молекул в биологических структурах.
Во внутренние структуры фибрилл целлюлозы не могут проникать целлюлолитические ферменты. Они действуют на их поверхности. Число гликозидных связей, доступных действию ферментов, зависит в значительной мере от степени набухания целлюлозы. Увеличение степени набухания можно достичь за счет предобработки – пропаривания, размалывания, обработки ультразвуком и т.д. Набухание всего волокна увеличивают минеральные кислоты и щелочи в высоких концентрациях, так как они способны разрывать водородные связи и проникать в кристаллические участки. Сравнительно легко поддаются действию кислот и щелочей аморфные участки целлюлозы.
При действии на целлюлозу концентрированных растворов щелочи изменяются ее физико-химические и структурные свойства: отмечается интенсивное набухание, изменяется степень кристалличности. Высокомолекулярная фракция целлюлозы, нерастворимая в 17,5%- ной NaOH, называется a- целлюлозой. Хлопок состоит преимущественно из a- целлюлозы, а другие растения и древесина содержит преимущественно b- целлюлозы, растворимые в концентрированном растворе щелочи.
Микрофибриллы погружены в аморфный матрикс оболочки, который представляет собой сложную смесь полимеров, среди которых преобладают полисахариды различной молекулярной массы. Предполагается, что на поверхности всех микрофибрилл имеется слой молекул ксилоглюкана, сорбированный за счет водородных связей. Лигнин и гемицеллюлозы заполняют пространство между элементарными фибриллами целлюлозы. Таким образом, одревесневшее вещество является полимерной структурой, состоящей из целлюлозной арматуры, погруженной в лигноцеллюлозную матрицу.
Целлюлоза и гемицеллюлозы характеризуются различной кинетикой образования, что свидетельствует, по-видимому, о разных местах их синтеза. Синтез целлюлозы происходит, по-видимому, вне клетки на поверхности цитоплазматической мембраны при участии ГДФ или УДФ. Донорами гликозильных групп являются нуклеозиддифосфаты сахаров, которые образуются из глюкозы и сахарозы разными путями. Например, из нуклеозидтрифосфатов и сахар-1-фосфатов с участием пирофосфорилазы. Гликозильные остатки присоединяются к остаткам сахаров у невосстанавливающих концов растущей цепи. Между первым углеродом «добавляемого» остатка и гидроксильной группой четвертого углеродного атома концевого остатка при участии синтетазы возникает гликозидная связь.
N (Нуклеозиддифосфатсахар) + (сахар) n синтетаза n (Нуклеозиддифосфат) + (сахар) n +1
В клеточной стенке обнаружены ферменты синтеза целлюлозы и гранулы диаметром 8 нм, представляющие, по-видимому, мультиферментные комплексы, которые располагаются на наружной поверхности плазмалеммы и в матриксе стенки. Целлюлозосинтетаза (Е) в плазмалемме связана с активирующим белком Р, способным связывать ГТФ и переходить в активную форму с образованием комплекса Р-Е-ГТФ.
В процессе синтеза целлюлозы происходит образование не только одиночных цепей целлюлозы, но и формирование микрофибрилл целлюлозы. Такой синтез осуществляется, вероятно, при участии многих молекул целлюлозосинтетаз, сосредоточенных в отдельных местах плазмалеммы, причем целлюлозосинтетазы состоят из множества субъединиц, каждая из которых отвечает за синтез одиночной цепи глюкана. На внутренней стороне мембран были найдены «розетки» из шести белковых глобул. Розетки секретируются в плазмолемму с помощью эндомембранной системы везикул. На основании особенностей структурной связи розеток, терминальных комплексов и образующихся микрофибрилл целлюлозы возникло предположение о механизме формирования микрофибрилл в комплементарной трансмембранной структуре: розетки – терминальный комплекс – растущий конец микрофибриллы. С концом вновь синтезированной микрофибриллы связан терминальный комплекс, а розетки являются местом связывания предшественников целлюлозы и выведения «носителей» остатков глюкозы.
Одиночные розетки вместе с соответствующими, терминальными комплексами отвечают за синтез беспорядочно ориентированных микрофибрилл первичной клеточной стенки (рис.12), а ориентированная совокупность розеток до 175 штук на отдельных участках плазмалеммы определяет образование параллельно расположенных микрофибрилл вторичной клеточной стенки (рис.13).
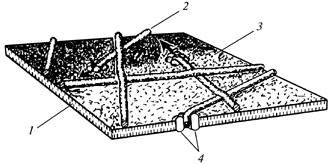
Рис.12.Микрофибриллы целлюлозы в период образования
первичной клеточной стенки:1
- двойной слой липида; 2
- микрофибрилла; 3
- внешняя поверхность плазмалеммы; 4
- розетка
Каждая розетка формирует микрофибриллу диаметром 5 нм. Расстояние между рядами розеток в плазмалемме соответствует рас-стоянию между микрофибриллами в клеточной стенке. Тонкие микрофибриллы содержат около 50 полиглюкановых цепей. Эти микрофибриллы образуют фибриллы вторичной клеточной стенки, диаметр которых может достигать 28 нм. Целлюлоза в отличие от крахмала является труднометаболируемым соединением. Ее усвояемость определяется происхождением, содержанием в пищевом рационе, характером предварительной обработки и колеблется в среднем от 6 до 23%. Так, перевариваемость целлюлозы пшеничных отрубей порядка 15%.
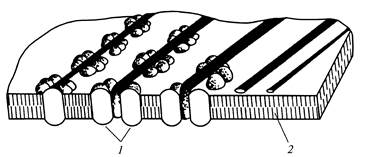
Рис. 13.Микрофибриллы целлюлозы в период образования
вторичной клеточной стенки: 1
- розетка; 2
- двойной слой липида
В пищевых волокнах целлюлоза составляет примерно одну треть. Ее эффективное разложение происходит в результате симбиотического взаимодействия различных микроорганизмов толстого кишечника и их целлюлолитических ферментов. В пищеварительном тракте целлюлоза стимулирует деятельность кишечника, усиливая его перистальтику, нормализует деятельность кишечной микрофлоры, сорбирует стерины, препятствует их всасыванию и способствует выделению холестерина. Широкое применение в качестве добавки к пище нашла микрокристаллическая целлюлоза (МКЦ) – не волокнистая порошкообразная модификация природной целлюлозы. Она не только снижает калорийность пищи, но и является ее загустителем и диспергатором, улучшающим качество блюд.
У грамотрицательныхэубактерий строение клеточной стенки намного сложнее, чем у грамположительных. В ее состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями.
Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий - чрезвычайно низкая сшитость ее пептидогликанового компонента.
Грамотрицательные прокариоты имеют наружную мембрану, в состав которой входят липиды (22%), белки, полисахариды, липопротеиды.
Липополисахариды (ЛПС) – гетерополимеры с комплексной структурой, обладающие разнообразной биологической активностью. Липоидный комплекс обуславливает токсичность (воспалительные реакции, лихорадка, эндотоксиновый шок), полисахаридный компонент ответственен за О-антигенспецифичность. ЛПС индуцирует синтез Jg М-антител, в иммунологии используется в качестве адъюванта и поликлонального активатора В-клеток.
Клеточная стенка у бактерий выполняет, в основном, формообразующую и защитную функции, обеспечивает ригидность, формирует капсулу, определяет способность клеток к адсорбции фагов.
№15 Фильтрующиеся, инволютивные формы бактерий, протопласты, сферопласты, L-формы.
протопласты, сферопласты, L-формы-это формы бактерий без КС
L-формы могут возникать в естеств-ых условиях в орг-зме чел-кав результате длительного лечения некот-ым антибиотиками, чаще всего пенициллином.
Различают нестабильные и стабильные L-формы бактерий. Первые способны к реверсии в исходный вид при устранении причины, вызвавшей их образование. Они восстан-ют способность синтезировать пептидогликан КС. Вторые, как правило, не способны к реверсии. L-формы разных бактерий играют существенную роль в патогенезе многих инфекционных заболеваний.
Основныесв-ва L-форм бактерий:
постоянное превращение из грампол-ых в грамотриц-е.
изменение антигенных св-в
снижение вирулентности
спос-ть к длительной персистенции
спос-ть при неполной утрате синетза КС к возврату в исходную форму.
№16 Тинкториальные свойства бактерий. Сложные методы окраски. Принцип окраски бактерий по методу Грама.
Методика окраски по Граму
На мазок кладут фильтровальную бумагу и наливают карболовый раствор генциановогофиолетового на 1-2 мин.
Снимают бумагу, сливают краситель и, не промывая мазок водой, наливают раствор Люголя на 1 мин.
Сливают раствор Люголя и обесцвечивают препарат в 96 0 спирте в течение 30 сек.
Промывают водой.
Красят 1-2 мин водным раствором фуксина.
Принцип окр. По Грамму
Грамположительные бактерии удерживают генциановыйфиолетовый в комплексе с йодом – фиолетовая окраска бактерий;
Грамотрицательные бактерии после воздействия спирта утрачивают краситель, обесцвечиваются и при обработке фуксином окрашиваются в красный цвет .
Сложные методы окраски применяют для изучения структуры клетки и дифференциации микроорганизмов. Окрашенные мазки микроскопируют в иммерсионной системе. Последовательно нанести на препарат определенные красители, различающиеся по химическому составу и цвету, протравы, спирты, кислоту и др.
Тинкториальныесвойства бактерий (лат. tinctura, от tingo - окрашиваю) способность к окрсаке: восприимчивость к окраске, кислото-спирто-щелочеустойчивость, равномерность окраски, метахроматичность, отношение к окраске по методу Грама.
№17 Особенности химического состава клеточной стенки кислотоустойчивых бактерий. Принцип окраски по методу Циля-Нильсена.
Кислотоустойчивые бактерии. Клеточная стенка некоторых бактерий содержит большое количество липидов и восков, делающих их устойчивыми к последующему после окрашивания обесцвечиванию кислотами, щелочами или этанолом (например, виды Mycobacterium или Nocardia). Подобные бактерии называют кислотоустойчивыми, их трудно окрашивать по Граму (хотя кислотоустойчивые бактерии рассматривают как грамположительные). Для их окраски применяют метод Циля-Нильсена.
Для микобактерий и нокардий характерна усложненная структура клеточной стенки. Основу у них, также как и у грамположительных бактерий, составляет муреиновый каркас, однако последний связан с полисахаридами и липидами. Липиды представлены миколовыми кислотами, которые придают клеточной поверхности гидрофобность. Гидрофобность, с одной стороны делает клетку устойчивой к действию различных химических веществ (такие бактерии называются кислотоустойчивыми), с другой стороны тормозит обмен клетки с окружающей средой и замедляет ее рост. Поэтому в питательные среды для культивирования микобактерий туберкулеза добавляют поверхностно-активные вещества. Кислотоустойчивость микобактерий является важным диагностическим признаком, для ее определения пользуются окраской по методу Циля-Нильсена.
Методика окраски кислотоустойчивых бактерий
по методу Циля-Нильсена
На фиксированный мазок помещают фильтровальную бумагу и наливают карболовый фуксин Циля и осторожно нагревают на горелке до появления паров. Операцию повторяют 2-3 раза.
Когда препарат остынет, снимают фильтровальную бумагу, сливают краситель и промывают препарат водой.
Препарат погружают 2-3 раза в стакан с 5% серной кислотой на 1-2 сек.
Тщательно промывают препарат водой и докрашивают щелочным метиленовым синим 3-5 мин.
Промывают водой и подсушивают.
Кислотоустойчивые бактерии не обесцвечиваются серной кислотой и сохраняют красный цвет, некислотоустойчивые теряют краситель и докрашиваются метиленовым синим в голубой цвет.
№18 Включения бактерий, их состав и биологическая роль. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. (по Нейссеру окраш-ся в темно-синий цвет)
№19 Споры бактерий, их строениеНекоторые бактерии в конце периода активного роста способны образовывать споры. Этому предшествует обеднение среды питательными веществами, изменение ее рН, накопление ядовитых продуктов метаболизма. Как правило, одна бактериальная клетка образует одну спору – локализация спор различна (центральная, терминальная, субтерминальная)
Если размеры спор не превышают поперечного размера палочковидной бактерии, то последняя называется бациллой (возбудитель сибирской язвы). Когда диаметр споры больше – бактерии имеют форму веретена и носят название клостридий (возбудители анаэробной инфекции). Клостридии столбняка имеют круглую спору и напоминают барабанные палочки. Клостридии ботулизма отличаются большими овальными спорами, что придает им вид теннисной ракетки.
По химическому составу различие спор от вегетативных клеток состоит лишь в количественном содержании химических соединений. Споры содержат меньше воды и больше липидов.
Формирование спор связано с уплотнением и обособлением определенного участка цитоплазмы вегетативной клетки с последующим образованием внутри бактерии круглого или овального тельца, покрытого плотной многослойной оболочкой, которая пропитана большим количеством липидов, кальция, дипиколиновой кислоты (рис. 12).
Попадая в организм человека и животных, споры прорастают в вегетативные клетки. Процесс прорастания спор включает три стадии: активации, начальной стадии и стадии роста. К активирующим агентам, нарушающим состояние покоя, относят повышенную температуру, кислую реакцию среды, механические повреждения и др. Спора начинает поглощать воду, освобождается от дипиколата кальция, с помощью гидролитических ферментов разрушает многие собственные структурные компоненты. После разрушения наружных слоев наступает период формирования вегетативной клетки с активацией биосинтеза, заканчивающейся делением клетки.
Метод окраски спор по Ожешко
Споры прочно удерживают карболовый фуксин и окрашиваются в красный цвет, цитоплазма бактерий обесцвечивается 5% серной кислотой и после докрашивания метиленовым синим приобретает синий цвет.
№20 Капсула и микрокапсула бактерийКапсула – слизистый слой клеточной стенки бактерий, состоящий из полисахаридов (пневмококк) или полипептидов (бацилла сибирской язвы). Микрокапсулу (толщиной менее 0,2 мкм) способны формировать большинство бактерий, четко выраженную макрокапсулу (толщиной более 0,2 мкм) формируют пневмококк, клебсиеллы, возбудитель сибирской язвы и некоторые другие. У патогенных бактерий капсула образуется в макроорганизме, на искусственных питательных средах она обычно утрачивается (за исключением клебсиелл).
В организме человека и животных капсула защищает патогенные бактерии от бактериофага, фагоцитоза и гуморальных факторов иммунитета, определяет антигенную специфичность микроорганизмов.
Капсулы, имея консистенцию геля, плохо удерживают краситель, и для их выявления чаще всего применяют методы негативного контрастирования.
Метод выявления капсулы по Бурри-Гинса
Бактерии окрашиваются в красный цвет, неокрашенные капсулы контрастно выделяются на темном фоне препарата.
№21 Жгутики бактерийЖгутики выполняют роль органа движения, позволяющего бактериям передвигаться со скоростью 20-60 мкм/сек. Бактерии могут иметь один (монотрихи) или несколько жгутиков, располагающихся по всей поверхности тела (перитрихи), либо собранные в пучки (лофотрихи).
Перитрихиальное расположение жгутиков характерно для энтеробактерий, возбудителей анаэробной инфекции, столбняка, ботулизма; монотрихом является холерный вибрион, лофотрихом - псевдомонас. У некоторых видов спирилл различают амфитрихиальное расположение жгутиков. Толщина жгутиков в среднем составляет 10-30 нм, а длина достигает 10-20 мкм.
Основу жгутика составляет длинная спиральная нить (фибрилла), которая у поверхности клеточной стенки переходит в утолщенную изогнутую структуру- крюк и прикрепляется к базальной грануле, вмонтированной в клеточную стенку и ЦПМ (рис. 10).
Базальные гранулы имеют диаметр около 40 нм и состоят из нескольких колец (одна пара у грамположительных бактерий, четыре - у грамотрицательных прокариот). Удаление пептидогликанового слоя клеточной стенки ведет к потере способности бактерий к движению, хотя жгутики при этом остаются неповрежденными.
Жгутики почти полностью состоят из белка флагеллина с некоторым содержанием углеводов и РНК.
Под микроскопом жгутики можно увидеть лишь после специальных методов протравливания и импрегнации солями серебра и ртути с последующей окраской метиленовой синью (метод Леффлера). При этом необходимо учитывать, что жгутики очень чувствительны к различным механическим воздействиям. О наличии жгутиков можно косвенно судить по направленному характеру движения в «висячей» и «раздавленной» капле в темнопольном и фазово-контрастном микроскопах, либо при светлопольной микроскопии при опущенном конденсоре и частично прикрытой диафрагме микроскопа.
Окраска жгутиков методом Леффлера
При микроскопии готового препарата жгутики видны как тонкие нитевидные структуры.
№22 ПилиПоверхность энтеробактерий и нескольких других микроорганизмов покрыта большим числом (от 10 до нескольких тысяч) ворсинок – нитевидных образований белковой природы. Как и жгутики, они построены из одного вида белка – пилина, субъединицы которого организованны в виде полой внутри нити и берут начало от ЦПМ. Они короче и тоньше жгутиков, их ширина 10-12 нм и длина до 12 мкм.
Ворсинки полифункциональны: обеспечивают трансмиссивную передачу генов (конъюгация), являются рецепторами для фагов, органом прикрепления бактерий к питательному субстрату (адгезия), участвуют в транспорте метаболитов.
У стрептококков имеется наружный слой протеиновых волосков (фимбрий), которые получили название белок М (М-протеин). Этот белок играет важную роль в процессах взаимоотношений бактерий с макроорганизмом.
№23 Актиномицеты
Представляют своеобразную группу бактерий, имеющих вид небольших или длинных несептированных ветвящихся нитей. Скопление гифов называют мицелием. Сходство с грибами чисто внешнее, так как актиномицеты имеют прокариотический тип клетки с наличием клеточной стенки не содержащей хитина и целлюлозы. Актиномицеты грамположительны, многие формы кислотоустойчивы, некоторые актиномицеты вокруг нитей имеют капсулу.
Субстратный мицелий образуется в результате врастания мицелия в питательную среду и воздушный, растущий на поверхности среды (рис. 19). В пораженных тканях (тканевая форма) актиномицеты могут образовывать друзы-гранулы, из плотно переплетенных нитей в виде лучей, отходящих от центра и заканчивающихся колбовидными утолщениями.
Актиномицеты размножаются бесполым путем, образую конидии или спороносцы со спорангиями на концах воздушного мицелия. Спороносцы могут быть прямыми, волнистыми, спиральными. Споры – овальными, круглыми, цилиндрическими, с гладкой поверхностью или шипами, иногда подвижные за счет жгутиков (зооспоры). Споры служат для размножения актиномицетов, они не термостойки, но выдерживают высушивание. Кроме того, возможно почкование и фрагментация мицелия на палочковидные или кокковидные формы.
Актиномицеты широко распространены в природе, обитают в воде, почве богатой перегноем. Они участвуют в круговороте веществ в природе. Отдельные виды актиномицетов используются как продуценты антибиотиков, витаминов, липидов, протеаз, аминокислот, стероидов.
Актиномицеты относятся к порядку Аctinomycetalеs, включающего семейства: Actinomycetaceae, Nocardiaceae, Streptomycetaceae, Mycobacteriaceae.
Патогенные для человека виды встречаются среди представителей семейства Actinomycetaceae и Nocardiaceae. Первые имеют вид длинных или коротких разветвленных палочек, не образующих воздушного мицелия. Они являются возбудителями актиномикоза человека и образуют друзы в пораженных тканях.
Представители семейства Nocardiaceae напоминают микобактерии, имеют нитевидную форму клеток и образуют на питательных средах воздушный и субстратный мицелий. Гифы фрагментируются на кокковидные и палочковидные клетки. Патогенные нокардии вызывают нокардиоз.
Методы исследования . окрашивают по Граму и Цилю-Нильсену. Друзу извлекают из патологического материала петлей, помещают в каплю воды на предметное стекло, слегка придавливают покровным, затем вводят под стекло каплю щелочного раствора метиленового синего и микроскопируют, можно использовать фазовый контраст.
№24 Спирохеты
спирально извитые, обладающие активной подвижностью бактерии. Размеры спирохет колеблются в толщину от 0,1-0,3 мкм, в длину от 7-500 мкм. Движения разнообразные – от винтообразных до сгибательных. Электронно-микроскопическое исследование позволило различить у спирохет протоплазматический цилиндр (тело клетки), аксиальную (опорную) нить и трехслойную наружную оболочку. Аксиальная нить находится в периплазматическом пространстве между наружной оболочкой и протоплазматическим цилиндром и состоит из отдельных фибрилл (эндофлагелл), число которых у разных видов различно: у трепонем и лептоспир – 3-4; у борелий – до 30. Каждая из фибрилл (эндожгутиков) закрепляется в области прикрепительных дисков на концах протоплазматического цилиндра и тянется к противоположному его концу, обвивая его и заканчиваясь свободно. Химический состав фибрилл аналогичен составу жгутиков (рис. 20).
В протоплазматическом цилиндре содержатся: нуклеоид, рибосомы, мезосомы, включения. Наружная оболочка (клеточная стенка) содержит тонкий слой пептидогликана, эластична и не обладает ригидностью. Эндоспор, капсул и экзожгутиков эти бактерии не образуют, грамотрицательны, в мазке располагаются беспорядочно.
Спирохеты относятся к порядку Spirochaetales, семейство Spirochaetaceae, которое включает три рода:
Borrelia - имеет 3-10 неравномерных отлогих завитков, концы заострены, длиной 10-30 мкм. Движение толчкообразное, по Романовскому-Гимзе окрашиваются в сине-фиолетовый цвет (представитель Borreliarecurrentis – вызывает эпидемический возвратный тиф; Borreliaburgdorferi - вызывает лаймоборрелиоз).
Treponema - имеет 8-14 туго закрученных, одинаковых по амплитуде завитков, длина 5-15 мкм. Движение плавное, медленное с вращением вокруг продольной оси, по Романовскому-Гимзе окрашиваются в бледно розовый цвет (представитель Treponemapallidum – возбудитель сифилиса).
Leptospira - имеет до двух десятков мелких частых завитков, заканчивающихся крючком с пуговчатым утолщением, длиной 5-15 мкм. Движение очень активное, поступательное перемещение вперед, сгибание и вращение вокруг оси. По Романовскому-Гимзе окрашиваются слабо в розовато-сиреневый цвет (представитель Leptospirainterrogans – возбудитель лептоспироза).
Методы исследования . В живом состоянии спирохеты изучают в фазово-контрастном микроскопе и темнопольном микроскопе, наблюдая за активным характерным движением спирохет, особенностями их формы.
Готовят препараты по Бурри (на темном фоне препарата становятся видимыми светлые извитые нити спирохет), окрашивают по Романовскому-Гимзе, по методу Морозова.
№25 Риккетсии
Class « Alphaproteobacteria»
Order Rickettsiales
Family Rickettsiaceae
Genus Rickettsia
Family Ehrlichiaceae
Риккетсии разнообразны по форме и выделяют следующие типы:
кокковидные однозернистые (до 0,5 мкм);
палочковидные двухзернистые (1-1,5 мкм);
бациллярные трех-четырехзернистые (3-4 мкм);
нитевидные многозернистые (10-40 мкм).
Зерна (нуклеопротеиды) обнаруживаются при окраске по Романовскому-Гимзе. Все формы взаимообратимы. Структурно имеют все компоненты бактериальной клетки: клеточную стенку, липоидную капсулу, цитоплазму, нуклеоид, рибосомы, пили. Риккетсии содержат как ДНК, так и РНК, обладают высоким содержанием фосфолипидов, содержание углеводов невелико.
В большинстве случаев (кроме вида Rochalimaeaguintana) на искусственных питательных средах риккетсии не растут. Жизненный цикл риккетсий зависит от жизнедеятельности клетки-хозяина и складывается из двух стадий: вегетативной и покоящейся (элементарные тельца). Риккетсии, находящиеся в вегетативной стадии (рис. 23) активно размножаются путем бинарного деления и обладают активной подвижностью, по-видимому, обусловленной жгутиковыми структурами. Риккетсии покоящейся стадии (элементарные тельца) имеют сферическую форму и они не активны.
Методы исследования. Риккетсии хорошо окрашиваются по Романовскому-Гимзе в сиреневый цвет, по Морозову (методом серебрения) в черный цвет. Для дифференциации риккетсий применяется метод окраски, предложенный П.Ф. Здродовским:
Риккетсии окрашиваются в рубиново-красный цвет и легко обнаруживаются на фоне голубой цитоплазмы и синего ядра клеток.
№26 Хламидии
Class «Chlamydiae»
Order Chlamydiales
Familia Chlamydiaceae
Genus: Chlamydia
1). Элементарные тельца – мелкие (0,2-0,5 мкм) электронноплотные шаровидные структуры, лишенные метаболитной активности, имеющие компактный нуклеоид и ригидную клеточную стенку, которые фильтруются через бактериальные фильтры. Они являются инфекционным началом хламидий и обеспечивают их выживание во внеклеточной среде и заражение новых клеток.
2). Ретикулярные тельца – более крупные (0,8-1,5 мкм), сферические образования, имеющие сетчатую структуру с тонкой клеточной стенкой и фибриллярным нуклеоидом. Они вырастают из элементарных телец внутри клеток, лишены инфекционности и, подвергаясь делению, обеспечивают репродукцию хламидий. Отсюда другое, исторически первое название ретикулярных телец – «инициальное тело». Ретикулярные тельца являются вегетативной формой хламидий.
3). Промежуточные тельца – промежуточная стадия между элементарными и ретикулярными тельцами.
Жизненный цикл хламидий начинается с того, что элементарные тельца фагоцитируются клеткой-хозяином, а затем в течение нескольких часов реорганизуются, увеличиваются в размерах и превращаются в ретикулярные формы, которые размножаются путем поперечного деления. Жизненный цикл заканчивается, когда возникающие промежуточные формы уплотняются, уменьшаются в размерах и превращаются в элементарные тельца. Размножаясь внутри цитоплазматических вакуолей, хламидии образуют микроколонии (включения), окруженные мембраной. В составе микроколоний обнаруживаются все три стадии развития хламидий. После разрыва стенки вакуоли (везикулы) и мембраны клетки-хозяина, вновь образовавшиеся хламидии высвобождаются, и элементарные тельца, инфицируя другие клетки, повторяют цикл развития. В оптимальных условиях роста в эукариотических клетках жизненный цикл хламидий составляет 17-40 часов (рис. 24).
Своеобразие хламидий проявляется и в строении их клеточной стенки. Она лишена пептидогликана и представляет собой двухслойную мембрану, ригидность которой определяют пептиды, перекрестно сшитые дисульфидными мостиками. В остальном хламидии напоминают грамотрицательные бактерии, так как содержат гликолипиды, сходные с липополисахаридами
Порядок Chlamydialesвключает одно семействоChlamydiaceaeс единственным родомChlamydia. Для человека патогенны видыC.trachomatis,C.psittaci,C.pneumoniae. Хламидии вызывают у людей заболевания глаз, дыхательной и мочеполовой систем и объединяются под общим названием «хламидиозы».
Методы исследования. Для микроскопического обнаружения телец включений (микроколоний) хламидий в инфицированных клетках (тканях) применяют различные методы окрашивания: Романовского-Гимзы, Маккиавелло и другие. При окрашивании по Романовскому-Гимзе они приобретают голубой или фиолетовый цвет. Кроме того, хламидии хорошо видны и в неокрашенном состоянии при микроскопии влажных препаратов под стеклом с помощью фазовоконтрастной оптической системы. В последнее время наиболее часто используется прямая реакция иммунофлюоресценции, окраска акридин –оранжевым.
№27 МикоплазмыClass Mollicutes
Order Mycoplasmatales
Family Mycoplasmataceae
Genus Mycoplasma
Genus Ureaplasma
Пять видов являются патогенными для человека: M . pneumoniae , M . hominis , M . genitalium , M . incognitus и U . urealyticum .
Микоплазмы – самые мелкие прокариоты (125-150 нм) способные самостоятельно размножаться. Полагают, что микоплазмы являются наиболее близкими потомками исходных прокариотических клеток. Геном микоплазм минимален для клетки, он в пять раз меньше генома кишечной палочки и составляет 0,45 МД. Главная особенность микоплазм – отсутствие клеточной стенки. Они окружены капсулоподобным слоем, под которым находится лишь тонкая трехслойная мембрана толщиной 7,5- 10 нм, содержащая в значительном количестве холестерин. Вследствие этого, микоплазмы выделяют в особый отделTenericutes, классMollicutes(«нежная кожа»), порядокMycoplasmatales.
Из-за отсутствия клеточной стенки микоплазмы (рис. 21) осмотически чувствительны и имеют разнообразную форму:
а) мелкие сферические или овоидные клетки размером 0,2 мкм (элементарные тельца) которые фильтруются через бактериальные фильтры;
б) более крупные шаровидные, размером до 1,5 мкм;
в) нитевидные, ветвящиеся клетки размером до 150 мкм.
Микоплазмы не образуют спор, жгутиков, некоторые виды обладают скользящей подвижностью.
Размножаются путем бинарного деления шаровидных и нитевидных клеток, почкования и высвобождения множества элементарных телец, образующихся в нитях.
Что касается энергии, то микоплазмы получают ее обычным для факультативных анаэробов способом, ферментируя углеводы или аминокислоты. Вследствие малого генома микоплазмы обладают ограниченными биосинтетическими способностями, и их приходится культивировать на питательных средах обогащенных липидами, белками, предшественниками нуклеиновых кислот. Растут медленно, колонии с плотным врастающим в среду центром, напоминающие «яичницу-глазунью» (темный центр и более светлая ажурная периферия). Размеры колоний мелкие, не превышающие 600 мкм.
В патологии человека наибольшую роль играют несколько представителей рода Mycoplasma:M.pneumoniae,M.hominis,M.anthritidisи единственный вид родаUreaplasma–U.urealyticum(названный так из-за уреазной активности). Патогенные микоплазмы вызывают заболевания (микоплазмозы) дыхательных, мочеполовых путей и суставов с разнообразными клиническими проявлениями. При лечении этих заболеваний следует помнить, что микоплазмы не чувствительны к бета-лактамным антибиотикам и другим лекарственным препаратам, угнетающим синтез клеточной стенки (из-за ее отсутствия у возбудителя).
Методы исследования . В световом микроскопе обнаруживаются лишь самые крупные формы микоплазм. В живом состоянии их изучают в темнопольном и фазово-контрастном микроскопе, ультраструктурные компоненты выявляют при помощи электронной микроскопии.
№28 Грибы
Классы (7) : · Basidiomycetes (шляпочные грибы). · Zygomycetes (род Mucor – мукоромикоз человека и животных) · Ascomycetes (сумчатые грибки, роды Aspergillus , Penicillium , дрожжевые грибки) · Deuteromycetes – несовершенные грибки, не размножаются половым путем (Candida )
Грибы – большая и успешно развивающаяся группа организмов, включающая около 80 000 идентифицированных видов. Размеры их колеблются от одноклеточных дрожжей до больших поганок, дождевиков и рожков. Грибы занимают самые различные местообитания – как в воде, так и на суше. Кроме того, они имеют важное значение и в связи с той ролью, которую играют в биосфере, и в связи с тем, что используются человеком в медицине и в хозяйстве
Методы исследования . Для микроскопического исследования готовят как нативные (неокрашенные), так и окрашенные препараты.
Исследование неокрашенных препаратов
Чтобы яснее различить элементы грибка, производят просветление препарата, для этого патологический материал (корочки, кусочки ногтя, волос, соскобы со слизистых, содержимое гранулематозных очагов) помещают на часовое стекло или чашку Петри, куда наливают 10-15% раствор едкого натрия или калия и ставят в термостат при 37 0 С на 20-30 мин. Затем материал извлекают и помещают в каплю 50% раствора глицерина на предметное стекло и закрывают покровным стеклом, микроскопируют в фазово-контрастном или световом микроскопе. Можно использовать другой метод: на патологический материал наносят каплю глицерина с добавлением 10% едкого калия и исследуют через 4-5 мин, закрыв покровным стеклом.
Гной из абсцессов, содержимое язв, мокроту разбавляют физиологическим раствором или водно-спиртовым (1:1) или 50% водным раствором глицерина, готовят препарат «раздавленная капля» и рассматривают при увеличении х200, х400 используя фазовый контраст.
Исследование окрашенных препаратов
Из гноя, крови, ликвора, осадка бронхиальных смывов и мочи готовят тонкие мазки, которые фиксируют в смеси Никифорова, Карнуа, спирт-формоле, высушивают и окрашивают:
Окраска лактофуксином , содержащим кислого фуксина – 0,1 г, молочной кислоты – 100 мл. Окрашивают в течение 3-5 мин. Фон препарата розовый, мицелий опалесцирует голубым цветом. Хорошо окрашиваются грибы при мукормикозе и аспергилезе.
№29 Дрожжи и дрожеподобные Дрожжевые клетки имеют округлую, пиальную или вытянутую форму размером 8-10 мкм, двухконтур-ную оболочку. В цитоплазме отмечаются включения в виде гранул гликогена, волютина, липидов. Размножение происходит почкованием и аскоспорами. Дрожеподобные сходны с истинными дрожжами, отличием служат отсутствие аскоспор и способность к образо-1шшю псевдомицелия. При образовании псевдомицелия клетки вытягиваются в длину и соприкасаются узким основанием. Они вызы-пают кандидозы, которые развиваются у больных людей при резком снижении резистентности организма и длительном применении антибиотиков Методы изучения дрожжей
Современный период изучения биологического разнообразия характеризуется интенсивным развитием филогенетической систематики, которая направлена на реконструкцию конкретных путей исторического развития организмов. В микробиологии филогенетическая систематика получила мощный импульс развития лишь в самом конце XX в. в связи со сравнительным изучением консервативных нуклеотидных последовательностей в рРНК. У дрожжей такая систематика строится главным образом на изучении двух участков рДНКдлиной около 600 нуклеотидных пар: D1/D2 домена на 5"-конце гена, кодирующего 26S рРНК и внутреннего транскрибируемого спейсерного региона (ITS), включающего ген 5.8S рРНК. Считается, что вследствие консервативности этих участков различия между ними прямо пропорциональны филогенетическому расстоянию, степени эволюционного родства.Секвенированиенеклеотидных последовательностей рДНК оказалось мощным инструментом для построения филогенетической классификации дрожжей, определения их места в общей системе грибов.
К настоящему времени расшифрованы и помещены в компьютерные банки данных, доступные по сети Интернет, нуклеотидные последовательности рРНК у представителей всех известных видов дрожжей. Это позволяет строить филогенетические деревья, отражающие эволюцию их рибосомальных генов. Оказалось, что группирование дрожжей на основе сходства нуклеотидных последовательностей рРНК во многих случаях не совпадает с группированием по фенотипическим признакам. Многие традиционные признаки, используемые в классификации дрожжей, такие как характеристикивегетативного размножения,форма аскоспор, способность к сбраживанию и ассимиляции сахаров, стали считаться ненадежными, не пригодными для определения филогенетического родства. Секвенирование рРНК (рДНК) сейчас считается необходимым при описании новых видов дрожжей.
Особенно сильное влияние на изучение дрожжей, также как и большинства других групп микроорганизмов, оказало бурное развитие в конце XX в. молекулярной биологии. В современной систематике дрожжей широко используются методы геносистематики, основанные на непосредственном сравнении геномов и секвенировании нуклеотидных последовательностей. Применение единых молекулярно-биологических методов позволило еще больше сблизить подходы к таксономии дрожжевых и мицелиальных грибов, установить связи между дрожжевыми анаморфами и мицелиальными телеоморфами, разработать новые критерии для создания единой филогенетической системы всего царства Mycota. В то же время, новые знания породили и новые научные проблемы, в частности, проблему соотношения новейших молекулярных методов с традиционными, основанными на морфологических и физиологических подходах к изучению дрожжей. Практически полностью расшифрован геном Saccharomyces cerevisiae, что открывает огромные перспективы геномики дрожжей, новые горизонты их биотехнологического использования. Таким образом, наука о дрожжах, проделав более чем полуторавековой путь, продолжает интенсивно развиваться и в XXI в.
Для микроскопического исследования готовят как нативные (неокрашенные), так и окрашенные препараты. Для изучения грибков в тканях проводится патогистологическое исследование.
клеточный дрожжи мицелий спора
30.Морфология и методы исследования нитчатых грибов. У грибов существует 2 типа роста: гифальный рост (гифомицеты) и дрожжевой рост (бластомицеты). Обычно вегетативное тело гифомицетов состоит из нитей толщиной около 5 мкм, сильно разветвленных и называемых гифами. Гифы либо не имеют поперечных перегородок (у низших грибов), либо разделены перегородками (септами) на клетки (у высших грибов). Стенка клеток может быть различной толщины, часто хорошо видна двухконтурность, среди включений в цитоплазме наиболее характерны зерна волютина, гликогена, пигмента меланина. Зрелые старые клетки грибов богаты липидами. Ядро содержит ядрышко и хроматиновую сеть, клетки могут быть многоядерными. Совокупность гифов образует мицелий (грибницу). Мицелий может быть субстратный, образующийся в результате врастания гифов в питательную среду и воздушный, растущий на поверхности среды. Мицелий представляет ветвящиеся трубки, ветвление осуществляется боковыми выростами гиф. Мицеальные нити иногда располагаются параллельными рядами, тесно прилегая друг к другу, напоминая «фитиль», отсюда и название «коремии» (у дерматофитов).
31.Основные принципы классификация и морфология простейших. Методы их исследования и медицинское значение.
Простейшие- одноклеточные эукариоты, близкие по строению к клеткам сложно организованных животных. Форма может быть грушевидной (трихомонады, лямблии), яйцевидной (балантидий), веретенообразной (трипаносомы, лейшма-нии), могут принимать самую причудливую конфигурацию (амебы)
Большинство простейших подвижно и передвижение осуществляется с помощью псевдоподий (амебы, малярийный плазмодий), жгутиков (лямблии, лейшмании), ресничек (балантидий).
Псевдоподии – временные выпячивания цитоплазмы, выпуская которые простейшие все время меняют форму тела.
Простейших относят к царству Protozoa (protos - первый, zoa - животные). Медицинское значение имеют:
Тип Sarcomastigophora, подтип Sarcodina (саркодовые). Тело их лишено пелликулы, передвигаются с помощью псевдоподий. К этому классу относятся различные виды амеб, в том числе дизентерийная амеба (Entamоeba histolytica).
Методы исследования . Для изучения простейших готовят временные и постоянные (окрашенные) препараты. Временный препараты готовят методом «раздавленной капли» или «висячей капли» с добавлением теплого физиологического раствора или витальных прижизненных красителей. Из крови готовят препараты «толстая капля». Для этого палец, обработанный эфиром, поворачивают проколом вниз и к выступающим каплям подносят предметное стекло, на которое берут 2-3 капли крови, и затем иглой или углом другого предметного стекла кровь распределяют, чтобы получить овал около 1 см, для ускорения высыхания препарата, его можно поместить в термостат при 35-37 0 С.
Для обнаружения цист применяют крепкий раствор Люголя, окрашивающий структуры цист в темно-коричневый цвет.
Окраска железным гематоксилином по методу Гейденгайна
Мазки после фиксации помещают в 2,5% раствор железоаммиачных квасцов на 1 час.
После трехкратного ополаскивания в воде окрашивают красителем (0,5 г гематоксилина, 10 мл 96 0 спирта и после растворения добавляют 90 мл дистиллированной воды) в течение 5-10 мин.
Промывают водой и высушивают.
Окраска незаменима в тех случаях, когда нужно выявить тончайшие детали строения ядра и цитоплазмы простейших.
Клеточные стенки растений обладают необычайной прочностью, и в процессе роста растения меняют свою структуру и состав. Основными компонентами клеточных стенок растений являются полисахариды, причем среди них преобладает целлюлоза, которая в значительной мере определяет архитектуру стенки.
Целлюлоза . Этот гомополисахарид является самым распространенным на Земле углеводом (растения образуют в год до 1011 т целлюлозы). Мономерами целлюлозы служат остатки глюкозы, соединенные в длинные цепочки (до 10 000 остатков глюкозы в каждой) с помощью b(1→4)-гликозидных связей (рис. 5.3). В такой молекуле отсутствует полная свобода вращения вокруг 1 С-О- и О- 4С-связей, и полимер приобретает конформацию, благоприятную для образования межцепочечных водородных связей, в случае, когда цепочки располагаются антипараллельно. В результате молекулы целлюлозы объединяются в микрофибриллы толщиной примерно от 10 до 25 нм. Микрофибриллы перевиваются и образуют тонкие нити, которые, в свою очередь, могут обматываться одна вокруг другой, как пряди в канате, формируя макрофибриллы. Каждая макрофибрилла имеет толщину около 0,5 мкм и может достигать в длину 6-8 мкм. Прочность макрофибрилл сопоставима с прочностью равной по толщине стальной проволоки. Кроме того, отдельные участки микрофибрилл имеют упорядоченное строение и придают клеточной стенке кристаллические свойства. Таким образом, можно отметить сложность и высокую упорядоченность целлюлозы в составе клеточных стенок, что неслучайно: этот полимер выполняет защитную и опорную функции в растении.
В таком виде полисахариды недоступны действию собственных ферментов, и целлюлоза не может использоваться растением в качестве резервного вещества. Лишь немногие организмы (некоторые бактерии, грибы, простейшие и редкие животные) обладают ферментными системами, способными расщеплять целлюлозу.
Микро- и макрофибриллы целлюлозы в клеточной стенке растений погружены в матрикс , который также состоит в основном из полисахаридов и меняет свою структуру в процессе роста растения. На начальных этапах развития матрикс состоит из пектиновых веществ , а в дальнейшем в нем появляются ксиланы и различные нейтральные полисахариды («гемицеллюлоза»). Пектиновые вещества представляют собой полимеры a-галактуроновой кислоты, в которых некоторые водородные атомы замещены метильными группами (-СН 3) (рис. 5.3). Ксиланы представляют собой полимеры ксилозы (рис. 5.3).
На более поздних этапах развития, когда происходит одревеснение клеточных стенок, в клетках откладывается лигнин - химически устойчивый полимер, содержащий большое число ароматических колец. Кроме этого, в составе клеточных стенок растений обнаруживаются небольшие количества гликопротеинов, нерастворимых липидных полимеров различного строения и восков.
Клеточные стенки некоторых растений содержат редкие полисахариды, имеющие необычное строение. Например, в стенках и межклеточном
веществе морских красных водорослей содержится сложный гетерополисахарид агар, представляющий собой смесь сульфатированных полисахаридов -агарозы и агаропектина. Агароза построена из чередующихся остатков D-галактозы и 3,6-ангидро-L-лактозы, связанных попеременно b(1→4)- и a(1→3)-связями. Агаропектин имеет более сложное строение: в его состав входят D-галактоза, 3,6-ангидрогалактоза, уроновые кислоты и сульфат. Агар используется в качестве наиболее распространенного уплотнителя для твердых сред, незаменимых в микробиологии, а также в пищевой промышленности для желирования продуктов. Следует отметить, что подавляющее большинство микроорганизмов не способно расщеплять агар, и это одно из главных его преимуществ перед другим уплотнителем питательных сред - желатиной. Агароза находит широкое применение в биохимических исследованиях: она в водной среде образует гель с большими порами, размер которых определяется ее концентрацией. Агарозные гели используются для фрак-ционирования белков и нуклеиновых кислот, а также для иммобилизации клеток.
- 20.44 КбКлеточная стенка, строение, химический состав. Видоизменения клеточной стенки.
Наличие прочной оболочки характерная черта растительной клетки, отличающая ее от клетки животной. Оболочка придает клетке определенную форму и прочность и защищает живое содержимое - протопласт и прежде всего плазмалемму, плотно прижатую к оболочке изнутри. Культивируемые на специальных питательных средах клетки высших растений, у которых ферментативным путем удаляется оболочка, всегда принимают сферическую форму. Без оболочки существование растительной клетки в обычных условиях невозможно, так как характерное для нее тургорное давление, уравновешиваемое противодавлением оболочки, неминуемо привело бы к разрыву плазмалеммы и разрушению протопласта. Совокупность клеточных оболочек в составе тела наземного растения, возвышающегося над поверхностью почвы и не имеющего внутреннего скелета, представляет собой своего рода поддерживающий остов, придающий растению механическую прочность.
Оболочка, как правило, бесцветна и прозрачна, легко пропускает солнечный свет. По ней могут передвигаться вода и растворенные низкомолекулярные вещества. У каждой клетки есть собственная оболочка; оболочки соседних клеток как бы сцементированы межклеточными веществами, образующими так называемую срединную пластинку. Вследствие этого соседние клетки оказываются отделенными друг от друга стенкой, образованной двумя оболочками и срединной пластинкой, что дает основание называть оболочку также клеточной стенкой.
Оболочка строится протопластом клетки и поэтому может расти, только находясь в контакте с ним. Очень часто оболочка сохраняется дольше протопласта (когда клетка рано отмирает), но это обычно не ведет к прекращению существования клетки как структурной единицы, ибо форма клетки благодаря прочности оболочки не изменяется. Поэтому растительные клетки и после отмирания могут выполнять важные функции передвижения растворов или механической опоры. Многие типы клеток растения – волокна, трахеиды, членики сосудов, клетки пробки – во взрослом состоянии представляют собой одни клеточные оболочки. В основном из оболочек отмерших клеток состоит древесина.
Оболочки клеток столь же разнообразны, как и сами клетки. По составу и строению оболочки часто можно судить о происхождении и функции клеток различных типов. Так, строение ископаемых растений в основном изучают путем исследования оболочек их клеток.
Первоначально к наружи от плазмалеммы возникает первичная клеточная стенка. Она состоит из полисахаридов – пектина и целлюлозы. Первичные клеточные стенки соседних клеток соединены протопектиновой срединной пластинкой. В клеточной стенке линейные очень длинные молекулы целлюлозы, состоящие из глюкозы, собраны в пучки – мицеллы, которые, в свою очередь, объединяются в фибриллы – тончайшие волоконца неопределенной длины. Целлюлоза образует многомерный каркас, который погружен в аморфный сильно обводненный матрикс из нецеллюлозных углеводов: пектинов, гемицеллюлоз и др. именно целлюлоза обусловливает прочность клеточной стенки. Микрофибриллы эластичны и по прочности на разрыв сходны со сталью. Полисахариды матрикса определяют такие свойства стенки, как высокая проницаемость для воды, растворенных мелких молекул и ионов, сильная набухаемость. Благодаря матриксу по стенкам, примыкающим друг к другу, могут передвигаться вода и вещества от клетки к клетке. Некоторые гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ.
Материал оболочек, включающий целлюлозу, широко используется промышленностью. Свойства древесины, бумаги, текстильных материалов в большой степени зависят от молекулярной структуры и химического состава оболочки, и знание их помогает улучшать технологию производства и качество продукции.
Химический состав и молекулярная организация оболочки. Клеточная оболочка построена в основном из полисахаридов, мономеры которых – сахара - связаны между собой гликозидной связью (-О-) в виде цепи. Называют полисахариды по составу образующих их мономеров с прибавлением окончания «ан». Например, полисахарид ксилан состоит из остатков сахара ксилозы, маннан - маннозы, глюкан - глюкозы и т.д.
Кроме полисахаридов, в состав оболочки могут входить белки, минеральные соли, лигнин, пигменты, липиды. Обычно оболочки пропитаны водой. Полисахариды оболочки по своей роли могут быть подразделены на скелетные вещества и вещества матрикса. Скелетным веществом оболочки высших растений является целлюлоза (клетчатка), в химическом отношении представляющая собой β-1, 4-D глюкан. Число глюкозных остатков в молекуле и, следовательно, длина самих молекул могут резко различаться в клеточных оболочках различных растений, что оказывает влияние на свойства целлюлозы. Ее нитчатые молекулы в оболочке располагаются параллельно друг к другу и группируются по нескольку десятков таким образом, что возникает вытянутая трехмерная решетка, характерная для кристаллов. Эти кристаллические группировки, в образовании которых участвуют ковалентные и водородные связи, составляют основу микрофибрилл – тончайших волокон, создающих структурный каркас оболочки, ее «арматуру».
Кристаллическое состояние молекул целлюлозы в микрофибриллах обусловливает двойное лучепреломление клеточной оболочки, наблюдаемое в поляризационном микроскопе. В микрофибриллах встречаются паракристаллические участки, т е. участки с более рыхлым параллельным, но не имеющим трехмерной ориентации расположениемI целлюлозных молекул. Степень кристалличности микрофибрилл, соотношение кристаллической и паракристаллической зон довольно сильно варьируют в зависимости от вида растения, состояния клетки и типа оболочки, что оказывает влияние на механические и физические целлюлозы как сырья. Диаметр микрофибрилл также колеблется у разных клеток (обычно в пределах 10-30 нм), длина может достигать нескольких микрометров. Микрофибриллы могут объединяться в отдельные группы или слои – макрофибриллы толщиной 0,4-0,5 мкм, видимые в световой микроскоп.
Рисунок 25
В процессе жизнедеятельности клеток, особенно в зависимости от выполнения какой-либо специализированной функции, наступают вторичные изменения клеточной стенки. Нередко они связаны с радикальными изменениями ее химического состава, структуры и физико-химических свойств.
Одревеснение. Клеточная стенка инкрустируется (пропитывается) особым веществом – лигнином, что повышает твердость, калорийность, плотность клеточной стенки и понижает ее пластичность и способность расти. Одревесневшие клеточные стенки не теряют способности пропускать воду и воздух. Протопласт их может оставаться живым, хотя обычно отмирает. Одревеснение очень широко распространено в природе. Оно обеспечивает крепость стволов и ветвей деревьев. Древесина хвойных и лиственных пород содержит целлюлозы до 50% и лигнина 20…30%. Одревесневают клеточные стенки и многих трав, особенно к концу вегетации.
Опробковение, или суберинизация. В результате обильного наслаивания в стенке клетки химически стойкого вещества – суберина наступает ее опробковение. По своей химической природе суберин близок к жирам. Он представляет собой аморфное гидрофобное соединение, которое состоит из высокополимерных насыщенных жирных кислот и оксикислот сложного состава. Суберин откладывается преимущественно в стенках клеток вторичной покровной ткани – пробки, которая является прекрасным водо- и термоизолятором. В малых количествах он накапливается в стенках клеток разнообразных тканей. При полном опробковении протопласт клетки отмирает, так как ламеллы суберина препятствуют проникновению воды и воздуха, при частичном – протопласт долгое время сохраняется.
Минерализация. В процессе жизнедеятельности стенки поверхностных клеток побегов могут накапливать минеральные вещества, особенно кремнезем и углекислый кальций.некоторые группы растений, напрмер многие виды осок, злаков, хвощей, всегда имеют сильно инкрустированные солями клетки покровной ткани – эпидермы. Причем инкрустации может подвергаться не только клеточная стенка, но и разнообразные выросты эпидермы – трихомы. Кроме того, минеральные вещества откладываются и на поверхности клеток покровной ткани – эпидермы. В результате минерализации осевые органы приобретают большую механическую прочность.
Ослизнение. Превращение целлюлозы и пектина в слизи м близкие к ним камеди, представляющие собой полимерные углеводы, которые отличаются способностью к сильному набуханию при соприкосновении с водой. Ослизнение наблюдается в клеточных сиенках кожуры семян, например у льна. Образование слизей имеет большое приспособительное значение. При прорастании семян слизь закрепляет их на определенном месте, легко поглощает и удерживает влагу, защищает семена от высыхания, улучшает водный режим всходов. Ослизнение клеточных стенок корневых влосков обеспечивает прочное склеивание их с частицами почвы.
Мацерация
– растворение
межклеточного вещества,
приводящее к разъединению
клеток. Естественная
мацерация происходит
в зрелых плодах.
Искусственно ее проводят,
например, при мочке
льна для освобождения
прядильного сырья
– групп клеток
лубяных волокон.
Описание работы
Наличие прочной оболочки характерная черта растительной клетки, отличающая ее от клетки животной. Оболочка придает клетке определенную форму и прочность и защищает живое содержимое - протопласт и прежде всего плазмалемму, плотно прижатую к оболочке изнутри. Культивируемые на специальных питательных средах клетки высших растений, у которых ферментативным путем удаляется оболочка, всегда принимают сферическую форму.