Вопрос 1. Каковы отличия в строении эукариотической и прокариотической клеток?
У прокариот нет настоящего оформленного ядра (греч. karyon - ядро). Их ДНК представляет собой одну кольцевую молекулу, свободно располагающуюся в цитоплазме и не окруженную мембраной. У прокариотических клеток отсутствуют пластиды, митохондрии, эндоплазматическая сеть, аппарат Гольджи, Лизосомы. Рибосомы есть как у прокариот, так и у эукариот (у ядерных - более крупные). Жгутик прокариотической клетки тоньше и работает по иному принципу, чем жгутик эукариотов. Эукариотическими организмами являются грибы, растения, животные - одноклеточные и многоклеточные; прокариотами - бактерии и синезеленые водоросли (цианобактерии).
Вопрос 2. Расскажите о пино - и фагоцитозе. Чем различаются эти процессы?
Мембрана клетки - подвижное образование, способное путем формирования впячиваний и выростов захватывать объекты внешней среды. Этот процесс называют эндоцитозом. Причина эндоцитоза - сложные биохимические реакции, происходящие в цитоплазме и связанные в первую очередь с изменением третичной структуры внутриклеточных белков. Если клетка захватывает каплю жидкости - это пиноцитоз, если твердую частицу - фагоцитоз. В результате образуются пиноцитарные или фагоцитарные вакуоли (мембранные пузырьки). Процесс, обратный эндоцитозу (выброс из клетки содержимого вакуолей), называют экзоцитозом.
Вопрос 3. Раскройте взаимосвязь строения и функций мембраны клеток.
Известно, что основой любой мембраны является бислой (двойной слой) фосфолипидов, в котором гидрофильные "головки" молекул (глицерин) обращены наружу, а гидрофобные остатки жирных кислот - внутрь. С липидным бислоем связаны молекулы белков, которые могут примыкать к мембране с любой из сторон, погружаться в нее или даже пронизывать. Положение клеточной мембраны на границе клетки и окружающей среды определяет ее основные функции. Прочный, эластичный, легко восстанавливающийся бислой является барьером, обеспечивающим постоянство внутриклеточной среды и предохраняющим цитоплазму от проникновения чужеродных веществ. Транспортная функция мембраны имеет избирательный характер. Мелкие незаряженные молекулы (02, N2) легко проникают непосредственно через бислой. Более крупные и/или заряженные частицы (Na+, К+, некоторые гормоны) проходят через специальные белковые поры (каналы) или транспортируются белками-переносчиками. Будучи подвижной структурой, клеточная мембрана может также осуществлять транспорт веществ путем эндо - и экзоцитоза.
Вопрос 4. Какие органоиды клетки находятся в цитоплазме?
Органоиды, расположенные в цитоплазме эукариотической клетки, можно разделить на три группы: одномембранные, двухмембранные и немембранные. К одномембранным органоидам относят эндоплазматическую сеть (гладкую и шероховатую), аппарат Гольджи, Лизосомы и вакуоли. Двухмембранные органоиды - это пластиды и митохондрии; немембранные - рибосомы, цитоскелет и клеточный центр.
Вопрос 5. Охарактеризуйте органоиды цитоплазмы и их значение в жизнедеятельности клетки.
Эндоплазматическая сеть (ЭПС) представляет собой совокупность вакуолей, каналов и трубочек. Она образует внутри цитоплазмы единую сеть, объединенную с наружной мембраной ядерной оболочки. Различают гладкую и шероховатую ЭПС. Гладкая ЭПС участвует в синтезе липидов и углеводов, а также обезвреживает токсичные вещества. На поверхности мембран шероховатой ЭПС располагаются рибосомы.
Аппарат Гольджи - одномембранный органоид, входящий в состав единой мембранной сети клетки и представляющий собой стопку плоских цистерн. В нем происходит окончательная сортировка и упаковка продуктов жизнедеятельности клетки в мембранные пузырьки (вакуоли). В числе прочего аппарат Гольджи формирует Лизосомы и обеспечивает экзоцитоз.
Лизосомы - мелкие мембранные пузырьки, которые содержат ферменты для переваривания питательных веществ. Лизосомы сливаются с эндоцитозной вакуолью, формируя пищеварительную вакуоль. Если содержимое лизосом высвобождается внутри самой клетки, наступает ее автолиз (самопереваривание клетки).
Митохондрии относят к двухмембранным органоидам. Их внешняя мембрана гладкая, а внутренняя образует складки (кристы). Митохондрии - энергетические станции клетки, их основная функция - синтез АТФ.
Пластиды представляют собой двухмембранные органоиды растительных клеток. Существует три типа пластид: хлоропласты, хромопласты и лейкопласты. Бесцветные лейкопласты запасают крахмал; зеленые хлоропласты осуществляют фотосинтез; оранжевые, желтые и красные хромопласты обеспечивают окраску плодов и цветов (привлечение опылителей и распространителей семян). Считается установленным, что в далеком прошлом митохондрии и пластиды произошли от прокариот, "проглоченных" эукариотической клеткой и вступивших с нею в симбиоз. Митохондрии и пластиды имеют кольцевую ДНК, самостоятельно синтезируют часть белков, а их рибосомы мельче эукариотических.
Рибосомы - мелкие многочисленные немембранные органоиды, образованные двумя субъединицами - большой и малой. Субъединицы состоят из белка и рибосомальной РНК. Функцией рибосом является синтез белка. Часть рибосом находится непосредственно в цитоплазме, а часть - на мембранах шероховатой ЭПС.
Клеточный центр - органоид немембранного строения клеток животных, грибов и низших растений. Состоит из двух центриолей, по форме сходных с цилиндрами и состоящих из мельчайших белковых трубочек; участвует в образовании веретена деления.
Вакуоль представляет собой мембранный пузырек, заполненный клеточным соком. Она обязательно присутствует в растительной клетке. Функция вакуоли - накопление воды, солей, питательных веществ. Здесь могут также содержаться пигменты (синие, фиолетовые) и накапливаться отходы обмена веществ.
Цитоскелет - немембранный органоид, представляющий собой белковые тяжи-трубочки, расположенные как рядом с мембраной, так и в цитоплазме. Их функция - поддержание формы клетки, обеспечение внутриклеточного транспорта веществ, а также активного движения клетки (амеба, фагоцит). Мембранные пузырьки, содержащие какие - либо вещества, могут двигаться по трубочкам цитоскелета, как по рельсам.
Как скачать бесплатное сочинение? . И ссылка на это сочинение; Эукариотическая клетка. Цитоплазма. Органоиды уже в твоих закладках.Дополнительные сочинения по данной теме
Вопрос 1. Где формируется лизосома? Лизосомы - мембранные структуры, содержащие множество активных ферментов, участвующих в расщеплении высокомолекулярных соединений: белков, липидов, углеводов. Лизосомы образуются в комплексе Гольджи, куда поступают и где накапливаются ферменты. Вопрос 2. Какова функция митохондрий? Митохондрии - клеточные структуры, покрытые двойной мембраной. На внутренней мембране, имеющей многочисленные выросты, расположено огромное количество ферментов, принимающих участие в синтезе АТФ. Следовательно, главная функция митохондрий - обеспечение клетки энергией за счет синтеза АТФ. Вопрос 3. Какие
Вопрос 1. Какие признаки примитивности прокариот по сравнению с эукариотами вы можете назвать? У прокариот отсутствуют ядро и хромосомы. Наследственная информация прокариот представлена единственной молекулой ДНК, которая замкнута в виде кольца и расположена в цитоплазме. Рибосомы у прокариот очень мелкие. Л органоидов, окруженных мембранами (эндоплазматическая сеть, аппарат Гольджи, митохондрии, пластиды), прокариоты и вовсе не имеют. В эукариотической клетке эти структуры предназначены для синтеза разнообразных соединений. В силу их отсутствия у прокариот реакции
Вопрос 1. Чем образованы стенки эндоплазматической сети и комплекса Гольджи? Стенки эндоплазматической сети и комплекса Гольджи образованы однослойной мембраной. Вопрос 2. Назовите функции эндоплазматической сети. Эндоплазматическая сеть (ЭПС) образует транспортную систему клетки. На гладкой ЭПС осуществляется синтез жиров и углеводов. На шероховатой (гранулярной) ЭПС происходит синтез белков за счет работы рибосом, прикрепленных к мембранам ЭПС. Вопрос 3. Какую функцию выполняют рибосомы? Основная функция рибосом - синтез белка. Вопрос 4. Почему большинство рибосом расположены на каналах эндоплазматической
Тест на тему «Лишайники. Водоросли» 1. Фотосинтез у водорослей происходит в: а) ядре в) цитоплазме б) хроматофорах г) хлоропластах 2. Лишайнику как единому организму присущ способ размножения: а) половой в) партеногенеза б) бесполый г) нет правильного ответа 3. Лишайники называют индикаторами чистоты воздуха, так как они: а) очищают воздух б) загрязняют воздух в) создают среду обитания для других растений и животных г) растут только в местах с чистым воздухом 4. Способны ли к активному передвижению водоросли? а) к передвижению способны все водоросли б) передвигаются
Вопрос 1. Каковы функции наружной мембраны клетки? Наружная клеточная мембрана состоит из двойного липидного слоя и молекул белков, часть которых расположена на поверхности, а некоторые пронизывают оба слоя липидов насквозь. Наружная клеточная мембрана выполняет защитную функцию, отделяя клетку от внешней среды, препятствует повреждению ее содержимого. Кроме того, наружная клеточная мембрана обеспечивает транспорт веществ внутрь клетки и из нее, позволяет клеткам взаимодействовать между собой. Вопрос 2. Какими способами различные вещества могут проникать внутрь клетки? Сквозь наружную
Вопрос 1. Какие основные этапы можно выделить в возникновении и развитии жизни на Земле? Существует много гипотез, пытающихся объяснить возникновение и развитие жизни на нашей планете. И хотя они предлагают различные подходы к решению данной проблемы, большинство из них предполагает наличие трех эволюционных этапов: химической, предбиологической и биологической эволюции. На этапе химической эволюции происходил абиогенный синтез органических полимеров. На втором этапе формировались белково-нуклеиново-липоидные комплексы (ученые называли их по-разному: коацерваты, гиперциклы, пробионы, прогеноты и
Вопрос 1. Каковы функции клеточного центра? Клеточный центр выполняет функцию формирования внутреннего скелета клетки (цитоскелета). Цитоскелет представляет собой сеть микротрубочек, пронизывающих цитоплазму, поддерживающих форму клетки, обеспечивающих движение органоидов клетки, а также работу специализированных органоидов движения - ресничек и жгутиков. Клеточный центр обеспечивает также и нормальное деление клетки. Центриоли клеточного центра расходятся к полюсам делящейся клетки и образуют веретено деления, благодаря которому из одной материнской впоследствии образуются две дочерние клетки. Центриоли представлены цилиндриками, образованными
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром; подразделяется на гиалоплазму (основное вещество цитоплазмы), органоиды (постоянные компоненты цитоплазмы) и включения (временные компоненты цитоплазмы). Химический состав цитоплазмы: основу составляет вода (60-90% всей массы цитоплазмы), различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Характерная особенность цитоплазмы эукариотической клетки — постоянное движение (циклоз ). Оно обнаруживается, прежде всего, по перемещению органоидов клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции.
Гиалоплазма (цитозоль ) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимопереходы: гель превращается в золь и наоборот.
Функции цитоплазмы:
- объединение всех компонентов клетки в единую систему,
- среда для прохождения многих биохимических и физиологических процессов,
- среда для существования и функционирования органоидов.
Клеточные оболочки
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны . Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).
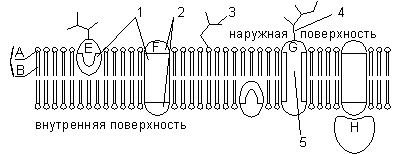
А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Функции мембран
Мембраны выполняют такие функции:
- отделение клеточного содержимого от внешней среды,
- регуляция обмена веществ между клеткой и средой,
- деление клетки на компартаменты («отсеки»),
- место локализации «ферментативных конвейеров»,
- обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
- распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ . Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na + , K + , Ca 2+ , Cl -); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na + /К + -насос (натрий-калиевый насос), 2) эндоцитоз, 3) экзоцитоз.
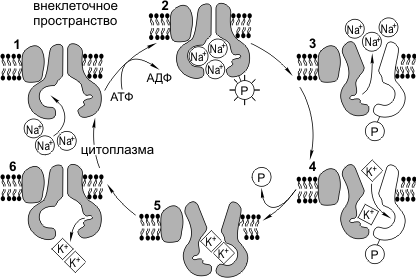
Работа Na + /К + -насоса . Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К + и Na + в цитоплазме и во внешней среде. Концентрация К + внутри клетки должна быть значительно выше, чем за ее пределами, а Na + — наоборот. Следует отметить, что Na + и К + могут свободно диффундировать через мембранные поры. Na + /К + -насос противодействует выравниванию концентраций этих ионов и активно перекачивает Na + из клетки, а K + в клетку. Na + /К + -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как K + , так и Na + . Цикл работы Na + /К + -насоса можно разделить на следующие фазы: 1) присоединение Na + с внутренней стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na + во внеклеточном пространстве, 4) присоединение K + с внешней стороны мембраны, 5) дефосфорилирование белка-насоса, 6) высвобождение K + во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл работы насос выкачивает из клетки 3Na + и закачивает 2К + .
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны. Таким способом из клеток желез внутренней секреции выводятся гормоны, у простейших — непереваренные остатки пищи.
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Перейти к лекции №7 «Эукариотическая клетка: строение и функции органоидов»
Содержание реферата
2.
Цитоплазма эукариотической клетки
4. Мембраны цитоплазмы
4.1. Химия и свойства клеточных мембран
5. Плазматическая мембрана
5.1. Рост плазматической мембраны
6.1. Транспортные функции
6.2. Рецепторные функций плазматической мембраны
6.3. Межклеточные контакты
6.4. Простой контакт. Соединение типа «замка»
6.6. Промежуточный контакт
6.7. Щелевидный контакт
Список литературы
1. Роль ученых в открытии клеточных мембран
До конца XVIII века предполагали, что клетки разделены общей стенкой и не могут
быть изолированными друг от друга. В дальнейшем развитии данной науки большую роль
сыграли исследования французского ботаника Бриссо де Мирбеля (1776-1854), в которых
впервые высказывалась идея сравнительного изучения растительных тканей. Мирбель
впервые предположил, что соприкасающиеся клетки разделены общей перегородкой - мембраной.
Немецкий естествоиспытатель Рудольфи пришел к заключению об обособленности клеток,
о наличии у них собственных мембран, окружающих их со всех сторон. Всякие сомнения
о том, что клетки могут быть выделены и изолированы, окончательно исчезли после
работы ботаника И. Молденгауера (1766-1827), из которой следовало, что клетка является
самостоятельной структурой, покрытой оболочкой. Итак, сформировалось новое представление
о клетке.
Наиболее четко его сформулировал несколько позже Ф. Мейен. Его определение таково:
«Клетка растительного организма представляет собой пространство, вполне замкнутое
вегетативной мембраной». Ф. Мейен ясно изложил этот взгляд, в начале ХIХ в. сменивший
первоначальное воззрение на клетку как на пору. Но конкретное содержание анатомии
растений по-прежнему принципиально мало чем отличалось от старого. Исследователи
продолжали системно описывать строение различных органов растений, а также детализировать
и уточнять изученное. Этот период справедливо считается периодом собирания материала,
периодом накопления многочисленных сведений о тончайшей структуре растений.
Итак, сам факт наличия клеток в различных тканях сомнения не вызывал. Но исчерпывается
ли все разнообразие структур растительных организмов только клетками? Вопрос оставался
еще не до конца ясным. Более того, большинство ученых отвечало на этот вопрос отрицательно,
указывая, наряду с клетками, на сосуды и волокна растений как на вполне своеобразные
структуры. Обобщая все знания в этой области, Мейен прямо писал, что растительные
ткани состоят из клеток, волокон и сосудов. Возникновение этих последних из метаморфизирующихся
клеток было окончательно установлено лишь несколько позднее - в 30-х гг. прошлого
века.
2. Цитоплазма эукариотической клетки
Цитоплазма
представляет собой метаболический рабочий аппарат клетки. В ней сосредоточены
общие и специальные органоиды, в цитоплазме протекают основные метаболические процессы.
В последние десятилетия достигнуты особенно большие успехи в дискретном морфофункциональном
анализе отдельных органоидов и мембранных систем метаболического аппарата цитоплазмы.
Это делает возможным представить себе работу метаболического аппарата как целостной
системы.
В современных исследованиях метаболического аппарата клетки особенно ярко проявляется
тенденция к изучению функционального значения морфологических структур - анализу
функции через структурно-биохимическую организацию этих структур. Важнейшие успехи
в этом плане достигнуты в изучении основных органоидов белкового синтеза (рибосом)
и систем, обеспечивающих энергетический обмен (митохондрий, хлоропластов и сопрягающих
мембран прокариотных клеток). Крупным обобщением в изучении метаболического аппарата
цитоплазмы являются представления о нем как о пластичной дифференцированной трехфазной
системе, состоящей из основной цитоплазмы, внутриклеточных мембран и содержимого
мембранной системы. Таким образом метаболический аппарат цитоплазмы, с одной стороны,
сложен специализированными структурами, выполняющими частные функции, а с другой
стороны, является единой целостной системой, находящейся в тесной взаимосвязи и
с поверхностным, и с ядерным аппаратами клетки.
Начало такому подходу положил еще в 60-х гг. Де Дюв в учении об экзоплазматических
мембранных системах. В дальнейшем этот подход развивался и в учении о лизосомах,
и в современных представлениях об аппарате Гольджи и эндоплазматической сети, а
также в представлениях о динамичном взаимодействии внутриклеточных мембранных систем.
Весьма перспективным, на наш взгляд, является также оформившееся в последнее время
представление об основной цитоплазме - гиалоплазме. В противовес бытовавшему раньше
взгляду на гиалоплазму как на однородную живую коллоидную систему оно трактует основную
цитоплазму как дифференцированную, гетерогенную фазу цитоплазмы, способную к формированию
сложных структур. С этим свойством гиалоплазмы мы уже частично познакомились на
примере субмембранной системы поверхностного аппарата с ее динамичными механо-химическими
и опорными структурами.
В настоящее время становится ясно, что такие примембранные системы гиалоплазмы формируются
не только в области плазматических мембран, но и в области всех мембранных, а иногда
даже и немембранных (клеточный центр) структур цитоплазмы. Эту часть цитоплазмы
предлагается выделить в особую субсистему гиалоплазмы, обозначив ее термином «цитозоль».
Естественно, что функции и морфобиохимическая организация такого цитозоля в области
шероховатой и гладкой эндоплазматической сети (ЭПС), мембран аппарата Гольджи, митохондрий,
клеточного центра, ядерной оболочки и т. д. будут различны и специфичны для тех
структур, вокруг которых формируются соответствующие участки цитозоля.
С учетом структурного и функционального разнообразия органоидов, обеспечивающих
внутриклеточный метаболизм в цитоплазме клеток, функция соответствующих каждому
конкретному органоиду и практически входящих в его состав участков цитозоля не будет
ограничиваться только опорно-сократительной. Структуры и ферменты, входящие в состав
цитозоля, специфичны для каждого органоида и принимают непосредственное участие
в обеспечении протекающих в данном органоиде биохимических процессов.
Рассмотрение цитоплазмы целесообразно начать с характеристики проблемы структурно-биохимической
организации рибосом - молекулярных машин белкового синтеза. В этом случае особенно
ярко проявляется значение структурной организации биохимических процессов в клеточных
системах и достигнуты большие успехи в ее изучении.
Второй областью биологии клетки, где структурная организация биохимических процессов
также изучена весьма детально, является анализ так называемых сопрягающих мембран
- аппарата энергетического обмена клеток. Их мы рассмотрим при характеристике митохондрий,
пластид и других структур цитоплазмы, отвечающих за энергетический обмен в клетке.
Далее мы обсудим результаты дискретного анализа основных мембран.
Собственно тело клетки и ее содержимое отделено от внешней среды или от соседних
элементов у многоклеточных организмов плазматической мембраной. Кнаружи от плазматической
мембраны, экстрацеллюлярно, расположена клеточная оболочка, или стенка, особенно
хорошо выраженная у растений и прокариотических организмов; у клеток животных она
отсутствует или выражена очень слабо. Все внутреннее содержимое клетки, за исключением
ядра, носит название цитоплазмы. Это общий термин, который подчеркивает разделение
клетки на два главных компонента: цитоплазму и ядро.
Цитоплазма эукариотических клеток
неоднородна по своему строению и составу и включает
в себя гиалоплазму, мембранные и немембранные компоненты. К мембранным компонентам
относится вакуолярная система (эндоплазматическая сеть, аппарат Гольджи, лизосомы,
вакуоли растений), мембранные органеллы (митохондрии и пластиды). К немембранным
компонентам относятся центриоли, характерные для животных клеток, и немембранные
макромолекулярные комплексы и структуры, такие как рибосомы, микротрубочки и микрофиламенты.
Такое разделение цитоплазмы на отдельные компоненты не означает их структурной и
функциональной обособленности друг от друга.
3. Гиалоплазма - внутренняя среда клетки
Термины «гиалоплазма» (от hyaline - просвечивающийся, прозрачный), «основная плазма»,
или «матрикс цитоплазмы», обозначают очень важную часть клетки, ее истинную, внутреннюю
среду. В электронном микроскопе матрикс цитоплазмы имеет вид гомогенного или тонкозернистого
вещества с низкой электронной плотностью. Из старых наблюдений за физико-химическими
свойствами цитоплазмы было получено представление о том, что гиалоплазма является
сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые
кислоты, полисахариды и др. Эта система способна переходить из золеобразного (жидкого)
состояния в гель и обратно. Так, например, при высоких гидростатических давлениях
цитоплазма
не уплотняется, а обратимо разжижается. Это явление объясняется нарушением
связей между молекулами в составе гиалоплазмы.
Благодаря электронно-микроскопическим и физико-химическим методам исследования возникли
представления об основной плазме как об организованной, упорядоченной многокомпонентной
системе. Отдельные зоны гиалоплазмы могут менять свое агрегатное состояние - в зависимости
от условий или от функциональной задачи. Так, известно, что отдельные молекулы белков-тубулинов
могут быть диспергированы в гиалоплазме, но в определенные моменты они начинают
собираться и строить длинные трубчатые структуры - микротрубочки.
Этот процесс самосборки микротрубочек обратим: при изменении условий жизни клетки
(повышение давления или изменение проницаемости мембран клетки) микротрубочки распадаются
до мономерных молекул тубулинов. Таким же образом в бесструктурной, на взгляд, гиалоплазме
могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых
молекул. Основная плазма может принимать участие в образовании клеточных мембран,
нитей и микрофиламентов. У амеб внешние слои цитоплазмы (эктоплазма) или зона цитоплазмы
в области псевдоподии практически состоят из одной гиалоплазмы, там редко обнаруживаются
мембраны, митохондрии и другие структуры. Однако после воздействия ионов, способствующих
набуханию клетки происходят быстрые и резкие перестройки этих участков цитоплазмы
- в них появляются мембранные мелкие вакуоли. Следовательно, основная плазма должна
содержать все строительные компоненты (липиды и белки), которые необходимы для образования
мембран.
Фибриллярные цитоплазматические структуры также могут возникать в результате специфических
агрегационных процессов в гиалоплазме. Если из гомогенатов клеток осадить крупные
ядра, мембранные структуры, а затем рибосомы, то оставшаяся надосадочная жидкость
будет заключать в себе основные химические компоненты гиалоплазмы. В состав гиалоплазмы
из микромолекул входят главным образом различные глобулярные белки и ферменты цитоплазматического
матрикса. Они составляют 20-25 % общего содержания белков в эукариотической клетке
.
В бактериальных клетках, бедных мембранными элементами, на долю белков гиалоплазмы
может приходиться около 50 % всех белков. К важнейшим ферментам матрикса относятся
ферменты гликолиза, ферменты метаболизма сахаров, азотистых оснований, аминокислот,
липидов и других важных соединений. В матриксе располагаются ферменты активации
аминокислот при синтезе белка, трансферные РНК. Осмотические и буферные свойства
клетки в значительной степени определяются составом и структурой гиалоплазмы.
Важнейшая роль гиалоплазмы заключается в том, что эта полужидкая среда объединяет
все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом.
Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов:
перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В гиалоплазме идет постоянный
поток ионов к плазматической мембране и от нее, к митохондриям, ядру и вакуолям.
Гиалоплазма является основным вместилищем и зоной перемещения массы молекул АТФ.
В ней же происходит отложение запасных продуктов: гликогена, жировых капель. В структурном
и морфологическом отношении гиалоплазма еще изучена плохо.
4. Мембраны цитоплазмы
Общей чертой всех мембран клетки, внешней плазматической мембраны и всех внутриклеточных
мембран и мембранных органоидов является то, что они представляют собой тонкие
(6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками), замкнутые
сами на себя. В клетке нет открытых мембран со свободными концами. Мембраны клетки
всегда ограничивают полости или участки, закрывая их со всех сторон и тем самым
отделяя содержимое таких полостей от окружающей их среды. Так, плазматическая мембрана,
покрывая всю поверхность клетки, имеющей сложную форму и многочисленные выросты,
нигде не прерывается, она замкнута. Она отделяет содержимое цитоплазмы от окружающей
клетку среды.
Внутриклеточные замкнутые мембраны образуют пузырьки - вакуоли шаровидной или уплощенной
формы. В последнем случае образуются плоские мембранные мешки, или цистерны. Часто
полости, отграниченные мембранами, имеют сложную форму, напоминающую губку или сеть,
но и в этом случае такие полости без перерывов отграничены мембраной. В подобных
вариантах мембраны также разделяют две структурные фазы цитоплазмы: гиалоплазму
от содержимого вакуолей и цистерн. Такое же свойство имеют мембраны митохондрий
и пластид: они разделяют внутреннее содержимое от межмембранных полостей и от гиалоплазмы.
Ядерная оболочка тоже может быть представлена в виде перфорированного полого двойного
мембранного мешка шаровидной формы. Мембраны ядерной оболочки разграничивают, отделяют
друг от друга кариоплазму и хромосомы от полости перинуклеарного пространства и
от гиалоплазмы. Эти общие морфологические свойства клеточных мембран определяются
их химическим составом, их липопротеидной природой.
4.1. Химия и свойства клеточных мембран
Основными химическими компонентами клеточных мембран служат липиды (40 %) и белки
(60 %), кроме того, во многих мембранах обнаружены углеводы. К липидам относится
большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность)
и растворимостью в органических растворителях и жирах (липофильность).
Состав липидов, входящих в мембраны клетки, очень разнообразен. Характерными представителями
липидов, встречающихся в клеточных мембранах, являются фосфолипиды, сфингомиелины
и из стероидных липидов - холестерин. Глицеролипиды, представляют собой сложные
эфиры трехатомного спирта, глицерина с двумя жирными кислотами и с фосфорной кислотой,
которая, в свою очередь, может быть связана с различными химическими группами (холин,
серин, инозит, этаноламин и др.).
Другой группой мембранных липидов являются сфингомиелины, где глицерин замещен аминоспиртом
сфингозином. Из липидов, относящихся к стероидам, больше всего в мембранах холестерина.
В растительных клетках холестерин не обнаружен, его там заменяют фитостерины. У
бактерий стерины отсутствуют. Характерной особенностью мембранных липидов является
разделение их молекулы на две функционально различные части: неполярные хвосты,
не несущие зарядов и состоящие из жирных кислот, и заряженные полярные головки.
Полярные головки несут на себе отрицательные заряды или могут быть нейтральными
(в случае, если они имеют одновременно положительные и отрицательные заряды).
Было обнаружено, что клеточные мембраны сильно отличаются друг от друга по составу
липидов. Так, плазматические мембраны клеток животных богаты холестерином (до
30 %), в них мало лецитина, в то время как мембраны митохондрий, наоборот, богаты
фосфолипидами и бедны холестерином. Из общего количества липидов содержание лецитина
во фракциях эндоплазматической сети составляет 60-70 % от всех фосфолипидов, в то
время как в плазматической мембране его может быть 25-35 %.
В целом для плазматической мембраны характерно высокое содержание холестерина и
сфинголипидов, а также преобладание насыщенных и мононенасыщенных жирных кислот
в составе фосфолипидов, тогда как в митохондриях, эндоплазматической сети и во многих
других цитоплазматических мембранах содержится мало холестерина и сфинголипидов
и сравнительно много полиненасыщенных жирных кислот. Видимо, в связи с этим мембраны
цитоплазмы менее жесткие, чем плазматическая мембрана, они более легкоплавки.
Количество белков в клеточных мембранах также различно. Так, в митохондриях, по
сравнению с другими мембранами, белков больше всего. Очень разнятся мембраны набором
белковых молекул. Часть из них связана с липидными головками с помощью ионных связей
и поэтому легко экстрагируются из мембран растворами солей. Другие образуют солевые
связи с полярными участками липидов через взаимодействие с ионами Mg2+ или Са2+.
Белки также экстрагируются с помощью хелатных соединений, таких как версен (ЭДТА).
Большая же часть белков взаимодействует с липидами в составе мембран на основе гидрофобных
связей.
Стало известно, что многие мембранные белки состоят как бы из двух частей: из участков,
богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными
аминокислотами (глицином, аланином, валионом, лейцином). Такие белки в липидных
слоях мембран располагаются так, что их неполярные участки как бы погружаются в
«жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная)
же часть таких белков взаимодействует с головками липидов и обращена в сторону водной
фазы. Поэтому такие белки, связанные с липидами путем гидрофобных взаимодействий,
практически не экстрагируются в водных фазах. Их можно выделить, лишь разрушая мембрану,
экстрагируя из нее липиды органическими растворителями или детергентами.
По биологической роли мембранные белки можно разделить на три группы: ферменты,
рецепторные белки и структурные белки. Набор ферментов в составе мембран может быть
очень велик и разнообразен (например, в плазматической мембране клеток печени обнаружено
не менее 24 различных ферментов).
В разных мембранах существует характерный набор ферментов. Иными словами, химическая
разнокачественность мембран определяется не только липидами, но и белками. Общим
для всех клеточных мембран является их липопротеидное строение; несходство же заключается
в количественных и качественных различиях по липидному и белковому составу. Например,
состав и количество липидов и белков будут совсем разными в мембранах эндоплазматической
сети и внутренней мембране митохондрий.
Углеводный компонент мембран представлен главным образом гликопротеинами - молекулами
белков, ковалентно (в отличие от нуклеопротеидов и липопротеидов) связанных с цепочками
углеводов.
4.2. Структура клеточных мембран
Представление о том, что в основе клеточных мембран лежит двойной липидный слой,
получено еще в 20-х гг. В 1925 г. была опубликована работа, которая указывала на
образование сплошного билипидного слоя плазматической мембраны. Несмотря на ряд
ошибок, общая идея о существовании билипидного слоя, оказалась справедливой. Выяснилось,
что если экстрагировать липиды из оболочки эритроцитов, а затем поместить их на
поверхность водного мениска, то можно рассчитать площадь, занимаемую образовавшимся
монослоем липидов. Оказалось, что эта площадь вдвое больше площади, занимаемой поверхностью
эритроцитов, из которых были экстрагированы липиды.
Ученые предположили, что в мембранах эритроцитов липиды располагаются в два слоя.
К тому же оказалось, что поверхностное натяжение клетки значительно ниже, чем поверхностное
натяжение искусственного липидного слоя. Далее исследователи обнаружили, что при
добавлении белка к маслу поверхностное натяжение снижается до величины, характерной
для поверхностного натяжения клеток. Исходя из этого заключили, что в состав мембраны
входит белок.
Так возникла гипотеза о структуре мембраны, согласно которой она представляет собой
трехслойный «сэндвич» (белок-липид-белок). Липидный слой в центре - это бимолекулярный
слой, в котором гидрофобные концы молекул липидов направлены друг к другу, а гидрофильные
- к белковым слоям, лежащим по обе стороны от липидного слоя. Эта гипотеза получила
прямые подтверждения, когда стало возможным использовать для изучения мембран клетки
электронную микроскопию. Благодаря ей удалось увидеть особенно хорошо на мембранах
миелиновой оболочки нервов, что мембрана действительно трехслойная: два темных слоя
по 2,5 нм каждый и более широкий светлый слой между ними.
Трехслойность клеточных мембран удается показать в электронном микроскопе с помощью
метода замораживания-скалывания, что особенно важно, т. к. в этом случае есть возможность
наблюдать специфические ферменты или даже ферментные системы. В начале 60-х гг.
Робертсоном была сформулирована гипотеза о единой, унитарной «элементарной» биологической
мембране. Вопрос о возникновении, образовании мембран в живой клетке еще недостаточно
изучен. Известно, что количество и величина мембранных элементов в клетках может
возрастать и уменьшаться, т. е. можно говорить о мембраногенезе и о разрушении мембран.
5. Плазматическая мембрана
Плазматическая мембрана, или плазмалемма, среди различных клеточных мембран занимает
особое место. Это поверхностная периферическая структура, ограничивающая клетку
снаружи, что обусловливает ее непосредственную связь с внеклеточной средой, а следовательно,
со всеми веществами и стимулами, воздействующими на клетку. В химическом отношении
плазматическая мембрана представляет собой липопротеиновый комплекс.
Главные принципы организации мембран были уже обсуждены в предыдущем разделе. Она
имеет толщину около 10 нм и представляет собой самую толстую из клеточных мембран.
Основными компонентами плазматических мембран являются липиды (около 40 %), белки
(более 60 %) и углеводы (около 1 %). Как уже указывалось, плазмалемма по сравнению
с другими мембранами более богата холестерином, в ее фосфолипидах преобладают насыщенные
жирные кислоты.
Состав белков, входящих в плазматическую мембрану, очень разнообразен, что определяется
многообразием ее функциональных нагрузок. Среди ферментов обнаружены 5"- нуклеотидаза,
Mg-зависимая АТФаза, активируемая ионами Na и К, щелочная и кислая фосфатаза, аденилатциклаза,
РНКаза, обнаружена протеолитическая активность. Ферменты дыхательной цепи и гликолитические
ферменты в плазматической мембране эукариотических клеток полностью отсутствуют.
Зато у прокариотических клеток именно в составе плазматической мембраны локализованы
элементы цепи переноса электронов и окислительного фосфорилирования.
Необходимо подчеркнуть, что часть белков, обнаруживаемых во фракциях плазматической
мембраны, может быть и не связана непосредственно с билипидным слоем. Многие ферменты,
обнаруживающие свою активность во фракциях плазматической мембраны, могут быть локализованы
в гликокаликсе, гликопротеиновом комплексе, ассоциированном с плазматической мембраной.
В состав именно гликокаликса входят различные углеводы, которые могут составлять
около 1 % от сухого веса мембран.
Углеводы образуют длинные ветвящиеся цепочки полисахаридов, связанных с белками
слоя, часто определяющего форму клетки. К плазматической мембране здесь изнутри
могут примыкать мембранные мешочки; в этом случае у поверхности клеток имеются три
мембранных слоя: собственно плазматическая мембрана и две мембраны пелликулярных
альвеол.
У инфузории туфельки пелликула образует утолщения, располагающиеся в виде шестиугольников,
в центре которых выходят реснички. Жесткость пелликулярных образований может быть
связана также с элементами цитоплазмы, подстилающими плазматическую мембрану, с
кортикальным слоем. Так, в гребнях пелликулы эвглены вблизи мембраны обнаруживаются,
кроме мембранных вакуолей, параллельные пучки микротрубочек и микрофиламентов. Такая
фибриллярная периферическая арматура вместе со складчатой многослойной мембранной
периферией создает жесткую структуру пелликулы.
5.1. Рост плазматической мембраны
После деления клеток происходит увеличение объемов растущих дочерних клеток, и в
связи с этим происходит рост клеточной поверхности, увеличение площади плазматической
мембраны. Но это не единственный пример быстрого роста объема и поверхности. Поверхность
быстро растущих клеток в тычиночных нитях злаков может за 1 ч увеличиться в 65 раз,
т. е. каждую минуту плазмалемма нарастает на ее первоначальную величину. Такую большую
скорость роста плазматической мембраны можно объяснить только тем, что происходит
быстрое встраивание готовых мембранных предшественников, липидов и белков, в старый
липопротеидный слой. Подобный способ роста вполне возможен, т. к. известно, что
в клетке происходит постоянная перестройка мембран - замена одних липидов на другие,
изменение их количества, изменение состава белков и т. д. Следовательно, мембраны
представляют собой очень лабильные соединения.
Но описан и другой случай образования новой плазматической мембраны. Плазмодий миксомицета
Physarum pоlycephalum протыкали стеклянным капилляром и полученный цилиндр плазмы,
лишенный плазматической мембраны, фиксировали сразу же и далее через 1, 2, 3 и
6 с после начала эксперимента, затем изучали в электронном микроскопе. Для цитоплазмы
этого гриба характерно наличие большого числа мелких пузырьков, содержащих слизь,
которая покрывает весь плазмодий снаружи. Можно было видеть, что такие пузырьки,
рассеянные в основной плазме, собирались у незащищенного мембраной края плазмы,
образуя уплощенные вакуоли, располагающиеся параллельно наружной поверхности обнаженного
кусочка. Такие плоские вакуоли сливались и образовывали сплошную мембрану - новую
плазматическую мембрану, изолирующую компоненты плазмы от внешней среды.
По-видимому, такой процесс обновления плазматической мембраны происходит не только
при больших ее повреждениях, но и постоянно, в процессе жизнедеятельности клетки.
Так, в результате экзоцитоза мембраны вакуолей, сливаясь с плазматической мембраной,
как бы встраиваются в нее, тем самым увеличивая общую площадь поверхности клетки.
Наблюдения за процессом секреции (одна из форм экзоцитоза) привели многих исследователей
к представлению о том, что в клетке постоянно имеются потоки мембранных элементов:
от плазматической мембраны внутрь цитоплазмы (эндоцитоз) и, наоборот, ток мембранных
структур из цитоплазмы к поверхности клетки (экзоцитоз). В этом круговороте мембран
ведущая роль отводится системе мембранных вакуолей аппарата Гольджи.
6. Функции плазматической мембраны
Плазматическая мембрана выполняет целый ряд важнейших клеточных функций, ведущими
из которых являются функция разграничения веществ цитоплазмы от внешней среды и
функции транспорта различных веществ как внутрь клетки, так и из нее. K транспортным
функциям относятся пассивный транспорт воды, ионов, низкомолекулярных веществ и
активный перенос этих веществ против градиентов концентрации, а также разные формы
транспорта высокомолекулярных соединений и комплексов (эндоцитоз).
Плазматическая мембрана, кроме того, участвует в выведении из клеток продуктов,
образованных в ней. Она же участвует в процессах внеклеточного расщепления биополимеров.
На поверхности плазмалеммы располагаются различные рецепторные структуры, специфически
взаимодействующие с внеклеточными факторами и с соседними клетками. Тем самым клеточная
мембрана участвует в передаче сигналов внутрь клетки. Плазматическая мембрана принимает
участие в межклеточных взаимодействиях в многоклеточных организмах. Отдельные участки
плазматической мембраны в специализированных клетках животных принимают участие
в построении специальных отростков клетки, таких как микроворсинки, реснички, рецепторные
выросты и др. Наконец, плазматическая мембрана играет важную роль при делении клетки.
Это краткое перечисление функций плазматической мембраны далеко от полноты, однако
из него видна чрезвычайная важность этой периферической структуры любой клетки,
будь то клетка бактерий или эукариотических организмов.
6.1. Транспортные функции
Плазматическая мембрана, как и другие клеточные липопротеидные мембраны, полупроницаема.
Это значит, что через нее с различной скоростью проходят разные молекулы, и чем
больше размер молекул, тем меньше скорость их прохождения через мембрану. Это свойство
определяет плазматическую мембрану как осмотический барьер.
Максимальной проникающей способностью обладает вода и растворенные в ней газы, значительно
медленнее проникают сквозь мембрану ионы (примерно в 104 раза медленнее). Поэтому
если клетку, например эритроцит, поместить в среду, где концентрация солей будет
ниже, чем в клетке (гипотония), то вода снаружи устремится внутрь клетки, что приведет
к увеличению ее объема и к разрыву плазматической мембраны (гипотонический «шок»).
Наоборот, при помещении эритроцита в растворы солей более высокой концентрации,
чем в клетке, произойдет выход воды из нее во внешнюю среду. Клетка при этом сморщится,
уменьшится в объеме. Такой пассивный транспорт воды из клетки и в клетку все же
идет с низкой скоростью. Следовательно, в клеточной мембране, в ее липопротеидном
слое существуют специальные поры для проникновения воды и ионов.
6.2. Рецепторные функции плазматической мембраны
Эти функции связаны с локализацией на плазматической мембране специальных структур,
нацеленных на специфическое узнавание химических или физических факторов. Клеточная
поверхность обладает большим набором компонентов - рецепторов, определяющих возможность
специфических реакций с различными агентами. В качестве таких рецепторов на поверхности
клетки могут выступать белки мембраны или элементы гликокаликса (полисахариды, гликопротеиды).
Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны
по поверхности клетки или собраны в небольшие зоны. Так, на поверхности бактериальных
клеток или клеток животных существует ограниченное число мест, с которыми могут
связываться вирусные частицы. Интересно, что разные вирусы связываются обычно с
разными участками клеточной периферии, причем одна и та же бактериальная клетка
может иметь на поверхности несколько разных типов рецепторов.
Применение различных иммуннологических методов показало, что на поверхности клетки
локализуются антигенно активные компоненты, специфически реагирующие с антителами
или иммунными клетками. Большая часть таких поверхностных антигенов содержит углеводные
группировки (гликолипиды, гликопротеиды). Антигенными компонентами бактериальной
стенки также чаще всего являются липополисахариды и гликолипиды. На поверхности
клетки, часто прямо в ее липопротеидной мембране, располагаются различные рецепторы,
связывающие физиологически активные вещества, такие как разные гормоны, медиаторы
и пр.
Разные клетки животных организмов могут обладать разными наборами рецепторов или
же разной чувствительностью одного и того же рецептора. Роль многих клеточных рецепторов
заключается в передаче сигналов с поверхности внутрь клетки. В настоящее время хорошо
изучена система передачи стимула клеткам с помощью некоторых гормонов, в состав
которых входят пептидные цепочки. Обнаружено, что эти гормоны связываются со специфическими
рецепторами на поверхности плазматической мембраны клетки.
Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к
созданию очень сложной системы маркеров, позволяющих отличать свои клетки (той же
особи или того же вида) от чужих. Сходные клетки вступают друг с другом во взаимодействия,
приводящие к слипанию поверхностей (конъюгация у простейших и бактерий, образование
тканевых клеточных комплексов). При этом клетки, отличающиеся набором детерминантных
маркеров или не воспринимающие их, либо исключаются из такого взаимодействия, либо
у высших животных уничтожаются в результате иммуннологических реакций.
С плазматической мембраной связана локализация специфических рецепторов, реагирующих
на физические факторы. Так, в плазматической мембране или в ее производных у фотосинтетических
бактерий и сине-зеленых водорослей локализованы белки - рецепторы (хлорофиллы),
взаимодействующие с квантами света. В плазматической мембране светочувствительных
клеток животных расположена специальная система фоторецепторных белков (родопсин),
с помощью которых световой сигнал превращается в химический, что, в свою очередь,
приводит к генерации электрического импульса.
6.3. Межклеточные контакты
Плазматическая мембрана, как уже говорилось, принимает активное участие в межклеточных
контактах, связанных с конъюгацией одноклеточных организмов. У многоклеточных организмов
за счет межклеточных взаимодействий образуются сложные клеточные ансамбли, поддержание
которых может осуществляться разными путями. В зародышевых, эмбриональных тканях,
особенно на ранних стадиях развития, клетки остаются в связи друг с другом за счет
способности их поверхностей слипаться. Это свойство соединения клеток может определяться
свойствами их поверхности, которые специфически взаимодействуют друг с другом.
Механизм этих связей еще недостаточно изучен, но, вероятнее всего, он обеспечивается
взаимодействием между липопротеидами и гликокаликсом плазматических мембран. При
таком межклеточном взаимодействии эмбриональных клеток между плазматическими мембранами
всегда остается щель шириной около 20 нм, заполненная гликокаликсом. Обработка ткани
ферментами, нарушающими целостность гликокаликса (муказы, действующие гидролитически
на муцины, мукополисахариды) или повреждающими плазматическую мембрану (протеазы),
приводит к обособлению клеток друг от друга, к их диссоциации. Однако если удалить
фактор диссоциации, то клетки могут снова собираться, реагрегировать.
Так, можно диссоциировать клетки разных по окраске губок - оранжевых и желтых. Оказалось,
что в смеси этих клеток образуются два типа агрегатов: состоящие только из желтых
и только из оранжевых клеток. При этом смешанные клеточные суспензии самоорганизуются,
восстанавливая исходную многоклеточную структуру. Сходные результаты были получены
с суспензиями разделенных клеток эмбрионов амфибий; в этом случае происходит избирательное
пространственное обособление клеток эктодермы от энтодермы и от мезенхимы.
Более того, если для реагрегации используются ткани поздних стадий развития зародышей,
то в пробирке самостоятельно собираются различные клеточные ансамбли, обладающие
тканевой и органной специфичностью, образуются эпителиальные агрегаты, сходные с
почечными канальцами, и т. д. Соединения между клетками в составе тканей и органов
многоклеточных животных организмов могут образовываться сложными специальными структурами,
которые называют собственно межклеточными контактами. Эти структурированные межклеточные
контакты особенно выражены в покровных пограничных тканях, в эпителиях.
Возможно, что первичное обособление пласта клеток, связанных друг с другом с помощью
специальных структурированных межклеточных контактов, в филогенезе животных обеспечило
образование и развитие тканей и органов. Благодаря электронной микроскопии накопилось
множество данных об ультраструктуре этих соединительных образований. К сожалению,
их биохимический состав и молекулярная структура еще недостаточно точно изучены.
Изучая соединения клеток в эпителиальных пластах, можно обнаружить следующие структуры,
связывающие клетки друг с другом: простой контакт, соединение типа «замка», плотный
контакт, промежуточный контакт, или зона слипания, десмосомный контакт, щелевидный
контакт.
6.4. Простой контакт. соединение типа «замка»
Простой контакт встречается среди большинства прилежащих друг к другу клеток различного
происхождения. Большая часть поверхности контактирующих клеток эпителия также связана
с помощью простого контакта, где плазматические мембраны соприкасающихся клеток
разделены пространством 15-20 нм. Как уже говорилось, это пространство представляет
собой надмембранные компоненты клеточных поверхностей. Ширина щели между мембранами
клеток может быть и больше 20 нм, образуя расширения, полости, но не меньше 10 нм.
Со стороны цитоплазмы к этой зоне плазматической мембраны не примыкают никакие специальные
дополнительные структуры.
Соединение типа «замка» являет собой выпячивание плазматической мембраны одной клетки
в инвагинат (впячивание) другой. На срезе такой тип соединения напоминает плотничий
шов. Межмембранное пространство и цитоплазма в зоне «замков» имеют те же характеристики,
что и в областях простого контакта.
6.5. Плотный замыкающий контакт
Он представляет собой зону, где внешние слои двух плазматических мембран максимально
сближены. Часто видна трехслойность мембраны в этом контакте: два внешних осмофильных
слоя обеих мембран сливаются в общий слой толщиной в 2-3 нм. Слияние мембран происходит
не по всей площади плотного контакта, а представляет собой ряд точечных соединений.
Со стороны цитоплазмы в этой зоне часто встречаются многочисленные фибриллы около
8 нм в диаметре, располагающиеся параллельно поверхности плазмалеммы.
Такого типа контакты были обнаружены между фибробластами в культуре ткани, между
эмбриональным эпителием и клетками мезенхимы.
6.6. Промежуточный контакт (или зона слипания)
В этом месте межмембранное расстояние несколько расширено (до 25-30 нм) и, в отличие от простого контакта, заполнено плотным содержимым, вероятнее всего, белковой природы.
6.7. Щелевидный контакт
Он представляет собой область протяженностью 0,5-3 мкм, где плазматические мембраны разделены промежутком в 2-3 нм, что после осмирования придает всей этой структуре семислойный вид. Со стороны цитоплазмы никаких специальных примембранных структур не обнаруживается. Этот тип соединения встречается во всех типах тканей. Функциональная роль щелевидного контакта заключается, видимо, в передаче ионов и молекул от клетки к клетке. Например, в сердечной мышце передача потенциала действия от клетки к клетке происходит через тип контакта, где ионы могут свободно переходить по межклеточным соединениям. Поддержание такой ионной связи между клетками зависит от энергии, получаемой благодаря окислительному фосфорилированию.
Список литературы
1. Вермель Е. М. История учения о клетке. М.: Наука, 1970. 259 с.
2. Гуляев Г. В., Мальченко В. В. Словарь терминов по генетике, цитологии, селекции,
семеноводству и семеноведению. М.: Россельхозиздат, 1983. 240 с.
3. Де Дюв К. Путешествие в мир живой клетки. М.: Мир, 1987. 253 с.
4. Заварзин А. А., Харазова А. Д. Основы общей цитологии: Учебное пособие. Л.: Изд-во
Ленингр. ун-та, 1982. 240 с.
5. От молекул до человека./ Под. ред. Наумова Н. П. М.: Просвещение, 1973. 480 с.
6. Паушева З. П. Практикум по цитологии растений. М.: Агропромиздат, 1988. 271 с.
7. Трошин А. С., Браун А. Д., Вахтин Ю. Б., Жилкин Л. Н., Суханова К. М. Цитология.
М.: Просвещение, 1970. 304 с.
8. Ченцов Ю. С. Общая цитология. М.: МГУ, 1978. 344 с.
Реферат плюс
Плазмалемма (клеточная оболочка) животных клеток образована мембраной, покрытой снаружи слоем гликокаликса толщиной 10-20 нм.Плазмалемма выполняет отграничивающую, барьерную, транспортную и рецепторную функции. Благодаря свойству избирательной проницаемости плазмалемма регулирует химический состав внутренней среды клетки. В плазмалемме размещены молекулы рецепторов, которые избирательно распознают определенные биологически активные вещества (гормоны). В пластах и слоях соседние клетки удерживаются благодаря наличию разного вида контактов, которые представлены участками плазмалеммы, имеющими особое строение. Изнутри к мембране примыкает кортикальный (корковый) слой цитоплазмы толщиной 0,1-0,5 мкм.
Цитоплазма. В цитоплазме находится целый ряд оформленных структур, имеющих закономерные особенности строения и поведения в разные периоды жизнедеятельности клетки. Каждая из этих структур несёт определенную функцию. Отсюда возникло сопоставление их с органами целого организма, в связи с чем они получили названиеорганеллы , илиорганоиды . В цитоплазме откладываются различные вещества - включения (гликоген, капли жира, пигменты). Цитоплазма пронизана мембранами эндоплазматической сети .
Эндоплазматическая сеть (ЭДС) . Эндоплазматическая сеть - это разветвлённая сеть каналов и полостей в цитоплазме клетки, образованная мембранами. На мембранах каналов находятся многочисленные ферменты, обеспечивающие жизнедеятельность клетки. Различают 2 вида мембран ЭДС - гладкие и шероховатые. На мембранах гладкой эндоплазматической сети находятся ферментные системы, участвующие в жировом и углеводном обмене. Основная функцияшероховатой эндоплазматической сети - синтез белков, который осуществляется в рибосомах, прикрепленных к мембранам.Эндоплазматическая сеть - это общая внутриклеточная циркуляционная система, по каналам которой транспортируются вещества внутри клетки и из клетки в клетку.
Рибосомы осуществляют функцию синтеза белков. Рибосомы представляют собой сферические частицы диаметром 15-35нм, состоящие из 2 субъединиц неравных размеров и содержащие примерно равное количество белков иРНК. Рибосомы в цитоплазме располагаются или прикрепляются к наружной поверхности мембран эндоплазматической сети. В зависимости от типа синтезируемого белка рибосомы могут объединяться в комплексы -полирибосомы . Рибосомы присутствуют во всех типах клеток.
Комплекс Гольджи. Основным структурным элементомкомплекса Гольджи является гладкая мембрана, которая образует пакеты уплощенных цистерн, или крупные вакуоли, или мелкие пузырьки. Цистерны комплекса Гольджи соединены с каналами эндоплазматической сети. Синтезированные на мембранах эндоплазматической сети белки, полисахариды, жиры транспортируются к комплексу, конденсируются внутри его структур и "упаковываются" в виде секрета, готового к выделению, либо используются в самой клетке в процессе её жизнедеятельности.
Митохондрии. Всеобщее распространение митохондрий в животном и растительном мире указывают на важную роль, которуюмитохондрии играют в клетке.Митохондрии имеют форму сферических, овальных и цилиндрических телец, могут быть нитевидной формы. Размеры митохондрий 0,2-1мкм в диаметре, до 5-7мкм в длину. Длина нитевидных форм достигает 15-20мкм. Количество митохондрий в клетках различных тканей неодинаково, их больше там, где интенсивны синтетические процессы (печень) или велики затраты энергии. Стенка митохондрий состоит из 2-х мембран - наружной и внутренней. Наружная мембрана гладкая, а от внутренней внутрь органоида отходят перегородки - гребни, или кристы. На мембранах крист находятся многочисленные ферменты, участвующие в энергетическом обмене.Основная функция митохондрий - синтезАТФ.
Лизосомы - небольшие овальные тельца диаметром около 0,4мкм, окруженные одной трехслойной мембраной. В лизосомах находится около 30 ферментов, способных расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и др. вещества. Расщепление веществ с помощью ферментов называетсялизисом , поэтому и органоид названлизосомой . Полагают, что лизосомы образуются из структур комплекса Гольджи либо непосредственно из эндоплазматической сети.Функции лизосом : внутриклеточное переваривание пищевых веществ, разрушение структуры самой клетки при её отмирании в ходе эмбрионального развития, когда происходит замена зародышевых тканей на постоянные, и в ряде других случаев.
Центриоли. Клеточный центр состоит из 2-х очень маленьких телец цилиндрической формы, расположенных под прямым углом друг к другу. Эти тельца называютсяцентриолями . Стенка центриоли состоит из 9-ти пар микротрубочек. Центриоли способны к самосборке и относятся к самовоспроизводящимся органоидам цитоплазмы. Центриоли играют важную роль в клеточном делении: от них начинается рост микротрубочек, образующих веретено деления.
Ядро. Ядро - важнейшая составная часть клетки. Оно содержит молекулыДНКи поэтому выполняет две главные функции: 1) хранение и воспроизведение генетической информации, 2) регуляция процессов обмена веществ, протекающих в клетке. Клетка утратившаяядро , не может существовать. Ядро также неспособно к самостоятельному существованию. Большинство клеток имеет одно ядро, но можно наблюдать 2-3ядра в одной клетке, например в клетках печени. Известны многоядерные клетки с числом ядер в несколько десятков. Формы ядер зависят от формы клетки. Ядра бывают шаровидные, многолопастные. Ядро окружено оболочкой, состоящей из двух мембран, имеющих обычное трёхслойное строение. Наружная ядерная мембрана покрыта рибосомами, внутренняя мембрана гладкая. Главную роль в жизнедеятельности ядра играет обмен веществ между ядром и цитоплазмой. Содержимое ядра включает ядерный сок, или кариоплазму, хроматин и ядрышко. В состав ядерного сока входят различные белки, в том числе большинство ферментов ядра, свободные нуклеотиды, аминокислоты, продукты деятельности ядрышка и хроматина, перемещающиеся из ядра в цитоплазму.Хроматин содержит ДНК, белки и представляет собой спирализованные и уплотненные участки хромосом.Ядрышко представляет собой плотное округлое тельце, располагающееся в ядерном соке. Число ядрышек колеблется от 1 до 5-7 и более. Ядрышки есть только в неделящихся ядрах, во время митоза они исчезают, а после завершения деление образуются вновь. Ядрышко не является самостоятельным органоидом клетки, оно лишено мембраны и образуется вокруг участка хромосомы, в котором закодирована структура рРНК. В ядрышке формируются рибосомы, которые затем перемещаются в цитоплазму.Хроматином называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличные по форме от ядрышка.
2) 1. Клеточная теория
Клеточная теория – это обобщенные представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточной теории предшествовал довольно длительный период накопления наблюдений над строением различных одноклеточных и многоклеточных организмов растений и животных. Этот период был связан с развитием применения и усовершенствования различных оптических методов исследований.
Роберт Гук первым наблюдал с помощью увеличительных линз подразделение тканей пробки на «ячейки», или «клетки». Его описания послужили толчком для появления систематических исследований анатомии растений, которые подтвердили наблюдения Роберта Гука и показали, что разнообразные части растений состоят из тесно расположенных «пузырьков», или «мешочков». Позднее А. Левенгук открыл мир одноклеточных организмов и впервые увидел клетки животных. Позднее клетки животных были описаны Ф. Фонтана; но эти и другие многочисленные исследования не привели в то время к пониманию универсальности клеточного строения, к четким представлениям о том, что же являет собой клетка. Прогресс в изучении микроанатомии и клетки связан с развитие микроскопирования в XIX в. К этому времени изменились представления о строении клеток: главным в организации клетки стала считаться не клеточная стенка, а собственно ее содержимое, протоплазма. В протоплазме был открыт постоянный компонент клетки – ядро. Все эти многочисленные наблюдения позволили Т. Шванну в 1838 г. сделать ряд обобщений. Он показал, что клетки растений и животных принципиально сходны между собой. «Заслуга Т. Шванна заключалась не в том, что он открыл клетки как таковые, а в том, что он научил исследователей понимать их значение». Дальнейшее развитие эти представления получили в работах Р. Вирхова. Создание клеточной теории стало важнейшим событием в биологии, одним из решающих доказательств единства всей живой природы. Клеточная теория оказала значительное влияние на развитие биологии, послужили главным фундаментом для развития таких дисциплин, как эмбриология, гистология и физиология. Она дала основы для понимания жизни, для объяснения родственной взаимосвязи организмов, для понимания индивидуального развития.
Основные положения клеточной теории сохранили свое значение и на сегодняшний день, хотя более чем за сто пятьдесят лет были получены новые сведения о структуре, жизнедеятельности и развитии клеток. В настоящее время клеточная теория постулирует:
1) Клетка – элементарная единица живого: – вне клетки нет жизни.
2) Клетка – единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование, состоящее из сопряженных функциональных единиц – органелл или органоидов.
3) Клетки сходны – гомологичны – по строению и по основным свойствам.
4) Клетки увеличиваются в числе путем деления исходной клетки после удвоения ее генетического материала: клетка от клетки.
5) Многоклеточный организм представляет собой новую систему, сложный ансамбль из множества клеток, объединенных и интегрированных в системы тканей и органов, связанных друг с другом с помощью химических факторов, гуморальных и нервных.
6) Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией различных генов, что приводит к их морфологическому и функциональному разнообразию – к дифференцировке.
Представление о клетке как о самостоятельной жизнедеятельной единице было дано еще в работах Т. Шванна. Р. Вирхов также считал, что каждая клетка несет в себе полную характеристику жизни: «Клетка есть последний морфологический элемент всех живых тел, и мы не имеем права искать настоящей жизнедеятельности вне ее».
Современная наука полностью доказала это положение. В популярной литературе клетку часто называют «атомом жизни», «квантом жизни», подчеркивая тем самым, что клетка – это наименьшая единица живого, вне которой нет жизни.
Такая общая характеристика клетки должна в свою очередь опираться на определение живого – что такое живое, что такое жизнь. Очень трудно дать окончательное определение живого, жизни.
М.В. Волькенштейн дает следующее определение жизни: «живые организмы представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, важнейшими функционирующими веществами которых являются белки и нуклеиновые кислоты». Живому свойствен ряд совокупных признаков, таких, как способность к воспроизведению, использование и трансформация энергии, метаболизм, чувствительность, изменчивость. И такую совокупность этих признаков можно обнаружить на клеточном уровне. Нет меньшей единицы живого, чем клетка. Мы можем выделить из клетки отдельные ее компоненты или даже молекулы и убедиться, что многие из них обладают специфическими функциональными особенностями. Так, выделенные актомиозиновые фибриллы могут сокращаться в ответ на добавление АТФ; вне клетки прекрасно «работают» многие ферменты, участвующие в синтезе или распаде сложных биоорганических молекул; выделенные рибосомы в присутствии необходимых факторов могут синтезировать белок, разработаны неклеточные системы ферментативного синтеза нуклеиновых кислот и т.д. Можно ли считать все эти клеточные компоненты, структуры, ферменты, молекулы живыми? Можно ли считать живым актомиозиновый комплекс? Думается, что нет, хотя бы потому, что он обладает лишь частью набора свойств живого. То же относится и к остальным примерам. Только клетка как таковая является наименьшей единицей, обладающей всеми вместе взятыми свойствами, отвечающими определению «живое».
3) Основу поверхностного аппарата клеток (ПАК) составляет наружная клеточная мембрана, или плазмалемма. Кроме плазмалеммы в ПАК имеется надмембранный комплекс, а у эукариот - и субмембранный комплекс. Основными биохимическими компонентами плазмалеммы (от греч. плазма - образование и лемма - оболочка, корка) являются липиды и белки. Их количественное соотношение у большинства эукариот составляет 1:1, а у прокариот в плазмалемме преобладают белки. В наружной клеточной мембране обнаруживается небольшое количество углеводов и могут встречаться жироподобные соединения (у млекопитающих - холестерол, жирорастворимые витамины). В 1925 г. Е. Гортер и Ф. Грендел (Голландия) предположили, что основу мембраны составляет двойной слой липидов - билипидный слой. В 1935 г. Дж.Даниэли и Г.Даусон предложили первую пространственную модель организации мембран, получившую название "сэндвич", или "бутербродная " модель. По их мнению, основой мембраны является билипидный слой, а обе поверхности слоя покрыты сплошными слоями белков. Дальнейшее изучение клеточных мембран, включая плазмалемму, показало, что почти во всех случаях они имеют сходное строение. В 1972 г. С.Зингер и Г.Николсон (США) сформулировали представление о жидкостно-мозаичном строении клеточных мембран (рис.). Согласно этой модели, основу мембран составляет билипидный слой, но белки в нем расположены отдельными молекулами и комплексами, т.е. мозаично (от франц. mosaique - мозаика; изображение, составленное из отдельных кусков). В частности, молекулы интегральных (от лат. интегер - целый) белков могут пересекать билипидный слой, полуинткгральных - частично погружаться в него, а периферических (от греч. периферия - окружность) - располагаться на его поверхности (рис.). Современная молекулярная биология подтвердила справедливость жидкостно-мозаичной модели, хотя были обнаружены и другие варианты клеточных мембран. В частности, у архебактерий основу мембраны составляет монослой сложного по строению липида, а некоторые бактерии содержат в цитоплазме мембранные пузырьки, стенки которых представлены белковым монослоем. Надмембранный комплекс поверхностного аппарата клеток характеризуется многообразием строения (рис.). У прокариот надмембранный комплекс в большинстве случаев представлен клеточной стенкой различной толщины, основу которой составляет сложный гликопротеин муреин (у архебактерий - псевдомуреин). У целого ряда эубактерий наружная часть надмембранного комплекса состоит из еще одной мембраны с большим содержанием липополисахаридов.У эукариот универсальным компонентом надмембранного комплекса являются углеводы - компоненты гликолипидов и гликопротеинов плазмалеммы. Благодаря этому его исходно называли гликокаликсом (от греч. гликос - сладкий, углевод и лат. каллум - толстая кожа, оболочка). Кроме углеводов, в состав гликокаликса относят периферические белки над билипидным слоем. Более сложные варианты надмембранного комплекса встречаются у растений (клеточная стенка из целлюлозы), грибов и членистоногих (наружный покров из хитина). Субмембранный (от лат. суб - под) комплекс характерен только для эукариотических клеток. Он состоит из разнообразных белковых нитевидных структур: тонких фибрилл (от лат. фибрилла - волоконце, ниточка), микрофибрилл (от греч. микрос - малый), скелетных (от греч. скелетон - высушенное) фибрилл и микротрубочек. Они связаны друг с другом белками и формируют опорно-сократительный аппарат клетки. Субмембранный комплекс взаимодействует с белками плазмалеммы, которые, в свою очередь, связаны с надмембранным комплексом. В результате ПАК представляет собой структурно целостную систему. Это позволяет ему выполнять важные для клетки функции: изолирующую, транспортную, каталитическую, рецепторно-сигнальную и контактную.
4) В мембранах содержатся также гликолипиды и холестерол. Гликолипиды - это липиды с присоединенными к ним углеводами. Как и у фосфолипидов, угликолипидов имеются полярные головы и неполярные хвосты. Холестерол близок к липидам; в его молекуле также имеется полярная часть.
1.Создателем клеточной теории является:
Р.Вирхов
В.И. Вернадский
Т.Шванн
2.Клеточная теория была создана в:
1839 году
3.Группа мембранных органелл цитоплазмы эукариотических клеток:
Рибосомы, митохондрии, пластиды, лизосомы
сферосомы, вакуоли, пластиды
Микротрубочки, микрофиламенты, рибосомы, центросома
ЭПС, рибосомы, лизосомы, аппарат Гольджи
митохондрии, ЭПС, аппарат Гольджи
4.Органеллы общего назначения:
Миофибриллы
рибосомы, митохондрии
ЭПС, аппарат Гольджи
Жгутики, рибосомы
Жгутики, миофибриллы
5.В интерфазном ядре различают:
Рибосомы
ядрышки
кариолемму
ядерный сок
хроматин
6.Виды хромосом в зависимости от расположения центромеры:
cубцентрические
Телоцентрические
акроцентрические
метацентрические
неравноплечие
7.Каждая хромосома в метафазе митоза состоит из:
ДНК, белков гистонов, негистоновых белков
2х микрофибрилл
2х хроматид
4х полухроматид
хромонем
8.Соматические клетки имеют набор хромосом:
Полиплоидный
диплоидный
Гаплоидный
Тетраплоидный
9.На гладкой ЭПС идет синтез:
углеводов
10.Хромопласты содержат пигмент:
Гемоглобин
ликопин
каротин
ксантофилл
Хлорофилл
11.Функции рибосом:
Гормонов
Синтез углеводов
Нуклеиновых кислот
синтез белков
Синтез жиров
12.Группы включений:
витаминные
трофические
минеральные
секреторные, экскреторные
пигментные
13.Половые клетки имеют набор хромосом:
Диплоидный
Полиплоидный
гаплоидный
14.К эукариотам относятся:
Цианобактерии
животные
лишайники
растения
15.Группа немебранных органелл цитоплазмы эукариотических клеток:
Рибосомы, вакуоли, микротрубочки
рибосомы, центросома
ЭПС, митохондрии, рибосомы
микротрубочки, микрофибриллы
Микрофиламенты, пластиды, лизосомы
16.Функции гиалоплазмы:
участие в обмене веществ
Синтетическая
Разграничительная
гомеостатическая
транспортная
17.Функции ядрышек:
Синтез белков
формирование субъединиц рибосом
синтез транспортной РНК
Образование микрофибрилл
синтез рибосомальной РНК
18.Функции ЭПС:
Рецепторная
транспортная
разграничительная
Защитная
синтетическая
19.Функции лизосом:
Синтез и накопление углеводов
разрушение временных органов эмбрионов и личинок
Синтез белков, жиров, углеводов
аутолиз, разрушение инородных частиц
внутриклеточное пищеварение
20.Вакуоли – это:
Немембранные органеллы животных клеток
мембранные органеллы клеток растений
Мембранные органеллы бактериальных клеток
мембранные органеллы одноклеточных животных
Немембранные органеллы клеток растений
21.Функции вакуолей:
осуществляют удаление излишков воды
поддерживают осмотическое давление клетки
осуществляют внутриклеточное пищеварение
поддерживают тургорное давление в клетках растений
Осуществляют синтез и накопление жиров в растительных клетках
22.Хлоропласты содержат пигмент:
Ксантофилл
хлорофилл
23.В состав рибосом входят:
Т- РНК, магний
Белки, ДНК, магний
Только белки
Белки, и- РНК
белки, р- РНК, магний
24.В состав микрофиламентов входит белок:
25.В состав микротрубочек входит белок:
тубулин
26.В состав микрофибрилл входит белок:
Коллаген
кератин
27.Ядро характерно для клеток:
Всех живых организмов
эукариот
растений
Прокариот