Jądro komórkowe jest podstawową strukturą każdego organizmu eukariotycznego. pełni różnorodne funkcje, ale jego głównym celem jest przechowywanie i przekazywanie dziedzicznego materiału genetycznego.
Prawie każda komórka w ludzkim ciele ma jądro. Jedynymi wyjątkami są płytki krwi i czerwone krwinki. Większość komórek jest jednojądrzasta, ale na przykład włókna mięśniowe i neurony mogą mieć kilka takich organelli. Jądro w komórce może mieć różne rozmiary - są to największe struktury jądrowe w żeńskim jaju.
Jądro komórkowe: struktura
Jądro ma dość złożoną strukturę i składa się z otoczki jądrowej, chromatyny, jąderka i nukleoplazmy. Przyjrzyjmy się każdej jego części bardziej szczegółowo.
- Karoteca, czyli otoczka jądrowa, to struktura oddzielająca wewnętrzne środowisko jądra od cytoplazmy. Powłoka ta składa się z błony zewnętrznej i wewnętrznej, pomiędzy którymi znajduje się tzw. przestrzeń okołojądrowa. Co ciekawe, zewnętrzna błona skorupy przechodzi bezpośrednio do błony ziarnistej retikulum endoplazmatycznego, dzięki czemu wnęki cystern EPS i jądro są ze sobą połączone. A skorupa ma pory jądrowe zamknięte przeponą. Są przeznaczone do penetracji dużych cząsteczek, a także do wymiany substancji między karioplazmą a cytoplazmą.
- Karioplazma jest jednorodną substancją wypełniającą wewnętrzną jamę jądra. Zawiera jąderko i chromatynę.
- Chromatyna jest materiałem genetycznym komórki. Jego jednostką strukturalną jest nukleosom, czyli nić DNA owinięta wokół określonego białka – histonu. W komórce występują dwa stany materiału genetycznego. Heterochromatyna to małe, gęste granulki osmofilowe. Euchromatyna, czyli luźna chromatyna, to obszar, w którym aktywnie zachodzą procesy syntetyczne. Podczas tego procesu chromatyna ulega kondensacji, tworząc chromosomy.
- Jąderko to mała, owalna struktura składająca się z nici RNA i cząsteczek białka. W tym miejscu następuje tworzenie podjednostek rybosomów. W jądrze może znajdować się jedno lub więcej jąder, ale można je zobaczyć tylko w komórkach niedzielących się.
Jądro komórkowe: funkcje
Funkcje można określić zapoznając się z jego strukturą. Po pierwsze, jądro jest odpowiedzialne za przekazywanie dziedzicznego zestawu informacji podczas podziału komórki, zarówno mitozy, jak i mejozy. Podczas mitozy komórki potomne otrzymują genom identyczny z komórką macierzystą. Podczas mejozy (powstawania człowieka) każda komórka otrzymuje tylko połowę zestawu chromosomów - pełny zestaw chromosomów powstaje dopiero po połączeniu z komórką zarodkową innego organizmu.
Ponadto jądro komórkowe odpowiada za jeden z najważniejszych etapów metabolizmu – syntezę białek. Faktem jest, że to w jądrze powstaje informacyjny lub informacyjny RNA. Następnie wchodzi do retikulum endoplazmatycznego, przyłącza się do rybosomu i służy jako model tworzenia sekwencji aminokwasowej cząsteczki peptydu.
I jak już wspomniano, synteza podjednostek rybosomów odbywa się w jądrze.
Jądro komórkowe: pochodzenie
Obecnie istnieje kilka zupełnie różnych hipotez, za pomocą których naukowcy próbują wyjaśnić, jak dokładnie powstało jądro w komórce. Ale niestety żadne z tych stwierdzeń nie znalazło jeszcze potwierdzenia faktycznego.
Istnieje teoria, że jądro jako struktura komórkowa powstało w wyniku symbiozy i archeonów. Inni naukowcy uważają, że jądro powstaje w wyniku zakażenia komórki określonym wirusem.
Najbardziej kompletne wyjaśnienie zawiera tzw. hipotezę egzobłonową. Według niej w procesie ewolucji komórka wykształciła kolejną zewnętrzną błonę komórkową. Jednocześnie stara, wewnętrzna błona zamieniła się w otoczkę jądra - z czasem pojawił się w niej złożony system porów, a następnie w jej wnęce zaczęły gromadzić się cząsteczki chromatyny.
Wszyscy wiedzą, że ludzie są eukariontami. Oznacza to, że wszystkie jego komórki mają organellę zawierającą całą informację genetyczną – jądro. Są jednak wyjątki. Czy w organizmie człowieka istnieją komórki bezjądrowe i jakie jest ich znaczenie dla życia?
Jądrowane komórki ludzkie
Nie można ich porównać z prokariotami, które mają typową budowę. Jakiego rodzaju są to erytrocyty bezjądrowe? Zamiast tej organelli zawierają złożony kompleks chemiczny substancji, który pozwala im pełnić najważniejsze funkcje dla organizmu. Płytki krwi – płytki krwi i limfocyty – są również komórkami pozbawionymi jądra. W komórkach zwanych komórkami macierzystymi nie ma jądra. Wszystkie powyższe konstrukcje łączy jeszcze jedna cecha wspólna. Ponieważ brakuje im jądra, nie są w stanie się rozmnażać. Oznacza to, że komórki pozbawione jądra, których przykłady zostały podane, po spełnieniu swojej funkcji umierają, a w wyspecjalizowanych narządach powstają nowe.
Czerwone krwinki
To oni decydują o kolorze naszej krwi. Bezjądrzaste krwinki czerwone mają nietypowy kształt – dwuwklęsły krążek, co znacznie zwiększa ich powierzchnię przy stosunkowo niewielkich rozmiarach. Ale ich liczba jest po prostu niesamowita: na 1 m2. Krwi jest aż 5 milionów mm! Czerwona krwinka żyje średnio do czterech miesięcy, po czym obumiera i ulega neutralizacji w śledzionie i wątrobie. Nowe komórki powstają co sekundę w czerwonym szpiku kostnym.
Funkcje czerwonych krwinek
Co zamiast jądra zawierają te pozbawione jądra komórki? Substancje te nazywane są hemem i globiną. Pierwszy zawiera żelazo. Nie tylko zabarwia krew na czerwono, ale także tworzy niestabilne związki z tlenem i dwutlenkiem węgla. Globina jest substancją białkową. Jego duża cząsteczka zawiera hem, który zawiera naładowany jon żelaza. Zgodnie z mechanizmem działania komórki te można porównać do minibusa. W płucach dodają tlen. Wraz z przepływem krwi rozprzestrzenia się do wszystkich komórek i tam jest uwalniany. Przy udziale tlenu zachodzi proces utleniania substancji organicznych, uwalniając pewną ilość energii, którą człowiek wykorzystuje do wykonywania czynności życiowych. Zwolnioną przestrzeń natychmiast zajmuje dwutlenek węgla, który przemieszcza się w przeciwnym kierunku - do płuc, gdzie jest wydychany. Proces ten jest niezbędnym warunkiem życia. Jeśli tlen nie dotrze do komórek, stopniowo obumierają. Może to być niebezpieczne dla życia całego organizmu.
Czerwone krwinki pełnią jeszcze jedną ważną funkcję. Na ich błonach znajduje się marker białkowy zwany czynnikiem Rh. Wskaźnik ten, podobnie jak grupa krwi, jest bardzo ważny podczas transfuzji krwi, ciąży, oddawania krwi i operacji. Należy go zainstalować, ponieważ w przypadku niezgodności może wystąpić tak zwany konflikt Rh. Jest to reakcja ochronna, ale może prowadzić do odrzucenia płodu lub narządu.
Złe odżywianie, złe nawyki i zanieczyszczone powietrze mogą powodować zniszczenie czerwonych krwinek. W rezultacie pojawia się poważna choroba, która nazywa się anemią lub anemią. W takim przypadku osoba odczuwa zawroty głowy, osłabienie, duszność i szum w uszach. Niedobór tlenu negatywnie wpływa na aktywność fizyczną i umysłową człowieka. Jest to szczególnie niebezpieczne w czasie ciąży. Jeśli przez pępowinę do płodu nie dotrze wystarczająca ilość tlenu, może to prowadzić do poważnych problemów w jego rozwoju.
Struktura płytek krwi
Płytki krwi, komórki bezjądrowe, nazywane są również płytkami krwi. W stanie nieaktywnym mają w rzeczywistości płaski kształt, przypominający soczewkę. Ale kiedy naczynia ulegają uszkodzeniu, puchną, zaokrąglają się i tworzą niestabilne narośla zewnętrznej warstwy - pseudopodia. Płytki krwi tworzą się i nie żyją długo - do 10 dni, są neutralizowane w śledzionie.
Proces tworzenia się skrzepów krwi
Macierz płytek krwi zawiera enzym zwany tromboplastyną. Kiedy integralność naczyń krwionośnych zostaje naruszona, trafia do osocza. Pod jego działaniem białko krwi protrombina przekształca się w swoją aktywną formę, działając z kolei na fibrynogen. W rezultacie substancja ta staje się nierozpuszczalna. Zamienia się w fibrynę białkową. Jego nici są ściśle ze sobą powiązane i tworzą skrzep krwi. Ochronna reakcja krzepnięcia krwi zapobiega utracie krwi. Jednakże utworzenie się skrzepu krwi w naczyniu jest bardzo niebezpieczne. Może to doprowadzić do jego pęknięcia, a nawet śmierci organizmu. Zaburzenie krwawienia nazywa się hemofilią. Ta dziedziczna choroba charakteryzuje się niewystarczającą liczbą płytek krwi i prowadzi do nadmiernej utraty krwi.
Komórki macierzyste
Te komórki bezjądrowe nie bez powodu nazywane są komórkami macierzystymi. Oni naprawdę są podstawą dla wszystkich innych. Nazywa się je również „czystymi genetycznie”. Komórki macierzyste znajdują się we wszystkich tkankach i narządach, ale najwięcej ich zawiera szpik kostny. Pomagają przywrócić integralność tam, gdzie jest to potrzebne. Te łodygowe zmieniają się w dowolne inne, gdy zostaną zniszczone. Wydawałoby się, że dzięki takiemu magicznemu mechanizmowi człowiek powinien żyć wiecznie. Dlaczego tak się nie dzieje? Rzecz w tym, że wraz z wiekiem intensywność różnicowania komórek macierzystych znacząco maleje. Nie są już w stanie przywrócić uszkodzonej tkanki. Ale istnieje inne niebezpieczeństwo. Istnieje duże prawdopodobieństwo, że komórki macierzyste zamienią się w komórki nowotworowe, co nieuchronnie doprowadzi do śmierci każdego żywego organizmu.
Komórki bezjądrowe: przykłady i różnice
W naturze komórki bezjądrowe są dość powszechne. Na przykład sinice i bakterie są prokariotami. Jednak w przeciwieństwie do pozbawionych jądra komórek ludzkich, nie umierają one po spełnieniu swojej biologicznej roli. Faktem jest, że prokarioty mają materiał genetyczny. Dlatego są zdolne do podziału, który zachodzi dzięki dwóm kopiom genetycznym komórki macierzystej. Informacja dziedziczna prokariotów jest reprezentowana przez kolistą cząsteczkę DNA, która podwaja się przed podziałem. Ten analog jądra nazywany jest również nukleoidem. W roślinach żywe komórki – rurki sitowe – są wolne od jąder.
Zatem pozbawione jądra komórki ludzkie nie są zdolne do podziału, dlatego istnieją przez krótki okres czasu, zanim spełnią swoją funkcję. Następnie są niszczone i trawione wewnątrzkomórkowo. Należą do nich elementy formowane (czerwone krwinki), płytki krwi (płytki krwi) i komórki macierzyste.
Brown nie poczynił żadnych założeń na temat funkcji jądra. W 1838 roku Matthias Schleiden zasugerował, że jądro bierze udział w tworzeniu nowych komórek, dlatego wprowadził termin „cytoblast” (budowniczy komórek) w odniesieniu do jąder. Był pewien, że zaobserwował gromadzenie się nowych komórek wokół „cytoblastów”. Zagorzałym przeciwnikiem tego poglądu był Franz Meyen, który odkrył, że komórki rozmnażają się przez podział i uważał, że wiele komórek może nie mieć jądra. Idea powstawania komórek od nowa , czyli od zera, poprzez cytoblasty lub w inny sposób, zaprzeczało pracom Roberta Remaka (1852) i Rudolfa Virchowa (1855), którzy ostatecznie ustalili nowy paradygmat stwierdzający, że komórki mogą powstawać jedynie z komórek („Omnis cellula e cellula” ). Funkcje jądra pozostały niejasne.
Struktury
Jądro jest największą organellą komórek zwierzęcych. U ssaków średnica jądra wynosi w przybliżeniu 6 µm, a samo jądro stanowi około 10% objętości komórki. Lepka ciecz wypełniająca jądro nazywana jest nukleoplazmą i jest chemicznie podobna do cytozolu otaczającego jądro.
Otoczka jądrowa i pory jądrowe
Otoczka jądrowa składa się z dwóch membran (zewnętrznej i wewnętrznej), które są umieszczone równolegle w odległości 10 ton 50 nm. Otoczka jądrowa całkowicie otacza jądro, oddzielając materiał genetyczny komórki od cytoplazmy i stanowiąc barierę uniemożliwiającą swobodną dyfuzję makrocząsteczek pomiędzy nukleoplazmą a cytoplazmą. Zewnętrzna błona jądrowa przechodzi w błonę szorstkiej siateczki śródplazmatycznej (ER) i jest pokryta rybosomami. Przestrzeń pomiędzy błonami jądrowymi nazywa się przestrzenią okołojądrową i biegnie do światła ER.
Pory jądrowe, czyli wypełnione wodą kanały w otoczce jądrowej, składają się z wielu białek zwanych nukleoporynami. U ludzi masa porów wynosi ok 120 000 kDa, co stanowi 40-krotność masy rybosomu; Ponadto u drożdży pory jądrowe zawierają około 50 białek, a u kręgowców - kilkaset. Chociaż średnica porów wynosi 100 nm, szerokość szczeliny, przez którą mogą przejść cząsteczki, ze względu na obecność układów regulacyjnych wewnątrz porów, wynosi zaledwie 9 nm. Przez taką szczelinę mogą przechodzić małe cząsteczki rozpuszczalne w wodzie, ale nie duże cząsteczki, takie jak kwasy nukleinowe i duże białka; Do transportu tych cząsteczek do jądra wymagany jest aktywny (to znaczy energochłonny) transport. Na błonie jądrowej typowej komórki ssaka znajduje się od 3000 do 4000 porów, a każdy z nich ma strukturę pierścieniową z 8 osiami symetrii na styku dwóch błon jądrowych. Do pierścienia przymocowana jest specjalna struktura zwana koszykiem jądrowym, która wnika do nukleoplazmy, a kilka jej włókien wystaje do cytoplazmy. Obie struktury są wymagane do pośredniczenia w wiązaniu białek transportujących jądro.
Większość białek, podjednostek rybosomów i część DNA jest transportowana przez pory jądrowe za pomocą rodziny czynników transportu znanych jako karioferyny. Nazywa się także karioferyny, które pośredniczą w transporcie do jądra importowanie, a te, które pośredniczą w transporcie z jądra, to eksportyny. Większość karioferyn bezpośrednio oddziałuje ze swoim ładunkiem, ale niektóre z nich korzystają adapter wiewiórki Hormony steroidowe (takie jak kortyzol i aldosteron) oraz inne małe cząsteczki rozpuszczalne w tłuszczach mogą dyfundować do cytoplazmy do komórki przez błonę komórkową; w cytoplazmie wiążą się z białkowymi receptorami jądrowymi, które dostarczają je do jądra. Tutaj receptory jądrowe związane ze swoimi ligandami, działają jako czynniki transkrypcyjne, a przy braku ligandu wiele receptorów działa jako deacetylazy histonowe, tłumiąc ekspresję niektórych genów.
Blaszka jądrowa
W komórkach zwierzęcych mechaniczne wsparcie jądra zapewniają dwie sieci włókien pośrednich: blaszka jądrowa, która jest siecią włókien pośrednich na wewnętrznej powierzchni jądra, oraz mniej zorganizowane włókna na powierzchni cytozolowej jądra. Obydwa systemy włókien zapewniają wsparcie dla jądra i służą do zakotwiczenia chromosomów i porów jądrowych.
Blaszka jądrowa składa się głównie z białek zwanych laminami. Podobnie jak wszystkie białka, laminy są syntetyzowane w cytoplazmie, a następnie transportowane do jądra, gdzie są wstawiane do blaszki jądrowej. Blaszki znajdujące się na zewnątrz otoczki jądrowej (np Emerin I nespryna), wiążą się z elementami cytoszkieletu, co zapewnia wsparcie strukturalne jądru. Laminy znajdują się również w nukleoplazmie, gdzie tworzą inną regularną strukturę zwaną zasłoną nukleoplazmatyczną. zasłona nukleoplazmatyczna) ; ten ostatni można uwidocznić za pomocą mikroskopii fluorescencyjnej. Funkcja zasłony nie jest znana, wiadomo jednak, że nie występuje ona w jąderku i jest obecna podczas interfazy cyklu komórkowego. Laminy tworzące zasłonę (takie jak LEM3) wiążą się z chromatyną, a zaburzenia w ich strukturze hamują transkrypcję genów kodujących białka.
Podobnie jak inne białka włókien pośrednich, monomery laminowe zawierają domenę α-helikalną, wykorzystywaną przez dwa monomery do owijania się wokół siebie, tworząc dimer o strukturze bispirale. Dwa dimery dalej łączą się po swoich bokach w orientacji antyrównoległej, tworząc tetramer znany jako protofilament. Osiem tetramerów łączy się, tworząc skręcone włókno przypominające linę. Filamenty mogą składać się i demontować w sposób dynamiczny, to znaczy długość włókna zależy od względnych szybkości jego montażu i demontażu.
Chromosomy
Jądro zawiera większość materiału genetycznego komórki, składającego się z wielu liniowych cząsteczek DNA zorganizowanych w struktury zwane chromosomami. Całkowita długość cząsteczek DNA w komórce ludzkiej wynosi około 2 m. Przez większą część cyklu komórkowego cząsteczki te w połączeniu z białkami tworzą tzw. chromatynę, a podczas podziału komórki chromosomy pojawiają się jako oddzielne, wyraźnie rozróżnialne chromosomy, które składają się na kariotyp. Niewielka ilość materiału genetycznego komórki zlokalizowana jest w mitochondriach, a w przypadku komórki roślinnej w chloroplastach.
Znane są dwa rodzaje chromatyny. W euchromatynie DNA jest najmniej gęsto zorganizowane; zawiera geny, które ulegają najczęściej transkrypcji. Inny rodzaj chromatyny, heterochromatyna, jest bardziej zwarty i zawiera DNA, które rzadko lub nigdy nie ulega transkrypcji. Heterochromatynę dzielimy na fakultatywną, która występuje tylko w komórkach określonego typu i na pewnym etapie cyklu komórkowego oraz konstytutywną, reprezentowaną przez struktury chromosomowe, takie jak telomery i centromery. Podczas interfazy chromatyna każdego chromosomu zajmuje ściśle określony obszar jądra - terytorium chromosomowe. Aktywne geny, które zwykle są zlokalizowane w euchromatynie, są zwykle zlokalizowane na granicy terytorium chromosomu.
Ciała jądrowe
Jądro komórek ssaków zawiera szereg odrębnych podprzedziałów zwanych ciałami jądrowymi. Dzielą rdzeń, tworząc w nim oddzielne przestrzenie, które mają określone właściwości. Wiele ciał jądrowych pełni określone funkcje - na przykład syntezę i przetwarzanie prerybosomalnego RNA w jąderku, akumulację i składanie składników spliceosomalnych w plamkach (patrz poniżej) lub akumulację cząsteczek RNA w parakropki. Mechanizmy zapewniające, że ciała jąderkowe pełnią te funkcje, są bardzo zróżnicowane. W niektórych przypadkach ciało jądrowe może służyć jako miejsce pewnych procesów, takich jak transkrypcja. W innych przypadkach ciała jądrowe najwyraźniej pośrednio regulują lokalne stężenia swoich składników w nukleoplazmie. Podobnie jak organelle cytoplazmatyczne, ciała jądrowe zawierają specyficzny zestaw białek, które określają ich strukturę na poziomie molekularnym. Jednakże w przeciwieństwie do organelli cytoplazmatycznych, ciała jądrowe nie są otoczone błonami lipidowymi, a ich integralność strukturalna jest w całości zapewniona przez interakcje białko-białko i RNA-białko. Poniższa tabela zawiera listę głównych cech ciał jądrowych.
| Ciało nuklearne | Funkcje | Charakterystyczne komponenty | Typowy rozmiar (w mikronach) | Ilość na rdzeń |
|---|---|---|---|---|
| Jądro | Biogeneza rybosomów | Maszyneria Polimeraza RNA I, czynniki przetwarzania rRNA i czynniki składania podjednostek rybosomów | 3-8 | 1-4 |
| Plamy | Akumulacja i montaż czynników splicingowych | Czynniki splicingu pre-mRNA | 2-3 | 20-50 |
| Naprężenia ciał jądrowych | Regulacja transkrypcji i splicingu w warunkach stresowych | HSF1,HAP | 1-2 | 3-6 |
| Ciało loci histonów | Przetwarzanie pre-mRNA histonów | NPA,BŁYSK, U7 snRNP | 0,2-1,2 | 2-4 |
| ciałko Cajala | Biogeneza, dojrzewanie i obrót małych RNA | Coilina, SMN | 0,2-1,5 | 1-10 |
| Organ PML | Regulacja stabilności genomu, naprawa DNA, kontrola transkrypcji, ochrona przed wirusami | PML | 0,1-1 | 10-30 |
| Paraspeckles | Regulacja mRNA, edycja RNA | Niekodujące RNA NEAT1/MENε/β, białka PSP1, p54 nrb/NONO | 0,2-1 | 2-20 |
| Przedział okołojądrowy | Potranskrypcyjna regulacja zestawu syntetyzowanych RNA Polimeraza RNA III | PTB | 0,2-1 | 1-2 |
Jądro
Jąderko jest odrębną, gęstą strukturą w jądrze. Nie jest otoczony błoną i powstaje w regionie, w którym znajduje się rDNA – tandemowe powtórzenia genów rybosomalnego RNA (rRNA), zwane organizatorami jąderkowymi. Główną funkcją jąderka jest synteza rRNA i tworzenie rybosomów. Integralność strukturalna jąderka zależy od jego aktywności, a inaktywacja genów rRNA prowadzi do pomieszania struktur jąderkowych.
W pierwszym etapie tworzenia rybosomów enzym polimeraza RNA I transkrybuje rDNA i wytwarza pre-rRNA, który jest dalej cięty na rRNA 5.8S, 18S i 28S. Transkrypcja i przetwarzanie potranskrypcyjne rRNA zachodzą w jąderku przy udziale małych jąderkowych RNA (snoRNA), z których część pochodzi ze splicowanych intronów mRNA genów kodujących białka związane z funkcją rybosomów. Zmontowane podjednostki rybosomów są największymi strukturami przechodzącymi przez pory jądrowe.
Oglądając pod mikroskopem elektronowym, w jąderku można wyróżnić trzy składniki: centra włókniste (FC), otaczający gęsty składnik włóknisty (DFC) i składnik ziarnisty (GC), który z kolei otacza PFC. Transkrypcja rRNA zachodzi w PC oraz na granicy PC i PFC, dlatego też, gdy aktywowane jest tworzenie rybosomów, PC stają się wyraźnie rozróżnialne. Cięcie i modyfikacje rRNA zachodzą w PFC, a kolejne etapy tworzenia podjednostek rybosomalnych, w tym ładowanie białek rybosomalnych, zachodzą w GC.
ciałko Cajala
Ciało Cajala (CB) to ciało jądrowe występujące u wszystkich eukariontów. Można go rozpoznać po obecności białka sygnatury cewki i specyficznych RNA (scaRNA). MC zawiera również białko SMN. przetrwanie neuronów ruchowych). MC zawierają wysokie stężenia małych jądrowych rybonukleoprotein splicingowych (snRNP) i innych czynników przetwarzających RNA i uważa się, że MC służą jako miejsca składania i/lub potranskrypcyjnej modyfikacji czynników splicingu. TK jest obecny w jądrze podczas interfazy, ale znika w mitozie. W biogenezie MC można prześledzić właściwości samoorganizującej się struktury.
Kiedy po raz pierwszy badano wewnątrzkomórkową lokalizację SMN za pomocą immunofluorescencji, białko znaleziono w całej cytoplazmie, a także w ciałku jąderkowym o wielkości podobnej do MC i często zlokalizowanej w sąsiedztwie MC. Z tego powodu ciało to nazwano „bliźniakiem TK” (ang. bliźnięta CB) lub po prostu klejnot. Okazało się jednak, że linia komórkowa HeLa, w której odkryto nowe ciało, była nietypowa: w innych liniach komórkowych człowieka, a także u muszki owocowej muszka owocowa SMN kolokalizowany z cewką w MC. Dlatego ogólnie SMN można uznać za ważny składnik MC, a nie za marker odrębnego ciała jądrowego.
Ciało loci histonów
Organy PML
Plamy
Paraspeckles
Paraspeckles to ciała jądrowe o nieregularnym kształcie zlokalizowane w przestrzeni międzychromatycznej jądra. Po raz pierwszy opisano je w komórkach HeLa, które mają 10–30 parapekles na jądro, ale obecnie paraspeckles można znaleźć we wszystkich pierwotnych komórkach ludzkich, w transformowanych liniach komórkowych i w skrawkach tkanek. Swoją nazwę otrzymali ze względu na położenie w jądrze – w pobliżu plamek.
Paraspeckles to dynamiczne struktury, które zmieniają się w odpowiedzi na zmiany w aktywności metabolicznej komórki. Zależą od transkrypcji i przy braku transkrypcji przez polimerazę RNA II, paraplemki znikają, a wszystkie białka składowe (PSP1, p54nrb, PSP2, CFI(m)68 i PSF) tworzą czapeczkę okołojądrową w kształcie półksiężyca. Zjawisko to obserwuje się podczas cyklu komórkowego: paracętki występują w interfazie i we wszystkich fazach mitozy z wyjątkiem telofazy. Podczas telofazy powstają jądra potomne, a polimeraza RNA II niczego nie transkrybuje, więc białka paraspeckle tworzą czapeczkę okołojądrową. Paraspeckles biorą udział w regulacji ekspresji genów, gromadząc te RNA, w których znajdują się regiony dwuniciowe, które podlegają edycji, czyli konwersji adenozyny do inozyny. Dzięki temu mechanizmowi paracętki biorą udział w kontroli ekspresji genów podczas różnicowania, infekcji wirusowej i stresu.
Przedział okołojądrowy
Przedział okołojądrowy (UC) jest ciałem jądrowym o nieregularnym kształcie, charakteryzującym się umiejscowieniem na obrzeżach jąderka. Pomimo fizycznego połączenia oba przedziały różnią się strukturalnie. Zwykle OK znajdują się w komórkach nowotworów złośliwych. OK jest strukturą dynamiczną i zawiera wiele białek wiążących RNA i polimerazę III RNA. Stabilność strukturalną OC zapewnia transkrypcja przeprowadzana przez polimerazę RNA III i obecność kluczowych białek. Ponieważ obecność TC jest zwykle związana z nowotworem złośliwym i możliwością tworzenia przerzutów, uważa się je za potencjalne markery raka i innych nowotworów złośliwych. Wykazano związek OK z określonymi loci DNA.
Naprężenia ciał jądrowych
Podczas szoku cieplnego w jądrze powstają ciała jądrowe naprężeniowe. Powstają w wyniku bezpośredniego oddziaływania czynnika transkrypcyjnego szoku cieplnego 1 ( HSF1) i perycentryczne powtórzenia tandemowe w sekwencji satelity III, które odpowiadają miejscom aktywnej transkrypcji niekodujących transkryptów satelity III. Powszechnie uważa się, że ciała takie odpowiadają bardzo gęsto upakowanym formom kompleksów rybonukleoproteinowych. Uważa się, że w komórkach poddanych stresowi biorą one udział w szybkich, przejściowych i globalnych zmianach w ekspresji genów poprzez różne mechanizmy – na przykład przebudowę chromatyny oraz wychwyt czynników transkrypcyjnych i splicingowych. W komórkach w normalnych (bezstresowych) warunkach rzadko spotyka się ciała jądrowe stresu, ale ich liczba gwałtownie wzrasta pod wpływem szoku cieplnego. Ciała jądrowe stresu znajdują się tylko w komórkach ludzi i innych naczelnych.
Nuklearne ciała sierot
| Ciało nuklearne | Opis | Typowy rozmiar (w mikronach) | Ilość na rdzeń |
|---|---|---|---|
| Klastosom | Koncentruje kompleksy proteasomów 20S i 19S oraz białka związane z ubikwityną. Jest wykrywany przede wszystkim w przypadku stymulacji aktywności proteasomu i jest eliminowany w przypadku hamowania aktywności proteasomu. | 0,2-1,2 | 0-3 |
| Korpuskuła podziału dekoltowe ciało) | Wzbogacony o czynniki rozszczepialne CstF I CPSF, a także białko DDX1 zawierający MARTWE pudełko. Występuje głównie w fazie S i nie podlega inhibicji transkrypcji. | 0,2-1,0 | 1-4 |
| Domena OPC | Wzbogacony o czynniki transkrypcyjne 1 października i PTF. Częściowo kolokalizuje z miejscami transkrypcji. Występuje głównie w późnej fazie G1 i ulega rozkładowi w przypadku zahamowania transkrypcji. | 1,0-1,5 | 1-3 |
| Korpus Polycomb | Występuje w komórkach człowieka i Drosophila, jest wzbogacony w białko PcG. U ludzi gromadzą się białka RING1, BMI1, HPC, może być powiązany z perycentromeryczną heterochromatyną. | 0,3-1,0 | 12-16 |
| Byk Sam68 | Gromadzi białko Sam68 i podobne białka SLM-1 i SLM-2. Rozkładany, gdy transkrypcja jest hamowana. Prawdopodobnie wzbogacony w RNA. | 0,6-1,0 | 2-5 |
| Byk SUMO | Wzbogacony o białka SUMO i enzym koniugujący SUMO Ubc9. Koncentruje czynniki transkrypcyjne pCREB, CBP, c-czerwiec. | 1-3 | 1-3 |
Funkcje
Otoczka jądrowa chroni DNA komórki i bierze udział w znacznie bardziej złożonej regulacji ekspresji genów w porównaniu z komórką prokariotyczną. U prokariotów transkrypcja i translacja są procesami sprzężonymi, a translacja mRNA na białko rozpoczyna się jeszcze przed jego pełną syntezą. W komórkach eukariotycznych cytoplazma, w której zachodzi translacja, i transkrypcja, która zachodzi w jądrze, są przestrzennie oddzielone, dlatego istnieje potrzeba zapewnienia transportu cząsteczek pomiędzy jądrem a cytoplazmą.
Otoczka jądrowa daje jądru możliwość kontrolowania swojej zawartości i oddziela ją od reszty cytoplazmy. Ma to znaczenie dla regulacji procesów zachodzących po obu stronach otoczki jądrowej. Kiedy proces cytoplazmatyczny musi zostać w jakiś sposób ograniczony, wówczas zazwyczaj jego kluczowy uczestnik zostaje przeniesiony do jądra, gdzie oddziałuje z czynnikami transkrypcyjnymi i tym samym powoduje zahamowanie powstawania niektórych enzymów biorących udział w procesie cytoplazmatycznym. Na przykład taki mechanizm regulacyjny istnieje w glikolizie, procesie, podczas którego komórka pobiera energię z cząsteczki glukozy. Pierwszą reakcję glikolizy przeprowadza enzym heksokinaza, przekształcając cząsteczkę glukozy w glukozo-6-fosforan. Gdy wzrasta stężenie fruktozo-6-fosforanu (substancji powstałej z glukozo-6-fosforanu podczas glikolizy), białko regulatorowe wysyła heksokinazę do jądra, gdzie tworzy kompleks represyjny transkrypcję, który hamuje ekspresję genów kodujących enzymy glikolityczne.
Aby kontrolować, które geny ulegają transkrypcji, czynniki transkrypcyjne w komórce nie mają fizycznego dostępu do DNA, dopóki nie zostaną aktywowane przez specyficzny szlak sygnalizacyjny. Zapobiega to nawet niskiej ekspresji nieprawidłowych genów. W szczególności w przypadku genów kontrolowanych przez NF-κB, które biorą udział w procesie zapalnym, transkrypcja jest indukowana przez szlak sygnalizacyjny, na przykład rozpoczynający się od związania cząsteczki sygnalizacyjnej TNF-α z jej receptorem na błonie komórkowej i ostatecznie prowadząc do aktywacji czynnika transkrypcyjnego NF-κB. Sygnał lokalizacji jądrowej obecny w NF-κB umożliwia mu przedostawanie się do i z jądra przez pory jądrowe; w jądrze stymuluje transkrypcję genów docelowych.
Kompartmentalizacja zapobiega transkrypcji niesplicowanego mRNA przez komórkę. Eukariotyczne mRNA zawierają introny, które należy usunąć przed translacją mRNA. Splicing, czyli usuwanie intronów, zachodzi w jądrze, co uniemożliwia dostęp do pre-mRNA rybosomom znajdującym się poza jądrem. Gdyby nie było jądra, rybosomy zaczęłyby tłumaczyć niedojrzały mRNA, co doprowadziłoby do powstania nieprawidłowych produktów białkowych.
Ponieważ transkrypcja zachodzi w jądrze, jądro zawiera wiele białek, które bezpośrednio biorą udział w transkrypcji lub regulują ten proces. Do białek tych należą helikazy, które rozwijają podwójną helisę DNA, ułatwiając dostęp do niej innym białkom, polimerazy RNA syntetyzujące RNA, topoizomerazy wpływające na topologię DNA oraz różnorodne czynniki transkrypcyjne.
Transport nuklearny
Wyjście i wejście do jądra dużych cząsteczek jest kontrolowane przez pory jądrowe. Chociaż małe cząsteczki mogą przedostawać się do jądra bez jakiejkolwiek regulacji, makrocząsteczki – takie jak białka i RNA – muszą wiązać się z karioferynami w celu transportu do jądra (importyny) i z jądra (eksportyny). Białka, które muszą zostać przetransportowane z cytoplazmy do jądra, zawierają specjalną sekwencję aminokwasów, zwaną sygnałem lokalizacji jądrowej, z którą wiążą się importyny. Podobnie zawierają białka, które muszą opuścić jądro sygnał eksportu broni nuklearnej, uznawany przez eksporterów. Zdolność importerów i eksporterów do przenoszenia ładunku jest regulowana przez GTPazy, enzymy, które hydrolizują GTP w celu uwolnienia energii. Kluczowa GTPaza transportu nuklearnego - Biegł, który może wiązać się z GTP lub PKB, w zależności od jego lokalizacji (w jądrze lub w cytoplazmie). W jądrze oddziaływanie Ran-GTP z importyną powoduje zmianę konformacyjną tej ostatniej, w wyniku czego zostaje ona oddzielona od transportowanego ładunku. Powstały kompleks Ran-GTP i importyny jest transportowany do cytoplazmy, gdzie białko RanBP oddziela Ran-GTP od importyny. Oddzielenie od importyny pozwala na białko LUKA wiążą się z Ran-GTP i katalizują hydrolizę GTP do PKB. Kompleks Ran-GDP jest następnie rozpoznawany przez białko NUTF2, który zwraca go do nukleoplazmy. Białko w jądrze GEF zastępuje PKB GTP, tworząc Ran-GTP i zamykając cykl.
Montaż i demontaż
W trakcie życia komórki jądro może zostać rozłożone (podczas podziału komórki lub apoptozy). Podczas tych procesów ulegają zniszczeniu elementy strukturalne jądra – otoczka jądrowa i blaszka jądrowa. W większości komórek rozkład jądra następuje podczas profazy mitozy. Jednakże rozkład jądra nie ogranicza się wyłącznie do mitozy i nie zachodzi we wszystkich komórkach. Niektóre jednokomórkowe eukarionty (na przykład drożdże) przechodzą tak zwaną mitozę zamkniętą, w której otoczka jądrowa pozostaje nienaruszona. W zamkniętej mitozie chromosomy przemieszczają się na różne strony jądra, które następnie dzieli się na dwie części. Natomiast komórki wyższych eukariontów zwykle przechodzą otwartą mitozę, podczas której rozpada się otoczka jądrowa. Chromosomy migrują do różnych biegunów wrzeciona, a wokół nich ponownie tworzą się dwa jądra. Blaszka jądrowa ulega również demontażowi w wyniku fosforylacji lamin przez kinazy takie jak kinaza białkowa zależna od cyklin 1. Montaż blaszki jądrowej w jądra potomne rozpoczyna się po defosforylacji lamin.
Apoptoza to kontrolowany proces niszczenia składników komórkowych, prowadzący do śmierci komórki. Zmiany związane z apoptozą zachodzą bezpośrednio w jądrze i jego zawartości. Należą do nich kondensacja chromatyny, a także rozpad otoczki jądrowej i blaszki jądrowej. Zakłócenie sieci lamin następuje poprzez udział apoptotycznych proteaz zwanych kaspazami, które niszczą lamin i w ten sposób wpływają na integralność strukturalną jądra. Rozerwanie laminatu jest czasami używane jako wskaźnik aktywności kaspazy w badaniach apoptozy. Komórki wykazujące ekspresję zmutowanych lamin, które są odporne na działanie kaspaz, nie tracą integralności jądrowej podczas apoptozy, dlatego laminy odgrywają kluczową rolę w zapoczątkowaniu zmian, jakim ulega jądro podczas apoptozy. Dodatkowo hamowanie składania laminów w sieć wyzwala apoptozę.
Cechy jąder różnych eukariontów
Rozmiary, kształty i morfologia jąder eukariotycznych różnią się w najszerszym zakresie. Jeśli piroplazmid i Leishmania średnica rdzenia wynosi 1-3 mikrony, następnie u niektórych radiolarystów średnica rdzenia sięga 400 mikronów, a nawet 1 mm. Z reguły kształt jądra u większości eukariontów jest zbliżony do kulistego, ale czasami może przybierać dość dziwaczne kształty (dotyczy to w szczególności makrojąder orzęsków). Chociaż u wszystkich eukariontów otoczka jądrowa składa się z dwóch membran, liczba w niej porów różni się znacznie u różnych gatunków, a czasami mogą przylegać do niej dodatkowe warstwy (zarówno na zewnątrz, jak i wewnątrz); na przykład u wielu wolno żyjących ameb warstwa włóknista o strukturze komórkowej przylega do wewnętrznej strony muszli, która jest znacznie grubsza niż grubość błony jądrowej, a u radiolarianów dodatkowe warstwy włókniste znajdują się na zewnątrz skorupy.
Organizacja jądra u protistów z rodzaju bruzdnic jest znacząco wyjątkowa. Większość ich przedstawicieli ma jądro, w którym znajdują się chromosomy skondensowany przez cały cykl komórkowy (łącznie z interfazą) i są praktycznie pozbawione histonów. Ten typ jądra nazywa się dinokarion. Co więcej, ilość DNA w dinokarionie jest dziesiątki i setki razy większa niż ilość DNA na komórkę u przedstawicieli innych grup eukariontów. Jednakże niektóre bruzdnice ( Noctiluca, Oodin ) mają zwykłe jądra eukariotyczne; u innych przedstawicieli tego typu jądra w komórkach wegetatywnych są normalne, a dinokarion jest obecny na innych etapach cyklu komórkowego (na przykład w gametach).
Komórki protistów mają co najmniej jedno jądro. Jednocześnie w organizmach Metazoan występują również komórki bezjądrowe, które pozbawione jądra utraciły zdolność do podziału wraz z utworzeniem dwóch komórek potomnych. Najbardziej znanym przykładem komórek bezjądrowych są czerwone krwinki ssaków, którym brakuje innych organelli, takich jak mitochondria. Czerwone krwinki dojrzewają w szpiku kostnym w procesie erytropoezy, podczas którego tracą jądra, inne organelle i rybosomy. Jądro jest wypychane z komórki w procesie różnicowania erytroblastu do retikulocytu, który pełni rolę bezpośredniego prekursora erytrocytu. Pod wpływem niektórych mutagenów niedojrzałe czerwone krwinki zawierające mikrojądra mogą zostać uwolnione do krwi.
Większość protistów ma tylko jedno jądro; u protistów charakteryzujących się złożonym cyklem życiowym (na przykład u przedstawicieli typu apicomplexa) występują etapy jednojądrzaste i wielojądrowe.
Komórki wielojądrowe protistów
Zawierają chloroplasty glonów kryptofitów i chlorarachniofitów nukleomorf- zredukowany rdzeń fototroficznego endosymbiontu, wchłonięty przez przodków tych glonów podczas wtórnej endosymbiozy (inkorporacja krasnorostów nastąpiła u Cryptophyta, a zielonych u Chlorarachnea)
JĄDRO KOMÓRKOWE
Rdzeń(jądro, s. karion) występuje we wszystkich komórkach człowieka, z wyjątkiem erytrocytów i płytek krwi. Funkcje jądra polegają na przechowywaniu i przekazywaniu informacji dziedzicznych do nowych (córek) komórek. Funkcje te są związane z obecnością DNA w jądrze. Synteza białek – RNA kwasu rybonukleinowego i materiałów rybosomalnych – zachodzi również w jądrze.
Większość komórek ma jądro kuliste lub jajowate, ale występują również inne formy jądra (w kształcie pierścienia, w kształcie pręta, wrzecionowate, w kształcie fasoli, segmentowane, w kształcie gruszki, polimorficzne). Wymiary jądra są bardzo zróżnicowane - od 3 do 25 mikronów. Komórka jajowa ma największe jądro. Większość ludzkich komórek jest jednojądrzasta, ale zdarzają się komórki dwujądrowe (niektóre neurony, hepatocyty, kardiomiocyty). Niektóre struktury są wielojądrowe (włókna mięśniowe). Jądro dzieli się na otoczkę jądrową, chromatynę, jąderko i nukleoplazmę (ryc. 4).
Otoczka jądrowa, czyli karioteka, oddzielająca zawartość jądra od cytoplazmy, składa się z wewnętrznej i zewnętrznej błony jądrowej, każda o grubości 8 nm. Błony oddzielone są przestrzenią okołojądrową (cystern karyoteca) o szerokości 20-50 nm, w której znajduje się drobnoziarnisty materiał o umiarkowanej gęstości elektronowej. Zewnętrzna błona jądrowa przechodzi do ziarnistej siateczki śródplazmatycznej. Poeto Przestrzeń okołojądrowa mu tworzy pojedynczą wnękę z retikulum endoplazmatycznym. Wewnętrzna błona jądrowa jest połączona od wewnątrz z rozgałęzioną siecią włókienek białkowych, składających się z pojedynczych podjednostek.
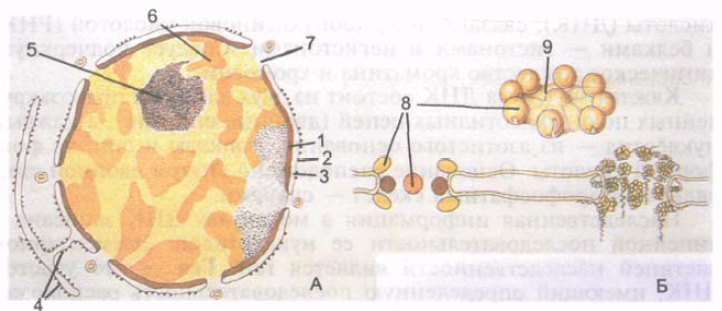
Ryż. 4. Jądro komórkowe (A) i kompleks porów jądrowych (B). 1 - wewnętrzna błona jądrowa; 2 - zewnętrzna błona jądrowa; 3 - przestrzeń okołojądrowa; 4 - błony siateczki śródplazmatycznej; 5 - jąderko; 6 - zdekondensowana chromatyna; 7 - czas nuklearny; 8 - granulki porów jądrowych; 9 - membrana porów jądrowych.
Otoczka jądrowa zawiera wiele okrągłych porów jądrowych, każdy o średnicy 50-70 nm. Pory jądrowe zajmują do 25% powierzchni jądra. Liczba porów w jednym jądrze sięga 3000-4000. Na krawędziach porów membrana zewnętrzna i wewnętrzna są ze sobą połączone i tworzą tzw. pierścień porów. Każdy por jest zamknięty przeponą, zwaną także kompleksem porów. Membrany porów mają złożoną strukturę, tworzą je połączone ze sobą granulki białkowe. Przez pory jądrowe następuje selektywny transport dużych cząstek, a także wymiana substancji pomiędzy jądrem a cytozolem komórki.
Pod otoczką nuklearną znajdują się nukleoplazma (karioplazma)(jądroplazma, s. karioplazma), który ma jednorodną strukturę i jąderko. W nukleoplazmie jądra niedzielącego się, w jego jądrowej macierzy białkowej, znajdują się osmiofilne granulki (grudki) tzw. heterochromatyny. Obszary luźniejszej chromatyny znajdujące się pomiędzy ziarnistościami nazywane są euchromatyną. Chromatyna luźna zwana jest także chromatyną zdekondensowaną, w niej zachodzą najintensywniej procesy syntetyczne. Podczas podziału komórki chromatyna zagęszcza się, kondensuje i tworzy chromosomy.
Chromatyna(chromatyna) jądra niedzielącego się, a chromosomy jądra dzielącego się tworzą cząsteczki kwasu dezoksyrybonukleinowego (DNA) związanego z kwasem rybonukleinowym (RNA) oraz białkami - histonami i niehistonami. Należy podkreślić chemiczną tożsamość chromatyny i chromosomów.
Każda cząsteczka DNA składa się z dwóch długich, prawoskrętnych łańcuchów polinukleotydowych (podwójnych helis), a każdy nukleotyd składa się z zasady azotowej, glukozy i reszty kwasu fosforowego. Podstawa znajduje się wewnątrz podwójnej helisy, a szkielet cukrowo-fosforanowy znajduje się na zewnątrz.
Informacja dziedziczna w cząsteczkach DNA jest zapisana w liniowej sekwencji jej nukleotydów. Elementarną cząstką dziedziczności jest gen. Gen to odcinek DNA posiadający określoną sekwencję nukleotydów odpowiedzialnych za syntezę jednego, określonego białka.
Cząsteczka DNA w jądrze jest zwarta. Zatem jedna cząsteczka DNA zawierająca 1 milion nukleotydów przy ich liniowym układzie zajmowałaby odcinek o długości zaledwie 0,34 mm. Długość jednego ludzkiego chromosomu po rozciągnięciu wynosi około 5 cm, ale w stanie zwartym chromosom ma objętość około 10-15 cm 3.
Cząsteczki DNA związane z białkami histonowymi tworzą nukleosomy, które są jednostkami strukturalnymi chromatyny. Nukleosom ma postać kulki o średnicy 10 nm. Każdy nukleosom składa się z histonów, wokół których skręcony jest odcinek DNA, obejmujący 146 par nukleotydów. Pomiędzy nukleosomami znajdują się liniowe odcinki DNA składające się z 60 par nukleotydów.
Chromatyna jest reprezentowana przez włókienka tworzące pętle o długości około 0,4 μm, zawierające od 20 000 do 30 000 par nukleotydów.
W wyniku zagęszczenia (kondensacji) i skręcenia (superspecjalizacji) deoksyrybonukleoprotein (DNP) w dzielącym się jądrze, chromosomy stają się widoczne. Struktury te są chromosomy(chromasomae, od greckiego chruma – farba, soma – ciało) – to wydłużone formacje w kształcie pręta, posiadające dwa ramiona oddzielone tzw. przewężeniem – centromerem. W zależności od położenia centromeru oraz względnego położenia i długości ramion (nog) wyróżnia się trzy typy chromosomów: metacentryczny, posiadający w przybliżeniu te same ramiona; submetacentryczny, w którym długość ramion jest inna; akrocentryczny, w którym jedno ramię jest długie, a drugie bardzo krótkie, ledwo zauważalne. Chromosom ma regiony eu- i heterochromatyczne. Te ostatnie pozostają zwarte w niedzielącym się jądrze i we wczesnej profazie mitozy. Do identyfikacji chromosomów wykorzystuje się naprzemienność regionów eu i heterochromatycznych.
Powierzchnia chromosomów pokryta jest różnymi cząsteczkami, głównie rybonukleoproteinami (RNP). W komórkach somatycznych występują 2 kopie każdego chromosomu, nazywane są one homologicznymi. Są identyczne pod względem długości, kształtu, budowy i niosą te same geny, które są zlokalizowane w ten sam sposób. Cechy strukturalne, liczba i wielkość chromosomów nazywane są kariotypem. Prawidłowy ludzki kariotyp obejmuje 22 pary autosomów i jedną parę chromosomów płciowych (XX lub XY). Ludzkie komórki somatyczne (diploidalne) mają podwójną liczbę chromosomów – 46. Komórki płciowe zawierają haploidalny (pojedynczy) zestaw – 23 chromosomy. Dlatego w komórkach rozrodczych jest 2 razy mniej DNA niż w diploidalnych komórkach somatycznych.
Jądro(jąderko), jeden lub więcej, wykrywa się we wszystkich niedzielących się komórkach. Ma wygląd intensywnie zabarwionego okrągłego korpusu, którego wielkość jest proporcjonalna do intensywności syntezy białek. Jąderko składa się z jąderka o dużej gęstości elektronowej (od greckiego peta - nić), w którym wyróżnia się część nitkowata (fibrylarna), składająca się z wielu przeplatających się nici RNA o grubości około 5 nm oraz część ziarnista. Część ziarnistą (ziarnistą) tworzą ziarna o średnicy około 15 nm, które są cząstkami RNP - prekursorami podjednostek rybosomalnych. Chromatyna okołojądrowa jest osadzona w zakamarkach jąderka. Rybosomy powstają w jąderku.
PODZIAŁ KOMÓREK. CYKL KOMÓRKOWY
Wzrost organizmu następuje w wyniku wzrostu liczby komórek w wyniku podziału. Głównymi metodami podziału komórek w organizmie człowieka są mitoza i mejoza. Procesy zachodzące podczas tych metod podziału komórek przebiegają w ten sam sposób, ale prowadzą do różnych rezultatów.
Mitotyczny podział komórek(mitoza) prowadzi do wzrostu liczby komórek, wzrostu organizmu. W ten sposób zapewniona jest odnowa komórek w przypadku ich zużycia lub śmierci. Obecnie wiadomo, że komórki naskórka żyją 10-30 dni, erytrocyty - do 4-5 miesięcy. Komórki nerwowe i mięśniowe (włókna) żyją przez całe życie człowieka.
Podczas rozmnażania (podziału) wszystkie komórki podlegają zmianom mieszczącym się w ramach cyklu komórkowego. Cykl komórkowy nazywaj procesy zachodzące w komórce od podziału do podziału lub od podziału do śmierci (śmierci) komórki. Cykl komórkowy dzieli się na przygotowanie komórki do podziału (interfaza) i mitozę (proces podziału komórki).
W interfazie, która trwa około 20-30 godzin, zwiększa się tempo procesów biosyntezy i zwiększa się liczba organelli. W tym czasie masa komórki i wszystkich jej elementów strukturalnych, w tym centrioli, podwaja się.
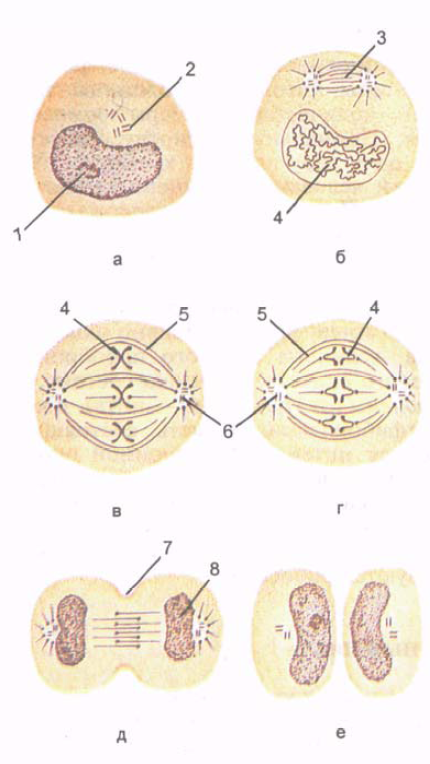
Ryż. 5. Podział komórek. Etapy mitozy. Pokazano powstawanie chromosomów, tworzenie wrzeciona oraz równomierne rozmieszczenie chromosomów i centrioli w dwóch komórkach potomnych.
A - interfaza; B - profaza; B - metafaza; G - anafaza; D - telofaza; E - późna telofaza. 1 - rdzeń; 2 - centrum komórki (centriola); 3 - wrzeciono podziału komórkowego; 4 - chromosomy; 5 - ciągłe mikrotubule; 6 - centrum komórki; 7 - bruzda podziału komórek; 8 - tworzenie rdzenia.
Następuje replikacja (powtórzenie, podwojenie) cząsteczek kwasu nukleinowego. Jest to proces przekazywania informacji genetycznej zapisanej w DNA rodzicielskim poprzez dokładne odtworzenie jej w komórkach potomnych. Macierzysta nić DNA służy jako matryca do syntezy DNA potomnego. W wyniku replikacji każda z dwóch cząsteczek potomnego DNA składa się z jednej starej i jednej nowej nici. Przygotowując się do mitozy, komórka syntetyzuje białka niezbędne do podziału komórki. Pod koniec interfazy chromatyna w jądrze ulega kondensacji.
Mitoza(mitoza; od greckiego mitos – nić) to okres, w którym komórka macierzysta dzieli się na dwie komórki potomne (ryc. 5). Mitotyczny podział komórek zapewnia równomierny rozkład struktur komórkowych i ich substancji jądrowej – chromatyny – pomiędzy dwiema komórkami potomnymi. Czas trwania mitozy wynosi od 30 minut do 3 godzin.Mitozę dzieli się na profazę, metafazę, anafazę i telofazę.
W profazie jąderko stopniowo się rozpada, a centriole rozprzestrzeniają się na bieguny komórki. Mikrotubule centrioli są skierowane w stronę równika, a w obszarze równikowym zachodzą na siebie.
W metafazie błona jądrowa ulega zniszczeniu, nici chromosomalne kierowane są do biegunów, utrzymując połączenie z równikiem
nalny obszar komórki. Struktury retikulum endoplazmatycznego i kompleksu Golgiego rozpadają się na małe pęcherzyki (pęcherzyki), które wraz z mitochondriami są rozmieszczone w obu połówkach dzielącej się komórki. Pod koniec metafazy każdy chromosom zaczyna się dzielić na dwa nowe chromosomy potomne.
W anafazie chromosomy oddzielają się od siebie i przemieszczają w kierunku biegunów komórki z prędkością do 0,5 μm/min. Pod koniec anafazy błona komórkowa wnika wzdłuż równika komórki, prostopadle do jej osi podłużnej, tworząc bruzdę szczelinową.
W telofazie chromosomy, które rozproszyły się na biegunach komórki, ulegają dekondensacji, przechodzą do chromatyny i rozpoczyna się transkrypcja (produkcja) RNA. Tworzą się otoczka jądrowa i jąderko oraz szybko tworzą się struktury błonowe przyszłych komórek potomnych. Na powierzchni komórki, wzdłuż jej równika, zwężenie pogłębia się, komórka dzieli się na dwie komórki potomne.
Dzięki podziałowi mitotycznemu komórki potomne otrzymują zestaw chromosomów identyczny z komórką matki. Mitoza zapewnia stabilność genetyczną, wzrost liczby komórek, a co za tym idzie, wzrost organizmu i procesy regeneracyjne.
Mejoza(z greckiej mejozy - redukcja) obserwuje się w komórkach rozrodczych. W wyniku podziału tych komórek powstają nowe komórki z pojedynczym (haploidalnym) zestawem chromosomów, co jest istotne dla przekazywania informacji genetycznej. Kiedy jedna komórka płciowa łączy się z komórką płci przeciwnej (podczas zapłodnienia), zestaw chromosomów podwaja się, stając się kompletnym, podwójnym (diploidalnym). Diploidalna (dwujądrowa) zygota powstała po fuzji komórek rozrodczych zawiera dwa zestawy identycznych (homologicznych) chromosomów. Każda para homologicznych chromosomów organizmu diploidalnego (zygoty) pochodzi z jądra komórki jajowej i jądra plemnika.
W wyniku mejozy komórek rozrodczych w dojrzałym organizmie każda komórka potomna zawiera tylko jedną ze wszystkich par homologicznych chromosomów komórek wyjściowych. Staje się to możliwe, ponieważ podczas mejozy zachodzą jedynie replikacja DNA i dwa kolejne podziały jądrowe. W rezultacie z jednej komórki diploidalnej powstają dwie komórki haploidalne. Każda z tych komórek potomnych zawiera o połowę mniej chromosomów (23) niż jądro komórki macierzystej (46). W wyniku mejozy haploidalne komórki rozrodcze mają nie tylko liczbę chromosomów zmniejszoną o połowę, ale także inny układ genów na chromosomach. Dlatego nowy organizm niesie ze sobą nie tylko sumę cech swoich rodziców, ale także własne (indywidualne) cechy.
Pytania do powtórzenia i samokontroli
1. Z jakich elementów składa się jądro komórkowe? Jakie funkcje pełni?
2. Opowiedz nam o budowie cząsteczek DNA.
3. Podaj cechy morfologiczne chromosomów, podaj ich klasyfikację.
4. Czym jest cykl komórkowy, jakie okresy (fazy) wyróżniają się w tym cyklu?
5. Czym jest mejoza, czym różni się od mitozy?