Цитоплазма каждой клетки окружена мембраной, которая ограничивает ее от окружающей среды. Цитоплазматическая мембрана (ЦПМ) является исключительно полифункциональной структурой:
1. ЦПМ воспринимает всю химическую информацию , поступающую в клетку из внешней среды.
2. Она является основным осмотическим барьером , благодаря которому внутри клетки поддерживается определенное осмотическое давление.
Клеточная стенка выполнена в основном из пептидогликана, перемежающегося с тейхоиновой кислотой, которая соединяет разные слои вместе. Количество сшивки выше, а стенка толще, чем в грамотрицательных клеточных стенках. Грамположительные клетки состоят почти полностью пептидогликаном, но важной структурой, обнаруженной в грамположительных клеточных стенках, является тейхоиновая кислота. Это фосфодиэфирный полимер глицерина или рибитола, соединенный фосфатными группами. Тейхоевая кислота ковалентно связана с мураминовой кислотой и стекает вместе с различными слоями пептидогликановой сетки.
3. ЦПМ совместно с клеточной стенкой участвует в регуляции роста и клеточного деления бактерий.
4. ЦПМ участвует в регуляции процессов репликации и сегрегации хромосом и плазмид (они связаны с ее рецепторами).
5. В ЦПМ содержится значительное количество ферментов, в том числе системы переноса электронов (ЦПМ - место генерации энергии у бактерий).
Тейхоевая кислота стабилизирует клеточную стенку и делает ее сильнее. Химическая формула тейхоевой кислоты показана на рисунке 25. Тейхоевая кислота представляет собой длинную тонкую молекулу, которая ткут через пептидогликан. Грамотрицательные клеточные стенки имеют более сложную структуру, чем у грамположительных организмов. Вне цитоплазматической мембраны находится периплазма, содержащая тонкий слой пептидогликана. Пептидогликан в грамотрицательных клетках содержит меньше сшивки, чем в грамположительных клетках без пептидного линкера.
6. С ЦПМ связаны жгутики и аппарат регуляции их движения.
7. ЦПМ участвует в процессах транспорта (в том числе активного) питательных веществ в клетку и продуктов жизнедеятельности, включая ферменты и экзотоксины, из клетки в окружающую среду. В ней содержатся белки, участвующие в облегченной диффузии и активном транспорте.
Ковалентно связанным с пептидогликаном является липопротеин Брауна, который имеет гидрофобный анкер, который помогает сильно связывать пептидогликан с наружной мембраной. На рисунке 26 показано расположение грамотрицательной клеточной стенки. Клеточная стенка в грамотрицательных бактериях содержит гораздо меньше пептидогликана и окружена наружной мембраной. Между пептидогликаном гораздо меньше сшивок.
Наружная мембрана грамотрицательных бактерий представляет собой другой липидный бислой, подобный цитоплазматической мембране, и содержит белки, а также. Это барьер для белков и предотвращает отложение ферментов, выделяемых в периплазму. Это отталкивание позволяет этим бактериям выживать в этой среде.
8.
ЦПМ играет важную роль в компартментализации и стабилизации рибосом
.
9.
ЦПМ участвует в синтезе компонентов клеточной стенки.
10.
ЦПМ участвует в образовании мезосом (мезосомы образуются в результате инвагинации участка ЦПМ в цитоплазму, они открыты в периплазматическое пространство).
Каким образом мембрана осуществляет на молекулярном уровне свои многочисленные функции - один из актуальнейших вопросов современной биологии. На долю ЦПМ приходится около 10% сухого веса бактерий. Она содержит 25-40% фосфолипидов, образующих два слоя , 20-75% белков и до 6% утлеводов. Молекулы фосфолипидов асимметричны : головки, несущие электрический заряд, гидрофильны; хвостики - нейтральны и гидрофобны. Фосфолипиды упакованы в мембране следующим образом: их полярные гидрофильные головки обращены наружу и образуют два слоя ЦПМ - внутренний и внешний, а неполярные гидрофобные хвостики скрыты в толще мембраны. На электронограммах ЦПМ имеет вид трехслойной структуры, состоящей из двух 2 параллельных темных слоев и разделяющего их светлого слоя. Этот слой более проницаем для электронов, чем слои, состоящие из полярных концов фосфолипидов, ассоциированных с белками. Специфичность функций ЦПМ во многом зависит от набора содержащихся в них белков. Расположение их в ЦПМ своеобразно: некоторые белки пронизывают весь двойной липидный слой, определенная часть белков связана или только с внутренней, или только с наружной поверхностью мембраны. Это вытекает из того, что взаимодействие между мембраной и цитоплазмой, с одной стороны, мембраной и внешней средой, с другой - определяет различные, хотя и взаимосвязанные процессы ее жизнеобеспечения: облегченная диффузия, активный транспорт, элементарной биологической мембраны.
Он создает широкий спектр физиологических реакций, включая индукцию лихорадки, изменения количества лейкоцитов, утечку из кровеносных сосудов, некроз опухолей и пониженное кровяное давление, приводящее к обрушению сосудов и, в конечном итоге, шоку. В наружной мембране меньше белков и меньше уникальных типов белков, чем в цитоплазматической мембране. Порины являются особенно важными компонентами из-за их роли в проницаемости внешней мембраны для малых молекул. Порины - это белки, которые образуют поры во внешней мембране, достаточно большие, чтобы пропускать самые маленькие гидрофильные молекулы.
При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее, считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т. е. в процессах с высокой затратой энергии.
На рис. 28 показана структура порина в молекулярном масштабе. Все известные порины имеют сходную структуру, причем белок содержит центральный канал, который позволяет проходить молекулы. Это позволяет переносить эти молекулы в периплазматическое пространство для возможного переноса через цитоплазматическую мембрану. Некоторые порины в наружной мембране являются общими, делают простую дискриминацию по размеру и заряду, но имеют небольшую специфичность. Другие порины более специфичны.
Очень большие или гидрофобные молекулы не могут проникать во внешнюю мембрану, поэтому наружная мембрана служит барьером проницаемости по меньшей мере для некоторых молекул. Молекулярная структура порина. Вид внутри находится снаружи ячейки, смотрящей на поверхность мембраны. Взгляд - это перспектива со стороны. Порин имеет три субъединицы белка, и фактическая пора является центральной треугольной областью в верхней панели, образованной тремя субъединицами.
Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 8OS-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы -- 50S и 30S. Рибосомные РНК (рРНК.) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК. входит в состав малой субъединицы рибосом, a 23S рРНК- в состав большой субъединицы рибосом. Изучение 16S рРНК. является основой геносистематики, позволяя оценить степень родства организмов.
Существуют также другие типы белков наружной мембраны, которые участвуют в различных функциях. Некоторые патогены содержат белки наружной мембраны, которые помогают им нейтрализовать защиту хозяев. Различия между стенками клеток грамотрицательных и грамотрицательных бактерий в значительной степени влияют на успех микробов в их среде. Толстая клеточная стенка грамотрицательных клеток позволяет им лучше справляться в сухих условиях, потому что это уменьшает потерю воды. В таблице 3 суммируется разница между грамотрицательными и грамотрицательными клеточными стенками.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Все запасные вещества присутствуют в клетки в химически инертной форме. Такое состояние препятствует нарушению осмостаза клеточного содержимого. Некоторые включения просто лежат в цитоплазме, другие окружены тонкой мембраной. Мембрана обычно белковой природы, но иногда может содержать и липиды. Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей.
Таблица 3 Свойства клеточных стенок
Резюме различий между грамотрицательными и грамотрицательными клеточными стенками. Для большинства бактериальных клеток клеточная стенка имеет решающее значение для выживаемости клеток, но есть некоторые бактерии, у которых нет клеточных стенок. Виды микоплазмы являются широко распространенными примерами, а некоторые могут быть внутриклеточными патогенами, которые растут внутри их хозяев. Стены клеток здесь не нужны, потому что клетки живут только в контролируемой осмотической среде других клеток.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в красно-фиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронно-плотных гранул размером 0,1-1,0 мкм.
Вероятно, у них была способность сформировать клеточную стену в какой-то момент в прошлом, но поскольку их образ жизни стал одним из бытия внутри других клеток, они потеряли способность создавать стены. В соответствии с этим очень ограниченным образом жизни в других клетках эти микробы также очень малы. Они не нуждаются в генах для всех видов биосинтетических ферментов, так как они могут украсть конечные компоненты этих путей от хозяина. Точно так же они не нуждаются в генах, кодирующих много разных путей для различных источников углерода, азота и энергии, поскольку их внутриклеточная среда полностью предсказуема.
Нуклеоид - эквивалент ядра у бактерий. В 1956 году внутри бактериальных клеток была обнаружена «ядерная зона», или нуклеоид, где размещена бактериальная хромосома. Бактериальная ДНК обнаружена в кольцевой и линейной формах. Для клеток E.coli доказано, что ДНК существует в виде кольцевой молекулы, в то время как для Borrelia burgdorferi в 1989 г. показано, что клетки содержат ДНК в линейной форме. Нуклеоид, в отличие от ядра эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, но при нарушении деления в ней может находиться 4 и более копий одной хромосомы. У некоторых видов в клетки обнаружены две и три неидентичных хромосомы. У видов Vibrio, Leptospira interrogans обнаружены две кольцевые хромосомы. Поэтому идея, что бактерии содержат только одну хромосому, считается устаревшей. Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Наличие стеринов в мембране способствует их долговечности, помогая увеличить силы, удерживающие мембрану вместе. Потому что не было бы людей, растений, животных, грибов или простейших. Все эти организмы - каждое живое существо, помимо бактерий, - состоят из эукариотических клеток.
И генезис этого типа клеток не был бы возможен без симбиоза. Образование эукариотических клеток объясняется в науке так называемой теорией эндосимбионтов. Согласно этой теории, сложные клетки с клеточным ядром, отделенным от плазмы и клеточных органелл, были сформированы из симбиоза более простых структурированных прокариотов. Только бактерии, которые жили как эндосимбионтен в большей клетке, образовали органеллы сегодняшних эукариотических клеток.
Кроме нуклеоида в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды , представляющие собой ковалентно замкнутые кольца ДНК.Они кодируют не основные для жизнедеятельности бактериальной клетки функции, но придающие бактерии преимущества при попадании в неблагоприятные условия существования.
Образование митохондрий
Это накопление органического вещества является долгожданным источником питательных веществ для гетеротрофных клеток. Клетки приобретают размеры и, помимо того, что они могут поглощать органическое вещество фагоцитозом, способны поглощать органические клетки, а также меньшие клетки фагоцитозом. Как правило, эти клетки оказываются в виде пищи, они перевариваются.
Иногда, однако, бывает, что бактерии, взятые в живых, выживают внутри большей клетки и взаимодействуют со своим хозяином. Развивается симбиоз, в котором клетка-хозяин может более эффективно использовать пищевые продукты с помощью ферментов из дыхательной цепи аэробных бактерий. С другой стороны, эндосимбионт более защищен внутри большей клетки от изменения окружающей среды. Другим возможным источником митохондрий будет вторжение бактериальных паразитов в клетку-хозяина и полученный симбиоз.
Капсула, микрокапсула, слизь . Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. Поверхность колоний клеток с капсулами выглядит гладкой, влажной, блестящей.
Органные клетки с несколькими мембранами
Неизвестно с абсолютной уверенностью, действительно ли возникли эукариоты таким образом, но существует множество фактов, которые поддерживают теорию эндосимбионта. Кроме того, органеллы не могут быть сформированы из генетической информации ядра, но умножаются на разделение. Процесс деления похож на процесс деления бактерий. Рибосомы клеточных органелл сходны по структуре с бактериальными, а не эукариотическими рибосомами.
Эндосимбиозы все еще встречаются сегодня. Таким образом, некоторые морские диатомовые водоросли могут, по-видимому, жить в симбиозе с дефицитными цианобактериями. Опять же, симбионт интегрирован в плазму клетки-хозяина. Поэтому даже сегодня этот вид симбиоза повторяется как процесс, который стоял в начале всей высшей жизни.
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют гликокаликсом. Кроме синтеза экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Характеризуется отсутствием ядра. Их основное разделение происходит в двух и Археях. В повседневной жизни прокариоты известны нам как бактерии. Часто слово «бактерии» или бактерии прямо приравнивается к патогенам. Меньше всего прокариотов являются патологическими - то есть патогенными.
Клеточная стенка окружает цитоплазматическую мембрану и внутреннюю часть клетки. Внутренняя отделка отсутствует. Все прокариоты имеют одну и ту же основную структуру. Большинство прокариотов имеют клеточную стенку, расположенную вне плазматической мембраны и состоящую из производного углевода. В зависимости от количества и расположения слоев мюреина проводится различие между грамположительными и грамотрицательными бактериями.
- Существует плазменная мембрана, окружающая клетку.
- После этого типа архея не имеет клеточной стенки.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка . Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей :
В принципе, клетка, окруженная мембраной, далее не подразделяется. В общем, есть механизм, который позволяет двигаться. Он имеет на цитоплазматической мембране, лежащей на другом клеточном слое. Это сделано из мурейна. На ней лежит внешняя оболочка. Осажденная одноклеточная ввела зеленую водоросль для субтензии. Симбиоз вызывает резкие изменения в обеих клетках. Одноклеточный организм переключается между фазой хищника и фазой хозяина с эндосимбионтом, переключая свой энергетический баланс с гетеротрофного на автотрофный, т.е. от подачи к солнечной энергии.
1. - спиралевидной нити,
2. - крюка
3. - базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары - у грамотрицательных бактерий) и моторными белками (рис).
Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. У бактерий жгутики правовращающиеся. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
В процессе адаптации двух клеток растение появляется из простейших с частицами; Исследователи назвали их Хатена, слишком таинственной на немецком языке. Это наблюдение является дополнительной поддержкой эндосимбиотической теории, согласно которой предшественники всех клеток растений и животных были образованы эндосимбиозом.
Какие органеллы получили предшественники растительных клеток в соответствии с Эндосомбионтенторией Эндосимбиозой? Назовите четыре черты этих органелл, которые поддерживают эндосимбиотическую теорию! В каждом случае оправдываем, почему вы рассматриваете характеристику как доказательство теории эндосимбионтов. В какой мере можно рассмотреть открытие новых водорослей Хатена в качестве основы теории эндосимбионтов? При эндосимбиозе с аэробными бактериями первичные эукариоты приобрели способность к окислительной деградации путем симбиоза с цианобактериями, способностью к фотосинтезу. Они не могут возникнуть снова. Это говорит о том, что они первоначально не происходили как структуры клетки-хозяина. Митохондрии и хлоропласты имеют свой собственный механизм синтеза белка, и их рибосомы построены как прокариоты. Оба они окружены двумя мембранами, внутренняя одна из которых похожа на прокариотическую мембрану, внешнюю из клеточной мембраны. Важным признаком для теории эндосинбионта является тот факт, что между прокариотическими и эукариотическими клетками нет промежуточных форм, но между эндосимбиотическими организмами и органеллами существуют промежуточные формы. Такая переходная форма представляет собой наблюдаемую одноклеточную водоросль: после преобразования диеты одноклеточная зависит от симбиоза с зеленой водорослью, тогда как только зеленая водоросль является жизнеспособной.
- Какие особые навыки вы приобрели?
- Митохондрии и хлоропласты являются потомками микроорганизмов.
- Митохондрии происходят от бактерий, хлоропластов из цианобактерий.
- Оба возникают только путем разделения их сверстников.
Жгутики состоят из белка - флагеллина (от. flagellum - жгутик), являющегося антигеном - так называемый Н-антиген . Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих ) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих ), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования, более тонкие и короткие, чем жгутики . Пили отходят от поверхности клетки и состоят из белка пилина, который организован в спиральную структуру. Архитектура пилей варьирует от тонких нитевидных до толстых прочных палочкообразных образований с осевыми отверстиями. Они обладают антигенной активностью. Различают пили, ответственные за адгезию, т. е. за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водно-солевой обмен, и половые (F-пили) или пили 4-го типа, или конъюгационные, пили.
Обычно пили многочисленны - несколько сотен на клетку. Однако половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Со/-плазмиды). Отличительной особенностью половых пилей является их взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях (рис.).
Основное предназначение пилей поддерживать на своих концах специфические прикрепительные структуры (адгезины). Адгезины являются посредниками при бактериальных контактах, при контактах с неживыми объектами, тканями и клетками восприимчивых организмах. Колонизация тканей хозяина бактериальными патогенами обычно зависит от стереохимического подобия между архитектурой адгезина и соответствующего рецептора клетки хозяина (рис). Таким образом, изучение функционирования пилей позволит не только глубже понять механизм колонизации и передачи сигналов, но и вести разработку новых поколений антимикробных препаратов.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки (рис.).
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных вешеств и др.). Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов.
Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (лат. clostridium - веретено). Споры кислотоустойчивы , поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена в красный, а вегетативная клетка - в синий.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга.
Форма спор может быть овальной, шаровидной; расположение в клетке :
1. терминальное, т. е. на конце палочки (у возбудителя столбняка),
2. субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены)
3. центрально е (у сибиреязвенной бациллы).
Процесс спорообразованя (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора.
Проспору окружают две цитоплазматические мембраны, между которыми формируется толстый измененный пептидогликановый слой кортекса (коры). Изнутри он соприкасается с клеточной стенкой споры, а снаружи - с внутренней оболочкой споры. Наружная оболочка споры образована вегетативной клеткой.
Споры некоторых бактерий имеют дополнительный покров - экзоспориум.
Таким образом формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойную оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии (рис):
1. активацию
2. инициацию
3 . вырастание
При этом из одной споры образуется одна бактерия.
Активация - это готовность к прорастанию. При температуре 60-80°С спора активируется для прорастания.
Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.
Вопросы для самоконтроля
| | | | | | | | | 10 | | | | | |
Строение типичной бактериальной клетки показано на рис. 2.3. На рис. 2.4 представлена электронная микрофотография среза палочковидной бактерии. Можно видеть, насколько просто устроена бактериальная клетка, особенно если сравнить ее с клетками эукариот (рис. 7.5 и 7.6).
Рис. 2.3. Обобщенная схема строения клетки палочковидной бактерии. Справа перечислены структуры, встречающиеся в каждой клетке, слева - встречающиеся не во всех клетках. Жгутик бывает один, как у Rhizobium, или несколько, как у Azotobacter; обычно он длиннее клетки. Капсула может быть слизистой, как у Azotobacter; если капсула рыхлая, то она называется слизистым слоем . Трубчатые или мешковидные фотосинтетические мембраны , содержащие пигменты, представляют собой впячивания плазматической мембраны; у фотосинтезирующих бактерий, например у Chromatium, такие мембраны рассеяны по всей цитоплазме. Число пилей , или фимбрий , может достигать от одной до нескольких сотен, как, например, у Escherichia coli, Salmonella. Мезосома представляет собой многоскладчатое впячивание плазматической мембраны, как, например, у Bacillus subtilis. Клеточная стенка жесткая и содержит муреин. Рибосомы , располагающиеся по всей цитоплазме, по размеру меньше, чем у эукариот. Из запасных питательных веществ в бактериальных клетках можно обнаружить липиды, гликоген, полифосфаты (волютиновые гранулы). Цитоплазма не содержит никаких органелл; содержит ферменты и т. п
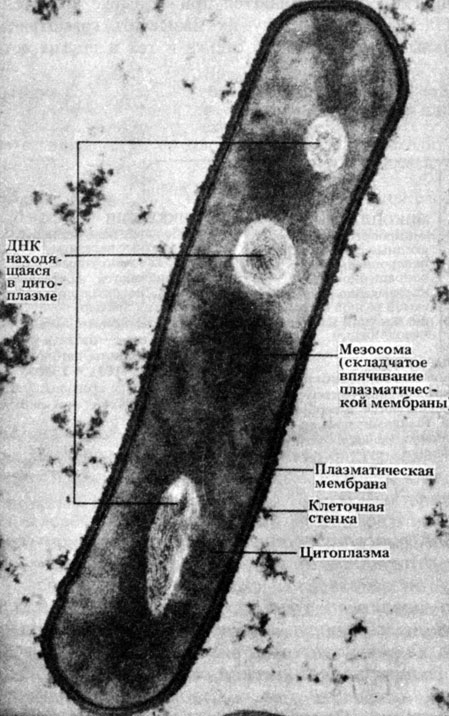
Рис. 2.4. Электронная микрофотография среза типичной палочковидной бактерии Bacillus subtilis. В светлых зонах находится ДНК. × 50000
Капсулы и слизистые слои
Капсулы и слизистые слои - это слизистые или клейкие выделения некоторых бактерий; такие выделения хорошо видны после негативного контрастирования (когда окрашивают не препарат, а фон). Капсула представляет собой относительно толстое и компактное образование, а слизистый слой намного рыхлее. В некоторых случаях слизь служит для формирования колоний из отдельных клеток. И капсула, и слизистые слои служат дополнительной защитой для клеток. Так, например, инкапсулированные штаммы пневмококков свободно размножаются в организме человека и вызывают воспаление легких, а некапсулированные штаммы легко атакуются и уничтожаются фагоцитами и поэтому совершенно безвредны.
Клеточная стенка
Клеточная стенка придает клетке определенную форму и жесткость. Ее хорошо видно на срезе (рис. 2.4). Как и у растений, клеточная стенка бактерий препятствует осмотическому набуханию и разрыву клеток, когда они, как это часто случается, попадают в гипотоническую среду (Приложение разд. П.1.5). Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот. Кроме того, клеточная стенка обладает антигенными свойствами, которые ей придают содержащиеся в ней белки и полисахариды.
По строению клеточной стенки бактерий можно разделить на две группы. Одни окрашиваются по Граму, поэтому их называют грамположительными , а другие обесцвечиваются при отмывке красителя (разд. 2.7), и поэтому их называют грамотрицательными . В клеточной стенке и тех и других есть особая жесткая решетка, состоящая из муреина . Молекула муреина представляет собой правильную сеть из параллельно расположенных полисахаридных цепей, сшитых друг с другом короткими цепями пептидов. Таким образом, каждая клетка окружена сетевидным мешком, составленным всего из одной молекулы. (Полисахаридная часть муреина описана в табл. 5.7).
У грамположительных бактерий, например у Lactobacillus, в муреиновую сетку встроены другие вещества, главным образом полисахариды и белки. Так вокруг клетки создается сравнительно толстая и жесткая упаковка. У грамотрицательных бактерий, скажем у Escherichia coli или у Azotobacter, клеточная стенка гораздо тоньше, но устроена она сложнее. Муреиновый слой у этих бактерий снаружи покрыт мягким и гладким слоем липидов. Это защищает их от лизоцима . Лизоцим обнаружен в слюне, слезах и других биологических жидкостях, а также в белке куриного яйца. Он катализирует гидролиз определенных связей между остатками углеводов и таким образом расщепляет полисахаридную основу муреина. Клеточная стенка разрывается, и, если клетка находится в гипотоническом растворе, происходит ее лизис (клетка осмотически набухает и лопается). Липидный слой придает клетке устойчивость и к пенициллину. Этот антибиотик препятствует образованию сшивок в клеточной стенке грамположительных бактерий, что делает растущие клетки более чувствительными к осмотическому шоку.
Жгутики
Многие бактерии подвижны, и эта подвижность обусловлена наличием у них одного или нескольких жгутиков. Жгутики у бактерий устроены гораздо проще, чем у эукариот (разд. 17.6.2, табл. 2.1), и по своей структуре напоминают одну из микротрубочек эукариотического жгутика. Жгутики состоят из одинаковых сферических субъединиц белка флагеллина (похожего на мышечный актин), которые расположены по спирали и образуют полый цилиндр диаметром около 10-20 нм. Несмотря на волнистую форму жгутиков, они довольно жестки.
Жгутики приводятся в движение посредством уникального механизма. Основание жгутика, по-видимому, вращается так, что жгутик как бы ввинчивается в среду, не совершая беспорядочных биений, и таким образом продвигает клетку вперед. Это, очевидно, единственная известная в природе структура, где используется принцип колеса. Другая интересная особенность жгутиков - это способность отдельных субъединиц флагеллина спонтанно собираться в растворе в спиральные нити. Спонтанная самосборка - очень важное свойство многих сложных биологических структур. В данном случае самосборка целиком обусловлена аминокислотной последовательностью (первичной структурой) флагеллина.
Подвижные бактерии могут передвигаться в ответ на определенные раздражители, т. е. они способны к таксису. Так, например, аэробные бактерии обладают положительным аэротаксисом (т. е. плывут туда, где среда богаче кислородом), а подвижные фотосинтезирующие бактерии - положительным фототаксисом (т. е. плывут к свету).
Жгутики легче всего рассмотреть в электронном микроскопе (рис. 2.5), применив технику напыления металлом (разд. П.2.5).
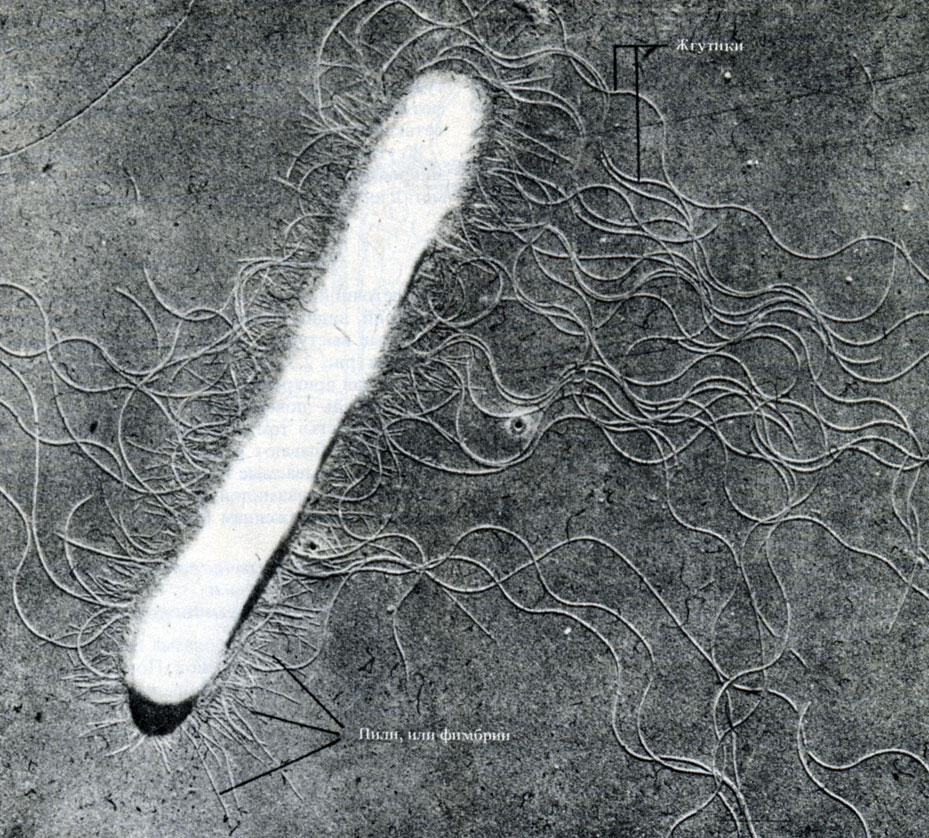
Рис. 2.5. Микрофотография палочковидной бактерии, полученная с помощью просвечивающего электронного микроскопа. Хорошо видны клеточная стенка, фимбрии и длинные волнистые жгутики, × 28000
Пили, или фимбрии
На клеточной стенке некоторых грамотрицательных бактерий видны тонкие выросты (палочковидные белковые выступы), которые называются пили или фимбрии (рис. 2.5). Они короче и тоньше жгутиков и служат для прикрепления клеток друг к другу или к какой-нибудь поверхности, придавая специфическую "липкость" тем штаммам, которые ими обладают. Пили бывают разного типа. Наиболее интересны так называемые F-пили, которые кодируются специальной плазмидой (разд. 2.2.4) и связаны с половым размножением бактерий.
Плазматическая мембрана, мезосомы и фотосинтетические мембраны
Как у всех клеток, протоплазма бактерий окружена полупроницаемой мембраной. По структуре и функциям плазматические мембраны бактерий не отличаются от мембран эукариотических клеток (разд. 7.2.1). У некоторых бактерий плазматическая мембрана впячивается внутрь клетки и образует мезосомы и (или) фотосинтетические мембраны.
Мезосомы - складчатые мембранные структуры (рис. 2.3 и 2.4), на поверхности которых находятся ферменты, участвующие в процессе дыхания. Следовательно, мезосомы можно назвать примитивными органеллами. Во время клеточного деления мезосомы связываются с ДНК, что, по-видимому, облегчает разделение двух дочерних молекул ДНК после репликации и способствует образованию перегородки между дочерними клетками.
У фотосинтезирующих бактерий в мешковидных, трубчатых или пластинчатых впячиваниях плазматической мембраны находятся фотосинтетические пигменты (в том числе бактериохлорофилл). Сходные мембранные образования участвуют и в фиксации азота.
Генетический материал
ДНК бактерий представлена одиночными кольцевыми молекулами длиной около 1 мм. Каждая такая молекула состоит примерно из 5·10 6 пар нуклеотидов. Суммарное содержание ДНК (геном) в бактериальной клетке намного меньше, чем в эукариотической, а следовательно, меньше и объем закодированной в ней информации. В среднем такая ДНК содержит несколько тысяч генов, что примерно в 500 раз меньше, чем в клетке человека (см. также табл. 2.1 и рис. 2.3).
Рибосомы
См. табл. 2.1 (биосинтез белка) и рис. 2.3.
Споры
Некоторые бактерии (в основном принадлежащие к роду Clostridium или Bacillus) образуют эндоспоры, т. е. споры, находящиеся внутри клетки. Эндоспоры - толстостенные долгоживущие образования, крайне устойчивые к нагреванию и коротковолновому излучению. Они по-разному располагаются внутри клетки, что служит очень важным признаком для идентификации и систематики таких бактерий (рис. 2.6). Если покоящаяся, устойчивая структура образуется из целой клетки, то она называется цистой. Цисты образуют некоторые виды Azotobacter.
Рис. 2.6. Различные формы бактерий на примере нескольких наиболее распространенных типов полезных и болезнетворных микробов.
А. Кокки (сферические)
Пример - Staphylococcus aureus, живущий в носоглотке; разные штаммы стафилококков вызывают фурункулез, воспаление легких, пищевые отравления и другие заболевания.
Пример - многие виды Streptococcus; некоторые вызывают инфекционные заболевания верхних дыхательных путей; например, S. pyogenes вызывает ангину и скарлатину; S. thermophilus придает йогурту его пикантный вкус; S. lactis - см. разд. 2.3.4
К этому роду относятся елинственный вид Diplococcus pneumoniae (пневмококк), возбудитель пневмонии * .
* (D. pneumoniae вызывает крупозное воспаление легких и многочисленные тяжелые острые пиогенные инфекции: менингит, септицемию, эмпиему и перитонит. - Прим. перев. )
Б. Бациллы (палочковидные)
![]()
Примеры - Escherichia coli (обычный кишечный симбионт); Lactobacillus см. разд. 2.3.4; Salmonella typhi - возбудитель брюшного тифа.
Примеры - Azotobacter, азотфиксирующая бактерия; Bacillus anthracis - возбудитель сибирской язвы.
Бациллы с эндоспорами (споры находятся в разном положении, имеют разные размеры и форму)
Находится в центре и не вызывает набухания клетки, например у Bacillus anthracis - возбудителя сибирской язвы.
Находится на конце материнской клетки, придает ей характерную форму барабанной палочки, например у Clostridium tetani - возбудителя столбняка.
Спора находится в субтерминальном положении, вызывая набухание клетки, например у Clostridium botulinum (споры могут занимать и центральное положение) - возбудителя смертельного пищевого отравления - ботулизма.
В. Спириллы (спиралевидные)
Пример - Spirillum; Форма клеток у спирохет очень схожа, но есть различия по спосубу передвижения, например Treponema pallidum - возбудитель сифилиса.
Г. Вибрионы (короткие палочки, всегда изогнутые в виде запятой)
Пример - Vibrio cholerae - возбудитель холеры; имеет один жгутик.