1. Bakterinės ląstelės anatomija. Bakterijų paviršiaus struktūros. Bakterijų kapsulė. Kapsulių organizavimas. Bakterinių kapsulių dažymas. Kapsulių sudėtis. Antigeninės kapsulių savybės.
2. Bakterijų vėliavėlė. Žvynelinės vieta. Peritrichsas. Monotrichai. Politrichai. Lophotrichous. Amfitrichai. Spiečius reiškinys. Bakterijų judrumo diagnostika.
3. Bakterijų mikrovileliai. bakterijų fimbrija. F-drink (sekso gėrimo) bakterijos. Bakterijų ląstelės sienelė. Glikokaliksas.
4. Bakterijų ląstelės sienelė. Ląstelės sienelės funkcijos. Bakterijų ląstelės sienelės struktūra. Peptidoglikanas. Pelės krepšys. Peptidoglikano (mureino) struktūra
5. Gramneigiamos bakterijos. Gramneigiamų bakterijų ląstelių sienelė. Gramneigiamų bakterijų ląstelės sienelės struktūra.
6. Gramteigiamos bakterijos. Gramteigiamų bakterijų ląstelių sienelė. Gramteigiamų bakterijų ląstelės sienelės struktūra. Bakteriniai autolizinai. Sferoplastai. Protoplastai.
7. Bakterijų citoplazminė membrana (CPM). Bakterijų citoplazminės membranos sudėtis. Transporto sistemos. Mezosomos. periplazminė erdvė.
9. Bakterijų fiziologija. Bakterijų mityba. Bakterijų maitinimo tipas. Holozojaus. Holofitai. Vanduo. Vandens svarba bakterijoms.
10. Bakterinės ląstelės virškinami junginiai. Medžiagų patekimo į bakterijų ląstelę keliai. Pasyvus perkėlimas. Difuzija.
Bakterijų citoplazma yra koloidinė matrica, kuri atlieka gyvybines funkcijas. Citoplazma daugumoje bakterijų yra DNR, ribosomų ir saugojimo granulių; likusią erdvės dalį užima koloidinė fazė. Pagrindiniai jo komponentai yra tirpūs fermentai ir tirpi RNR (mRNR ir tRNR). Įvairių eukariotinei ląstelei būdingų organelių bakterijose nėra, o jų funkcijas atlieka bakterinė CPM, atskirianti citoplazmą nuo ląstelės sienelės. Didžioji dauguma bakterijų citoplazma santykinai nejudrios, tačiau Streptococcus, Proteus, Clostridium rūšys turi specialius kanalėlius – rapidosomas, panašias į pirmuonių mikrovamzdelius.
bakterijų genomas
Bakterijų ląstelės neturi branduolinės membranos, joje koncentruojasi DNR citoplazma rutulio pavidalu. Kadangi eukariotinėse ląstelėse DNR paprastai randama šerdis, tada pagal analogiją su bakterijų DNR jie vadino nukleoidas[iš lat. branduolys, šerdis + graikų k. eidos, panašumas]. Ji taip pat vadinama genoforas, arba bakterinė chromosoma. Genoforas bakterijas vaizduoja dviguba spiralinė, apskrita, kovalentiškai uždaryta superspiralinė DNR molekulė. Jis sudaro 2-3% sausos ląstelės masės (daugiau nei 10% tūrio). Genoforas nėra histonų. Užkoduotos genetinės informacijos kiekis genoforas, skiriasi priklausomai nuo bakterijų tipo (pavyzdžiui, Escherichia coli genomas koduoja maždaug 4000 skirtingų polipeptidų). Bakterijos gali turėti papildomos DNR inkliuzų pavidalu. Šie intarpai arba plazmidės, turi daugybę skirtingų genų, koduojančių papildomas bakterijų savybes, tačiau plazmidėse esanti informacija bakterinei ląstelei nėra absoliučiai būtina.
bakterinė ribosoma
bakterinė ribosoma- sudėtingi rutuliniai dariniai, susidedantys iš įvairių RNR molekulių ir su jomis susijusių baltymų. Visas formavimas veikia kaip polipeptidų sintezės vieta. Priklausomai nuo augimo intensyvumo, bakterijų ląstelėje gali būti nuo 5000 iki 50000 ribosomų. Skersmuo bakterinės ribosomos apie 16-20 nm. Jų nusėdimo greitis ultracentrifugavimo metu yra 70 S (Swedbergo vienetai), o eukariotinėse ląstelėse – 80 S. Bakterijų ribosomos susideda iš dviejų subvienetų, kurių sedimentacijos koeficientas yra 50 S ir 30 S (eukariotuose 40 S ir 60 S). Subvienetų susiejimas įvyksta prieš pradedant vertimą. Prokariotų ir eukariotų ribosomos turi panašią molekulinę struktūrą ir veikimo mechanizmus, tačiau skiriasi ne tik dydžiu, bet ir baltymų sudėtimi bei baltymų faktoriais. Dėl šių skirtumų eukariotinės ribosomos tampa praktiškai atsparios antibiotikų, blokuojančių baltymų sintezę bakterijose, poveikiui.
Atsarginės bakterijų granulės
Atsarginės granulės yra laikinas metabolitų perteklius; granulių buvimas ir skaičius skiriasi priklausomai nuo bakterijų tipo ir jų metabolinio aktyvumo. Granulių pavidalu gali būti laikomi polisacharidai (krakmolas, glikogenas, granulozė), riebalai (trigliceridai, panašūs į aukštesniųjų gyvūnų riebalus, Candida genties mielėse), vaškas mikobakterijose ir nokardijose; p-hidroksisviesto rūgšties polimerai (pvz., Bacillus megaterium ląstelėse), polifosfatai (volutinas) Spirillum volutans ir Corynebacterium diphtheriae, siera (sulfidą oksiduojančiose bakterijose), kristalizuoti baltymai (pvz., vabzdžiams toksiškas Bacillus thuringiensis prototoksinas).
Prokariotinė ląstelė turi keletą pagrindinių savybių, susijusių su jos ultrastruktūrine ir chemine organizacija.
Pagrindinis prokariotinės ląstelės bruožas yra branduolio, kurį nuo citoplazmos riboja dviguba membrana, nebuvimas. Paveldima medžiaga yra koncentruota bakterijų chromosomoje, paprastai vaizduojama kaip apskrita dvigrandė DNR molekulė. Kai kuriose spirochetuose, rodokokuose ir agrobakterijose buvo aptiktos linijinio tipo chromosomos, o aktinomicetuose – „pseudožiedinės“ chromosomos. Kai kurių rūšių ląstelėje randamos dvi ar net trys neidentiškos chromosomos. Srieginė DNR molekulė atlieka branduolio funkciją ir yra centrinėje ląstelės zonoje, vadinamoje " nukleoidas Elektroniniame mikroskope ant bakterinės ląstelės dalies ši zona atrodo šviesesnė nei likusi citoplazmos dalis.
Bakterijos gali turėti papildomą DNR molekulę ekstrachromosominių elementų pavidalu arba integruotos į genoforą. tokie intarpai žymimi terminu " plazmidės"(atitinkamai epizodinis ir integruotas). Epizomų DNR taip pat būdinga apskritimo forma, tačiau epizodo dydis yra mažesnis nei bakterijų chromosomos. Plazmidės turi daugybę skirtingų genų ir dažnai lemia bakterijų virulentiškumą, tačiau plazmidėse esanti informacija nėra absoliučiai būtina, bakterinė ląstelė.
citoplazminė membrana, supantis kiekvieną ląstelę, lemia jos dydį ir užtikrina reikšmingų ląstelių turinio ir aplinkos skirtumų išsaugojimą. Bakterijos ląstelėje nėra membraninio tinklo – endoplazminis tinklas, citoplazminė membrana, esanti po ląstelės sienele ir iš išorės apgaubianti protoplastą, yra vienintelė membrana ląstelėje.
Tačiau membrana nėra tik mechaninė pertvara. Jis taip pat tarnauja kaip labai selektyvus filtras, kuris palaiko skirtingą jonų koncentraciją abiejose membranos pusėse ir leidžia maistinėms medžiagoms patekti į ląstelę ir pašalinti atliekas. Citoplazminė membrana yra lipidų ir baltymų molekulių visuma. palaikomos nekovalentinės sąveikos. Elektroniniame mikroskope membranos atrodo kaip apie 7 nm storio lakštai su ryškia trijų sluoksnių struktūra, dvigubas lipidų molekulių sluoksnis lemia membranos struktūrines ypatybes, o baltymai atsakingi už daugumą membranos funkcijų.
Prokariotų citoplazminėje membranoje yra daugybė invaginacijų – invaginacijų, kurios atlieka skirtingas funkcijas. Juose gali būti energijos apykaitos fermentų ir atitinkamai atlikti mitochondrijų funkcijas arba būti mezosomomis ir dalyvauti ląstelių dalijimuisi. Endosporų dangteliai susidaro iš citoplazminės membranos. Tai taip pat yra pagrindinė kliūtis medžiagoms prasiskverbti į ląstelę.
Už citoplazminės membranos ribų esančios struktūros – ląstelės sienelė, kapsulė, gleivinė, žvyneliai, fimbrijos, piliai, gaureliai – yra paviršinės struktūros.
ląstelių sienelės- svarbus ir privalomas daugumos prokariotinių ląstelių struktūrinis komponentas (tikrosios bakterijos, neskaitant archebakterijų ir mikoplazmų). Jis yra po kapsule ar gleivine arba (jei pastarosios nėra) liečiasi su aplinka. Ląstelės sienelė tarnauja kaip mechaninis barjeras tarp protoplasto ir išorinės aplinkos ir suteikia ląstelei tam tikrą jai būdingą formą – standumą. Ląstelės sienelės sudėtis ir struktūra yra svarbus sisteminis požymis, pagal kurį visi prokariotai skirstomi į šias grupes: gramteigiamus, gramneigiamus ir be ląstelės sienelės.
Šių grupių pavadinimai kilę iš skirtingų bakterijų gebėjimo arba nesugebėjimo nudažyti gramų. 1884 metais danų gydytojas K. Gramas, užsiėmęs audinių dažymu, pasiūlė dažymo būdą, kuris vėliau buvo panaudotas bakterijoms. Jo esmę išsamiai apsvarstysime laboratorinėje pamokoje, tačiau kol kas pastebime, kad gramteigiamų ir gramneigiamų bakterijų ląstelių sienelės labai skiriasi tiek chemine sudėtimi, tiek ultrastruktūra.
G+ bakterijos gerai išlaiko gencijonų violetinės spalvos spalvinį kompleksą su jodu ląstelės viduje, o G - – po apdorojimo alkoholiu praranda spalvą.
Nustatyta, kad empiriškai nustatyti skirtumai tarp dviejų bakterijų grupių, susijusių su Gramo dėmėmis, koreliuoja su daugeliu kitų svarbių bruožų. Pavyzdžiui, G+ bakterijos dauginasi dvejetainiu dalijimusi dėl to, kad ląstelėje susidaro skersinė pertvara, gramneigiamos – susidarius susiaurėjimui. Pirmieji jautrūs penicilinui, antrieji – ne. G-bakterijos nesudaro endosporų, bet kai kurios G+ sudaro.
Pagal struktūrą ir cheminę sudėtį prokariotų ląstelių sienelė labai skiriasi nuo eukariotinių organizmų. Prokariotų ląstelės sienelė susideda iš kelių sluoksnių, iš kurių pagrindinis – rėmo – sluoksnis, atsakingas už stiprumą, susidaro iš specialios medžiagos – peptidoglikano. mureina, kurią sintetina tik prokariotinė ląstelė ir nėra randama kituose organizmuose. Tai yra susietas biopolimeras, heteropolisacharidas, kuris sudaro uždarą maišelį, kuris visiškai uždengia ląstelės išorę. Mureinas sudaro didžiąją dalį G+ bakterijų ląstelės sienelės (40 kartų daugiau nei G-). Molekulės polisacharidinė sudėtis yra sudaryta iš kintančių likučių N-acetilgliukozaminas ir N-acetilmuramo rūgštis sujungtos viena su kita glikozidiniais ryšiais. Mureino maišelis yra gana elastingas ir veikiamas išorinių veiksnių gali išsitempti ir susitraukti iki 4 kartų. Įdomu tai, kad tokia biologinė struktūra kaip mureinas evoliucijos eigoje susidarė du kartus, nes kai kuriuose archeuose yra panašus biopolimeras, susidedantis iš kitų pradinių medžiagų.
Ląstelės sienelėje taip pat yra techo rūgščių ir polisacharidų. Teichoinės rūgštys yra polimerai, sudaryti iš polihidroksilių alkoholių (ribitolio ir glicerolio), tarpusavyje sujungtų fosfodiesterio jungtimis. Manoma, kad teicho rūgštys suteikia mureino maišeliui tam tikrą įtempimo ir suspaudimo laisvę ir veikia kaip spyruoklės čiužinyje.
G-eubakterijose ląstelės sienelės struktūra yra daug sudėtingesnė. Jame yra daug daugiau skirtingų cheminių tipų makromolekulių. Peptidoglikanas sudaro tik vidinį ląstelės sienelės sluoksnį, laisvai prilipdamas prie citoplazminės membranos. Už peptidoglikano yra papildomas ląstelės sienelės sluoksnis - išorinė membrana, kuris susideda iš fosfolipidų, baltymų, lipoproteinų ir lipopolisacharidų. Tarp citoplazminės ir išorinės membranos yra unikalus periplazminė erdvė. Išorinėje membranoje yra porino baltymų, kurie sudaro poras. Be to, jis užtikrina ląstelių kontaktą tarpusavyje, su substrato paviršiumi ir laiko daugybę išorinių struktūrinių darinių, tokių kaip pilis.
gerti- baltyminio pobūdžio siūlinės polimerinės organelės, lokalizuotos ląstelių paviršiuje. Žymi visų tipų nelipliuojančius darinius ląstelės paviršiuje (fimbrija). Pili veikia kaip bakteriofagų akceptoriai. Jie padeda ląstelėms priimti ir perduoti DNR konjugacijos metu ir dalyvauti ląstelės judėjime. Pagrindinis tikslas yra palaikyti specifines ląstelių tvirtinimo struktūras.
Prokariotų ląstelės sienelė mechaniškai apsaugo ląstelę nuo aplinkos poveikio, išlaiko išorinę formą ir leidžia ląstelei egzistuoti hipotoniniuose tirpaluose. Medžiagų pralaidumas ir transportavimas į ląstelę yra susijęs su ląstelės sienele.
Po citoplazmine membrana bakterijose yra citoplazma. Tai koloidinė sistema, susidedanti iš vandens, baltymų, riebalų, angliavandenių, mineralinių junginių ir kitų medžiagų, kurių santykis kinta priklausomai nuo bakterijų rūšies ir jų amžiaus. Citoplazmoje gali būti inkliuzų (mezosomų, dujų vakuolių) ir atsarginių medžiagų (β-hidroksibutirato, glikogeno ir sieros granulių, karboksizomų, parosporinių kūnų). Kai kurie inkliuzai tiesiog glūdi citoplazmoje, kiti yra apsupti plona membrana (2-4 nm).
Prokariotinės ląstelės citoplazma nėra pavaldi cikliniam judėjimui, kaip augaluose, ar susitraukimo reiškiniams, kaip amebose. Bakterijų ląstelės juda dėl kitų mechanizmų: arba paprasčiausiai išdėstytų žvynelių pagalba, kurios skiriasi nuo sudėtingų eukariotų žvynelių, jos aktyviai plaukia skystoje terpėje arba slysta, išskirdamos gleives ir judėdamos tankaus substrato paviršiumi. Šiek tiek pakeistas judėjimo tipas pastebimas spirochetuose, turinčiuose „ašinį siūlą“.
bakterinės žvyneliai esantys poliuose arba skirtingose ląstelės pusėse. Priklausomai nuo žvynelių skaičiaus ir išsidėstymo, išskiriami monotrichiniai (vienas poliarinis žiuželis), amfitrichas (žvyneliai dviejuose poliuose), lofotrichas (žvynelių ryšulėlis) ir peritrichas (žvyneliai per visą ląstelės paviršių). Flagelos statomos skirtingai. Nei eukariotų (9+2). Bakterijos turi dešinę besisukančias žvynelius, o archėjos – į kairę besisukančias žvynelius. Žulyklas susideda iš bazinio kūno, kurį sudaro 4 arba 2 žiedai, strypas, motoriniai baltymai, taip pat kabliukas ir filomentas (cilindras susideda iš flagellino baltymo).Judrios bakterijų ląstelės aktyviai juda tam tikra kryptimi. faktoriai. Tokie nukreipti bakterijų judėjimai vadinami „taksi“.
pailsėjusios bakterijų ląstelės G+ bakterijose atstovauja endosporos, G- – cistos. Bakterijų sporos turi unikalų atsparumą neigiamam poveikiui. Sporų stabilumas yra susijęs su specialiu sluoksniu jų ląstelės sienelėje - žievė.
Endosporos išlieka gyvybingos 500 metų, aktinomicetų sporos iki 7500 metų, bacilos – 25-30 milijonų metų (bitės žarnyne gintaro gabalėlyje).
Sporų susidarymo tipas (baciliarinis, klostridinis ir plektridinis) dažnai padeda atpažinti pasėlius.
Cista- ląstelė, praradusi mobilumą su sustorėjusia gleivine. Atsparus džiūvimui ir ekstremalioms temperatūroms. Cistoms būdinga ramybės būsena su žymiai sumažėjusiu metaboliniu aktyvumu. Funkcijos: apsaugo nuo žalingo aplinkos poveikio, tarnauja kaip būdas išsaugoti DNR, būdas perduoti informaciją iš šeimininko šeimininkui (patogenuose). Dygimą skatina palankios išorės sąlygos.
Išorėje prokariotų ląstelių sienelės dažnai yra apsuptos gleivinės medžiagos. Tokie dariniai, priklausomai nuo struktūrinių ypatybių, vadinami kapsulės, gleivinės sluoksniai, apvalkalai ar makštys(gijinėse bakterijose). Visi jie yra prokariotų organinių polimerų, daugiausia polisacharidinio pobūdžio, biosintezės ir jų nusėdimo aplink ląsteles rezultatas. Paprastai bakterijų suformuotų kapsulių cheminė sudėtis yra būdinga genčiai ir rūšiai. Kapsulės ir dangteliai apsaugo ląstelę nuo pažeidimo, išsausėjimo, sukuria papildomą barjerą, tarnauja kaip kliūtis fagų prasiskverbimui, sukuria aplink ląsteles specialią aplinką, užtikrinančią tam tikrą vandens režimą.
Prokariotinės ląstelės forma yra labai įvairi. Yra trys pagrindinės bakterijų formos – sferinės arba ovalios – kokos, lazdelės (bacilos) ir spiralinės.
Cocci skirstomi į poras - diplokokai, tetrakokai, pakelių cocci arba Sarcinas, esantis aukštuose, streptokokai- grandinėse stafilokokai, formuojant beformius kekes primenančius vynuogių kekes.
Tarp lazdelių išskiriami viengungis, atsitiktine tvarka diplobacilos, išdėstyti poromis ir streptobacilos- grandinėse
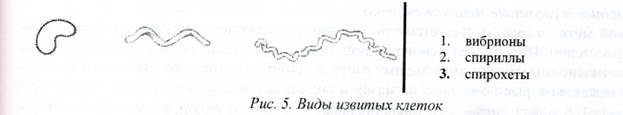
Spiralinės bakterijos skirstomos į dvi grupes - vibrijos Ir spirochetos Ir spirilla turintys posūkius, lygius vienam ar keliems spiralės posūkiams.
Daugialąsčiai prokariotai yra siūliniai arba kirmėlės formos.
Taigi, bakterijos yra morfologiškai gana silpnai diferencijuotos, todėl tarp jų galima išskirti tik ribotą skaičių formų. Toks išorinis „vienodumas“ stebėtinai kontrastuoja su ypatinga medžiagų apykaitos procesų įvairove.
Pagrindinė bakterijų klasifikacija
Klasifikuojant mikroorganizmus, priskiriamus prokariotams, visų pirma atsižvelgiama į praktinius aspektus ir jie padeda atpažinti aprašytas formas. Klasifikacija suprantama kaip vienetų paskirstymas į aukštesnės eilės grupes. Tai atliekama hierarchine sistema. Pagrindinis vienetas yra gryna izoliuotų bakterijų kultūra – „štamas“. Padermės grupuojamos į rūšis (rūšis), rūšys į gentis (genus, pl. gena), o gentys į šeimas. Klasifikavimo pagrindas yra tinkamas štamų aprašymas. pagal kuriuos jie lygina ir skiria nagrinėjamus vienetus. Užsiimama atskirų organizmų grupių pavaldumo nustatymu taksonomija.
Yra dviejų tipų klasifikacijos: filogenetinė „natūrali“ ir dirbtinė. Natūralios klasifikacijos sukūrimas yra galutinis bakterijų taksonomijos tikslas – sujungti giminingas formas, susijusias su bendra kilme, ir šiuo pagrindu sukurti filogenetinį bakterijų medį. Be jokios abejonės, kada nors tai bus galima padaryti remiantis cheminėmis savybėmis – tokiomis kaip aminorūgščių seka funkciškai panašiuose baltymuose arba nukleotidų seka konservuotose nukleorūgščiuose, pavyzdžiui, ribosominėje RNR.
Dirbtinė klasifikacija kelia sau kuklesnius tikslus. Ji yra skirta sugrupuoti organizmus į atskiras grupes pagal jų panašumą ir naudojama organizmams identifikuoti ir apibrėžti. Dirbtinė sistema skirta naudoti kaip raktas nustatant. Pats išsamiausias darbas, kuriame aprašytos bakterijos, yra Bergi bakterijų apibrėžimo vadovas. Pirmasis vadovo leidimas, kurį parengė Amerikos mikrobiologijos draugija, buvo išleistas 1928 m., paskutinis – devintas – 1994 m. Tai visame pasaulyje pripažintas kelių tomų veikalas, reguliariai perspausdinamas, parengtas tarptautinės komandos geriausi tam tikros mikroorganizmų grupės specialistai (apie 300 autorių). 9-asis amerikietiškas leidimas buvo išverstas į rusų kalbą 1997 m. pasiaukojančio Rusijos mikrobiologų darbo dėka, kurie noriai atidavė savo laiką ir žinias atitinkamiems skyriams išversti. Tai yra bakterijų identifikavimo nuoroda, būtina kiekvienoje laboratorijoje.
Bakterijų skirstymas į grupes pagal fenotipinius požymius (morfologiją, fiziologiją) patogus praktiniais tikslais greitam organizmų identifikavimui. Organizmai grupuojami į dirbtines grupes arba „skyrius“, kurie neturi taksonominio statuso ir dažnai vadinami jiems būdingiausiu genties ar pavadinimu. Burgee Determinant (9-asis leidimas, 1997) sukurtas šiuo principu.
Bakterijų „rūšies“ sąvoka, kuri yra kertinis kiekvienos klasifikacijos akmuo, vis dar neturi aiškaus apibrėžimo. Įvairūs bakterijų kintamumo mechanizmai sukelia tam tikrą požymių nestabilumą, kurių visuma lemia tam tikrą rūšį. Todėl bakterijų taksonomijoje sąvoka " variantas“. Yra morfologinių, biologinių, fermentinių, serologinių ir daug kitų variantų
Mikrobiologijoje taip pat vartojamos specializuotos sąvokos ir terminai - " padermė“ ir „klonas“. Terminas " įtampa" apibrėžia mikroorganizmų kultūrą, išskirtą iš tam tikro konkretaus šaltinio. Klonas yra mikroorganizmų kultūra, gauta iš vienos motininės ląstelės.
Pagrindiniai skirtumai tarp prokariotinės (bakterinės) ląstelės ir eukariotinės yra: formalizuoto branduolio (t. y. branduolinės membranos) nebuvimas, tarpląstelinių membranų, branduolių, Golgi komplekso, lizosomų ir mitochondrijų nebuvimas.
Pagrindinės bakterijų ląstelės struktūros yra:
Nukleoidas – tai paveldima (genetinė) bakterinės ląstelės medžiaga, kurią vaizduoja 1 DNR molekulė, uždaryta žiedu ir superspiralinė (susisukta į laisvą rutulį). DNR ilgis yra apie 1 mm. Informacijos kiekis – apie 1000 genų (ypatybių). Nukleoidas nuo citoplazmos nėra atskirtas membrana.
Citoplazma yra koloidas, t.y. vandeninis baltymų, angliavandenių tirpalas. Lipidai, mineralai, kuriuose yra ribosomų, inkliuzų, plazmidžių.
Baltymų sintezė vyksta ribosomose. Prokariotų ribosomos skiriasi nuo eukariotų mažesniais dydžiais (70 S).
Inkliuzai yra rezervinės bakterinės ląstelės maistinės medžiagos, taip pat pigmentų sankaupos. Rezervinės maistinės medžiagos yra: volutino (neorganinio polifosfato) granulės, glikogenas, granulozė, krakmolas, riebalų lašeliai, pigmento sankaupos, siera, kalcis. Inkliuzai, kaip taisyklė, susidaro, kai bakterijos auga ant turtingos maistinių medžiagų terpės ir išnyksta badaujant.
Ląstelių membrana – riboja citoplazmą. Susideda iš dvigubo fosfolipidų sluoksnio ir įterptųjų membraninių baltymų. CM, be barjerinių ir transportavimo funkcijų, atlieka metabolinio aktyvumo centro vaidmenį (priešingai nei eukariotinės ląstelės). Membraniniai baltymai, atsakingi už esminių medžiagų transportavimą į ląstelę, vadinami permeazėmis. Vidiniame CM paviršiuje yra fermentų ansambliai, t.y. sutvarkytos fermentų molekulių sankaupos, atsakingos už energijos nešėjų - ATP molekulių - sintezę. CM gali suformuoti invaginacijas į citoplazmą, kurios vadinamos mezosomomis. Yra dviejų tipų mezosomos:
Pertvara - formuoja skersines pertvaras ląstelių dalijimosi procese.
Šoninis - padeda padidinti CM paviršių ir padidinti medžiagų apykaitos procesų greitį.
Nukleoidas, CP ir CM sudaro protoplastą.
Viena iš išskirtinių bakterijų savybių yra labai aukštas viduląstelinis osmosinis slėgis (nuo 5 iki 20 atm), kuris yra intensyvios medžiagų apykaitos rezultatas. Todėl, siekiant apsisaugoti nuo osmosinio šoko, bakterijos ląstelė yra apsupta stipria ląstelės sienele.
Pagal ląstelės sienelės sandarą visos bakterijos skirstomos į 2 grupes: Turinčios vienasluoksnę ląstelės sienelę – gramteigiamos. Turinti dviejų sluoksnių ląstelės sienelę – gramneigiama. Vardai „Gram+“ ir „Gram-“ turi savo istoriją. 1884 metais danų mikrobiologas Hansas Christianas Gramas sukūrė originalų mikrobų dažymo metodą, dėl kurio vienos bakterijos nusidažė mėlynai (gram+), o kitos raudonai (gram-). Skirtingos bakterijų spalvos pagal Gramo metodą cheminis pagrindas buvo išaiškintas palyginti neseniai – maždaug prieš 35 metus. Paaiškėjo, kad G- ir G+ bakterijos turi skirtingą ląstelių sienelių struktūrą. G+ bakterijų ląstelės sienelės sandara. G+ bakterijų ląstelės sienelės pagrindą sudaro 2 polimerai: peptidoglikanas ir teikhoinės rūgštys. Peptidoglikanas yra linijinis polimeras su kintamomis muramo rūgšties ir acetilgliukozamino liekanomis. Tetrapeptidas (baltymas) yra kovalentiškai surištas su muramo rūgštimi. Peptidoglikano gijos yra tarpusavyje sujungtos peptidais ir sudaro tvirtą karkasą – ląstelės sienelės pagrindą. Tarp peptidoglikano gijų yra kitas polimeras – teikhoinės rūgštys (glicerolis TK ir ribitolis TK) – polifosfatų polimeras. Teichoinės rūgštys veikia ląstelės sienelės paviršių ir yra pagrindiniai G+ bakterijų antigenai. Be to, Mg ribonukleatas yra įtrauktas į G+ bakterijų ląstelės sienelę. G bakterijų sienelė susideda iš 2 sluoksnių: vidinį sluoksnį vaizduoja vienas arba dvisluoksnis peptidoglikanas (plonas sluoksnis). Išorinis sluoksnis susideda iš lipopolisacharidų, lipoproteinų, baltymų, fosfolipidų. Visų G bakterijų LPS turi toksinių ir slenkstinių savybių ir yra vadinami endotoksinais.
Veikiant tam tikroms medžiagoms, pavyzdžiui, penicilinui, sutrinka peptidoglikano sluoksnio sintezė. Tuo pačiu metu iš G+ bakterijų susidaro protoplastas, o iš G bakterijų – sferoplastas (nes išsaugomas išorinis ląstelės sienelės sluoksnis).
Tam tikromis auginimo sąlygomis ląstelės, neturinčios ląstelės sienelės, išlaiko gebėjimą augti ir dalytis, o tokios formos vadinamos L formomis (pagal Listerio instituto, kuriame šis reiškinys buvo atrastas, pavadinimo). Kai kuriais atvejais, pašalinus faktorių, kuris slopina ląstelės sienelės sintezę, L formos gali virsti pirminėmis formomis.
Daugelis bakterijų sintetina gleivinę medžiagą, susidedančią iš mukopolisacharidų, kurie nusėda ląstelės sienelės išorėje ir supa bakterinę ląstelę gleivine apvalkalu. Tai kapsulė. Kapsulės funkcija yra apsaugoti bakterijas nuo fazocitozės.
Bakterijų ląstelės paviršiaus struktūros.
Tvirtinimo prie pagrindo (sukibimo) organai yra piliai (fimbrijos) arba blakstienėlės. Jie prasideda nuo ląstelės membranos. Susideda iš pilino baltymo. Pilų skaičius gali siekti 400 ląstelėje.
Paveldimos informacijos perdavimo organai yra F-drank arba sex-drank. F tabletės susidaro tik tuo atveju, jei ląstelė yra nelyginė plazmidei, nes F-pili baltymai koduoja plazmidinę DNR. Jie yra plonas ilgas vamzdelis, kuris prisitvirtina prie kitos bakterinės ląstelės. Per susidariusį kanalą plazmidė patenka į kaimyninę bakterinę ląstelę.
Judėjimo organai – žvyneliai – yra spiraliniai siūlai. Jų ilgis gali viršyti skersmenį 10 ar daugiau kartų. Žvyneliai sudaryti iš baltymo flagellino. Žulyklės pagrindas yra sujungtas su ląstelės membrana per bazinį kūną. Bazinis kūnas susideda iš žiedų sistemos, kuri besisukdama perduoda sukimosi judesį žvyneliui. Pagal žvynelio išsidėstymą bakterijos skirstomos į mono-, lofo-, amfi-, peritricines.
Darbo pabaiga -
Ši tema priklauso:
Medicinos mikrobiologija, jos dalykas, metodai, ryšys su kitais mokslais
Liuminescencinė mikroskopija liuminescencija veikiant šviesos energijai e-spinduliai jonizatorius spinduliuojantis Sava bespalvė indukuota aplinka ... Mikroskopinio metodo metodas morfams ir tinctorial šviesai tirti bakterijose ant ... Bakterijos ląstelės cheminė sudėtis Vanduo ...
Jei jums reikia papildomos medžiagos šia tema arba neradote to, ko ieškojote, rekomenduojame pasinaudoti paieška mūsų darbų duomenų bazėje:
Ką darysime su gauta medžiaga:
Jei ši medžiaga jums pasirodė naudinga, galite ją išsaugoti savo puslapyje socialiniuose tinkluose:
| tviteryje |
Visos temos šiame skyriuje:
Medicinos mikrobiologija, jos dalykas, metodai, ryšys su kitais mokslais
Mikrobiologija – mokslas apie mikroorganizmus, t.y. apie gyvas būtybes, kurių matmenys mažesni nei 0,1 mm. Mikroorganizmai yra labai įvairūs. Tai apima kai kuriuos daugialąsčius organizmus, pirmuonius, ne
Pagrindiniai mikrobiologijos raidos etapai
I. Euristika – Hipokratas – ligų priežastis yra nematomos medžiagos – miazmos II. Morfologinis – Leeuwenhoekas – mikroskopu atrado gyvus organizmus. Ivanovskis - atrado virusus. III
Mikroorganizmai ir jų padėtis gyvojo pasaulio sistemoje
Mikroorganizmų klasifikacija. Atstovauja ikiląstelinės (karalystė Vira – virusai) ir ląstelinės formos (bakterijos, grybai, pirmuonys). Tarp ląstelių gyvybės formų išskiriamos 3 sritys: Bakterijos
Cheminė bakterinės ląstelės sudėtis
Bakterijų cheminė sudėtis Vanduo - 70%, Sausoji medžiaga - 30%. Baltymų – 52, polisacharidų – 16. Lipidų – 9,4, RNR – 16, DNR – 3,2, neorganinių junginių – 0,4. Bakterijų poreikis
Sporuliacija.
Esant nepalankioms sąlygoms, kai kurios bakterijos sugeba suformuoti endosporas – ramybės ląsteles. Viena ląstelė sudaro vieną endosporą, t.y. sporuliacija nėra dauginimosi forma, bet a
Fizinių veiksnių poveikis mikroorganizmams
Sterilizacija – tai visiškas bet kokios medžiagos ar objekto išlaisvinimas iš vegetatyvinių mikroorganizmų formų ir sporų, veikiant juos fiziniams ir cheminiams veiksniams. Sterilizuotas chirurgijoje
Cheminių veiksnių poveikis mikroorganizmams. Dezinfekcija
Dezinfekavimas (dezinfekavimas) – (pranc. „des“ reiškia kažko pašalinimą, sunaikinimą; lot. „inficere“ – užkrėsti, sugadinti, nuodyti) – vegetatyvinio fo pašalinimas ir sunaikinimas.
Bakterijų mityba
Pagal mitybos tipą bakterijos skirstomos į autotrofus ir heterotrofus. Autotrofai sugeba sugerti anglį iš CO2. Heterotrofai sugeria anglį tik iš organinių junginių.
Maistinės medžiagos
Laboratorinėmis sąlygomis bakterijos auginamos maistinėse terpėse. Temperatūros sąlygos turi didelę reikšmę bakterijų augimui ir dauginimuisi. Visi mikroorganizmai temperatūros atžvilgiu p
Kvėpavimo bakterijos
Kvėpavimas reiškia katabolizmo reakcijas. Dėl kvėpavimo sudėtingos molekulės suskaidomos į paprastas, kai išsiskiria energija, kuri kaupiama ATP molekulėse (efektyvumas apie 40%). A
Bakterijų fermentai, jų biologinis vaidmuo
Fermentai yra didelės molekulinės struktūros biologiniai katalizatoriai, kuriuos gamina gyva ląstelė. Jie turi baltyminį pobūdį, yra griežtai specifiniai ir atlieka svarbų vaidmenį mikroorganizmų apykaitoje.
Mikroorganizmų augimas ir dauginimasis
Bakterijų dauginimasis reiškia jų gebėjimą daugintis, padidinti individų skaičių tūrio vienete. Augimas reiškia bakterijų masės padidėjimą dėl ląstelių sintezės
Aktinomicetai, jų morfologija
Aktinomicetai (aeyg - ray, mum - fungus) - spinduliuojantys grybai, yra didelė mikroorganizmų grupė, įtraukta į Acipomycetes būrį. Dauguma aktinomicetų yra laisvi
Riketijos, jų morfologija ir biologinės savybės
Riketijos yra polimorfiniai mikroorganizmai, kurie gyvena ir dauginasi tik gyvūnų, žmonių ir nešiotojų audinių ląstelėse (citoplazmoje ir branduolyje). Kokoidinės formos atrodo labai mažos
Mikoplazmų morfologija ir ultrastruktūra
Mikoplazmos priklauso Mollicutes klasei, Mycoplasmatales būriui, Mycoplasmasmaceae šeimai. Tai mažos 100-150 nm dydžio, kartais 200-700 nm dydžio bakterijos, nesudarančios sporų, nejudrios, gramtrios
Chlamidijos, morfologija ir kitos biologinės savybės
Chlamydia genčiai, Chlamydiaceae šeimai, Chlamydiales būriui priklauso trachomos, konjunktyvito (blenorėjos su inkliuzais), kirkšnies limfogranulomatozės (Nicolas-Favre liga), orn patogenai.
Grybai, jų morfologija ir biologinės savybės
Grybų sistematika ir klasifikacija. Grybai priskiriami prie augalų heterotrofinių eukariotinių organizmų, kuriems trūksta chlorofilo. Grybų rūšis (Fungi s. Mycetes) turi per 100 000 rūšių, kartu
Į mieles panašūs Candida genties grybai
kandidozės sukėlėjai. Tai apima į mieles panašius grybus iš Candida genties. Tai vienaląsčiai organizmai (134 pav.), kurie dauginasi pumpuruodami; nesusidaro konidijos ir askosporos,
Deuteromicetai
Deuteromicetai yra netobuli grybai (Fungi imperfecti), labai didelė grupė (25 000 rūšių) grybų, kurie turi daugialąstę grybieną, bet neturi nei marsupialinės, nei bazidinės sporuliacijos ir
Viruso sąveika su ląstele
virusų dauginimasis. Viruso dauginimosi ciklas susideda iš nuoseklių įvykių: Specifinė viruso adsorbcija ląstelės paviršiuje, prasiskverbimas į ląstelę.
Ląstelių kultūros, jų rūšys
Virusų auginimui naudojamos ląstelių kultūros, viščiukų embrionai ir laboratoriniai gyvūnai. Plačiausiai naudojamos viensluoksnės tripsinizuotų ląstelių kultūros, taip pat persodintos
Mikroorganizmų kintamumas
Tyrėjai, stovėję prie bakteriologijos ištakų, susidūrę su bakterijų rūšių įvairove ir tariamai metamorfozių galimybe, vienų bakterijų abipuse transformacija į kitas, sukūrė pleomorfizmo doktriną.
Mutacijos
Azoto bazių sekos pokyčiai DNR. Tipai: vienos ar kelių bazių praradimas (ištrynimas) arba įterpimas (įterpimas) su skaitymo rėmelio poslinkiu. Zame
Genetinės rekombinacijos
Rekombinacija – paveldimas kintamumas dėl DNR sekcijų apsikeitimo tarp mikroorganizmų, kurių vienas yra donoras, kitas – recipientas. Rekombinacijų tipai: Transformacija, į
Mikrobų plitimas gamtoje
Objektas Taršos gamta Sanitarinės-orientacinės bakterijos Vanduo Išmatos
Normali žmogaus organizmo mikroflora
Makroorganizmas ir aplinka yra viena ekologinė sistema, kurioje mikroorganizmams tenka svarbus fiziologinis vaidmuo. Makroorganizmas ir jo mikroflora (autoflora) normaliomis sąlygomis
Bakterinės ląstelės struktūrai tirti kartu su šviesos mikroskopu naudojami elektroniniai mikroskopiniai ir mikrocheminiai tyrimai, siekiant nustatyti bakterinės ląstelės ultrastruktūrą.
Bakterijos ląstelė (5 pav.) susideda iš šių dalių: trisluoksnės membranos, citoplazmos su įvairiais intarpais ir branduolinės medžiagos (nukleoido). Papildomi struktūriniai dariniai yra kapsulės, sporos, žvyneliai, piliai.
Ryžiai. 5. Scheminis bakterijos ląstelės sandaros pavaizdavimas. 1 - apvalkalas; 2 - gleivinis sluoksnis; 3 - ląstelės sienelė; 4 - citoplazminė membrana; 5 - citoplazma; 6 - ribosoma; 7 - polisomas; 8 - inkliuzai; 9 - nukleoidas; 10 - žvyneliai; 11 - gėrimas
apvalkalas Ląstelė susideda iš išorinio gleivinės sluoksnio, ląstelės sienelės ir citoplazminės membranos.
Gleivinis kapsulinis sluoksnis yra už ląstelės ribų ir atlieka apsauginę funkciją.
Ląstelės sienelė yra vienas pagrindinių ląstelės struktūrinių elementų, išlaikančių jos formą ir atskiriančių ląstelę nuo aplinkos. Svarbi ląstelės sienelės savybė – selektyvus pralaidumas, užtikrinantis būtinų maistinių medžiagų (aminorūgščių, angliavandenių ir kt.) prasiskverbimą į ląstelę ir medžiagų apykaitos produktų pasišalinimą iš ląstelės. Ląstelės sienelė palaiko pastovų osmosinį slėgį ląstelės viduje. Sienos tvirtumo suteikia mureinas – polisacharidinio pobūdžio medžiaga. Kai kurios medžiagos ardo ląstelės sienelę, pavyzdžiui, lizocimas.
Bakterijos, kuriose visiškai nėra ląstelės sienelės, vadinamos protoplastais. Jie išlaiko gebėjimą kvėpuoti, dalytis, sintetinti fermentus; išorinių veiksnių įtakai: mechaniniams pažeidimams, osmosiniam slėgiui, aeracijai ir kt. Protoplastus galima išsaugoti tik hipertoniniuose tirpaluose.
Bakterijos su iš dalies sunaikintomis ląstelių sienelėmis vadinamos sferoplastais. Jei ląstelės sienelės sintezės procesą slopinate penicilinu, susidaro L formos, kurios visų tipų bakterijose yra sferinės didelės ir mažos ląstelės su vakuolėmis.
Citoplazminė membrana tvirtai prilimpa prie ląstelės sienelės iš vidaus. Jis yra labai plonas (8-10 nm) ir susideda iš baltymų ir fosfolipidų. Tai pusiau pralaidus ribinis sluoksnis, per kurį maitinama ląstelė. Membranoje yra permeazės fermentų, kurie atlieka aktyvų medžiagų transportavimą, ir kvėpavimo fermentų. Citoplazminė membrana sudaro mezosomas, kurios dalyvauja ląstelių dalijimuisi. Kai ląstelė dedama į hipertoninį tirpalą, membrana gali atsiskirti nuo ląstelės sienelės.
Citoplazma- bakterinės ląstelės vidus. Tai koloidinė sistema, susidedanti iš vandens, baltymų, angliavandenių, lipidų, įvairių mineralinių druskų. Citoplazmos cheminė sudėtis ir konsistencija kinta priklausomai nuo ląstelės amžiaus ir aplinkos sąlygų. Citoplazmoje yra branduolinė medžiaga, ribosomos ir įvairūs intarpai.
Nukleoidas, ląstelės branduolinė medžiaga, jos paveldimas aparatas. Prokariotų branduolinė medžiaga, skirtingai nei eukariotai, neturi savo membranos. Brandžios ląstelės nukleoidas yra dviguba DNR grandinė, susukta į žiedą. DNR molekulė koduoja ląstelės genetinę informaciją. Pagal genetinę terminologiją branduolinė medžiaga vadinama genoforu arba genomu.
Ribosomos yra ląstelės citoplazmoje ir atlieka baltymų sintezės funkciją. Ribosomoje yra 60% RNR ir 40% baltymų. Ribosomų skaičius ląstelėje siekia 10 000. Susijungusios kartu, ribosomos sudaro polisomas.
Inkliuzai – granulės, kuriose yra įvairių rezervinių maistinių medžiagų: krakmolo, glikogeno, riebalų, volutino. Jie yra citoplazmoje.
Bakterijų ląstelės gyvybės procese suformuoja apsauginius organelius – kapsules ir sporas.
Kapsulė- išorinis sutankintas gleivinis sluoksnis, esantis greta ląstelės sienelės. Tai apsauginis organas, atsirandantis kai kuriose bakterijose, kai jos patenka į žmonių ir gyvūnų organizmą. Kapsulė apsaugo mikroorganizmą nuo apsauginių organizmo faktorių (plaučių uždegimo ir juodligės sukėlėjų). Kai kurie mikroorganizmai turi nuolatinę kapsulę (Klebsiella).
ginčų randama tik lazdelės formos bakterijose. Jie susidaro mikroorganizmui patekus į nepalankias aplinkos sąlygas (aukštą temperatūrą, džiūvimą, pH pokyčius, aplinkoje sumažėjusį maistinių medžiagų kiekį ir kt.). Sporos yra bakterijos ląstelės viduje ir yra sutankinta citoplazmos sritis su nukleoidu, apsirengusiu savo tankiu apvalkalu. Pagal cheminę sudėtį jie skiriasi nuo vegetatyvinių ląstelių nedideliu kiekiu vandens, padidėjusiu lipidų ir kalcio druskų kiekiu, kuris prisideda prie didelio sporų atsparumo. Sporuliacija įvyksta per 18-20 valandų; mikroorganizmui patekus į palankias sąlygas, sporos per 4-5 valandas išdygsta į vegetatyvinę formą. Bakterijos ląstelėje susidaro tik viena spora, todėl sporos nėra dauginimosi organai, o tarnauja išgyventi nepalankiomis sąlygomis.
Sporas formuojančios aerobinės bakterijos vadinamos bacilomis, o anaerobinės – klostridijomis.
Sporos skiriasi forma, dydžiu ir vieta ląstelėje. Jie gali būti išdėstyti centre, po galais ir galinėje dalyje (6 pav.). Juodligės sukėlėjo sporos yra centre, jos dydis neviršija ląstelės skersmens. Botulizmo sukėlėjo sporos yra arčiau ląstelės galo – po galais ir viršija ląstelės plotį. Stabligės sukėlėjo ląstelės gale yra suapvalinta spora - galutinai ir žymiai viršija ląstelės plotį.
Flagella- judėjimo organai, būdingi lazdelės formos bakterijoms. Tai plonos siūlinės fibrilės, susidedančios iš baltymo – flagelino. Jų ilgis gerokai viršija bakterinės ląstelės ilgį. Flagela tęsiasi nuo bazinio kūno, esančio citoplazmoje, ir išeina į ląstelės paviršių. Jų buvimą galima nustatyti nustatant ląstelių mobilumą mikroskopu, pusiau skystoje maistinėje terpėje arba dažant specialiais metodais. Žievelių ultrastruktūra buvo tiriama naudojant elektroninį mikroskopą. Pagal žvynelių išsidėstymą bakterijos skirstomos į grupes (žr. 6 pav.): monotrichines – su vienu žvyneliais (choleros sukėlėjas); amfitrichas – su ryšuliais arba pavieniais žiuželiais abiejuose ląstelės galuose (spirilė); lophotrichous - su žiuželių ryšuliu viename ląstelės gale (išmatų šarminis formuotojas); peritrichoziniai – žvyneliai išsidėstę visame ląstelės paviršiuje (žarnyno bakterijos). Bakterijų judėjimo greitis priklauso nuo žvynelių skaičiaus ir vietos (aktyviausios yra monotrichinės), nuo bakterijų amžiaus ir aplinkos veiksnių įtakos.
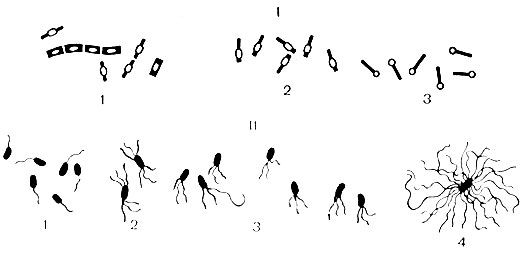
Ryžiai. 6. Sporų ir žvynelių išsidėstymo bakterijose variantai. I - ginčai: 1 - centrinis; 2 - subterminalas; 3 - terminalas; II - žvyneliai: 1 - vienspalviai; 2 - amfitrichai; 3 - lophotrichous; 4 - peritrichas
Pili arba fimbriae- gaureliai, esantys bakterijų ląstelių paviršiuje. Jie yra trumpesni ir plonesni nei žvyneliai, taip pat turi spiralinę struktūrą. Susideda geriant iš baltymų – pilino. Vieni piliai (jų yra keli šimtai) skirti bakterijoms pritvirtinti prie gyvūnų ir žmogaus ląstelių, o kiti (pavieniai) yra susiję su genetinės medžiagos perkėlimu iš ląstelės į ląstelę.
Mikoplazmos
Mikoplazmos yra ląstelės, kurios neturi ląstelės sienelės, bet yra apsuptos trijų sluoksnių lipoproteinų citoplazminės membranos. Mikoplazmos gali būti sferinės, ovalios, siūlų ir žvaigždžių formos. Mikoplazmos pagal Bergi klasifikaciją išskiriamos į atskirą grupę. Šiuo metu šie mikroorganizmai sulaukia vis didesnio dėmesio kaip uždegiminių ligų sukėlėjai. Jų dydžiai skirtingi: nuo kelių mikrometrų iki 125-150 nm. Mažos mikoplazmos praeina per bakterijų filtrus ir vadinamos filtruojamomis formomis.
Spirochetes
Spirochetai (žr. 52 pav.) (iš lot. speira – lenkimas, chaite – plaukai) – ploni, vingiuoti, judrūs vienaląsčiai organizmai, kurių ilgis nuo 5 iki 500 mikronų, o plotis – 0,3-0,75 mikronų. Paprasčiausiai jie yra susiję judėjimo būdu, sutrumpinant vidinį ašinį sriegį, susidedantį iš pluoštų pluošto. Spirochetų judėjimo pobūdis yra skirtingas: transliacinis, sukamasis, lenkiamasis, banguotas. Likusi ląstelių struktūra būdinga bakterijoms. Kai kurios spirochetos silpnai nusidažo anilino dažais. Spirochetai skirstomi į gentis pagal siūlų garbanų skaičių ir formą bei jo galą. Be saprofitinių formų, paplitusių gamtoje ir žmogaus organizme, tarp spirochetų yra ir patogenų – sifilio ir kitų ligų sukėlėjų.
Riketsija
Virusai
Tarp virusų išskiriama fagų grupė (iš lot. phagos – ryjantys), sukeliantys mikroorganizmų ląstelių lizę (sunaikinimą). Išlaikydami virusams būdingas savybes ir sudėtį, fagai skiriasi viriono struktūra (žr. 8 skyrių). Jie nesukelia žmonių ir gyvūnų ligų.
Kontroliniai klausimai
1. Papasakokite apie mikroorganizmų klasifikaciją.
2. Kokios yra pagrindinės prokariotų karalystės atstovų savybės.
3. Išvardykite ir apibūdinkite pagrindines bakterijų formas.
4. Įvardykite pagrindines ląstelės organeles ir jų paskirtį.
5. Trumpai apibūdinkite pagrindines bakterijų ir virusų grupes.
Vienas iš pagrindinių prokariotinės ląstelės bruožų yra vidinio atskyrimo, kurį užtikrina elementarios membranos, nebuvimas. Tiesą sakant, vienintelė prokariotų membranų sistema yra CPM, kuri yra ląstelės sienelės dalis, dažnai sudėtinga ir skersinėmis raukšlėmis prasiskverbia giliai į citoplazmą.
Bakterijų struktūra gerai ištirta naudojant ištisų ląstelių ir jų itin plonų pjūvių elektroninę mikroskopiją bei kitus metodus. Bakterinę ląstelę supa membrana, susidedanti iš ląstelės sienelės ir citoplazminės membranos. Po apvalkalu yra protoplazma, kurią sudaro citoplazma su ribosomomis ir nesusiformavęs branduolys, vadinamas nukleoidu. Gali būti papildomų struktūrų: kapsulė, mikrokapsulė, gleivės, žvyneliai, inkliuzai. Kai kurios bakterijos gali formuoti sporas, kad išgyventų nepalankiomis sąlygomis. Visi bakterinės ląstelės struktūriniai elementai gali būti sąlyginai suskirstyti į privalomus (nukleoidą, CPM, ląstelės sienelę, citoplazmą, 70S ribosomas) ir pasirenkamus, kurių kai kuriose rūšyse gali nebūti (kapsulė, plazmidės, inkliuzai, žvyneliai, pilis ir kt.)
Kapsulė. Daugelio bakterijų ląstelių membraną gaubia amorfinės, hidrofilinės medžiagos sluoksnis. Kapsulę sudaro polisacharidai (egzopolisacharidai), kartais polipeptidai; pavyzdžiui, B. anthracis kapsulė sudaryta iš D-glutamo rūgšties polimerų. Pagrindinį vaidmenį organizuojant kapsules atlieka CPM. Paskirstykite mikrokapsules (atskleidžiamos tik tada, kai
elektroninė mikroskopija mukopolisacharidinių fibrilių sluoksnio pavidalu) ir makrokapsulės (aptinkamos šviesos mikroskopu). Kai kuriose bakterijose ląstelės membranos polimerai, kurie išsiskiria į išorę, yra laisvai išsidėstę aplink ją, sudarydami gleivinį sluoksnį. Glebės tirpsta vandenyje. Gleiviniai egzopolisacharidai būdingi Pseudomonas aeruginosa gleivinės padermėms, kurios dažnai randamos cistine fibroze sergančių pacientų skrepliuose. Kapsulė ir gleivės apsaugo bakterijas nuo pažeidimų, išsausėjimo, fagocitus. veikiant baktericidiniams veiksniams, užtikrina sukibimą su įvairiais pagrindais, gali turėti maistinių medžiagų atsargų. Antigeno kapsulė (K-antigenas). Norint vizualizuoti kapsulę, naudojami neigiamo dažymo metodai (pagal Burri-Gins).
Ląstelių sienelės. Ląstelės sienelė yra unikali savo chemine sudėtimi prokariotinės ląstelės struktūra. Ji atlieka nemažai svarbių funkcijų: saugo bakterijas nuo išorinio poveikio, suteikia joms būdingą formą, palaiko vidinės aplinkos pastovumą, dalyvauja dalijime, ląstelės sienelės paviršiuje yra bakteriofagų, bakteriocinų ir įvairių cheminių medžiagų receptorių. , maistinės medžiagos pernešamos ir išsiskiria per ląstelės sienelę.. metabolitai. Tai tvirta, elastinga struktūra su sudėtinga struktūra. Bakterijos ląstelės sienelės atraminis karkasas yra peptidoglikanas (mureinas, mukopeptidas) – heteropolimeras, susidedantis iš pasikartojančių disacharidų grupių, sujungtų skersinėmis ir šoninėmis grandinėmis. Disacharidas, kuris yra peptidoglikano struktūrinis vienetas, susideda iš N-acetilgliukozamino ir N-acetilmuramo rūgšties, sujungtų β-glikozidinėmis jungtimis. Oligopeptidai yra prijungti prie N-acetilmuramo rūgšties molekulės, sudarydami šonines grandines. Peptidoglikano fragmentų surišimas susideda iš peptidinės jungties susidarymo tarp aminorūgščių tilto galinės liekanos (D-alanino) su priešpaskutine gretimo aminorūgščių tilto liekana (L-lizinu arba diaminopimelio rūgštimi). Šoniniai tilteliai sudaro keturias aminorūgštis, skersiniai (vertikaliai jungiantys peptidoglikano sluoksnius) – penkias aminorūgštis. Šoniniuose ir skersiniuose tiltuose yra unikalių aminorūgščių, randamų tik prokariotuose, todėl šios jungtys yra daugelio antibakterinių vaistų taikinys.
Pagal ląstelės sienelės struktūrą ir sudėtį mikroorganizmai skirstomi į gramteigiamus ir gramneigiamus, o tai yra svarbi dirbtinės mikrobų sistemos ypatybė. Pagrindinis gramteigiamų bakterijų storosios ląstelės sienelės komponentas yra daugiasluoksnis peptidoglikanas, kuris sudaro 40-90% ląstelės sienelės masės. Teichoinės rūgštys (iš graikų k. teichos- sienelė), kurios molekulės yra 8-50 glicerolio ir ribitolio likučių grandinės, sujungtos fosfatiniais tilteliais. Gramteigiamų bakterijų ląstelės sienelėje yra nedidelis kiekis polisacharidų, lipidų, baltymų.
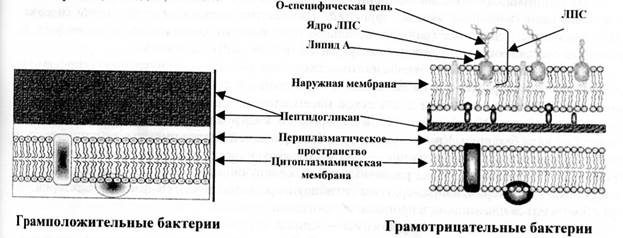
Ryžiai. 2. Bakterijų ląstelės sienelės sandara
Gramneigiamų bakterijų ląstelės sienelėje yra išorinė membrana, lipoproteinu sujungta su apatiniu peptidoglikano sluoksniu. Itin plonose bakterijų dalyse išorinė membrana turi banguotą trijų sluoksnių struktūrą, panašią į vidinę membraną, kuri vadinama citoplazmine. Pagrindinis šių membranų komponentas yra bimolekulinis (dvigubas) lipidų sluoksnis. Vidinį išorinės membranos sluoksnį sudaro fosfolipidai, o išoriniame sluoksnyje yra lipopolisacharido. Išorinės membranos lipopolisacharidas susideda iš 3 fragmentų: lipidas A – konservatyvi struktūra, beveik tokia pati gramneigiamose bakterijose; šerdis, arba strypas, žievės dalis (iš lat. šerdis -šerdis), santykinai konservatyvi oligosacharidinė struktūra (pastoviiausia LPS šerdies dalis yra ketodeoksioktono rūgštis); labai kintama O specifinė polisacharidų grandinė, susidaranti pasikartojančioms identiškoms oligosacharidų sekoms (O-antigenas). Išorinės membranos matricoje esantys baltymai prasiskverbia į ją taip, kad baltymų molekulės, vadinamos porinais, ribojasi su hidrofilinėmis poromis, pro kurias praeina vanduo ir mažos hidrofilinės molekulės.
Pažeidus bakterijų ląstelės sienelės sintezę, veikiant įvairiems veiksniams, susidaro modifikuotos formos (dažniausiai sferinės) ląstelės: protoplastai - bakterijos visiškai neturi ląstelės sienelės; sferoplastai - bakterijos su iš dalies išsaugota ląstelės sienele. Sfero arba protoplasto tipo bakterijos, praradusios gebėjimą sintetinti peptidoglikaną ir gebančios daugintis, vadinamos. L formos. Yra nestabilių L formų, kurios gali sugrįžti, „grįždamos“ į pradinę bakterinę ląstelę, ir stabilių L formų, kurios negali grįžti.
citoplazminė membrana. CPM yra privaloma ląstelinė struktūra, kuri yra fizinis, osmosinis, metabolinis barjeras tarp vidinio bakterinės ląstelės turinio ir išorinės aplinkos. CPM susideda iš dviejų sluoksnių fosfolipidų ir baltymų molekulių, įterptų į lipidų membraną (kaip ir daugumą biologinių membranų). Baltymai sudaro 20-75%, lipidai - 25-40%, o angliavandeniai ir RNR yra nedideliais kiekiais CPM CPM baltymai skirstomi į struktūrinius ir funkcinius. Pirmieji sudaro įvairias CPM struktūras, antruosius atstovauja fermentai, dalyvaujantys sintetinėse reakcijose membranos paviršiuje ir redokso procesuose, taip pat kai kurie specializuoti fermentai (permeazės dalyvauja medžiagų pernešime). Esant pernelyg dideliam augimui (palyginti su ląstelės sienelės augimu), citoplazminėje membranoje susidaro invaginacijos - invaginacijos sudėtingai susuktų membraninių struktūrų, vadinamų mezosomomis, pavidalu. Mezosomos sudaro skersines pertvaras tarp besidalijančių ląstelių ir yra bakterinės chromosomos prisitvirtinimo vieta.
Kai kuriose bakterijose tarp CPM ir ląstelės sienelės yra periplazminė erdvė – maždaug 10 nm pločio ertmė. Išorėje į šią erdvę atsiveria ląstelės sienelės poros, o iš vidaus išeina kai kurie ląstelių fermentai (ribonukleazės, fosfatazės, |3-laktamazės).
Citoplazma. Bakterijų citoplazma yra koloidinė matrica, kuri atlieka gyvybines funkcijas. Daugumos bakterijų citoplazmoje yra DNR, ribosomų ir saugojimo granulių; likusią erdvės dalį užima koloidinė fazė. Pagrindiniai jo komponentai yra tirpūs fermentai ir RNR (mRNR ir tRNR).
bakterijų genomas. Branduolio atitikmuo bakterijose yra nukleoidas (genoforas). Jis yra centrinėje bakterijų zonoje dvigubos superspiralinės žiedinės DNR molekulės pavidalu. Tai yra 2-3% sausos ląstelės masės (daugiau nei 10% tūrio). Genofor sudėtyje nėra histonų. Bakterijų chromosomoje užkoduotos genetinės informacijos kiekis priklauso nuo bakterijų tipo. Be nukleoido, bakterijos ląstelėje yra ekstrachromosominių genetinės informacijos nešėjų – plazmidžių, kurios yra kovalentiškai uždari DNR žiedai. Plazmidės turi daugybę skirtingų genų, koduojančių papildomus (neprivalomus) bakterijų požymius, tokius kaip atsparumo antibiotikams (K faktoriaus) genai. Konjugacijos proceso metu bakterijos gali keistis plazmidėmis.
Ribosomos. Bakterijų ribosomos yra sudėtingi rutuliniai dariniai, susidedantys iš RNR molekulių ir su jomis susijusių baltymų. Ribosomos yra būtinos polipeptidų sintezei. Ribosomų skaičius įvairiose bakterijų ląstelėse svyruoja nuo 5 iki 50 tūkst. Ribosomos skersmuo yra apie 16-20 nm. Jų nusėdimo greitis ultracentrifugavimo metu yra 70S (Swedbergo vienetai), o eukariotinėse ląstelėse – 80S. Bakterijų ribosomos susideda iš dviejų subvienetų, kurių sedimentacijos koeficientas yra 50S ir 30S (eukariotuose nuo 40S ir 60S). Subvienetų susiejimas įvyksta prieš pradedant vertimą. Ribosominė RNR (rRNR) yra konservatyvūs bakterijų elementai (evoliucijos „molekulinis laikrodis“). 16S rRNR yra mažo ribosomų subvieneto dalis, o 23S rRNR yra didelio ribosomų subvieneto dalis. 16S rRNR tyrimas yra genų sistematikos pagrindas, leidžiantis įvertinti organizmų giminingumo laipsnį. Dėl ribosomų struktūros skirtumų pro- ir eukariotinėse ląstelėse eukariotinės ribosomos tampa praktiškai atsparios antibiotikų, blokuojančių baltymų sintezę bakterijose, poveikiui.
Atsarginės granulės. Bakterijų citoplazmoje yra įvairių intarpų, kuriuose yra laikinas metabolitų perteklius. Granulių, polisacharidų (krakmolo, glikogeno), riebalų (trigliceridų, laikomų į mieles panašiuose ^1 genties grybuose) pavidalu SapsHc1a),β-hidroksisviesto rūgšties polimerai, polifosfatai (volutinas) C. difterija siera, kristalizuoti baltymai ir kt.
Flagella. Bakterinės žvyneliai yra bakterijų judėjimo (judėjimo) organai. Žvynelinės vieta yra būdingas požymis, turintis taksonominę reikšmę. Pagal žvynelių skaičių ir išsidėstymą išskiriami vienarūšiai – vienas žiuželis (V. cholerae), peritrichi (iš graikų k. peri- aplink ir trichosas- plaukai) – žvyneliai visame bakterijos ląstelės paviršiuje (E.coli), lophotrichs (iš graikų k. 1ophos - kuokštas ir trichosas - plaukai) – žvynelių pluoštelis viename ląstelės gale (Pseudomonas), amfitrichai (iš graikų k. atrhi - dvipusis, dvišalis ir trichosas- plaukai) – pavienės žvyneliai arba žvynelių ryšuliai skirtinguose ląstelės poliuose (Spirillum).Žyželis yra spirališkai išlenktas tuščiaviduris siūlas, sudarytas iš flagellino baltymų subvienetų. Žvyneliai yra 12–20 nm storio ir 3–15 µm ilgio (daugiau nei ląstelės ilgis). Žvyneliai susideda iš 3 dalių: spiralinio siūlelio, kabliuko ir bazinio korpuso. Baziniame korpuse yra lazdelė su specialiais diskais: viena pora diskų gramteigiamoms bakterijoms ir 2 poros diskų gramneigiamoms bakterijoms. Žvynelių diskai yra pritvirtinti prie citoplazminės membranos ir ląstelės sienelės. Taip sukuriamas elektrinio variklio su strypu efektas – varikliu, kuris suka žvynelį. Bakterinės žiogelės atlieka transliacinius ir sukamuosius judesius, stumdamos bakterijas per terpę kaip laivo sraigtą. Flagella yra H-antigenai, naudojami serologiniam identifikavimui.
Bakterijų mobilumas nustatomas mikroskopuojant „susmulkintus“ arba „pakabintus“ preparatus.
kieno" lašai. Gebėjimas judėti taip pat gali būti nustatomas įvedus bakterijų kultūrą įšvirkščiant į pusiau skysto agaro kolonėlę (judrios rūšys auga per visą terpės storį, nejudrios – įpurškiant) arba pasėjus bakterijas į vandeninį. nuožulnios agaro kolonėlės kondensatas.
(judrios rūšys nuplaukia iš kondensato į terpės paviršių ir ją kolonizuoja).
Microvilli. Be žvynelių, daugelio bakterijų paviršius padengtas citoplazminėmis ataugomis – mikrovilliais, randamais judriose ir nejudriose rūšyse. Pili (fimbriae, villi) – gijiniai dariniai, plonesni ir trumpesni (3-10 nm x 0,3-10 mikronų) už žiuželius. Pili tęsiasi nuo ląstelės paviršiaus ir susideda iš pilino baltymo, kuris turi antigeninį aktyvumą. Yra pilių, atsakingų už sukibimą, bakterijų prijungimą prie paveiktos ląstelės; pili atsakingi už mitybą, vandens-druskų apykaitą ir seksą (F-gėrė), arba konjugaciją, gėrė. Gėrimų gausu – po kelis šimtus narve. vienas-
ląstelėje dažniausiai būna 1-3 lyties piliai: juos formuoja vadinamieji „vyriški“
donorų ląstelės, turinčios pernešamų plazmidžių (F-, R-, Co1- plazmidės). Išskirtinis lyties pilių bruožas yra sąveika su specialiais „vyriškais“ sferiniais bakteriofagais, kurie intensyviai adsorbuojami ant lytinių pilių. Ginčai. Sporos susidaro nepalankiomis bakterijoms egzistuoti sąlygomis (džiūvimas, maistinių medžiagų trūkumas ir kt.). Bakterijos ląstelės viduje susidaro viena spora (endospora). Sporų susidarymas prisideda prie rūšies išsaugojimo ir nėra dauginimosi būdas, kaip grybuose. Tik genčių bakterijos turi galimybę sporuliuoti. bacila Ir Clostridium,įskaitant žmonėms patogeniškas rūšis ir daug saprofitų. Genties bakterijose Vasilijus sporos skersmuo neviršija bakterinės ląstelės skersmens (skersmens). Genties bakterijose Clostridium(iš lot. closter, verpstė) sporos skersmuo viršija bakterijos skersmenį, todėl joms suteikiama verpstės forma (C.perfringens), būgnų lazdelė (C. tetani) arba teniso raketė (C. botulinum) atitinkamai su centrine, galine ir subterminalia sporų lokalizacija. Pagrindinę sporų reikšmę bakterijų išlikimui lemia jų šiluminė varža ir atsparumas kitiems faktoriams (dezinfekcinėms medžiagoms ir kt.), turintiems neigiamą poveikį bakterijoms. Didelis sporų atsparumas yra susijęs su mažu laisvo vandens kiekiu, didele kalcio koncentracija, letarginiais medžiagų apykaitos procesais, dipikolino rūgšties ir baltymo, kuriame gausu cisteino (dėl kurio jis panašus į keratiną), buvimu, taip pat keletu lukštai, kurie yra papildoma apsauga nuo neigiamo išorės poveikio. Kai sporos patenka į palankias sąlygas, jos sudygsta, pereina tris iš eilės stadijas: aktyvacijos, iniciacijos, augimo.
Sporos blogai dažosi ir išlieka bespalvės dažytose ląstelėse naudojant įprastinius dažymo metodus (paprasti metodai, Gramo metodas). Sporos dažomos Orzeszko (Auesky) metodu.
5. Pagrindinės bakterijų formos
Tam tikros bakterijų rūšys, turinčios pakankamai pastovumo, yra būdingos: tam tikrai formai, dydžiui ir vietai. Bakterijų ląstelių ilgis svyruoja nuo 0,1-0,2 μm (Mucoplasma rūšys) iki 10-15 μm (Mucoplasma rūšys). Clostridium) storis - nuo 0,1 iki 2,5 mikronų. Vidutinis bakterijų dydis yra 2-3x0,3-0,8 µm. Yra trys pagrindinės bakterijų formos – sferinės (kokai), lazdelės formos (cilindrinės) ir vingiuotos (spiralinės).
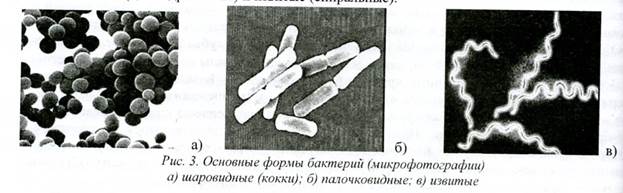
Ryžiai. 3. Pagrindinės bakterijų formos (mikrografai) a) sferinės (kokai); 6) strypo formos: c) vingiuotas
Dauguma cocci (iš graikų k. kokkos- uogos, grūdai) yra sferinės arba ovalios formos, kai kurių rūšių ląstelės gali būti lancetiškos (pneumokokai) arba pupelės formos (neisseria). Pagal ląstelių išsidėstymo tepinėliuose pobūdį diplokokai (išdėlioti poromis), streptokokai (susidūrę grandinėmis), stafilokokai (išdėstyti vynuogių kekių pavidalu), sarkinai (išdėlioti pakuotėse po 8. 16. 32 ir daugiau ląstelės) ir kt.
Strypo formos bakterijos skiriasi ląstelės galų forma ir santykine padėtimi. Lazdelės gali būti taisyklingos (E. coli), netaisyklingos (Corynebacteria), šakotos (Actinomycetes). Lazdelės formos bakterijos gali išsidėstyti taurėse pavieniui ir atsitiktinai (monobakterijos), poromis (diplobakterijos) arba grandinėje (streptobakterijos, streptobacilos) (4 pav.).

Ryžiai. 4. Pagrindinės bakterijų formos
Išlenktos formos – vaizduojamos lenktomis lazdelėmis. Atsižvelgiant į garbanų formą ir skaičių, išskiriami trys ląstelių tipai: vibrijos(iš graik. vibrio- vingiuoti, lenkti) turėti vieną garbanę. neviršijantys ketvirtadalio spiralės apsisukimo (išlenktos ląstelės kaip kablelis): spirilla(iš graikų kalbos. speira- spirale) turi 3-5 dideles garbanas ir spirochetos - daug smulkių garbanų (5 pav.).