Ląstelės branduolys yra esminė kiekvieno eukariotinio organizmo struktūra. atlieka labai įvairias funkcijas, tačiau pagrindinė jos paskirtis – paveldimos genetinės medžiagos saugojimas ir perdavimas.
Beveik kiekviena žmogaus kūno ląstelė turi branduolį. Vienintelės išimtys yra trombocitai ir raudonieji kraujo kūneliai. Dauguma ląstelių yra vienabranduolinės, tačiau, pavyzdžiui, raumenų skaidulos ir neuronai gali turėti keletą šių organelių. Ląstelėje esantis branduolys gali būti įvairaus dydžio – didžiausios branduolinės struktūros patelės kiaušinėlyje.
Ląstelės branduolys: sandara
Branduolys yra gana sudėtingos struktūros ir susideda iš branduolinės membranos, chromatino, branduolio ir nukleoplazmos. Pažvelkime į kiekvieną jo dalį išsamiau.
- Karioteka arba branduolio apvalkalas yra struktūra, skirianti vidinę branduolio aplinką nuo citoplazmos. Šis apvalkalas susideda iš išorinės ir vidinės membranų, tarp kurių yra vadinamoji perinuklearinė erdvė. Įdomu tai, kad išorinė apvalkalo membrana patenka tiesiai į granuliuoto endoplazminio tinklo membraną, todėl EPS rezervuarų ir branduolio ertmės yra tarpusavyje susijusios. O apvalkalas turi branduolines poras, uždarytas diafragma. Jie skirti prasiskverbti į dideles molekules, taip pat keistis medžiagomis tarp karioplazmos ir citoplazmos.
- Karioplazma yra vienalytė medžiaga, užpildanti vidinę branduolio ertmę. Jame yra branduolys ir chromatinas.
- Chromatinas yra genetinė ląstelės medžiaga. Jo struktūrinis vienetas yra nukleosoma, kuri yra DNR grandinė, apvyniota aplink konkretų baltymą, histoną. Ląstelėje yra dvi genetinės medžiagos būsenos. Heterochromatinas yra mažos, tankios osmofilinės granulės. Euchromatinas arba atsipalaidavęs chromatinas yra sritys, kuriose aktyviai vyksta sintetiniai procesai. Laikui bėgant chromatinas kondensuojasi ir sudaro chromosomas.
- Branduolys yra maža, ovali struktūra, sudaryta iš RNR ir baltymų molekulių gijų. Būtent čia susidaro ribosomų subvienetai. Branduolys gali būti vienas ar daugiau branduolių, tačiau juos galima pamatyti tik nesidalijančiose ląstelėse.
Ląstelės branduolys: funkcijos
Funkcijas galima nustatyti susipažinus su jo struktūra. Pirma, branduolys yra atsakingas už paveldimos informacijos perdavimą ląstelių dalijimosi metu, tiek mitozės, tiek mejozės metu. Mitozės metu dukterinės ląstelės gauna genomą, identišką motininei ląstelei. Mejozės (žmogaus formavimosi) metu kiekviena ląstelė gauna tik pusę chromosomų rinkinio – visas chromosomų rinkinys susidaro tik susiliejus su kito organizmo lytinėmis ląstelėmis.
Be to, ląstelės branduolys yra atsakingas už vieną svarbiausių medžiagų apykaitos etapų – baltymų sintezę. Faktas yra tas, kad būtent branduolyje susidaro informacija arba matricinė RNR. Tada jis patenka į endoplazminį tinklelį, prisitvirtina prie ribosomos ir yra peptido molekulės aminorūgščių sekos formavimo modelis.
Ir kaip jau minėta, ribosomų subvienetų sintezė vykdoma branduolyje.
Ląstelės branduolys: kilmė
Iki šiol yra kelios visiškai skirtingos hipotezės, kurių pagalba mokslininkai bando paaiškinti, kaip tiksliai ląstelėje susiformavo branduolys. Tačiau, deja, nė vienas iš šių teiginių dar nerado tikro patvirtinimo.
Yra teorija, kad branduolys, kaip ląstelinė struktūra, susidarė dėl simbiozės ir archėjos. Kiti mokslininkai mano, kad branduolys yra ląstelės užsikrėtimo konkrečiu virusu rezultatas.
Išsamiausiame paaiškinime yra vadinamoji egzomembranos hipotezė. Anot jos, evoliucijos procese ląstelė turėjo dar vieną išorinę ląstelės membraną. Tuo pat metu senoji, vidinė membrana virto branduolio apvalkalu – laikui bėgant joje susidarė sudėtinga porų sistema, o tada jos ertmėje pradėjo telktis chromatino molekulės.
Visi žinome, kad žmonės yra eukariotai. Tai reiškia, kad visos jo ląstelės turi organelę, kurioje yra visa genetinė informacija – branduolį. Tačiau yra išimčių. Ar žmogaus organizme yra ląstelių be branduolių ir kokia jų reikšmė gyvybei?
Branduolinės žmogaus ląstelės
Jų negalima lyginti su prokariotais, kurie turi tipišką struktūrą. Kokio branduolio neturinčio nėra - eritrocituose. Vietoj šios organelės juose yra sudėtingas cheminis medžiagų kompleksas, leidžiantis atlikti svarbiausias organizmo funkcijas. Trombocitai – trombocitai ir limfocitai – taip pat yra nebranduolinės ląstelės. Ląstelėse, kurios vadinamos kamieninėmis ląstelėmis, nėra branduolio. Visas šias struktūras vienija dar viena savybė. Kadangi jiems trūksta branduolio, jie negali daugintis. Tai reiškia, kad nebranduolinės ląstelės, kurių pavyzdžiai buvo pateikti, atlikusios savo funkciją miršta, o specializuotuose organuose formuojasi naujos.
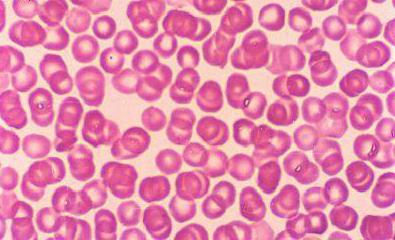
raudonieji kraujo kūneliai
Jie nustato mūsų kraujo spalvą. Branduolinės kraujo ląstelės, eritrocitai, turi neįprastą formą – abipus įgaubtą diską, kuris, esant santykinai mažam dydžiui, žymiai padidina jų paviršių. Tačiau jų skaičius tiesiog nuostabus: 1 kvadrate. mm jų kraujo yra iki 5 milijonų! Vidutiniškai eritrocitas gyvena iki keturių mėnesių, po to miršta ir neutralizuojamas blužnyje bei kepenyse. Kas sekundę raudonuosiuose kaulų čiulpuose susidaro naujos ląstelės.
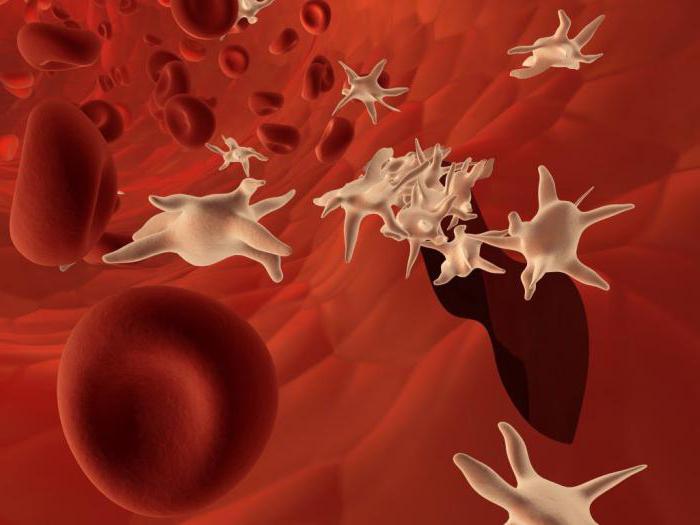
Raudonųjų kraujo kūnelių funkcijos
Kas šiose nebranduolinėse ląstelėse yra vietoj branduolio? Šios medžiagos vadinamos hemu ir globinu. Pirmasis yra geležies turintis. Jis ne tik nudažo kraują raudonai, bet ir sudaro nestabilius junginius su deguonimi ir anglies dioksidu. Globinas yra baltyminė medžiaga. Hemas, kuriame yra įkrautas geležies jonas, yra panardintas į didelę jo molekulę. Pagal veikimo mechanizmą šias ląsteles galima palyginti su fiksuoto maršruto taksi. Plaučiuose jie prideda deguonies. Su kraujotaka jis nunešamas į visas ląsteles ir ten išleidžiamas. Dalyvaujant deguoniui, vyksta organinių medžiagų oksidacijos procesas, kai išsiskiria tam tikras energijos kiekis, kurį žmogus naudoja gyvybei vykdyti. Atsilaisvinusią erdvę iš karto užima anglies dioksidas, kuris juda priešinga kryptimi – į plaučius, kur iškvepiamas. Šis procesas yra būtina gyvenimo sąlyga. Jei ląstelės nepateikiamos deguonimi, jos palaipsniui miršta. Tai gali kelti pavojų viso organizmo gyvybei.
Eritrocitai atlieka dar vieną svarbią funkciją. Jų membranose yra baltymų žymeklis, vadinamas Rh faktoriumi. Šis rodiklis, kaip ir kraujo grupė, labai svarbus kraujo perpylimo, nėštumo, donorystės ir chirurginių operacijų metu. Jis turi būti įdiegtas, nes nesuderinamumo atveju gali kilti vadinamasis Rh konfliktas. Tai apsauginė reakcija, tačiau gali sukelti vaisiaus ar organų atmetimą.
Netinkama mityba, blogi įpročiai, užterštas oras gali sukelti raudonųjų kraujo kūnelių sunaikinimą. Dėl to susergama sunkia liga, kuri vadinama anemija, arba anemija. Tokiu atveju žmogus jaučia galvos svaigimą, silpnumą, dusulį, spengimą ausyse. Deguonies trūkumas neigiamai veikia fizinę ir protinę žmogaus veiklą. Tai ypač pavojinga nėštumo metu. Jei per virkštelę vaisiui tiekiamas nepakankamas deguonies kiekis, tai gali sukelti rimtų jo vystymosi sutrikimų.
Trombocitų struktūra
Nebranduolinės ląstelės, trombocitai, dar vadinamos trombocitais. Neaktyvios būsenos jie tikrai turi plokščią formą, primenančią objektyvą. Bet kai kraujagyslės yra pažeistos, jos išsipučia, apvalėja, susidaro nestabilios išorinio sluoksnio ataugos - pseudopodijos. Trombocitai susidaro ir negyvena ilgai – iki 10 dienų, neutralizuojami blužnyje.
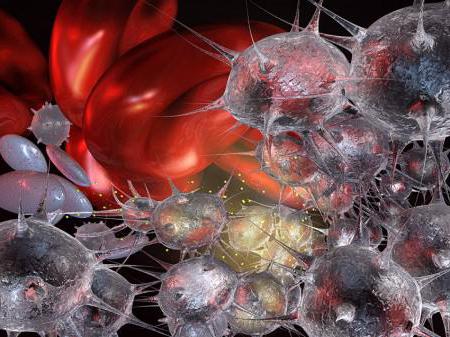
Trombų susidarymo procesas
Trombocitų matricoje yra fermento, vadinamo tromboplastinu. Jei pažeidžiamas kraujagyslių vientisumas, jis pasirodo plazmoje. Veikdamas kraujo baltymas protrombinas pereina į aktyvią formą, savo ruožtu veikdamas fibrinogeną. Dėl to ši medžiaga pereina į netirpią būseną. Jis virsta baltymu fibrinu. Jos siūlai yra glaudžiai susipynę ir sudaro trombą. Apsauginė kraujo krešėjimo reakcija apsaugo nuo kraujo netekimo. Tačiau kraujo krešulio susidarymas kraujagyslės viduje yra labai pavojingas. Tai gali sukelti jo plyšimą ir net kūno mirtį. Krešėjimo proceso pažeidimas vadinamas hemofilija. Šiai paveldimai ligai būdingas nepakankamas trombocitų skaičius ir dėl to netenkama daug kraujo.
kamieninės ląstelės
Šios ląstelės be branduolio ne veltui vadinamos kamieninėmis ląstelėmis. Jie iš tikrųjų yra visų kitų pagrindas. Jie taip pat vadinami „genetiškai grynais“. Kamieninių ląstelių yra visuose audiniuose ir organuose, tačiau daugiausia jų yra kaulų čiulpuose. Prireikus jie prisideda prie vientisumo atkūrimo. Kamieninės virsta bet kokiais kitais, kai yra sunaikintos. Atrodytų, kad esant tokiam magiškam mechanizmui, žmogus turėtų gyventi amžinai. Kodėl taip neatsitinka? Reikalas tas, kad su amžiumi kamieninių ląstelių diferenciacijos intensyvumas žymiai mažėja. Jie nebesugeba atkurti sunaikintų audinių. Tačiau yra ir kitas pavojus. Didelė tikimybė, kad kamieninės ląstelės virs vėžinėmis ląstelėmis, kurios neišvengiamai sukels bet kurio gyvo organizmo mirtį.
Celės be branduolių: pavyzdžiai ir skirtumai
Gamtoje ląstelės be branduolių yra gana dažnos. Pavyzdžiui, melsvadumbliai ir bakterijos yra prokariotai. Tačiau, skirtingai nei žmogaus ląstelės be branduolio, jos nemiršta atlikusios savo biologinį vaidmenį. Faktas yra tas, kad prokariotai turi genetinę medžiagą. Todėl jie gali dalytis, o tai įvyksta darant dvi genetines motininės ląstelės kopijas. Paveldima prokariotų informacija yra apvali DNR molekulė, kuri prieš dalijimąsi padvigubėja. Šis branduolio analogas dar vadinamas nukleoidu. Augaluose gyvos ląstelės yra be branduolių – sieto vamzdeliai.
Taigi, branduolio neturinčios žmogaus ląstelės nepajėgios dalytis, todėl jos egzistuoja trumpą laiką prieš atlikdamos savo funkciją. Po to vyksta jų sunaikinimas ir tarpląstelinis virškinimas. Tai yra suformuoti elementai (eritrocitai), trombocitai (trombocitai) ir kamieninės ląstelės.
Brownas nepadarė jokių prielaidų apie branduolio funkcijas. 1838 m. Matthiasas Schleidenas pasiūlė, kad branduolys dalyvauja formuojant naujas ląsteles, todėl įvedė terminą „citoblastas“ (ląstelių statybininkas), nurodant branduolius. Jis buvo įsitikinęs, kad stebi naujų ląstelių kaupimąsi aplink „citoblastus“. Tvirtas šio požiūrio priešininkas buvo Franzas Meyenas, kuris atrado, kad ląstelės dauginasi dalijantis, ir manė, kad daugelis ląstelių gali neturėti branduolio. Ląstelių formavimosi idėja de novo ty nuo nulio, per citoblastus ar kitaip, prieštaravo Roberto Remacko (1852) ir Rudolfo Virchow (1855) darbui, kurie pagaliau patvirtino naują paradigmą, kad ląstelės gali būti suformuotos tik iš ląstelių ("Omnis cellula e cellula"). . Šerdies funkcijos liko neaiškios.
struktūros
Branduolys yra didžiausia gyvūnų ląstelių organelė. Žinduolių branduolio skersmuo yra apytikslis 6 µm, o pats branduolys sudaro apie 10% ląstelės tūrio. Klampus skystis, užpildantis branduolį, vadinamas nukleoplazma ir yra chemiškai panašus į citozolį, kuris supa branduolį.
Branduolinis apvalkalas ir branduolio poros
Branduolinį apvalkalą sudaro dvi membranos (išorinė ir vidinė), kurios yra lygiagrečiai 10–10 atstumu. 50 nm. Branduolinis apvalkalas visiškai supa branduolį, atskirdamas ląstelės genetinę medžiagą nuo citoplazmos ir tarnaudamas kaip barjeras, neleidžiantis laisvai makromolekulėms sklisti tarp nukleoplazmos ir citoplazmos. Išorinė branduolio membrana tęsiasi į grubią endoplazminio tinklo (ER) membraną ir yra išklota ribosomomis. Erdvė tarp branduolinių membranų vadinama perinuklearine erdve ir tęsiasi į ER spindį.
Branduolinės poros, kurios yra vandens užpildyti kanalai branduolio apvalkale, yra sudarytos iš įvairių baltymų, vadinamų nukleoporinais. Žmonėms porų masė yra apie 120 000 kDa, kuri yra 40 kartų didesnė už ribosomos masę; tuo pačiu metu mielių branduolinėse porose yra apie 50 baltymų, o stuburinių – keli šimtai. Nors porų skersmuo yra 100 nm, tarpo plotis, per kurį gali praeiti molekulės, dėl porų viduje esančių reguliavimo sistemų yra tik 9 nm. Pro tokį tarpą gali prasiskverbti vandenyje tirpios mažos molekulės, bet ne didelės molekulės, tokios kaip nukleino rūgštys ir dideli baltymai; Norint perkelti šias molekules į branduolį, reikalingas aktyvus (tai yra energiją vartojantis) transportas. Tipiškos žinduolių ląstelės branduolio apvalkale yra nuo 3000 iki 4000 porų, o kiekviena dviejų branduolinių membranų sandūroje turi žiedinę struktūrą su 8 simetrijos ašimis. Prie žiedo pritvirtinta speciali struktūra, žinoma kaip branduolinis krepšelis, kuris išsikiša į nukleoplazmą, o keletas jo siūlų išsikiša į citoplazmą. Abi struktūros yra reikalingos tarpininkauti transportuojančių branduolinių baltymų prisijungimui.
Daugumą baltymų, ribosomų subvienetų ir kai kurių DNR per branduolio poras perneša transportavimo faktorių šeima, vadinama karioferinai. Taip pat vadinami karioferinai, kurie tarpininkauja pernešimui į branduolį importins, ir tarpininkaujant transportui iš branduolio – eksportinų. Dauguma karioferinų tiesiogiai sąveikauja su savo kroviniu, tačiau kai kurie naudoja adapteris voverės. Steroidiniai hormonai (pvz., kortizolis ir aldosteronas), taip pat kitos riebaluose tirpios mažos molekulės gali pasklisti į citoplazmą į ląstelės vidų per ląstelės membraną; citoplazmoje jie jungiasi prie baltymų branduolio receptorių, kurie juos pristato į branduolį. Čia branduoliniai receptoriai surišti su jų ligandais veikia kaip transkripcijos faktoriai, o nesant ligando daugelis receptorių veikia kaip histono deacetilazės, slopinančios tam tikrų genų ekspresiją.
branduolinė lamina
Gyvūnų ląstelėse branduolio mechaninę atramą užtikrina du tarpinių gijų tinklai: branduolio sluoksnis, kuris yra tarpinių gijų tinklas vidiniame branduolio paviršiuje, ir mažiau organizuoti siūlai citozoliniame branduolio paviršiuje. Abi gijų sistemos palaiko branduolį ir padeda įtvirtinti chromosomas bei branduolines poras.
Branduolinę sluoksnį daugiausia sudaro baltymai, žinomi kaip laminatas. Kaip ir visi baltymai, laminai sintetinami citoplazmoje ir toliau transportuojami į branduolio vidų, kur įterpiami į branduolio sluoksnį. Laminos, esančios išorinėje branduolio apvalkalo pusėje (pvz emerinas Ir nesprin), jungiasi su citoskeleto elementais, kurie suteikia branduoliui struktūrinę paramą. Laminai taip pat randami nukleoplazmoje, kur jie sudaro kitą taisyklingą struktūrą, žinomą kaip nukleoplazminis šydas. nukleoplazminis šydas) ; pastarąjį galima vizualizuoti naudojant fluorescencinę mikroskopiją. Šydo funkcija nežinoma, tačiau žinoma, kad jos branduolyje nėra ir yra ląstelės ciklo tarpfazėje. Laminai, sudarantys šydą (pvz., LEM3), jungiasi prie chromatino, o jų struktūros sutrikimai slopina baltymus koduojančių genų transkripciją.
Kaip ir kiti tarpiniai gijų baltymai, laminato monomerai turi α-spiralinį domeną, kurį abu monomerai naudoja, kad susivyniotų vienas aplink kitą, kad susidarytų dimeras, turintis tokią struktūrą. bispiralės. Abu dimerai yra toliau sujungti savo šoniniais paviršiais antilygiagrečiai, sudarydami tetramerą, žinomą kaip protofilamentas. Aštuoni tetramerai yra sujungti į susuktą, virvę primenantį siūlą. Kaitinimo siūlus galima surinkti ir išardyti dinamiškai, t.y. gijos ilgis priklauso nuo santykinių jo surinkimo ir išmontavimo greičių.
Chromosomos
Branduolys turi didžiąją dalį ląstelės genetinės medžiagos, kurią sudaro daugybė linijinių DNR molekulių, kurios yra suskirstytos į struktūras, žinomas kaip chromosomos. Bendras DNR molekulių ilgis žmogaus ląstelėje yra apie 2 m. Per didžiąją ląstelių ciklo dalį šios molekulės kartu su baltymais sudaro vadinamąjį chromatiną, o dalijantis chromosomos atrodo kaip atskiros, gerai išsiskiriančios chromosomos, sudarančios kariotipą. Nedidelis ląstelių genetinės medžiagos kiekis yra mitochondrijose, o augalo ląstelės atveju – chloroplastuose.
Yra du chromatino tipai. Euchromatine DNR yra mažiausiai tankiai organizuota; jame yra dažniausiai transkribuojami genai. Kita chromatino rūšis, heterochromatinas, yra kompaktiškesnė ir jame yra retai arba niekada netranskribuojama DNR. Heterochromatinas skirstomas į fakultatyvinį, kuris yra tik tam tikro tipo ląstelėse ir tam tikrame ląstelių ciklo etape, ir konstitucinį, atstovaujamą chromosomų struktūromis, tokiomis kaip telomerai ir centromerai. Interfazės metu kiekvienos chromosomos chromatinas užima griežtai apibrėžtą branduolio sritį - chromosomų teritorija. Aktyvūs genai, kurie dažniausiai būna euchromatine, dažniausiai yra ties chromosomos teritorijos riba.
branduoliniai kūnai
Žinduolių ląstelių branduolyje yra keletas atskirų pogrupių, vadinamų branduoliniais kūnais. Jie atlieka branduolio suskaidymą, sukurdami jame atskiras erdves, turinčias tam tikrų savybių. Daugelis branduolinių kūnų atlieka specifines funkcijas, tokias kaip ikiribosominės RNR sintezė ir apdorojimas branduolyje, susiliejimo komponentų kaupimasis ir surinkimas dėmėse (žr. toliau) arba RNR molekulių kaupimasis paraspeckles. Mechanizmai, užtikrinantys branduolinių kūnų šių funkcijų atlikimą, yra labai įvairūs. Kai kuriais atvejais branduolinis kūnas gali būti tam tikrų procesų, pavyzdžiui, transkripcijos, vieta. Kitais atvejais branduoliniai kūnai, matyt, netiesiogiai reguliuoja vietines jų komponentų koncentracijas nukleoplazmoje. Kaip ir citoplazminėse organelėse, branduoliniuose kūnuose yra specifinis baltymų rinkinys, lemiantis jų struktūrą molekuliniu lygmeniu. Tačiau, skirtingai nuo citoplazminių organelių, branduoliniai kūnai nėra apsupti lipidų membranų, o jų struktūrinį vientisumą visiškai užtikrina baltymų ir baltymų bei RNR ir baltymų sąveika. Žemiau esančioje lentelėje pateikiamos pagrindinės branduolinių kūnų charakteristikos.
| branduolinis kūnas | Funkcijos | Būdingi komponentai | Tipinis dydis (µm) | Kiekis viename šerdyje |
|---|---|---|---|---|
| branduolys | Ribosomų biogenezė | Mašinos RNR polimerazė I, rRNR apdorojimo faktoriai ir ribosomų subvienetų surinkimas | 3-8 | 1-4 |
| Dėmės | Sujungimo faktorių kaupimas ir surinkimas | Pre-mRNR splaisingo faktoriai | 2-3 | 20-50 |
| Branduolinių kūnų įtampa | Transkripcijos ir sujungimo reguliavimas esant stresui | HSF1, HAP | 1-2 | 3-6 |
| Histono lokusų kūnas | Histono išankstinis mRNR apdorojimas | NPAT, BLYKSTĖ, U7 snRNP | 0,2-1,2 | 2-4 |
| Cajal kūnas | Mažų RNR biogenezė, brendimas ir cirkuliacija | ritė, SMN | 0,2-1,5 | 1-10 |
| PML kūnas | Genomo stabilumo reguliavimas, DNR atstatymas, transkripcijos kontrolė, apsauga nuo virusų | PML | 0,1-1 | 10-30 |
| Paraspeckles | mRNR reguliavimas, RNR redagavimas | Nekoduojančios RNR NEAT1/MENε/β, PSP1 baltymai, p54 nrb /NONO | 0,2-1 | 2-20 |
| Perinukleolinis skyrius | Susintetintų RNR rinkinio post-transkripcijos reguliavimas RNR polimerazė III | PTB | 0,2-1 | 1-2 |
branduolys
Branduolys yra atskira tanki struktūra branduolyje. Jis nėra apsuptas membranos ir susidaro toje vietoje, kur yra rDNR – ribosominės RNR (rRNR) genų, vadinamų branduolio organizatoriais, tandeminiai pasikartojimai. Pagrindinė branduolio funkcija yra rRNR sintezė ir ribosomų susidarymas. Branduolio struktūrinis vientisumas priklauso nuo jo aktyvumo, o rRNR genų inaktyvacija veda į branduolinių struktūrų mišinį.
Pirmajame ribosomų susidarymo etape fermentas RNR polimerazė I transkribuoja rDNR ir suformuoja pre-rRNR, kuri toliau suskaidoma į 5,8S, 18S ir 28S rRNR. RRNR transkripcija ir potranskripcijos apdorojimas vyksta branduolyje, dalyvaujant mažoms nukleolinėms RNR (snoRNR), kai kurios iš jų yra kilusios iš susijungusių genų mRNR intronų, koduojančių baltymus, susijusius su ribosomų funkcija. Surinkti ribosomų subvienetai yra didžiausios struktūros, einančios per branduolio poras.
Žiūrint pro elektroninį mikroskopą, branduolyje galima išskirti tris komponentus: fibrilinius centrus (FC), juos supantį tankų pluoštinį komponentą (CFC) ir granuliuotą komponentą (GC), kuris, savo ruožtu, supa CFC. rRNR transkripcija vyksta FC ir FC bei PFC ribose, todėl suaktyvėjus ribosomų susidarymui FC tampa aiškiai atskiriamas. RRNR pjaustymas ir modifikavimas vyksta PFC, o vėlesni ribosomų subvienetų susidarymo etapai, įskaitant ribosomų baltymų įkėlimą, vyksta HA.
Cajal kūnas
Cajal kūnas (TC) yra branduolinis kūnas, randamas visuose eukariotuose. Jis identifikuojamas pagal kodinio baltymo ir specifinių RNR (scaRNR) buvimą. TK taip pat yra SMN baltymo. motorinių neuronų išlikimas). MA turi didelę susiliejančių mažų branduolinių ribonukleoproteinų (snRNP) ir kitų RNR apdorojimo faktorių koncentraciją, todėl manoma, kad MA tarnauja kaip susijungimo faktorių surinkimo ir (arba) po transkripcijos modifikavimo vietos. TK yra branduolyje tarpfazės metu, bet išnyksta mitozės metu. TC biogenezėje atsekamos savaime besitvarkančios struktūros savybės.
Kai SMN tarpląstelinė lokalizacija pirmą kartą buvo ištirta imunofluorescencijos būdu, baltymas buvo rastas visoje citoplazmoje, taip pat branduoliniame kūne, panašaus dydžio į MC ir dažnai greta MC. Dėl šios priežasties šis kūnas buvo vadinamas „TK dvyniu“ (angl. CB dvyniai) arba tiesiog brangakmenis. Tačiau paaiškėjo, kad HeLa ląstelių linija, kurioje buvo aptiktas naujas kūnas, buvo neįprasta: kitose žmogaus ląstelių linijose, taip pat vaisinėje muselėje. Drosophila melanogaster SMN kolokalizuotas su coilin TC. Todėl bendru atveju SMN gali būti laikomas svarbiu TC komponentu, o ne kaip atskiro branduolinio kūno žymekliu.
Histono lokusų kūnas
PML kūnai
Dėmės
Paraspeckles
Paraspeckles yra netaisyklingos formos branduoliniai kūnai, esantys tarpchromatinėje branduolio erdvėje. Pirmiausia jie buvo aprašyti HeLa ląstelėse, kurių viename branduolyje yra 10–30 paraspeckles, tačiau dabar paraspeckles randamos visose pirminėse žmogaus ląstelėse, transformuotų linijų ląstelėse ir audinių pjūviuose. Jie gavo savo vardą dėl savo vietos šerdyje - šalia dėmių.
Paraspeckles yra dinamiškos struktūros, kurios keičiasi reaguodamos į ląstelės metabolinio aktyvumo pokyčius. Jie priklauso nuo transkripcijos, o nesant RNR polimerazės II transkripcijos, paraspeckles išnyksta ir visi jų baltymai (PSP1, p54nrb, PSP2, CFI(m)68 ir PSF) sudaro pjautuvo formos perinukleolinį dangtelį. Šis reiškinys stebimas ląstelių ciklo metu: tarpfazėje ir visose mitozės fazėse yra paraspeckles, išskyrus telofazę. Telofazės metu susidaro dukteriniai branduoliai, o RNR polimerazė II nieko netranskribuoja, todėl paraspeckle baltymai sudaro perinukleolinį dangtelį. Paraspeckles dalyvauja reguliuojant genų ekspresiją, kaupdamos tas RNR, kuriose yra dvigrandžių regionų, kuriuos reikia redaguoti, ty adenozino pavertimą inozinu. Per šį mechanizmą paraspeckles dalyvauja kontroliuojant genų ekspresiją diferenciacijos, virusinės infekcijos ir streso metu.
Perinukleolinis skyrius
Perinukleolinis skyrius (OK) yra netaisyklingos formos branduolinis kūnas, kuriam būdinga tai, kad jis yra branduolio periferijoje. Nepaisant to, kad abu skyriai yra fiziškai susiję, jie struktūriškai skiriasi. TC paprastai randama piktybinių navikų ląstelėse. OK yra dinamiška struktūra, joje yra daug RNR surišančių baltymų ir RNR polimerazės III. OK struktūrinį stabilumą užtikrina RNR polimerazės III vykdoma transkripcija ir pagrindinių baltymų buvimas. Kadangi TC buvimas dažniausiai siejamas su piktybiniais navikais ir gebėjimu metastazuoti, jie laikomi potencialiais vėžio ir kitų piktybinių navikų žymenimis. Buvo parodytas OK ryšys su specifiniais DNR lokusais.
Branduolinių kūnų įtampa
Šilumos šoko metu branduolyje susidaro stresiniai branduoliniai kūnai. Jie susidaro tiesiogiai sąveikaujant šilumos šoko transkripcijos faktoriui 1 ( HSF1) ir pericentriniai tandemo pakartojimai III palydovo sekoje, atitinkantys III palydovo nekoduojančių transkriptų aktyvios transkripcijos vietas. Plačiai manoma, kad tokie kūnai atitinka labai tankiai supakuotas ribonukleoproteinų kompleksų formas. Manoma, kad streso patiriančiose ląstelėse jie dalyvauja greituose, laikinuose ir visuotiniuose genų ekspresijos pokyčiuose dėl įvairių mechanizmų, tokių kaip chromatino remodeliavimas ir transkripcijos bei splaisingo faktorių įsisavinimas. Ląstelėse normaliomis (ne stresinėmis) sąlygomis įtemptų branduolinių kūnų randama retai, tačiau jų skaičius smarkiai padidėja veikiant šilumos šokui. Streso branduoliniai kūnai randami tik žmogaus ir kitų primatų ląstelėse.
Našlaičiai branduoliniai kūnai
| branduolinis kūnas | apibūdinimas | Tipinis dydis (µm) | Kiekis viename šerdyje |
|---|---|---|---|
| Klastosoma | Koncentruoja 20S ir 19S proteasomų kompleksus ir su ubikvitinu susijusius baltymus. Jis randamas daugiausia, kai stimuliuojamas proteasomų aktyvumas, ir pašalinamas, kai proteasomų aktyvumas slopinamas. | 0,2-1,2 | 0-3 |
| Skyriaus korpusas klirenso korpusas) | Praturtintas dalijimosi faktoriais CstF Ir CPSF, taip pat baltymų DDX1 kuriuose yra DEAD dėžė. Jis randamas daugiausia S fazėje ir nėra paveiktas transkripcijos slopinimo. | 0,2-1,0 | 1-4 |
| OPT domenas | Praturtintas transkripcijos faktoriais spalio 1 d ir PTF. Iš dalies kolokalizuojasi su transkripcijos vietomis. Daugiausia randama vėlyvoje G1 fazėje, išardyta slopinant transkripciją. | 1,0-1,5 | 1-3 |
| Polycomb korpusas | Aptinkama žmogaus ir Drosophila ląstelėse, praturtinta PcG baltymu. Kaupia RING1 baltymus žmonėms, KMI1, HPC, gali būti susijęs su pericentromeriniu heterochromatinu. | 0,3-1,0 | 12-16 |
| Jautis Sam68 | Kaupia Sam68 baltymą ir panašius baltymus SLM-1 ir SLM-2. Išardyta slopinant transkripciją. Turbūt gausu RNR. | 0,6-1,0 | 2-5 |
| SUMO korpusas | Praturtintas SUMO baltymais ir SUMO konjuguojančiu fermentu Ubc9. Koncentruoja transkripcijos faktorius pCREB, CBP, c-birželio mėn. | 1-3 | 1-3 |
Funkcijos
Branduolinis apvalkalas apsaugo ląstelės DNR ir dalyvauja daug sudėtingesniame genų ekspresijos reguliavime, palyginti su prokariotine ląstele. Prokariotuose transkripcija ir transliacija yra susieti procesai, o mRNR transliacija į baltymą prasideda dar prieš jai visiškai susintetinant. Eukariotinėse ląstelėse citoplazma, kurioje vyksta transliacija ir branduolyje vykstanti transkripcija, yra erdviškai atskirta, todėl reikia užtikrinti molekulių transportavimą tarp branduolio ir citoplazmos.
Branduolinis apvalkalas suteikia branduoliui kontroliuoti jo turinį ir atskiria jį nuo likusios citoplazmos. Tai svarbu reguliuoti procesus, vykstančius abiejose branduolinio apvalkalo pusėse. Kai citoplazminį procesą reikia kažkaip apriboti, tada dažniausiai pagrindinis jo dalyvis perkeliamas į branduolį, kur jis sąveikauja su transkripcijos faktoriais ir taip sukelia kai kurių citoplazminiame procese dalyvaujančių fermentų susidarymo slopinimą. Pavyzdžiui, glikolizė, procesas, kurio metu ląstelė išgauna energiją iš gliukozės molekulės, turi tokį reguliavimo mechanizmą. Pirmąją glikolizės reakciją vykdo fermentas heksokinazė, paverčiant gliukozės molekulę į gliukozės-6-fosfatą. Padidėjus fruktozės-6-fosfato (medžiagos, susidarančios iš gliukozės-6-fosfato glikolizės metu) koncentracijai, reguliuojantis baltymas siunčia heksokinazę į branduolį, kur susidaro transkripcijos represinis kompleksas, slopinantis glikolitinių fermentų koduojančių genų ekspresiją.
Siekiant kontroliuoti, kurie genai yra transkribuojami, transkripcijos faktoriai ląstelėje neturi fizinės prieigos prie DNR, kol jie nėra aktyvuojami tam tikrame signalizacijos kelyje. Tai užkerta kelią net žemai netinkamų genų ekspresijai. Visų pirma, NF-κB kontroliuojamų genų, dalyvaujančių uždegiminiame procese, atveju transkripciją sukelia signalizacijos kelias, pavyzdžiui, pradedant TNF-α signalizacijos molekulės prisijungimu prie jos receptorių ląstelės membranoje. ir galiausiai suaktyvina transkripcijos faktorių NF-κB. Branduolinės lokalizacijos signalas, esantis NF-κB, leidžia jam patekti į branduolį ir iš jo per branduolio poras; branduolyje jis skatina tikslinių genų transkripciją.
Dalijimas neleidžia ląstelei perrašyti nesujungtos mRNR. Eukariotinėse mRNR yra intronų, kurie turi būti pašalinti prieš prasidedant mRNR transliacijai. Sujungimas, tai yra intronų pašalinimas, vyksta branduolyje, o tai neleidžia ribosomoms, esančioms už branduolio, patekti į pre-mRNR. Jei nebūtų branduolio, ribosomos pradėtų versti nesubrendusią iRNR, o tai lemtų neteisingų baltymų produktų susidarymą.
Kadangi transkripcija vyksta branduolyje, branduolyje yra daug baltymų, tiesiogiai dalyvaujančių arba reguliuojančių transkripciją. Šie baltymai apima helikazes, kurios išvynioja DNR dvigubą spiralę, palengvindamos kitų baltymų patekimą į ją, RNR polimerazes, kurios sintetina RNR, topoizomerazes, kurios veikia DNR topologiją, ir įvairius transkripcijos faktorius.
branduolinis transportas
Didelių molekulių išėjimą iš branduolio ir patekimą į branduolį kontroliuoja branduolio poros. Nors mažos molekulės gali patekti į branduolį be jokio reguliavimo, makromolekulės, tokios kaip baltymai ir RNR, turi prisijungti prie karioferinų, kad galėtų patekti į branduolį (importinai) ir iš branduolio (eksportinai). Baltymai, kurie turi būti transportuojami iš citoplazmos į branduolį, turi specifinę aminorūgščių seką, žinomą kaip branduolio lokalizacijos signalas, prie kurios jungiasi importinai. Panašiai yra baltymų, kurie turi išeiti iš branduolio branduolinės energijos eksporto signalas, pripažino eksportins. Importinų ir eksportinų gebėjimą gabenti savo krovinį reguliuoja GTPazės – fermentai, hidrolizuojantys GTP, kad išskirstytų energiją. Pagrindinė branduolinio transporto GTPazė yra bėgo, kuris gali prisijungti prie GTP arba GDP, priklausomai nuo jo vietos (branduolyne arba citoplazmoje). Iš esmės Ran-GTP sąveika su importinu sukelia pastarojo konformacinius pokyčius, kad jis atsiskirtų nuo vežamo krovinio. Susidaręs Ran-GTP ir importino kompleksas pernešamas į citoplazmą, kur RanBP baltymas atskiria Ran-GTP nuo importino. Atskyrimas nuo importino leidžia baltymams tarpas susisiekite su Ran-GTP ir katalizuoja GTP hidrolizę į BVP. Be to, Ran-BVP kompleksą atpažįsta baltymas NUTF2, kuris grąžina jį į nukleoplazmą. baltymas branduolyje GEF pakeičia BVP GTP, sudarydamas Ran-GTP ir uždarydamas ciklą.
Surinkimas ir išmontavimas
Ląstelės gyvavimo metu branduolys gali būti išardomas (ląstelių dalijimosi ar apoptozės metu). Šių procesų metu sunaikinami branduolio struktūriniai komponentai – branduolio apvalkalas ir branduolio sluoksnis. Daugumoje ląstelių branduolio išardymas vyksta mitozės fazės metu. Tačiau branduolio išardymas neapsiriboja tik mitoze ir vyksta ne visose ląstelėse. Kai kurie vienaląsčiai eukariotai (pvz., mielės) patiria vadinamąją uždarą mitozę, kurios metu branduolinis apvalkalas lieka nepažeistas. Esant uždarai mitozei, chromosomos persikelia į skirtingas branduolio puses, kurios vėliau dalijasi į dvi dalis. Priešingai, aukštesniųjų eukariotų ląstelėse paprastai vyksta atvira mitozė, kurios metu suyra branduolio apvalkalas. Chromosomos migruoja į skirtingus veleno polius, o aplink juos vėl susidaro du branduoliai. Branduolinė plokštelė taip pat yra išardoma dėl laminų fosforilinimo kinazėmis, tokiomis kaip nuo ciklino priklausoma proteinkinazė 1. Branduolinės plokštelės surinkimas dukteriniuose branduoliuose prasideda po sluoksnių defosforilinimo.
Apoptozė yra kontroliuojamas ląstelių komponentų sunaikinimo procesas, sukeliantis ląstelių mirtį. Su apoptoze susiję pokyčiai vyksta tiesiogiai branduolyje ir jo turinyje. Tai apima chromatino kondensaciją, taip pat branduolio apvalkalo ir branduolio sluoksnio suirimą. Laminų tinklo skilimą skatina apoptotinės proteazės, žinomos kaip kaspazės, kurios suardo laminatus ir taip paveikia branduolio struktūrinį vientisumą. Laminų sunaikinimas kartais naudojamas kaip kaspazės aktyvumo rodiklis apoptozės tyrimuose. Ląstelės, ekspresuojančios kaspazei atsparius mutantus, apoptozės metu nepraranda branduolio vientisumo, todėl laminai vaidina pagrindinį vaidmenį inicijuojant pokyčius, kuriuos branduolys patiria apoptozės metu. Be to, laminų surinkimo į tinklą slopinimas sukelia apoptozę.
Įvairių eukariotų branduolių ypatumai
Eukariotų branduolių dydžiai, formos ir morfologija labai skiriasi. Jeigu piroplazmidas ir Leishmania, branduolio skersmuo yra 1-3 mikronai, tada kai kuriuose radiolariuose branduolių skersmuo siekia 400 mikronų ir net 1 mm. Paprastai daugumos eukariotų branduolio forma yra artima sferinei, tačiau kartais jis gali įgauti gana keistas formas (tai ypač pasakytina apie blakstienas makrobranduolius). Nors visuose eukariotuose branduolio apvalkalas susideda iš dviejų membranų, jame esančių porų skaičius įvairiose rūšyse labai skiriasi, o kartais prie jo gali priglusti papildomi sluoksniai (tiek išorėje, tiek viduje); pavyzdžiui, daugelyje laisvai gyvenančių amebų prie apvalkalo vidinės pusės ribojasi pluoštinis sluoksnis, turintis ląstelinę struktūrą, kuris savo storiu gerokai viršija branduolio apvalkalą, o radiolarijose išorinėje apvalkalo pusėje yra papildomi fibriliniai sluoksniai. .
Dinoflagellate tipo protistų (Dinoflagellata) branduolio organizacija išsiskiria dideliu originalumu. Dauguma jų atstovų turi branduolį, kuriame yra chromosomos kondensuotas per visą ląstelių ciklą (įskaitant tarpfazę) ir praktiškai neturi histonų. Šis branduolio tipas vadinamas dinokarionas. Tuo pačiu metu DNR kiekis dinokarione yra dešimtis ir šimtus kartų didesnis nei DNR kiekis ląstelėje kitų eukariotų grupių atstovuose. Tačiau kai kurie dinoflagellatai ( Noctiluca, Oodinium ) turi normalius eukariotų branduolius; kitų tipo atstovų vegetacinėse ląstelėse branduoliai yra įprasti, o dinokarionas yra kituose ląstelių ciklo etapuose (pavyzdžiui, gametose).
Protistų ląstelės turi bent vieną branduolį. Tuo pačiu metu nebranduolinių ląstelių randama ir Metazoa organizmuose, kurie, neturėdami branduolio, prarado gebėjimą dalytis susidarant dviem dukterinėms ląstelėms. Žinomiausias ląstelių neturinčių branduolių pavyzdys yra žinduolių eritrocitai, kuriems taip pat trūksta kitų organelių, tokių kaip mitochondrijos. Raudonieji kraujo kūneliai bręsta kaulų čiulpuose eritropoezės proceso metu, kurio metu netenka branduolių, kitų organelių ir ribosomų. Branduolys išstumiamas iš ląstelės eritroblasto diferenciacijos į retikulocitą proceso metu, kuris veikia kaip tiesioginis eritrocito pirmtakas. Kai kurių mutagenų įtakoje į kraują gali patekti nesubrendę eritrocitai, kuriuose yra mikrobranduolių.
Dauguma protistų turi tik vieną branduolį; protistams, kuriems būdingas sudėtingas gyvenimo ciklas (pavyzdžiui, apicomplexa tipo (Apicomplexa) atstovai turi vieno branduolio ir kelių branduolių stadijas).
Daugiabranduolės protistų ląstelės
Chloroplastų sudėtis kriptofituose ir chlorarachniofituose yra nukleomorfas- sumažintas fototrofinio endosimbiono branduolys, įtrauktas šių dumblių protėvių antrinės endosimbiozės metu (Cryptophyta buvo įtraukta raudonųjų dumblių, o Chlorarachnea - žaliųjų dumblių)
LĄSTELĖS BRANDUOLIS
Šerdis(branduolys, s. karionas) yra visose žmogaus ląstelėse, išskyrus eritrocitus ir trombocitus. Branduolio funkcijos – paveldimos informacijos saugojimas ir perdavimas naujoms (dukterinėms) ląstelėms. Šios funkcijos yra susijusios su DNR buvimu branduolyje. Baltymų sintezė taip pat vyksta branduolyje – ribonukleino rūgšties RNR ir ribosominėse medžiagose.
Daugumoje ląstelių branduolys yra sferinis arba kiaušiniškas, tačiau yra ir kitų branduolio formų (žiedinis, lazdelės formos, fusiforminis, karoliuko formos, pupelės formos, segmentuotas, kriaušės formos, polimorfinis). Branduolio dydis labai įvairus – nuo 3 iki 25 mikronų. Kiaušialąstė turi didžiausią branduolį. Dauguma žmogaus ląstelių yra vienabranduolinės, tačiau yra ir dvibranduolių (kai kurie neuronai, hepatocitai, kardiomiocitai). Kai kurios struktūros yra daugiabranduolės (raumenų skaidulos). Branduolys turi branduolio apvalkalą, chromatiną, branduolį ir nukleoplazmą (4 pav.).
Branduolinė membrana arba karioteka (caryothéca), atskirianti branduolio turinį nuo citoplazmos, susideda iš vidinės ir išorinės 8 nm storio branduolio membranos. Membranas skiria 20-50 nm pločio perinuklearinė erdvė (kariotekos cisterna), kurioje yra smulkiagrūdė vidutinio elektronų tankio medžiaga. Išorinė branduolio membrana pereina į granuliuotą endoplazminį tinklą. Poeto Mu perinuklearinė erdvė yra viena ertmė su endoplazminiu tinklu. Vidinė branduolio membrana viduje yra sujungta su plačiu baltymų fibrilių tinklu, susidedančiu iš atskirų subvienetų.
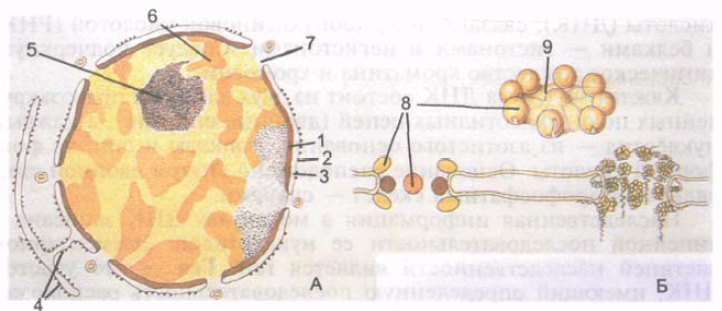
Ryžiai. 4. Ląstelės branduolys (A) ir branduolių porų kompleksas (B). 1 - vidinė branduolio membrana; 2 - išorinė branduolinė membrana; 3 - perinuklearinė erdvė; 4 - endoplazminio tinklo membranos; 5 - branduolys; 6 - dekondensuotas chromatinas; 7 - branduolinis laikas; 8 - branduolinės poros granulės; 9 - branduolinių porų diafragma.
Branduoliniame apvalkale yra daug suapvalintų branduolinių porų, kurių kiekvienos skersmuo 50–70 nm. Branduolinės poros iš viso užima iki 25% branduolio paviršiaus. Porų skaičius viename branduolyje siekia 3000-4000. Išilgai porų kraštų išorinė ir vidinė membranos yra sujungtos viena su kita ir sudaro vadinamąjį porų žiedą. Kiekviena pora uždaroma diafragma, dar vadinama porų kompleksu. Porų diafragmos turi sudėtingą struktūrą, jas sudaro tarpusavyje sujungtos baltymų granulės. Per branduolines poras atliekamas selektyvus didelių dalelių pernešimas, taip pat medžiagų apykaita tarp ląstelės branduolio ir citozolio.
Po branduoliniu apvalkalu yra nukleoplazma (karioplazma)(nukleoplazma, s. karyoplasma), kuri turi vienalytę struktūrą, ir branduolys. Nesidalijančio branduolio nukleoplazmoje, jo branduolinio baltymo matricoje, yra vadinamojo heterochromatino osmiofilinės granulės (gumbeliai). Labiau atsipalaidavusios chromatino sritys, esančios tarp granulių, vadinamos euchromatinu. Birus chromatinas dar vadinamas dekondensuotu chromatinu, jame sintetiniai procesai vyksta intensyviausiai. Ląstelių dalijimosi metu chromatinas sutirštėja, kondensuojasi ir susidaro chromosomos.
Chromatinas nesidalijančio branduolio (chromatiną) ir besidalijančiojo chromosomas sudaro dezoksiribonukleino rūgšties (DNR) molekulės, susietos su ribonukleino rūgštimi (RNR) ir baltymais – histonais ir nehistonais. Reikėtų pabrėžti cheminę chromatino ir chromosomų tapatybę.
Kiekvieną DNR molekulę sudaro dvi ilgos dešiniosios polinukleotidų grandinės (dvigubos spiralės), o kiekvienas nukleotidas susideda iš azoto bazės, gliukozės ir fosforo rūgšties liekanos. Pagrindas yra dvigubos spiralės viduje, o cukraus-fosfato skeletas yra išorėje.
Paveldima informacija DNR molekulėse rašoma tiesine jos nukleotidų seka. Paveldimumo elementarioji dalelė yra genas. Genas yra DNR dalis, turinti specifinę nukleotidų seką, atsakingą už vieno konkretaus baltymo sintezę.
DNR molekulė branduolyje yra kompaktiškai supakuota. Taigi viena DNR molekulė, turinti 1 milijoną nukleotidų, su jų linijiniu išsidėstymu, užimtų tik 0,34 mm ilgio segmentą. Vienos žmogaus chromosomos ilgis ištemptoje formoje yra apie 5 cm, tačiau sutankintoje būsenoje chromosomos tūris yra apie 10–15 cm 3.
DNR molekulės, susijusios su histono baltymais, sudaro nukleosomas, kurios yra struktūriniai chromatino vienetai. Nukleosoma turi 10 nm skersmens karoliuko formą. Kiekviena nukleosoma sudaryta iš histonų, aplink kuriuos susisukusi 146 bp DNR grandinė. Tarp nukleozomų yra linijinės DNR dalys, susidedančios iš 60 nukleotidų porų.
Chromatiną vaizduoja fibrilės, kurios sudaro apie 0,4 μm ilgio kilpas, kuriose yra nuo 20 000 iki 30 000 bazinių porų.
Dėl dezoksiribonukleoproteinų (DNP) sutankinimo (kondensacijos) ir sukimosi (superspecializacijos) besidalijančiame branduolyje išryškėja chromosomos. Šios struktūros yra chromosomos(chromasomae, iš graikų chruma – dažai, soma – kūnas) – tai pailgos strypo formos dariniai, kurių dvi rankos atskirtos vadinamuoju susiaurėjimu – centromera. Priklausomai nuo centromeros vietos ir santykinės rankų (kojų) padėties bei ilgio, išskiriami trys chromosomų tipai: metacentrinės, turinčios maždaug vienodas rankas; submetacentrinis, kuriame pečių ilgis skiriasi; akrocentrinis, kurio vienas petys ilgas, o kitas labai trumpas, vos pastebimas. Chromosomoje yra eu- ir heterochromatiniai regionai. Pastarieji nesidalijančiame branduolyje ir ankstyvoje mitozės fazėje išlieka kompaktiški. Chromosomoms identifikuoti naudojamas eu- ir heterochromatinių sričių kaitaliojimas.
Chromosomų paviršius yra padengtas įvairiomis molekulėmis, daugiausia ribonukleoproteinų (RNP). Somatinės ląstelės turi po 2 kiekvienos chromosomos kopijas, jos vadinamos homologinėmis. Jie yra vienodo ilgio, formos, struktūros, turi tuos pačius genus, kurie yra vienodai išsidėstę. Struktūrinės ypatybės, chromosomų skaičius ir dydis vadinami kariotipu. Normalus žmogaus kariotipas apima 22 poras autosomų ir vieną porą lytinių chromosomų (XX arba XY). Somatinės žmogaus ląstelės (diploidinės) turi dvigubą chromosomų skaičių – 46. Lytinėse ląstelėse yra haploidinis (vienas) rinkinys – 23 chromosomos. Todėl lytinėse ląstelėse DNR yra 2 kartus mažiau nei diploidinėse somatinėse ląstelėse.
branduolys(nucleolus), vienas ar daugiau, yra visose nesidalijančiose ląstelėse. Jis turi intensyviai dažyto apvalaus kūno formą, kurio dydis yra proporcingas baltymų sintezės intensyvumui. Branduolys susideda iš elektronų tankios nukleolonemos (iš graikų peta – siūlas), kurioje išskiriama siūlinė (fibrilinė) dalis, susidedanti iš daugybės susipynusių apie 5 nm storio RNR gijų, ir granuliuota dalis. Granuliuotą (granuliuotą) dalį sudaro apie 15 nm skersmens grūdeliai, kurie yra RNP dalelės – ribosomų subvienetų pirmtakai. Perinukleolinis chromatinas yra įterptas į nukleolonemos įdubas. Ribosomos susidaro branduolyje.
LĄSTELIŲ SKYRIMAS. LĄSTELĖS CIKLAS
Organizmas auga dalijantis didindamas ląstelių skaičių. Pagrindiniai ląstelių dalijimosi būdai žmogaus organizme yra mitozė ir mejozė. Procesai, vykstantys taikant šiuos ląstelių dalijimosi metodus, vyksta vienodai, tačiau lemia skirtingus rezultatus.
Mitozinių ląstelių dalijimasis(mitozė) lemia ląstelių skaičiaus padidėjimą, organizmo augimą. Tokiu būdu užtikrinamas ląstelių atsinaujinimas, kai jos susidėvi ir miršta. Dabar žinoma, kad epidermio ląstelės gyvena 10-30 dienų, eritrocitai – iki 4-5 mėnesių. Nervų ir raumenų ląstelės (skaidulos) gyvena visą žmogaus gyvenimą.
Visose ląstelėse dauginimosi (dalijimosi) metu pastebimi pokyčiai, kurie patenka į ląstelės ciklo rėmus. ląstelių ciklas vadinami procesai, vykstantys ląstelėje nuo dalijimosi iki dalijimosi arba nuo dalijimosi iki ląstelės mirties (mirties). Ląstelių cikle išskiriamas ląstelės paruošimas dalijimuisi (interfazė) ir mitozė (ląstelių dalijimosi procesas).
Interfazėje, kuri trunka maždaug 20-30 valandų, didėja biosintezės procesų greitis, daugėja organelių. Šiuo metu ląstelės ir visų jos struktūrinių komponentų, įskaitant centrioles, masė padvigubėja.
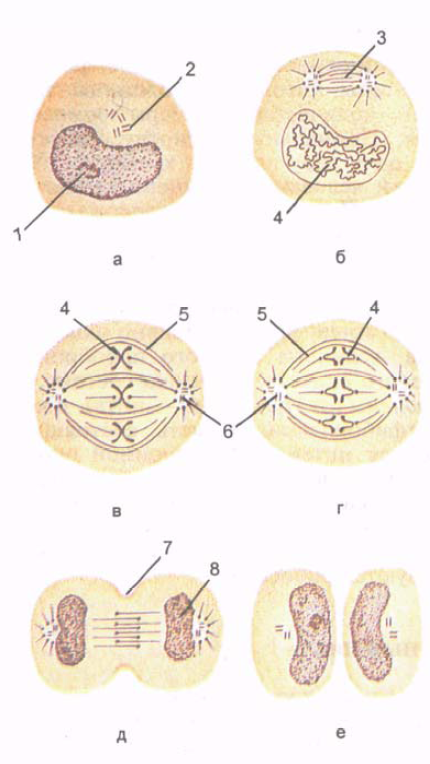
Ryžiai. 5. Ląstelių dalijimasis. mitozės stadijos. Parodytas chromosomų susidarymas, dalijimosi veleno susidarymas ir vienodas chromosomų bei centriolių pasiskirstymas dviejose dukterinėse ląstelėse.
A - tarpfazė; B - profazė; B - metafazė; G - anafazė; D - telofazė; E – vėlyvoji telofazė. 1 - šerdis; 2 - ląstelės centras (centriolė); 3 - ląstelių dalijimosi velenas; 4 - chromosomos; 5 - ištisiniai mikrovamzdeliai; 6 - ląstelių centras; 7 - ląstelių dalijimosi vaga; 8 - branduolio susidarymas.
Vyksta nukleorūgščių molekulių replikacija (kartojimasis, padvigubėjimas). Tai yra genetinės informacijos, saugomos tėvų DNR, perdavimo procesas, tiksliai atkuriant ją dukterinėse ląstelėse. Pirminė DNR grandinė yra dukterinės DNR sintezės šablonas. Dėl replikacijos kiekviena iš dviejų dukterinių DNR molekulių susideda iš vienos senos ir vienos naujos grandinės. Pasiruošimo mitozei laikotarpiu ląstelėje sintetinami baltymai, reikalingi ląstelių dalijimuisi. Pasibaigus tarpfazei, chromatinas branduolyje kondensuojasi.
Mitozė(mitozė; iš graikų mitos – siūlas) – laikotarpis, kai motininė ląstelė dalijasi į dvi dukterines ląsteles (5 pav.). Mitozinis ląstelių dalijimasis užtikrina vienodą ląstelių struktūrų pasiskirstymą, jos branduolinę medžiagą – chromatiną – tarp dviejų dukterinių ląstelių. Mitozės trukmė nuo 30 minučių iki 3 valandų Mitozė skirstoma į profazę, metafazę, anafazę, telofazę.
Profazėje branduolys palaipsniui suyra, centrioliai nukrypsta link ląstelės polių. Centrolių mikrovamzdeliai yra nukreipti į pusiaują, o pusiaujo srityje jie persidengia vienas su kitu.
Metafazėje sunaikinamas branduolio apvalkalas, chromosomų gijos siunčiamos į ašigalius, išlaikant ryšį su pusiauju
visos ląstelės ploto. Endoplazminio tinklo ir Golgi komplekso struktūros suyra į mažas pūsleles (pūsleles), kurios kartu su mitochondrijomis pasiskirsto abiejose besidalijančios ląstelės pusėse. Pasibaigus metafazei, kiekviena chromosoma išilginiu plyšiu pradeda skilti į dvi naujas dukterines chromosomas.
Anafazėje chromosomos atsiskiria viena nuo kitos ir nukrypsta link ląstelės polių iki 0,5 µm/min greičiu. Anafazės pabaigoje plazminė membrana invaginuoja išilgai ląstelės pusiaujo, statmenai jos išilginei ašiai, sudarydama dalijimosi vagą.
Telofazėje chromosomos, kurios išsiskyrė į ląstelės polius, dekondensuojasi, pereina į chromatiną ir prasideda RNR transkripcija (gamyba). Susidaro branduolio apvalkalas, branduolys, sparčiai formuojasi būsimų dukterinių ląstelių membraninės struktūros. Ląstelės paviršiuje, išilgai jos pusiaujo, susiaurėjimas gilėja, ląstelė dalijama į dvi dukterines ląsteles.
Dėl mitozinio dalijimosi dukterinės ląstelės gauna chromosomų rinkinį, identišką motinai. Mitozė užtikrina genetinį stabilumą, ląstelių skaičiaus padidėjimą ir atitinkamai organizmo augimą bei regeneracijos procesus.
Mejozė(iš graikų meiosis – redukcija) stebimas lytinėse ląstelėse. Dėl šių ląstelių dalijimosi susidaro naujos ląstelės su vienu (haploidiniu) chromosomų rinkiniu, kuris svarbus genetinės informacijos perdavimui. Kai viena lytinė ląstelė susilieja su priešingos lyties ląstele (apvaisinimo metu), chromosomų rinkinys padvigubėja, tampa pilnas, dvigubas (diploidinis). Diploidinėje (dvibranduolėje) zigotoje, susidariusioje susiliejus lytinėms ląstelėms, yra du identiškų (homologinių) chromosomų rinkiniai. Kiekviena diploidinio organizmo (zigotos) homologinių chromosomų pora kilusi iš kiaušinėlio branduolio ir iš spermos branduolio.
Dėl lytinių ląstelių mejozės brandžiame organizme kiekvienoje dukterinėje ląstelėje yra tik viena iš visų pirminių ląstelių homologinių chromosomų porų. Tai tampa įmanoma, nes mejozės metu vyksta tik DNR replikacija ir du iš eilės branduolio dalijimasis. Dėl to iš vienos diploidinės ląstelės susidaro dvi haploidinės ląstelės. Kiekvienoje iš šių dukterinių ląstelių yra perpus mažiau chromosomų (23) nei motininės ląstelės branduolyje (46). Dėl mejozės haploidinės lytinės ląstelės turi ne tik perpus mažiau chromosomų, bet ir skirtingą genų išsidėstymą chromosomose. Todėl naujasis organizmas turi ne tik savo tėvų savybių sumą, bet ir savo (individualius) bruožus.
Klausimai kartojimui ir savikontrolei
1. Iš kokių elementų susideda ląstelės branduolys? Kokias funkcijas jis atlieka?
2. Papasakokite apie DNR molekulių sandarą.
3. Pateikite chromosomų morfologinę charakteristiką, nurodykite jų klasifikaciją.
4. Kas yra ląstelės ciklas, kokie periodai (fazės) skiriami šiame cikle?
5. Kas yra mejozė, kuo ji skiriasi nuo mitozės?