V jedrih protozojev, pa tudi v jedrih večceličnih organizmov, obstaja lupina, jedrski sok (kariolimfa), kromatin (kromosomi) in nukleoli. Glede na velikost in strukturo jedra pa so različne praživali zelo raznolike (sl.). Te razlike so posledica razmerja strukturne komponente jedra: količina jedrnega soka, število in velikost nukleolov (nukleola), stopnja ohranjenosti strukture kromosomov v interfaznem jedru itd.
riž. Jedra praživali.
A - mikronukleus ciliatov Paramecium aurelia;
B - flagelat Trypanosoma brucei,
B - ameba Amoeba sphaeronuclei;
G - oklepni flagelat Geratium fuscus;
E - radiolarija Aulacantha scolymantha;
E - makrogameta kokcidije Aggregata eberthi;
G - makronukleus ciliatov Epidinium ecaudatum
Večina praživali ima eno jedro. Vendar pa obstajajo tudi večjedrne vrste praživali.
Pri nekaterih praživalih, in sicer infuzorijah in nekaj korenonožcih - foraminiferah, opazimo zanimiv pojav dualizma (dvojnosti) jedrnega aparata. Izhaja iz dejstva, da v telesu najpreprostejšega obstajata dve jedri dveh kategorij, ki se razlikujeta tako po svoji zgradbi kot po fiziološki vlogi v celici. Infuzorije imajo na primer dve vrsti jeder: veliko, s kromatinom bogato jedro – makronukleus in majhno jedro – mikronukleus. Prvi je povezan z izvajanjem vegetativnih funkcij v celici, drugi pa s spolnim procesom.
Najenostavnejši, kot vsi organizmi, se nagibajo k razmnoževanju. Pri protozojih obstajata dve glavni obliki razmnoževanja: nespolno in spolno. Oba temeljita na procesu celične delitve.
Pri nespolnem razmnoževanju se število osebkov poveča zaradi delitve. Na primer, ameba med nespolnim razmnoževanjem se s zožitvijo telesa razdeli na dve amebi. Ta proces se začne z jedrom in nato zajame citoplazmo. včasih nespolno razmnoževanje pridobi značaj večkratne delitve. V tem primeru se jedro predhodno večkrat razdeli in najpreprostejše postane večjedrno. Nato citoplazma razpade na več predelkov, ki ustrezajo številu jeder.
Kot rezultat, organizem najpreprostejših takoj povzroči precejšnje število majhni posamezniki. Tako pride na primer do nespolnega razmnoževanja malarijskega plazmodija, povzročitelja malarije pri ljudeh.
spolno razmnoževanje za praživali je značilno, da pred dejanskim razmnoževanjem (povečanjem števila osebkov) sledi spolni proces, znak ki je zlitje dveh spolnih celic (gamet) ali dveh spolnih jeder, kar povzroči nastanek ene celice - zigote, iz katere nastane nova generacija.
Oblike spolnega procesa in spolnega razmnoževanja pri praživalih so zelo raznolike.
Za razmnoževanje protozojev sta značilni dve vrsti procesov. Eden od njih - agamno ali nespolno razmnoževanje - se zmanjša na povečanje števila posameznikov določene vrste in se lahko pojavi na različne načine. Individui, ki nastanejo kot posledica takšnega razmnoževanja, imajo lahko tudi drugačno usodo. Lahko neposredno zrastejo v organizem, podoben materi, gredo skozi metamorfozo (kot potepuhi suctoria ali peritrih), se popolnoma spremenijo v gameto ali gametocit (kot gregarinski sporozoiti).
Poleg tega je spolni proces vključen v življenjski cikel mnogih protistov. Povezana je z nastankom posebnih stopenj – gamet, ki se lahko oblikujejo različne poti. Spolni proces morda sploh ne spremlja razmnoževanje (konjugacija, avtogamija, partenogeneza). Najpomembnejši trenutek spolnega procesa je prestrukturiranje jedra in drugih sestavin celice, ki poteka skozi vrsto faz in vodi do nastanka nove generacije, ki se po svoji sposobnosti preživetja in dednih lastnosti razlikuje od prejšnjih generacij. Kot smo že omenili, vse vrste spolnega procesa ne vodijo do povečanja števila osebkov, zato se glede na praživali običajno reče, da imajo na eni strani nespolno razmnoževanje in spolni proces na drugi strani.
^
Nespolno razmnoževanje
Enostavno razpolovitev ali monotomija. Ta način razmnoževanja je najpogostejši pri praživalih in celicah večceličnih tkiv. V tipičnem primeru predstavlja pravilno enotno delitev telesa najpreprostejšega na dva enaka hčerinska posameznika, pred tem pa obdobje prehranjevanja in rasti. Klasičen primer razmnoževanje ameb. Pri amebah delitev nima določene orientacije. Telo amebe se raztegne, na ekvatorju se začrta prečna zožitev, ki se postopoma poglablja in končno se celica razdeli na dve polovici, ki se takoj začneta sami premikati in hraniti. Delitev citoplazme spremlja mitotična delitev jedrca. drugi celične organele bodisi razdelijo sami ali pa se preprosto porazdelijo med hčerinske osebke, manjkajoče pa se oblikujejo na novo. Enako neorientirano delitev opazimo pri delu sončnic in radiolarij.
Prisotnost okostja seveda nekoliko oteži ta proces. Pri testnih amebah - testacey (Arcella) in gromiid (Euglypha) pred delitvijo so zametki kremenčevih lupinskih plošč položeni v posebne vakuole. Nato se začne delitev, katere ravnina je pravokotna na vzdolžno os lupine. Jedro se deli in približno polovica citoplazme, skupaj s hčerinskim jedrom in ploščami silicijevega dioksida, izstopi iz ustja navzven in je obdana z novo lupino. Pri radiolarjih, ki imajo skelet iz dveh zaklopk, je položaj cepitvene ravnine določen s položajem zaklopk. Pri delitvi se lističi razpršijo skupaj s polovico citoplazme in manjkajoči listič se dopolni.
Pri bičkovcih in migetalkah je položaj delitvene ravnine bolj konstanten in stabilen. Pri flagelatih se običajno pojavi v vzdolžni smeri, pri ciliatih - v prečni smeri. Pri flagelatih se delitev začne na sprednjem koncu telesa in nadaljuje proti zadnjemu koncu. V tem primeru se zgodi, da sta ločevalni polovici celice upognjeni v nasprotnih smereh, kar ustvarja vtis prečne delitve. Pri dinoflagelatih je delitvena ravnina odklonjena od vzdolžne - delitev poteka v poševni smeri. Tudi v poševni smeri je delitvena ravnina opalinov.
Pri kompleksno organiziranih mastigoforjih je delitev povezana z reorganizacijskimi pojavi. Hkrati se pri hčerinskih posameznikih resorbirajo in na novo oblikujejo nekateri organeli. Podobni pojavi se lahko pojavijo pri bazalnih zrncih flageluma, aksostila in polimastiginskih bičkov.
Pri migetalkah vodi proces delitve do še bolj zapletene reorganizacije kot pri bičkovcih. Tako se pri hipotrihih v sprednjem hčerinskem osebku stare adoralne membrane postopoma zmanjšujejo, hkrati pa se na njihovem dnu pojavijo zametki novih membran, ki rastejo, ko se stare resorbirajo. Podobno se zamenjajo ventralni cirusi, tako da obe hčeri prejmeta popolnoma nov sklop lokomotornih organelov.
Poseben primer reorganizacija migetalk - stomatogeneza. Stomatogeneza je morfogenetski proces, ki ga lahko opazimo med delitvijo, regeneracijo ciliatov, pa tudi pri preoblikovanju mikrostomskih oblik v makrostomske. Prva stopnja stomatogeneze je tvorba novih kinetosomov, ki tvorijo rudiment perioralnih cilij. Ta stopnja pri različne skupine ciliati potekajo na različne načine, kar služi kot osnova za filogenetske in sistematske konstrukcije.
Začetna oblika stomatogeneze se šteje za telokinetalno - tvorba začetkov ciliature, v kateri kinetosomi izvirajo iz sprednjih segmentov somatskih kinetov ali sosednjih fragmentov kinetov. Značilen je za predstavnike razreda Kinetofragminophorea.
Mnogi himenostomati, spirotrichus in peritrichus imajo bukokinetalno stomatogenezo - novi kinetosomi nastanejo poleg kinetosomov starega ustnega aparata. pri čemer bazalna telesa specializirani deli peristoma povzročajo različne komponente novih ust. Ločitev dveh novonastalih kompleksov cilij poteka skozi niz kompleksnih gibov, kot so premiki in rotacije.
Parakinetalna stomatogeneza se pojavi pri nekaterih himenostomah in spirotrihih. Pri tem se stare ustne strukture raztopijo, nastanejo nove zaradi (ali v bližini) somatskih ciliarnih vrst na ventralni strani telesa.
Če ni mogoče ugotoviti povezave med novimi kinetosomi in nekdanjimi oralnimi strukturami, govorimo o apokinetični stomatogenezi.
Takoj po nastanku bazalnih teles se že pokažejo različne sisteme mikrotubule, kot so ogorčice, postciliarne in transverzalne mikrotubule. Kinetosomi se nato preselijo v plazmalemo, kjer začnejo rasti migetalke.
Vklopljeno naslednji korak stomatogenezi se kinetosomi zvrstijo znotraj rudimenta v določen vrstni red. Ta proces poteka v določeni smeri in v strogo določenem zaporedju. Na koncu stomatogeneze se razvije aparat za zajemanje hrane - cona, iz katere se raztezajo vakuole hrane.
palintomija. Palintomija se imenuje delitev na dvoje, večkrat ponovljena, brez vmesne stopnje prehrane in rasti. Značilnosti palintomije so: 1. Predhodna hipertrofična rast materinega osebka, ki ji omogoča večkratno delitev. 2. Palintomija se zelo pogosto pojavi v obdobjih življenjskega cikla, pred katerimi je okrepljena prehrana telesa. 3. Za palintomijo je običajno značilna začasna dediferenciacija cepitvenih produktov, saj cepitveni produkti nimajo časa, da bi šli skozi proces reorganizacije. Zato se palintomija pogosto izvaja na stopnji ciste. Palintomija je razdeljena na linearno, ko so hčerinske celice razporejene v obliki verige (dinoflagelati, ciliati-apostomati) in tabularno, ki nastane kot posledica izmeničnih vzdolžnih delitev v dveh medsebojno pravokotnih ravninah, zaradi česar produkti cepitve nastanejo. so razporejeni vzporedno drug z drugim v ravni plošči. Takšna palintomija je značilna za fitomonade, nekatere dinoflagelate in evglenide. Palintomijo pogosto opazimo med tvorbo gametocitov in gamet.
brstenje. Brstenje izhaja iz neenakomerne delitve. Hkrati se morfološko razlikujeta materinski posameznik (trofon) in hčerka (potepuh, telotroh, ledvica, tomit). Obstaja monotomno brstenje, ko se oblikuje en potepuh, in večkratno brstenje. Poleg tega imajo različne skupine ciliatov notranje brstenje, zunanje brstenje in nekatere prehodne oblike brstenja. Brstenje je značilno za sesilne ciliate - suctorium, peritrich, chonotrich, rhynchodid, najdemo ga v sončnicah. značilna lastnost brstenje je odsotnost reorganizacij pri materinih posameznikih - praviloma ohranijo vse nespremenjeno, vključno z lokomotornimi ali ustnimi organeli. Med mastigoforji je brstenje izjemno redko, zlasti preprosto zunanje brstenje poznamo pri Haplozoonu in nekaterih krizomonadah. Pri sončnicah poznamo večkratno zunanje brstenje. Pri nekaterih miksosporidijah opazimo ločevanje in ločevanje delov telesa plazmodialnih stopenj.
^ Sintomija ali večkratna delitev . Pri tej vrsti razmnoževanja žival na določeni stopnji življenski krog(običajno po obdobju prehranjevanja in rasti) gre skozi vrsto ponavljajočih se jedrskih delitev in postane začasno večjedrno, nato pa takoj razpade na ustrezno število celic. Odvisno od tega, ali zaradi procesa nastanejo vegetativni posamezniki ali zarodne celice, se stopnja razmnoževanja s sintomijo imenuje shizont ali gamont, in če večkratna delitev sledi neposredno po spolnem procesu, sporont. Produkti shizogonije se imenujejo merozoiti, gamogonije - gamete in sporogonije - sporozoiti za sporozoane in zoospore za druge skupine. Sintomija je zelo razširjena med sporozoji, manj pogosta pri mastigoforjih (dinoflagelati - Noctiluca) in odsotna pri migetalkah.
Plazmatomija. Plazmatomija je vrsta nespolnega razmnoževanja, pri kateri telo večjedrne praživali razdelimo na dva oz. več večjedrne hčerinske osebe. V tem primeru pride do delitve celic v nasprotju z jedrskimi delitvami in neodvisno od njih. Plastomija vključuje delitev plazmodijev polžev, miksosporidija, ameb - pelomiksina itd. Nekateri avtorji dvomijo o dodelitvi te metode razmnoževanja posebnemu tipu.
^ Razmnoževanje kolonij. Druga vrsta nespolnega razmnoževanja. Obstajajo trije načini razmnoževanja kolonij. Prvi je, da celotno kolonijo razdelimo na dvoje. Na ta način se razmnožujejo kolonialni radiolariji in krizomonade. Drugi način je izločanje osebkov (potepuhov) iz kolonije, ki tvorijo nove kolonije. To metodo najdemo pri sedečih bičkovcih in peritrihih (Zoothamnium arbuscula). Tretja metoda je podobna drugi, vendar se tu ne oblikujejo potepuhi, ustanovni posamezniki pa ostanejo v stari koloniji in ustvarijo hčerinsko kolonijo. Značilen je za kolonialne volvocine.
Opozoriti je treba, da kolonialnost obravnavamo tudi kot rezultat nespolnega razmnoževanja, ki še ni dokončano, in ker kolonialnost velja za evolucijsko osnovo večceličnosti, lahko slednja v določeni meri izhaja tudi iz nespolnega razmnoževanja.
Glavna oblika razmnoževanja praživali je monotomija- preprosta delitev na dvoje. Med dvema delitvama žival pridobi vse strukture, izgubljene v procesu delitve, se aktivno hrani, raste in do začetka nove delitve ima čas, da doseže velikost starševskega posameznika (ameba, euglena, ceratium itd.) .
posebno obliko razmnoževanje praživali je palintomija ali prerazporeditev. Delitev se večkrat ponovi brez nadaljnjega obdobja prehranjevanja in rasti, vmesni produkti palintomije pa ostanejo nediferencirani, njihova velikost pa se z vsako delitvijo zmanjša. Samo zadnja generacija doseže polni razvoj, se hrani in raste. Palintomska delitev običajno poteka pod pokrovom zaščitnih membran (delitev oociste v malarijskem plazmodiju, zigota v volvoksu itd.).
Pri protozojih je pogosta tudi večkratna delitev - sintomija(shizogonija). V tem primeru se najprej pojavi večkratna delitev jedra brez delitve citoplazme, zaradi česar celica začasno postane polienergična, nato pa citoplazma takoj razpade na številne (glede na število jeder) hčerinske celice (tripanozome). , malarijski plazmodij, foraminifere).
Spolni proces pri protozojih
Spolni proces lahko poteka pri praživalih v obliki kopulacija in konjugacije. pri kopulacija pride do popolne fuzije dveh spolnih osebkov - gamet. Proces, ko se gamete med seboj ne razlikujejo, se imenuje izogamija. V drugih primerih nastanejo velike makrogamete (ženske) in majhne mikrogamete (moške); pride do anizogamije. Proces, ko makrogamete postanejo nepremične in se kopičijo rezerve hranila se imenuje oogamija.
Konjugacija je bolj zapletena oblika spolnega procesa in jo najdemo le pri migetalkah. V tem primeru pride le do začasne povezave dveh posameznikov in izmenjave delov mikrojeder. Prave gamete v tem primeru ne nastanejo, ampak je selitveno jedro oblečeno v tanko plast citoplazme in ustreza mikrogametu, preostali del migetalnega telesa s stacionarnim jedrom pa je makrogamet. Selitveno (moško) jedro vsakega konjugata preide v partnerjevo telo in se kopulira s svojim stacionarnim (ženskim) jedrom - pride do navzkrižne oploditve. Makronukleusi niso vključeni v ta proces.
jedrski cikli
Jedrski cikel je povezan s spolnim procesom. Vsem jedrskim ciklom je skupno menjavanje haploidnih in diploidnih generacij. Gamete so vedno haploidne; ko se združijo, dobimo diploidno zigoto; na neki stopnji življenjskega cikla pride do redukcijske delitve (mejoze), ki vrne jedra v haploidno stanje.
Obstajajo tri vrste jedrskih ciklov, od katerih je za vsakega značilno mesto v tem ciklu, ki ga zavzema redukcijska cepitev (slika 3).
1) Ciklusi z zigotično redukcijo. Večinaživljenjski cikel pade na haploidno generacijo. Diploidno generacijo predstavlja ena zigota, ki se redukcijsko deli, haploidija pa se obnovi. Takšni cikli so značilni za Phytomonadida, Sporozoa, del Polymastigida, del Hypermastlgida.
2) Cikli z gametsko redukcijo. Večina cikla pade na diploidno generacijo. Haploidno generacijo predstavljajo samo gamete, redukcijska delitev se zgodi tik pred njihovim nastankom. Takšni cikli so značilni za preostale Polymastigida in Hypermaatlgida, Heliozoa, Ciliophora, Opalinida, Cnidosporidia in skoraj vse Metazoa.
3) Cikli z vmesnim zmanjšanjem. Haploidna in diploidna generacija zavzameta približno enake dele cikla: zigota se deli mitotično. Pojavijo se bolj razvite stopnje diploidne generacije (agamont). Na neki točki pride do redukcijske delitve s tvorbo enojedrnih haploidnih agametov - agamno (nespolno) razmnoževanje. Agamete se razvijejo v naprednejše stopnje haploidne generacije. Nato ta odrasla haploidna generacija (gamont) razpade na gamete, ki se kopulirajo - pride do spolnega procesa. Z nastankom zigote se cikel sklene. Tak cikel se pojavi le pri foraminiferah in kot redka izjema pri nekaterih kolobarjih.
Življenjski cikli
Življenjski cikli praživali so zelo raznoliki. Obstajajo praživali, ki imajo cikel brez spolnega procesa, tj. brez jedrnega cikla (npr. pri tripanozah, pri katerih se izmenjujejo različne vegetativne stopnje).
V najbolj diferenciranem življenjskem ciklu ločimo štiri obdobja:
1) progamno, ki se konča s tvorbo gameta;
2) singamni, vključno s kopulacijo gamet in tvorbo zigote;
3) metagamni, ki zajema več generacij sporozoitov;
4) vegetativno, med katero pride do rasti, povečanje mase posameznikov.
V določenem delu življenjskega cikla pride do zmanjšanja števila kromosomov.
Za udobje primerjave življenjskih ciklov bomo sestavili tabelo, ki jo bomo navpično razdelili na 4 dele, ki ustrezajo navedenim štirim obdobjem. V to tabelo smo postavili več ciklov najpreprostejših in Cikel večcelični organizem, pri čemer so obdobja njihovih ciklov postavljena v ustrezne stolpce (slika 4). Izberemo najpreprostejše z najbolj preprostimi in razčlenjenimi cikli.
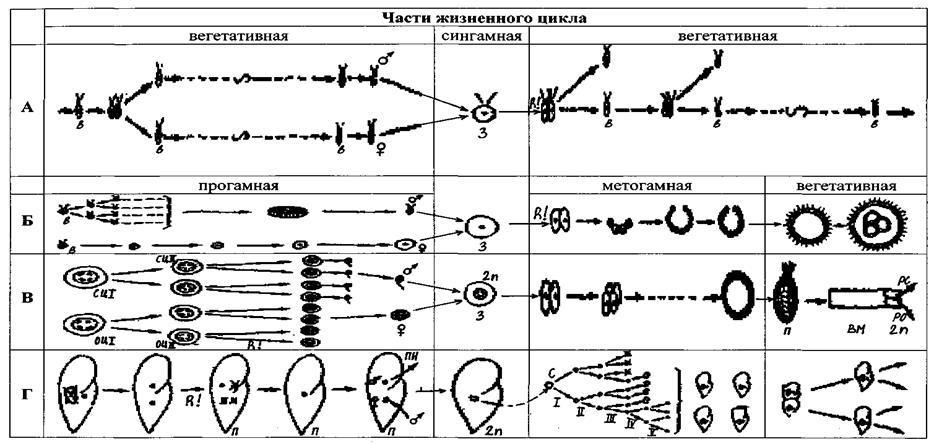
Slika 4. Primerjava življenjskih ciklov različnih protozojev in večceličnih organizmov (iz K.V. Beklemisheva, 1979):
A – neomejeno večpovezan cikel nižjih fitomonad s fakultativnim spolnim procesom; B - Volvoxov cikel z delitvijo cikla na vegetativni, progamni in metagamni del; C - cikel večceličnih organizmov s povečanimi razlikami med deli cikla; D - cikel ciliatov (v metagamnem delu cikla je prikazana samo jedrna delitev, celične konture niso prikazane). 3 - zigota, C - sinkarion, R! - redukcijska delitev, PN - pronukleus, P - parenhimula, rs - razmnoževanje in rast sporogonija, rho - razmnoževanje in rast oogonia, VM - odrasli mnogocelični, sI in ssP - spermatociti I in II reda, otsI in otsP - oociti I. in II
Od vse raznolikosti življenjskih ciklov protozojev s spolnim procesom imajo monoenergetski flagelati, na primer Chlamydomonas, najpreprostejši cikel. Pri nekaterih vrstah tega rodu je veliko nespolnih generacij, ki nastanejo z monotomijo, spolni proces se pojavi, ko se pogoji obstoja poslabšajo. Gamete se morfološko malo razlikujejo od posameznikov nespolnih generacij. Po kopulaciji se zigota obleče v lupino, pod pokrovom katere s palintomično delitvijo nastane 4–32 osebkov, ki izstopijo iz lupine in se začnejo nespolno razmnoževati. Zmanjšanje števila kromosomov se pojavi med prvimi delitvami zigote. V tem ciklu so vse stopnje haploidne, edina diploidna stopnja je zigota. To je zigotski redukcijski cikel. Pri drugih vrstah rodu Chlamydomonas pride do nespolnega razmnoževanja s palintomijo, gamete nastanejo tudi s palintomijo.
Najbolj razčlenjen življenjski cikel je pri »večceličnem« praživali – Volvoxu. V tečaju zoologije se temu predmetu posveča malo pozornosti, ker. ima avtotrofni tip prehrane. Vendar Volvox vzame pomembno mesto v primerjalni anatomiji nevretenčarjev, saj je njegov življenjski cikel najbližji ciklu večceličnih organizmov. Za življenjski cikel Volvoxa je značilno strogo določeno mesto spolnega procesa, pred katerim je progamni del cikla, med katerim pride do homogonije, tj. preoblikovanje posebnih generativnih celic v gamete: makrogamont raste, kopiči hranila v sebi in se spremeni v veliko nepremično makrogameto, mikrogamont se palintomsko deli, pri čemer nastane veliko število mikrogamete. Sledi zlitje gamet (singamični del cikla) in redukcijska delitev. Naslednji - metagamni del - zigota, se deli palintomsko, kar vodi v nastanek blastule podobne kolonije, ki zelo spominja na stopnjo blastule večceličnih organizmov; ta kolonija raste, v njej se oblikujejo nove hčerinske kolonije - po metagamu se pojavi vegetativni del cikla. V življenjskem ciklu Volvoxa je treba biti pozoren na dejstvo, da se razvoj hčerinske kolonije iz zigote najprej pojavi v stanju inkarvacije (tj. celice, ki nastanejo zaradi palintomske delitve zigote, se spremenijo z bičkovimi konci v votla krogla), šele pred izhodom nastale hčerinske kolonije pride do ekskurvacije od matere - everzije, zaradi česar celice prevzamejo končno razporeditev z bički navzven. Redukcijska delitev pri Volvoxu, tako kot pri sporozojih, se pojavi po nastanku zigote - cikel z zigotsko redukcijo. Cikel ciliatov je zelo svojevrsten. Učbeniki zoologije opisujejo cikel ciliatov brez upoštevanja jedrskega cikla. Zato je treba biti pozoren na dejstvo, da v programskem delu sodeluje le mikronukleus, ki je bil podvržen redukcijski delitvi, od štirih nastalih haploidnih jeder se tri reducirajo in ostane eno, ki se deli s tvorbo dveh pronukleusov. Ženski pronukleus se spremeni v stacionarno jedro, ostane v ciliatu in tako vse ustreza makrogametu, moški pronukleus pa se spremeni v selitveno jedro, ki ustreza mikrogametu, in preide v partnerja, ki se združi s svojim stacionarnim jedrom ( singamski del cikla). V metagamnem delu cikla ciliatov palintomije ni opaziti; pride do delitve sinkariona, ki po vrsti preureditev pripelje do ponovne vzpostavitve binuklearnosti vsakega posameznika in do ponovne vzpostavitve poliploidije makronukleusa. V vegetativnem delu cikla migetalke rastejo in se množijo. aseksualno. To je cikel z gametsko redukcijo.
V življenjskem ciklu katerega koli večceličnega organizma, tako kot pri Volvoxu, obstajajo progamni, singamni, metagamni in vegetativni deli. Toda programski del postane bolj zapleten zaradi dejstva, da so večcelični organizmi v tem obdobju podvrženi kompleksnemu prestrukturiranju jedrskega aparata: nastanek tetraploidnih oo- in spermatocitov prvega reda, od tega diploidnih oo- in spermatocitov drugega reda. in končno redukcijska delitev, ki povzroči nastanek haploidnih gamet.
Metagamni palintomski del cikla metazojev je zelo podoben delu fitomonad. Vendar pa je vegetativni del cikla drugačen: označen je visoko stopnjo integracijo telesa in veliko različnih celic.
Iz tabele je razvidno, da je od obravnavanih praživali Volvoxov cikel najbližji življenjskemu ciklu večceličnih organizmov. Njen metagamni del se konča s tvorbo sferične blastule podobne stopnje; ima tako kot Metazoa vegetativni del, med katerim organizem (ali kolonija) raste, se njegove celice diferencirajo in le majhen del se jih specializira za generativne. . Glavna razlika med življenjskim ciklom Volvoxa in večceličnim ciklom je kraj redukcijska delitev. Tako podrobna primerjava ciklov različnih praživali in metazojev je bistvena za razumevanje izvora metazojev. Vendar je treba jasno razumeti, da Volvoxa nikakor ne bi smeli obravnavati kot neposrednega prednika večceličnih organizmov. Ta predmet se uporablja le kot model, ki je primeren za razumevanje procesov, ki bi se lahko pojavili v evoluciji kolonialnih heterotrofnih flagellatov, najverjetneje blizu sodobnih Choanoflagellata, ki trenutno veljajo za prednike metazojev. Med sodobnimi bičkovarji z ovratnikom ni vrst s spolnim postopkom, vendar se domneva, da so bili prav takšni kolonialni Choanoflagellata, ki so imeli obliko krogle in življenjski cikel tipa Volvox, vendar z gametsko redukcijo, predniki večcelični organizmi.
Aseksualno - z mitozo jedra in delitvijo celice na dvoje (pri amebi, eugleni, ciliatih), pa tudi z shizogonija- večkratna delitev (pri sporozojih).
spolno - kopulacija. Celica protozoja postane funkcionalna gameta; Kot rezultat zlitja gamet nastane zigota.
Za ciliate je značilen spolni proces - konjugacija. Gre za to, da si celice izmenjujejo genetske informacije, vendar ne pride do povečanja števila osebkov.
Mnoge praživali lahko obstajajo v dveh oblikah - trofozoit(vegetativna oblika, ki je sposobna aktivna prehrana in gibanje) in ciste, ki nastane v neugodnih razmerah. Celica je imobilizirana, dehidrirana, prekrita gosta lupina metabolizem se močno upočasni. V tej obliki živali, veter zlahka prenašajo praživali na velike razdalje in jih razpršijo. Ob udarcu noter ugodni pogoji habitat, pride do ekscistacije, celica začne delovati v stanju trofozoita. Tako encistacija ni način razmnoževanja, ampak pomaga celici preživeti neugodne razmere okolju.
Za številne predstavnike vrste Praživali značilna prisotnost življenski krog, ki je sestavljen iz rednega menjavanja življenjske oblike. Praviloma pride do menjave generacij z nespolnim in spolnim razmnoževanjem. Tvorba ciste ni del običajnega življenjskega cikla.
Čas generacije za praživali je 6-24 ur, kar pomeni, da se celice, ko so v telesu gostitelja, začnejo eksponentno razmnoževati in teoretično lahko povzročijo njegovo smrt. Vendar se to ne zgodi, saj začnejo veljati zaščitni mehanizmi gostiteljskega organizma.
Medicinsko pomembni so predstavniki praživali, ki spadajo v razrede sarkodov, flagelatov, ciliatov in sporozojev.
Tip Protozoa vključuje organizme, katerih telo je sestavljeno iz ene same celice, ki pa deluje kot celega organizma. Protozojske celice so sposobne samooskrba, gibanje, zaščita pred sovražniki in doživljanje neugodnih razmer.
Prehrana protozojev poteka s pomočjo prebavnih vakuol, ki jih izvaja fag - ali pinocitoza. Ostanki neprebavljena hrana izvržen. Nekatere praživali vsebujejo kloroplaste in se lahko hranijo s fotosintezo.
Večina praživali ima organele za gibanje: bičke, migetalke in psevdopodije (začasne mobilne izrastke citoplazme). Oblike organelov gibanja so osnova taksonomije praživali.
Običajno se izvaja razmnoževanje protozojev različne oblike delitve so različice mitoze. Značilen je tudi spolni proces: v obliki zlitja celic - kopulacije - ali izmenjave dela dednega materiala - konjugacije.
Večina praživali ima eno jedro, obstajajo pa tudi večjedrne oblike.
Medicinsko pomembne so praživali, ki spadajo v razrede Sarcodaceae, Flagellates, Ciliates in Spores.