Pregunta 1. ¿Cuáles son las diferencias en la estructura de las células eucariotas y procarióticas?
Los procariotas no tienen un núcleo realmente formado (del griego karyon, núcleo). Su ADN es una única molécula circular, ubicada libremente en el citoplasma y no rodeada por una membrana. Ud. células procariotas no hay plastidios, mitocondrias, retículo endoplásmico, aparato de Golgi ni lisosomas. Tanto los procariotas como los eucariotas tienen ribosomas (los nucleares los tienen más grandes). El flagelo de una célula procariota es más delgado y funciona según un principio diferente al del flagelo de los eucariotas. Los organismos eucariotas son hongos, plantas y animales, unicelulares y multicelulares; procariotas: bacterias y algas verdiazules (cianobacterias).
Pregunta 2. Cuéntanos sobre el pinot y la fagocitosis. ¿En qué se diferencian estos procesos?
La membrana celular es una formación móvil capaz de capturar objetos del entorno externo mediante la formación de invaginaciones y excrecencias. Este proceso se llama endocitosis. La causa de la endocitosis son reacciones bioquímicas complejas que ocurren en el citoplasma y se asocian principalmente con cambios en la estructura terciaria de las proteínas intracelulares. Si una célula capta una gota de líquido, se trata de pinocitosis; si es una partícula sólida, se trata de fagocitosis. Como resultado, se forman vacuolas pinocíticas o fagocíticas (vesículas de membrana). El proceso inverso de la endocitosis (la liberación del contenido de la vacuola de la célula) se llama exocitosis.
Pregunta 3. Explique la relación entre la estructura y funciones de la membrana celular.
Se sabe que la base de cualquier membrana es una bicapa (doble capa) de fosfolípidos, en la que las "cabezas" hidrofílicas de las moléculas (glicerol) miran hacia afuera y los residuos hidrofóbicos ácidos grasos- adentro. Asociadas a la bicapa lipídica hay moléculas de proteínas que pueden estar adyacentes a la membrana en ambos lados, sumergirse en ella o incluso penetrarla. La posición de la membrana celular en el límite celular y ambiente define sus principales funciones. Una bicapa fuerte, elástica y de fácil restauración es una barrera que asegura la constancia del entorno intracelular y protege el citoplasma de la penetración de sustancias extrañas. La función de transporte de la membrana es selectiva. Pequeñas moléculas sin carga (O2, N2) penetran fácilmente directamente a través de la bicapa. Las partículas más grandes y/o cargadas (Na+, K+, algunas hormonas) pasan a través de poros (canales) proteicos especiales o son transportadas por proteínas transportadoras. Al ser una estructura móvil, la membrana celular también puede transportar sustancias mediante endo y exocitosis.
Pregunta 4. ¿Qué orgánulos celulares se encuentran en el citoplasma?
Los orgánulos ubicados en el citoplasma de una célula eucariota se pueden dividir en tres grupos: de membrana única, de doble membrana y sin membrana. Los orgánulos de membrana única incluyen el retículo endoplásmico (liso y rugoso), el aparato de Golgi, los lisosomas y las vacuolas. Dos organelos de membrana- estos son plastidios y mitocondrias; no membrana: ribosomas, citoesqueleto y centro celular.
Pregunta 5. Describe los orgánulos del citoplasma y su importancia en la vida de la célula.
El retículo endoplasmático (RE) es un conjunto de vacuolas, canales y túbulos. Forma una única red dentro del citoplasma, combinada con la membrana externa. membrana nuclear. Hay EPS lisos y rugosos. El EPS suave participa en la síntesis de lípidos y carbohidratos, y también neutraliza sustancias toxicas. Los ribosomas se encuentran en la superficie de las membranas rugosas del RE.
El aparato de Golgi es un orgánulo de membrana única que forma parte de una red de membrana única de la célula y es una pila de cisternas planas. Es donde tiene lugar la clasificación final y el envasado de los productos de desecho celular en vesículas de membrana (vacuolas). Entre otras cosas, el aparato de Golgi forma lisosomas y proporciona exocitosis.
Los lisosomas son pequeñas vesículas de membrana que contienen enzimas para la digestión. nutrientes. Los lisosomas se fusionan con la vacuola endocítica para formar una vacuola digestiva. Si el contenido de los lisosomas se libera dentro de la propia célula, se produce la autólisis (autodigestión de la célula).
Las mitocondrias se clasifican como orgánulos de doble membrana. Su membrana exterior es lisa y la interior forma pliegues (crestas). Las mitocondrias son las estaciones de energía de la célula, su función principal es la síntesis de ATP.
Los plástidos son orgánulos de células vegetales de doble membrana. Hay tres tipos de plastidios: cloroplastos, cromoplastos y leucoplastos. Los leucoplastos incoloros almacenan almidón; los cloroplastos verdes realizan la fotosíntesis; Los cromoplastos anaranjados, amarillos y rojos proporcionan color a frutos y flores (atrayendo a polinizadores y dispersores de semillas). Se considera establecido que en el pasado lejano las mitocondrias y los plastidios se originaban a partir de procariotas que eran “tragados” por una célula eucariota y entraban en simbiosis con ella. Las mitocondrias y los plastidios tienen ADN circular, sintetizan de forma independiente algunas proteínas y sus ribosomas son más pequeños que los eucariotas.
Los ribosomas son pequeños y numerosos orgánulos sin membrana formados por dos subunidades: una grande y una pequeña. Las subunidades están formadas por proteínas y ARN ribosómico. La función de los ribosomas es la síntesis de proteínas. Algunos de los ribosomas están ubicados directamente en el citoplasma y otros en las membranas del RE rugoso.
Centro celular: un orgánulo de estructura no membranosa en animales, hongos y plantas inferiores. Consta de dos centriolos, de forma similar a cilindros y formados por diminutos tubos de proteínas; Participa en la formación del huso de fisión.
Una vacuola es una vesícula de membrana llena de savia celular. Está necesariamente presente en la célula vegetal. La función de la vacuola es la acumulación de agua, sales y nutrientes. También puede contener pigmentos (azul, violeta) y acumular desechos metabólicos.
El citoesqueleto es un orgánulo sin membrana, que consta de túbulos y cadenas de proteínas ubicados junto a la membrana y en el citoplasma. Su función es mantener la forma de la célula, asegurar el transporte intracelular de sustancias, así como el movimiento activo de la célula (ameba, fagocito). Las vesículas de membrana que contienen cualquier sustancia pueden moverse a lo largo de los tubos citoesqueléticos, como sobre rieles.
¿Cómo descargar un ensayo gratis? . Y un enlace a este ensayo; Célula eucariota. Citoplasma. organoides ya en tus marcadores.Ensayos adicionales sobre este tema.
Pregunta 1. ¿Dónde se forma el lisosoma? Los lisosomas son estructuras de membrana que contienen muchas enzimas activas involucradas en la descomposición de compuestos de alto peso molecular: proteínas, lípidos, carbohidratos. Los lisosomas se forman en el complejo de Golgi, donde entran y se acumulan las enzimas. Pregunta 2: ¿Cuál es la función de las mitocondrias? mitocondrias - estructuras celulares, cubierto con una doble membrana. En la membrana interna, que tiene numerosas proyecciones, hay gran cantidad enzimas implicadas en la síntesis de ATP. Por eso, función principal mitocondrias: proporcionan energía a la célula mediante la síntesis de ATP. Pregunta 3. ¿Qué
Pregunta 1. ¿Qué signos del carácter primitivo de los procariotas en comparación con los eucariotas puedes nombrar? Los procariotas carecen de núcleo y cromosomas. La información hereditaria de los procariotas está representada por una única molécula de ADN, que está cerrada en forma de anillo y ubicada en el citoplasma. Los ribosomas en los procariotas son muy pequeños. Los procariotas no tienen orgánulos rodeados de membranas (retículo endoplásmico, aparato de Golgi, mitocondrias, plastidios). En una célula eucariota, estas estructuras están diseñadas para sintetizar una variedad de compuestos. Debido a su ausencia en procariotas, las reacciones
Pregunta 1. ¿De qué están formadas las paredes del retículo endoplásmico y el complejo de Golgi? Las paredes del retículo endoplasmático y del complejo de Golgi están formadas por una membrana monocapa. Pregunta 2. Nombra las funciones del retículo endoplásmico. El retículo endoplásmico (RE) forma el sistema de transporte de la célula. La síntesis de grasas y carbohidratos tiene lugar en el RE liso. En el RE rugoso (granular), la síntesis de proteínas se produce debido al trabajo de los ribosomas adheridos a las membranas del RE. Pregunta 3. ¿Qué función realizan los ribosomas? La función principal de los ribosomas es la síntesis de proteínas. Pregunta 4. ¿Por qué la mayoría de los ribosomas están ubicados en canales endoplasmáticos?
Prueba sobre el tema “Líquenes. Algas" 1. La fotosíntesis en las algas ocurre en: a) núcleo c) citoplasma b) cromatóforos d) cloroplastos 2. Cómo liquen un solo organismo método inherente de reproducción: a) sexual c) partenogénesis b) asexual d) no hay respuesta correcta 3. Los líquenes se denominan indicadores de pureza del aire, ya que: a) purifican el aire b) contaminan el aire c) crean un hábitat para otras plantas y animales d) crecen sólo en lugares con aire limpio 4. ¿Son capaces de movimiento activo¿algas marinas? a) todas las algas son capaces de moverse b) moverse
Pregunta 1. ¿Cuáles son las funciones? membrana externa¿células? La membrana celular externa consta de una doble capa lipídica y moléculas de proteínas, algunas de las cuales se encuentran en la superficie y otras penetran a través de ambas capas de lípidos. La membrana celular externa realiza una función protectora, separando la célula del ambiente externo y evitando daños a su contenido. Además, la membrana celular externa asegura el transporte de sustancias dentro y fuera de la célula y permite que las células interactúen entre sí. Pregunta 2. ¿De qué manera pueden penetrar diversas sustancias en la célula? Por el exterior
Pregunta 1. ¿Cuáles son las principales etapas del surgimiento y desarrollo de la vida en la Tierra? Existen muchas hipótesis que intentan explicar el surgimiento y desarrollo de la vida en nuestro planeta. Y aunque ofrecen diferentes enfoques para resolver este problema, la mayoría asume la presencia de tres etapas evolutivas: evolución química, prebiológica y biológica. En la etapa de evolución química tuvo lugar la síntesis abiogénica de polímeros orgánicos. En la segunda etapa, se formaron complejos proteína-ácido nucleico-lipoide (los científicos los llamaron de otra manera: coacervados, hiperciclos, probiones, progenotes y
Pregunta 1. ¿Cuáles son las funciones del centro celular? El centro celular cumple la función de formar el esqueleto interno de la célula (citoesqueleto). El citoesqueleto es una red de microtúbulos que penetran en el citoplasma, manteniendo la forma de la célula, asegurando el movimiento de los orgánulos celulares, así como el trabajo de orgánulos de movimiento especializados: cilios y flagelos. El centro celular también asegura la división celular normal. Los centríolos del centro celular divergen hacia los polos de la célula en división y forman un huso de división, por lo que posteriormente se forman dos células hijas a partir de una célula madre. Los centriolos están representados por cilindros formados
Citoplasma- una parte obligatoria de la célula, encerrada entre la membrana plasmática y el núcleo; se divide en hialoplasma (la sustancia principal del citoplasma), orgánulos (componentes permanentes del citoplasma) e inclusiones (componentes temporales del citoplasma). Composición química del citoplasma: la base es el agua (60-90% de la masa total del citoplasma), diversos compuestos orgánicos e inorgánicos. El citoplasma tiene una reacción alcalina. Un rasgo característico del citoplasma de una célula eucariota es el movimiento constante ( ciclosis). Se detecta principalmente por el movimiento de orgánulos celulares, como los cloroplastos. Si el movimiento del citoplasma se detiene, la célula muere, ya que solo estando en movimiento constante, puede realizar sus funciones.
hialoplasma ( citosol) es una solución coloidal incolora, viscosa, espesa y transparente. Es en él donde tienen lugar todos los procesos metabólicos, asegura la interconexión del núcleo y todos los orgánulos. Dependiendo del predominio de la parte líquida o de moléculas grandes en el hialoplasma, se distinguen dos formas de hialoplasma: Sol- hialoplasma más líquido y gel- hialoplasma más espeso. Son posibles transiciones mutuas entre ellos: el gel se convierte en sol y viceversa.
Funciones del citoplasma:
- combinar todos los componentes de la celda en un solo sistema,
- ambiente para el paso de muchos procesos bioquímicos y fisiológicos,
- Ambiente para la existencia y funcionamiento de los orgánulos.
Membranas celulares
Membranas celulares limitar las células eucariotas. En cada membrana celular se pueden distinguir al menos dos capas. La capa interna está adyacente al citoplasma y está representada por membrana de plasma(sinónimos: plasmalema, membrana celular, membrana citoplasmática), sobre la cual se forma la capa externa. EN célula animal es delgado y se llama glicocalix(formado por glicoproteínas, glicolípidos, lipoproteínas), en una célula vegetal - espesa, llamada pared celular(formado por celulosa).
Todo membranas biológicas tienen características y propiedades estructurales comunes. Actualmente es generalmente aceptado modelo de mosaico fluido de la estructura de la membrana. La base de la membrana es una bicapa lipídica formada principalmente por fosfolípidos. Los fosfolípidos son triglicéridos en los que un residuo de ácido graso se reemplaza por un residuo de ácido fosfórico; la sección de la molécula que contiene el residuo de ácido fosfórico se llama cabeza hidrofílica, las secciones que contienen residuos de ácido graso se llaman colas hidrofóbicas. En la membrana, los fosfolípidos están dispuestos de forma estrictamente ordenada: las colas hidrófobas de las moléculas se enfrentan entre sí y las cabezas hidrófilas miran hacia afuera, hacia el agua.
Además de los lípidos, la membrana contiene proteínas (en promedio ≈ 60%). Determinan la mayoría de funciones específicas de la membrana (transporte de determinadas moléculas, catálisis de reacciones, recepción y conversión de señales del medio ambiente, etc.). Hay: 1) proteínas periféricas(ubicado en la superficie externa o interna de la bicapa lipídica), 2) proteínas semiintegrales(sumergido en la bicapa lipídica a diferentes profundidades), 3) Proteínas integrales o transmembrana.(perforar la membrana, contactando tanto con el entorno externo como interno de la célula). Las proteínas integrales se denominan en algunos casos proteínas formadoras de canales o proteínas de canal, ya que pueden considerarse canales hidrófilos a través de los cuales pasan moléculas polares al interior de la célula (el componente lipídico de la membrana no las dejaría pasar).
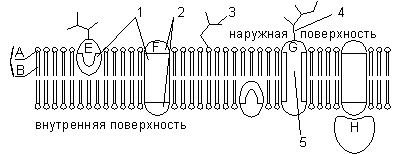
A - cabeza de fosfolípidos hidrófilos; B - colas de fosfolípidos hidrófobos; 1 - regiones hidrofóbicas de las proteínas E y F; 2 — regiones hidrofílicas de la proteína F; 3 - cadena de oligosacárido ramificada unida a un lípido en una molécula de glicolípido (los glicolípidos son menos comunes que las glicoproteínas); 4 - cadena de oligosacárido ramificada unida a una proteína en una molécula de glicoproteína; 5 - canal hidrófilo (funciona como un poro a través del cual pueden pasar iones y algunas moléculas polares).
La membrana puede contener carbohidratos (hasta un 10%). El componente carbohidrato de las membranas está representado por cadenas de oligosacáridos o polisacáridos asociados con moléculas de proteínas (glicoproteínas) o lípidos (glicolípidos). Los carbohidratos se encuentran principalmente en la superficie exterior de la membrana. Los carbohidratos proporcionan funciones receptoras de la membrana. En las células animales, las glicoproteínas forman un complejo supramembrana, el glicocálix, que tiene un grosor de varias decenas de nanómetros. Contiene muchos receptores celulares y con su ayuda se produce la adhesión celular.
Las moléculas de proteínas, carbohidratos y lípidos son móviles y capaces de moverse en el plano de la membrana. El espesor de la membrana plasmática es de aproximadamente 7,5 nm.
Funciones de las membranas
Las membranas realizan las siguientes funciones:
- separación del contenido celular del entorno externo,
- regulación del metabolismo entre la célula y el medio ambiente,
- dividir la celda en compartimentos ("compartimentos"),
- lugar de localización de “transportadores enzimáticos”,
- Asegurar la comunicación entre las células en los tejidos. organismos multicelulares(adhesión),
- reconocimiento de señales.
El más importante propiedad de la membrana— permeabilidad selectiva, es decir Las membranas son muy permeables a algunas sustancias o moléculas y poco permeables (o completamente impermeables) a otras. Esta propiedad subyace a la función reguladora de las membranas, asegurando el intercambio de sustancias entre la célula y el entorno externo. El proceso por el que las sustancias atraviesan la membrana celular se llama transporte de sustancias. Hay: 1) transporte pasivo- el proceso de paso de sustancias sin consumo de energía; 2) transporte activo- el proceso de paso de sustancias que se produce con el gasto de energía.
En transporte pasivo Las sustancias se mueven desde un área con más alta concentración a un área con un más bajo, es decir. a lo largo del gradiente de concentración. En cualquier solución hay moléculas de solvente y soluto. El proceso de mover las moléculas del soluto se llama difusión y el movimiento de las moléculas del disolvente se llama ósmosis. Si la molécula está cargada, su transporte también se ve afectado por el gradiente eléctrico. Por lo tanto, la gente suele hablar de un gradiente electroquímico, que combina ambos gradientes. La velocidad del transporte depende de la magnitud del gradiente.
Puedes elegir los siguientes tipos transporte pasivo: 1) difusión simple— transporte de sustancias directamente a través de la bicapa lipídica (oxígeno, dióxido de carbono); 2) difusión a través de canales de membrana— transporte a través de proteínas formadoras de canales (Na +, K +, Ca 2+, Cl -); 3) difusión facilitada- transporte de sustancias mediante proteínas de transporte especiales, cada una de las cuales es responsable del movimiento de determinadas moléculas o grupos de moléculas relacionadas (glucosa, aminoácidos, nucleótidos); 4) ósmosis— transporte de moléculas de agua (en todos sistemas biológicos El disolvente es agua.)
Necesidad transporte activo Ocurre cuando es necesario asegurar el transporte de moléculas a través de una membrana contra un gradiente electroquímico. Este transporte se realiza mediante proteínas portadoras especiales, cuya actividad requiere un gasto energético. La fuente de energía son las moléculas de ATP. El transporte activo incluye: 1) bomba Na + /K + (bomba de sodio-potasio), 2) endocitosis, 3) exocitosis.
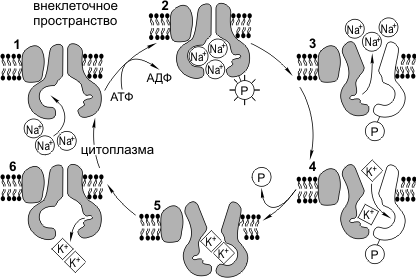
Funcionamiento de la bomba Na+/K+. Para funcionamiento normal la célula debe mantener una cierta proporción de iones K + y Na + en el citoplasma y en el ambiente externo. La concentración de K + dentro de la célula debe ser significativamente mayor que fuera de ella, y Na +, viceversa. Cabe señalar que Na+ y K+ pueden difundir libremente a través de los poros de la membrana. La bomba Na+/K+ contrarresta la igualación de las concentraciones de estos iones y bombea activamente Na+ fuera de la célula y K+ hacia el interior de la célula. La bomba Na+/K+ es una proteína transmembrana capaz de realizar cambios conformacionales, como resultado de los cuales puede unir tanto K+ como Na+. El ciclo de funcionamiento de la bomba Na+ /K+ se puede dividir en las siguientes fases: 1) conexión de Na+ con adentro membranas, 2) fosforilación de la proteína bomba, 3) liberación de Na + en el espacio extracelular, 4) adición de K + a afuera membranas, 5) desfosforilación de la proteína bomba, 6) liberación de K + en el espacio intracelular. Casi un tercio de toda la energía necesaria para el funcionamiento de las células se gasta en el funcionamiento de la bomba de sodio y potasio. En un ciclo de operación, la bomba bombea 3Na + de la celda y bombea 2K +.
Endocitosis- el proceso de absorción de partículas grandes y macromoléculas por parte de la célula. Hay dos tipos de endocitosis: 1) fagocitosis- captura y absorción de partículas grandes (células, partes de células, macromoléculas) y 2) pinocitosis— captura y absorción de material líquido (solución, solución coloidal, suspensión). El fenómeno de la fagocitosis fue descubierto por I.I. Mechnikov en 1882. Durante la endocitosis, la membrana plasmática forma una invaginación, sus bordes se fusionan y las estructuras delimitadas del citoplasma por una sola membrana se unen al citoplasma. Muchos protozoos y algunos leucocitos son capaces de fagocitosis. La pinocitosis se observa en las células epiteliales intestinales y en el endotelio de los capilares sanguíneos.
Exocitosis- proceso inverso a la endocitosis: excreción varias sustancias desde la celda. Durante la exocitosis, la membrana de la vesícula se fusiona con la membrana citoplasmática externa, el contenido de la vesícula se elimina fuera de la célula y su membrana se incluye en la membrana citoplasmática externa. De esta manera a partir de células glandulares. secreción interna las hormonas se excretan, en los más simples - restos no digeridos alimento.
Ir a conferencias número 5"Teoría celular. Tipos de organización celular"
Ir a conferencias número 7“Célula eucariota: estructura y funciones de los orgánulos”
Contenido del resumen
2.
Citoplasma de una célula eucariota.
4. Membranas citoplasmáticas
4.1. Química y propiedades membranas celulares
5. Membrana plasmática
5.1. Crecimiento de la membrana plasmática
6.1. Funciones de transporte
6.2. Funciones receptoras de la membrana plasmática.
6.3. contactos intercelulares
6.4. Contacto sencillo. Conexión tipo bloqueo
6.6. contacto intermedio
6.7. Contacto de ranura
Bibliografía
1. El papel de los científicos en el descubrimiento de las membranas celulares
Hasta finales del siglo XVIII se suponía que las celdas estaban separadas por una pared común y no podían aislarse unas de otras. En el desarrollo posterior de esta ciencia, la investigación del botánico francés Brissot de Mirbel (1776-1854) jugó un papel importante, en la que se expresó por primera vez la idea de un estudio comparativo de los tejidos vegetales. Mirbel fue el primero en sugerir que las células adyacentes están separadas por una partición común: una membrana.
El naturalista alemán Rudolphi llegó a la conclusión de que las células están aisladas, que tienen sus propias membranas que las rodean por todos lados. Cualquier duda de que las células puedan aislarse y aislarse finalmente desapareció después del trabajo del botánico I. Moldenhauer (1766-1827), de donde se dedujo que la célula es una estructura independiente cubierta por una membrana. Entonces, se formó una nueva idea de la célula.
Fue formulado con mayor claridad un poco más tarde por F. Meyen. Su definición es la siguiente: “La célula de un organismo vegetal es un espacio completamente encerrado por una membrana vegetativa”. F. Meyen expresó claramente esta opinión a principios del siglo XIX. reemplazando la vista original de la celda como una época. Pero el contenido específico de la anatomía vegetal todavía no se diferenciaba fundamentalmente del anterior. Los investigadores continuaron describiendo sistemáticamente la estructura. varios órganos plantas, así como detallar y aclarar lo estudiado. Este período se considera, con razón, un período de recolección de material, un período de acumulación de abundante información sobre la estructura más sutil de las plantas.
Entonces, el hecho mismo de la presencia de células en varias telas no había duda. Pero, ¿toda la diversidad de estructuras de los organismos vegetales se limita a las células? La cuestión todavía no estaba del todo clara. Además, la mayoría de los científicos respondieron negativamente a esta pregunta, señalando, junto con las células, los vasos y las fibras de las plantas como estructuras bastante únicas. Resumiendo todo el conocimiento en esta área, Meyen escribió directamente que los tejidos vegetales están formados por células, fibras y vasos. La aparición de estas últimas células en metamorfosis se confirmó finalmente sólo un poco más tarde, en los años 30. el siglo pasado.
2. Citoplasma de una célula eucariota.
Citoplasma Representa el aparato de trabajo metabólico de la célula. En él se concentran orgánulos generales y especiales y los principales procesos metabólicos tienen lugar en el citoplasma. En las últimas décadas se han logrado éxitos especialmente grandes en el análisis morfofuncional discreto de orgánulos individuales y sistemas de membranas del aparato metabólico del citoplasma. Esto permite imaginar el trabajo del aparato metabólico como un sistema integral.
En los estudios modernos del aparato metabólico de la célula, una tendencia particularmente pronunciada es estudiar el significado funcional de las estructuras morfológicas, analizar la función a través de la organización estructural y bioquímica de estas estructuras. Los avances más importantes en este sentido se han logrado en el estudio de los principales orgánulos de síntesis de proteínas (ribosomas) y de los sistemas que proporcionan el metabolismo energético (mitocondrias, cloroplastos y membranas conjugadoras de las células procarióticas). Una generalización importante en el estudio del aparato metabólico del citoplasma es la idea de él como un sistema trifásico diferenciado plástico, que consta del citoplasma principal, las membranas intracelulares y el contenido del sistema de membranas. Así, el aparato metabólico del citoplasma, por un lado, está compuesto por estructuras especializadas que realizan funciones particulares y, por otro lado, es un sistema integral único que está en estrecha relación tanto con el aparato superficial como nuclear del celúla.
Este enfoque comenzó allá por los años 60. De Duve en la doctrina de los sistemas de membranas exoplásmicas. Este enfoque se desarrolló posteriormente tanto en el estudio de los lisosomas como en ideas modernas sobre el aparato de Golgi y el retículo endoplásmico, así como en ideas sobre la interacción dinámica de los sistemas de membranas intracelulares. En nuestra opinión, el concepto recientemente desarrollado del citoplasma principal, el hialoplasma, también es muy prometedor. A diferencia de la visión predominante hasta ahora del hialoplasma como un sistema coloidal vivo y homogéneo, ésta considera el citoplasma principal como una fase diferenciada y heterogénea del citoplasma, capaz de formar estructuras complejas. Ya nos hemos familiarizado parcialmente con esta propiedad del hialoplasma utilizando el ejemplo del sistema submembrana del aparato de superficie con sus estructuras dinámicas mecanoquímicas y de soporte.
Ahora está quedando claro que tales sistemas hialoplásmicos perimembranosos se forman no sólo en el área membranas plasmáticas, pero también en el área de todas las estructuras de membrana y, a veces, incluso de las estructuras del citoplasma que no son de membrana (centro celular). Se propone separar esta parte del citoplasma en un subsistema especial de hialoplasma, denotándolo con el término "citosol". Naturalmente, las funciones y la organización morfobioquímica de dicho citosol en la zona del retículo endoplásmico (RE) rugoso y liso, las membranas del aparato de Golgi, las mitocondrias, el centro celular, la envoltura nuclear, etc. serán diferentes y específicas de aquellos. estructuras alrededor de las cuales se forman las correspondientes secciones del citosol.
Teniendo en cuenta la diversidad estructural y funcional de los orgánulos que aseguran el metabolismo intracelular en el citoplasma de las células, la función de las secciones del citosol correspondientes a cada orgánulo específico y prácticamente incluidas en su composición no se limitará únicamente a la función contráctil de soporte. Las estructuras y enzimas que forman el citosol son específicas de cada orgánulo y participan directamente en asegurar los procesos que ocurren en este orgánulo. procesos bioquímicos.
Es aconsejable comenzar a considerar el citoplasma caracterizando el problema de la organización estructural y bioquímica de los ribosomas, las máquinas moleculares de síntesis de proteínas. En este caso, queda especialmente demostrada la importancia de la organización estructural de los procesos bioquímicos en los sistemas celulares y se ha logrado un gran éxito en su estudio.
La segunda área de la biología celular, donde también se ha estudiado con gran detalle la organización estructural de los procesos bioquímicos, es el análisis de las llamadas membranas de apareamiento, el aparato de metabolismo energético de las células. Los consideraremos al caracterizar mitocondrias, plastidios y otras estructuras citoplasmáticas responsables del metabolismo energético en la célula. A continuación, discutimos los resultados del análisis discreto de las membranas principales.
El propio cuerpo celular y su contenido están separados del entorno externo o de los elementos vecinos en los organismos multicelulares por una membrana plasmática. Fuera de la membrana plasmática, extracelular, ubicada membrana celular, o pared, especialmente bien expresada en plantas y organismos procarióticos; en las células animales está ausente o se expresa muy débilmente. Todo el contenido interno de la célula, a excepción del núcleo, se denomina citoplasma. Este es un término general que enfatiza la división de una célula en dos componentes principales: el citoplasma y el núcleo.
Citoplasma de células eucariotas. es heterogéneo en su estructura y composición e incluye hialoplasma, componentes membranosos y no membranosos. Los componentes de la membrana incluyen el sistema vacuolar (retículo endoplásmico, aparato de Golgi, lisosomas, vacuolas vegetales), orgánulos de membrana (mitocondrias y plastidios). Los componentes fuera de la membrana incluyen centriolos, característicos de las células animales, y estructuras y complejos macromoleculares fuera de la membrana, como ribosomas, microtúbulos y microfilamentos. Esta división del citoplasma en componentes separados no significa su aislamiento estructural y funcional entre sí.
3. Hialoplasma: el entorno interno de la célula.
Los términos "hialoplasma" (de hialino - translúcido, transparente), "plasma básico" o "matriz citoplasmática" designan una parte muy importante de la célula, su verdadero entorno interno. EN microscopio electrónico la matriz citoplasmática tiene la apariencia de una sustancia homogénea o de grano fino con baja densidad electrónica. A partir de antiguas observaciones de las propiedades fisicoquímicas del citoplasma, se obtuvo la idea de que el hialoplasma es un sistema coloidal complejo, que incluye varios biopolímeros: proteínas, ácidos nucleicos, polisacáridos, etc. Este sistema es capaz de pasar de un sol (líquido) a estado a gel y viceversa. Por ejemplo, a altas presiones hidrostáticas. citoplasma no espesa, sino que se licua reversiblemente. Este fenómeno se explica por la ruptura de los enlaces entre las moléculas del hialoplasma.
Gracias a los métodos de investigación fisicoquímica y microscópica electrónica, surgieron ideas sobre el plasma principal como un sistema multicomponente organizado y ordenado. Las zonas individuales del hialoplasma pueden cambiar su estado de agregación, dependiendo de las condiciones o de la tarea funcional. Por lo tanto, se sabe que las moléculas individuales de proteínas tubulina pueden dispersarse en el hialoplasma, pero en ciertos momentos comienzan a ensamblarse y construir estructuras tubulares largas: los microtúbulos.
Este proceso de autoensamblaje de microtúbulos es reversible: cuando cambian las condiciones de vida de la célula (aumento de presión o cambios en la permeabilidad de las membranas celulares), los microtúbulos se desintegran en moléculas monoméricas de tubulina. De la misma manera, en un hialoplasma aparentemente sin estructura, pueden surgir y desintegrarse varios complejos fibrilares y filamentosos de moléculas de proteínas. El plasma principal puede participar en la formación de membranas celulares, filamentos y microfilamentos. En las amebas, las capas externas del citoplasma (ectoplasma) o la zona del citoplasma en la región del pseudópodo consisten prácticamente solo en hialoplasma; rara vez se encuentran allí membranas, mitocondrias y otras estructuras. Sin embargo, después de la exposición a iones que promueven la inflamación celular, se producen reordenamientos rápidos y bruscos de estas áreas del citoplasma: en ellas aparecen pequeñas vacuolas de membrana. Por lo tanto, el plasma a granel debe contener todos los componentes estructurales (lípidos y proteínas) necesarios para la formación de membranas.
Las estructuras citoplasmáticas fibrilares también pueden surgir como resultado de procesos de agregación específicos en el hialoplasma. Si se precipitan núcleos grandes, estructuras de membrana y luego ribosomas a partir de homogenados celulares, el sobrenadante restante contendrá los principales componentes químicos del hialoplasma. La micromolécula hialoplasma se compone principalmente de varias proteínas globulares y enzimas de la matriz citoplasmática. Representan el 20-25% contenido general Proteínas en una célula eucariota..
EN células bacterianas, pobres en elementos de membrana, las proteínas hialoplásmicas pueden representar aproximadamente el 50% de todas las proteínas. Las enzimas de matriz más importantes incluyen enzimas glicolíticas, enzimas para el metabolismo de azúcares, bases nitrogenadas, aminoácidos, lípidos y otros compuestos importantes. La matriz contiene enzimas para activar aminoácidos durante la síntesis de proteínas y la transferencia de ARN. Las propiedades osmóticas y tampón de la célula están determinadas en gran medida por la composición y estructura del hialoplasma.
La función más importante del hialoplasma es que este medio semilíquido une todas las estructuras celulares y asegura su interacción química entre sí. La mayoría de los procesos de transporte intracelular se llevan a cabo a través del hialoplasma: la transferencia de aminoácidos, ácidos grasos, nucleótidos y azúcares. En el hialoplasma hay un flujo constante de iones hacia y desde la membrana plasmática, hacia las mitocondrias, el núcleo y las vacuolas. El hialoplasma es el principal receptáculo y área de movimiento de la masa de moléculas de ATP. En él se produce el depósito de productos de reserva: glucógeno, gotas de grasa. Estructural y morfológicamente, el hialoplasma aún está poco estudiado.
4. Membranas citoplasmáticas
Una característica común de todas las membranas celulares, la membrana plasmática externa y todas las membranas intracelulares y orgánulos de membrana es que son capas delgadas (6-10 nm) de naturaleza lipoproteica (lípidos en complejo con proteínas), cerradas sobre sí mismas. No hay membranas abiertas con extremos libres en la célula. Las membranas celulares siempre limitan las cavidades o áreas, cerrándolas por todos lados y separando así el contenido de dichas cavidades del entorno que las rodea. Así, la membrana plasmática, que recubre toda la superficie de la célula, que tiene una forma compleja y numerosas extensiones, no se interrumpe por ningún lado, sino que está cerrada. Separa el contenido del citoplasma del entorno que rodea la célula.
Las membranas cerradas intracelulares forman vesículas: vacuolas esféricas o aplanadas. En este último caso se forman bolsas o depósitos de membrana plana. A menudo, las cavidades delimitadas por membranas tienen una forma compleja, que se asemeja a una esponja o una red, pero incluso en este caso, tales cavidades están delimitadas por una membrana sin interrupción. En tales variantes, las membranas también separan dos fases estructurales del citoplasma: el hialoplasma del contenido de las vacuolas y las cisternas. Las membranas de las mitocondrias y los plastidios tienen la misma propiedad: separan el contenido interno de las cavidades intermembrana y del hialoplasma.
La envoltura nuclear también puede presentarse en forma de un saco hueco de doble membrana perforado de forma esférica. Las membranas de la membrana nuclear delimitan, separan el carioplasma y los cromosomas entre sí de la cavidad del espacio perinuclear y del hialoplasma. Estas propiedades morfológicas generales de las membranas celulares están determinadas por su composición química, su naturaleza lipoproteica.
4.1. Química y propiedades de las membranas celulares.
Principal componentes químicos Las membranas celulares están compuestas por lípidos (40%) y proteínas (60%), además, en muchas membranas se encuentran carbohidratos. Los lípidos incluyen un grupo grande materia orgánica, teniendo poca solubilidad en agua (hidrofobicidad) y solubilidad en disolventes orgánicos y grasas (lipofilicidad).
La composición de los lípidos incluidos en las membranas celulares es muy diversa. Los representantes típicos de los lípidos que se encuentran en las membranas celulares son los fosfolípidos, las esfingomielinas y los lípidos esteroides: el colesterol. Los glicerolípidos son ésteres de un alcohol trihídrico, el glicerol, con dos ácidos grasos y con ácido fosfórico, que, a su vez, pueden estar asociados a diversos grupos químicos (colina, serina, inositol, etanolamina, etc.).
Otro grupo de lípidos de membrana son las esfingomielinas, donde el glicerol se reemplaza por el aminoalcohol esfingosina. De los lípidos relacionados con los esteroides, el colesterol es el más abundante en las membranas. EN células vegetales El colesterol no se detecta, allí es reemplazado por fitoesteroles. Las bacterias no tienen esteroles. Característica distintiva Los lípidos de membrana es la división de su molécula en dos partes funcionalmente diferentes: colas no polares, que no llevan cargas y están formadas por ácidos grasos, y cabezas polares cargadas. Las cabezas polares llevan cargas negativas o pueden ser neutras (si tienen cargas tanto positivas como negativas).
Se descubrió que las membranas celulares difieren mucho entre sí en la composición de lípidos. Así, las membranas plasmáticas de las células animales son ricas en colesterol (hasta un 30%) y tienen poca lecitina, mientras que las membranas mitocondriales, por el contrario, son ricas en fosfolípidos y pobres en colesterol. De numero total Lípidos, el contenido de lecitina en las fracciones del retículo endoplásmico es del 60 al 70% de todos los fosfolípidos, mientras que en la membrana plasmática puede ser del 25 al 35%.
En general, la membrana plasmática se caracteriza alto contenido colesterol y esfingolípidos, así como el predominio de ácidos grasos saturados y monoinsaturados en la composición de los fosfolípidos, mientras que las mitocondrias, el retículo endoplásmico y muchas otras membranas citoplasmáticas contienen poco colesterol y esfingolípidos y relativamente muchos ácidos grasos poliinsaturados. Al parecer, en este sentido, las membranas citoplasmáticas son menos rígidas que la membrana plasmática y son más fusibles.
La cantidad de proteínas en las membranas celulares también varía. Por tanto, las mitocondrias, en comparación con otras membranas, contienen la mayor cantidad de proteínas. Las membranas varían mucho en su conjunto de moléculas de proteínas. Algunos de ellos están asociados a cabezas lipídicas mediante enlaces iónicos y, por tanto, se extraen fácilmente de las membranas con soluciones salinas. Otros forman enlaces salinos con regiones polares de lípidos mediante la interacción con iones Mg2+ o Ca2+. Las proteínas también se extraen utilizando quelatos como el verseno (EDTA). La mayoría de las proteínas interactúan con los lípidos en las membranas basándose en enlaces hidrofóbicos.
Se sabe que muchas proteínas de membrana constan de dos partes: secciones ricas en aminoácidos polares (portadores de carga) y secciones enriquecidas en aminoácidos no polares (glicina, alanina, valion, leucina). Estas proteínas en las capas lipídicas de las membranas están ubicadas de tal manera que sus secciones no polares parecen estar sumergidas en la parte "grasa" de la membrana, donde se encuentran las secciones hidrófobas de los lípidos. La parte polar (hidrófila) de tales proteínas interactúa con las cabezas lipídicas y se enfrenta a la fase acuosa. Por tanto, dichas proteínas asociadas con lípidos mediante interacciones hidrófobas prácticamente no se extraen en fases acuosas. Solo se pueden aislar destruyendo la membrana y extrayendo los lípidos con disolventes orgánicos o detergentes.
Por papel biológico Las proteínas de membrana se pueden dividir en tres grupos: enzimas, proteínas receptoras y proteínas estructurales. El conjunto de enzimas en las membranas puede ser muy grande y diverso (por ejemplo, se encontraron al menos 24 enzimas diferentes en la membrana plasmática de las células hepáticas).
Las diferentes membranas contienen un conjunto característico de enzimas. En otras palabras, la calidad química de las membranas está determinada no sólo por los lípidos, sino también por las proteínas. Todas las membranas celulares tienen en común su estructura lipoproteica; La diferencia radica en las diferencias cuantitativas y cualitativas en lípidos y composición proteica. Por ejemplo, la composición y cantidad de lípidos y proteínas será muy diferente en las membranas del retículo endoplásmico y la membrana interna de las mitocondrias.
El componente de carbohidratos de las membranas está representado principalmente por glicoproteínas, moléculas de proteínas asociadas covalentemente (a diferencia de las nucleoproteínas y lipoproteínas) con cadenas de carbohidratos.
4.2. Estructura de las membranas celulares.
La idea de que las membranas celulares se basan en una bicapa lipídica se obtuvo allá por los años 20. En 1925 se publicó un trabajo que indicaba la formación de una capa bilípida continua de la membrana plasmática. A pesar de una serie de errores, la idea general de la existencia de una capa bilipídica resultó ser correcta. Resultó que si los lípidos se extraen de la membrana de los glóbulos rojos y luego se colocan en la superficie del menisco acuoso, se puede calcular el área ocupada por la monocapa de lípidos resultante. Resultó que esta área es el doble de la que ocupa la superficie de los glóbulos rojos de los que se extrajeron los lípidos.
Los científicos han sugerido que en las membranas de los glóbulos rojos, los lípidos se encuentran en dos capas. Además, resultó que la tensión superficial de la célula es significativamente menor que la tensión superficial de la capa lipídica artificial. Los investigadores descubrieron además que cuando se añadía proteína al aceite, la tensión superficial se reducía a un valor similar a la tensión superficial de las células. En base a esto, se concluyó que la membrana contiene proteínas.
Así surgió la hipótesis sobre la estructura de la membrana, según la cual se trata de un “sándwich” de tres capas (proteína-lípido-proteína). La capa lipídica en el centro es una capa bimolecular en la que los extremos hidrófobos de las moléculas lipídicas están dirigidos entre sí y los extremos hidrófilos están dirigidos hacia las capas de proteínas que se encuentran a ambos lados de la capa lipídica. Esta hipótesis recibió confirmación directa cuando fue posible utilizar la microscopía electrónica para estudiar las membranas celulares. Gracias a ella, en las membranas de la vaina de mielina de los nervios se pudo ver especialmente que la membrana tiene realmente tres capas: dos capas oscuras de 2,5 nm cada una y una capa clara más ancha entre ellas.
La estructura de tres capas de las membranas celulares se puede demostrar en un microscopio electrónico mediante el método de escisión por congelación, lo cual es especialmente importante, ya que en este caso es posible observar enzimas específicas o incluso sistemas enzimáticos. A principios de los años 60. Robertson formuló una hipótesis sobre una única membrana biológica unitaria "elemental". La cuestión de la aparición y formación de membranas en una célula viva aún no se ha estudiado suficientemente. Se sabe que el número y tamaño de los elementos de la membrana en las células puede aumentar y disminuir, es decir, podemos hablar de membranogénesis y destrucción de membranas.
5. Membrana plasmática
La membrana plasmática, o plasmalema, ocupa un lugar especial entre varias membranas celulares. Se trata de una estructura periférica superficial que limita a la célula desde el exterior, lo que determina su conexión directa con el entorno extracelular y, en consecuencia, con todas las sustancias y estímulos que afectan a la célula. Químicamente, la membrana plasmática es un complejo de lipoproteínas.
Los principios fundamentales de la organización de las membranas ya se han discutido en la sección anterior. Tiene unos 10 nm de espesor y es la más gruesa de las membranas celulares. Los principales componentes de las membranas plasmáticas son los lípidos (alrededor del 40%), proteínas (más del 60%) y carbohidratos (alrededor del 1%). Como ya se indicó, el plasmalema, en comparación con otras membranas, es más rico en colesterol y en sus fosfolípidos predominan los ácidos grasos saturados.
La composición de las proteínas incluidas en la membrana plasmática es muy diversa, lo que está determinado por la variedad de sus cargas funcionales. Entre las enzimas se encontraron 5"-nucleotidasa, ATPasa dependiente de Mg, activada por iones Na y K, fosfatasa alcalina y ácida, adenilato ciclasa, RNasa y actividad proteolítica. Las enzimas de la cadena respiratoria y las enzimas glicolíticas están completamente ausentes en la membrana plasmática. de las células eucariotas, pero en las células procarióticas es dentro de la membrana plasmática donde se localizan los elementos de la cadena de transporte de electrones y la fosforilación oxidativa.
Cabe destacar que algunas de las proteínas que se encuentran en las fracciones de la membrana plasmática pueden no estar directamente asociadas con la bicapa lipídica. Muchas enzimas que se encuentran activas en fracciones de la membrana plasmática pueden localizarse en el glicocálix, un complejo de glicoproteína asociado con la membrana plasmática. La composición del glicocálix incluye diversos carbohidratos, que pueden constituir aproximadamente el 1% del peso seco de las membranas.
Los carbohidratos forman cadenas largas y ramificadas de polisacáridos unidos a proteínas en una capa que a menudo determina la forma de la célula. Los sacos de membrana pueden estar adyacentes a la membrana plasmática desde el interior; en este caso, la superficie celular tiene tres capas de membrana: la propia membrana plasmática y dos membranas de los alvéolos peliculares.
En los ciliados, la película forma engrosamientos dispuestos en forma de hexágonos, en cuyo centro emergen los cilios. La rigidez de las formaciones peliculares también puede estar asociada con los elementos del citoplasma subyacentes a la membrana plasmática y la capa cortical. Así, en las crestas de la película de euglena cerca de la membrana, además de las vacuolas de membrana, se encuentran haces paralelos de microtúbulos y microfilamentos. Este refuerzo periférico fibrilar, junto con la periferia de la membrana multicapa plegada, crea la estructura rígida de la película.
5.1. Crecimiento de la membrana plasmática
Después de la división celular, el volumen de células hijas en crecimiento aumenta y, en este sentido, la superficie celular crece y aumenta el área de la membrana plasmática. Pero este no es el único ejemplo. crecimiento rápido volumen y superficie. La superficie de las células de rápido crecimiento en los filamentos estaminales de los cereales puede aumentar 65 veces en 1 hora, es decir, cada minuto el plasmalema aumenta a su tamaño original. Una tasa de crecimiento tan alta de la membrana plasmática sólo puede explicarse por la rápida incorporación de precursores de membrana ya preparados, lípidos y proteínas, en la antigua capa de lipoproteínas. Método similar El crecimiento es bastante posible, ya que se sabe que en la célula hay una reestructuración constante de las membranas: sustitución de unos lípidos por otros, cambios en su cantidad, cambios en la composición de las proteínas, etc. En consecuencia, las membranas son compuestos muy lábiles.
Pero también se ha descrito otro caso de formación de una nueva membrana plasmática. Se perforó el plasmodio del mixomiceto Physarum polycephalum con un capilar de vidrio y el cilindro de plasma resultante, desprovisto de membrana plasmática, se fijó inmediatamente y luego 1, 2, 3 y 6 s después del inicio del experimento, y luego se estudió en un electrón. microscopio. El citoplasma de este hongo se caracteriza por la presencia de una gran cantidad de pequeñas vesículas que contienen moco, que recubre todo el plasmodio desde el exterior. Se pudo observar que dichas burbujas, dispersas en el plasma principal, se acumulaban en el borde del plasma desprotegido por la membrana, formando vacuolas aplanadas ubicadas paralelas a la superficie exterior de la pieza expuesta. Estas vacuolas planas se fusionaron y formaron una membrana continua, una nueva membrana plasmática, que aislaba los componentes plasmáticos del entorno externo.
Al parecer, este proceso de renovación de la membrana plasmática se produce no sólo cuando ésta está gravemente dañada, sino también de forma constante, durante la vida de la célula. Así, como resultado de la exocitosis, las membranas de las vacuolas se fusionan con la membrana plasmática y parecen estar incrustadas en ella, aumentando así la superficie total de la célula. Las observaciones del proceso de secreción (una de las formas de exocitosis) llevaron a muchos investigadores a la idea de que en la célula hay flujos constantes de elementos de membrana: desde la membrana plasmática al citoplasma (endocitosis) y, a la inversa, un flujo de membrana. estructuras desde el citoplasma hasta la superficie celular (exocitosis). En este recambio de membrana, el papel principal lo desempeña el sistema de vacuolas de membrana del aparato de Golgi.
6. Funciones de la membrana plasmática
La membrana plasmática realiza linea entera las funciones celulares más importantes, cuyas principales son la función de separar sustancias citoplasmáticas del entorno externo y la función de transportar diversas sustancias tanto dentro como fuera de la célula. Las funciones de transporte incluyen el transporte pasivo de agua, iones, sustancias de bajo peso molecular y el transporte activo de estas sustancias contra gradientes de concentración, así como diversas formas de transporte de compuestos y complejos de alto peso molecular (endocitosis).
La membrana plasmática, además, participa en la eliminación de los productos formados en ella a partir de las células. También participa en los procesos de descomposición extracelular de biopolímeros. En la superficie del plasmalema existen varias estructuras receptoras que interactúan específicamente con factores extracelulares y células vecinas. Por tanto, la membrana celular participa en la transmisión de señales al interior de la célula. La membrana plasmática participa en las interacciones intercelulares en organismos multicelulares. Ciertas secciones de la membrana plasmática en células animales especializadas participan en la construcción de procesos celulares especiales, como microvellosidades, cilios, procesos receptores, etc. Finalmente, la membrana plasmática juega un papel importante en la división celular.
Esta breve enumeración de las funciones de la membrana plasmática está lejos de ser completa, pero muestra la extrema importancia de esta estructura periférica de cualquier célula, ya sea una célula bacteriana o un organismo eucariota.
6.1. Funciones de transporte
La membrana plasmática, al igual que otras membranas lipoproteicas celulares, es semipermeable. Esto significa que diferentes moléculas la atraviesan a diferentes velocidades, y cuanto mayor es el tamaño de las moléculas, menor es la velocidad de su paso a través de la membrana. Esta propiedad define a la membrana plasmática como una barrera osmótica.
El agua y los gases disueltos en ella tienen la máxima capacidad de penetración, los iones penetran en la membrana mucho más lentamente (unas 104 veces más lento). Por lo tanto, si una célula, por ejemplo un eritrocito, se coloca en un ambiente donde la concentración de sales es menor que en la célula (hipotensión), entonces el agua del exterior entrará en la célula, lo que provocará un aumento de su volumen y rotura de la membrana plasmática (“shock” hipotónico).
Por el contrario, cuando se coloca un eritrocito en soluciones salinas de mayor concentración que en la célula, el agua escapará de él hacia ambiente externo. La célula se encogerá y disminuirá de volumen. Este transporte pasivo de agua desde y hacia la célula todavía se produce a un ritmo bajo. En consecuencia, en la membrana celular, en su capa de lipoproteínas, existen poros especiales para la penetración de agua e iones.
6.2. Funciones receptoras de la membrana plasmática.
Estas funciones están asociadas con la localización en la membrana plasmática de estructuras especiales destinadas al reconocimiento específico de factores químicos o físicos. La superficie celular tiene un gran conjunto de componentes: receptores, que determinan la posibilidad de reacciones específicas con diversos agentes. Como tales receptores en la superficie celular pueden actuar proteínas de membrana o elementos del glicocalix (polisacáridos, glicoproteínas).
Se cree que estas zonas sensibles a sustancias individuales pueden estar dispersas sobre la superficie celular o agrupadas en pequeñas zonas. Por tanto, en la superficie de las células bacterianas o animales existe un número limitado de lugares a los que pueden unirse las partículas virales. Curiosamente, diferentes virus normalmente se unen a diferentes partes de la periferia celular, y la misma célula bacteriana puede tener varias diferentes tipos receptores.
El uso de diversos métodos inmunológicos ha demostrado que las células se localizan antigénicamente en la superficie. ingredientes activos, reaccionando específicamente con anticuerpos o células inmunes. La mayoría de estos antígenos de superficie contienen grupos de carbohidratos (glicolípidos, glicoproteínas). Los componentes antigénicos de la pared bacteriana también suelen ser lipopolisacáridos y glicolípidos. En la superficie de la célula, a menudo directamente en su membrana lipoproteica, hay varios receptores que se unen fisiológicamente sustancias activas, como diversas hormonas, mediadores, etc.
Diferentes celdas Los organismos animales pueden tener diferentes conjuntos de receptores o diferentes sensibilidades del mismo receptor. La función de muchos receptores celulares es transmitir señales desde la superficie a la célula. Actualmente, se ha estudiado bien el sistema de transmisión de estímulos a las células mediante determinadas hormonas, entre las que se encuentran las cadenas peptídicas. Se ha descubierto que estas hormonas se unen a receptores específicos en la superficie de la membrana plasmática celular.
La diversidad y especificidad de los conjuntos de receptores en la superficie de las células conduce a la creación de un sistema muy complejo de marcadores que permite distinguir las células propias (del mismo individuo o de la misma especie) de las extrañas. Células similares interactúan entre sí, lo que conduce a la adhesión de superficies (conjugación en protozoos y bacterias, formación de complejos de células tisulares). En este caso, las células que difieren en un conjunto de marcadores determinantes o no los perciben quedan excluidas de dicha interacción o, en animales superiores, se destruyen como resultado de reacciones inmunológicas.
La membrana plasmática está asociada con la localización de receptores específicos que responden a factores físicos. Por lo tanto, las proteínas, receptores (clorofilas), que interactúan con los cuantos de luz, se localizan en la membrana plasmática o sus derivados en las bacterias fotosintéticas y las algas verdiazules. En la membrana plasmática de las células animales sensibles a la luz hay un sistema especial de proteínas fotorreceptoras (rodopsina), con la ayuda de las cuales la señal luminosa se convierte en una señal química, lo que, a su vez, conduce a la generación de un impulso eléctrico. .
6.3. contactos intercelulares
La membrana plasmática, como ya se mencionó, participa activamente en los contactos intercelulares asociados con la conjugación. organismos unicelulares. En los organismos multicelulares, debido a las interacciones intercelulares, se forman conjuntos celulares complejos, cuyo mantenimiento se puede realizar. En maneras diferentes. En tejidos germinales y embrionarios, especialmente en primeras etapas Durante el desarrollo, las células permanecen conectadas entre sí debido a la capacidad de sus superficies para unirse. Esta propiedad de conexión de las células puede estar determinada por las propiedades de su superficie, que interactúan específicamente entre sí.
El mecanismo de estas conexiones aún no se ha estudiado lo suficiente, pero lo más probable es que se deba a la interacción entre las lipoproteínas y el glicocálix de las membranas plasmáticas. Con tal interacción intercelular de células embrionarias, siempre queda un espacio de aproximadamente 20 nm de ancho, lleno de glicocálix, entre las membranas plasmáticas. El tratamiento del tejido con enzimas que alteran la integridad del glicocálix (mucasas que actúan hidrolíticamente sobre mucinas, mucopolisacáridos) o que dañan la membrana plasmática (proteasas) conduce a la separación de las células entre sí y su disociación. Sin embargo, si se elimina el factor de disociación, las células pueden volver a ensamblarse y reagregarse.
De este modo, es posible disociar células de esponjas de diferentes colores: naranja y amarillo. Resultó que en una mezcla de estas células se forman dos tipos de agregados: que consisten solo en células amarillas y solo en naranja. En este caso, las suspensiones de células mixtas se autoorganizan, restaurando la estructura multicelular original. Se obtuvieron resultados similares con suspensiones de células separadas de embriones de anfibios; en este caso, se produce una separación espacial selectiva de las células del ectodermo del endodermo y del mesénquima.
Además, si se utilizan tejidos para la reagregación últimas etapas desarrollo de embriones, luego en un tubo de ensayo se ensamblan de forma independiente varios conjuntos celulares con especificidad de tejido y órgano, se forman agregados epiteliales similares a los túbulos renales, etc. Las conexiones entre las células de los tejidos y órganos de organismos animales multicelulares pueden formarse mediante estructuras especiales complejas eso llamado contactos intercelulares propiamente dichos. Estos contactos intercelulares estructurados son especialmente pronunciados en los tejidos fronterizos tegumentarios, en los epitelios.
Es posible que la separación primaria de una capa de células conectadas entre sí a través de contactos intercelulares estructurados especiales en la filogénesis de los animales asegurara la formación y el desarrollo de tejidos y órganos. Gracias a la microscopía electrónica se han acumulado muchos datos sobre la ultraestructura de estas formaciones conectivas. Desafortunadamente, ellos composición bioquímica y la estructura molecular aún no se comprende bien.
Al estudiar las conexiones celulares en las capas epiteliales, se pueden detectar las siguientes estructuras que conectan las células entre sí: contacto simple, conexión tipo "cerradura", contacto estrecho, contacto intermedio o zona de adhesión, contacto desmosómico, unión en hendidura.
6.4. Contacto sencillo. conexión tipo bloqueo
El contacto simple ocurre entre la mayoría de las células adyacentes de diferentes orígenes. La mayor parte de la superficie de las células epiteliales en contacto también está conectada por contacto simple, donde las membranas plasmáticas de las células en contacto están separadas por un espacio de 15 a 20 nm. Como ya se mencionó, este espacio representa los componentes supramembranosos de las superficies celulares. El ancho del espacio entre las membranas celulares puede ser superior a 20 nm, formando expansiones y cavidades, pero no inferior a 10 nm. En el lado citoplasmático, no hay estructuras adicionales especiales adyacentes a esta zona de la membrana plasmática.
Una conexión tipo "cerradura" es una protuberancia de la membrana plasmática de una célula hacia la intususcepción (invaginación) de otra. En el corte, este tipo de conexión se asemeja a una costura de carpintero. El espacio intermembrana y el citoplasma en la zona “lock” tienen las mismas características que en las áreas de contacto simple.
6.5. Contacto estrecho
Representa la zona donde las capas externas de las dos membranas plasmáticas están lo más cerca posible. La estructura de tres capas de la membrana en este contacto es a menudo visible: las dos capas osmófilas exteriores de ambas membranas se fusionan en una capa común de 2-3 nm de espesor. La fusión de membranas no ocurre en toda el área de contacto estrecho, sino que representa una serie de conexiones puntuales. En el lado citoplasmático, en esta zona suele haber numerosas fibrillas de unos 8 nm de diámetro, situadas paralelas a la superficie del plasmalema.
Se han encontrado contactos de este tipo entre fibroblastos en cultivos de tejidos y entre epitelio embrionario y células mesenquimales.
6.6. Contacto intermedio (o zona de adhesión)
En este lugar, la distancia intermembrana está algo ampliada (hasta 25-30 nm) y, a diferencia del simple contacto, está llena de un contenido denso, probablemente de naturaleza proteica.
6.7. Contacto de ranura
Representa una región de 0,5 a 3 µm de largo, donde las membranas plasmáticas están separadas por un espacio de 2 a 3 nm, que, después de la ósmosis, le da a toda la estructura una apariencia de siete capas. En el lado citoplasmático no se detectan estructuras especiales cercanas a la membrana. Este tipo de conexión se encuentra en todo tipo de tejidos. La función funcional de la unión gap parece ser la transferencia de iones y moléculas de una célula a otra. Por ejemplo, en el músculo cardíaco, la transmisión de potenciales de acción de una célula a otra se produce a través de un tipo de contacto en el que los iones pueden moverse libremente a través de las uniones intercelulares. El mantenimiento de dicha comunicación iónica entre células depende de la energía obtenida mediante la fosforilación oxidativa.
Bibliografía
1. Vermel E. M. Historia de la doctrina de la célula. M.: Nauka, 1970. 259 p.
2. Gulyaev G.V., Malchenko V.V. Diccionario de términos sobre genética, citología, selección, producción de semillas y ciencia de las semillas. M.: Rosselkhozizdat, 1983. 240 p.
3. De Duve K. Viaje al mundo de una célula viva. M.: Mir, 1987. 253 p.
4. Zavarzin A. A., Kharazova A. D. Fundamentos de citología general: Libro de texto. L.: Editorial Leningr. Universidad, 1982. 240 p.
5. De las moléculas a los humanos./Under. ed. Naumova N. P. M.: Educación, 1973. 480 p.
6. Pausheva Z. P. Taller sobre citología vegetal. M.: Agropromizdat, 1988. 271 p.
7. Troshin A. S., Brown A. D., Vakhtin Yu. B., Zhilkin L. N., Sukhanova K. M. Citología. M.: Educación, 1970. 304 p.
8. Chentsov Yu. S. Citología general. Moscú: Universidad Estatal de Moscú, 1978. 344 p.
Resumen más
plasmalema(membrana celular) de las células animales está formada por una membrana cubierta por fuera con una capa de glicocálix de 10-20 nm de espesor. plasmalema Realiza funciones delimitadoras, de barrera, de transporte y receptoras. Debido a la propiedad de permeabilidad selectiva, el plasmalema regula la composición química del ambiente interno de la célula. El plasmalema contiene moléculas receptoras que reconocen selectivamente determinadas sustancias biológicamente activas (hormonas). En capas y capas, las células vecinas se retienen debido a la presencia diferentes tipos contactos, que están representados por áreas del plasmalema que tienen una estructura especial. La capa cortical está adyacente a la membrana desde el interior. citoplasma espesor 0,1-0,5 micras.
Citoplasma. El citoplasma contiene una serie de estructuras formadas que tienen características estructurales y de comportamiento regulares en diferentes períodos de la vida de la célula. Cada una de estas estructuras tiene una función específica. De ahí surgió su comparación con los órganos de todo el organismo, y por eso recibieron el nombre organelos, o organoides. En el citoplasma se depositan varias sustancias: inclusiones (glucógeno, gotitas de grasa, pigmentos). El citoplasma está impregnado de membranas. retículo endoplásmico.
Retículo endoplásmico (EDR). El retículo endoplasmático es una red ramificada de canales y cavidades en el citoplasma de una célula, formada por membranas. En las membranas de los canales se encuentran numerosas enzimas que aseguran la actividad vital de la célula. Hay 2 tipos membranas EMF- liso y áspero. Sobre membranas retículo endoplasmático liso Hay sistemas enzimáticos involucrados en la grasa y metabolismo de los carbohidratos. Función principal retículo endoplasmático rugoso- síntesis de proteínas, que se produce en los ribosomas adheridos a las membranas. Retículo endoplásmico- este es el sistema circulatorio intracelular general, a través de cuyos canales se transportan sustancias dentro de la célula y de una célula a otra.
ribosomas Realizar la función de síntesis de proteínas. Los ribosomas son partículas esféricas con un diámetro de 15 a 35 nm, que constan de 2 subunidades de tamaños desiguales y que contienen cantidades aproximadamente iguales de proteínas de ARNm. Los ribosomas en el citoplasma están ubicados o adheridos a la superficie exterior de las membranas del retículo endoplásmico. Dependiendo del tipo de proteína sintetizada, los ribosomas se pueden combinar en complejos: polirribosomas. Los ribosomas están presentes en todo tipo de células.
Complejo de Golgi. Elemento estructural principal complejo de Golgi Es una membrana lisa que forma paquetes de cisternas aplanadas, o grandes vacuolas, o pequeñas vesículas. Las cisternas del complejo de Golgi están conectadas a los canales del retículo endoplásmico. Las proteínas, polisacáridos y grasas sintetizadas en las membranas del retículo endoplásmico son transportadas al complejo, condensadas dentro de sus estructuras y “envasadas” en forma de secreción, listas para ser liberadas o utilizadas en la propia célula durante su vida.
Mitocondrias. La distribución universal de las mitocondrias en animales y flora indicar el importante papel que mitocondrias jugando en una jaula. mitocondrias Tienen forma de cuerpos esféricos, ovalados y cilíndricos, y pueden ser filamentosos. El tamaño de las mitocondrias es de 0,2 a 1 micrones de diámetro y de hasta 5 a 7 micrones de longitud. La longitud de las formas filamentosas alcanza las 15-20 micrones. El número de mitocondrias en las células de diferentes tejidos no es el mismo, hay más allí donde los procesos de síntesis son intensos (hígado) o los costes energéticos son elevados. La pared mitocondrial consta de 2 membranas: la exterior y la interior. La membrana externa es lisa y los septos (crestas o crestas) se extienden desde la membrana interna hasta el organoide. Las membranas de las crestas contienen numerosas enzimas implicadas en el metabolismo energético. Función principal de las mitocondrias. - Síntesis de ATP.
lisosomas- pequeños cuerpos ovalados con un diámetro de aproximadamente 0,4 µm, rodeados por una membrana de tres capas. Los lisosomas contienen alrededor de 30 enzimas que pueden descomponer proteínas, ácidos nucleicos, polisacáridos, lípidos y otras sustancias. La descomposición de sustancias mediante enzimas se llama lisis, por lo que el organoide se llama lisosoma. Se cree que los lisosomas se forman a partir de las estructuras del complejo de Golgi o directamente del retículo endoplásmico. Funciones de los lisosomas : digestión intracelular de nutrientes, destrucción de la estructura de la propia célula cuando muere durante el desarrollo embrionario, cuando los tejidos embrionarios se reemplazan por tejidos permanentes y en varios otros casos.
Centríolos. El centro de la celda consta de 2 cuerpos cilíndricos muy pequeños ubicados en ángulo recto entre sí. Estos cuerpos se llaman centríolos. La pared del centríolo consta de 9 pares de microtúbulos. Los centríolos son capaces de autoensamblarse y pertenecen a los orgánulos autorreplicantes del citoplasma. Los centriolos juegan un papel importante en la división celular: inician el crecimiento de microtúbulos que forman el huso de división.
Centro. El núcleo es lo más importante. componente células. Contiene moléculas de ADN y por tanto realiza dos funciones principales: 1) almacenamiento y reproducción de información genética, 2) regulación de los procesos metabólicos que ocurren en la célula. celular perdido centro, no puede existir. El núcleo también es incapaz de existir de forma independiente. La mayoría de las células tienen un núcleo, pero se pueden observar 2-3 núcleos en una célula, por ejemplo en las células del hígado. Se conocen células multinucleadas con un número de núcleos de varias docenas. La forma de los núcleos depende de la forma de la célula. Los granos son esféricos y multilobulados. El núcleo está rodeado por una envoltura que consta de dos membranas que tienen la estructura habitual de tres capas. La membrana nuclear externa está cubierta de ribosomas, la interna es lisa. El papel principal en la vida del núcleo lo desempeña el intercambio de sustancias entre el núcleo y el citoplasma. El contenido del núcleo incluye la savia nuclear o carioplasma, la cromatina y el nucléolo. La composición del jugo nuclear incluye varias proteínas, incluidas la mayoría de las enzimas nucleares, nucleótidos libres, aminoácidos, productos de la actividad del nucleolo y cromatina, que pasan del núcleo al citoplasma. cromatina Contiene ADN, proteínas y representa secciones de cromosomas en espiral y compactadas. nucleolo Es un cuerpo denso y redondo ubicado en el jugo nuclear. El número de nucléolos varía de 1 a 5-7 o más. Los nucléolos están presentes sólo en núcleos que no se dividen; durante la mitosis desaparecen y, una vez completada la división, se forman nuevamente. El nucléolo no es un orgánulo celular independiente; carece de membrana y se forma alrededor de la región del cromosoma en la que está codificada la estructura del ARNr. Los ribosomas se forman en el nucleolo, que luego pasan al citoplasma. cromatina Se denominan bultos, gránulos y estructuras del núcleo en forma de red, que se tiñen intensamente con algunos tintes y difieren en forma del nucléolo.
2)1. Teoría celular
La teoría celular es una idea generalizada de la estructura de las células como unidades vivas, su reproducción y su papel en la formación de organismos multicelulares.
El surgimiento y formulación de disposiciones individuales. teoría celular Fue precedido por un período bastante largo de acumulación de observaciones sobre la estructura de varios organismos unicelulares y multicelulares de plantas y animales. Este período estuvo asociado con el desarrollo del uso y mejora de diversos métodos de investigación óptica.
Robert Hooke fue el primero en observar, utilizando lentes de aumento, la subdivisión del tejido del corcho en “células” o “células”. Sus descripciones inspiraron estudios sistemáticos de la anatomía de las plantas, que confirmaron las observaciones de Robert Hooke y demostraron que varias partes de las plantas estaban compuestas de "vesículas" o "sacos" muy próximos entre sí. Más tarde, A. Leeuwenhoek descubrió el mundo de los organismos unicelulares y vio por primera vez células animales. Las células animales fueron descritas posteriormente por F. Fontana; pero estos y otros numerosos estudios no condujeron en ese momento a una comprensión de la universalidad de la estructura celular, a una idea clara de lo que es una célula. Los avances en el estudio de la microanatomía y las células están asociados con el desarrollo de la microscopía en el siglo XIX. En ese momento, las ideas sobre la estructura de las células habían cambiado: lo principal en la organización de una célula comenzó a considerarse no la pared celular, sino su contenido real, el protoplasma. Se descubrió en el protoplasma un componente permanente de la célula, el núcleo. Todas estas numerosas observaciones permitieron a T. Schwann hacer una serie de generalizaciones en 1838. Demostró que las células vegetales y animales son fundamentalmente similares entre sí. "El mérito de T. Schwann no fue que descubriera las células como tales, sino que enseñó a los investigadores a comprender su significado". Estas ideas se desarrollaron aún más en los trabajos de R. Virchow. La creación de la teoría celular se convirtió en el acontecimiento más importante de la biología, una de las pruebas decisivas de la unidad de toda la naturaleza viva. La teoría celular tuvo un impacto significativo en el desarrollo de la biología y sirvió como base principal para el desarrollo de disciplinas como la embriología, la histología y la fisiología. Proporcionó la base para comprender la vida, explicar las relaciones entre organismos y comprender el desarrollo individual.
Los principios básicos de la teoría celular han conservado su importancia hasta el día de hoy, aunque a lo largo de más de ciento cincuenta años se ha obtenido nueva información sobre la estructura, la actividad vital y el desarrollo de las células. Actualmente, la teoría celular postula:
1) La célula es la unidad elemental de la vida: - no hay vida fuera de la célula.
2) Una célula es un sistema único que consta de muchos elementos que están naturalmente interconectados entre sí, lo que representa una determinada formación integral que consta de unidades funcionales conjugadas: orgánulos u orgánulos.
3) Las células son similares (homólogas) en estructura y propiedades básicas.
4) Las células aumentan en número al dividirse la célula original después de duplicar su material genético: célula por célula.
5) Un organismo multicelular es nuevo sistema, un conjunto complejo de muchas células unidas e integradas en sistemas de tejidos y órganos, conectadas entre sí a través de factores químicos, humorales y nerviosos.
6) Las células de organismos multicelulares son totipotentes, es decir. Tienen el potencial genético de todas las células de un organismo determinado, son equivalentes en información genética, pero se diferencian entre sí en la expresión diferente de varios genes, lo que conduce a su diversidad morfológica y funcional: a la diferenciación.
La idea de una celda como unidad de vida independiente surgió en las obras de T. Schwann. R. Virchow también creía que cada célula lleva dentro de sí una característica completa de la vida: "La célula es el último elemento morfológico de todos los cuerpos vivos, y no tenemos derecho a buscar actividad de vida real fuera de ella".
La ciencia moderna ha demostrado plenamente esta posición. En la literatura popular, a la célula a menudo se la llama "átomo de vida", "cuánto de vida", enfatizando así que la célula es la unidad más pequeña de seres vivos, fuera de la cual no hay vida.
Esta característica general de una célula debe, a su vez, basarse en la definición de lo vivo: qué es vivir, qué es vida. Es muy difícil dar una definición definitiva de los seres vivos, de la vida.
MV Wolkenstein da la siguiente definición de vida: "los organismos vivos son sistemas abiertos, autorreguladores y autorreproductores, cuyas sustancias funcionales más importantes son las proteínas y los ácidos nucleicos". Los seres vivos se caracterizan por una serie de características combinadas, como la capacidad de reproducirse, el uso y transformación de la energía, el metabolismo, la sensibilidad y la variabilidad. Y tal combinación de estos signos se puede encontrar en nivel celular. No existe unidad de vida más pequeña que una célula. Podemos aislar componentes individuales o incluso moléculas de una célula y asegurarnos de que muchos de ellos tengan características funcionales específicas. Por tanto, las fibrillas de actomiosina aisladas pueden contraerse en respuesta a la adición de ATP; fuera de la célula, muchas enzimas implicadas en la síntesis o degradación de moléculas bioorgánicas complejas “funcionan” perfectamente; los ribosomas aislados, en presencia de los factores necesarios, pueden sintetizar proteínas, se han desarrollado sistemas no celulares para la síntesis enzimática de ácidos nucleicos, etc. ¿Se pueden considerar vivos todos estos componentes celulares, estructuras, enzimas y moléculas? ¿Se puede considerar vivo el complejo de actomiosina? Parece que no, aunque sólo sea porque posee sólo una parte del conjunto de propiedades de un ser vivo. Lo mismo se aplica a los otros ejemplos. Sólo la célula como tal es la unidad más pequeña que tiene todas las propiedades combinadas que cumplen con la definición de “vivo”.
3) La base del aparato de superficie de las células (SAC) es externo membrana celular o plasmalema. Además de la membrana plasmática, el PAA tiene un complejo supramembrana y en los eucariotas también hay un complejo submembrana. Los principales componentes bioquímicos del plasmalema (del griego plasma - formación y lema - cáscara, corteza) son los lípidos y las proteínas. Su proporción cuantitativa en la mayoría de los eucariotas es 1:1, y en los procariotas predominan las proteínas en el plasmalema. En la membrana celular externa se encuentra una pequeña cantidad de carbohidratos y se pueden encontrar compuestos similares a las grasas (en los mamíferos: colesterol, vitaminas liposolubles). En 1925, E. Gorter y F. Grendel (Holanda) sugirieron que la base de la membrana es una doble capa de lípidos: una capa bilípida. En 1935, J. Danieli y G. Dawson propusieron el primer modelo espacial de organización de membranas, llamado modelo “sándwich” o “sándwich”. En su opinión, la base de la membrana es una capa bilípida y ambas superficies de la capa están cubiertas con capas continuas de proteínas. Un estudio más detallado de las membranas celulares, incluido el plasmalema, mostró que en casi todos los casos tienen una estructura similar. En 1972, S. Singer y G. Nicholson (EE.UU.) formularon la idea de estructura de mosaico líquido membranas celulares (Fig.). Según este modelo, la base de las membranas es capa bilipídica, pero las proteínas que contiene están ubicadas en moléculas y complejos individuales, es decir. mosaico (del francés mosaico - mosaico; una imagen formada por piezas separadas). En particular, las moléculas de proteínas integrales (del latín entero - enteras) pueden atravesar la capa bilipídica, semiintegrales - parcialmente sumergidas en ella y periféricas (de la periferia griega - círculo) - ubicadas en su superficie (Fig.). La biología molecular moderna ha confirmado la validez del modelo del mosaico fluido, aunque se han descubierto otras variantes de las membranas celulares. En particular, en las arqueobacterias, la base de la membrana es una monocapa de una estructura lipídica compleja, y algunas bacterias contienen vesículas de membrana en el citoplasma, cuyas paredes están representadas por una monocapa de proteínas. complejo supramembrana El aparato de superficie de las células se caracteriza por una variedad de estructuras (Fig.). En los procariotas, el complejo supramembrana en la mayoría de los casos está representado por una pared celular de espesor variable, cuya base es el complejo glicoproteína mureína (en arqueobacterias, pseudomureína). En varias eubacterias, la parte exterior del complejo supramembrana consiste en otra membrana con un alto contenido de lipopolisacáridos. En los eucariotas, el componente universal del complejo supramembrana son los carbohidratos, componentes de los glicolípidos y las glicoproteínas del plasmalema. Debido a esto, originalmente se llamó glicocalix (del griego glicos - dulce, carbohidrato y del latín callum - piel gruesa, cáscara). Además de los carbohidratos, el glicocálix incluye proteínas periféricas por encima de la capa bilípida. Se encuentran variantes más complejas del complejo supramembrana en plantas (pared celular hecha de celulosa), hongos y artrópodos (cubierta externa hecha de quitina). submembrana(del latín sub - under) el complejo es característico únicamente de las células eucariotas. Consiste en una variedad de estructuras proteicas en forma de hilos: fibrillas delgadas (del latín fibrilla - fibra, hilo), microfibrillas (del griego micros - pequeño), fibrillas esqueléticas (del griego esqueleto - seco) y microtúbulos. Están conectados entre sí por proteínas y forman el aparato musculoesquelético de la célula. El complejo submembrana interactúa con las proteínas del plasmalema, que, a su vez, están asociadas con el complejo supramembrana. Como resultado, el PAK es un sistema estructuralmente integral. Esto le permite realizar funciones importantes para la célula: aislante, de transporte, catalítica, de señalización del receptor y de contacto.
4) Las membranas también contienen glicolípidos y colesterol. Glicolípidos- Son lípidos con carbohidratos unidos a ellos. Como los fosfolípidos, glicolípidos Hay cabezas polares y colas apolares. El colesterol está cerca de los lípidos; su molécula también tiene una parte polar.
1. El creador de la teoría celular es:
R. Virchow
Y EN. Vernadsky
T.Schwann
2. La teoría celular se creó en:
1839
3. Grupo de orgánulos de membrana del citoplasma de células eucariotas:
Ribosomas, mitocondrias, plastidios, lisosomas.
esferosomas, vacuolas, plastidios
Microtúbulos, microfilamentos, ribosomas, centrosoma.
RE, ribosomas, lisosomas, aparato de Golgi
mitocondrias, RE, aparato de Golgi
4. Organelos de propósito general:
Miofibrillas
ribosomas, mitocondrias
ER, aparato de Golgi
Flagelos, ribosomas
Flagelos, miofibrillas
5. En el núcleo en interfase se encuentran:
ribosomas
nucleolos
cariolema
jugo nuclear
cromatina
6.Tipos de cromosomas según la localización del centrómero:
subcéntrico
telocéntrico
acrocéntrico
metacéntrico
hombros desiguales
7.Cada cromosoma en metafase de la mitosis consta de:
ADN, proteínas histonas, proteínas no histonas.
2x microfibrillas
2x cromátidas
4x hemicrómátidas
cromonema
8. Las células somáticas tienen un conjunto de cromosomas:
poliploide
diploide
haploide
tetraploide
9. La síntesis se produce en EPS lisos:
carbohidratos
10.Los cromoplastos contienen pigmento:
Hemoglobina
licopeno
caroteno
xantofila
Clorofila
11.Funciones de los ribosomas:
hormonas
Síntesis de carbohidratos
Ácidos nucleicos
síntesis de proteínas
Síntesis de grasas
12. Grupos de inclusiones:
vitamina
trófico
mineral
secretor, excretor
pigmento
13. Las células sexuales tienen un conjunto de cromosomas:
diploide
poliploide
haploide
14. Los eucariotas incluyen:
Cianobacterias
animales
líquenes
plantas
15. Grupo de orgánulos no membranosos del citoplasma de células eucariotas:
Ribosomas, vacuolas, microtúbulos.
ribosomas, centrosoma
EPS, mitocondrias, ribosomas
microtúbulos, microfibrillas
Microfilamentos, plastidios, lisosomas.
16.Funciones del hialoplasma:
participación en el metabolismo
Sintético
Demarcación
homeostático
transporte
17.Funciones de los nucléolos:
Síntesis de proteínas
formación de subunidades ribosómicas
síntesis de ARN de transferencia
Formación de microfibrillas
síntesis de ARN ribosómico
18. Funciones EPS:
Receptor
transporte
delimitando
Protector
sintético
19.Funciones de los lisosomas:
Síntesis y acumulación de carbohidratos.
destrucción de órganos temporales de embriones y larvas.
Síntesis de proteínas, grasas, carbohidratos.
autólisis, destrucción de partículas extrañas.
digestión intracelular
20. Las vacuolas son:
Organelos no membranarios de células animales.
orgánulos de membrana de las células vegetales
Organelos de membrana de células bacterianas.
orgánulos de membrana de animales unicelulares
Organelos no membranosos de las células vegetales.
21.Funciones de las vacuolas:
eliminar el exceso de agua
mantiene la presión osmótica de la célula
realizar la digestión intracelular
Mantiene la presión de turgencia en las células vegetales.
Realizar la síntesis y acumulación de grasas en las células vegetales.
22.Los cloroplastos contienen pigmento:
Xantofila
clorofila
23. La composición de los ribosomas incluye:
ARN-T, magnesio
Proteínas, ADN, magnesio.
solo ardillas
Proteínas, ARNm
proteínas, r-RNA, magnesio
24. Los microfilamentos contienen proteínas:
25. Los microtúbulos contienen proteínas:
tubulina
26. La composición de las microfibrillas incluye proteínas:
colágeno
queratina
27. El núcleo es característico de las células:
Todos los organismos vivos
eucariotas
plantas
procariota