Микроорганизмы (от лат. micros - малый) - организмы, невидимые невооруженным глазом. К ним относятся простейшие, спирохеты, грибы, бактерии, вирусы, изучением которых занимается микробиология. Величина микроорганизмов измеряется в микрометрах (мкм). В микромире существует большое разнообразие форм, которые делятся на группы с учетом общих принципов биологической классификации.
Первой общей биологической классификацией была созданная в XVIII веке система шведского ученого К. Линнея, основанная на морфологических признаках и включавшая животный и растительный мир. С развитием науки в классификации стали учитывать не только морфологические, но и физиологические, биохимические и генетические особенности микроорганизмов. В настоящее время невозможно говорить об единой классификации всех живых организмов: сохраняя единые принципы, классификации макро- и микроорганизмов имеют свои особенности.
Основными ступенями всех классификаций являются: царство - отдел - класс (группа) - порядок - семейство - род - вид. Главной классификационной категорией является вид - совокупность организмов, имеющих общее происхождение, сходные морфологические и физиологические признаки и обмен веществ.
Микроорганизмы относятся к царству прокариотов, представители которых, в отличие от эукариотов, не обладают оформленным ядром. Наследственная информация у прокариотов заключена в молекуле ДНК, располагающейся в цитоплазме клетки.
Для микроорганизмов принята в 1980 г. единая международная классификация, в основе которой лежит система, предложенная американским ученым Берги.
Для того чтобы определить, к какому виду относится микроорганизм, необходимо с помощью различных методов изучить его особенности (форму клетки, спорообразование, подвижность, ферментативные свойства) и по определителю найти его систематическое положение - идентифицировать.
Внутри вида существуют варианты: морфоварианты отличаются по морфологии, биоварианты - по биологическим свойствам, хемоварианты - по ферментативной активности, сероварианты - по антигенной структуре, фаговарианты - по чувствительности к фагам.
Для обозначения микроорганизмов принята общебиологическая бинарная или биноминальная (двойная) номенклатура, введенная К.Линнеем. Первое название обозначает род и пишется с прописной буквы. Второе название обозначает вид и пишется со строчной буквы. Например, Staphylococcus aureus - стафилококк золотистый. В названиях могут быть отражены имена исследователей, открывших микроорганизмы: бруцеллы - в честь Брюса, эшерихии - в честь Эшериха и т. д. В ряд наименований включены органы, которые поражает данный микроорганизм: пневмококки - легкие, менингококки - мозговую оболочку и т. д.
Бактерии
Бактерии - это одноклеточные организмы, лишенные хлорофилла. Средние размеры бактериальной клетки - 2-6 мкм. Размеры и форма клеток бактерий, присущие микроорганизмам определенного вида, могут изменяться под влиянием различных факторов (в зависимости от возраста бактериальной культуры, среды обитания и пр). Это явление называется полиморфизмом.
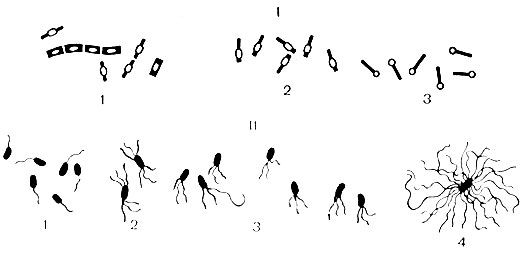
По форме клетки бактерии делятся на три группы: шаровидные, палочковидные и извитые (рис. 4).
Шаровидные бактерии называются кокки (от лат. coccus - ягода) и имеют диаметр клетки от 0,5 до 1 мкм. Форма кокков разнообразна: сферическая, ланцетовидная, бобовидная. По взаимному расположению клеток после деления среди кокков выделяют: микрококки (от лат. micros - малый) - клетки делятся в разных плоскостях и располагаются поодиночке; диплококки (от лат. diploos - двойной) - клетки делятся в одной плоскости и затем располагаются попарно; к ним относятся ланцетовидные пневмококки и бобовидные гонококки и менингококки; стрептококки (от лат. streptos - цепочка) - клетки делятся в одной плоскости и не расходятся, образуя цепочку; стафилококки (от лат. staphyle - гроздь) - клетки делятся в различных плоскостях, образуя скопления в виде грозди винограда; тетракокки (от лат. tetra - четыре) - клетки делятся в двух взаимно перпендикулярных плоскостях и располагаются по четыре; сарцины (от лат. sarcio - соединяю) - клетки делятся в трех взаимно перпендикулярных плоскостях и располагаются в виде тюков или пакетов по 8 или 16 клеток в каждом.
Кокки широко распространены во внешней среде, а также в организме человека и животных. Почти все группы кокков, исключая микрококки, тетракокки и сарцины, включают возбудителей инфекционных заболеваний.
Палочковидные формы называются бактериями. Средние размеры их от 1 до 6 мкм в длину и от 0,5 до 2 мкм в толщину.
Бактерии различаются по внешнему виду: концы их могут быть закругленными (кишечная палочка), обрубленными (возбудитель сибирской язвы), заостренными (возбудитель чумы) или утолщенными (возбудитель дифтерии). После деления бактерии могут располагаться попарно - диплобактерии (клебсиеллы), цепочкой (возбудитель сибирской язвы), иногда под углом друг к другу или крест-накрест (возбудитель дифтерии). Большинство бактерий располагается беспорядочно.
Среди бактерий встречаются изогнутые формы - вибрионы (возбудитель холеры).
К извитым формам относятся спириллы и спирохеты. Форма их клетки напоминает спираль. Большинство спирилл неболезнетворны.
Строение бактериальной клетки
Для изучения строения бактериальной клетки наряду со световым микроскопом применяют электронно-микроскопические и микрохимические исследования, позволяющие определить ультраструктуру бактериальной клетки.
Бактериальная клетка (рис. 5) состоит из следующих частей: трехслойной оболочки, цитоплазмы с различными включениями и ядерного вещества (нуклеоида). Дополнительными структурными образованиями являются капсулы, споры, жгутики, пили.
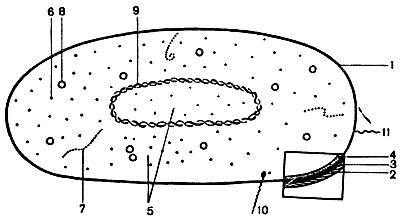
Оболочка клетки состоит из наружного слизистого слоя, клеточной стенки и цитоплазматической мембраны.
Слизистый капсульный слой находится снаружи клетки и выполняет защитную функцию.
Клеточная стенка - один из основных структурных элементов клетки, сохраняющий ее форму и отделяющий клетку от окружающей среды. Важным свойством клеточной стенки является избирательная проницаемость, которая обеспечивает проникновение в клетку необходимых питательных веществ (аминокислот, углеводов и др.) и выведение из клетки продуктов обмена. Клеточная стенка сохраняет внутри клетки постоянное осмотическое давление. Прочность стенки обеспечивает муреин, вещество полисахаридной природы. Некоторые вещества разрушают клеточную стенку, например лизоцим.
Бактерии, полностью лишенные клеточной стенки, называются протопластами. Они сохраняют способность к дыханию, делению, синтезу ферментов; к воздействию внешних факторов: механическому повреждению, осмотическому давлению, аэрации и др. Сохранить протопласты можно только в гипертонических растворах.
Бактерии с частично разрушенной клеточной стенкой называются сферопластами. Если подавить процесс синтеза клеточной стенки с помощью пенициллина, то образуются L-формы, которые у всех видов бактерий представляют шаровидные крупные и мелкие клетки с вакуолями.
Цитоплазматическая мембрана плотно прилегает к клеточной стенке с внутренней стороны. Она очень тонкая (8-10 нм) и состоит из белков и фосфолипидов. Это пограничный полупроницаемый слой, через который осуществляется питание клетки. В мембране находятся ферменты пермеазы, осуществляющие активный перенос веществ, и ферменты дыхания. Цитоплазматическая мембрана образует мезосомы, принимающие участие в делении клетки. При помещении клетки в гипертонический раствор мембрана может отделиться от клеточной стенки.
Цитоплазма - внутреннее содержимое бактериальной клетки. Она представляет собой коллоидную систему, состоящую из воды, белков, углеводов, липидов, различных минеральных солей. Химический состав и консистенция цитоплазмы изменяются в зависимости от возраста клетки и условий окружающей среды. В цитоплазме находятся ядерное вещество, рибосомы и различные включения.
Нуклеоид, ядерное вещество клетки, ее наследственный аппарат. Ядерное вещество прокариотов в отличие от эукариотов не имеет собственной мембраны. Нуклеоид зрелой клетки представляет собой двойную нить ДНК, свернутую в кольцо. В молекуле ДНК закодирована генетическая информация клетки. По генетической терминологии ядерное вещество получило название генофор или геном.
Рибосомы находятся в цитоплазме клетки и выполняют функцию синтеза белка. В состав рибосомы входит 60% РНК и 40% белка. Количество рибосом в клетке достигает 10000. Соединяясь вместе, рибосомы образуют полисомы.
Включения - гранулы, содержащие различные запасные питательные вещества: крахмал, гликоген, жир, волютин. Они расположены в цитоплазме.
Клетки бактерий в процессе жизнедеятельности образуют защитные органеллы - капсулы и споры.
Капсула - внешний уплотненный слизистый слой, примыкающий к клеточной стенке. Это защитный орган, который появляется у некоторых бактерий при попадании их в организм человека и животных. Капсула предохраняет микроорганизм от защитных факторов организма (возбудители пневмонии и сибирской язвы). Некоторые микроорганизмы имеют постоянную капсулу (клебсиеллы).
Споры встречаются только у палочковидных бактерий. Они образуются при попадании микроорганизма в неблагоприятные условия внешней среды (действие высоких температур, высыхание, изменение рН, уменьшение количества питательных веществ в среде и т. д.). Споры находятся внутри бактериальной клетки и представляют уплотненный участок цитоплазмы с нуклеоидом, одетый собственной плотной оболочкой. По химическому составу они отличаются от вегетативных клеток малым количеством воды, увеличенным содержанием липидов и солей кальция, что способствует высокой устойчивости спор. Спорообразование происходит в течение 18-20 ч; при попадании микроорганизма в благоприятные условия спора в течение 4-5 ч прорастает в вегетативную форму. В бактериальной клетке образуется только одна спора, следовательно, споры не являются органами размножения, а служат для переживания неблагоприятных условий.
Спорообразующие аэробные бактерии называются бациллами, а анаэробные - клостридиями.
Споры отличаются по форме, размерам и расположению в клетке. Они могут располагаться центрально, субтерминально и терминально (рис. 6). У возбудителя сибирской язвы спора располагается центрально, ее размер не превышает поперечника клетки. Спора возбудителя ботулизма расположена ближе к концу клетки - субтерминально и превышает ширину клетки. У возбудителя столбняка округлая спора располагается на конце клетки - терминально и значительно превышает ширину клетки.
Жгутики - органы движения, характерны для палочковидных бактерий. Это тонкие нитевидные фибриллы, состоящие из белка - флагеллина. Длина их значительно превышает длину бактериальной клетки. Жгутики отходят от базального тельца, расположенного в цитоплазме, и выходят на поверхность клетки. Наличие их можно обнаружить по определению подвижности клеток под микроскопом, в полужидкой питательной среде или при окраске специальными методами. Ультраструктура жгутиков изучена в электронном микроскопе. По расположению жгутиков бактерии делят на группы (см. рис. 6): монотрихи - с одним жгутиком (возбудитель холеры); амфитрихи - с пучками или единичными жгутиками на обоих концах клетки (спириллы); лофотрихи - с пучком жгутиков на одном конце клетки (фекальный щелочеобразователь); перитрихи - жгутики расположены по всей поверхности клетки (кишечные бактерии). Скорость движения бактерий зависит от количества и расположения жгутиков (наиболее активны монотрихи), от возраста бактерий и влияния окружающих факторов.
Пили или фимбрии - ворсинки, расположенные на поверхности бактериальных клеток. Они короче и тоньше жгутиков и также имеют спиральную структуру. Состоят пили из белка - пилина. Одни пили (их несколько сотен) служат для прикрепления бактерий к клеткам животных и человека, с другими (единичными) связана передача генетического материала из клетки в клетку.
Микоплазмы
Микоплазмы - клетки, не имеющие клеточной стенки, но окруженные трехслойной липопротеидной цитоплазматической мембраной. Микоплазмы могут быть сферической, овальной формы, в виде нитей и звезд. Микоплазмы по классификации Берги выделены в отдельную группу. В настоящее время этим микроорганизмам уделяется все большее внимание как возбудителям заболеваний воспалительного характера. Размеры их различны: от нескольких микрометров до 125-150 нм. Мелкие микоплазмы проходят через бактериальные фильтры и называются фильтрующимися формами.
Спирохеты
Спирохеты (см. рис. 52) (от лат. speira - изгиб, chaite - волосы) - тонкие, извитые, подвижные одноклеточные организмы, имеющие размеры от 5 до 500 мкм в длину и 0,3-0,75 мкм в ширину. С простейшими их роднит способ движения путем сокращения внутренней осевой нити, состоящей из пучка фибрилл. Характер движения спирохет различен: поступательное, вращательное, сгибательное, волнообразное. В остальном строение клетки типичное для бактерий. Некоторые спирохеты слабо окрашиваются анилиновыми красителями. Спирохеты разделяют на роды по количеству и форме завитков нити и ее окончанию. Кроме сапрофитных форм, распространенных в природе и организме человека, среди спирохет имеются болезнетворные - возбудители сифилиса и других заболеваний.
Риккетсии
Вирусы
Среди вирусов выделяют группу фагов (от лат. phagos - пожирающий), вызывающих лизис (разрушение) клеток микроорганизмов. Сохраняя присущие вирусам свойства и состав, фаги отличаются структурой вириона (см. главу 8). Они не вызывают заболеваний человека и животных.
Контрольные вопросы
1. Расскажите о классификации микроорганизмов.
2. Назовите основные свойства представителей царства прокариотов.
3. Перечислите и охарактеризуйте основные формы бактерий.
4. Назовите основные органеллы клетки и их назначение.
5. Дайте краткую характеристику основных групп бактерий и вирусов.
Изучение морфологии микроорганизмов
Для изучения морфологии микроорганизмов применяют микроскопический метод исследования. Важным условием успешного использования этого метода является правильное приготовление мазка из исследуемого материала или бактериальной культуры. Культурой называются микроорганизмы, выращенные на питательных средах в лабораторных условиях.
Техника приготовления мазка
Для работы необходимо иметь чистые и обезжиренные предметные и покровные стекла. Новые стекла кипятят 15-20 мин в 2-5% растворе соды или мыльной воде, споласкивают водой и помещают в слабую хлороводородную кислоту, затем тщательно промывают водой.
Стекла, бывшие в употреблении и загрязненные красителями или иммерсионным маслом, можно обработать двумя способами: 1) погрузить на 2 ч в концентрированную серную кислоту или хромовую смесь, а затем тщательно промыть; 2) кипятить 30-40 мин в 5% растворе соды или щелочи. Необработанные стекла можно обезжирить, натерев их мылом, а затем очистить от него сухой тканью.
Внимание! Если стекло хорошо обезжирено, то капля воды растекается на нем равномерно, не распадаясь на мелкие капли.
Хранят стекла в сосудах с притертыми пробками в смеси Никифорова (равные объемы спирта и эфира) или в 96% спирте. Из растворов стекла извлекают пинцетом.
Внимание! При работе стекла держат пальцами за грани.
Материал для исследования наносят на предметное стекло бактериальной петлей, иглой или пастеровской пипеткой. Чаще всего применяют бактериальную петлю (рис. 7), сделанную из платиновой или нихромовой нити длиной 5-6 см. Петлю закрепляют в петледержателе или впаивают в стеклянную палочку. Конец проволоки сгибают в виде кольца размером 1×1,5 или 2×3 мкм.
Внимание! Правильно приготовленная петля при погружении в воду и извлечении оттуда сохраняет водную пленку.
Перед приготовлением мазка рабочую часть петли прожигают в пламени горелки в вертикальном положении: сначала саму петлю, а затем металлический стержень. Эту манипуляцию проводят и после окончания посева.
Приготовление мазка из культуры, выращенной на жидкой питательной среде . Обезжиренное предметное стекло прожигают в пламени горелки и охлаждают. На предметное стекло, помещенное на подставку (чашку Петри, штатив), наносят культуру. Пробирку с культурой держат большим и указательным пальцами левой руки. Петлю держат в правой руке. Не выпуская петли, мизинцем правой руки прижимают пробку к ладони и осторожно вынимают ее из пробирки. Движения должны быть плавными и спокойными. Горло пробирки обжигают в пламени горелки. Вводят петлю в пробирку. Охлаждают петлю о стенку пробирки и затем погружают ее в культуру. Вынимают петлю, не касаясь ею стенок пробирки. Закрывают пробку, предварительно проведя ее через пламя горелки. Ставят пробирку в штатив. Петлей наносят культуру на предметное стекло, круговыми движениями равномерно распределяя ее. Затем петлю прожигают в пламени горелки. Мазок оставляют для высыхания.
Внимание! Мазок должен быть равномерно растертым, тонким и небольшим (с двухкопеечную монету).
Приготовление мазка из культуры, выращенной на плотной питательной среде . На подготовленное предметное стекло наносят пастеровской пипеткой или петлей каплю изотонического раствора натрия хлорида (0,9%). Культуру осторожно снимают петлей с агара в пробирке или чашке Петри и эмульгируют в капле на стекле. Приготовленный мазок должен быть равномерным и не густым. При его высыхании на предметном стекле остается слабый налет.
Приготовление мазка из гноя или мокроты . Материал забирают стерильной пипеткой или петлей и наносят на середину предметного стекла. Вторым предметным стеклом покрывают первое так, чтобы свободными остались треть первого и второго стекол. Стекла с усилием раздвигают в стороны. Получают два больших мазка.
Приготовление мазка из крови . Каплю крови наносят на предметное стекло на расстоянии одной трети от левого края. Затем краем специально отшлифованного стекла, наклонив его под углом 45°, прикасаются к капле крови. Прижимая отшлифованное стекло к предметному продвигают его вперед. Правильно приготовленный мазок имеет желтоватый цвет и просвечивает.
Приготовление мазков-отпечатков из внутренних органов трупов и пищевых продуктов твердой консистенции . Поверхность органа или пищевого продукта прижигают раскаленным скальпелем и из этого участка вырезают кусочек материала. Пинцетом осторожно захватывают этот кусочек и поверхностью среза прикасаются к предметному стеклу в двух - трех местах, делая ряд мазков-отпечатков.
Высушивание мазка
Мазок высушивают на воздухе при комнатной температуре. В случае необходимости его можно высушить около пламени горелки, держа стекло в горизонтальном положении за края большим и указательным пальцами мазком вверх.
Внимание! При высокой температуре может произойти нарушение структуры клеток.
Фиксация мазка
Мазки фиксируют после полного высыхания с целью: 1) закрепить микроорганизмы на стекле; 2) обезвредить материал; 3) убитые микроорганизмы лучше воспринимают окраску. Фиксированный мазок называется препаратом.
Способы фиксации. 1. Физический - в пламени горелки: стекло берут пинцетом или большим и указательным пальцами и троекратно проводят через верхнюю часть пламени горелки в течение 6 с.
2. Химический - в жидкости: клеточные элементы в мазках из крови и мазках-отпечатках при действии высоких температур разрушаются, поэтому их обрабатывают одной из фиксирующих жидкостей: а) метиловым спиртом- 5 мин; б) этиловым спиртом - 10 мин; в) смесью Никифорова - 10-15 мин; г) ацетоном - 5 мин; д) парами кислоты и формалина - несколько секунд.
Окраска препаратов
После фиксации приступают к окраске препарата.
Окраску препаратов производят на специально оборудованном столе, покрытом линолеумом, пластиком, стеклом и т. д. На столе необходимы сосуд с дистиллированной водой; подставка из двух трубочек или палочек, соединенных резиновыми трубками с обеих сторон (для размещения препаратов); пинцеты, цилиндры, пипетки, фильтровальная бумага, набор красителей, емкость для их слива. Стол для окраски должен находиться рядом с водопроводным краном.
Отношение микроорганизмов к красителям называется их тинкториальными свойствами. В микробиологии широко используют анилиновые красители. Большинство микроорганизмов лучше воспринимает основные красители.
Наиболее употребительны следующие красители: красные (фуксин основной, фуксин кислый, конго красный, нейтральный красный); синие (метиленовый и толуидиновый); фиолетовые (генциановый, метиловый, кристаллический); коричнево-желтые (везувин, хризоидин); зеленые (бриллиантовый, малахитовый).
Все красители выпускают в виде аморфных или кристаллических порошков. Из них готовят насыщенные спиртовые и феноловые растворы, а затем для работы используют водно-спиртовые или водно-феноловые растворы красителей. Если при окраске используют концентрированные растворы красителей, то препарат предварительно накрывают фильтровальной бумагой, на которую наносят краситель. При этом кусочки красителя остаются на бумаге.
Внимание! Каплю красителя наносят пипеткой так, чтобы он покрыл весь препарат.
Рецепты красителей
1. Насыщенные спиртовые растворы (исходные):
Красителя - 1 г спирта 96% - 10 мл
Смесь помещают в термостат до полного растворения на несколько дней. Взбалтывают ежедневно. Хранят в склянках с притертыми пробками.
2. Карболовый фуксин Циля (для окраски кислотоустойчивых микроорганизмов, спор и капсул):
Насыщенного спиртового раствора основного фуксина - 10 мл раствора карболовой кислоты 5% - 90 мл
Внимание! Карболовую кислоту вливают в краситель, а не наоборот.
Смесь в течение нескольких минут энергично встряхивают, фильтруют и сливают во флакон для хранения.
3. Фуксин Пфейффера (для окраски по Граму и для простого метода окраски):
Фуксина Циля - 1 мл воды дистиллированной - 9 мл
Краситель готовят непосредственно перед применением.
4. Карболовый генциановый фиолетовый (для окраски по Граму):
насыщенного спиртового раствора
генцианового фиолетового - 10 мл
карболовой кислоты 5% - 100 мл
Растворы смешивают и фильтруют через бумажный фильтр.
5. Раствор Люголя (для окраски по Граму и реактив на крахмал):
Йодида калия - 2 г кристаллического йода - 1 г дистиллированной воды - 10 мл
Смесь помещают в бутыль матового стекла, хорошо закупоривают и ставят на сутки в термостат, затем добавляют 300 мл дистиллированной воды.
6. Щелочной раствор метиленового синего Леффлера:
Насыщенного спиртового раствора метиленового синего - 30 мл раствора гидроксида калия 1% - 1 мл дистиллированной воды - 100 мл
7. Бумажки по Синеву (для окраски по Граму):
1% спиртовой раствор кристаллического фиолетового
Полоски фильтровальной бумаги пропитывают раствором и высушивают.
Методы окраски делят на ориентировочные (простые) и дифференциальные (сложные), выявляющие химические и структурные особенности бактериальной клетки.
Простой метод окраски
Препарат помещают на подставку для окраски, исследуемым материалом вверх. Пипеткой наносят на него раствор красителя. По истечении указанного времени краситель осторожно сливают, препарат промывают водой и высушивают фильтровальной бумагой. При простом методе используют один краситель. Метиленовым синим и щелочным синим Леффлера окрашивают препарат в течение 3-5 мин, фуксином Пфейффера - 1-2 мин (см. рис. 4).
На окрашенный и высушенный препарат наносят каплю иммерсионного масла и
Сложные методы окраски
Окраска по Граму (универсальный метод) . Наиболее распространенным методом дифференциальной окраски является окраска по Граму.
В зависимости от результатов окраски все микроорганизмы делят на две группы - грамположительные и грамотрицательные.
Грамположительные бактерии содержат в клеточной стенке магниевую соль РНК, которая образует комплексное соединение с йодом и основным красителем (генциановым, метиловым или кристаллическим фиолетовым). Этот комплекс не разрушается при действии спирта, и бактерии сохраняют фиолетовый цвет.
Грамотрицательные бактерии не способны удержать основной краситель, так как не содержат магниевой соли РНК. Под действием спирта краситель вымывается, клетки обесцвечиваются и окрашиваются дополнительным красителем (фуксином) в красный цвет.
1. На препарат накладывают бумажку по Синеву и наносят несколько капель воды или раствор генцианового фиолетового. Окрашивают 1-2 мин. Снимают бумагу или сливают краситель.
2. Не промывая водой, наносят раствор Люголя до почернения (1 мин), затем краситель сливают.
3. Не промывая водой, наносят 96% спирт до отхождения красителя (30-60 с). Можно опустить препарат в стаканчик со спиртом на 1-2 с.
4. Промывают препарат водой.
5. Докрашивают фуксином Пфейффера 3 мин, промывают водой и высушивают.
Микроскопируют с помощью иммерсионной системы.
Окраска по Цилю - Нильсену (для кислотоустойчивых бактерий) . Этот метод применяют для выявления бактерий туберкулеза и проказы, имеющих в оболочке клеток большое количество липидов, воска и оксикислот. Бактерии кислото-, щелоче- и спиртоустойчивы. Для увеличения проницаемости клеточной стенки первый этап окрашивания проводят при подогревании.
1. Фиксированный препарат покрывают фильтровальной бумагой и наносят фуксин Циля. Удерживая стекло пинцетом, препарат подогревают над пламенем горелки до отхождения паров. Добавляют новую порцию красителя и подогревают еще 2 раза. После охлаждения снимают бумагу и промывают препарат водой.
2. Препарат обесцвечивают 5% раствором серной кислоты, погружая 2-3 раза в раствор или наливая кислоту на стекло, затем несколько раз промывают водой.
3. Окрашивают водно-спиртовым раствором метиленового синего в течение 3-5 мин, промывают водой и высушивают.
Микроскопируют с помощью иммерсионной системы.
Кислотоустойчивые бактерии окрашиваются в красный цвет, остальные - в синий (см. рис. 4).
Окраска по Ожешко (выявление спор) . 1. На высушенный на воздухе мазок наливают несколько капель 0,5% раствора хлороводородной кислоты и подогревают до образования паров. Препарат высушивают и фиксируют над пламенем.
2. Окрашивают по способу Циля - Нильсена. Кислотоустойчивые споры окрашиваются в розово-красный, а бактериальная клетка - в голубой цвет (см. рис. 4).
Окраска по Бурри - Гинсу (выявление капсулы) . Этот метод назван негативным, так как окрашивается фон препарата и бактериальная клетка, а капсула остается неокрашенной.
1. На предметное стекло наносят каплю черной туши, разведенной в 10 раз. В нее вносят каплю культуры. Ребром шлифовального стекла делают мазок, так же как мазок крови, и высушивают.
2. Фиксируют химическим способом спиртом или сулемой. Осторожно промывают водой.
3. Окрашивают фуксином Пфейффера 3-5 мин. Осторожно промывают и высушивают на воздухе.
Внимание! Фильтровальной бумагой не пользоваться, чтобы не повредить препарат.
Микроскопируют с помощью иммерсионной системы. Фон препарата черный, клетки - красные, капсулы - неокрашенные (см. рис. 4).
Прижизненная окраска микроорганизмов
Для изучения живой культуры используют чаще всего метиленовый синий и другие красители в больших разведениях (1:10000). Каплю исследуемого материала смешивают на предметном стекле с каплей красителя и накрывают покровным стеклом. Микроскопируют с помощью объектива 40×.
Изучение подвижности микроорганизмов
Для исследования используют культуру бактерий, выращенных в жидкой питательной среде, или взвесь бактерий в изотоническом растворе натрия хлорида.
Метод раздавленной капли . На предметное стекло наносят пипеткой каплю культуры и покрывают ее покровным стеклом. Чтобы не образовывалось пузырьков воздуха, покровное стекло подводят ребром к краю капли и резко опускают его. Для предохранения препарата от высыхания его помещают во влажную камеру.
Влажная камера представляет собой чашку Петри, на дне которой находится влажная фильтровальная бумага. На бумагу кладут две спички и на них помещают препарат. Чашку закрывают крышкой.
Микроскопируют при увеличении объектива 40х в темном поле (см. главу 2).
Метод висячей капли (рис. 8). Для приготовления препарата необходимы стекло с лункой, покровное стекло и вазелин. Края лунки покрывают тонким слоем вазелина.

На покровное стекло наносят каплю культуры. Затем осторожно накрывают покровное стекло стеклом с лункой так, чтобы капля оказалась в центре. Склеившиеся стекла быстро переворачивают покровным стеклом вверх. Капля находится в герметической камере и сохраняется долгое время. При микроскопии сначала при малом увеличении (8×) находят край капли, а затем проводят изучение препарата при большом увеличении.
Контрольные вопросы
1. Как приготовить бактериальную петлю?
2. Назовите цели и способы фиксации мазков.
3. Назовите основные красители.
4. Какими методами изучают подвижность микроорганизмов?
Задание
1. Возьмите готовые препараты, изучите их и зарисуйте основные формы микроорганизмов.
2. Приготовьте мазки из различного материала (культуры, гноя, крови, мазки-отпечатки).
3. Окрасьте препараты сложными методами (по Граму, Цилю - Нильсену, Ожешко, Бурри - Гинсу).
Бактерии - это очень простая форма растительной жизни, которая состоит из одной живой клетки. Размножение осуществляется делением клетки. При достижении стадии зрелости бактерия делится на две равные клетки . В свою очередь каждая из этих клеток достигает зрелости и также делится на две равные клетки. В идеальных условиях бактерия достигает состояния зрелости и размножается менее чем за 20–30 минут. При такой скорости размножения одна бактерия теоретически может произвести 34 триллиона потомков за 24 часа! К счастью, жизненный цикл бактерий относительно короток и продолжается от нескольких минут до нескольких часов. Поэтому даже в идеальных условиях они не могут размножаться с такой скоростью.Скорость роста и размножения бактерий и других микроорганизмов зависит от условий окружающей среды. Температура, свет, наличие кислорода, влажность и рН-фактор (уровень кислотности или щелочности) наряду с наличием питания влияют на скорость развития бактерий. Из них особый интерес у техников и инженеров вызывает температура. Для каждой разновидности бактерий существует минимальная температура, при которой они могут развиваться. При температуре ниже данного порога бактерии впадают в спячку и не способны к воспроизводству. Точно так же для каждой разновидности бактерий существует порог максимальной температуры. При температуре выше этого предела бактерии разрушаются. Между этими пределами находится оптимальная температура, при которой бактерии размножаются с максимальной скоростью. Оптимальная температура для большинства бактерий, которые питаются пометом животных и мертвой тканью животных и растений (сапрофиты), от 24 до 30°С. Оптимальная температура для большинства бактерий, которые являются причиной инфекций и болезней носителя (патогенные бактерии), около 38°С. В большинстве случаев можно значительно снизить скорость размножения бактерий , если понизить температуру окружающей среды. Наконец, существует несколько разновидностей бактерий, которые лучше всего чувствуют себя при температуре воды, в то время как другие — при температуре ее замерзания.
Дополнение к изложенному выше
Происхождение, эволюция, место в развитии жизни на Земле
Бактерии наряду с археями были одними из первых живых организмов на Земле, появившись около 3,9—3,5 млрд лет назад. Эволюционные взаимоотношения между этими группами ещё до конца не изучены, есть как минимум три основные гипотезы: Н. Пэйс предполагает наличие у них общего предка протобактерии, Заварзин считает архей тупиковой ветвью эволюции эубактерий, освоившей экстремальные местообитания; наконец, по третьей гипотезе археи — первые живые организмы, от которых произошли бактерии.Эукариоты возникли в результате симбиогенеза из бактериальных клеток намного позже: около 1,9—1,3 млрд лет назад. Для эволюции бактерий характерен ярко выраженный физиолого-биохимический уклон: при относительной бедности жизненных форм и примитивном строении, они освоили практически все известные сейчас биохимические процессы. Прокариотная биосфера имела уже все существующие сейчас пути трансформации вещества. Эукариоты, внедрившись в неё, изменили лишь количественные аспекты их функционирования, но не качественные, на многих этапах элементов бактерии по-прежнему сохраняют монопольное положение.
Одними из древнейших бактерий являются цианобактерии. В породах, образованных 3,5 млрд лет назад, обнаружены продукты их жизнедеятельности — строматолиты, бесспорные свидетельства существования цианобактерий относятся ко времени 2,2—2,0 млрд лет назад. Благодаря ним в атмосфере начал накапливаться кислород, который 2 млрд лет назад достиг концентраций, достаточных для начала аэробного дыхания. К этому времени относятся образования, свойственные облигатно аэробной Metallogenium.
Появление кислорода в атмосфере нанесло серьёзный удар по анаэробным бактериям. Они либо вымирают, либо уходят в локально сохранившиеся бескислородные зоны. Общее видовое разнообразие бактерий в это время сокращается.
Предполагается, что из-за отсутствия полового процесса, эволюция бактерий идёт по совершенно иному механизму, нежели у эукариот. Постоянный горизонтальный перенос генов приводит к неоднозначностям в картине эволюционных связей, эволюция протекает крайне медленно (а, возможно, с появлением эукариот и вовсе прекратилась), зато в изменяющихся условиях происходит быстрое перераспределение генов между клетками при неизменном общем генетическом пуле.
Строение
Подавляющее большинство бактерий (за исключением актиномицетов и нитчатых цианобактерий) одноклеточны. По форме клеток они могут быть округлыми (кокки), палочковидными (бациллы, клостридии, псевдомонады), извитыми (вибрионы, спириллы, спирохеты), реже — звёздчатыми, тетраэдрическими, кубическими, C- или O-образными. Формой определяются такие способности бактерий, как прикрепление к поверхности, подвижность, поглощение питательных веществ. Отмечено, например, что олиготрофы, то есть бактерии, живущие при низком содержании питательных веществ в среде, стремятся увеличить отношение поверхности к объёму, например, с помощью образования выростов (т. н. простек).Из обязательных клеточных структур выделяют три:
- нуклеоид
- рибосомы
- цитоплазматическая мембрана (ЦПМ)
Строение протопласта
ЦПМ ограничивает содержимое клетки (цитоплазму) от внешней среды. Гомогенная фракция цитоплазмы, содержащая набор растворимых РНК, белков, продуктов и субстратов метаболических реакций, названа цитозолем. Другая часть цитоплазмы представлена различными структурными элементами.Одним из основных отличий клетки бактерий от клетки эукариот является отсутствие ядерной мембраны и, строго говоря, отсутствие вообще внутрицитоплазматических мембран, не являющихся производными ЦПМ. Однако у разных групп прокариот (особенно часто у грамположительных бактерий) имеются локальные впячивания ЦПМ — мезосомы, выполняющие в клетке разнообразные функции и разделяющие её на функционально различные части. У многих фотосинтезирующих бактерий существует развитая сеть производных от ЦПМ фотосинтетических мембран. У пурпурных бактерий они сохранили связь с ЦПМ, легко обнаруживаемую на срезах под электронным микроскопом, у цианобактерий эта связь либо трудно обнаруживается, либо утрачена в процессе эволюции. В зависимости от условий и возраста культуры фотосинтетические мембраны образуют различные структуры — везикулы, хроматофоры, тилакоиды.
Вся необходимая для жизнедеятельности бактерий генетическая информация содержится в одной ДНК (бактериальная хромосома), чаще всего имеющей форму ковалентно замкнутого кольца (линейные хромосомы обнаружены у Streptomyces и Borrelia). Она в одной точке прикреплена к ЦПМ и помещается в структуре, обособленной, но не отделённой мембраной от цитоплазмы, и называемой нуклеоид. ДНК в развёрнутом состоянии имеет длину более 1 мм. Бактериальная хромосома представлена обычно в единственном экземпляре, то есть практически все прокариоты гаплоидны, хотя в определённых условиях одна клетка может содержать несколько копий своей хромосомы, а Burkholderia cepacia имеет три разных кольцевых хромосомы (длиной 3,6; 3,2 и 1,1 млн пар нуклеотидов). Рибосомы прокариот также отличны от таковых у эукариот и имеют константу седиментации 70 S (80 S у эукариот).
Помимо этих структур, в цитоплазме также могут находиться включения запасных веществ.
Клеточная оболочка и поверхностные структуры
Клеточная стенка — важный структурный элемент бактериальной клетки, однако необязательный. Искусственным путём были получены формы с частично или полностью отсутствующей клеточной стенкой (L-формы), которые могли существовать в благоприятных условиях, однако иногда утрачивали способность к делению. Известна также группа природных не содержащих клеточной стенки бактерий — микоплазм.У бактерий существует два основных типа строения клеточной стенки, свойственных грамположительным и грамотрицательным видам.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой толщиной 20—80 нм, построенный в основном из пептидогликана с меньшим количеством тейхоевых кислот и небольшим количеством полисахаридов, белков и липидов (так называемый липополисахарид). В клеточной стенке имеются поры диаметром 1—6 нм, которые делают её проницаемой для ряда молекул.
У грамотрицательных бактерий пептидогликановый слой неплотно прилегает к ЦПМ и имеет толщину лишь 2—3 нм. Он окружён наружной мембраной, имеющей, как правило, неровную, искривлённую форму. Между ЦПМ, слоем пептидогликана и внешней мембраной имеется пространство, называемое периплазматическим, и заполненное раствором, включающим в себя транспортные белки и ферменты.
С внешней стороны от клеточной стенки может находиться капсула — аморфный слой, сохраняющий связь со стенкой. Слизистые слои не имеют связи с клеткой и легко отделяются, чехлы же не аморфны, а имеют тонкую структуру. Однако между этими тремя идеализированными случаями есть множество переходных форм.
Бактериальных жгутиков может быть от 0 до 1000. Возможны как варианты расположения одного жгутика у одного полюса (монополярный монотрих), пучка жгутиков у одного (монополярный перитрих или лофотрихиальное жгутикование) или двух полюсов (биполярный перитрих или амфитрихиальное жгутикование), так и многочисленные жгутики по всей поверхности клетки (перитрих). Толщина жгутика составляет 10—20 нм, длина — 3—15 мкм. Его вращение осуществляется против часовой стрелки с частотой 40—60 об/с.
Помимо жгутиков, среди поверхностных структур бактерий необходимо назвать ворсинки. Они тоньше жгутиков (диаметр 5—10 нм, длина до 2 мкм) и необходимы для прикрепления бактерии к субстрату, принимают участие в метаболитов, а особые ворсинки — F-пили —нитевидные образования, более тонкие и короткие (3—10 нм х 0, 3—10 мкм), чем жгутики — необходимы клетке-донору для передачи реципиенту ДНК при конъюгации.
Размеры
Размеры бактерий в среднем составляют 0,5—5 мкм. Escherichia coli, например, имеет размеры 0,3—1 на 1—6 мкм, Staphylococcus aureus — диаметр 0,5—1 мкм, Bacillus subtilis 0,75 на 2—3 мкм. Крупнейшей из известных бактерий является Thiomargarita namibiensis, достигающая размера в 750 мкм (0,75 мм). Второй является Epulopiscium fishelsoni имеющая диаметр 80 мкм и длину до 700 мкм и обитающая в пищеварительном тракте хирурговой рыбы Acanthurus nigrofuscus. Achromatium oxaliferum достигает размеров 33 на 100 мкм, Beggiatoa alba — 10 на 50 мкм. Спирохеты могут вырастать в длину до 250 мкм при толщине 0,7 мкм. В то же время к бактериям относятся самые мелкие из имеющих клеточное строение организмов. Mycoplasma mycoides имеет размеры 0,1—0,25 мкм, что соответствует размеру крупных вирусов, например, табачной мозаики, коровьей оспы или гриппа. По теоретическим подсчётам сферическая клетка диаметром менее 0,15—0,20 мкм становится неспособной к самостоятельному воспроизведению, поскольку в ней физически не помещаются все необходимые биополимеры и структуры в достаточном количестве.Однако были описаны нанобактерии, имеющие размеры меньше «допустимых» и сильно отличающиеся от обычных бактерий. Они, в отличие от вирусов, способны к самостоятельному росту и размножению (чрезвычайно медленным). Они пока мало изучены, живая их природа ставится под сомнение.
При линейном увеличении радиуса клетки её поверхность возрастает пропорционально квадрату радиуса, а объём — пропорционально кубу, поэтому у мелких организмов отношение поверхности к объёму выше, чем у более крупных, что означает для первых более активный обмен веществ с окружающей средой. Метаболическая активность, измеренная по разным показателям, на единицу биомассы у мелких форм выше, чем у крупных. Поэтому небольшие даже для микроорганизмов размеры дают бактериям и археям преимущества в скорости роста и размножения по сравнению с более сложноорганизованными эукариотами и определяют их важную экологическую роль.
Многоклеточность у бактерий
Одноклеточные формы способны осуществлять все функции, присущие организму, независимо от соседних клеток. Многие одноклеточные прокариоты склонны к образованию клеточных , часто скреплённых выделяемой ими слизью. Чаще всего это лишь случайное объединение отдельных организмов, но в ряде случаев временное объединение связано с осуществлением определённой функции, например, формирование плодовых тел миксобактериями делает возможным развитие цист, при том что единичные клетки не способны их образовывать. Подобные явления наряду с образованием одноклеточными эубактериями морфологически и функционально дифференцированных клеток — необходимые предпосылки для возникновения у них истинной многоклеточности.Многоклеточный организм должен отвечать следующим условиям:
- его клетки должны быть агрегированы,
- между клетками должно осуществляться разделение функций,
- между агрегированными клетками должны устанавливаться устойчивые специфические контакты.
Размножение бактерий
Некоторые бактерии не имеют полового процесса и размножаются лишь равновеликим бинарным поперечным делением или почкованием. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Для обеспечения необходимой для эволюции и приспособления к изменчивой окружающей среде пластичности генотипа у них существуют иные механизмы.При делении большинство грамположительных бактерий и нитчатых цианобактерий синтезируют поперечную перегородку от периферии к центру при участии мезосом. Грамотрицательные бактерии делятся путём перетяжки: на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних. Почкование имеется у разных групп бактерий и, предположительно, возникало несколько раз в процессе эволюции.
У бактерий наблюдается и половое размножение, но в самой примитивной форме. Половое размножение бактерий отличается от полового размножения эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесённая ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смещением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового размножения. Известны 3 способа получения рекомбинантов. Это — в порядке их открытия — трансформация, конъюгация и трансдукция.
2.1. Систематика и номенклатура микробов
Мир микробов можно разделить на клеточные и неклеточные формы. Клеточные формы микробов представлены бактериями, грибами и простейшими. Их можно называть микроорганизмами. Неклеточные формы представлены вирусами, вироидами и прионами.
Новая классификация клеточных микробов включает следующие таксономические единицы: домены, царства, типы, классы, порядки, семейства, роды, виды. В основу классификации микроорганизмов положены их генетическое родство, а также морфологические, физиологические, антигенные и молекулярнобиологические свойства.
Вирусы нередко рассматриваются не как организмы, а как автономные генетические структуры, поэтому они будут рассмотрены отдельно.
Клеточные формы микробов разделены на три домена. Домены Bacteria и Archaebacteria включают микробы с прокариотическим типом строения клетки. Представители домена Eukarya являются эукариотами. Он состоит из 4 царств:
Царства грибов (Fungi, Eumycota);
царства простейших (Protozoa);
царства Chromista (хромовики);
Микробов с неуточненным таксономическим положением (Microspora, микроспоридии).
Различия в организации прокариотической и эукариотической клеток представлены в табл. 2.1.
Таблица 2.1. Признаки прокариотической и эукариотической клетки
2.2. Классификация и морфология бактерий
Термин «бактерия» происходит от слова bacterion, что означает палочка. Бактерии относятся к прокариотам. Их разделяют на два домена: Bacteria и Archaebacteria. Бактерии, входящие в домен Archaebacteria, представляют одну из древнейших форм жизни. Они имеют особенности строения клеточной стенки (у них отсутствует пептидогликан) и рибосомальной РНК. Среди них отсутствуют возбудители инфекционных заболеваний.
Внутри домена бактерии подразделяются на следующие таксономические категории: класс, тип, порядок, семейство, род, вид. Одной из основных таксономических категорий является вид (species). Вид - это совокупность особей, имеющих единое происхождение и генотип, объединенные по близким свойствам, отличающим их от других представителей рода. Название вида соответствует бинарной номенклатуре, т.е. состоит из двух слов. Например, возбудитель дифтерии пишется как Corynebacterium diphtheriae. Первое слово - название рода и пишется с прописной буквы, второе слово обозначает вид и пишется со строчной буквы.
При повторном упоминании вида родовое название сокращается до начальной буквы, например C. diphtheriae.
Совокупность однородных микроорганизмов, выделенных на питательной среде, характеризующихся сходными морфологическими, тинкториальными (отношение к красителям), культуральными, биохимическими и антигенными свойствами, называется чистой культурой. Чистая культура микроорганизмов, выделенных из определенного источника и отличающихся от других представителей вида, называется штаммом. Близким к понятию «штамм» является понятие «клон». Клон представляет собой совокупность потомков, выращенных из единственной микробной клетки.
Для обозначения некоторых совокупностей микроорганизмов, отличающихся по тем или иным свойствам, употребляется суффикс «вар» (разновидность), поэтому микроорганизмы в зависимости от характера различий обозначают как морфовары (отличие по морфологии), резистентовары (отличие по устойчивости, например, к антибиотикам), серовары (отличие по антигенам), фаговары (отличие по чувствительности к бактериофагам), биовары (отличие по биологическим свойствам), хемовары (отличие по биохимическим свойствам) и т.д.
Раньше основу классификации бактерий составляла особенность строения клеточной стенки. Подразделение бактерий по особенностям строения клеточной стенки связано с возможной вариабельностью их окраски в тот или иной цвет по методу Грама. Согласно этому методу, предложенному в 1884 г. датским ученым Х. Грамом, в зависимости от результатов окраски бактерии делятся на грамположительные, окрашиваемые в сине-фиолетовый цвет, и грамотрицательные, окрашиваемые в красный цвет.
В настоящее время основу классификации составляет степень генетического родства, основанная на изучении строения генома рибосомных РНК (рРНК) (см. главу 5), определении процентного содержания в геноме гуанинцитозиновых пар (ГЦ-пары), построении рестрикционной карты генома, изучении степени гибридизации. Также учитываются и фенотипические показатели: отношение к окраске по Граму, морфологические, культуральные и биохимические свойства, антигенная структура.
Домен Bacteria включает 23 типа, из которых медицинское значение имеют нижеизложенные.
Большинство грамотрицательных бактерий объединены в тип Proteobacteria (по имени греческого бога Proteus, способного принимать различные облики). Тип Proteobacteria подразделен на 5 классов:
Класс Alphaproteobacteria (роды Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
класс Betaproteobacteria (роды Вordetellа, Burholderia, Neisseria, Spirillum);
Класс Gammaproteobacteria (представители семейства Enterobacteriaceae, роды Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Класс Deltaproteobacteria (род Bilophila);
Класс Epsilonproteobacteria (роды Campilobacter, Helicobacter). Грамотрицательные бактерии входят также в следующие типы:
тип Chlamydiae (роды Chlamydia, Chlamydophila), тип Spirochaetes (роды Spirocheta, Borrelia, Treponema, Leptospira); тип Bacteroides (роды Bacteroides, Prevotella, Porphyromonas).
Грамположительные бактерии входят в следующие типы:
Тип Firmicutes включает класс Clostridium (роды Clostridium, Peptococcus), класс Bacilli (Listeria, Staphylococcus, Lactobacillus, Streptococcus) и класс Mollicutes (роды Mycoplasma, Ureaplasma), которые являются бактериями, не имеющими клеточную стенку;
тип Actinobacteria (роды Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. Морфологические формы бактерий
Различают несколько основных форм бактерий: кокковидные, палочковидные, извитые и ветвящиеся (рис. 2.1).
Сферические формы, или кокки - шаровидные бактерии размером 0,5-1 мкм, которые по взаимному расположению делятся на микрококки, диплококки, стрептококки, тетракокки, сарцины и стафилококки.
Микрококки (от греч. micros - малый) - отдельно расположенные клетки.
Диплококки (от греч. diploos - двойной), или парные кокки, располагаются парами (пневмококк, гонококк, менингококк), так как клетки после деления не расходятся. Пневмококк (возбудитель пневмонии) имеет с противоположных сторон ланцетовидную форму, а гонококк (возбудитель гонореи) и менингококк (возбу-
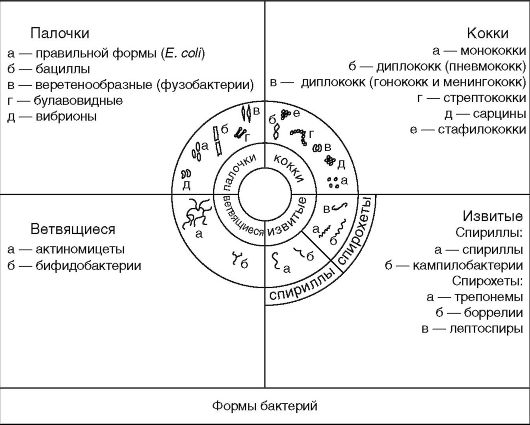 Рис. 2.1.
Формы бактерий
Рис. 2.1.
Формы бактерий
дитель эпидемического менингита) имеют форму кофейных зерен, обращенных вогнутой поверхностью друг к другу.
Стрептококки (от греч. streptos - цепочка) - клетки округлой или вытянутой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления.
Сарцины (от лат. sarcina - связка, тюк) располагаются в виде пакетов из 8 кокков и более, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях.
Стафилококки (от греч. staphyle - виноградная гроздь) - кокки, расположенные в виде грозди винограда в результате деления в разных плоскостях.
Палочковидные бактерии различаются по размерам, форме концов клетки и взаимному расположению клеток. Длина клеток 1-10 мкм, толщина 0,5-2 мкм. Палочки могут быть правильной
(кишечная палочка и др.) и неправильной булавовидной (коринебактерии и др.) формы. К наиболее мелким палочковидным бактериям относятся риккетсии.
Концы палочек могут быть как бы обрезанными (сибиреязвенная бацилла), закругленными (кишечная палочка), заостренными (фузобактерии) или в виде утолщения. В последнем случае палочка похожа на булаву (коринебактерии дифтерии).
Слегка изогнутые палочки называются вибрионами (холерный вибрион). Большинство палочковидных бактерий располагается беспорядочно, так как после деления клетки расходятся. Если после деления клетки остаются связанными общими фрагментами клеточной стенки и не расходятся, то они располагаются под углом друг к другу (коринебактерии дифтерии) или образуют цепочку (сибиреязвенная бацилла).
Извитые формы - спиралевидные бактерии, которые бывают двух видов: спириллы и спирохеты. Спириллы имеют вид штопорообразно извитых клеток с крупными завитками. К патогенным спириллам относятся возбудитель содоку (болезнь укуса крыс), а также кампилобактерии и хеликобактерии, имеющие изгибы, напоминающие крылья летящей чайки. Спирохеты представляют тонкие длинные извитые бактерии, отличающиеся от спирилл более мелкими завитками и характером движения. Особенность их строения описана ниже.
Ветвящиеся - палочковидные бактерии, которые могут иметь разветвление в форме латинской буквы Y, встречающиеся у бифидобактерий, также быть представленными в виде нитевидных разветвленных клеток, способных переплетаться, образуя мицелий, что наблюдается у актиномицет.
2.2.2. Структура бактериальной клетки
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и наследственного аппарата - аналога ядра, называемого нуклеоидом (рис. 2.2). Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
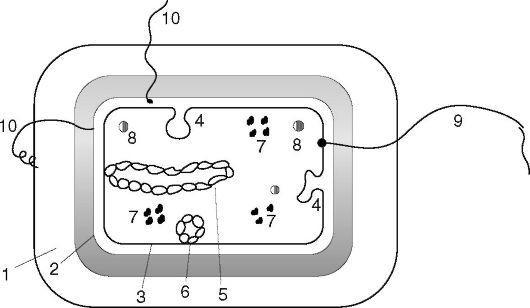 Рис. 2.2.
Структура
бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 -
цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7
- рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Рис. 2.2.
Структура
бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 -
цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7
- рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной сдерживающая высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов, имеет рецепторы для бактериофагов, бактериоцинов и различных веществ. Наиболее толстая клеточная стенка у грамположительных бактерий (рис. 2.3). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более.
Основу клеточной стенки бактерий составляет пептидогликан. Пептидогликан является полимером. Он представлен параллельными полисахаридными гликановыми цепями, состоящими из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эту связь разрывает лизоцим, являющийся ацетилмурамидазой.
К N-ацетилмурамовой кислоте ковалентными связями присоединен тетрапептид. Тетрапептид состоит из L-аланина, который связан с N-ацетилмурамовой кислотой; D-глутамина, который у грамположительных бактерий соединен с L-лизином, а у грамотри-
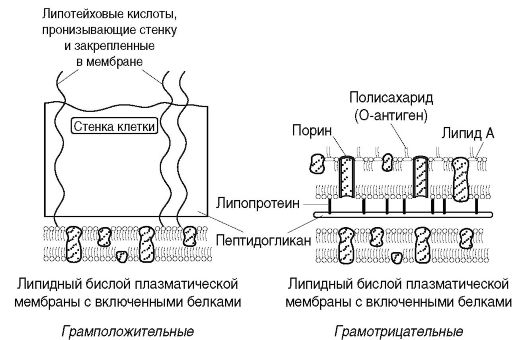 Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
цательных бактерий - с диаминопимелиновой кислотой (ДАП), которая представляет собой предшественник лизина в процессе бактериального биосинтеза аминокислот и является уникальным соединением, присутствующим только у бактерий; 4-й аминокислотой является D-аланин (рис. 2.4).
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов и белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. Тетрапептиды разных слоев пептидогликана у грамположительных бактерий соединены друг с другом полипептидными цепочками из 5 остатков глицина (пентаглицина), что придает пептидогликану жесткую геометрическую структуру (рис. 2.4, б). С пептидогликаном ктеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. tekhos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного, с поперечными пептидными сшивками пептидогликана.
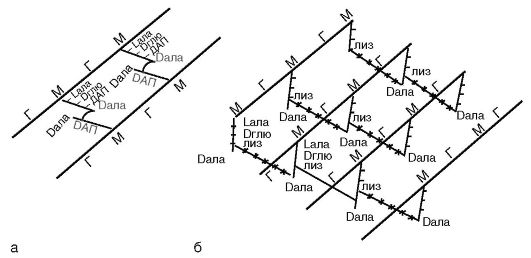 Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого последующая обработка мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краситель в клеточной стенке.
Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10% массы клеточной стенки); они обесцвечиваются спиртом, и при обработке фуксином или сафранином приобретают красный цвет. Это связано с особенностями строения клеточной стенки. Пептидогликан в клеточной стенке грамотрицательных бактерий представлен 1-2 слоями. Тетрапептиды слоев соединены между собой прямой пептидной связью между аминогруппой ДАП одного тетрапептида и карбоксильной группой D-аланина тетрапептида другого слоя (рис. 2.4, а). Кнаружи от пептидогликана расположен слой липопротеина, соединенный с пептидогликаном через ДАП. За ним следует наружная мембрана клеточной стенки.
Наружная мембрана является мозаичной структурой, представленной липополисахаридами (ЛПС), фосфолипидами и белками. Внутренний слой ее представлен фосфолипидами, а в наружном слое расположен ЛПС (рис. 2.5). Таким образом, наружная мем-
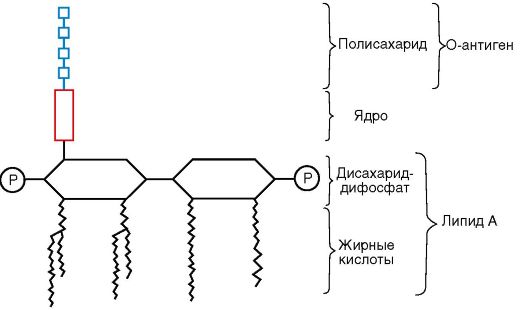 Рис. 2.5.
Структура липополисахарида
Рис. 2.5.
Структура липополисахарида
брана асимметрична. ЛПС наружной мембраны состоит из трех фрагментов:
Липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий. Липид А состоит из фосфорилированных глюкозоаминовых дисахаридных единиц, к которым прикреплены длинные цепочки жирных кислот (см. рис. 2.5);
Ядра, или стержневой, коровой части (от лат. core - ядро), относительно консервативной олигосахаридной структуры;
Высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями.
ЛПС заякорен в наружной мембране липидом А, обусловливающим токсичность ЛПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота. О-специфическая полисахаридная цепь, отходящая от стержневой части молекулы ЛПС,
состоящая из повторяющихся олигосахаридных единиц, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Генетические изменения могут привести к дефектам, укорочению ЛПС бактерий и появлению в результате этого шероховатых колоний R-форм, теряющих О-антигенную специфичность.
Не все грамотрицательные бактерии имеют полноценную О-специфическую полисахаридную цепь, состоящую из повторяющихся олигосахаридных единиц. В частности, бактерии рода Neisseria имеют короткий гликолипид, который называется липоолигосахаридом (ЛОС). Он сравним с R-формой, потерявшей О-антигенную специфичность, наблюдаемой у мутантных шероховатых штаммов E. coli. Структура ЛОС напоминает структуру гликосфинголипида цитоплазматической мембраны человека, поэтому ЛОС мимикрирует микроб, позволяя ему избегать иммунного ответа хозяина.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы с относительной массой до 700 Д.
Между наружной и цитоплазматической мембраной находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, β-лактамазы), а также компоненты транспортных систем.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сфероили протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия Института им. Д. Листера, где они впер-
вые были изучены). L-формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, возвращаясь в исходную бактериальную клетку. L-формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым - промежуточным). По структуре она похожа на плазмолемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. В отличие от эукариотических клеток, в цитоплазматической мембране бактериальной клетки отсутствуют стеролы (за исключением микоплазм).
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы - АТФазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии. Цитоплазма занимает основной объем бактери-
альной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы: 50S и 30S. рРНК - консервативные элементы бактерий («молекулярные часы» эволюции). 16S-рРНК входит в состав малой субъединицы рибосом, а 23S-рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, β-оксимасляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в краснофиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронноплотных гранул размером 0,1-1 мкм.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, плотно уложенной наподобие клубка. Нуклеоид бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У большинства бактерий содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Но у некоторых бактерий имеются две хромосомы кольцевой формы (V. cholerae) и линейные хромосомы (см. раздел 5.1.1). Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК
методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды (см. раздел 5.1.2), представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь. Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка по Бурри- Гинсу, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Она препятствует фагоцитозу бактерий. Капсула антигенна: антитела к капсуле вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии.
От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют гликокаликсом.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие на-
чало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из трех частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (одна пара дисков у грамположительных и две пары у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
Жгутики состоят из белка - флагеллина (flagellum - жгутик), являющегося антигеном - так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий разных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования, более тонкие и короткие (3-10 нм * 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Известно несколько типов пилей. Пили общего типа отвечают за прикрепления к субстрату, питание и водно-солевой обмен. Они многочисленны - несколько сотен на клетку. Половые пили (1-3 на клетку) создают контакт между клетками, осуществляя между ними передачу генетической информации путем конъюгации (см. главу 5). Особый интерес представляют пили IV типа, у которых концы обладают гидрофобностью, в результате чего они закручиваются, эти пили называют еще кудряшками. Располага-
ются они по полюсам клетки. Эти пили встречаются у патогенных бактерий. Они обладают антигенными свойствами, осуществляют контакт бактерии с клеткой-хозяином, участвуют в образовании биопленки (см. главу 3). Многие пили являются рецепторами для бактериофагов.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки. Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (от лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена в красный, а вегетативная клетка - в синий цвет.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор бывает овальной и шаровидной, расположение в клетке - терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены) и центральное (у сибиреязвенной бациллы).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора.
В протопласте проспоры находятся нуклеоид, белоксинтезирующая система и система получения энергии, основанная на гликолизе. Цитохромы отсутствуют даже у аэробов. Не содержится АТФ, энергия для прорастания сохраняется в форме 3-глицеринфосфата.
Проспору окружают две цитоплазматические мембраны. Слой, окружающий внутреннюю мембрану споры, называется стенкой споры, он состоит из пептидогликана и является главным источником клеточной стенки при прорастании споры.
Между наружной мембраной и стенкой споры формируется толстый слой, состоящий из пептидогликана, имеющего много сшивок, - кортекс.
Кнаружи от внешней цитоплазматической мембраны расположена оболочка споры, состоящая из кератиноподобных белков, со-
держащих множественные внутримолекулярные дисульфидные связи. Эта оболочка обеспечивает резистентность к химическим агентам. Споры некоторых бактерий имеют дополнительный покров - экзоспориум липопротеиновой природы. Таким образом формируется многослойная плохо проницаемая оболочка.
Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: активации, инициации, вырастания. При этом из одной споры образуется одна бактерия. Активация - это готовность к прорастанию. При температуре 60-80 °С спора активируется для прорастания. Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.
2.2.3. Особенности строения спирохет, риккетсий, хламидий, актиномицет и микоплазм
Спирохеты - тонкие длинные извитые бактерии. Они состоят из наружной мембранной клеточной стенки, которая окружает цитоплазматический цилиндр. Поверх наружной мембраны располагается прозрачный чехол гликозаминогликановой природы. Под наружной мембранной клеточной стенки располагаются фибриллы, закручивающиеся вокруг цитоплазматического цилиндра, придавая бактериям винтообразную форму. Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Число и расположение фибрилл варьируют у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками. Спирохеты плохо воспринимают красители. Обычно их окрашивают по Романовскому-Гимзе или серебрением. В живом
виде спирохеты исследуют с помощью фазово-контрастной или темнопольной микроскопии.
Спирохеты представлены тремя родами, патогенными для человека: Treponema, Borrelia, Leptospira.
Трепонемы (род Treponema) имеют вид тонких штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками. Вокруг протопласта трепонем расположены 3-4 фибриллы (жгутики). В цитоплазме имеются цитоплазматические филаменты. Патогенными представителями являются Т. pallidum - возбудитель сифилиса, T. pertenue - возбудитель тропической болезни - фрамбезии. Имеются и сапрофиты - обитатели полости рта человека, ила водоемов.
Боррелии (род Borrelia), в отличие от трепонем, более длинные, имеют по 3-8 крупных завитков и 7-20 фибрилл. К ним относятся возбудитель возвратного тифа (В. recurrentis) и возбудители болезни Лайма (В. burgdorferi) и других заболеваний.
Лептоспиры (род Leptospira) имеют завитки неглубокие и частые в виде закрученной веревки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв S или С; имеют две осевые фибриллы. Патогенный представитель L. interrogans вызывает лептоспироз при попадании в организм с водой или пищей, приводя к кровоизлияниям и желтухе.
Риккетсии обладают независимым от клетки хозяина метаболизмом, однако, возможно, они получают от клетки хозяина макроэргические соединения для своего размножения. В мазках и тканях их окрашивают по Романовскому-Гимзе, по Маккиавелло- Здродовскому (риккетсии красного цвета, а инфицированные клетки - синего).
У человека риккетсии вызывают эпидемический сыпной тиф (R. prowazekii), клещевой риккетсиоз (R. sibirica), пятнистую лихорадку Скалистых гор (R. rickettsii) и другие риккетсиозы.
Строение их клеточной стенки напоминает таковую грамотрицательных бактерий, хотя имеются отличия. Она не содержит типичного пептидогликана: в его составе полностью отсутствует N-ацетилмурамовая кислота. В состав клеточной стенки входит двойная наружная мембрана, которая включает липополисахарид и белки. Несмотря на отсутствие пептидогликана, клеточная стенка хламидий обладает ригидностью. Цитоплазма клетки ограничена внутренней цитоплазматической мембраной.
Основным методом выявления хламидий является окраска по Романовскому-Гимзе. Цвет окраски зависит от стадии жизненного цикла: элементарные тельца окашиваются в пурпурный цвет на фоне голубой цитоплазмы клетки, ретикулярные тельца - в голубой цвет.
У человека хламидии вызывают поражения глаз (трахома, конъюнктивит), урогенитального тракта, легких и др.
Актиномицеты - ветвящиеся, нитевидные или палочковидные грамположительные бактерии. Свое название (от греч. actis - луч, mykes - гриб) они получили в связи с образованием в пораженных тканях друз - гранул из плотно переплетенных нитей в виде
лучей, отходящих от центра и заканчивающихся колбовидными утолщениями. Актиномицеты, как и грибы, образуют мицелий - нитевидные переплетающиеся клетки (гифы). Они формируют субстратный мицелий, образующийся в результате врастания клеток в питательную среду, и воздушный, растущий на поверхности среды. Актиномицеты могут делиться путем фрагментации мицелия на клетки, похожие на палочковидные и кокковидные бактерии. На воздушных гифах актиномицетов образуются споры, служащие для размножения. Споры актиномицетов обычно не термостойки.
Общую филогенетическую ветвь с актиномицетами образуют так называемые нокардиоподобные (нокардиоформные) актиномицеты - собирательная группа палочковидных бактерий неправильной формы. Их отдельные представители образуют ветвящиеся формы. К ним относят бактерии родов Corynebacterium, Mycobacterium, Nocardia и др. Нокардиоподобные актиномицеты отличаются наличием в клеточной стенке сахаров арабинозы, галактозы, а также миколовых кислот и больших количеств жирных кислот. Миколовые кислоты и липиды клеточных стенок обусловливают кислотоустойчивость бактерий, в частности микобактерий туберкулеза и лепры (при окраске по Цилю-Нельсену они имеют красный цвет, а некислотоустойчивые бактерии и элементы ткани, мокроты - синий цвет).
Патогенные актиномицеты вызывают актиномикоз, нокардии - нокардиоз, микобактерии - туберкулез и лепру, коринебактерии - дифтерию. Сапрофитные формы актиномицетов и нокардиоподобных актиномицетов широко распространены в почве, многие из них являются продуцентами антибиотиков.
Микоплазмы - мелкие бактерии (0,15-1 мкм), окруженные только цитоплазматической мембраной, содержащей стеролы. Они относятся к классу Mollicutes. Из-за отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют разнообразную форму: кокковидную, нитевидную, колбовидную. Эти формы видны при фазово-контрастной микроскопии чистых культур микоплазм. На плотной питательной среде микоплазмы образуют колонии, напоминающие яичницу-глазунью: центральная непрозрачная часть, погруженная в среду, и просвечивающая периферия в виде круга.
Микоплазмы вызывают у человека атипичную пневмонию (Mycoplasma pneumoniae) и поражения мочеполового тракта
(М. hominis и др.). Микоплазмы вызывают заболевания не только у животных, но и у растений. Достаточно широко распространены и непатогенные представители.
2.3. Строение и классификация грибов
Грибы относятся к домену Eukarya, царству Fungi (Mycota, Mycetes). Недавно грибы и простейшие были разделены на самостоятельные царства: царство Eumycota (настоящие грибы), царство Chromista и царство Protozoa. Некоторые микроорганизмы, ранее считавшиеся грибами или простейшими, были перемещены в новое царство Chromista (хромовики). Грибы - многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с толстой клеточной стенкой. Они имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннаны, глюканы, целлюлоза, хитин), а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы (в отличие от холестерина - главного стерола тканей млекопитающих). Большинство грибов - облигатные или факультативные аэробы.
Грибы широко распространены в природе, особенно в почве. Некоторые грибы содействуют производству хлеба, сыра, молочнокислых продуктов и алкоголя. Другие грибы продуцируют антимикробные антибиотики (например, пенициллин) и иммунодепрессивные лекарства (например, циклоспорин). Грибы используют генетики и молекулярные биологи для моделирования различных процессов. Фитопатогенные грибы наносят значительный ущерб сельскому хозяйству, вызывая грибковые болезни злаковых растений и зерна. Инфекции, вызываемые грибами, называются микозами. Различают гифальные и дрожжевые грибы.
Гифальные (плесневые) грибы, или гифомицеты, состоят из тонких нитей толщиной 2-50 мкм, называемых гифами, которые сплетаются в грибницу или мицелий (плесень). Тело гриба называется талломом. Различают демациевые (пигментированные - коричневые или черные) и гиалиновые (непигментированные) гифомицеты. Гифы, врастающие в питательный субстрат, отвечают за питание гриба и называются вегетативными гифами. Гифы, ра-
стущие над поверхностью субстрата, называются воздушными или репродуктивными гифами (отвечают за размножение). Колонии из-за воздушного мицелия имеют пушистый вид.
Различают низшие и высшие грибы: гифы высших грибов разделены перегородками, или септами с отверстиями. Гифы низших грибов не имеют перегородок, представляя собой многоядерные клетки, называемые ценоцитными (от греч. koenos - единый, общий).
Дрожжевые грибы (дрожжи) в основном представлены отдельными овальными клетками диаметром 3-15 мкм, а их колонии, в отличие от гифальных грибов, имеют компактный вид. По типу полового размножения они распределены среди высших грибов - аскомицет и базидиомицет. При бесполом размножении дрожжи образуют почки или делятся. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток - «сарделек». Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом - почкованием или делением. Понятия «дрожжеподобные грибы» часто идентифицируют с понятием «дрожжи».
Многие грибы обладают диморфизмом - способностью к гифальному (мицелиальному) или дрожжеподобному росту в зависимости от условий культивирования. В инфицированном организме они растут в виде дрожжеподобных клеток (дрожжевая фаза), а на питательных средах образуют гифы и мицелий. Диморфизм связан с температурным фактором: при комнатной температуре образуется мицелий, а при 37 °С (при температуре тела человека) - дрожжеподобные клетки.
Грибы размножаются половым или бесполым способом. Половое размножение грибов происходит с образованием гамет, половых спор и других половых форм. Половые формы называются телеоморфами.
Бесполое размножение грибов происходит с образованием соответствующих форм, называемых анаморфами. Такое размножение происходит почкованием, фрагментацией гиф и бесполыми спорами. Эндогенные споры (спорангиоспоры) созревают внутри округлой структуры - спорангия. Экзогенные споры (конидии) формируются на кончиках плодоносящих гиф, так называемых конидиеносцах.
Различают разнообразые конидии. Артроконидии (артроспоры), или таллоконидии, образуются при равномерном септировании и расчленении гиф, а бластоконидии образуются в результате почкования. Небольшие одноклеточные конидии называются микроконидиями, большие многоклеточные конидии - макроконидиями. К бесполым формам грибов относят также хламидоконидии, или хламидоспоры (толстостенные крупные покоящиеся клетки или комплекс мелких клеток).
Различают совершенные и несовершенные грибы. Совершенные грибы имеют половой способ размножения; к ним относят зигомицеты (Zygomycota), аскомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Несовершенные грибы имеют только бесполый способ размножения; к ним относят формальный условный тип/ группу грибов - дейтеромицеты (Deiteromycota).
Зигомицеты относятся к низшим грибам (мицелий несептированный). Они включают представителей родов Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Распространены в почве и воздухе. Могут вызывать зигомикоз (мукоромикоз) легких, головного мозга и других органов человека.
При бесполом размножении зигомицет на плодоносящей гифе (спорангиеносце) образуется спорангий - шаровидное утолщение с оболочкой, содержащее многочисленные спорангиоспоры (рис. 2.6, 2.7). Половое размножение у зигомицетов происходит с помощью зигоспор.
Аскомицеты (сумчатые грибы) имеют септированный мицелий (кроме одноклеточных дрожжей). Свое название они получили от основного органа плодоношения - сумки, или аска, содержащего 4 или 8 гаплоидных половых спор (аскоспор).
К аскомицетам относятся отдельные представители (телеоморфы) родов Aspergillus и Penicillium. Большинство грибов родов Aspergillus, Penicillium являются анаморфами, т.е. размножаются только беспо-
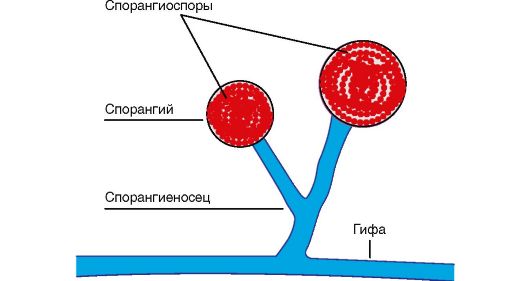 Рис. 2.6.
Грибы рода Mucor
(рис. А.С. Быкова)
Рис. 2.6.
Грибы рода Mucor
(рис. А.С. Быкова)
 Рис. 2.7.
Грибы рода Rhizopus.
Развитие спорангия, спорангиоспор и ризоидов
Рис. 2.7.
Грибы рода Rhizopus.
Развитие спорангия, спорангиоспор и ризоидов
лым путем с помощью бесполых спор - конидий (рис. 2.8, 2.9) и должны быть отнесены по этому признаку к несовершенным грибам. У грибов рода Aspergillus на концах плодоносящих гиф, конидиеносцах, имеются утолщения - стеригмы, фиалиды, на которых образуются цепочки конидий («леечная плесень»).
У грибов рода Penicillium (кистевик) плодоносящая гифа напоминает кисточку, так как из нее (на конидиеносце) образуются утолщения, разветвляющиеся на более мелкие структуры - стеригмы, фиалиды, на которых находятся цепочки конидий. Некоторые виды аспергилл могут вызывать аспергиллезы и афлатоксикозы, пенициллы могут вызывать пенициллиозы.
Представителями аскомицетов являются телеоморфы родов Trichophyton, Microsporum, Histoplasma, Blastomyces, а также дрож-
 Рис. 2.8.
Грибы рода Penicillium.
Цепочки конидий отходят от фиалид
Рис. 2.8.
Грибы рода Penicillium.
Цепочки конидий отходят от фиалид
 Рис. 2.9.
Грибы рода Aspergillus fumigatus.
От фиалид отходят цепочки конидий
Рис. 2.9.
Грибы рода Aspergillus fumigatus.
От фиалид отходят цепочки конидий
Базидиомицеты включают шляпочные грибы. Они имеют септированный мицелий и образуют половые споры - базидиоспоры путем отшнуровывания от базидия - концевой клетки мицелия, гомологичной аску. К базидиомицетам относятся некоторые дрожжи, например телеоморфы Cryptococcus neoformans.
Дейтеромицеты являются несовершенными грибами (Fungi imperfecti, анаморфные грибы, конидиальные грибы). Это условный, формальный таксон грибов, объединяющий грибы, не имеющие полового размножения. Недавно вместо термина «дейтеромицеты» предложен термин «митоспоровые грибы» - грибы, размножающиеся неполовыми спорами, т.е. путем митоза. При установлении факта полового размножения несовершенных грибов их переносят в один из известных типов - Ascomycota или Basidiomycota, присваивая название телеоморфной формы. Дейтеромицеты имеют септированный мицелий, размножаются только путем бесполого формирования конидий. К дейтеромицетам относятся несовершенные дрожжи (дрожжеподобные грибы), например некоторые грибы рода Candida, поражающие кожу, слизистые оболочки и внутренние органы (кандидоз). Они имеют овальную форму, диаметр 2-5 мкм, делятся почкованием, образуют псевдогифы (псевдомицелий) в виде цепочек из удлиненных клеток, иногда образуют гифы. Для Candida albicans характерно образование хламидоспор (рис. 2.10). К дейтеромицетам относят также другие грибы, не имеющие полового способа размножения, относящиеся к родам Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaeа, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria и др.
 Рис. 2.10.
Грибы рода Candida albicans
(рис. А.С. Быкова)
Рис. 2.10.
Грибы рода Candida albicans
(рис. А.С. Быкова)
2.4. Строение и классификация простейших
Простейшие относятся к домену Eukarya, царству животных (Animalia), подцарству Protozoa. Недавно предложено выделить простейшие в ранг царства Protozoa.
Клетка простейших окружена мембраной (пелликулой) - аналогом цитоплазматической мембраны клеток животных. Она имеет ядро с ядерной оболочкой и ядрышком, цитоплазму, содержащую эндоплазматический ретикулум, митохондрии, лизосомы и рибосомы. Размеры простейших колеблются от 2 до 100 мкм. При окраске по Романовскому-Гимзе ядро простейших имеет красный, а цитоплазма - голубой цвет. Простейшие передвигаются с помощью жгутиков, ресничек или псевдоподий, некоторые из них имеют пищеварительные и сократительные (выделительные) вакуоли. Они могут питаться в результате фагоцитоза или образования особых структур. По типу питания они разделяются на гетеротрофы и аутотрофы. Многие простейшие (дизентерийная амеба, лямблии, трихомонады, лейшмании, балантидии) могут расти на питательных средах, содержащих нативные белки и аминокислоты. Для их культивирования используют также культуры клеток, куриные эмбрионы и лабораторных животных.
Простейшие размножаются бесполым путем - двойным или множественным (шизогония) делением, а некоторые и половым путем (спорогония). Одни простейшие размножаются внеклеточно (лямблии), а другие - внутриклеточно (плазмодии, токсоплазма, лейшмании). Жизненный цикл простейших характеризуется стадийностью - образованием стадии трофозоита и стадии цисты. Цисты - покоящиеся стадии, устойчивые к изменению температуры и влажности. Кислотоустойчивостью отличаются цисты Sarcocystis, Cryptosporidium и Isospora.
Ранее простейшие, вызывающие заболевания у человека, были представлены 4 типами 1 (Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). Эти типы недавно реклассифицированы на большее количество, появились новые царства - Protozoa и Chromista (табл. 2.2). В новое царство Chromista (хромовики) вошли некоторые простейшие и грибы (бластоцисты, оомицеты и Rhinosporidium seeberi). Царство Protozoa включает амебы, жгутиконосцы, споровики и реснитчатые. Они подразделены на различные типы, среди которых различают амебы, жгутиконосцы, споровики и реснитчатые.
Таблица 2.2. Представители царств Protozoa и Chromista, имеющие медицинское значение
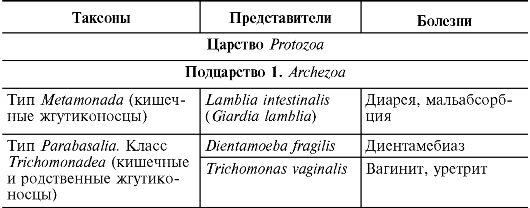 1 Тип Sarcomastigophora
состоял из подтипов Sarcodina
и Mastigophora.
Подтип Sarcodina
(саркодовые) включал дизентерийную амебу, а подтип Mastigophora
(жгутиконосцы) - трипаносомы, лейшмании, лямблию и трихомонады. Тип Apicomplexa
включал класс Sporozoa
(споровики), куда входили плазмодии малярии, токсоплазма, криптоспоридии и др. Тип Ciliophora
включает балантидии, а тип Microspora
- микроспоридии.
1 Тип Sarcomastigophora
состоял из подтипов Sarcodina
и Mastigophora.
Подтип Sarcodina
(саркодовые) включал дизентерийную амебу, а подтип Mastigophora
(жгутиконосцы) - трипаносомы, лейшмании, лямблию и трихомонады. Тип Apicomplexa
включал класс Sporozoa
(споровики), куда входили плазмодии малярии, токсоплазма, криптоспоридии и др. Тип Ciliophora
включает балантидии, а тип Microspora
- микроспоридии.
Окончание табл. 2.2
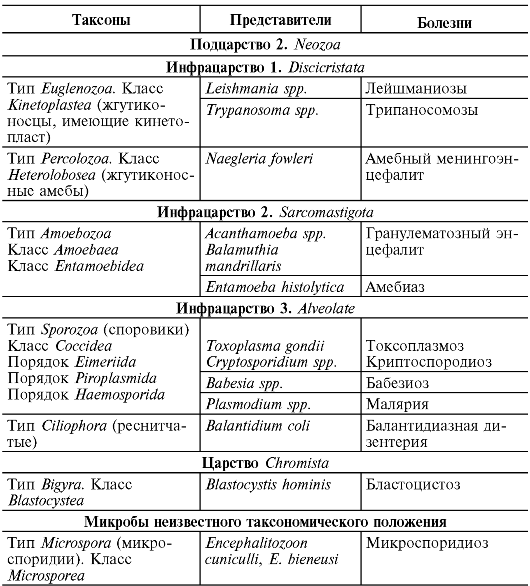 К амебам относятся возбудитель амебиаза человека - амебной дизентерии (Entamoeba histolytica),
свободно
живущие и непатогенные амебы (кишечная амеба и др.). Амебы размножаются
бинарно бесполым путем. Их жизненный цикл состоит из стадии трофозоита
(растущая, подвижная клетка, малоустойчивая) и стадии цисты. Трофозоиты
передвигаются с помощью псевдоподий, которые захватывают и погружают в
цитоплазму питательные вещества. Из
К амебам относятся возбудитель амебиаза человека - амебной дизентерии (Entamoeba histolytica),
свободно
живущие и непатогенные амебы (кишечная амеба и др.). Амебы размножаются
бинарно бесполым путем. Их жизненный цикл состоит из стадии трофозоита
(растущая, подвижная клетка, малоустойчивая) и стадии цисты. Трофозоиты
передвигаются с помощью псевдоподий, которые захватывают и погружают в
цитоплазму питательные вещества. Из
трофозоита образуется циста, устойчивая к внешним факторам. Попав в кишечник, она превращается в трофозоит.
Жгутиконосцы характеризуются наличием жгутиков: у лейшманий один жгутик, у трихомонад 4 свободных жгутика и один жгутик, соединенный с короткой ундулирующей мембраной. Ими являются:
Жгутиконосцы крови и тканей (лейшмании - возбудители лейшманиозов; трипаносомы - возбудители сонной болезни и болезни Шагаса);
Жгутиконосцы кишечника (лямблия - возбудитель лямблиоза);
Жгутиконосцы мочеполового тракта (трихомонада влагалищная - возбудитель трихомоноза).
Реснитчатые представлены балантидиями, которые поражают толстую кишку человека (балантидиазная дизентерия). Балантидии имеют стадию трофозоита и цисты. Трофозоит подвижен, имеет многочисленные реснички, более тонкие и короткие, чем жгутики.
2.5. Строение и классификация вирусов
Вирусы - мельчайшие микробы, относящиеся к царству Virae (от лат. virus - яд). Они не имеют клеточного строения и состоят
Структуру вирусов из-за их малых размеров изучают с помощью электронной микроскопии как вирионов, так и их ультратонких срезов. Размеры вирусов (вирионов) определяют напрямую с помощью электронной микроскопии или косвенно методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Размер вирусов колеблется от 15 до 400 нм (1 нм равен 1/1000 мкм): к маленьким вирусам, размер которых сходен с размером рибосом, относят парвовирусы и вирус полиомиелита, а к наиболее крупным - вирус натуральной оспы (350 нм). Вирусы отличаются по форме вирионов, которые имеют вид палочек (вирус табачной мозаики), пули (вирус бешенства), сферы (вирусы полиомиелита, ВИЧ), нити (филовирусы), сперматозоида (многие бактериофаги).
Вирусы поражают воображение своим разнообразием структуры и свойств. В отличие от клеточных геномов, которые содержат однородную двунитевую ДНК, вирусные геномы чрезвычайно разнообразны. Различают ДНК- и РНК-содержащие вирусы, которые гаплоидны, т.е. имеют один набор генов. Диплоидный геном имеют только ретровирусы. Геном вирусов содержит от 6 до 200 генов и представлен различными видами нуклеиновых кислот: двунитевыми, однонитевыми, линейными, кольцевыми, фрагментированными.
Среди однонитевых РНК-содержащих вирусов различают геномные плюс-нить РНК и минус-нить РНК (полярность РНК). Плюс-нить (позитивная нить) РНК этих вирусов, кроме геномной (наследственной) функции, выполняет функцию информационной, или матричной РНК (иРНК, или мРНК); она является матрицей для белкового синтеза на рибосомах инфицированной клетки. Плюс-нить РНК является инфекционной: при введении в чувствительные клетки она способна вызвать инфекционный про-
цесс. Минус-нить (негативная нить) РНК-содержащих вирусов выполняет только наследственную функцию; для синтеза белка на минус-нити РНК синтезируется комплементарная ей нить. У некоторых вирусов РНК-геном является амбиполярным (ambisense от греч. амби - с обеих сторон, двойная комплементарность), т.е. содержит плюс- и минус-сегменты РНК.
Различают простые вирусы (например, вирус гепатита А) и сложные вирусы (например, вирусы гриппа, герпеса, коронавирусы).
Простые, или безоболочечные, вирусы имеют только нуклеиновую кислоту, связанную с белковой структурой, называемой капсидом (от лат. capsa - футляр). Протеины, связанные с нуклеиновой кислотой, известны как нуклеопротеины, а ассоциация вирусных протеинов капсида вируса с вирусной нуклеиновой кислотой названа нуклеокапсидом. Некоторые простые вирусы могут формировать кристаллы (например, вирус ящура).
Капсид включает повторяющиеся морфологические субъединицы - капсомеры, скомпонованные из нескольких полипептидов. Нуклеиновая кислота вириона, связываясь с капсидом, образует нуклеокапсид. Капсид защищает нуклеиновую кислоту от деградации. У простых вирусов капсид участвует в прикреплении (адсорбции) к клетке хозяина. Простые вирусы выходят из клетки в результате ее разрушения (лизиса).
Сложные, или оболочечные, вирусы (рис. 2.11), кроме капсида, имеют мембранную двойную липопротеиновую оболочку (синоним: суперкапсид, или пеплос), которая приобретается путем почкования вириона через мембрану клетки, например через плазматическую мембрану, мембрану ядра или мембрану эндоплазматического ретикулума. На оболочке вируса расположены гликопротеиновые шипы,
или шипики, пепломеры. Разрушение оболочки эфиром и другими растворителями инактивирует сложные вирусы. Под оболочкой некоторых вирусов находится матриксный белок (М-белок).
Вирионы имеют спиральный, икосаэдрический (кубический) или сложный тип симметрии капсида (нуклеокапсида). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вирусов гриппа, коронавирусов): капсомеры уложены по спирали вместе с нуклеиновой кислотой. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вируса герпеса).
Капсид и оболочка (суперкапсид) защищают вирионы от воздействия окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) своими рецепторными белками с опреде-
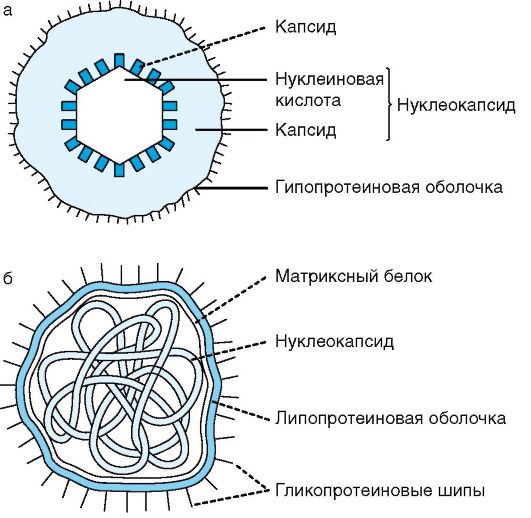 Рис. 2.11.
Строение оболочечных вирусов с икосаэдрическим (а) и спиральным (б) капсидом
Рис. 2.11.
Строение оболочечных вирусов с икосаэдрическим (а) и спиральным (б) капсидом
ленными клетками, а также антигенные и иммуногенные свойства вирионов.
Внутренние структуры вирусов называют сердцевиной. У аденовирусов сердцевина состоит из гистоноподобных белков, связанных с ДНК, у реовирусов - из белков внутреннего капсида.
Лауреат Нобелевской премии Д. Балтимор предложил систему балтиморской классификации, основанной на механизме синтеза мРНК. Эта классификация размещает вирусы в 7 группах (табл. 2.3). Международный комитет на таксономии вирусов (ICTV) принял универсальную систему классификации, которая использует такие таксономические категории, как семейство (название оканчивается на viridae), подсемейство (название оканчивается на virinae), род (название оканчивается на virus). Вид вируса не получил биноминального названия, как у бактерий. Вирусы классифицируют по типу нуклеиновой кислоты (ДНК или РНК), ее структуре и количеству нитей. Они имеют двунитевые или однонитевые нуклеиновые кислоты; позитивную (+), негативную (-) полярность нуклеиновой кислоты или смешанную полярность нуклеиновой кислоты, амбиполярную (+, -); линейную или циркулярную нуклеиновую кислоту; фрагментированную или нефрагментированную нуклеиновую кислоту. Учитывают также размер и морфологию вирионов, количество капсомеров и тип симметрии нуклеокапсида, наличие оболочки (суперкапсида), чувствительность к эфиру и дезоксихолату, место размножения в клетке, антигенные свойства и др.
Таблица 2.3. Основные вирусы, имеющие медицинское значение
 Продолжение табл. 2.3
Продолжение табл. 2.3
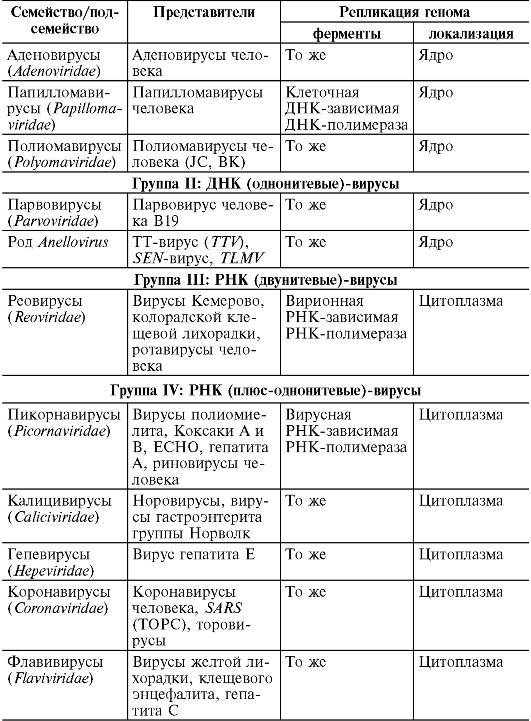 Окончание табл. 2.3
Окончание табл. 2.3
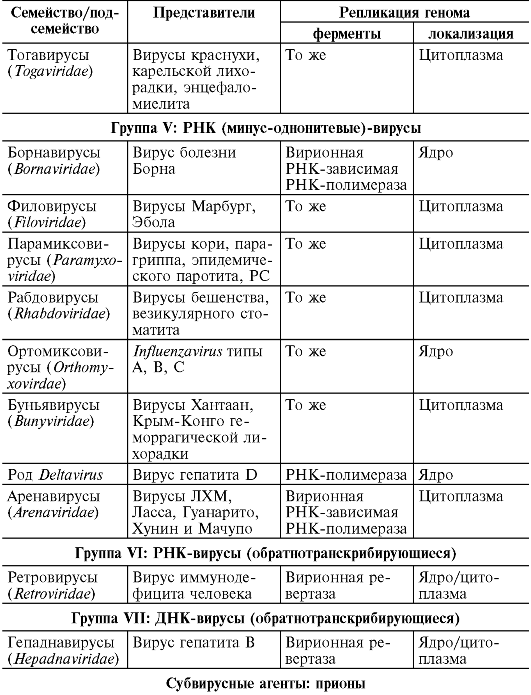 Вирусы
поражают животных, бактерии, грибы и растения. Являясь основными
возбудителями инфекционных заболеваний человека, вирусы также участвуют в
процессах канцерогенеза, могут передаваться различными путями, в том
числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая
плод человека. Они могут приводить и к постинфекционным осложнениям -
развитию миокардитов, панкреатитов, иммунодефицитов и др.
Вирусы
поражают животных, бактерии, грибы и растения. Являясь основными
возбудителями инфекционных заболеваний человека, вирусы также участвуют в
процессах канцерогенеза, могут передаваться различными путями, в том
числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая
плод человека. Они могут приводить и к постинфекционным осложнениям -
развитию миокардитов, панкреатитов, иммунодефицитов и др.
К неклеточным формам жизни, кроме вирусов, относят прионы и вироиды. Вироиды - небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белок и вызывающие заболевания растений. Патологические прионы - инфекционные белковые частицы, вызывающие особые конформационные болезни в результате изменения структуры нормального клеточного прионового протеина (PrP c ), который имеется в организме животных и человека. PrP с выполняет регуляторные функции. Его кодирует нормальный прионовый ген (PrP-ген), расположенный в коротком плече 20-й хромосомы человека. Прионные болезни протекают по типу трансмиссивных губкообразных энцефалопатий (болезнь Крейтцфельда-Якоба, куру и др.). При этом прионный протеин приобретает другую, инфекционную форму, обозначаемую как PrP sc (sc от scrapie - скрепи - прионная инфекция овец и коз). Этот инфекционный прионный протеин имеет вид фибрилл и отличается от нормального прионного протеина третичной или четвертичной структурой.
Задания для самоподготовки (самоконтроля)
А. Отметьте микробы, являющиеся прокариотами:
2. Вирусы.
3. Бактерии.
4. Прионы.
Б. Отметьте отличительные особенности прокариотической клетки:
1. Рибосомы 70S.
2. Наличие пептидогликана в клеточной стенке.
3. Наличие митохондрий.
4. Диплоидный набор генов.
В. Отметьте составные компоненты пептидогликана:
1. Тейхоевые кислоты.
2. N-ацетилглюкозоамин.
3. Липополисарид.
4. Тетрапептид.
Г. Отметьте особенности строения клеточной стенки грамотрицательных бактерий:
1. Мезодиаминопимелиновая кислота.
2. Тейхоевые кислоты.
4. Белки-порины.
Д. Назовите функции спор у бактерий:
1. Сохранение вида.
2. Жароустойчивость.
3. Расселение субстрата.
4. Размножение.
1. Риккетсии.
2. Актиномицеты.
3. Спирохеты.
4. Хламидии.
Ж. Назовите особенности актиномицет:
1. Имеют термолабильные споры.
2. Грамположительные бактерии.
3. Отсутствует клеточная стенка.
4. Имеют извитую форму.
З. Назовите особенности спирохет:
1. Грамотрицательные бактерии.
2. Имеют двигательный фибриллярный аппарат.
3. Имеют извитую форму.
И. Назовите простейшие, обладающие апикальным комплексом, позволяющим проникать внутрь клетки:
1. Малярийный плазмодий.
3. Токсоплазма.
4. Криптоспоридии.
К. Назовите отличительную особенность сложноорганизованных вирусов:
1. Два типа нуклеиновой кислоты.
2. Наличие липидной оболочки.
3. Двойной капсид.
4. Наличие неструктурных белков. Л. Отметьте высшие грибы:
1. Mucor.
2. Candida.
3. Penicillium.
4. Aspergillus.
Бактерии — различаемые только в микроскоп микроорганизмы, в большинстве одноклеточные и подвижные, благодаря наличию жгутиков.
Форма клетки различна у разных видов. Чаще встречаются бактерии, которые можно отнести к следующим формам: шаровидным - кокки; палочковидным прямым - бациллы и бактерии; палочковидным согнутым - вибрионы; спирально закрученным - спириллы. Клетка бактерии состоит из оболочки, заключающей протоплазму, а также иногда вакуоли. При соответствующей окраске в протоплазме можно обнаружить тельца, напоминающие ядра, по, возможно, и не соответствующие клеточным ядрам других организмов. Большинство бактерий бесцветно, в некоторых имеются красный и зелёный пигменты.
Многие бактерии, особенно палочковидные и некоторые шаровидные, образуют споры (эндоспоры), развивающиеся в уплотняющейся части протоплазменного содержимого и снабжённые на своей поверхности плотной, трудно проницаемой оболочкой. Эта оболочка придаёт спорам большую стойкость к неблагоприятным внешним условиям, и они очень долго сохраняют свою жизнеспособность. При благоприятные условиях споры растут, развиваясь в новую вегетативную клетку.
Размножаются бактерии поперечным делением клетки на две, причём интервал между двумя делениями нередко составляет всего полчаса, почему по быстроте размножения они далеко превосходят все другие организмы.
Большинство бактерий имеет гетеротрофное питание, т. е. покрывают свою потребность за счёт готового органического вещества, и лишь немногим свойственно автотрофное питание за счёт углерода, находящегося в углекислоте. Что касается азотного питания, то для некоторых из этих микробов необходим органический азот, большинство же может довольствоваться аммонийным и нитратным азотом, а некоторые способны усваивать элементарный азот.
Большинство бактерий это аэробные организмы, некоторые могут развиваться только при достаточном содержании кислорода в окружающей среде, но известны и анаэробные бактерии; им не нужен кислород. Они чрезвычайно распространены, особенно там, где есть органические вещества, которые они разрушают, образуя различные виды гниения или брожения. Многие микробы паразитируют в растительных или животных организмах. Многие являются возбудителями инфекционных заболеваний, например, туберкулёза, брюшного тифа, холеры, чумы, дифтерита. Другие обеспечивают очень важные в жизни человека процессы (различные виды брожения, накопление азота в почве клубеньковыми бактериями), не говоря уже о том, что без их участия немыслим процесс почвообразования и кругооборот веществ в природе. Некоторые микроорганизмы имеют непосредственное значение в медицине, например, те, из которых получаются сильные
Бактерии отличают по таким признакам:
- морфологическим;
- тинкториальным;
- физиологическим;
- культуральным;
- биохимическим;
- антигенным;
- молекулярно-генетическим.
Морфологические признаки
Бактерии – преимущественно . Структура клетки прокариот значительно отличается от строения клеток грибов, растений, животных. Они не имеют ядерной оболочки, митохондрий, аппарата Гольджи. В бактериальной клетке различают такие элементы:
- поверхностные структуры;
- клеточную оболочку;
- цитоплазму с нуклеотидом.
Поверхностными структурами называют образования, расположенные снаружи от цитоплазматической мембраны. К ним относят капсулу, слизистый слой, чехол, клеточную стенку, жгутики, ворсинки.
Капсула, чехол или слизистый слой необходимы для защиты бактерии от влияния окружающей среды. Они предохраняют от различных химических веществ, защищают от высыхания, служат препятствием для проникновения фагов. Бактерии, обитающие в желудке, защищены специальным слизистым веществом, предохраняющим клетку от воздействия хлорной кислоты, ферментов желудочного сока.
Жгутики определяют способность к передвижению. Их количество, расположение постоянно, поэтому они учитываются при систематизации прокариот.
Они могут находиться у полюсов или в полярной области бактериальной клетки. По наличию, местоположению подразделяют на:
- монотрихи – один жгутик расположен на полюсе клетки (холерный вибрион);
- лофотрихи – пучок из жгутиков расположен с одной стороны (синегнойная палочка);
- амфитрихи – пучки жгутиков находятся на разных полюсах бактериальной клетки (спириллы);
- перитрихи – жгутики распределены по всей поверхности клетки (сальмонеллы).
Клеточная стенка располагается непосредственно под капсулой, слизистым слоем или чехлом.
Внутри бактериальная клетка заполнена цитоплазмой, органеллами и нуклеотидом. От других прокариот, к которым относят царство Цианобактерий, они отличаются отсутствием хлорофилла.
Бактерии различают по форме. Они могут быть сферические (кокки, диплококки, стрепто-, стафилококки), цилиндрические (палочки), спиральные (спирохеты, спириллы, вибрионы).
Тинкториальные признаки (отношение к специальным красителям)
Метод Циля-Нельсона используют для выявления кислото- и спиртоустойчивых микобактерий туберкулеза, возбудителей лепры, бактерий, способствующих развитию язвы желудка.
Метод серебрения по Морозову используют для выделения спирохет, вирусных включений.
Окраска по Романовскому-Гимзе применяется для определения бактерий, грибов, простейших. Для проведения окраски используется сложный краситель, который окрашивает различные типы микроорганизмов в разные цвета.
Физиологические признаки
Физиологические признаки включают:
- биохимические;
- культуральные.
Биохимические признаки
Виды бактерий различают по биохимическим параметрам (по выделяемым ферментам, питанию, дыханию). Например, хеликобактер пилори, обитающие в желудке, способны выделять секрет, стимулирующий появление язв. Поэтому при появлении симптомов язвы назначают бактериальное исследование. При болезнях, вызываемых инфекцией, особенно у детей, возникает отравление токсинами, которые выделяют микроорганизмы. Ферменты бактерий добавляют в некоторые стиральные порошки. Используют для получения соединений уксусной, молочной кислот.
Особое значение при определении инфекции имеет тип дыхания. Различают:
- Облигатные анаэробные бактерии. Кислород для них токсичен. Для дыхания им необходимы нитраты, сульфаты или фумараты. Они обитают в безвоздушном пространстве. Поражают желудок, кишечник, мочеполовую систему. Особо опасными являются инфекции, вызывающие ботулизм, столбняк, газовую гангрену.
- Облигатные аэробы. Могут существовать только при наличии кислорода. Соответственно, являются возбудителями заболеваний дыхательной системы, кожных болезней у взрослых и детей.
- Факультативные анаэробы. К ним относятся некоторые виды бактерий и грибов, которым для роста и размножения наличие кислорода необязательно.
По типам питания бактерии делят на:
- автотрофы;
- гетеротрофы.
Некоторые бактерии составляют нормальную микрофлору организма человека. В таком случае болезни могут вызваться уменьшением количества полезных бактерий. Так, прием антибиотиков уменьшает количество кишечной палочки, в результате возникают расстройства желудка или кишечника. Сапрофиты не дают развиваться патогенным микроорганизмам, при уменьшении количества бактерий могут начинаться болезни, вызванные патогенными видами грибов.
Культуральные признаки
В зависимости от типов питания, среды обитания, бактерии отличают по культуральным признакам.
Чтобы выявить, к какому типу относятся микроорганизмы, производят посев. Для проведения бактериального исследования могут использовать не только частички кожи, слюну, мокроту, но и содержимое желудка.
Например, при симптомах ангины у детей и взрослых обязательно проводят микробиологическое обследование. Стерильной палочкой с ваткой снимают белый налет на гортани. Высаживают в специальные чашки Петри. Затем смотрят, на какой питательной среде лучше всего микроорганизмы размножились, их расположение (раскиданные одиночные точки или сосредоточенные в одном месте). Цвет, размер бактериальной колонии тоже имеют огромное значение.
Выявив тип инфекции, вызвавшей заболевание у детей и взрослых, проводят анализ на устойчивость штамма к антибиотикам. Потом назначают соответствующее лечение.
Антигенные признаки
Определяют вид бактериальной культуры, взяв анализ крови. Ведь при заболевании, вызванном инфекцией, наш организм выделяет антитела к возбудителю. Выявив их наличие, устанавливают штамм микроорганизмов, вызвавших болезнь.
Для определения типа инфекции, которая вызвала болезнь, проводят исследование сыворотки крови на наличие антител или антигена – серологические реакции. Их используют:
- для , токсинов, антигена с помощью иммунодиагностической сыворотки;
- для определения антитела с помощью известного антигена.
Серологический анализ используют при подозрении на скрытую форму болезни. Обследуют людей, которые контактировали с инфицированным, для выявления болезни на ранних сроках.
Иногда человек может сам не болеть, но стать переносчиком. Или же болезнь, вызванная инфекцией, протекает, не проявляя симптомов, носит хронический характер.
Для диагностирования туберкулеза детям делают кожную пробу – реакция Манту. Для ее проведения используют сухой туберкулин (экстракт бактериальных остатков). Покраснение, отечность кожи наблюдаются, если в организме ребенка есть антитела к палочке Коха. Они вырабатываются только в случае наличия инфекции.
Молекулярно-генетические признаки
В основу дифференциации микроорганизмов по генетическим признакам положено принципиальное различие строения генов. Так, у вирусов и фагов наследственный материал заключен в 1 молекуле ДНК или РНК. У простейших и грибов хромосомы расположены в клеточном ядре. Генетический аппарат бактерии отличается тем, что:
- Хромосома размещается в цитоплазме. Она состоит из 3000 генов.
- Содержимое ДНК изменяется, она может увеличиваться. Эта уникальная возможность регулирует скорость размножения, что положительно сказывается на выживаемости видов.
- Наследственная информация содержится в генах хромосомы и плазмидах.
- Передача информации происходит по вертикали (от родительской клетки к дочерней), по горизонтали (бактерии способны обмениваться генами друг с другом). Таким образом, от одной бактериальной клетки к другой передаются мутационные гены. Например, у одного произошла положительная генная мутация, обеспечившая инфекцию устойчивостью к антибиотику. При обмене информацией с другими бактериями другая колония получит такую же способность.
При проявлении симптомов болезни, если есть подозрение, что она вызвана бактериями, следует обязательно провести исследование, направленное на дифференциацию возбудителей заболевания, определение резистентности к антибиотикам.
При выявленном инфекционном характере болезни необходимо помнить, что бактерии могут вызвать эпидемии. Из-за слабого иммунитета инфекционным болезням чаще всего подвержены дети. Чтобы не допустить распространение болезни, предпринимают ряд мер:
- При возникновении нескольких случаев болезней в школах или садиках, чтобы не подвергать других детей опасности, учреждения закрывают на карантин.
- Для профилактики серьезных заболеваний, которые могут привести к летальному исходу, рекомендуют делать детям прививки.
- Во время вспышки заболевания не рекомендуют посещать общественные мероприятия.
- Детям и взрослым, особенно во время сезонных заболеваний, необходимо принимать иммуностимулирующие препараты.
При диагностике болезней, вызванных инфекцией, необходимо учитывать основные . Они различаются между собой строением, формой клетки, типами питания и дыхания. Проводя микробиологическое исследование, необходимо провести окраску по Граму, сделать посев на питательные среды (определяют культуральные признаки). Иногда назначают серологические исследование (анализ сыворотки крови определяет наличие антигена или антител). Также проводят молекулярно-генетическое исследование, которое основано на отличиях строения и месторасположения нуклеотида.