Rozmnażanie porostów tak idiosynkratyczny jak cała ich organizacja. Każdy ze składników plechy jest w stanie rozmnażać się indywidualnie, np. komórki glonów dzielą się od czasu do czasu, jednak późniejszy rozwój nowej plechy jest możliwy tylko wtedy, gdy grzyb i glony ponownie się spotkają. Dlatego bardziej obiecująca (przynajmniej dla mykobionta) będzie taka metoda rozmnażania, w której struktury generatywne zawierają oba składniki.
Wśród żywych organizmów można spotkać dwie formy rozmnażania - płciową i bezpłciową. Różnorodność genetyczna zachodzi poprzez mieszanie genów dwóch osobników poprzez rozmnażanie płciowe, podczas gdy w rozmnażaniu bezpłciowym takie mieszanie genów nie występuje. Wśród porostów można spotkać rozmnażanie płciowe i bezpłciowe. Mówiąc o roślinach, rozmnażanie bezpłciowe jest zwykle określane jako rozmnażanie wegetatywne. Chociaż porosty ogólnie mogą rozmnażać się zarówno płciowo, jak i wegetatywnie, istnieją gatunki, w których mogą rozprzestrzeniać się oba rodzaje rozmnażania, a także gatunki, w których jeden typ jest rzadki lub nawet nieznany.
Większość porostów rozmnaża się głównie wegetatywnie, a dla wielu z nich ta metoda rozmnażania pozostaje jedyną. Wysychając, plecha staje się bardzo krucha i łatwo odrywają się od niej kawałki, które następnie są podnoszone przez wiatr i rozprzestrzeniane. Dzieje się tak z wieloma porostami tundrowymi.
Rola porostów w przyrodzie i ich znaczenie gospodarcze
Jeśli chodzi o rozmnażanie porostów, pojawiają się dwa podstawowe pytania. Co to znaczy mówić o rozmnażaniu płciowym u porostów? Kolejne dwie części poświęcone będą tym zagadnieniom. Przecież porostów nie ma odrębny organizm i jest związkiem między dwoma organizmami. W rzeczywistości u porostów tylko partnerzy grzybowi mogą rozmnażać się płciowo. Stwierdzenie, że porosty mogą rozmnażać się płciowo, jest rzeczywiście skrótem myślowym, ponieważ „partnerzy grzybów w porostach mogą rozmnażać się płciowo”.
Soredia i isidia to wyspecjalizowane struktury rozmnażania wegetatywnego, z których każda zawiera elementy mykobiontu i fikobiontu. Soredia to mikroskopijne grudki komórek glonów otoczone strzępkami grzybów (). Nagromadzenie soredii (nazywane są soralami) przypomina sproszkowaną masę, która pokrywa wierzchołek plechy lub otacza plechę wzdłuż zewnętrznego obwodu. Powstawanie sorediów jest możliwe tylko w warstwie plechy zawierającej glony (warstwa gonidiów), a poprzez pęknięcia w górnej warstwie skorupy soredia wychodzą na zewnątrz, tworząc skupiska - sorale. Soredia rozmnażają się głównie porosty wysoko zorganizowane (liściaste i krzaczaste), które zwykle nie tworzą owocników.
Rozmnażanie bezpłciowe porostów
Wiele gatunków fotobiontów występujących w porostach można znaleźć jako wolno żyjące, a następnie rozmnażające się płciowo, ale rozmnażanie płciowe fotobiontów jest tłumione w porostach. Porosty rozmnażające się płciowo to ascomycetes lub basidiomycetes. Często askospory lub bazydiospory są po prostu określane jako zarodniki, zwłaszcza tam, gdzie nie ma potrzeby ich rozróżniania lub gdy nie ma możliwości pomylenia ich z rozmnażaniem wegetatywnym.
Najczęstszymi strukturami tworzącymi worki są aptekarze, zwykle przypominające dyski, przypominające miseczki, które wyrastają z powierzchni wzgórza. Wiele wiadomo o procesach rozmnażania płciowego u niezlichenizowanych ascomycetes lub basidiomycetes, ponieważ wiele z nich można hodować w ich laboratorium, można monitorować wszystkie etapy ich rozwoju. cykle życia i przeprowadzać eksperymenty krycia. Dzięki temu można było zobaczyć, jak przebiegają procesy rozmnażania płciowego u tych gatunków.
Isidia są również charakterystyczne dla form liściastych i krzaczastych, ale powstają rzadziej niż soredia. Morfologicznie, izydia są małymi wyrostkami na Górna powierzchnia thallus, wewnątrz którego znajdują się komórki glonów otoczone strzępkami. Na zewnątrz izydia pokryte są warstwą skorupy. Wygląd izydii może być bardzo różnorodny, ale dla każdego rodzaju porostów specyficzna jest pewna forma.
Takie badania wykazały, że rozmnażanie płciowe u niezlichenizowanych Ascomycetes lub Basidiomycetes ma podobieństwa i znaczące różnice z organizmami takimi jak ludzie czy rośliny. Nie udało się wyjaśnić większości procesów zachodzących w mykobiontach porostów. Niektóre z nich można do pewnego stopnia hodować w laboratorium, ale nie w takim stopniu, który pozwalałby na dobre zrozumienie wszystkich procesów rozmnażania płciowego. W tym kontekście warto zauważyć, że wśród niezlichenizowanych workowców lub podstawczaków badanie procesów u gatunków mikoryzowych jest trudniejsze niż u gatunków saprotroficznych.
rozmnażanie bezpłciowe porosty każdy ze składników występuje niezależnie. Mykobiont może tworzyć konidia, piknoconidia i stylospory, które tworzą się egzogennie. Konidia, które rozmnażają wiele wolnożyjących grzybów, bardzo rzadko powstają w porostach w przyrodzie, znacznie częściej można je zaobserwować podczas sztucznej uprawy mykobiontu. Pycnoconidia to bardzo małe komórki (około 1 µm szerokości i około 5 µm długości) różne kształty, który w duża liczba tworzą się w zamkniętych okrągłych pojemnikach z otworem w górnej części, zwanych pyknidiami (gr. pyknos - gęsty). Zwykle rozwijają się na górnej powierzchni plechy lub wzdłuż jej krawędzi. Stylospory rozwijają się również w piknidiach (tylko rzadziej), ale są duże (do 100 µm długości) i składają się z dwóch lub więcej komórek.
Wiesz, że porosty to skojarzenia grzybów z fotobiontami. Uprośćmy na razie sprawę i załóżmy, że mamy plechę, która zawiera tylko jeden gatunek fotobiontu i jeden gatunki grzybów. Odpowiedź na poprzednie fundamentalne pytanie pokazała, że rozmnażanie płciowe u porostów wiąże się zatem tylko z partnerem grzybowym ważne pytanie: Czy porosty plechowe mają tylko jednego człowieka-grzyba? Odpowiedź: Czasami tak, czasami nie.
Na tym etapie przydatne jest wprowadzenie słowa grzybnia, podstawowego pojęcia dla grzybów nielichenizowanych. Kiedy ascomycete lub basidiomycete rozprzestrzeniają się, zwykle dzieje się to poprzez wzrost strzępek, które poprzez ponowne rozszerzenie i rozgałęzienie utworzą sieć strzępek, a sieć ta nazywa się grzybnią. U grzybów nielichenizowanych grzybnia ta rozprzestrzenia się poprzez glebę, obornik lub drewno, nabywając składniki odżywcze od podłoża, ale pozostając poza zasięgiem wzroku. W grzybach zlichenizowanych grzybnia jest związana z gatunkami fotobiontów i jest zawarta w porostach, które są zwykle dobrze widoczne, chociaż talki niektórych gatunków czaszek mogą rosnąć w pęknięciach drewna lub skały.
rozmnażanie płciowe Charakterystyka mykobiont porostowy towarzyszy temu powstawanie owocników. W zależności od pozycji systematycznej mykobionta wyróżnia się porosty podstawkowe i torbacze. Porosty podstawne mają grzyby podstawkowe jako mykobionty. Ta bardzo mała grupa (tylko około 20 gatunków) ma niezależne pochodzenie i występuje głównie w tropikach, chociaż niektóre z nich żyją w klimacie umiarkowanym, a nawet w tundrze. Ich podstawki tworzą się na spodniej stronie plechy w otwartej warstwie generatywnej, gdzie podstawki przeplatane są jałowymi strzępkami - parafizami. Owocniki nietropikalnych porostów podstawnych przypominają miniaturowe grzyby kapeluszowe. Rozwój bazydiospor jest pod wieloma względami podobny do procesu sporulacji płciowej u niezależnych grzybów podstawkowych, który jest szczegółowo opisany w części niniejszego podręcznika poświęconej tym grzybom.
Załóżmy, że propagandę grzybów wystrzelono z krainy porostów obok żywych alg. Nie ma znaczenia, czy propaganda jest produkowana seksualnie, czy wegetatywnie. Załóżmy dalej, że kiełkowanie kiełkuje, dając początek grzybni, która zatrzymuje niektóre pobliskie glony iw ten sposób rozwija nowy porost. Załóżmy teraz, że para genetycznie różnych hodowli grzybów tego samego gatunku wylądowała blisko siebie iw pobliżu wolnych żywych alg.
Załóżmy, że oba wyrostki grzybów wykiełkują, przechwytują niektóre komórki alg, zamieniają się w grzybnię i tym samym dają początek dwóm plechom, ale w tym przypadku bliskość dwóch reprodukcji skutkuje połączeniem dwóch plech. Po fuzji gołym okiem widać tylko jedną plechę, ale wewnątrz tej plechy znajdują się dwa osobniki grzyba – tego samego gatunku, ale różniące się genetycznie. Załóżmy, że kilka zarodników w oktecie kiełkuje i wychwytuje komórki fotobiontów, tworząc nową plechę.
Większość porostów zawiera torbacze, więc są one łączone w grupę porostów torbaczy. Podczas sporulacji płciowej tworzą również owocniki, których rozwój w Różne formy dzieje się inaczej. W porostach ascolocular najpierw tworzy się zrąb, składający się ze splecionych strzępek, po czym tworzą się w nim żeńskie narządy płciowe, archikarpy. Rozwojowi woreczków z zarodnikami towarzyszy tworzenie się ubytków w zrębie wokół torebek - locules.
Nowy tal będzie składał się z genetycznie odrębnych osobników grzybów, jeśli same zarodniki są genetycznie odrębne. Nacisk położono na grzyby, ponieważ partnerzy grzybów mogą rozmnażać się płciowo. Warto jednak zauważyć, że gdy pojedyncza populacja grzybów rozprzestrzenia się wokół wolno żyjących fotobiontów i chwyta je, tworząc nowy porost, fotobionty mogą być również zróżnicowane genetycznie. Dlatego pojedyncza plecha porostu, nawet jeśli zawiera tylko jeden gatunek fotobionta, może nadal wykazywać różnorodność genetyczną wśród komórek fotobiontów.
U większości porostów torbaczy rozwój archikarpu rozpoczyna się na granicy rdzenia i warstwy glonów (warstwa błony dziewiczej), dlatego w owocnikach zawsze występuje warstwa błony dziewiczej. Proces zapłodnienia porostów nie został wystarczająco zbadany, wielu badaczy uważa, że \u200b\u200bowocnik rozwija się z niezapłodnionego askogonu.
O porostach zwykle zapomina się, patrząc na różnorodność gatunków na danym obszarze. Zwykle są ignorowane i na pewno źle rozumiane. Nadszedł czas, abyśmy wszyscy wyjęli lupy i dowiedzieli się o nich trochę więcej. Fotobiont wytwarza węglowodany w procesie fotosyntezy i dzieli się nimi z grzybem. To małżeństwo między dwojgiem całkowicie różne organizmy działa wyjątkowo dobrze.
Porosty występują w różnych formach wzrostu. Kształt porostów może pomóc w identyfikacji. Krzewiasta lub fruktozowata forma wzrostu jest przyczepiona do podłoża tylko w jednym miejscu. On nabywa wygląd miniaturowy krzew lub wahadło i ma kształt cylindryczny lub barkowy. Forma foliowa plechy przypomina kształtem liście, wyrasta w płatkach i jest luźno przyczepiona do podłoża za pomocą ryzin, które wyglądają jak korzenie, ale nie są nimi. Chociaż typy plech opisane powyżej można łatwo usunąć z podłoża, nie dotyczy to porostów prążkowanych, do których jest przyczepiona plecha.
W porostach torbaczy wyróżnia się następujące główne typy owocników: apothecia, gastrothecia i perithecia.
Apotecja to owocniki typu otwartego, które są najbardziej rozpowszechnione wśród porostów. Są małe (zwykle 1 - 2 mm, rzadko większe lub mniejsze) zaokrąglone lub owalny kształt zlokalizowane na górnej (bardzo rzadko, tylko u nephroma i nephromopsis, na dolnej) powierzchni wzgórza. Apotecjum składa się ze spłaszczonej części środkowej - krążka i wałka otaczającego ten krążek wzdłuż obwodu. funkcja reprodukcyjna apotecja posiada dysk, Górna warstwa która (warstwa błony dziewiczej) jest utworzona przez liczne pionowo zorientowane worki i sterylne strzępki (parafizy). Górne końce parafizy mają zgrubienia w kształcie maczugi, które zamykając się, tworzą warstwę ochronną nad workami (leżą nieco niżej) - nabłonek. Kolor apotecji zależy od nabłonka. Pod warstwą błony dziewiczej znajduje się warstwa gęsto splecionych strzępek - hipotecja, w której rozwijają się strzępki workowate i znajdują się podstawy przynasad.
Część podłoża odpadłaby, gdybyś próbował usunąć porosty. Niezwykłą formą są porosty w kształcie stożka. Mają plechę pierwotną, która składa się z drobnych struktur łuskowych, z których wyrastają plechy wtórne. Rozmnażanie grzybów jest takie samo, jakie znamy ze świata grzybów. W przypadku grzybów lichenizowanych zdecydowana większość to grzyby workowate, nazwane tak od sposobu, w jaki wytwarzają swoje zarodniki, niewielka liczba grzybów lichenizowanych to grzyby klubowe.
Tutaj zarodniki grzybów musiałyby znaleźć odpowiednie algi lub sinice, aby stworzyć nowe porosty, zamiast najprostszej metody osiągnięcia pomyślnych wyników. Pod mikroskopem lub dobrym szkłem powiększającym Isidia wygląda jak małe wypukłości wychodzące z plechy, podczas gdy soradia wychodzi z plechy lub wzdłuż krawędzi plechy.
Rolka otaczająca tarczę z boków i od dołu pełni funkcję ochronną. W zależności od budowy grzbietu wyróżnia się kilka rodzajów apotecji: porosty lecanor, lecideus i biator.
Apotecja Lecanora charakteryzujące się tym, że ich tarcza jest otoczona rolką podobną w Struktura wewnętrzna i ubarwienie plechą, a ubarwienie grzbietu i krążka jest zawsze inne. Grzbiet (podobnie jak plecha) zawiera warstwę skorupy, warstwę alg i luźniejszy rdzeń. Nazwa tego typu owocników pochodzi od łacińskiej nazwy rodzaju Lecanor.
Tylko jedna z izydii lub soradii powinna pęknąć i znaleźć odpowiednie warunki do powstania nowego porostu. Porosty potrzebują światła i wilgoci oraz podłoża, które pozostaje wystarczająco długo, aby plecha mogła rosnąć. Często spotykamy porosty na liniach ogrodzeń i dachówkach domów, ale ich miejsce w naturze jest na pniach i gałęziach drzew, na skałach, na ziemi.
Pozwala to naświetlić ich strukturę, sposób formowania się plechy, wygląd faktury powierzchni, sposób rozmnażania. Kolor zmienia się wraz z wilgotnością i dlatego nie jest niezawodny. Tak zwany test na miejscu, w którym małe ilości niektórych substancje chemiczne widoczne na wzgórzu lub mózgu, co może wywołać reakcję barwną. W zależności od wyniku można zidentyfikować porosty. Jeśli te testy są niewystarczające, można je rozpocząć dodatkowe procedury, takich jak chromatografia cienkowarstwowa, w której ekstrakty z porostów są przenoszone na płytkę i poddawane serii zabiegów mających na celu identyfikację ich składników chemicznych.
Lecidea apotecia posiadają wałek pomalowany w taki sam sposób jak dysk (najczęściej na czarno), natomiast charakteryzują się dużą twardością. Struktura grzbietu różni się od plechy głównej; tworzą go ściśle przylegające do siebie ciemno zabarwione strzępki, które razem tworzą wyrostek otaczający krążek ze wszystkich stron. Lecideal apotecia są utworzone wyłącznie przez strzępki grzybów i nie zawierają glonów. Nazwa tego typu owocników pochodzi od łacińskiej nazwy rodzaju Lecidea.
Porosty przeżywają równie ciężko jak pionierzy. Występują niemal wszędzie, od pustyni po Antarktydę. Czy wiesz, że na Antarktydzie znanych jest 350 porostów, ale tylko dwie rośliny kwitnące. Porosty wróciły na ziemię po 6 dniach i dalej funkcjonowały. Porosty zawierają kwasy, rozbijają skały i przez bardzo długi czas przyczyniają się do powstawania gleby. Porosty są również pierwszymi, które tworzą tak zwane skorupy glebowe w miejscach odsłonięcia gleby. Czynią to wraz z rogami rogowymi, wątrobowymi, mchami i sinicami oraz zapewniają ochronę i ochronę przed erozją.
Porosty biatoryczne pod wieloma względami podobne do lecideaceae, ale w przeciwieństwie do nich mają bardziej miękką teksturę, ponadto są pomalowane na jasne kolory i nigdy nie są czarne.
Gasterothia to również owocniki otwarte, ale bardzo rzadkie (tylko u dwóch rodzajów porostów łuskowatych: opegraph i sęp). Ich dysk, położony głęboko, wygląda jak wąski pasek. Wokół dysku wznosi się margines, który może należeć do plechy lub samej gasterotecji.
Jest to szczególnie ważne w obszarach suchych, takich jak Australia Południowa, ale można je łatwo zaobserwować w naszym obszarze zlewni. Wodochłonność porostów wynosi od 3 do 35% ich masy. Woda jest zwykle pobierana, gdy pada deszcz. Niektóre porosty pochłaniają tylko mgłę i mgłę.
Chociaż wiadomo, że bezkręgowce używają porostów jako kamuflażu, a ptaki używają materiału z porostów do następnego budynku, niewiele wiadomo, że porosty są ważne źródło pokarm nie tylko dla bezkręgowców, ale także dla małych ssaków.
W wielu krajach porosty są wykorzystywane jako bioindykatory. Większość krajów, przynajmniej na półkuli północnej, wykorzystuje porosty do monitorowania zanieczyszczeń ze względu na sposób, w jaki pochłaniają nie tylko wodę, ale także np. dwutlenek siarki i związki azotu. Spaliny, przemysł ciężki bez jakichkolwiek lub znikomych środków ochrony środowiska i nawozów mogą przyczynić się do wymierania porostów. Porównaj obszar Śródmieścia, na przykład, z miejscami z dobra jakość powietrza, a zauważysz znaczną zmianę we florze porostów.
Perithecia, w przeciwieństwie do apotecji i gasterotecji, są owocnikami. typ zamknięty. Morfologicznie są to okrągłe lub dzbaneczkowate struktury głęboko osadzone w plechach. Perytecja komunikuje się z środowisko przez małe otwory u góry, czasami nazywane aparatami szparkowymi (nie mylić z prawdziwymi aparatami szparkowymi Wyższe rośliny!). Dojrzałe zarodniki opuszczają peritecjum przez te otwory. Jako część ściany owocnika tworzy się warstwa zewnętrzna (ekscipule), utworzona z kilku warstw strzępek, czasami na zewnątrz znajduje się jeszcze jedna skorupa - zasnówka. Wewnątrz znajduje się błona dziewicza, składająca się z rozwijających się torebek i parafizy. Warstwa ta tworzy się na wąskiej wewnętrznej warstwie ściany perytecji - hipotecji. W pobliżu otworu znajdują się specjalne nitkowate strzępki - peryfizy, które pełnią funkcję ochronną.
Kształt worków może być bardzo różnorodny, liczba dojrzewających w nich zarodników różne rodzaje również jest zróżnicowana - od jednej dużej do 200. U większości gatunków ściana worka ma jedną muszlę, ale w niektórych formach (głównie u porostów skośnych) mogą występować dwie muszle.
Zarodniki są otoczone dwuwarstwową błoną, składającą się z warstwy zewnętrznej (exosporium) i warstwy wewnętrznej (endosporium). Każdy zarodnik zawiera jądro i niewielką ilość cytoplazmy. Kształt i wielkość zarodników są zróżnicowane. Wyjście zarodników może być bierne w wyniku zniszczenia ścianki worka, ale częściej są one wyrzucane z worka na skutek wzrostu ciśnienia osmotycznego wewnątrz worka. Całkowity zarodniki utworzone przez porosty są bardzo duże. Na przykład tylko jedna apotecja porostu Solorina wytwarza około 124 000 zarodników. Uwalnianie zarodników z worków uzależnione jest od warunków atmosferycznych i przede wszystkim wilgotności – nawilżanie stymuluje uwalnianie zarodników (sporulacja).
Większość zarodników umiera i tylko kilka z nich trafia do środka korzystne warunki i rosnąć. Strzępki wykiełkowane z zarodników przeżywają tylko wtedy, gdy w pobliżu spotkają odpowiednie algi. Następnie komórki glonów splatają się ze strzępkami grzyba i stopniowo tworzą plechę (patrz 276).
REPRODUKCJA POROSÓW
spór grzyby porostowe, wyrzucane z perytecji i apotecji, wyrastają na strzępki, które przeplatając się tworzą szczątkowa plecha porosty lub prototyp. Aby w przyszłości powstał z niego prawdziwy porost, konieczny jest kontakt pierwotniaka z określonym gatunkiem. wodorost, a także pewne warunki zewnętrzne. W przeciwnym razie prototalom szybko umiera. Tak więc zarodniki grzybów utworzone w apotecji i perytecji nie mają wielkiego znaczenia dla rozmnażania porostów, ponieważ połączenie niezbędnych do tego warunków jest rzadkie w przyrodzie.
Ważniejsze do reprodukcji porostów, takich formacji, w których jednocześnie obecne są strzępki grzybów i komórki glonów. Ten soredia I izydia. Służą do reprodukcji porostów jako całego organizmu. Gdy znajdą się w sprzyjających warunkach, bezpośrednio dają początek nowej plechy. Soredia i isidia występują częściej u porostów liściastych i fruktozowych.
Soredia to najmniejsze formacje w postaci cząstek pyłu, składające się z jednej lub więcej komórek glonów otoczonych strzępkami grzybów. Ich powstawanie zwykle rozpoczyna się w warstwie gonidialnej. Ze względu na masowe tworzenie się soredii zwiększa się ich liczba, naciskają na górną korę, rozdzierają ją i trafiają na powierzchnię plechy, skąd są łatwo wydmuchiwane przy każdym ruchu powietrza lub zmywane wodą. Nazywa się skupiska soredii sorale(widzieć zdjęcie). Obecność i brak sorediów i soralów, ich umiejscowienie, kształt i kolor są stałe dla niektórych porostów i służą cechą.
Czasami, gdy porosty umierają, ich plechy zamieniają się w sproszkowaną masę składającą się z soredii. Są to tak zwane formy trądu porostów (od greckiego słowa „lepros” - „szorstki”, „nierówny”). W takim przypadku określenie porostów jest prawie niemożliwe.
Soredia, niesione przez wiatr i wodę deszczową, raz w sprzyjających warunkach, stopniowo tworzą nową plechę. Odnowienie nowej plechy z soredii jest bardzo powolne. Tak więc u gatunków z rodzaju Cladonia normalne łuski plechy pierwotnej rozwijają się z soredii dopiero po okresie od 9 do 24 miesięcy. A rozwój plechy wtórnej z apotecją trwa od jednego do ośmiu lat, w zależności od rodzaju porostu i warunków zewnętrznych.
Jeśli warunki do rozwoju nowej plechy nie są do końca sprzyjające, ale nadal możliwy jest wzrost strzępek grzybów i rozmnażanie się glonów, to z soredii rozwijają się nowe soredia i pojawia się opisana powyżej postać trądu, która wygląda jak sproszkowana miejsce.
Izydia występują u mniejszej liczby gatunków porostów niż soredia i sorals. Są to proste lub rozgałęzione koralowce wyrostki, zwykle gęsto pokrywające górną część plechy (patrz rysunek). W przeciwieństwie do sorali, izydia są pokryte korą na zewnątrz, często ciemniejszą niż plecha. Wewnątrz, pod korą, zawierają glony i strzępki grzybów. Isidia łatwo odrywa się od powierzchni wzgórza. Odrywając się i rozprzestrzeniając za pomocą deszczu i wiatru, podobnie jak soredia mogą w sprzyjających warunkach tworzyć nowe thalli porostów.
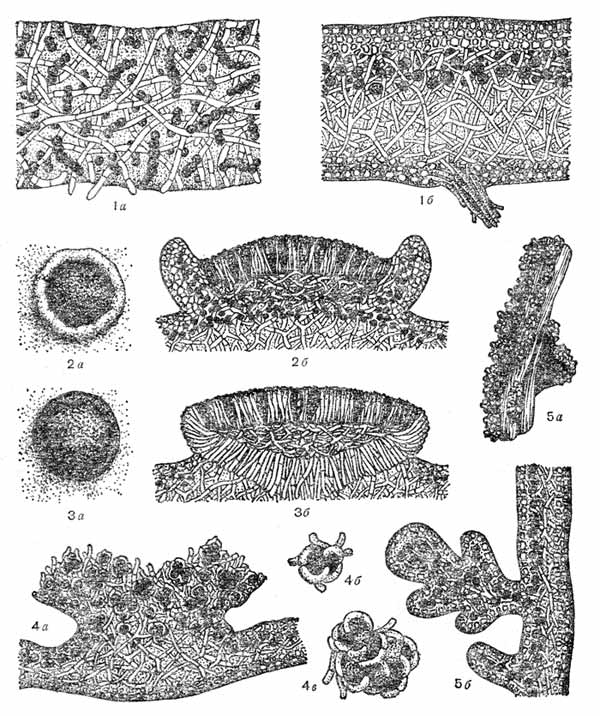
Przekrój plechy (a - homeomeryczny, 6 - heteromeryczny) 2 - apotek lekanorynowy (a - widok z góry, b - przekrój), 3 - apotek lecydynowy (a - widok z góry, b - przekrój), 4 - budowa soralów (a - soral, b, c - oddzielna soredia), 5 - isidia (a - wygląd, b - przekrój)
Wiele porostów nie tworzy apotecji, soredii i izydii i nie rozmnaża się części wzgórza, które łatwo odrywają się od porostów, które są kruche przy suchej pogodzie przez wiatr lub zwierzęta i również są przez nie przenoszone. Szczególnie rozpowszechnione jest rozmnażanie porostów w obszarach wzgórza. w regionach arktycznych. W największym stopniu rozmnażanie przez kawałki plechy jest nieodłączne od porostów epilitycznych łusek. Pojawienie się spękań w skałach i kamieniach porośniętych porostami, a także pojawienie się spękań w samym wzgórzu porosty łuskowate ze względu na nierówny stosunek do nagrzewania i nawilżania kamienia i samego porostu prowadzi do rozbicia plechy i oddzielenia jej drobnych odcinków. Te oderwane kawałki plechy są przenoszone przez wiatr i wodę w nowe miejsca, dając początek nowym plechom.
Niektóre porosty (około 100 gatunków z różnych rodzajów) charakteryzują się obecnością na powierzchni, a czasem wewnątrz wzgórza, formacji różnej wielkości w postaci rozsianych drobnych zgrubień, brodawek, maczugowatych, kulistych i koralowatych wyrostków . To są cefalodia. Mogą pokrywać górną lub dolną stronę plechy liściastej lub rozwijać się na bokach podecji lub gałązek plechy fruktozowej. W przeciwieństwie do izydiów są bardziej zróżnicowane pod względem kształtu i rzadziej rozproszone. Na przekroju pod mikroskopem widać, że mają one budowę homeomeryczną i składają się z przypadkowego nagromadzenia strzępek, wśród których równomiernie rozmieszczone są komórki lub włókna niebiesko-zielonych alg. Rzadziej cefalodia ma strukturę heteromeryczną. Glony w cefalodii są zawsze niebiesko-zielone, podczas gdy w plechach porostów niosących cefalodia algi są zawsze zielone. Powstawanie cefalodii jest spowodowane wnikaniem do plechy niebiesko-zielonych alg, które nie są charakterystyczne dla tego typu porostów, co powoduje lokalny wzrost strzępek grzyba „zarastającego” te glony. Rola cefalodii dla samego porostu nie jest do końca jasna. Zakłada się i częściowo udowodniono eksperymentalnie, że zwiększają one odżywianie porostów żyjących na ubogim podłożu. Niebiesko-zielone glony cefalodia są zdolne do wiązania azotu atmosferycznego, który jest następnie absorbowany w postaci związków azotowych przez całe plechy porostów - tworzące go grzyby i zielenice, które same nie mogą wiązać azotu atmosferycznego.
Cephalodia są specyficzne dla wielu gatunków i mają wartość cechy definiującej (systematycznej). Typowy przykład porosty z cefalodiami – naziemny gatunek pryszczatego peltigera (Peltigera aphthosa).
SynonimyŁacińskie nazwy porostów:
Na stronie ekologicznego centrum „Ekosystem” możesz zakup karta kolorów „Porosty centralnej Rosji”, jak i innych materiały dydaktyczne botanika(patrz poniżej).
Na naszej stronie internetowej można również zapoznać się z ogólne informacje o porostach: czym są porosty (wprowadzenie), budowa porostów, rozmnażanie porostów, rozmieszczenie i grupy ekologiczne porostów, znaczenie gospodarcze porostów, zbiór i identyfikacja porostów. A na stronie ekologicznego centrum „Ekosystem” można przeczytać artykuły popularnonaukowe z zakresu lichenologii Doktor nauk biologicznych A.V. Pchelkina: 1) Najprostsze metody badań lichenologicznych, 2) Wykorzystanie porostów i alg w monitoringu środowiska i badaniach bioindykacyjnych, 3) Popularna lichenologia: wstęp i spis treści (historia badań porostów, formy życia porostów, składniki porostów, sposób rozmnażania porostów, gdzie występują porosty, główne czynniki wpływające na porosty, jakie substancje wytwarzają porosty,