Citoplazma vsake celice je obdana z membrano, ki jo omejuje od okolju. Citoplazemska membrana (CPM) je izključno večnamenska struktura:
1. CPM zazna vse kemijske informacije vstop v celico iz zunanjega okolja.
2. Ona je glavna osmotska pregrada zaradi česar se znotraj celice vzdržuje določen osmotski tlak.
Celična stena je sestavljena predvsem iz peptidoglikana, prepletenega s teihojsko kislino, ki drži različne plasti skupaj. Količina zamreženja je večja in stena je debelejša kot pri stenah po Gramu negativnih celic. Gram-pozitivne celice so skoraj v celoti sestavljene iz peptidoglikana, vendar pomembna struktura najdemo v stenah po Gramu pozitivnih celic teihojsko kislino. Je fosfodiesterski polimer glicerola ali ribitola, povezan s fosfatnimi skupinami. Teihojska kislina je kovalentno povezana z muramsko kislino in teče skupaj z različnimi plastmi peptidoglikanske mreže.
3. CPM skupaj s celično steno sodeluje pri uravnavanje rasti in delitve celic bakterije.
4. CPM je vključen v regulacijo procesov replikacije in ločevanja kromosomov in plazmidov (povezani so z njegovimi receptorji).
5. CPM vsebuje pomemben znesek encimi vključno s sistemi za prenos elektronov (TsPM - mesto pridobivanja energije v bakterijah).
Teihoična kislina stabilizira celično steno in jo naredi močnejšo. Kemijska formula teihojska kislina je prikazana na sliki 25. Teihojska kislina je dolga, tanka molekula, ki je vtkana skozi peptidoglikan. Gramnegativne celične stene imajo več kompleksna struktura kot pri gram-pozitivnih organizmih. Zunaj citoplazemske membrane je periplazma, ki vsebuje tanko plast peptidoglikana. Peptidoglikan v gramnegativnih celicah vsebuje manj zamreževanja kot v grampozitivnih celicah brez peptidnega povezovalca.
6. S CPM povezane flagele in aparati za uravnavanje njihovega gibanja.
7. CPM sodeluje v procesih transport(vključno z aktivnimi) hranila v celico in odpadne produkte, vključno z encimi in eksotoksini, iz celice v okolje. Vsebuje beljakovine, ki sodelujejo pri olajšani difuziji in aktivnem transportu.
Kovalentno povezan s peptidoglikanom je Brownov lipoprotein, ki ima hidrofobno sidro, ki pomaga močno vezati peptidoglikan na zunanjo membrano. Slika 26 prikazuje lokacijo gramnegativne celične stene. Celična stena gramnegativnih bakterij vsebuje veliko manj peptidoglikana in je obdana z zunanjo membrano. Med peptidoglikanom je veliko manj navzkrižnih povezav.
Zunanja membrana po Gramu negativnih bakterij je še en lipidni dvosloj, podoben citoplazmatski membrani in vsebuje tudi beljakovine. Je ovira za beljakovine in preprečuje odlaganje encimov, ki se sproščajo v periplazmo. Ta odbojnost omogoča tem bakterijam, da preživijo v tem okolju.
8.
CPM igra pomembno vlogo pri kompartmentalizaciji in stabilizaciji ribosom.
9.
CPM sodeluje pri sintezi komponent celične stene.
10.
CPM sodeluje pri tvorbi mezosomov (mezosomi nastanejo kot posledica invaginacije mesta CPM v citoplazmo, so odprti v periplazmatski prostor).
Kako deluje membrana molekularni ravni njegovih številnih funkcij – ena od najbolj pereča vprašanja sodobna biologija. Delež CPM predstavlja približno 10 % suhe teže bakterij. Vsebuje 25-40% fosfolipidi, ki tvorijo dve plasti, 20-75% beljakovin in do 6% ogljikovih hidratov. molekule fosfolipidi so asimetrični: glave, ki nosijo električni naboj, so hidrofilne; repi so nevtralni in hidrofobni. Fosfolipidi so zapakirani v membrano na naslednji način: njihove polarne hidrofilne glave so obrnjene navzven in tvorijo dve plasti CPM - notranjo in zunanjo, nepolarni hidrofobni repi pa so skriti v debelini membrane. Na elektronskih difrakcijskih vzorcih ima CPM obliko troslojne strukture, sestavljene iz dveh 2 vzporedne temne plasti in svetla plast, ki ju ločuje. Ta plast je bolj prepustna za elektrone kot plasti, sestavljene iz polarnih koncev fosfolipidov, povezanih z beljakovinami. Specifičnost funkcij CPM je v veliki meri odvisna od nabora beljakovin, ki jih vsebujejo. Njihova lokacija v CPM je nenavadna: nekateri proteini prežemajo celotno dvojno lipidno plast, določen del proteinov je povezan samo z notranjim ali samo z zunanjo površino membrane. To izhaja iz dejstva, da je interakcija med membrano in citoplazmo na eni strani membrana in zunanje okolje, na drugi strani pa določa različne, čeprav medsebojno povezane procese njegovega vzdrževanja življenja: olajšano difuzijo, aktivni transport, elementarne biološke membrane.
On ustvarja širok spekter fiziološke reakcije vključno z indukcijo vročine, spremembami števila belih krvnih celic, uhajanjem iz krvne žile, tumorska nekroza in zmanjšana krvni pritisk vodi do vaskularnega kolapsa in na koncu do šoka. Zunanja membrana ima manj beljakovin in manj edinstvenih vrst beljakovin kot citoplazemska membrana. Porini so še posebej pomembne komponente zaradi njihove vloge pri prepustnosti zunanje membrane za majhne molekule. Porini so proteini, ki tvorijo pore v zunanji membrani, ki so dovolj velike, da omogočajo prehod najmanjših hidrofilnih molekul.
S prekomerno rastjo (v primerjavi z rastjo celične stene) citoplazemska membrana tvori invaginate – invaginacije v obliki kompleksno zavitih membranskih struktur, t.i. mezosomi. Manj kompleksne zvite strukture imenujemo intracitoplazmatske membrane. Vloga mezosomov in intracitoplazemske membrane še ni povsem razjasnjeno. Predvideva se celo, da so artefakt, ki nastane po pripravi (fiksaciji) preparata za elektronsko mikroskopijo. Kljub temu se domneva, da derivati citoplazemske membrane sodelujejo pri delitvi celic, zagotavljajo energijo za sintezo celične stene, sodelujejo pri izločanju snovi, nastajanju spor, to je v procesih z visoko porabo energije.
Na sl. 28 prikazuje strukturo porina na molekularnem merilu. Vsi znani porini imajo podobno strukturo, pri čemer protein vsebuje osrednji kanal, ki omogoča molekulam prehod. To omogoča, da se te molekule transportirajo v periplazmatski prostor za možen transport skozi citoplazmo. plazemska membrana. Nekateri porini v zunanji membrani so splošni, omogočajo preprosto razlikovanje velikosti in naboja, vendar imajo malo specifičnosti. Drugi porini so bolj specifični.
Zelo velike ali hidrofobne molekule ne morejo prodreti skozi zunanjo membrano, zato zunanja membrana služi kot prepustna ovira za vsaj nekatere molekule. Molekularna struktura porina. Notranji pogled je zunaj celice in gleda na površino membrane. Pogled je perspektiva od zunaj. Porin ima tri beljakovinske podenote in dejanska pora je osrednje trikotno območje na zgornji plošči, ki ga tvorijo tri podenote.
citoplazma zavzema večino bakterijske celice in je sestavljena iz topnih beljakovin, ribonukleinskih kislin, vključkov in številnih majhnih zrnc - ribosomov, odgovornih za sintezo (prevajanje) beljakovin.
Ribosomi bakterije imajo velikost približno 20 nm in koeficient sedimentacija 70S, v nasprotju z ribosomi 8OS, značilnimi za evkariontske celice. Zato se nekateri antibiotiki vežejo na bakterijske ribosome in zavirajo sintezo bakterijskih beljakovin, ne da bi vplivali na sintezo beljakovin v evkariontskih celicah. Bakterijski ribosomi lahko disociirajo na dve podenoti, 50S in 30S. Ribosomska RNA (rRNA.) - konzervativni elementi bakterij ("molekularna ura" evolucije). 16S rRNA. je del male podenote ribosomov, 23S rRNA pa del velike podenote ribosomov. Študija 16S rRNA. je osnova genosistematike, ki omogoča oceno stopnje sorodnosti organizmov.
Obstajajo tudi druge vrste beljakovin zunanja membrana ki sodelujejo pri različne funkcije. Nekateri patogeni vsebujejo beljakovine zunanje membrane, ki jim pomagajo nevtralizirati obrambo gostitelja. Razlike med celičnimi stenami gramnegativnih in gramnegativnih bakterij močno vplivajo na uspešnost mikrobov v njihovem okolju. Debela celična stena po Gramu negativnih celic jim omogoča, da se bolje soočajo v suhih razmerah, saj zmanjšuje izgubo vode. Tabela 3 povzema razlike med gramnegativnimi in gramnegativnimi celičnimi stenami.
Citoplazma vsebuje različne vključevanje v obliki granul glikogen, polisaharidi, beta-hidroksimaslena kislina in polifosfati (volutin). Vse rezervne snovi so v celicah prisotne v kemično inertni obliki. Ta pogoj preprečuje kršitev osmostaze celične vsebine. Nekateri vključki preprosto ležijo v citoplazmi, drugi so obdani s tanko membrano. Membrana je običajno beljakovinske narave, včasih pa lahko vsebuje lipide. Kopičijo se s presežkom hranil v okolju in služijo kot rezervne snovi za prehranske in energetske potrebe.
Tabela 3 Lastnosti celične stene
Povzetek razlik med gramnegativnimi in gramnegativnimi celičnimi stenami. Za večino bakterijskih celic ima celična stena ključnega pomena za preživetje celic, vendar obstajajo nekatere bakterije, ki nimajo celične stene. Pogosti primeri so vrste mikoplazme, nekatere pa so lahko znotrajcelični patogeni, ki rastejo znotraj svojih gostiteljev. Celične stene tukaj niso potrebne, ker celice živijo samo v nadzorovanem osmotskem okolju drugih celic.
Volyutin ima afiniteto do osnovnih barvil in se zlahka zazna s posebnimi metodami barvanja (na primer po Neisserju) v obliki metakromatskih zrnc. Toluidinsko modro ali metilensko modro obarvata volutin rdeče-vijolično, bakterijsko citoplazmo pa modro. Značilna razporeditev volutinskih zrnc se pri difteričnem bacilu kaže v obliki intenzivno obarvanih polov celice. Metakromatsko obarvanje volutina je povezano z visoka vsebnost polimeriziran anorganski polifosfat. Pod elektronsko mikroskopijo so videti kot zrnca z elektronsko gostoto velikosti 0,1-1,0 µm.
Verjetno so nekoč v preteklosti imeli možnost oblikovati celično steno, a ko je njihov življenjski slog postal bivanje znotraj drugih celic, so izgubili sposobnost ustvarjanja sten. V skladu s tem zelo omejenim načinom življenja v drugih celicah so tudi ti mikrobi zelo majhni. Ne potrebujejo genov za vse vrste biosintetskih encimov, saj lahko gostitelju ukradejo končne komponente teh poti. Prav tako ne potrebujejo genov, ki kodirajo veliko različne poti za različne vire ogljika, dušika in energije, saj je njihovo znotrajcelično okolje povsem predvidljivo.
Nukleoid je enakovredno jedru v bakterijah. Leta 1956 so v bakterijskih celicah odkrili "jedrsko cono" ali nukleoid, kjer se nahaja bakterijski kromosom. Bakterijska DNK se nahaja v krožni in linearni obliki. Za celice E. coli je bilo dokazano, da obstaja DNK v obliki krožne molekule, za Borrelio burgdorferi pa je bilo leta 1989 dokazano, da celice vsebujejo DNK v linearni obliki. Nukleoid, za razliko od evkariontskega jedra, nima jedrsko ovojnico, nukleolus in glavne beljakovine (histoni). Ponavadi v bakterijska celica vsebuje en kromosom, če pa je delitev motena, lahko vsebuje 4 ali več kopij enega kromosoma. Pri nekaterih vrstah so v celicah našli dva ali tri neidentične kromosome. Vrste Vibrio, Leptospira interrogans imajo dva krožna kromosoma. Zato se ideja, da bakterije vsebujejo samo en kromosom, šteje za zastarelo. Nukleoid se zazna v svetlobnem mikroskopu po barvanju z metodami, specifičnimi za DNK: po Feulgenu ali Romanovsky-Giemsa. Na elektronskih difrakcijskih vzorcih ultratankih odsekov bakterij ima nukleoid obliko svetlih con s fibrilarnimi, nitastimi strukturami DNA, ki so povezane z določenimi območji s citoplazmatsko membrano ali mezosomom, ki sodeluje pri replikaciji kromosomov.
Prisotnost sterolov v membrani prispeva k njihovi dolgoživosti, saj pomaga povečati sile, ki držijo membrano skupaj. Ker ne bi bilo ljudi, rastlin, živali, gliv ali praživali. Vsi ti organizmi so Živo bitje, poleg bakterij, sestavljajo evkariontske celice.
In nastanek te vrste celic ne bi bil mogoč brez simbioze. Nastanek evkariontskih celic v znanosti razlagajo s tako imenovano endosimbiontsko teorijo. Po tej teoriji kompleksne celice z celično jedro, ločeni od plazme in celičnih organelov, so nastali iz simbioze preprosteje strukturiranih prokariontov. Samo bakterije, ki so živele kot endosimbiont v večji celici, so oblikovale organele današnjih evkariontskih celic.
Poleg nukleoida ima bakterijska celica ekstrakromosomske dejavnike dednosti. - plazmidi, ki so kovalentno sklenjeni obroči DNA, ki ne kodirajo funkcij, ki so bistvene za življenje bakterijske celice, vendar dajejo bakteriji prednosti, ko je izpostavljena neugodnim pogojem obstoja.
Tvorba mitohondrijev
To kopičenje organske snovi je dobrodošel vir hranil za heterotrofne celice. Celice pridobijo velikost in poleg tega, da lahko absorbirajo organska snov fagocitozo, sposoben absorbirati organske celice pa tudi manjše celice se fagocitirajo. Praviloma so te celice v obliki hrane, se prebavijo.
Včasih pa se zgodi, da žive bakterije preživijo v notranjosti večjo celico in komunicirajo s svojim gospodarjem. Razvije se simbioza, v kateri lahko gostiteljska celica bolj učinkovito uporablja prehrambeni izdelki z uporabo encimov iz dihalne verige aerobnih bakterij. Po drugi strani pa je endosimbiont znotraj večje celice bolj zaščiten pred okoljskimi spremembami. drugo možen vir mitohondrije napadejo bakterijski paraziti v gostiteljski celici in posledična simbioza.
Kapsula, mikrokapsula, sluz. Kapsula- sluzna struktura z debelino več kot 0,2 mikrona, trdno povezana s celično steno bakterije in ima jasno določene zunanje meje. Kapsula je razločljiva v brisih-odtisih patološkega materiala. Površina celičnih kolonij s kapsulami je videti gladka, vlažna, sijoča.
Organske celice z več membranami
Ni povsem zanesljivo znano, ali so evkarionti dejansko nastali na ta način, vendar obstaja veliko dejstev, ki podpirajo teorijo o endosimbiontih. Prav tako organele ne morejo nastati iz genetske informacije jedra, ampak se pomnožijo z delitvijo. Postopek delitve je podoben procesu delitve bakterij. Ribosomi celičnih organelov so strukturno bolj podobni bakterijskim kot evkariontskim ribosomom.
Endosimbioze najdemo še danes. Tako lahko nekatere morske diatomeje očitno živijo v simbiozi s pomanjkljivimi cianobakterijami. Spet je simbiont integriran v plazmo gostiteljske celice. Zato se še danes tovrstna simbioza ponavlja kot proces, ki je stal na začetku vsega višjega življenja.
Nastane veliko bakterij mikrokapsula- tvorba sluznice z debelino manj kot 0,2 mikrona, odkrita le z elektronsko mikroskopijo. Sluz je treba razlikovati od kapsule - mukoidnih eksopolisaharidov, ki nimajo jasnih zunanjih meja. Sluz je topen v vodi.
Mukoidni eksopolisaharidi so značilni za mukoidne seve Pseudomonas aeruginosa, ki jih pogosto najdemo v sputumu bolnikov s cistično fibrozo. Bakterijski eksopolisaharidi sodelujejo pri adheziji (lepljenju na podlago); imenujemo jih tudi glikokaliks. Poleg sinteze eksopolisaharidov s strani bakterij obstaja še en mehanizem za njihov nastanek: z delovanjem zunajceličnih bakterijskih encimov na disaharide. Posledično nastanejo dekstrani in levani.
Zanj je značilna odsotnost jedra. Njihova glavna delitev se pojavlja v dveh in Archaea. IN Vsakdanje življenje Prokarioti so nam znani kot bakterije. Pogosto se beseda "bakterije" ali bakterije neposredno enači s patogeni. Še najmanj pa so prokarionti patološki – torej patogeni.
Celična stena obdaja citoplazmatsko membrano in notranji del celice. Notranje opreme ni. Vsi prokarionti imajo enako osnovno strukturo. Večina prokariotov ima celično steno, ki se nahaja zunaj plazemske membrane in je sestavljena iz derivata ogljikovih hidratov. Glede na število in razporeditev mureinskih plasti ločimo po Gramu pozitivne in po Gramu negativne bakterije.
- Celico obdaja plazemska membrana.
- Po tej vrsti arheje nimajo celične stene.
Kapsula in sluz ščitita bakterije pred poškodbami in izsušitvijo, saj kot hidrofilna dobro vežeta vodo in preprečujeta delovanje zaščitnih faktorjev makroorganizma in bakteriofagov.
Flagella bakterije določajo mobilnost bakterijske celice. Flagele so tanke niti, ki izvira iz citoplazme imajo membrane daljši od same celice. Bički so debeli 12–20 nm in dolgi 3–15 µm. Sestavljeni so iz 3 delov.:
Načeloma se celica, obdana z membrano, ne deli naprej. Na splošno obstaja mehanizem, ki vam omogoča premikanje. Ima na citoplazmatski membrani, ki leži na drugi celični plasti. Izdelan je iz muraina. Na njem leži zunanja lupina. Oblegani enocelični so vnesli zelene alge za subtenzijo. Simbioza povzroča drastične spremembe v obeh celicah. enocelični organizem preklaplja med fazo plenilca in fazo gostitelja z endosimbiontom, s čimer preklopi svojo energijsko bilanco iz heterotrofne v avtotrofno, tj. od dobave do sončne energije.
1. - spiralna nit,
2. - kavelj
3. - bazalno telo, ki vsebuje paličico s posebnimi diski (1 par diskov pri gram-pozitivnih in 2 para pri gram-negativnih bakterijah) in motorične beljakovine (sl.
diski flagele so pritrjene na citoplazmatsko membrano in celične stene. Tako nastane učinek elektromotorja s palico - rotorjem, ki vrti biček. Kot vir energije se uporablja razlika protonskih potencialov na citoplazmatski membrani. Rotacijski mehanizem zagotavlja protonska ATP sintetaza. Hitrost vrtenja flagelluma lahko doseže 100 vrt / min. Bakterije imajo desnosučne flagele. Če ima bakterija več flagel, se začnejo sinhrono vrteti, prepletati v en sam snop in tvoriti nekakšen propeler.
V procesu prilagajanja dveh celic rastlina nastane iz praživali z delci; Raziskovalci so jih poimenovali Hatena, preveč skrivnostni za nemški. Ta ugotovitev je dodatna podpora endosimbiotski teoriji, po kateri so predhodniki vseh rastlinskih in živalskih celic nastali z endosimbiozo.
Katere organele so dobili predhodniki rastlinske celice po Endosombiontentoria Endosimbioza? Navedite štiri značilnosti teh organelov, ki podpirajo endosimbiotično teorijo! V vsakem primeru utemeljimo, zakaj menite, da je značilnost dokaz endosimbiontske teorije. V kolikšni meri lahko odkritje novih alg Khaten velja za osnovo teorije o endosimbiontih? Med endosimbiozo z aerobnimi bakterijami so primarni evkarionti s simbiozo s cianobakterijami pridobili sposobnost oksidativne razgradnje, sposobnost fotosinteze. Ne morejo se ponovno pojaviti. To nakazuje, da prvotno niso nastale kot strukture gostiteljskih celic. Mitohondriji in kloroplasti imajo lasten mehanizem za sintezo beljakovin in njihovi ribosomi so zgrajeni kot prokarionti. Oba sta obdana z dvema membranama, od katerih je notranja podobna prokariontska membrana, zunaj celična membrana. Pomembna značilnost endosimbiotske teorije je dejstvo, da med prokariontskimi in evkariontskimi celicami ni vmesnih oblik, obstajajo pa vmesne oblike med endosimbiotskimi organizmi in organeli. Ta prehodna oblika je opazovana enocelična alga: po spremembi prehrane je enocelična odvisna od simbioze z zelenimi algami, medtem ko je le zelene alge je sposoben preživeti.
- Katere posebne veščine ste pridobili?
- Mitohondriji in kloroplasti so potomci mikroorganizmov.
- Mitohondriji izhajajo iz bakterij, kloroplasti iz cianobakterij.
- Oba nastaneta šele z ločitvijo svojih vrstnikov.
Flagele so sestavljene iz beljakovin flagelin(iz. flagellum- flagellum), ki je antigen - ti H-antigen. Podenote flagelina so zvite.
Število bičkov v bakterijah različne vrste razlikuje od enega (monotrich) pri vibrio cholerae do deset in več sto flagel, ki se raztezajo vzdolž oboda bakterije (peritrič), pri coli, protea itd. lofotričen imajo na enem koncu celice snop flagel. amfitrihi imajo en biček ali snop bičkov na nasprotnih koncih celice.
Flagele odkrivamo z elektronsko mikroskopijo preparatov, poškropljenih s težkimi kovinami, ali v svetlobnem mikroskopu po obdelavi. posebne metode temelji na luženju in adsorpciji različne snovi, kar vodi do povečanja debeline flagele (na primer po posrebrenju).
resice, ali pili(fimbrije) - nitaste tvorbe, več tanek in kratek, kot flagella. Pili segajo od površine celice in so sestavljeni iz pilina veverica, ki je organiziran v spiralno strukturo. Arhitektura pilijev se spreminja od tankih nitastih do debelih močnih paličastih formacij z aksialnimi luknjami. Imajo antigensko aktivnost. Razlikovati pitje, odgovoren za oprijem t.j. za pritrditev bakterij na prizadeto celico, pa tudi za pitje, ki je odgovorno za prehrano, izmenjava vode in soli, In spolno (F-pil) ali pili 4. vrsto, ali konjugirano, pili.
Običajno obilno pil- nekaj sto na celico. Vendar običajno so 1-3 spolni pili na celico: tvorijo jih tako imenovane "moške" donorske celice, ki vsebujejo transmisivne plazmide (F-, R-, Co/-plazmidi). Posebnost spolnih pilijev je njihova interakcija s posebnimi "moškimi" sferičnimi bakteriofagi, ki se intenzivno adsorbirajo na spolnih pilih (slika).
Glavni namen pilijev je podpiranje specifičnih pritrdilnih struktur (adhezinov) na njihovih koncih. Adhezini so posredniki pri stiku z bakterijami, v stiku z neživimi predmeti, tkivi in celicami dovzetnih organizmov. Kolonizacija gostiteljskih tkiv z bakterijskimi patogeni je na splošno odvisna od stereokemične podobnosti med arhitekturo adhezina in ustreznim receptorjem gostiteljske celice (slika). Tako bo preučevanje delovanja pili omogočilo ne le globlje razumevanje mehanizma kolonizacije in signalizacije, temveč tudi razvoj novih generacij protimikrobnih zdravil.
polemika - posebna oblika mirujočih bakterij z gram-pozitivno strukturo celične stene (slika).
Spore nastanejo, ko ne ugodni pogoji obstoj bakterij (sušenje, pomanjkanje hranil itd.). Znotraj bakterijske celice nastane ena trosa (endospora). Tvorba trosov prispeva k ohranjanju vrste in ne Xia način razmnoževanja, kot gobe.
bakterije iz rodu, ki tvorijo spore bacil, ki velikost spore ne presega premera celice, imenovane bacili. bakterije, ki tvorijo spore velikost trosov presega premer celice, zato imajo obliko vretena, imenujemo jih klostridija, kot so bakterije iz rodu Clostridium(lat. klostridij- vreteno). Spore so kislinsko odporne, zato so obarvane z metodo Aujeszkyja ali z metodo Ziehl-Nelsen v rdeči barvi, vegetativna celica pa v modri barvi.
Sporulacija, oblika in lokacija trosov v celici (vegetativno) je vrstna lastnost bakterij, ki jih omogoča razlikovanje med seboj.
Obrazec za spor lahko biti ovalen, sferičen; ureditev v celici :
1. terminal, na koncu paličice (pri povzročitelju tetanusa),
2. subterminal- bližje koncu palice (za povzročitelje botulizma, plinska gangrena)
3. centralno e (pri bacilu antraksa).
Proces tvorba spor (sporulacija) poteka skozi vrsto stopenj, med katerimi se loči del citoplazme in kromosom bakterijske vegetativne celice, obdan z vraščeno citoplazmatsko membrano, - a prospore.
Prosporus obdana z dvema citoplazemskima membranama, med katerima nastane debela spremenjena peptidoglikanska plast korteksa (korteks). Od znotraj pride v stik s celično steno spore, od zunaj pa z notranjo lupino spore. Zunanjo lupino spore tvori vegetativna celica.
Spore nekaterih bakterij imajo dodatno prevleko - eksosporij.
Tako nastane večplastna slabo prepustna lupina. Sporulacijo spremlja intenzivna poraba prospore, nato pa nastajajoča lupina spor dipikolinske kisline in kalcijevih ionov. Spore pridobi toplotna odpornost, kar je povezano s prisotnostjo kalcijevega dipikolinata v njem.
Spore lahko obstojajo dolgo časa zaradi prisotnosti večplastne lupine, kalcijevega dipikolinata, nizka vsebnost voda in počasni presnovni procesi. V tleh, na primer, patogeni antraks in tetanus lahko traja desetletja.
V ugodnih razmerah spore kalijo skozi tri zaporedne stopnje (slika):
1. aktiviranje
2. iniciacija
3 . rast
V tem primeru iz ene spore nastane ena bakterija.
Aktivacija - pripravljena je za kalitev. Pri temperaturi 60-80°C se tros aktivira za kalitev.
Iniciacija kalitev traja nekaj minut. Značilna je stopnja rasti hitra rast, ki ga spremlja uničenje lupine in sprostitev sadike.
Vprašanja za samokontrolo
| | | | | | | | | 10 | | | | | |
Struktura tipične bakterijske celice je prikazana na sl. 2.3. Na sl. 2.4 prikazuje elektronsko mikrografijo odseka paličaste bakterije. Vidite lahko, kako preprosta je bakterijska celica, zlasti v primerjavi z evkariontskimi celicami (sliki 7.5 in 7.6).
riž. 2.3. Posplošen diagram strukture paličaste bakterijske celice. Na desni so strukture, ki jih najdemo v vsaki celici, na levi - ne v vseh celicah. Flagellum včasih enega, kot Rhizobium, ali več, kot Azotobacter; običajno je daljši od kletke. Kapsula lahko je sluzast, kot pri Azotobacterju; če je kapsula ohlapna, potem se imenuje sluzna plast. cevaste ali vrečaste oblike fotosintetske membrane ki vsebujejo pigmente so invaginacije plazemske membrane; pri fotosintetskih bakterijah, kot je Chromatium, so takšne membrane razpršene po vsej citoplazmi. številka pili, oz pili, lahko doseže od enega do več sto, kot na primer v Escherichia coli, salmonela. mezosom je večkratna invaginacija plazemske membrane, kot na primer pri Bacillus subtilis. celične stenežilava in vsebuje murein. Ribosomi, ki se nahaja po celotni citoplazmi, je manjši kot pri evkariontih. Od rezervna hranila v bakterijskih celicah lahko zaznamo lipide, glikogen, polifosfate (volutinska zrnca). citoplazma ne vsebuje organelov; vsebuje encime itd.
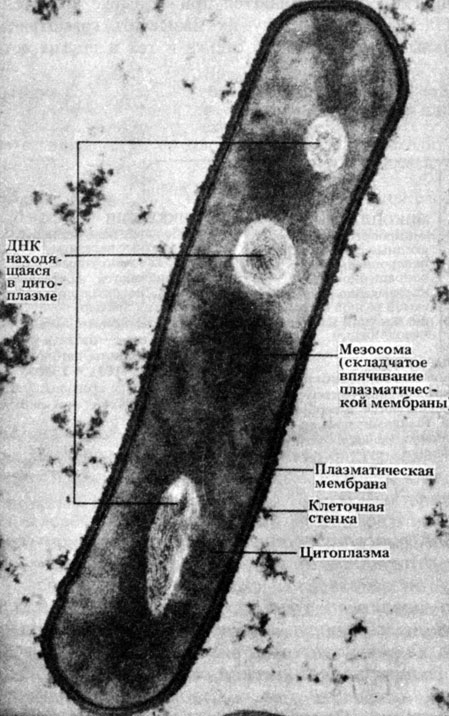
riž. 2.4. Elektronska mikrofotografija odseka tipične paličaste bakterije Bacillus subtilis. Svetla področja vsebujejo DNK. × 50000
Kapsule in sluznice
Kapsule in sluznice so sluzasti ali lepljivi izločki nekaterih bakterij; takšni izcedki so jasno vidni po negativnem kontrastu (ko barvanje ni pripravek, ampak ozadje). Kapsula je razmeroma debel in kompakten ter plast sluzi veliko bolj ohlapno. V nekaterih primerih sluz služi za oblikovanje kolonij iz posameznih celic. Tako kapsula kot sluznica služita kot dodatna zaščita celic. Inkapsulirani sevi pnevmokokov se na primer prosto razmnožujejo v človeškem telesu in povzročajo pljučnico, neinkapsulirane seve pa fagociti zlahka napadejo in uničijo in so zato popolnoma neškodljivi.
celične stene
Celična stena daje celici določeno obliko in togost. Na rezu je jasno viden (slika 2.4). Tako kot pri rastlinah celična stena bakterij preprečuje osmotsko nabrekanje in zlom celice, ko pridejo v hipotonično okolje, kar se pogosto zgodi (Dodatek, razdelek A.1.5). Voda, druge majhne molekule in različni ioni zlahka prehajajo skozi drobne pore v celični steni, velike molekule beljakovin in nukleinskih kislin pa ne prehajajo skoznje. Poleg tega ima celična stena antigenske lastnosti, ki ji jih dajejo beljakovine in polisaharidi, ki jih vsebuje.
Glede na zgradbo celične stene lahko bakterije razdelimo v dve skupini. Nekatere so obarvane po Gramu, tako se imenujejo gram-pozitiven, druge pa postanejo brezbarvne, ko barvilo speremo (razdelek 2.7), zato jih imenujemo gramnegativni. V celični steni obeh je posebna toga mreža, sestavljena iz mureina. Molekula mureina je pravilna mreža vzporednih polisaharidnih verig, ki so med seboj premrežene s kratkimi peptidnimi verigami. Tako je vsaka celica obdana z retikularno vrečko, ki jo sestavlja samo ena molekula. (Polisaharidni del mureina je opisan v tabeli 5.7).
Pri gram-pozitivnih bakterijah, kot je Lactobacillus, so v mureinsko mrežo vgrajene druge snovi, predvsem polisaharidi in beljakovine. To ustvari razmeroma debel in tog paket okoli celice. Gramnegativne bakterije, kot sta Escherichia coli ali Azotobacter, imajo veliko tanjšo celično steno, vendar je bolj zapletena. Mureinska plast teh bakterij je na zunanji strani prekrita z mehko in gladko plastjo lipidov. To jih ščiti pred lizocim. Lizocim se nahaja v slini, solzah in drugih telesnih tekočinah ter v beljakovinah piščančje jajce. Katalizira hidrolizo določenih vezi med ostanki ogljikovih hidratov in tako cepi polisaharidno ogrodje mureina. Celična stena poči, in če je celica v hipotonični raztopini, pride do njene lize (celica osmotsko nabrekne in poči). Lipidna plast daje celici odpornost na penicilin. Ta antibiotik preprečuje nastanek zamreženj v celični steni gram-pozitivnih bakterij, zaradi česar so rastoče celice bolj dovzetne za osmotski šok.
Flagella
Številne bakterije so gibljive in ta gibljivost je posledica prisotnosti enega ali več flagel v njih. Bakterijski bički so veliko enostavnejši od bičkov evkariontov (oddelek 17.6.2, tabela 2.1) in po svoji zgradbi spominjajo na enega od mikrotubulov evkariontskega bička. Flagele so sestavljene iz identičnih sferičnih beljakovinskih podenot flagelin(podobno kot mišični aktin), ki so razporejeni spiralno in tvorijo votel valj s premerom okoli 10-20 nm. Kljub valoviti obliki bičkov so precej togi.
Flagele poganja edinstven mehanizem. Osnova bička se očitno vrti tako, da se zdi, da je biček privit v medij brez naključnih udarcev, in tako premika celico naprej. To je očitno edina znana struktura v naravi, kjer se uporablja princip kolesa. drugo zanimiva lastnost flagela je sposobnost posameznih podenot flagelina, da se v raztopini spontano sestavijo v vijačne filamente. Spontana samosestava- Zelo pomembna lastnina veliko kompleksnih bioloških struktur. V tem primeru je samosestavljanje v celoti posledica aminokislinskega zaporedja (primarne strukture) flagelina.
Gibljive bakterije se lahko premikajo kot odgovor na določene dražljaje, tj. so sposobne taksiranja. na primer aerobne bakterije imajo pozitivno aerotakso (tj. plavajo tja, kjer je okolje bogatejše s kisikom), mobilne fotosintetske bakterije pa pozitivno fototaksijo (tj. plavajo proti svetlobi).
Flagele je najlažje videti v elektronskem mikroskopu (slika 2.5) s tehniko nanašanja kovin (razdelek A.2.5).
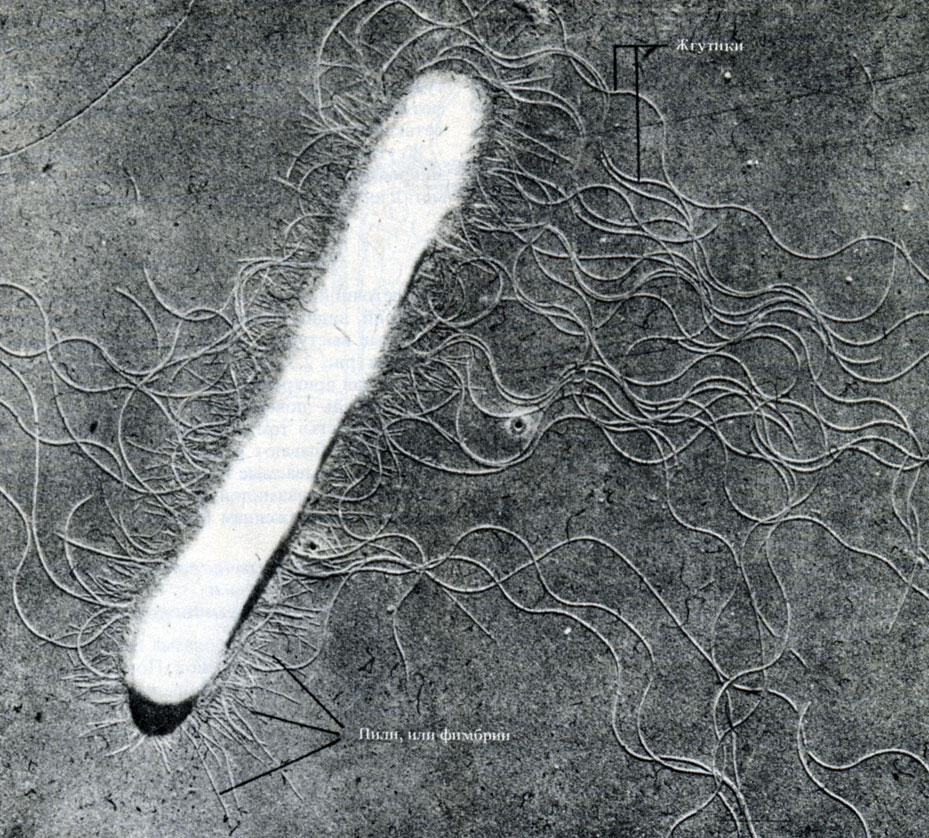
riž. 2.5. Mikrografija paličaste bakterije, pridobljena s prosojno svetlobo elektronski mikroskop. Celična stena, fimbrije in dolgi valoviti bički so jasno vidni, × 28000
Pili ali fimbrije
Na celični steni nekaterih gram-negativnih bakterij so vidni tanki izrastki (paličaste beljakovinske izbokline), ki jih imenujemo pil oz fimbrije(slika 2.5). So krajši in tanjši od bičkov in služijo za pritrditev celic med seboj ali na neko površino, kar daje posebno "lepljivost" tistim sevom, ki jih imajo. Obstaja pitje drugačen tip. Najbolj zanimivi so tako imenovani F-pili, ki jih kodira poseben plazmid (točka 2.2.4) in so povezani s spolnim razmnoževanjem bakterij.
Plazemska membrana, mezosomi in fotosintetske membrane
Kot vse celice je protoplazma bakterij obdana s polprepustno membrano. Bakterijske plazemske membrane se po strukturi in delovanju ne razlikujejo od membran evkariontskih celic (oddelek 7.2.1). Pri nekaterih bakterijah se plazemska membrana izboči v celico in tvori mezosome in/ali fotosintetske membrane.
mezosomi- zložene membranske strukture (sl. 2.3 in 2.4), na površini katerih so encimi, ki sodelujejo v procesu dihanja. Zato lahko mezosome imenujemo primitivni organeli. Med celično delitvijo se mezosomi vežejo na DNA, kar očitno olajša ločitev dveh hčerinskih molekul DNA po replikaciji in spodbuja nastanek septuma med hčerinskimi celicami.
Pri fotosintetskih bakterijah vrečaste, cevaste ali lamelarne invaginacije plazemske membrane vsebujejo fotosintetske pigmente (vključno z bakterioklorofilom). Podobne membranske tvorbe sodelujejo tudi pri fiksaciji dušika.
genetski material
Bakterijska DNK je predstavljena z enojnimi okroglimi molekulami, dolgimi približno 1 mm. Vsaka taka molekula je sestavljena iz približno 5 × 10 6 baznih parov. Skupna vsebnost DNK (genoma) v bakterijski celici je veliko manjša kot v evkariontski, posledično pa je manjša tudi količina informacij, ki so v njej zakodirane. V povprečju takšna DNK vsebuje nekaj tisoč genov, kar je približno 500-krat manj kot v človeški celici (glej tudi tabelo 2.1 in sliko 2.3).
Ribosomi
Glej tabelo. 2.1 (biosinteza beljakovin) in sl. 2.3.
polemika
Nekatere bakterije (večinoma iz rodu Clostridium ali Bacillus) tvorijo endospore, tj. spore, ki so znotraj celice. Endospore so debelostenske, dolgožive tvorbe, ki so izjemno odporne na vročino in kratkovalovno sevanje. V notranjosti celice se nahajajo različno, kar je zelo pomembna značilnost za identifikacijo in sistematiko tovrstnih bakterij (slika 2.6). Če mirujoča, stabilna struktura nastane iz cele celice, se imenuje cista. Ciste tvorijo nekatere vrste Azotobacter.
riž. 2.6. Različne oblike bakterije na primeru več najpogostejših vrst koristnih in patogenih mikrobov.
A. Cocci (krogla)
Primer je Staphylococcus aureus, ki živi v nazofarinksu; različni sevi stafilokokov povzročajo furunculozo, pljučnico, zastrupitev s hrano in druge bolezni.
Primer so številne vrste Streptococcus; neki vzrok nalezljive bolezni vrh dihalni trakt; na primer S. pyogenes povzroča vneto grlo in škrlatinko; S. thermophilus daje jogurtu pikanten okus; S. lactis - glej razd. 2.3.4
Ta rod vključuje edino vrsto Diplococcus pneumoniae (pnevmokok), povzročitelja pljučnice *.
* (D. pneumoniae povzroča lobarno pljučnico in številne hude akutne piogene okužbe: meningitis, septikemijo, empiem in peritonitis. - pribl. prevod)
B. Bacilli (v obliki palice)
![]()
Primeri so Escherichia coli (običajni črevesni simbiont); Lactobacillus glej razd. 2.3.4; Salmonella typhi je povzročitelj trebušnega tifusa.
Primeri so Azotobacter, bakterija, ki veže dušik; Bacillus anthracis je povzročitelj antraksa.
Bacili z endosporami (spore so v različnih položajih, imajo različne velikosti in oblika)
Nahaja se v središču in ne povzroča otekanja celice, na primer pri Bacillus anthracis, povzročitelju antraksa.
Nahaja se na koncu matične celice in jo daje značilna oblika bobnarska palčka, kot je Clostridium tetani, povzročitelj tetanusa.
Spora je v subterminalnem položaju, kar povzroča otekanje celice, na primer pri Clostridium botulinum (spore lahko zasedejo tudi osrednji položaj) - povzročitelj smrtonosne zastrupitev s hrano- botulizem.
B. Spirilli (spirala)
Primer je Spirillum; Oblika celic pri spirohetah je zelo podobna, razlike pa so v načinu premikanja, npr. Treponema pallidum- povzročitelj sifilisa.
D. Vibriosi (kratke palice, vedno ukrivljene v obliki vejice)
Primer - Vibrio cholerae - povzročitelj kolere; ima en flagellum.