هناك ثلاث فرضيات للأصل الخلايا حقيقية النواة:
- الفرضية التكافلية، أو التكافل,
- غزاوي,
- خيمري.
حتى الآن ، في العالم العلمي ، تم التعرف على الفرضية الرئيسية لأصل حقيقيات النوى التكافل.
وفقًا للتكوين التكافلي ، حدثت عضيات الخلية حقيقية النواة مثل البلاستيدات الخضراء والسوط عن طريق إدخال أحد بدائيات النوى إلى خلية بدائية النواة أخرى أكبر ، والتي لعبت دور الخلية المضيفة.
في الفرضية التكافلية ، توجد صعوبات في شرح أصل نواة الخلايا حقيقية النواة وفي مسألة أي بدائيات النوى كانت هي المضيف بعد كل شيء. تُظهر بيانات التحليل الجزيئي للجينوم والبروتينات حقيقية النواة أنه ، من ناحية ، كان كائنًا قريبًا من العتائق (كانوا ينتمون إلى البكتيريا ، ثم تم فصلهم إلى فرع منفصل). من ناحية أخرى ، توجد في حقيقيات النوى البروتينات (والجينات المسؤولة عن تركيبها) التي تتميز بمجموعات أخرى مثالية من بدائيات النوى.
وفقًا لفرضية الانغماس لأصل الخلايا حقيقية النواة ، تشكلت عضياتها عن طريق غزو العصارة الخلوية. غشاء بلازميمع الفصل اللاحق لهذه الهياكل. تم تشكيل شيء مثل الكرات ، محاطًا بغشاء ويحتوي داخل السيتوبلازم والمركبات والهياكل التي تم التقاطها هنا. اعتمادًا على ما دخل في الداخل ، تم تشكيل عضيات مختلفة.
لا تحتوي بدائيات النوى على عضيات حقيقية ، يتم تنفيذ وظائفها عن طريق غشاءات الغشاء. لذلك ، من السهل تخيل مثل هذا الجلد. أيضًا لصالح فرضية الانغلاف هي تشابه الغشاء السيتوبلازمي والأغشية المزدوجة للعضيات.
من وجهة نظر الانغماس ، يمكن تفسير أصل النواة بسهولة ، ولكن لا يمكن تفسير سبب اختلاف الجينوم والريبوزومات في المركب النووي السيتوبلازمي عن تلك الموجودة في البلاستيدات الخضراء والميتوكوندريا (تذكر أنها تحتوي أيضًا على الحمض النووي والريبوزومات). علاوة على ذلك ، في هذه العضيات ، يشبه نظام التخليق الحيوي للبروتين (DNA ، RNA ، الريبوسومات) نظام بدائيات النوى.
تم شرح هذا الاختلاف جيدًا من وجهة نظر الفرضية التكافلية الأولى. وفقا لها ، بطريقة أو بأخرى ، دخلت بدائيات النوى الهوائية في بدائيات النوى اللاهوائية. لم يتم هضمه ، بل بدأ ، على العكس من ذلك ، في تناول الطعام على حساب الخلية المضيفة. في المقابل ، استخدم الأكسجين للحصول على الطاقة ، وكانت طريقة الأكسدة هذه أكثر كفاءة ، فقد أعطى الطاقة الزائدة للمضيف بدائية النواة ، والتي استفادت أيضًا في هذه الحالة. نشأ التعايش. في وقت لاحق ، أصبحت بدائيات النوى المقدمة أبسط ، وهاجر جزء من جينومها إلى الخلية المضيفة ، ولم يعد من الممكن أن توجد بشكل مستقل.
بطريقة مماثلة ، يشرح التكاثر التكافلي أصل البلاستيدات الخضراء. تم إدخال بدائيات النوى فقط القادرة على التمثيل الضوئي (على غرار الطحالب الخضراء المزرقة).
أدت أول حقيقيات النوى بدون المتعايشات الضوئية إلى ظهور الحيوانات التي ظهرت فيها - النباتات.
حاليًا ، هناك كائنات أولية (كائنات حقيقية النواة وحيدة الخلية) لا تحتوي على الميتوكوندريا أو البلاستيدات الخضراء. ولكن بدلاً من ذلك ، تستقر بدائيات النوى المتعايشة في السيتوبلازم ، وتؤدي الوظائف المقابلة. هذه الحقيقة ، بالإضافة إلى تشابه نظام التخليق الحيوي للبروتين في الميتوكوندريا والبلاستيدات بدائيات النوى ، تعتبر دليلاً على التكاثر التكافلي. والدليل أيضًا أن الميتوكوندريا والبلاستيدات الخضراء تتكاثر من تلقاء نفسها ، ولا يتم بناؤها أبدًا بواسطة الخلية من نقطة الصفر.
لصالح الفرضية الثالثة ، الخيمرية ، عن أصل الخلايا حقيقية النواة حجم كبيرالجينوم الخاص بهم ، والذي يتجاوز الجينوم البكتيري بآلاف المرات أو أكثر ، بالإضافة إلى مجموعة متنوعة من البروتينات المركبة الموجودة في مجموعات مختلفةبدائيات النوى. من الواضح أنه خلال تطور حقيقيات النوى ، أصبح جينومها أكثر تعقيدًا ، وتضاعف ، وظهرت العديد من الجينات التنظيمية فيه. ولكن لا يزال من الممكن أن تحدث الزيادة الأولية في حجم الجينوم بسبب مزيج من جينومات العديد من بدائيات النوى.
ربما في العصور القديمة ، اكتسبت بدائيات نوى معينة القدرة على البلعمة ، وأكلت بهذه الطريقة ، من بين أشياء أخرى ، بدائيات النوى الأخرى ، التي لم يتم هضمها دائمًا. احتوى جينومهم على جينات مفيدة للمضيف ، وقام بتضمينها في جينومه. ربما أصبحت بعض بدائيات النوى التي كانت بالداخل عضيات ، والتي تجمع بين الفرضية الكيميرية والتكافل.
نظرًا لأن جميع الفرضيات المذكورة أعلاه لها قوية و الجوانب الضعيفة، وأيضًا لا تستبعد إلى حد كبير مواقف بعضنا البعض ، إذن ، في رأينا ، يمكن أن تلعب مجموعة من العوامل التي وصفتها فرضيات مختلفة دورًا في أصل هياكل الخلايا حقيقية النواة.
وتجدر الإشارة أيضًا إلى أنه وفقًا للفرضية التكافلية ، يمكن اعتبار أصل عضيات الغشاء الأخرى مثل الفجوات ، ومركب جولجي ، وما إلى ذلك ، بمثابة تبسيط إضافي ، على سبيل المثال ، للميتوكوندريا.
نلاحظ أيضًا أن مزيج البروتينات من بدائيات النوى المختلفة في حقيقيات النوى قد لا يشير على الإطلاق إلى خيمرية الأولى. من الممكن أن تؤدي بعض الوظائف الأنزيمية فقط بروتينات ذات بنية معينة. وقد وصل تطور حقيقيات النوى إلى هذا مرارًا وتكرارًا ، بغض النظر عن تطور مجموعات بدائيات النوى التي ليست أسلافها. لنفترض أنه كان هناك تقارب على المستوى الجزيئي.
ظهور حقيقيات النوى هو حدث كبير. لقد غيرت بنية المحيط الحيوي وفتحت فرصًا جديدة بشكل أساسي للتطور التدريجي. إن الخلية حقيقية النواة هي نتيجة تطور طويل لعالم بدائيات النوى ، عالم تكيفت فيه الميكروبات المتنوعة مع بعضها البعض وبحثت عن طرق للتعاون الفعال.
مخطط الخط الزمني (تكرار)

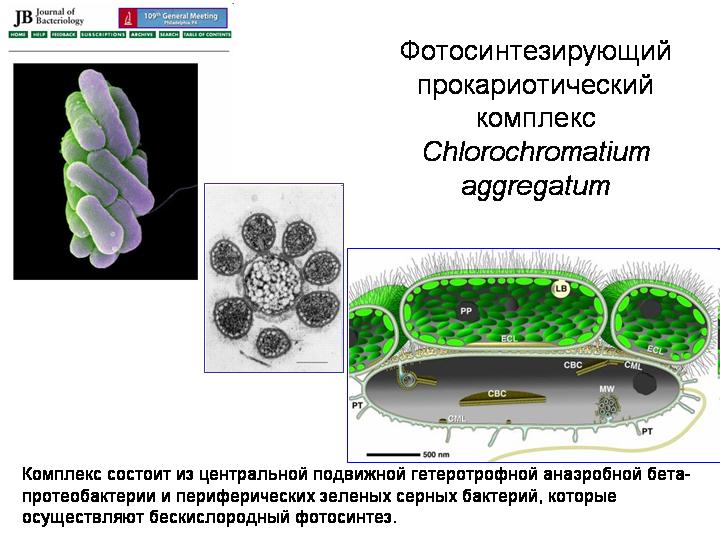
مجمع بدائيات النوى الضوئي مجمع الكلوروكروماتيوم.
نشأت حقيقيات النوى نتيجة تعايش عدة أنواع من بدائيات النوى. بدائيات النوى بشكل عام معرضة تمامًا للتكافل (انظر الفصل 3 في ولادة التعقيد). هنا نظام تكافلي مثير للاهتمام يُعرف باسم مجموع الكلوروكروماتيوم. يعيش في البحيرات العميقة ، حيث توجد ظروف نقص الأكسجين في العمق. المكون المركزي هو بكتيريا بيتا البروتينية غيرية التغذية المتنقلة. حولها ، تتكون الأكوام من 10 إلى 60 بكتيريا كبريتية خضراء ضوئية. جميع المكونات متصلة بواسطة النواتج الغشاء الخارجيالبكتيريا المركزية. معنى المجتمع هو أن بكتيريا بيتا المتنقلة تجر الشركة بأكملها إلى أماكن مواتية لحياة بكتيريا الكبريت شديدة الحساسية ، وتشارك بكتيريا الكبريت في عملية التمثيل الضوئي وتوفر الطعام لأنفسهم وبكتيريا بيتا. ربما كانت بعض الجمعيات الميكروبية القديمة من هذا النوع تقريبًا هي أسلاف حقيقيات النوى.
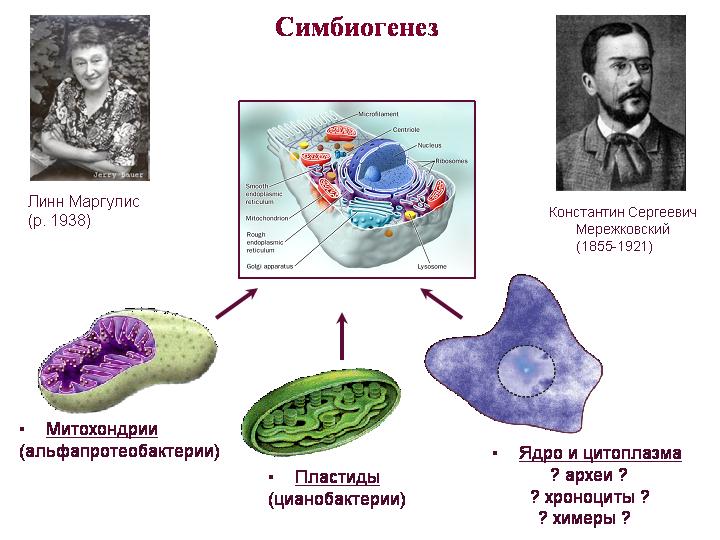
نظرية التكافل. ميريزكوفسكي ، مارغوليس. الميتوكوندريا هم من نسل ألفا بروتيوباكتيريا ، والبلاستيدات من نسل البكتيريا الزرقاء. من الصعب فهم من كان سلف كل شيء آخر ، أي السيتوبلازم والنواة. تجمع نواة وسيتوبلازم حقيقيات النوى بين سمات العتائق والبكتيريا ، ولها أيضًا العديد من الميزات الفريدة.
حول الميتوكوندريا. ربما كان اكتساب الميتوكوندريا (وليس النواة) هو اللحظة الحاسمة في تطور حقيقيات النوى. تم نقل معظم جينات الميتوكوندريا السلفية إلى النواة ، حيث أصبحت تحت سيطرة الأنظمة التنظيمية النووية. هذه الجينات النووية من أصل الميتوكوندريا لا ترمز فقط بروتينات الميتوكوندريا ، ولكن أيضًا العديد من البروتينات التي تعمل في السيتوبلازم. يشير هذا إلى أن تعايش الميتوكوندريا لعب دورًا أكثر أهمية في تكوين الخلية حقيقية النواة أكثر مما كان متوقعًا.
يتطلب التعايش بين جينومين مختلفين في خلية واحدة تطوير نظام فعال لتنظيمهما. ومن أجل إدارة عمل جينوم كبير بشكل فعال ، من الضروري عزل الجينوم عن السيتوبلازم ، الذي يحدث فيه التمثيل الغذائي والآلاف من تفاعلات كيميائية. يفصل الغلاف النووي فقط الجينوم عن العمليات الكيميائية المضطربة للسيتوبلازم. يمكن أن يصبح اكتساب المتكافلين (الميتوكوندريا) حافزًا مهمًا لتطوير النظم التنظيمية للنواة والجينات.
الأمر نفسه ينطبق على التكاثر الجنسي. يمكنك العيش بدون تكاثر جنسي طالما أن الجينوم الخاص بك صغير بما يكفي. الكائنات الحية ذات الجينوم الكبير ، ولكنها خالية من التكاثر الجنسي ، محكوم عليها بالانقراض السريع ، مع استثناءات نادرة.
![]()
بكتيريا ألفا - تضمنت هذه المجموعة أسلاف الميتوكوندريا.
Rhodospirillum هو كائن حي دقيق مذهل يمكن أن يعيش بسبب عملية التمثيل الضوئي ، بما في ذلك في ظل الظروف اللاهوائية ، وككائن هوائي غير متجانسة ، وحتى كيميائي كيميائي هوائي. يمكن ، على سبيل المثال ، أن ينمو بسبب الأكسدة أول أكسيد الكربونثاني أكسيد الكربون دون استخدام أي مصادر طاقة أخرى. بالإضافة إلى كل هذا ، فهو يعرف أيضًا كيفية إصلاح النيتروجين في الغلاف الجوي. هذا هو ، هو في أعلى درجةكائن عالمي.
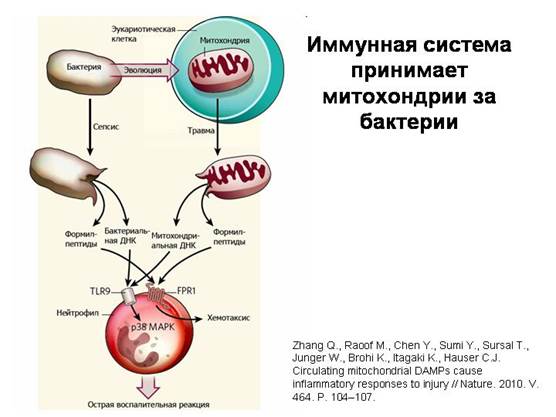
يخطئ جهاز المناعة في الميتوكوندريا على أنها بكتيريا. عندما تدخل الميتوكوندريا التالفة إلى مجرى الدم أثناء الإصابة ، فإنها تطلق جزيئات مميزة توجد فقط في البكتيريا والميتوكوندريا (DNA دائري من النوع البكتيري والبروتينات التي تحمل فورميل ميثيونين حمض أميني معدل خاص في أحد نهاياتها). هذا يرجع إلى حقيقة أن جهاز تخليق البروتين في الميتوكوندريا ظل كما هو الحال في البكتيريا. الخلايا الجهاز المناعي- العدلات - تتفاعل مع هذه المواد الميتوكوندريا بنفس الطريقة التي تتفاعل بها مع المواد البكتيرية ، وبمساعدة نفس المستقبلات. هذا هو أوضح تأكيد على الطبيعة البكتيرية للميتوكوندريا.
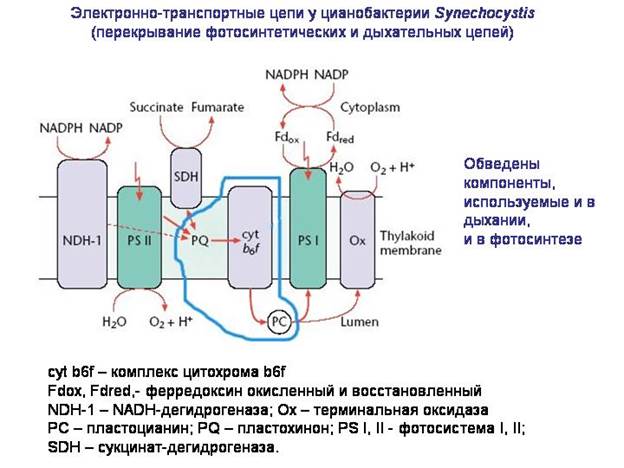
الوظيفة الأساسيةالميتوكوندريا - تنفس الأكسجين. على الأرجح ، كان الحافز لربط السلف اللاهوائي للنواة والسيتوبلازم مع "البروتوميتوكوندريا" هو الحاجة للحماية من عمل سامالأكسجين.
من أين حصلت البكتيريا ، بما في ذلك البكتيريا البروتينية ، على الأنظمة الجزيئية اللازمة لها تنفس الأكسجين؟ يبدو أنها كانت تستند إلى الأنظمة الجزيئية لعملية التمثيل الضوئي. تم تكييف سلسلة نقل الإلكترون ، التي تشكلت في البكتيريا كجزء من جهاز التمثيل الضوئي ، من أجل تنفس الأكسجين. في بعض البكتيريا ، لا تزال أجزاء من سلاسل نقل الإلكترون تُستخدم في وقت واحد في كل من التمثيل الضوئي والتنفس. على الأرجح ، كانت أسلاف الميتوكوندريا عبارة عن بكتيريا ألفا بروتينية غير متجانسة هوائية ، والتي بدورها تنحدر من بكتيريا ألفا البروتينية الضوئية ، مثل رودوسبيريلوم.
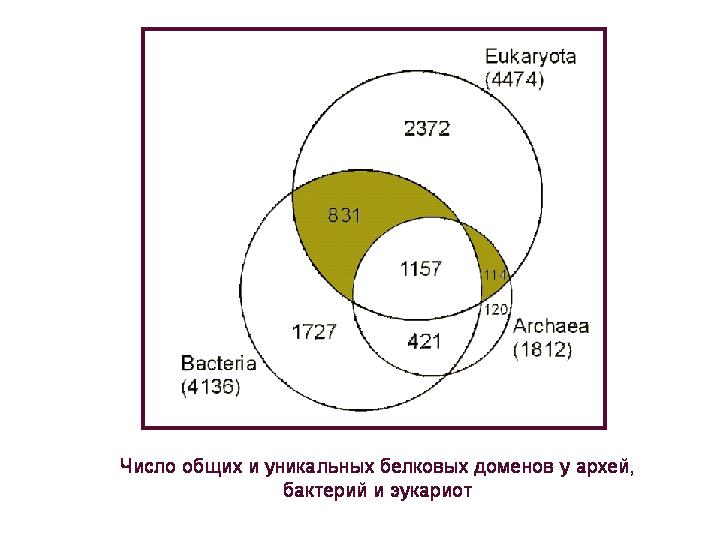
عدد مجالات البروتين الشائعة والفريدة من نوعها في العتائق والبكتيريا وحقيقيات النوى. مجال البروتين هو جزء من جزيء بروتين له وظيفة محددة وبنية مميزة ، أي سلسلة من الأحماض الأمينية. يحتوي كل بروتين ، كقاعدة عامة ، على واحد أو أكثر من هذه الوحدات أو المجالات الهيكلية والوظيفية.
4.5 ألف مجال من البروتينات التي تمتلكها حقيقيات النوى يمكن تقسيمها إلى 4 مجموعات: 1) متوفرة فقط في حقيقيات النوى ، 2) مشتركة بين جميع الملكات الثلاث ، 3) شائعة في حقيقيات النوى والبكتيريا ، ولكنها غائبة في العتائق ؛ 4) شائعة في حقيقيات النوى والعتائق ، ولكنها غائبة في البكتيريا. سننظر في اثنين المجموعات الأخيرة(يتم تمييزها بالألوان في الشكل) ، لأنه بالنسبة لهذه البروتينات يمكن للمرء أن يتحدث ببعض اليقين عن أصلها: جرثومي أو عتيق ، على التوالي.
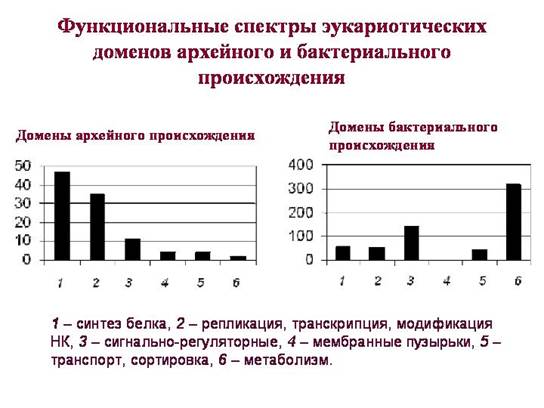
لحظة مهمةمن حيث أن المجالات حقيقية النواة التي يُفترض أنها موروثة من البكتيريا ومن العتائق لها وظائف مختلفة بشكل كبير. تلعب المجالات الموروثة من العتائق (يظهر طيفها الوظيفي في الرسم البياني الأيسر) دورًا رئيسيًا في حياة الخلية حقيقية النواة. من بينها ، المجالات المرتبطة بتخزين واستنساخ وتنظيم وقراءة المعلومات الجينية هي السائدة. تنتمي غالبية المجالات "البدائية" إلى تلك المجموعات الوظيفية التي يحدث فيها تبادل الجينات الأفقي في بدائيات النوى بشكل أقل تكرارًا. على ما يبدو ، استقبلت حقيقيات النوى هذا المركب عن طريق الوراثة المباشرة (الرأسية) من العتائق.
من بين مجالات الأصل البكتيري ، هناك أيضًا بروتينات مرتبطة بعمليات المعلومات ، لكنها قليلة. يعمل معظمهم فقط في الميتوكوندريا أو البلاستيدات. الريبوسومات حقيقية النواة في السيتوبلازم من أصل بدائي ، وريبوزومات الميتوكوندريا والبلاستيدات من أصل بكتيري.
من بين المجالات البكتيرية لحقيقيات النوى ، فإن حصة البروتينات المنظمة للإشارة أعلى بكثير. ورثت حقيقيات النوى من البكتيريا العديد من البروتينات المسؤولة عن آليات استجابة الخلية للعوامل بيئة خارجية. وأيضًا - العديد من البروتينات المرتبطة بعملية التمثيل الغذائي (لمزيد من التفاصيل ، انظر الفصل 3 ، "ولادة التعقيد").
حقيقيات النوى لها:
Archaeal "الأساسية" (آليات العمل مع المعلومات الجينية وتخليق البروتين)
"المحيط" البكتيري (التمثيل الغذائي ونظم تنظيم الإشارات)
أبسط سيناريو: ARCHEIA ابتلع BACTERIA (أسلاف الميتوكوندريا والبلاستيدات) وجميع ما لديها علامات بكتيريةتم شراؤها منهم.
· هذا السيناريو بسيط للغاية لأن حقيقيات النوى تحتوي على العديد من البروتينات البكتيرية التي لا يمكن استعارتها من أسلاف الميتوكوندريا أو البلاستيد.
تحتوي حقيقيات النوى على العديد من المجالات "البكتيرية" التي لا تتميز بأي من البكتيريا الزرقاء (أسلاف البلاستيدات) أو البكتيريا البروتينية (أسلاف الميتوكوندريا). تم الحصول عليها من بعض البكتيريا الأخرى.
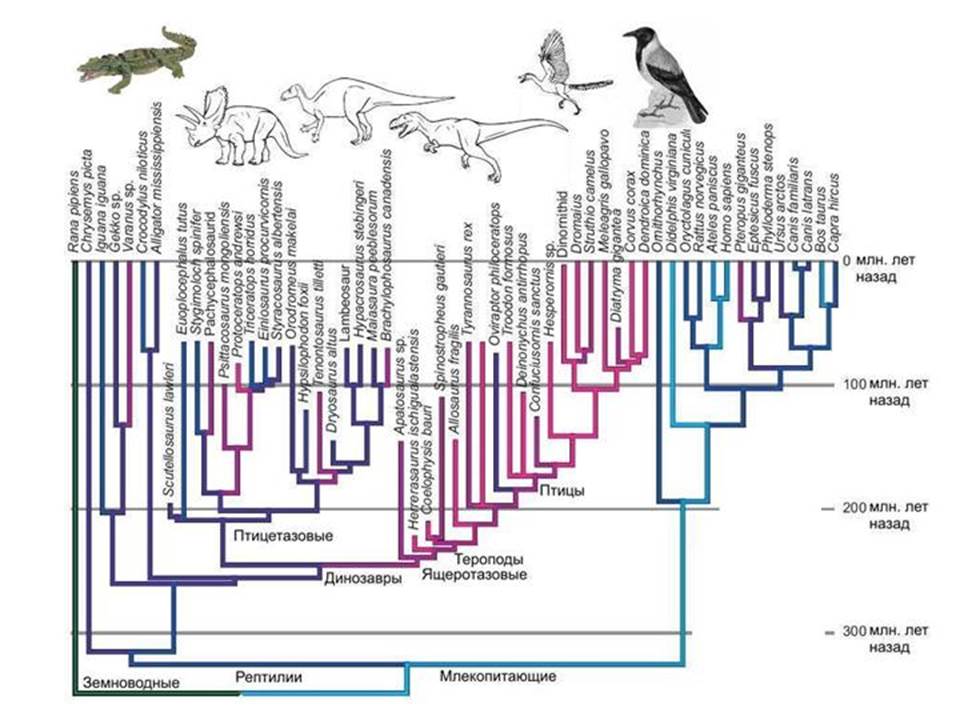
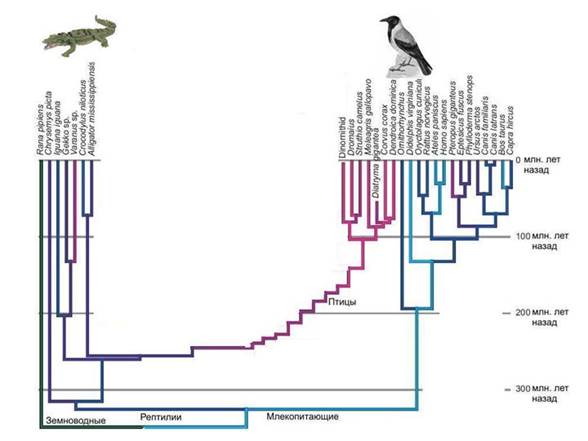
الطيور والديناصورات. من الصعب إعادة بناء حقيقيات النوى البدائية. من الواضح أن مجموعة بدائيات النوى القديمة التي أدت إلى ظهور النواة والسيتوبلازم لديها عدد من الميزات الفريدة التي لا تمتلكها بدائيات النوى التي نجت حتى يومنا هذا. وعندما نحاول إعادة بناء مظهر هذا السلف ، فإننا نواجه حقيقة أن مجال الفرضيات كبير جدًا.
تشبيه. من المعروف أن الطيور تنحدر من الديناصورات ، وليس من بعض الديناصورات المجهولة ، بل من الديناصورات بالكامل مجموعة معينة- الديناصورات maniraptor ، التي تنتمي إلى ذوات الأقدام ، والثيروبودات ، بدورها ، هي واحدة من مجموعات الديناصورات السحلية. تم العثور على العديد من الأشكال الانتقالية بين الديناصورات والطيور التي لا تطير.
لكن ماذا يمكن أن نقول عن أسلاف الطيور إذا لم يكن هناك سجل أحفوري؟ في أفضل حالةسنكتشف أن أقرب أقرباء الطيور هم التماسيح. لكن هل يمكننا إعادة إنشاء مظهر أسلاف الطيور المباشرة ، أي الديناصورات؟ بالكاد. ولكن في هذا الموقف بالتحديد نجد أنفسنا عندما نحاول استعادة مظهر سلف النواة والسيتوبلازم. من الواضح أن هذه كانت مجموعة من بعض الديناصورات بدائية النواة ، وهي مجموعة منقرضة ، على عكس الديناصورات الحقيقية ، لم تترك آثارًا مميزة في السجل الجيولوجي. العتائق الحديثة بالنسبة لحقيقيات النوى تشبه التماسيح الحديثةتجاه الطيور. حاول إعادة بناء هيكل الديناصورات التي لا تعرف إلا الطيور والتماسيح.
حجة لصالح حقيقة أن العديد من الميكروبات عاشت في عصر ما قبل الكمبري ، لا تشبه تلك الحالية. كانت ستراتوليتات البروتيروزويك أكثر تعقيدًا وتنوعًا من تلك الحديثة. الستروماتوليت هي نتاج نشاط حيوي للمجتمعات الميكروبية. ألا يعني هذا أن الميكروبات البدائية كانت أيضًا أكثر تنوعًا من الميكروبات الحديثة ، وأن العديد من مجموعات الميكروبات الأولية لم تنجو حتى يومنا هذا؟

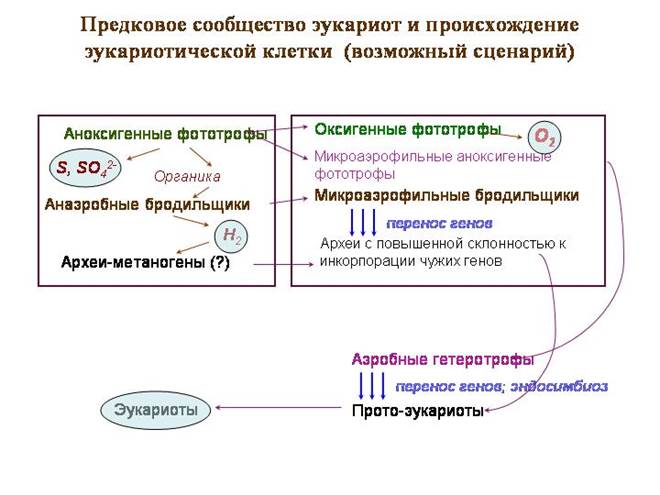
مجتمع أسلاف حقيقيات النوى وأصل الخلية حقيقية النواة (سيناريو محتمل)
إن "مجتمع الأسلاف" الافتراضي هو عبارة عن حصيرة بكتيرية نموذجية ، فقط في الجزء العلوي منها عاش أسلاف البكتيريا الزرقاء ، التي لم تتحول بعد إلى التمثيل الضوئي الأكسجين. كانوا يشتغلون في عملية التمثيل الضوئي غير المؤكسدة ، ولم يكن مانح الإلكترون من الماء ، ولكن كبريتيد الهيدروجين. مثل ثانويةتم إطلاق الكبريت والكبريتات.
كانت الطبقة الثانية مأهولة ببكتيريا التمثيل الضوئي الأرجواني ، بما في ذلك بكتيريا alphaproteobacteria ، أسلاف الميتوكوندريا. تستخدم البكتيريا الأرجوانية الضوء ذي الموجة الطويلة (الأحمر والأشعة تحت الحمراء). هذه الموجات لديها أفضل قوة اختراق. لا تزال البكتيريا البنفسجية تعيش غالبًا تحت طبقة من البكتيريا الزرقاء. تستخدم بكتيريا alphaproteobacteria الأرجواني أيضًا كبريتيد الهيدروجين كمانح للإلكترون.
في الطبقة الثالثة كانت هناك بكتيريا مخمرة تعالج مادة عضوية. البعض منهم ينبعث الهيدروجين كنفايات. هذا خلق قاعدة للبكتيريا التي تقلل الكبريتات. يمكن أن يكون هناك أيضًا عتائق ميثانوجينية. من بين الأركيا التي عاشت هنا أسلاف النواة والسيتوبلازم.
بدأت أحداث الأزمة بانتقال البكتيريا الزرقاء إلى عملية التمثيل الضوئي للأكسجين. كمتبرع للإلكترون ، بدأ استخدام البكتيريا الزرقاء بدلاً من كبريتيد الهيدروجين الماء العادي. لقد فتح هذا فرصًا كبيرة ، ولكن أيضًا عواقب سلبية. بدلاً من الكبريتات والكبريتات ، بدأ الأكسجين في إطلاقه أثناء عملية التمثيل الضوئي - وهي مادة شديدة السمية لجميع سكان الأرض القدامى.
أول من واجه هذا السم كان منتجه ، البكتيريا الزرقاء. ربما كانوا أول من طور وسائل الحماية ضده. تم تعديل سلاسل نقل الإلكترون التي تعمل في عملية التمثيل الضوئي وبدأت تعمل من أجل التنفس الهوائي. لم يكن الغرض الأصلي ، على ما يبدو ، هو الحصول على الطاقة ، ولكن فقط لتحييد الأكسجين.
بعد فترة وجيزة ، كان على سكان الطبقة الثانية من المجتمع - البكتيريا الأرجواني - تطوير أنظمة دفاع مماثلة. تمامًا مثل البكتيريا الزرقاء ، فقد طوروا أنظمة تنفس هوائية تعتمد على أنظمة التمثيل الضوئي. كانت البكتيريا ألفا بروتيوبروتي أرجوانية هي التي طورت أكثر سلسلة تنفسية مثالية ، والتي تعمل الآن في الميتوكوندريا في حقيقيات النوى.
في الطبقة الثالثة من المجتمع ، لا بد أن ظهور الأكسجين الحر قد تسبب في حدوث أزمة. تستخدم الميثانوجينات والعديد من مخفضات الكبريتات الهيدروجين الجزيئيبمساعدة إنزيمات الهيدروجين. لا تستطيع هذه الميكروبات أن تعيش في ظروف هوائية لأن الأكسجين يثبط إنزيمات الهيدروجين. العديد من البكتيريا التي تنتج الهيدروجين ، بدورها ، لا تنمو في بيئة لا توجد فيها كائنات دقيقة تستخدمه. من بين المخمرين ، يبدو أن المجتمع احتفظ بالأشكال التي تنبعث منها مركبات عضوية منخفضة (البيروفات ، واللاكتات ، والأسيتات ، وما إلى ذلك) كمنتجات نهائية. طورت هذه المخمرات وسائلها الخاصة للحماية من الأكسجين ، وهي أقل فعالية. وكان من بين الناجين العتائق - أسلاف النواة والسيتوبلازم.
ربما ، في هذه اللحظة من الأزمة ، حدث حدث رئيسي - إضعاف العزلة الجينية في أسلاف حقيقيات النوى وبداية الاقتراض النشط للجينات الأجنبية. دمجت حقيقيات النوى البدائية جينات العديد من المخمرات حتى أصبحت تخمر ميكرويروفيليك نفسها ، تخمر الكربوهيدرات إلى البيروفات وحمض اللاكتيك.
أصبح سكان الطبقة الثالثة - أسلاف حقيقيات النوى - الآن على اتصال مباشر مع السكان الجدد من الطبقة الثانية - البكتيريا الهوائية الهوائية ، التي تعلمت استخدام الأكسجين للحصول على الطاقة. أصبح استقلاب حقيقيات النوى البدائية والبكتيريا المتكوّنة من البكتيريا مكملين ، مما خلق المتطلبات الأساسية للتعايش. وقد حدد موقع البكتريا ألفا في المجتمع (بين الطبقة العليا ، التي تطلق الأكسجين ، والطبقة السفلية) دورها "كمدافعين" عن أسلاف حقيقية النواة من الأكسجين الزائد.
من المحتمل أن تكون حقيقيات النوى البدائية قد ابتلعت واكتسبت العديد من البكتيريا المختلفة مثل التعايش الداخلي. هذا النوع من التجارب لا يزال مستمرا. حقيقيات النوى أحادية الخلية، التي تحتوي على مجموعة كبيرة من المتعايشات داخل الخلايا. من بين هذه التجارب ، أثبت التحالف مع البكتيريا الهوائية الهوائية أنه الأكثر نجاحًا.
علم الأحياء وعلم الوراثة
وفقًا للفرضية التكافلية للعضيات الجسيمية للخلايا حقيقية النواة الشائعة حاليًا والتي لها جينوم خاص بها ، تتميز بأصل مستقل وتنشأ من خلايا بدائية النواةالمتعايشين. في البداية ، كانت كمية المعلومات في جينومات الخلية المضيفة ، من ناحية ، ومتلازمة الميتوكوندريا المفترضة للمريكزات والبلاستيدات الخضراء ، من ناحية أخرى ، قابلة للمقارنة على ما يبدو. في المستقبل ، يمكن أن تفقد جينومات المتعايشين جزءًا من وظائفها الجينية مع حركة الكتل الجينية في الجينوم ...
فرضيات أصل الخلايا حقيقية النواة.
1. التكافلية.
وفقًا للفرضية التكافلية الشائعة حاليًا ، تتميز العضيات الجسدية لخلية حقيقية النواة ، والتي لها جينوم خاص بها ، بأصل مستقل وتنشأ من خلايا بدائية النواة متعايشة. من المفترض أن تكون الخلية المضيفة عبارة عن بدائيات نوى لاهوائية ، قادرة فقط على الحركة الأميبية. نشأت الميتوكوندريا والسوط من خلال التغييرات في الخلايا المضيفة بدائيات النوى الهوائية والبكتيريا المتكافلة ، تشبه اللولبيات الحديثة. نشأت البلاستيدات الخضراء للخلايا النباتية الخضراء من المتعايشات - خلايا بدائية النواة من الطحالب الخضراء المزرقة. كان اكتساب الخلية أثناء تطور الأسواط بأجسام قاعدية قريبة من المريكزات ذات أهمية خاصة. هذا جعل من الممكن تكثيف النشاط الحركيوإنشاء آلية للانقسام.
مسألة أصل النواة صعبة. في البداية ، كانت كمية المعلومات في جينومات الخلية المضيفة ، من ناحية ، ومتعايشات الميتوكوندريا المفترضة ، والمريكزات ، والبلاستيدات الخضراء ، من ناحية أخرى ، قابلة للمقارنة على ما يبدو. في المستقبل ، يمكن أن تفقد جينومات المتعايشين بعض وظائفها الجينية مع نقل الكتل الجينية إلى جينوم الخلية المضيفة. رافق ذلك زيادة في حجم الجينوم النووي ، ومركزية التنظيم الجيني لوظائف الخلية ، وفقدان استقلالية الميتوكوندريا والعضيات الجسدية الأخرى. ومع ذلك ، لا يمكن استبعاد أن النواة هي أيضًا مشتق من تكافؤ داخل الخلايا. يفسر أصل الأغشية داخل البلازمية للشبكة السيتوبلازمية الخشنة والملساء والمركب الرقائقي والبنى الحويصلية الفراغية من خلال التحولات التطورية للغشاء الخارجي المغلف النووي. لا تشرح الفرضية التكافلية عددًا من الحقائق. وهكذا ، فإن بروتين عصيات ، الذي يتكون من أهداب وسواط من بدائيات النوى الحديثة ، يختلف عن بروتين توبولين لحقيقيات النوى. لم يكن للبكتيريا هياكل مع تكوين الأنابيب الدقيقة "9 + 2" أو "9 + 0" النموذجية للأسواط ، والأهداب ، والأجسام القاعدية أو المريكزات للخلايا حقيقية النواة.
2. الغزو.
تنطلق فرضية الانغماس لأصل الخلية حقيقية النواة من حقيقة أن شكل الأجداد كان بدائيات النوى الهوائية. احتوت على العديد من الجينومات ، كل منها مرتبط غشاء الخلية. نشأت العضيات الجسدية مع الحمض النووي ، وكذلك النواة ، نتيجة الانغماس وجلد شظايا القشرة جنبًا إلى جنب مع الجينوم ، تليها التخصص الوظيفي في النواة ، والميتوكوندريا ، والبلاستيدات الخضراء ، ومضاعفات الجينوم النووي ، و تطور الأغشية السيتوبلازمية. تشرح هذه الفرضية بشكل مرضٍ وجود غشاءين في غلاف النواة ، الميتوكوندريا و chporoplasts. واجهت صعوبات في شرح الاختلافات في تفاصيل عملية التخليق الحيوي للبروتين في العضيات الجسدية والسيتوبلازم للخلية حقيقية النواة. في الميتوكوندريا والبلاستيدات الخضراء ، تتوافق هذه العملية تمامًا مع تلك الموجودة في الخلايا بدائية النواة الحديثة. لا تستنفد فرضيات التكافؤ والابتلاع جميع وجهات النظر حول أصل نوع حقيقيات النوى للتنظيم الخلوي.
3. ظهور التعددية الخلوية.
كانت الأشكال الاستعمارية للأوليات غيرية التغذية هي سلف الكائنات متعددة الخلايا.
المرحلة 1: بالقرب من كل سوط كان هناك ثمرة من السيتوبلازم طوق أو جيب يتم دفع جزيئات الطعام إليه (البلعمة).
المرحلة 2: تخصص الخلايا البلعمية (وظيفة البلعمة الهضمية) والخلايا الحركية (حركة الوظيفة ؛ بقيت على المحيط). كثافة العمليات طبقة الخلايا البلعمية + تحويلة. kinocytoblast = البلعمة أو الحمة (متشنيكوف).
المرحلة 3: يتم تشكيل الفم في قطب واحد. الثقب => أدى إلى ظهور الإسفنج ، تجاويف الأمعاء ، إلخ.
وفقًا لهيجل: كرة ، مستعمرة من البحر ، عن طريق ملء السائل ، يتشكل كائن حي يشبه الفقاعة (blastea) ؛ الغزو مخلوق من طبقتين (gastrea) ؛ (يعتقد هيكل أن الأول كان الهضم البطني).
4. ميزات متعددة الخلايا.
تشكل الحيوانات متعددة الخلايا أكبر مجموعة من الكائنات الحية على هذا الكوكب ، حيث يبلغ عددها أكثر من 1.5 مليون نوع. يقود أصلهم من الأبسط ، فقد خضعوا لتحولات كبيرة في عملية التطور المرتبطة بتعقيد التنظيم.
أحد أهم سمات تنظيم الكائنات متعددة الخلايا هو الاختلاف المورفولوجي والوظيفي بين خلايا الجسم. في سياق التطور ، تخصصت خلايا متشابهة في جسم الحيوانات متعددة الخلايا في أداء وظائف معينة ، مما أدى إلى تكوينها.الأقمشة.
الأقمشة المختلفة مجتمعةللأعضاء والأعضاء في أنظمة الأعضاء.لتنفيذ العلاقة بينهم وتنسيق عملهم ،الأنظمة التنظيميةالجهاز العصبي والغدد الصماء. شكرا للعصبية التنظيم الخلطيأنشطة جميع الأنظمة ، يعمل الكائن متعدد الخلايا كنظام بيولوجي متكامل.
يرتبط ازدهار مجموعة من الحيوانات متعددة الخلايا بالمضاعفات الهيكل التشريحيو وظائف فسيولوجية. لذلك ، أدت زيادة حجم الجسم إلى التطور القناة الهضميةمما سمح لهم بتناول مواد غذائية كبيرة ، مما يوفر كمية كبيرة من الطاقة لتنفيذ جميع العمليات الحياتية. كفلت الأنظمة العضلية والهيكلية المتطورة حركة الكائنات الحية ، والحفاظ على شكل معين للجسم ، وحماية ودعم الأعضاء. القدرة على حركة نشطةسمح للحيوانات بالبحث عن الطعام والعثور على مأوى والاستقرار.
مع زيادة حجم جسم الحيوانات ، ظهرت الحاجة إلى المظهرأنظمة الدورة الدموية داخل النقل ،تقديم دعم الحياة للأنسجة البعيدة عن سطح الجسم "والأعضاء العناصر الغذائيةوالأكسجين ، وكذلك إزالة المنتجات النهائية الأيضية.
أصبح نظام نقل الدورة الدموية هذا عبارة عن دم من الأنسجة السائلة.
ذهب تكثيف النشاط التنفسي بالتوازي مع التطور التدريجيالجهاز العصبيوأجهزة الإحساس. كانت هناك حركة الإدارات المركزيةالجهاز العصبي إلى الطرف الأمامي لجسم الحيوان ، ونتيجة لذلك أصبح قسم الرأس معزولًا. سمح هذا الهيكل للجزء الأمامي من جسم الحيوان له بتلقي معلومات حول التغييرات في بيئةوالاستجابة لها بشكل مناسب.
تتنوع الحيوانات متعددة الخلايا بشكل كبير في البنية ، وخصائص الحياة ، ومختلفة في الحجم ، ووزن الجسم ، وما إلى ذلك.
يتوافق الدرس مع البرنامج والكتاب المدرسي لـ I.N. Ponomareva ، والتي تنص على دراسة علم الخلايا في الصف الحادي عشر. يتم عقد الدرس في شكل مناقشة علمية ومصمم لمدة 90 دقيقة.
أهداف الدرس: الإلمام أهم الفرضياتحول أصل الخلية حقيقية النواة والكائنات متعددة الخلايا ؛ تنمية القدرة على المجادلة والدفاع عن وجهة نظر المرء ؛ تعزيز ثقافة النقاش العام.
معدات: جداول تصور بنية الخلية وعضياتها الفردية ، رسم تخطيطي لظهور خلية حقيقية النواة ، رسومات تصور trichoplax وأشكال مختلفة من الطحالب.
أثناء الفصول
مدرس.مع تطور علم الخلية - علم الخلايا وظهور نتائج بحث جديدة لهذه المرحلة الابتدائية النظام البيولوجينشأت مناقشات ساخنة بين العلماء ، وتحولت إلى مناقشات علمية جادة. على مدار تاريخ علم الخلايا كعلم ، اكتسبت أي فرضية جديدة تقريبًا مؤيديها وخصومها.
المناقشة العلمية هي مناقشة عامة لسؤال أو مشكلة. اليوم في الدرس سنجري مناقشة نناقش خلالها اثنتين القضايا الحرجةعلم الخلايا: مشكلة أصل حقيقيات النوى الأولى ومشكلة أصل الكائنات متعددة الخلايا الأولى.
أنت تعلم أن الفصل تم تقسيمه سابقًا إلى مجموعات ستعرض وتدافع عن وجهات نظر مختلفة حول هاتين المسألتين اللتين نناقشهما. اسمحوا لي أن أذكركم بأسماء هذه المجموعات.
1. مؤيدو الفرضية الذاتية التولد عن أصل الخلية حقيقية النواة وعضياتها.
2. مؤيدو الفرضية التكافلية لأصل الخلية حقيقية النواة وعضياتها.
3. مؤيدو فرضية الأصل الاستعماري للكائنات أحادية الخلية الأولى.
4. مؤيدو فرضية أصل الكائنات متعددة الخلايا من الشركات العملاقة عن طريق خلوية خلاياها.
سيوضح ممثلو كل مجموعة في عرض تقديمي قصير جوهر وجهة نظر واحدة أو أخرى حول كل مشكلة من المشكلات التي تمت مناقشتها ، ثم يجيبون على الأسئلة والتعليقات والاعتراضات لكل من "خصومهم" العلميين وجميع الحاضرين في الفصل.
مشكلة الأصل عضيات الغشاءونواة الخلايا حقيقية النواة هي واحدة من أكثر موضوعات المناقشة الموضعية في علم الخلايا الحديث. لحل هذه المشكلة ، تم اقتراح عدة فرضيات مختلفة في العلم. دعنا نلقي نظرة على اثنين من أهمها.
فرضية التولد الذاتي لأصل الخلية حقيقية النواة
كلمة ممثلي الدفعة الأولى من الطلاب
ظهرت أولى حقيقيات النوى على الأرض منذ 1.1 - 1.4 مليار سنة (في عصر البروتيروزويك). كان هذا هو المعلم الرئيسي التالي في تطور ما قبل الكمبري بعد ظهور التمثيل الضوئي.
تنص الفرضية الذاتية ، أو التتابعية ، على أن خلية حقيقية النواة المعقدة تطورت مباشرة من خلية بدائية النواة: نتيجة لانغماس غشاء البلازما ، متبوعًا بإعادة ترتيبها لأداء وظائف معينة ، نشأت أهم العضيات. ومثل هذه العضيات المعقدة مثل الميتوكوندريا والبلاستيدات تنشأ من هياكل الغشاء داخل البلازمات للبنية الأنبوبية الموجودة في بدائيات النوى.
أسئلة
1. ما الذي يدعم وجهة النظر هذه؟ هل هناك أشكال انتقالية أي خلايا بدائية النواة مع "أساسيات" عضيات حقيقية النواة؟
إجابة.لا توجد مثل هذه الأشكال الأثرية بين الكائنات الحية. لكن دعونا نتذكر وجود الميزوزومات في الخلايا البكتيرية ، أي الهياكل الغشائية ذات الشكل الأنبوبي والحويصلي (الكروية) ، التي تشكلت عن طريق غزو غشاء البلازما في السيتوبلازم. من المفترض أن الميزوسومات تشارك في تكوين جدران الخلايا ، وتكرار الحمض النووي ، والعمليات الأخرى. من الممكن تمامًا أنه على أساس الميزوزومات نشأ مجمع الغشاء تدريجياً وتمايز ، مما أدى إلى ظهور عضيات مختلفة للخلية حقيقية النواة.
2. من المعروف أن التركيب الكيميائي الحيوي لعدد من العضيات ، ولا سيما الميتوكوندريا والبلاستيدات الخضراء ، يختلف اختلافًا كبيرًا عن تكوين غشاء البلازما في السيتوبلازم. ألا يشير هذا إلى أن أصلهم ليس من الأصل غشاء الخلية- أغشية البلازما؟
إجابة.في الواقع ، تركيبة البروتين في الميتوكوندريا والبلاستيدات الخضراء فريدة من نوعها. ولكن يمكن الحصول على هذه الأصالة مرة ثانية ، ويمكن أن تكون نتيجة للتكيف مع أداء وظائف معينة من قبل هذه العضيات.
مدرس.دعونا نستمع إلى حجج مؤيدي الأصل التكافلي لحقيقيات النوى وتحديد الفرضية الأكثر شيوعًا في الفصل.
الفرضية التكافلية لأصل الخلية حقيقية النواة
كلمة ممثلي المجموعة الثانية من الطلاب
تنطلق الفرضية التكافلية (تسمى الآن نظرية) من حقيقة أن حقيقيات النوى هي نتيجة تعايش بين بدائيات النوى المختلفة.
في وقت مبكر من بداية القرن العشرين. علماء النبات الروس أ. فامينتسين ، ب. كوزو بوليانسكي وك. طرح Merezhkovsky فرضية أن خلية النباتات الخضراء (حقيقيات النوى) تلقت البلاستيدات نتيجة تعايش خلية خالية من الكلوروفيل مع خلايا الطحالب الخضراء المزرقة. كانت هذه الفرضية الخاصة بالأصل التكافلي للخلية حقيقية النواة سابقة لعصرها ، وتم نسيانها ، وجذبت الانتباه مرة أخرى في منتصف القرن العشرين.
تم تطوير الفرضية المسماة بواسطة Lynn Sagan-Margulis (1983). وفقًا لهذه الفرضية ، فإن الخلية الأولية لبكتيريا بدائية النواة كبيرة ، بعد أن دخلت في تعايش مع خلايا الطحالب الخضراء المزرقة ، اكتسبت البلاستيدات. أدى التعايش مع الخلايا بدائية النواة غيرية التغذية إلى تحولها إلى ميتوكوندريا. بعض الخلايا ، كونها غيرية التغذية ، استولت على بكتيريا أخرى أصغر حجمًا أسباب غير معروفةلم يهضم. كانت الخلايا بدائية النواة الصغيرة الملتقطة قادرة على امتصاص الأكسجين. كانت هذه الخاصية مفيدة للخلية التي ابتلعت البكتيريا ، لأنها أعطتها قدرًا أكبر من الطاقة المخزنة في جزيئات ATP.
أسئلة
1. ما الدليل الذي يؤيد الأصل التكافلي للبلاستيدات والميتوكوندريا؟
إجابة.والدليل على هذا القول على النحو التالي.
1. الميتوكوندريا والبلاستيدات الخضراء محاطة بغشاء مزدوج.
2. تتكاثر هذه العضيات عن طريق الانقسام ، ولا تتبرعم من أي عضيات غشائية أخرى.
3. الميتوكوندريا والبلاستيدات الخضراء لها مادتها الجينية الخاصة ، والتي تشفر البروتينات الخاصة بها فقط.
4. الحمض النووي لهذه العضيات له بنية دائرية ، كما هو الحال في بدائيات النوى ، وليس خطيًا ، كما هو الحال في حقيقيات النوى.
5. تمتلك الميتوكوندريا والبلاستيدات الخضراء آلية تصنيع البروتينات والبروتينات الخاصة بها ، كما أن ريبوسوماتها بدائية النواة أكثر من حقيقيات النوى.
6. تتشابه بعض بروتينات هذه العضيات في تركيبها الأساسي مع البروتينات البكتيرية المماثلة ولا تشبه البروتينات السيتوبلازمية المقابلة.
2. من المعروف أنه لا يتم تصنيع جميع بروتينات الميتوكوندريا والبلاستيدات الخضراء وفقًا للحمض النووي الخاص بها ؛ يتم ترميز بعض بروتينات هذه العضيات في الحمض النووي للنواة. كيف يمكنك تفسير هذه الحقيقة؟
إجابة.في الواقع ، يتم تشفير جزء فقط من بروتيناتها في الحمض النووي الخاص بالميتوكوندريا والبلاستيدات الخضراء ، بينما يتم ترميز الباقي في الحمض النووي لنواة الخلية. على ما يبدو ، أثناء التطور ، "تدفق" جزء من المادة الوراثية من جينومات الميتوكوندريا والبلاستيدات الخضراء إلى الجينوم النووي. جينوم البلاستيدات الخضراء هو أكثر ضخامة من جينوم الميتوكوندريا ، وجينوم الميتوكوندريا لحقيقيات النوى الأدنى (على سبيل المثال ، الخميرة) هو أكثر ضخامة من الثدييات ، مما يؤكد بشكل غير مباشر الافتراض أعلاه.
3. نحن نعلم أنه بالإضافة إلى الميتوكوندريا والبلاستيدات الخضراء ، توجد أيضًا كمية صغيرة من الحمض النووي في الخلية حقيقية النواة في المريكزات وقاعدة الأسواط. هل نشأوا أيضًا بشكل تكافلي؟
إجابة.نعم. يمكن أن يؤدي التعايش مع البكتيريا الشبيهة باللولبيات إلى ظهور الأسواط ، من ناحية ، وإلى تحويل بعض هذه الخلايا الشبيهة باللولبيات إلى مريكزات من ناحية أخرى. ينص المفهوم الحديث للتكاثر التكافلي على أن الخلية حقيقية النواة قد تشكلت نتيجة لعدة أعمال متتالية من التكاثر التكافلي.
4. كيف نشأت بقية عضيات الخلية حقيقية النواة (مجمع جولجي ، EPS ، الجسيمات الحالة)؟
إجابة.من المفترض أن الشبكة الإندوبلازمية والليزوزومات وجهاز جولجي يمكن أن تكون قد نشأت عن طريق غزو غشاء البلازما.
5. كيف ، وفقًا لفرضية التكافؤ ، يمكن أن تنشأ نواة محاطة بغشاء مزدوج؟
إجابة.من المفترض أن النواة تطورت أيضًا نتيجة لعزل الحمض النووي من السيتوبلازم عن طريق غزو غشاء البلازما وتغليف المادة النووية.
6. من إجاباتك على السؤالين الأخيرين ، يترتب على ذلك أن ظهور خلية حقيقية النواة ليس فقط نتيجة تعايش وراثي للعديد من بدائيات النوى. ماذا تقول في ذلك؟
إجابة.نعم إنه كذلك. يعترف مؤيدو الفرضية التكافلية أن بعض هياكل الخلية حقيقية النواة نشأت نتيجة لتطورها الذاتي ومضاعفاتها.
7. لقد أحضرت بيانات كيميائية حيوية ووراثية ومجهرية إلكترونية السنوات الأخيرةلصالح الأصل التكافلي للخلية حقيقية النواة. ولكن هل هناك آثار تطورية ، تسمى الحفريات الحية ، تشير إلى روابط بين بدائيات النوى وحقيقيات النوى؟
إجابة.لوحظت هذه العلاقات بين الكائنات الحية النباتية والحيوانية. على سبيل المثال ، الأميبا التي تعيش بحرية بيلوميكسا بالستريس(مستنقع بيلوميكس) لا يحتوي على ميتوكوندريا ، ولكنه يحتوي على بكتيريا تكافلية تمدها بالتنفس.
مدرس.يبدو لي أنه تم طرح أسئلة كافية على ممثلي المجموعة الثانية. مؤيدو ما هي الفرضية التي عبرت ، في رأيك ، عن حجج أكثر إقناعًا وما هي الفرضية الأكثر شهرة في العلم؟
في الواقع ، تجد الفرضية التكافلية عددًا كبيرًا من المؤيدين. ولكن يجب أن نتذكر أن الاعتبارات المعبر عنها لصالح فرضية التكافؤ ليس لها تأكيد تجريبي ، وبالتالي لا يمكن اعتبارها نظرية. في رأيي ، من المهم أيضًا أن نتذكر أن التعايش لعب دورًا بارزًا ، ولكن ليس الدور الوحيد ، في ظهور الخلية حقيقية النواة. أهميةفي تكوين حقيقيات النوى ، كانت هناك أيضًا عمليات التطور الذاتي للخلايا بدائية النواة.
دعنا ننتقل إلى المشكلة التالية- مشكلة أصل الكائنات متعددة الخلايا.
فرضيات الأصل الاستعماري للميتازوان الأوائل
مدرس.أثبتت التطورات في علم الخلايا والبيولوجيا الجزيئية العلاقة الهيكلية والكيميائية الحيوية للكائنات أحادية الخلية ومتعددة الخلايا. كل هذا أكد فرضية أصل الحيوانات متعددة الخلايا من أشكال وحيدة الخلية. تتعلق الخلافات العلمية الرئيسية بالأسئلة التي تدور حول أي من الكائنات الأولية نشأت الحيوانات متعددة الخلايا ، وكيف بدت الحيوانات متعددة الخلايا الأولى؟ هناك العديد من الفرضيات حول أصل الحيوانات متعددة الخلايا من الأوليات. دعونا نفكر في بعضها.
كلمة ممثلي المجموعة الثالثة من الطلاب
لقد لوحظ بشكل صحيح أن هناك العديد من الفرضيات حول أصل الحيوانات متعددة الخلايا من البروتوزوا. الأكثر منطقية بينهم ، في رأينا ، يجب اعتباره الفرضية الاستعمارية.
اقترح إرنست هيجل أول فرضية استعمارية حول أصل الكائنات متعددة الخلايا. لقد طور "نظريته في المعدة" ، والتي بموجبها يكون السلف المشترك لجميع الحيوانات متعددة الخلايا مشابهًا ليرقة تجويف الأمعاء حرة السباحة - وهي الشعاب المرجانية ذات طبقتين من الخلايا.
اقترح عالم الأحياء الروسي إيليا إيليتش ميتشنيكوف فرضية أخرى: نشأت الحيوانات متعددة الخلايا من البروتوزوا السوطي الاستعماري القادر على البلعمة ، أي التقاط الطعام عن طريق الخلايا مع الهضم داخل الخلايا. تفترض هذه النظرية أن الخلايا السطحية التي استولت على الطعام تغرق في أعماق جسم المستعمرة من أجل الهضم ، مما يفسح المجال للخلايا الجائعة. والنتيجة هي كائن حي تقوم فيه خلايا الطبقة الداخلية بهضم الطعام ، وتلتقطه الطبقة الخارجية المكونة من الخلايا الجائعة. كما يؤدي وظائف الاستقبال والحركة والحماية. دعا متشنيكوف الكائن الحي المقترح البلعمة (كائن يتكون من خلايا تعمل في البلعمة).
أخيرًا ، تم طرح الفرضية الثالثة من قبل عالم الحيوان الألماني O. Byuchli. وفقا لها ، فإن الكائن الأصلي متعدد الخلايا يتكون من طبقتين من الخلايا ، علاوة على ذلك ، مع جانبه السفلي يزحف على طول الأرض ، و الجانب العلويلديها وظائف وقائية وحساسة. بعد أن التقى هذا الكائن الحي بأطعمة ذات أحجام كبيرة ، لف نفسه حوله وهضمه بالطبقة السفلية. أطلق Bütschli على هذا الكائن الحي المفترض اسم placula (حيوان على شكل كعكة مغلفة).
أسئلة
1. أي من هذه الفرضيات الثلاثة تفضل؟ أي منهم هو الأكثر شهرة في العلم؟
إجابة.من الصعب إعطاء الأفضلية لأي فرضية ، حيث لا توجد بيانات حفرية عن الخطوات الأولى في تطور الحيوانات متعددة الخلايا. يحاول العلماء العثور على تأكيد لفرضيات الأصل الاستعماري للحيوانات متعددة الخلايا من خلال مقارنة الكائنات الافتراضية مع الأشكال البدائية الحية للحيوانات متعددة الخلايا.
أكثر الحيوانات متعددة الخلايا بدائية المعروفة اليوم هي Trichoplax (الشكل 1). تم وصفه من قبل Artemy Vasilyevich Ivanov (1973). تم العثور على Trichoplax على الساحل الأوروبي للمحيط الأطلسي. هذا الحيوان له مظهر صفيحة رقيقة من طبقتين من الخلايا ذات الأسواط. يوجد في تجويف جسده خلايا هضمية منفصلة. بين طبقتين من الخلايا توجد خلايا تشبه الأميبا.
بعد أن درس Trichoplax ، A.V. توصل إيفانوف إلى استنتاج مفاده أنه يشبه إلى حد بعيد البلعمة الافتراضية phagocetella I.I. ميتشنيكوف هو ، في الواقع ، نموذجها الحي. من ناحية أخرى ، يبدو Trichoplax أيضًا وكأنه صفيحة. وهكذا ، فإن اكتشاف Trichoplax ، على غرار يرقة تجاويف الأمعاء ، يجمع بين فرضيات Haeckel (1866) و Mechnikov (1877) و Buchli (1884).
2. لفترة طويلة ، ارتبط أصل الحيوانات البدائية متعددة الخلايا بسوط استعمارية خضراء مثل eudorina و volvox و pandorina. ألا تعتبر الآن أشكالًا انتقالية إلى تعدد الخلايا؟
إجابة.في الواقع ، لم يعد أصل الحيوانات متعددة الخلايا مرتبطًا بالأسواط الخضراء الاستعمارية. تستند وجهة النظر هذه إلى حقيقة أن نموها الفردي وتكاثرها قريب من الطحالب ، وليس للحيوانات البدائية متعددة الخلايا.
3. ما الحيوانات البدائية متعددة الخلايا التي أدت إلى نشوء كائن استعماري افتراضي؟ هل كانت عبارة عن إسفنج أم تجاويف أمعاء أم ممثلين لنوع آخر من الحيوانات؟
إجابة.يُفترض أن عدة أنواع من الحيوانات متعددة الخلايا ذات مستويات مختلفة من التنظيم تنحدر من أسلاف تشبه البلعمة في آنٍ واحد: الإسفنج ، والمعيار المعوي ، والحيوانات البدائية ثلاثية الطبقات القريبة من مستورقات غير معوية تنتمي إلى الديدان المفلطحة. الإسفنج و تجاويف الأمعاء هي فروع التطور المسدودة. إن التخصص الضيق لهذه الأنواع ، وقدرتها على التكيف مع أسلوب حياة غير متحرك أو سلبي ، أغلق احتمالات تطورها التدريجي.
4. أنت تواصل الحديث عن أصل أول حيوانات متعددة الخلايا ، لكن تعددية الخلايا مميزة ليس فقط لممثلي مملكة الحيوان. كيف يمكن أن يكون هناك الفطريات متعددة الخلايا، النباتات؟
إجابة.يعتقد العلم الحديث أن النباتات ليس لديها خط حاد بين أحادية الخلية وتعدد الخلايا. نشأت تعدد الخلايا مرارًا وتكرارًا بشكل مستقل عن بعضها البعض في أقسام مختلفة من النباتات. لذلك ، في الطحالب الذهبية ، متعددة السوط ، الحرارية ، الخضراء ، يتم ملاحظة الأنواع ذات البنية الخيطية ، الاستعمارية متعددة الخلايا والسيفون (يتكون الجسم من خلية عملاقة متعددة النوى). يشير وجود مثل هذه الأشكال ، في رأينا ، إلى محاولات الاختراق إلى المستوى التالي من التنظيم - تعدد الخلايا ، حيث تؤدي الخلايا المختلفة وظائف مختلفة(الصورة 2).
أرز. 2. أشكال مختلفةالطحالب (1 - أحادية الخلية ؛
2 - استعماري 3 - متعدد النواة ؛ 4 - الخيطية)
المعلومات المتاحة للعلم الحديث عن تطور الأشكال متعددة الخلايا في عصر ما قبل الكمبري نادرة. هذا ينطبق بشكل خاص على الفطر. والسبب هو سوء الحفاظ على بقايا هذه الكائنات الحية. كما هو الحال في عدد من النباتات ، تخضع الفطريات للانتقال من بنية غير خلوية (سيفون ، على سبيل المثال ، في مخاط فطر العفن) إلى تعدد الخلايا. وهكذا ، في رأينا ، في مرحلة معينة من تطور الفطريات (منذ حوالي 1.3 مليار سنة) ، في بعضها ، تحولت الفطريات السيفون إلى تكوين متعدد الخلايا.
مدرس.دعونا نلخص ما قيل. الكائنات متعددة الخلايا ، وفقًا لـ العلم الحديث، ظهرت في وقت سابق بين النباتات والفطريات. في الحيوانات ، الأول أشكال متعددة الخلايانشأت منذ حوالي 700 مليون سنة. تم إعطاء بداية الحيوانات متعددة الخلايا من قبل الكائنات السوطية الاستعمارية القديمة غيرية التغذية.
فرضية الأصل غير الاستعماري للميتازوان الأوائل
كلمة ممثلي المجموعة الرابعة من الطلاب
يصر ممثلو المجموعة السابقة على أن أسلاف الحيوانات متعددة الخلايا البعيدة كانت مستعمرات من البروتوزوا. على الرغم من أن هذه وجهة نظر شائعة ، ولكنها ليست وجهة النظر الوحيدة.
نقدم فرضية تشير إلى أنه في عملية التطور ، تم تحويل الكائنات الأولية المفردة بالكامل إلى كائنات متعددة الخلايا. تم طرح هذه الفكرة من قبل عالم الحيوان الصربي الشهير إ. حاجي. في رأيه ، تطورت الحيوانات متعددة الخلايا من الأهداب متعددة النوى.
Ciliates لها هيكل معقد نوعًا ما. يتم تمثيل السيتوبلازم من خلال طبقتين - المحيطية والمركزية ، حيث يحدث الهضم داخل الخلايا. Ciliates لها فم خلوي ، بلعوم ، الفجوة المنقبضةمع القنوات المقربة. كل هذه الأجزاء المتباينة بشكل مختلف كائن وحيد الخلية- العضيات - يعتبر الحاج النموذج الأولي لأعضاء حيوان متعدد الخلايا. نعم ، هو يعتقد ذلك جلدنشأت الكائنات متعددة الخلايا من الطبقة الخارجية من السيتوبلازم (ectoplasm) ، وأمعائها - من الطبقة الداخلية من السيتوبلازم (Entoplasm).
يمكن أن يحدث الانتقال من حالة أحادية الخلية إلى حالة متعددة الخلايا في جسم الهدبية فورًا عن طريق تكوين حدود خلوية حول نوى فردية وأقسام متجاورة من السيتوبلازم. هذه العملية المفترضة تسمى "الخلوية" (من الكلمة اللاتينية السيلولا- الخلية) ، والفرضية نفسها تسمى نظرية الخلوية.
أسئلة
1. هل فرضية الحاجي لها أي دعم في علم أجنة الكائنات متعددة الخلايا السفلية ، مثل فرضية I.I. متشنيكوف؟
إجابة.لسوء الحظ ، لا توجد مثل هذه البيانات من علم الأجنة المقارن. لم يلاحظ أحد من قبل عملية الخلوية للشركات العملاقة في الطبيعة.
2. ألا تعتقد أن فرضية حاجي لا تصمد أمام النقد ، لأن المبدأ الأساسي الذي تقوم عليه هو شرير. تتعارض فرضية الخلوية ، في رأينا ، مع الأحكام الرئيسية للنظرية الخلوية ، وهي: أنها تعادل أجزاء الخلية الواحدة إلى أنسجة وأعضاء الكائنات متعددة الخلايا. ماذا يمكنك أن تقول عن هذا؟
إجابة.لن نكون قاطعين في الأحكام. لا تساوي فرضية حجي العضيات الهدبية مع أنسجة وأعضاء الكائنات الحية متعددة الخلايا. هي فقط تهم ممكن التعليمأعضاء مماثلة على أساس العضيات الهدبية.
3. على ماذا تستند فرضية حاجي ، إلى جانب حقيقة الشركات العملاقة بنية معقدةوهياكلها يمكن أن تصبح أعضاء لحيوانات متعددة الخلايا؟
إجابة.التأكيد غير المباشر للفرضية هو بعض التشابه بين الهدبيات والتوربينات - بدائية الديدان المفلطحة: أحجام متقاربة ، ونفس شكل الجسم ، وموقع عدد من الهياكل ، وما إلى ذلك. (الشكل 3 و 4).
مدرس. استمعنا إلى الحجج المؤيدة للأصل الاستعماري وغير الاستعماري لأول حيوانات متعددة الخلايا ، وتعرّفنا على وجهات النظر حول أصل النباتات والفطريات متعددة الخلايا. برأيك ، ما الفرضية - الأصل الاستعماري أو غير الاستعماري للحيوانات متعددة الخلايا - أكثر منطقية وثباتًا؟
بالتأكيد ، فرضية الأصل الاستعماري لأول حيوانات متعددة الخلايا. هذه الفرضية هي التي تجد أوسع اعتراف في العالم العلمي. ولكن يجب أن نتذكر أن الفرضية الثانية تتطور وتتغير باستمرار.
تلخيص الدرس
1. الدرجات للدرس.
2. الواجب المنزلي:
- دراسة فقرة "مشاكل علم الخلايا القابلة للنقاش" ؛
و - التعرف في المنزل على إحدى مشاكل علم الخلايا التي نوقشت في القرن الثامن عشر ، والتي تم إجراؤها بين مختلف مؤيدي التشكيل ؛ إعداد نظام الحجج لصالح وجهات النظر المختلفة.