organelle komórkowe
Organelle (organella) to niezbędne mikrostruktury dla wszystkich komórek, które pełnią określone funkcje życiowe. Rozróżnij organelle błoniaste i niebłonowe. Organelle błonowe oddzielone od otaczającej hialoplazmy błonami obejmują retikulum endoplazmatyczne, wewnętrzny aparat siatkowaty (kompleks Golgiego), lizosomy, peroksysomy i mitochondria.
Organelle komórkowe błony
Wszystkie organelle błonowe zbudowane są z błon elementarnych, których zasada organizacji jest podobna do budowy cytolematów. Procesy cytofizjologiczne wiążą się ze stałą adhezją, fuzją i separacją błon, podczas gdy możliwa jest adhezja i asocjacja tylko topologicznie identycznych monowarstw błon. Tak więc zewnętrzna warstwa dowolnej błony organelli skierowana w stronę hialoplazmy jest identyczna z wewnętrzną warstwą cytolemmy, a wewnętrzna warstwa skierowana w stronę jamy organelli jest podobna do zewnętrznej warstwy cytolemmy.
Retikulum endoplazmatyczne (retikulum endoplazmatyczne) to A ciągła struktura, utworzone przez system cysterny, kanaliki i spłaszczone woreczki. Na mikrografach elektronowych rozróżnia się retikulum endoplazmatyczne ziarniste (szorstkie, ziarniste) i nieziarniste (gładkie, ziarniste). Zewnętrzna strona sieci ziarnistej pokryta jest rybosomami, sieć nieziarnista jest pozbawiona rybosomów. Ziarnista retikulum endoplazmatyczne syntetyzuje (na rybosomach) i transportuje białka. Nieziarnista sieć syntetyzuje lipidy i węglowodany oraz uczestniczy w ich metabolizmie [np. hormony steroidowe w korze nadnerczy i komórkach Leydiga (sustenocytach) jąder; glikogen w komórkach wątroby]. Jedną z najważniejszych funkcji retikulum endoplazmatycznego jest synteza białek błonowych i lipidów dla wszystkich organelli komórkowych.
Wewnętrzny aparat siatkowaty lub zespół Golgiego (apparatus reticularis internus) to zbiór worków, pęcherzyków, cystern, rurek, płytek, ograniczonych błoną biologiczną. Elementy kompleksu Golgiego są połączone wąskimi kanałami. W strukturach kompleksu Golgiego dochodzi do syntezy i akumulacji polisacharydów, kompleksów białkowo-węglowodanowych, które są wydalane z komórek. W ten sposób powstają granulki wydzielnicze. Kompleks Golgiego jest obecny we wszystkich komórkach człowieka, z wyjątkiem erytrocytów i rogowych łusek naskórka. W większości komórek kompleks Golgiego znajduje się wokół lub w pobliżu jądra, w komórkach zewnątrzwydzielniczych - nad jądrem, w wierzchołkowej części komórki. Wewnętrzna wypukła powierzchnia struktur kompleksu Golgiego jest skierowana w stronę retikulum endoplazmatycznego, a zewnętrzna, wklęsła powierzchnia jest skierowana w stronę cytoplazmy.
Błony kompleksu Golgiego są utworzone przez ziarniste retikulum endoplazmatyczne i są przenoszone przez pęcherzyki transportujące. Z poza kompleks Golgiego stale pąkuje pęcherzyki wydzielnicze, a błony jego zbiorników są stale aktualizowane. Pęcherzyki wydzielnicze dostarczają materiału błonowego dla Błona komórkowa i glikokaliks. Zapewnia to odnowienie błony plazmatycznej.
Lizosomy (lysosomae) to pęcherzyki o średnicy 0,2-0,5 mikrona, zawierające około 50 rodzajów różnych enzymów hydrolitycznych (proteazy, lipazy, fosfolipazy, nukleazy, glikozydazy, fosfatazy). Enzymy lizosomalne są syntetyzowane na rybosomach ziarnistej retikulum endoplazmatycznego, skąd są transportowane przez pęcherzyki transportowe do kompleksu Golgiego. Pierwotne lizosomy pączkują z pęcherzyków kompleksu Golgiego. Obsługiwane w lizosomach kwaśne środowisko, jego pH waha się od 3,5 do 5,0. Błony lizosomów są odporne na zawarte w nich enzymy i chronią cytoplazmę przed ich działaniem. Naruszenie przepuszczalności błony lizosomalnej prowadzi do aktywacji enzymów i ciężkiego uszkodzenia komórki aż do jej śmierci.
W lizosomach wtórnych (dojrzałych) (fagolizosomach) biopolimery są trawione do monomerów. Te ostatnie są transportowane przez błonę lizosomalną do hialoplazmy komórki. Niestrawione substancje pozostają w lizosomie, w wyniku czego lizosom zamienia się w tzw. ciało szczątkowe o dużej gęstości elektronowej.
Peroksysomy (peroksysomy) to pęcherzyki o średnicy od 0,3 do 1,5 mikrona. Zawierają enzymy utleniające, które rozkładają nadtlenek wodoru. Peroksysomy biorą udział w rozkładzie aminokwasów, metabolizmie lipidów, w tym cholesterolu, puryn, w neutralizacji wielu substancje toksyczne. Uważa się, że błony peroksysomów powstają w wyniku pączkowania z nieziarnistej retikulum endoplazmatycznego, a enzymy są syntetyzowane przez polirybosomy.
Mitochondria (mitochondria), będące „stacjami energetycznymi komórki”, biorą udział w procesach oddychania komórkowego i przemiany energii w formy dostępne dla komórki. Ich główną funkcją jest utlenianie materia organiczna oraz syntezę trójfosforanu adenozyny (ATP). Mitochondria mają postać okrągłych, wydłużonych lub pręcikowatych struktur o długości 0,5-1,0 µm i szerokości 0,2-1,0 µm. Liczba, wielkość i lokalizacja mitochondriów zależy od funkcji komórki, jej potrzeb energetycznych. Wiele dużych mitochondriów w kardiomiocytach, włóknach mięśniowych przepony. Zlokalizowane są w grupach pomiędzy miofibrylami, otoczone ziarnistościami glikogenu i elementami nieziarnistej retikulum endoplazmatycznego. Mitochondria to organelle z podwójną błoną (każda o grubości około 7 nm). Pomiędzy zewnętrzną i wewnętrzną błoną mitochondrialną znajduje się przestrzeń międzybłonowa o szerokości 10-20 nm. Błona wewnętrzna tworzy liczne fałdy lub cristae. Zazwyczaj cristae są zorientowane wzdłuż długiej osi mitochondriów i nie docierają do przeciwnej strony błony mitochondrialnej. Dzięki cristae powierzchnia błony wewnętrznej dramatycznie wzrasta. Zatem powierzchnia cristae jednego mitochondrium hepatocytu wynosi około 16 μm. Wewnątrz mitochondrium, pomiędzy cristae, znajduje się drobnoziarnista macierz, w której widoczne są ziarnistości o średnicy około 15 nm (rybosomy mitochondrialne) i cienkie nici, które są cząsteczkami kwasu dezoksyrybonukleinowego (DNA).
Syntezę ATP w mitochondriach poprzedza tzw początkowe etapy występujący w hialoplazmie. W nim (pod nieobecność tlenu) cukry są utleniane do pirogronianu (kwasu pirogronowego). W tym samym czasie syntetyzowana jest niewielka ilość ATP. Główna synteza ATP zachodzi na błonach cristae w mitochondriach przy udziale tlenu (utlenianie tlenowe) i enzymów obecnych w matrix. Podczas tego utleniania generowana jest energia dla funkcji komórki, a także uwalniany jest dwutlenek węgla (CO 2) i woda (H 2 O). W mitochondriach cząsteczki informacyjnego, transportowego i rybosomalnego kwasu nukleinowego (RNA) są syntetyzowane na własnych cząsteczkach DNA.
Macierz mitochondrialna zawiera również rybosomy o wielkości do 15 nm. Jednak mitochondrialne kwasy nukleinowe i rybosomy różnią się od podobnych struktur w danej komórce. Tak więc mitochondria mają własny system niezbędny do syntezy białek i samoreprodukcji. Zwiększenie liczby mitochondriów w komórce następuje poprzez podzielenie jej na mniejsze części, które rosną, powiększają się i są zdolne do ponownego podziału.
Wykład 6. Organelle jednobłonowe
Organelle- trwałe, koniecznie obecne struktury komórkowe, które pełnią określone funkcje i mają określoną strukturę. W zależności od struktury organelle można podzielić na dwie grupy - błoniaste, które koniecznie obejmują membrany i niebłonowe. Z kolei organelle błonowe mogą być jednobłonowe - jeśli są zbudowane z jednej błony i dwubłonowe - jeśli otoczka organelli składa się z dwóch błon.
Organelle
http://pandia.ru/text/78/152/images/image002_207.gif" width="134" height="26">
Membrana Bez membrany
http://pandia.ru/text/78/152/images/image006_130.gif" width="146 height=26" height="26">Pojedyncza membrana Podwójna membrana
Rozważ budowę i funkcje organelli jednobłonowych.
Retikulum endoplazmatyczne (ER), retikulum endoplazmatyczne (EPR)- jeden błoniasty organoid. Jest to system membran tworzących zbiorniki i kanały, połączonych ze sobą i ograniczających pojedynczą przestrzeń wewnętrzną - wnęki EPS. Z jednej strony błony są połączone z błoną cytoplazmatyczną, z drugiej strony z zewnętrzną błoną jądrową. Istnieją trzy rodzaje EPS: szorstki (ziarnisty), zawierający rybosomy na swojej powierzchni i reprezentujący zestaw spłaszczonych woreczków połączonych ze sobą; gładki (ziarnisty), mające strukturę rurową, której błony nie zawierają rybosomów; I mediatorłączenie szorstkiego i gładkiego EPS.
ER odpowiada za transport substancji, tworzy przedziały („przedziały”), w których zachodzą różne reakcje. Na gładkim ER zachodzi synteza węglowodanów i lipidów, na szorstkim ER zachodzi synteza białek. Poprzez kanały ER cząsteczki białka są transportowane do kompleksu Golgiego, oddzielonego od ER w postaci pęcherzyków błonowych z cząsteczkami organicznymi, które łączą się z kompleksem Golgiego.
Aparat Golgiego, zespół Golgiego- organelle jednobłonowe. Jest to stos spłaszczonych „zbiorników” o rozszerzonych krawędziach, z którymi połączony jest układ małych jednobłonowych pęcherzyków (pęcherzyków Golgiego).
Pęcherzyki Golgiego koncentrują się głównie po stronie przylegającej do ER i wzdłuż obrzeży stosów. Uważa się, że przenoszą białka i lipidy do aparatu Golgiego, którego cząsteczki, przemieszczając się ze zbiornika do zbiornika, ulegają chemicznej modyfikacji. Najważniejszą funkcją kompleksu Golgiego jest usuwanie różnych tajemnic (enzymów, hormonów) z komórki, dlatego jest on dobrze rozwinięty w komórkach wydzielniczych - „system eksportu” komórki.
Aparat Golgiego ma dwie różne strony: formujące (proksymalne,cis-Polak) skierowany w stronę EPS, ponieważ stamtąd wchodzą małe pęcherzyki przenoszące białka i lipidy do aparatu Golgiego i dojrzały (dystalny,trans-Polak), z którego stale wyrastają pęcherzyki przenoszące białka i lipidy do różnych przedziałów komórki lub poza nią.
Każdy stos kompleksu Golgiego składa się zwykle z czterech do sześciu „cystern”, jest strukturalną i funkcjonalną jednostką aparatu Golgiego i nazywa się dictyosom. Liczba dictyosomów w komórce waha się od jednego do kilkuset. W komórkach roślinnych dictyosomy są izolowane. Aparat Golgiego zwykle znajduje się w pobliżu Jądro komórkowe(w komórkach zwierzęcych często w pobliżu centrum komórki).
Zewnętrzna część aparatu Golgiego ulega ciągłemu zużyciu w wyniku splatania się pęcherzyków, a część wewnętrzna jest stopniowo formowana w wyniku działania EPR.
Funkcje zespołu Golgiego: gromadzenie białek, lipidów, węglowodanów; modyfikacja i pakowanie w pęcherzyki membranowe (pęcherzyki) napływających substancji organicznych; wydzielanie białek, lipidów, węglowodanów; miejsce edukacji lizosomy. funkcja wydzielnicza jest najważniejsza, dlatego aparat Golgiego jest dobrze rozwinięty w komórkach wydzielniczych.
Lizosomy- organelle jednobłonowe. Są to małe pęcherzyki (średnica od 0,5 do 2 mikronów) zawierające zestaw enzymów hydrolitycznych. Enzymy są syntetyzowane na szorstkim ER, przenoszone do aparatu Golgiego, gdzie są modyfikowane i pakowane w pęcherzyki błonowe lizosomów. Lizosom może zawierać od 20 do 60 różnego rodzaju Enzymy hydrolizy. Rozkład substancji przez enzymy to tzw Liza.
Wyróżnić podstawowy I lizosomy wtórne. Pierwotne lizosomy nazywane są lizosomami, pączkującymi z aparatu Golgiego. Lizosomy wtórne nazywane są lizosomami, powstałymi w wyniku fuzji lizosomów pierwotnych z wakuolami endocytarnymi. W tym przypadku trawią substancje, które dostają się do komórki przez fagocytozę lub pinocytozę, więc można je nazwać wakuole trawienne.
autofagia- proces niszczenia struktur zbędnych dla komórki. Najpierw struktura przeznaczona do zniszczenia jest otoczona pojedynczą błoną, następnie powstały pęcherzyk błony łączy się z pierwotnym lizosomem, w wyniku czego powstaje również wtórny lizosom - autofagiczny wakuoli, w której ta struktura jest trawiona. Produkty trawienia są wchłaniane przez cytoplazmę komórki, ale część materiału pozostaje niestrawiona. Wtórny lizosom zawierający ten niestrawiony materiał nazywa się pozostałości ciała. W wyniku egzocytozy niestrawione cząsteczki są usuwane z komórki.autoliza- samozniszczenie komórki, które następuje w wyniku uwolnienia zawartości lizosomów. Normalnie autoliza zachodzi podczas metamorfoz (zanikanie ogona żabiej kijanki), inwolucji macicy po porodzie, w ogniskach martwicy tkanek.
Funkcje lizosomów. Zatem lizosomy są odpowiedzialne za wewnątrzkomórkowe niszczenie makrocząsteczek substancji organicznych... układ trawienny» komórek oraz do niszczenia zbędnych struktur komórkowych i pozakomórkowych.
Peroksysomy- organelle o budowie podobnej do lizosomów, pęcherzyki o średnicy do 1,5 μm z jednorodną matrycą zawierającą około 50 enzymów. Najważniejsze są enzymy oksydazy, katalizujący przeniesienie dwóch atomów wodoru z cząsteczek organicznych (aminokwasów, węglowodanów, Kwasy tłuszczowe) bezpośrednio do tlenu, tworząc w ten sposób nadtlenek wodoru, utleniacz niebezpieczny dla komórki:
AN2 + O2 → A + H2O2
Powstały nadtlenek wodoru katalaza wykorzystuje do utleniania różnych substratów: H2O2 + AN2 → A + 2H2O. W komórkach wątroby peroksysomy są duże i liczne; katalaza utlenia się etanol do aldehydu octowego. Nadmiar H2O2 i H2O2 powstały w innym miejscu komórki jest również niszczony przez katalazę (2H2O2 → 2H2O + O2) . Wraz z mitochondriami peroksysomy aktywnie wykorzystują tlen jako środek utleniający. Istnieje hipoteza, że peroksysomy to starożytne organelle, które pojawiły się przed mitochondriami: gdy w atmosferze pojawił się toksyczny dla większości komórek tlen, peroksysomy zmniejszyły jego stężenie w komórkach, jednocześnie wykorzystując go do reakcji oksydacyjnych.
Powstają peroksysomy początkujący od istniejących wcześniej, czyli należą do samoreprodukujących się organelli, mimo że nie zawierają DNA. Rosną dzięki przyjmowaniu w nich enzymów, enzymy peroksysomów powstają na szorstkim EPS iw hialoplazmie. W dużych peroksysomach wykrywa się gęsty rdzeń - nukleoid odpowiadający obszarowi kondensacji enzymu.
wakuole- organelle jednobłonowe. Vacuole to „pojemniki” wypełnione roztwory wodne organiczne i substancje nieorganiczne. ER i aparat Golgiego biorą udział w tworzeniu wakuoli.
Młody komórki roślinne zawierają wiele małych wakuoli, które następnie, gdy komórki rosną i różnicują się, łączą się ze sobą i tworzą jedną dużą centralna wakuola. centralna wakuola może zajmować do 95% objętości dojrzałej komórki, do której wypychane jest jądro i organelle Błona komórkowa. Błona otaczająca wakuolę rośliny nazywa się tonoplast. Nazywa się płyn wypełniający wakuole roślinne sok komórkowy. W skład soku komórkowego wchodzą rozpuszczalne w wodzie sole organiczne i nieorganiczne, monosacharydy, disacharydy, aminokwasy, końcowe lub toksyczne produkty przemiany materii (glikozydy, alkaloidy), niektóre barwniki (antocyjany). Z substancji organicznych częściej gromadzone są cukry i białka. Cukier – częściej w postaci roztworów, białka występują w postaci pęcherzyków EPR i aparatu Golgiego, po czym wakuole ulegają odwodnieniu, zamieniając się w ziarna aleuronowe.
Komórki zwierzęce zawierają małe wakuole trawienne i autofagiczne należący do grupy lizosomów wtórnych i zawierający enzymy hydrolityczne. Zwierzęta jednokomórkowe mają więcej kurczliwe wakuole , pełniąc funkcję osmoregulacji i wydalania.
Funkcje. Wakuole roślinne odpowiadają za gromadzenie wody i utrzymanie turgoru, gromadzenie rozpuszczalnych w wodzie metabolitów - zapas składniki odżywcze I sole mineralne, barwiące kwiaty i owoce, a tym samym przyciągające owady zapylające i rozsiewające nasiona. Wakuole trawienne i autofagiczne - niszczą makrocząsteczki organiczne; kurczliwe wakuole regulują ciśnienie osmotyczne komórki i usuwają z komórki zbędne substancje.
Retikulum endoplazmatyczne, aparat Golgiego, lizosomy, peroksysomy i wakuole tworzą pojedynczą sieć wakuolową komórki, których poszczególne elementy mogą się wzajemnie przekształcać.
|
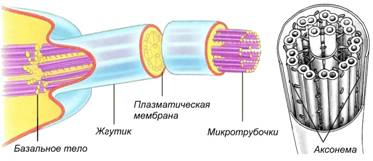
Aksonem tworzy 9 obwodowych par mikrotubul i jedna centralna para; przypomina koło rowerowe: oś to para mikrotubul, szprychy to specjalne białka, które łączą środkową parę mikrotubul z parami obwodowymi, a obręcz jest uformowana przez 9 par mikrotubul. Rzęski mają komórki drogi oddechowe człowiek, nabłonek jajowody. Wici u ludzi występują tylko w męskich komórkach rozrodczych - plemnikach. U prokariotów struktura wici jest inna i nie są one otoczone błoną.
Wiele komórek może mieć cienkie wypukłości na powierzchni - mikrokosmki zwiększyć powierzchnię (komórki jelito cienkie kręte kanaliki nerkowe). W przeciwieństwie do rzęsek nie są w stanie się poruszać; brakuje im mikrotubul aksonemów pod błoną.
Kluczowe terminy i pojęcia
1. Ziarnisty, gładki, pośredni EPS. 2. Cis-, trans-bieguny zespołu Golgiego. 3. Dictyosom. 4. Lizosomy pierwotne, wtórne. 5. Wakuole autofagiczne, autoliza. 6. Nukleoid peroksysomu. 7. Tonoplast. 8. Sieć wakuolowa komórki. 9. Ziarna aleuronowe. 10. Aksonem.
Podstawowe pytania przeglądowe
1. Charakterystyka EPR.
2. Budowa i funkcje zespołu Golgiego.
3. Charakterystyka lizosomów.
4. Budowa, funkcje i powstawanie peroksysomów.
5. Rodzaje wakuoli i ich funkcje.
6. Budowa wici i rzęsek u eukariontów.
7. Różnica między rzęskami a mikrokosmkami.
Wykład 6. Organelle jednobłonowe
Organelle- trwałe, koniecznie obecne struktury komórkowe, które pełnią określone funkcje i mają określoną strukturę. W zależności od struktury organelle można podzielić na dwie grupy - błoniaste, które koniecznie obejmują membrany i niebłonowe. Z kolei organelle błonowe mogą być jednobłonowe - jeśli są zbudowane z jednej błony i dwubłonowe - jeśli otoczka organelli składa się z dwóch błon.Organelle
Membrana Bez membrany
Pojedyncza membrana Podwójna membrana 

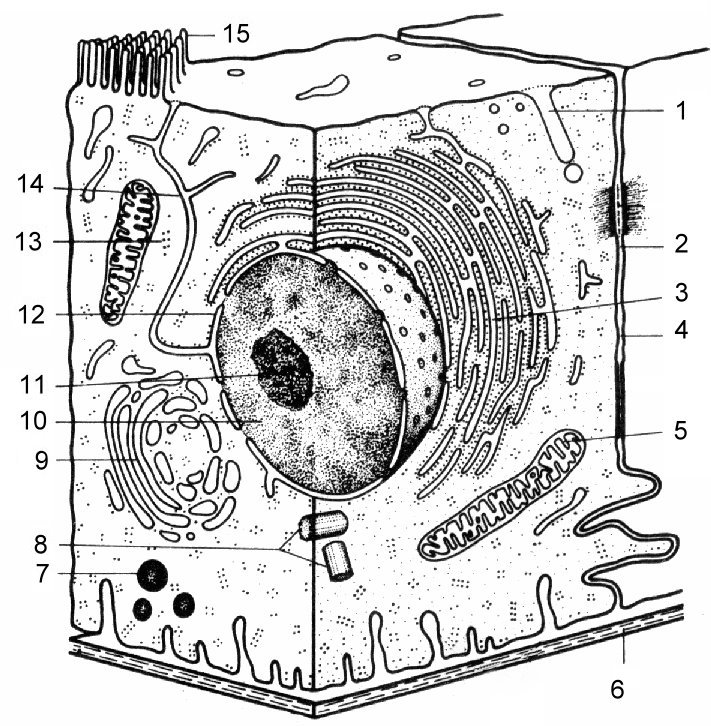
Ryż. . Schemat budowy komórki eukariotycznej
1 - kanalik pinocytarny; 2 - przerwa międzykomórkowa; 3 - szorstki EPS; 4 - plazmalemma; 5 - mitochondrium; 6 - błona podstawna; 7 - lizosom; 8 - centriole centrum komórki; 9 - zespół Golgiego; 10 - sok jądrowy, karioplasm z chromatyną; 11 - jąderko; 12 - otoczka jądrowa; 13 - rybosomy; 14 - kanaliki gładkie ER; 15 - mikrokosmki.
Rozważ budowę i funkcje organelli jednobłonowych. Retikulum endoplazmatyczne (ER), retikulum endoplazmatyczne (EPR)- organelle jednobłonowe. Jest to system membran tworzących zbiorniki i kanały, połączonych ze sobą i ograniczających pojedynczą przestrzeń wewnętrzną - wnęki EPS. Z jednej strony błony są połączone z błoną cytoplazmatyczną, z drugiej strony z zewnętrzną błoną jądrową. Istnieją trzy rodzaje EPS: szorstki (ziarnisty), zawierający rybosomy na swojej powierzchni i reprezentujący zestaw spłaszczonych woreczków połączonych ze sobą; gładki (ziarnisty), mające strukturę rurową, której błony nie zawierają rybosomów; I mediatorłączenie EPS szorstkiego i gładkiego EPS odpowiada za transport substancji, tworzy przedziały („przedziały”), w których zachodzą różne reakcje. Na gładkim ER zachodzi synteza węglowodanów i lipidów, na szorstkim ER zachodzi synteza białek. Poprzez kanały ER cząsteczki białka są transportowane do kompleksu Golgiego, oddzielonego od ER w postaci pęcherzyków błonowych z cząsteczkami organicznymi, które łączą się z kompleksem Golgiego. Aparat Golgiego, zespół Golgiego- organelle jednobłonowe. Jest to stos spłaszczonych „zbiorników” o rozszerzonych krawędziach, z którymi połączony jest system małych pęcherzyków jednobłonowych (pęcherzyków Golgiego).Pęcherzyki Golgiego koncentrują się głównie po stronie przylegającej do EPR i wzdłuż obwodu stosów. Uważa się, że przenoszą białka i lipidy do aparatu Golgiego, którego cząsteczki, przemieszczając się ze zbiornika do zbiornika, ulegają chemicznej modyfikacji. Najważniejszą funkcją kompleksu Golgiego jest usuwanie różnych tajemnic (enzymów, hormonów) z komórki, dlatego jest on dobrze rozwinięty w komórkach wydzielniczych - „system eksportu” komórki.Aparat Golgiego ma dwie różne strony: formujące (proksymalne,cis-Polak) skierowany w stronę EPS, ponieważ stamtąd wchodzą małe pęcherzyki przenoszące białka i lipidy do aparatu Golgiego i dojrzały (dystalny,trans-Polak), z którego stale pączkują pęcherzyki, przenosząc białka i lipidy do różnych przedziałów komórki lub poza nią.Każdy stos kompleksu Golgiego składa się zwykle z czterech do sześciu „zbiorników”, jest strukturalną i funkcjonalną jednostką aparatu Golgiego i jest nazywany dictyosom. Liczba dictyosomów w komórce waha się od jednego do kilkuset. W komórkach roślinnych dictyosomy są izolowane. Aparat Golgiego jest zwykle zlokalizowany w pobliżu jądra komórkowego (w komórkach zwierzęcych często w pobliżu środka komórki).Zewnętrzna część aparatu Golgiego jest stale zużywana w wyniku sznurowania pęcherzyków, a wewnętrzna część jest stopniowo formowana w wyniku działalność EPR.
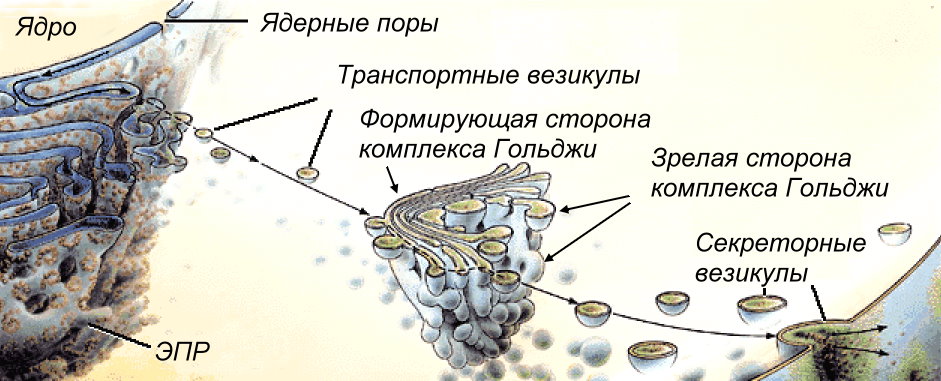
Ryż. kompleks Golgiego
Funkcje zespołu Golgiego: gromadzenie białek, lipidów, węglowodanów; modyfikacja i pakowanie w pęcherzyki membranowe (pęcherzyki) napływających substancji organicznych; wydzielanie białek, lipidów, węglowodanów; miejsce edukacji lizosomy. Najważniejsza jest funkcja wydzielnicza, dlatego aparat Golgiego jest dobrze rozwinięty w komórkach wydzielniczych. Lizosomy- organelle jednobłonowe. Są to małe pęcherzyki (średnica od 0,5 do 2 mikronów) zawierające zestaw enzymów hydrolitycznych. Enzymy są syntetyzowane na szorstkim ER, przenoszone do aparatu Golgiego, gdzie są modyfikowane i pakowane w pęcherzyki błonowe lizosomów. Lizosom może zawierać od 20 do 60 różnych typów enzymów hydrolitycznych. Rozkład substancji przez enzymy to tzw Liza.Wyróżnić podstawowy I lizosomy wtórne. Pierwotne lizosomy nazywane są lizosomami, pączkującymi z aparatu Golgiego. Lizosomy wtórne nazywane są lizosomami, powstałymi w wyniku fuzji lizosomów pierwotnych z wakuolami endocytarnymi. W tym przypadku trawią substancje, które dostają się do komórki przez fagocytozę lub pinocytozę, więc można je nazwać wakuole trawienne.A
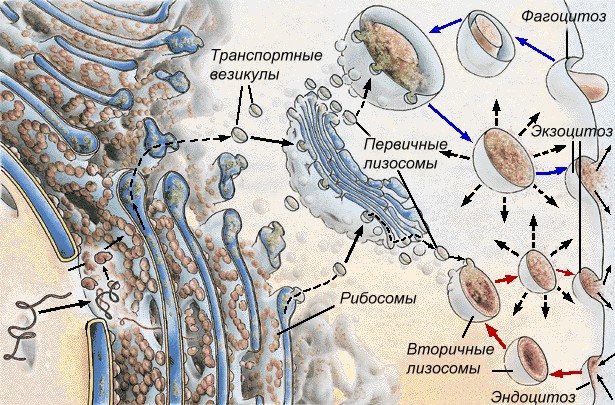
Ryż. Tworzenie lizosomów
Autofagia to proces niszczenia zbędnych struktur komórkowych. Najpierw struktura przeznaczona do zniszczenia jest otoczona pojedynczą błoną, następnie powstały pęcherzyk błony łączy się z pierwotnym lizosomem, w wyniku czego powstaje również wtórny lizosom - autofagiczny wakuola, w której ta struktura jest trawiona.P
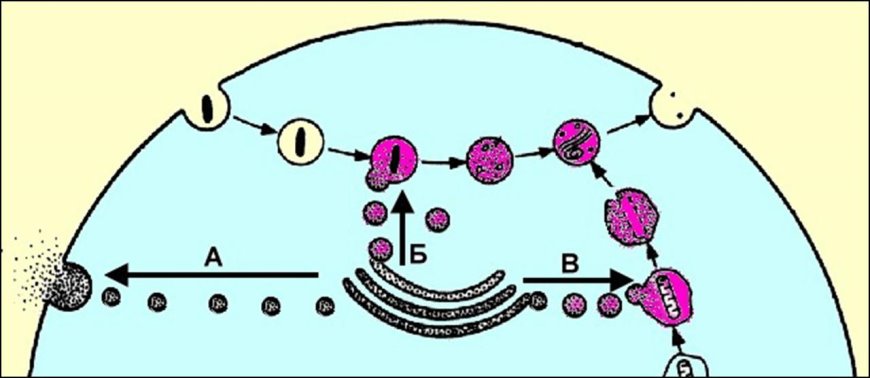
Ryż. . Tworzenie pęcherzyków egzocytarnych (A), wakuoli trawiennych (B) i wakuoli autofagicznych (C).
Produkty trawienia są przyswajane przez cytoplazmę komórki, ale część materiału pozostaje niestrawiona. Wtórny lizosom zawierający ten niestrawiony materiał nazywa się pozostałości ciała. W wyniku egzocytozy niestrawione cząsteczki są usuwane z komórki. autoliza- samozniszczenie komórki, które następuje w wyniku uwolnienia zawartości lizosomów. Normalnie autoliza zachodzi podczas metamorfoz (zanikanie ogona żabiej kijanki), inwolucji macicy po porodzie, w ogniskach martwicy tkanek. Funkcje lizosomów. Lizosomy są zatem odpowiedzialne za wewnątrzkomórkowe niszczenie makrocząsteczek substancji organicznych – „układu pokarmowego” komórki oraz za niszczenie zbędnych struktur komórkowych i pozakomórkowych.
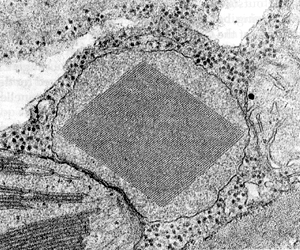
Ryż. . Peroksysom.
W centrum znajduje się nukleoid.
Peroksysomy- organelle o budowie podobnej do lizosomów, pęcherzyki o średnicy do 1,5 μm z jednorodną matrycą zawierającą około 50 enzymów. Najważniejsze są enzymy oksydazy, katalizujący przeniesienie dwóch atomów wodoru z cząsteczek organicznych (aminokwasów, węglowodanów, kwasów tłuszczowych) bezpośrednio do tlenu, z utworzeniem nadtlenku wodoru, utleniacza niebezpiecznego dla komórki:
AN 2 + O 2 → A + H 2 O 2
Powstały nadtlenek wodoru katalaza wykorzystuje do utleniania różnych substratów: H 2 O 2 + AN 2 → A + 2H 2 O. W komórkach wątroby peroksysomy są duże i wiele z nich katalaza utlenia alkohol etylowy do aldehydu octowego. Nadmiar H 2 O 2 i H 2 O 2, powstający w innym miejscu komórki, jest również niszczony przez katalazę (2H 2 O 2 → 2H 2 O + O 2) . Wraz z mitochondriami peroksysomy aktywnie wykorzystują tlen jako środek utleniający. Istnieje hipoteza, według której peroksysomy to starożytne organelle, które pojawiły się przed mitochondriami: kiedy w atmosferze pojawił się tlen, toksyczny dla większości komórek, peroksysomy zmniejszyły jego stężenie w komórkach, jednocześnie wykorzystując go do reakcji oksydacyjnych. początkujący z dotychczasowych, tj. są samoreplikującymi się organellami, mimo że nie zawierają DNA. Rosną dzięki przyjmowaniu w nich enzymów, enzymy peroksysomów powstają na szorstkim EPS iw hialoplazmie. W dużych peroksysomach wykrywa się gęsty rdzeń - nukleoid odpowiadający obszarowi kondensacji enzymu. wakuole- organelle jednobłonowe. Vacuole to „zbiorniki” wypełnione wodnymi roztworami substancji organicznych i nieorganicznych. EPS i aparat Golgiego biorą udział w tworzeniu wakuoli Młode komórki roślinne zawierają wiele małych wakuoli, które następnie w miarę wzrostu i różnicowania się komórek łączą się ze sobą i tworzą jedną dużą centralna wakuola. Centralna wakuola może zajmować do 95% objętości dojrzałej komórki, podczas gdy jądro i organelle są wypychane z powrotem do błony komórkowej. Błona otaczająca wakuolę rośliny nazywa się tonoplast. Nazywa się płyn wypełniający wakuole roślinne sok komórkowy. W skład soku komórkowego wchodzą rozpuszczalne w wodzie sole organiczne i nieorganiczne, monosacharydy, disacharydy, aminokwasy, końcowe lub toksyczne produkty przemiany materii (glikozydy, alkaloidy), niektóre barwniki (antocyjany). Z substancji organicznych częściej gromadzone są cukry i białka. Cukier – częściej w postaci roztworów, białka występują w postaci pęcherzyków EPR i aparatu Golgiego, po czym wakuole ulegają odwodnieniu, zamieniając się w ziarna aleuronowe.W komórkach zwierzęcych są małe wakuole trawienne i autofagiczne należący do grupy lizosomów wtórnych i zawierający enzymy hydrolityczne. Zwierzęta jednokomórkowe mają więcej kurczliwe wakuole, pełniąc funkcję osmoregulacji i wydalania. Funkcje. Wakuole roślinne odpowiadają za gromadzenie wody i utrzymywanie turgoru, gromadzenie rozpuszczalnych w wodzie metabolitów – rezerwowych składników pokarmowych i soli mineralnych, wybarwienie kwiatów i owoców, a tym samym przyciąganie zapylaczy i rozsiewaczy nasion. Wakuole trawienne i autofagiczne - niszczą makrocząsteczki organiczne; kurczliwe wakuole regulują ciśnienie osmotyczne komórki i usuwają z komórki zbędne substancje Retikulum endoplazmatyczne, aparat Golgiego, lizosomy, peroksysomy i wakuole formularzpojedyncza sieć wakuolowa komórki, których poszczególne elementy mogą się wzajemnie przekształcać. R
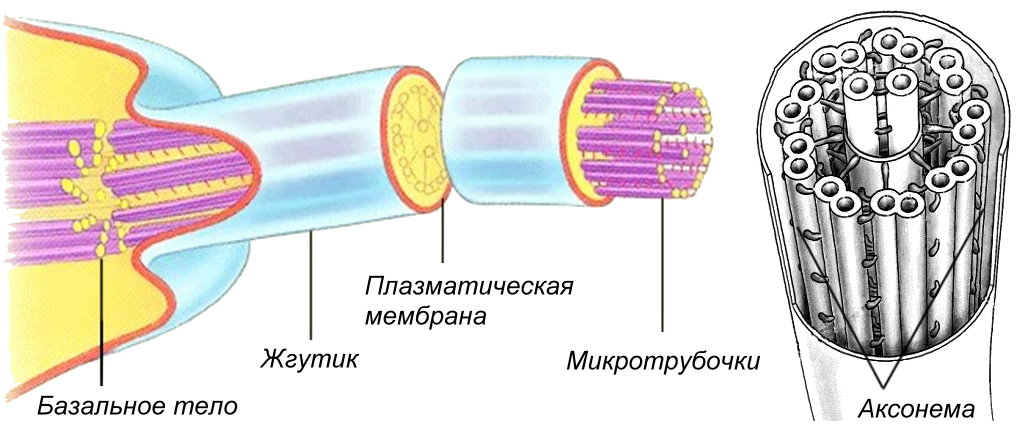
Ryż. Budowa wici i rzęsek u eukariontów
Jesnica i wici. Organelle te biorą udział w procesach ruchu i stanowią szkielet mikrotubul, tzw aksonem otoczony plazmalemmą. Długość rzęsek wynosi do 10 mikronów, wici różnią się tylko rozmiarem, a ich długość sięga 70 mikronów. U podstawy rzęsek i wici znajduje się ciało podstawowe, w którym znajduje się 9 trojaczków mikrotubul Aksonema jest utworzona z 9 obwodowych par mikrotubul i jednej centralnej pary, przypomina koło rowerowe: oś to para mikrotubul, szprychy to specjalne białka, które łączą środkową parę mikrotubule z parami obwodowymi, a brzeg tworzy 9 par mikrotubul. Rzęski mają komórki dróg oddechowych człowieka, nabłonek jajowodów. Wici u ludzi występują tylko w męskich komórkach rozrodczych - plemnikach. U prokariontów struktura wici jest inna i nie są one otoczone błoną.Wiele komórek może mieć cienkie wypustki na powierzchni - mikrokosmki w celu zwiększenia powierzchni (komórki jelita cienkiego, kręte kanaliki nerkowe). W przeciwieństwie do rzęsek nie są w stanie się poruszać; brakuje im mikrotubul aksonemów pod błoną. Kluczowe terminy i pojęcia 1. Ziarnisty, gładki, pośredni EPS. 2. Cis-, trans-bieguny zespołu Golgiego. 3. Dictyosom. 4. Lizosomy pierwotne, wtórne. 5. Wakuole autofagiczne, autoliza. 6. Nukleoid peroksysomu. 7. Tonoplast. 8. Sieć wakuolowa komórki. 9. Ziarna aleuronowe. 10. Aksonem. Podstawowe pytania przeglądowe
Charakterystyka EPR.
Wyjaśnienie.
Błona komórkowa: selektywna przepuszczalność; transport aktywny; zdolność do fagocytozy. Ściana komórkowa: zachowanie kształtu komórki; nadaje sztywność komórce.
Zewnętrzna błona komórkowa lub plazmalemma. Oddziela komórkę od otoczenie zewnętrzne, często razem ze ścianą komórkową. Jednak tylko prokarioty, rośliny i grzyby mają ścianę komórkową, podczas gdy zwierzęta nie. A membrana jest zawsze obecna. Grubość błony komórkowej wynosi 5–7 nm. Membrana jest otoczką o bardzo niezwykłych właściwościach. Ona nie ma stała forma, a ograniczona przez nią przestrzeń ma stałą objętość i ogólnie rzecz biorąc jest płynna, choć lepka. Fosfolipidy układają się w dwie warstwy - ogony do wewnątrz, głowy na zewnątrz. Nazywa się to dwuwarstwą lipidową. Ich ogony tworzą tę samą niemieszającą się z wodą fazę - warstwę hydrofobową, a ich głowy są skierowane w stronę środowisko wodne na zewnątrz i wewnątrz komórki.
W środowisku wodnym fosfolipidy zawsze układają się w dwuwarstwę i tworzą pęcherzyki. Ta właściwość zapewnia zamknięcie błony komórkowej: jeśli jej integralność zostanie naruszona, zostanie natychmiast przywrócona. Istnieją również białka błonowe, które dzielą się na obwodowe (słabo związane z błoną) i integralne (zlokalizowane w dwuwarstwie lipidowej).
Możliwy jest transport aktywny i bierny (dyfuzja) przez błonę.
Plazmalemma to bardzo złożona skorupa. Może zmieniać kształt i powierzchnię. Dzięki różnorodności białek może pomijać lub nie pomijać różnych zestawów substancji. Ale jest to półpłynna i nieuchronnie bardzo delikatna skorupa, która z trudem może uchronić komórkę przed poważnym uszkodzeniem uszkodzenie mechaniczne. Dlatego w wielu organizmach komórka jest również otoczona ścianą komórkową. Jest to sztywna, mała lub całkowicie nierozciągliwa formacja, znajdująca się na zewnątrz komórki. Z reguły zachowuje swój kształt do pewnego stopnia, jest elastyczny i mocny, w niektórych przypadkach jest bardzo mocny i ma odpowiednią grubość. Składa się z substancji wytwarzanych wewnątrz komórki, wydzielanych przez nią na zewnątrz i tam utwardzanych. Najczęściej podstawę ściany komórkowej stanowią polisacharydy. Ale czasami większośćściany są reprezentowane przez inne stałe substancje organiczne.
Ściana komórkowa to zewnętrzna ściana komórki błona cytoplazmatyczna oraz pełniących funkcje konstrukcyjne, ochronne i transportowe. Występuje w większości bakterii, archeonów, grzybów i roślin. Zwierzęta i wiele pierwotniaków nie ma ściany komórkowej.
W przeciwieństwie do plazmalemmy, ściana komórkowa jest nieprzepuszczalna dla większości substancji. Dlatego w niektórych miejscach dogodnych dla danej komórki w ściany komórkowe są pory. Przez pory przechodzą mostki cytoplazmatyczne, łączące ze sobą komórki roślinne - plazmodesmy.
Źródło. Kurs wykładów z biologii ogólnej. OE Kosterin. http://pisum.bionet.nsc.ru/kosterin/lectures/index.htm
Odpowiedź: 11221.
Organelle błonowe komórki.
Wypełnia uczeń 10 klasy „δ”.
Gurova Valeria
Kierownik: Sorokin Władimir Anatolijewicz
Wstęp…………………………3
Rozdział 1
Rozdział 2. PlastydyRozdział 3. Mitochondria
Rozdział 4. Retikulum endoplazmatyczne.
Rozdział 5
Rozdział 6
Rozdział 7
Wniosek
Bibliografia
Wstęp
Moja praca poświęcona jest badaniu organoidów błony komórkowej: ujawnieniu ich znaczenia, struktury i funkcji. Wybrany temat jest aktualny w naszych czasach, ponieważ jest integralną częścią biologii. Przedmiotem badań są m.in niezbędne elementy organizmy pełniące funkcje życiowe dla wszystkich żywych istot.
Cel: zbadanie organelli błonowych komórki, wyjaśnienie ich budowy i funkcji, poznanie roli, jaką odgrywają w życiu każdej komórki i organizmu jako całości.
Zadania:
1) Wyjaśnij, czym są organelle błonowe komórki i co zawierają.
2) mówić o budowie organelli błonowych komórki
3) określić ich znaczenie i funkcje Praca składa się z: wstępu, siedmiu rozdziałów, zakończenia oraz spisu piśmiennictwa.
Rozdział 1. Czym są błonowe organelle komórkowe.
Organelle to trwałe struktury wewnątrzkomórkowe, które mają określoną budowę i pełnią odpowiednie funkcje. Wszystkie organelle komórkowe dzielą się na dwie grupy: błonową i niebłonową. Organelle błonowe dzielą się na dwubłonowe i jednobłonowe. Składnikami dwubłonowymi są plastydy, mitochondria. Organelle jednobłonowe obejmują organelle układu wakuolowego - retikulum endoplazmatyczne, kompleks Golgiego, lizosomy, wakuole komórek grzybów roślinnych, wakuole pulsujące itp. Orgnoidy niebłonowe obejmują rybosomy i ośrodek komórkowy, stale obecne w komórce, a także cytoszkielet i organelle ruchu. Nasilenie elementów cytoszkieletu (stałego składnika komórki) może się znacznie zmieniać w trakcie cyklu komórkowego – od całkowitego zaniku jednego składnika (np. kanalików cytoplazmatycznych podczas podziału komórki) do pojawienia się nowych struktur (podział wrzeciono).
Wspólną właściwością organelli błonowych jest to, że wszystkie są zbudowane z warstw lipoproteinowych ( membrany biologiczne), zamykając się na sobie tak, że tworzą się zamknięte wnęki lub przedziały. Wewnętrzna zawartość tych przedziałów jest zawsze inna niż w hialoplazmie Organelle dwubłonowe.
Rozdział 2. Plastydy
Organelle dwubłonowe obejmują plastydy i mitochondria. Plastydy to charakterystyczne organelle komórkowe autotroficznych organizmów eukariotycznych. Ich kolor, kształt i rozmiar są bardzo zróżnicowane. Istnieją chloroplasty, chromoplasty i leukoplasty.
Chloroplasty mają zielony kolor ze względu na obecność głównego pigmentu - chlorofilu. Chloroplasty zawierają również pigmenty pomocnicze - karotenoidy (pomarańczowy). W kształcie chloroplasty są owalnymi soczewkowatymi ciałami o wymiarach (5-10) x (2-4) mikronów. W jednej komórce liścia może znajdować się 15-20 lub więcej chloroplastów, aw niektórych algach tylko 1-2 gigantyczne chloroplasty o różnych kształtach.
Chloroplasty są ograniczone dwiema membranami - zewnętrzną i wewnętrzną. Zewnętrzna błona ogranicza płynne wewnętrzne jednorodne środowisko chloroplastu - zrąb (matrycę). Zrąb zawiera białka, lipidy, DNA (cząsteczka okrężna), RNA, rybosomy i substancje rezerwowe (lipidy, ziarna skrobi i białka), a także enzymy biorące udział w wiązaniu dwutlenku węgla....