Cytoplazma każdej komórki jest otoczona błoną, która ją ogranicza środowisko. Błona cytoplazmatyczna (CPM) jest wyłącznie wielofunkcyjną strukturą:
1. CPM dostrzega wszystkie informacje chemiczne przedostanie się do komórki ze środowiska zewnętrznego.
2. Ona jest główna bariera osmotyczna dzięki czemu wewnątrz komórki utrzymuje się określone ciśnienie osmotyczne.
Ściana komórkowa jest zbudowana głównie z peptydoglikanu przeplatanego kwasem teichojowym, który utrzymuje razem różne warstwy. Stopień usieciowania jest wyższy, a ściana grubsza niż w ścianach komórkowych Gram-ujemnych. Komórki Gram-dodatnie składają się prawie wyłącznie z peptydoglikanu, ale ważna struktura występującym w ścianach komórkowych Gram-dodatnich jest kwas teichojowy. Jest to polimer fosfodiestrowy glicerolu lub rybitolu połączony grupami fosforanowymi. Kwas teichojowy jest kowalencyjnie związany z kwasem muraminowym i przepływa razem z różnymi warstwami sieci peptydoglikanu.
3. CPM wraz ze ścianą komórkową bierze udział w regulacja wzrostu i podziału komórek bakteria.
4. CPM bierze udział w regulacji procesów replikacji i segregacji chromosomów i plazmidów (są one związane z jego receptorami).
5. CPM zawiera znacząca ilość enzymy w tym systemy przenoszenia elektronów (TsPM - miejsce wytwarzania energii w bakteriach).
Kwas teichojowy stabilizuje ścianę komórkową i wzmacnia ją. Wzór chemiczny kwas teichojowy pokazano na rycinie 25. Kwas teichojowy jest długą, cienką cząsteczką, która jest utkana z peptydoglikanu. Ściany komórkowe Gram-ujemne mają więcej złożona struktura niż u organizmów Gram-dodatnich. Na zewnątrz błony cytoplazmatycznej znajduje się peryplazma zawierająca cienką warstwę peptydoglikanu. Peptydoglikan w komórkach Gram-ujemnych zawiera mniej sieciowania niż w komórkach Gram-dodatnich bez łącznika peptydowego.
6. Z CPM związana wici oraz urządzenia do regulowania ich ruchu.
7. CPM uczestniczy w procesach transport(w tym aktywne) składniki odżywcze do komórki, a produkty odpadowe, w tym enzymy i egzotoksyny, z komórki do środowiska. Zawiera białka biorące udział w ułatwionej dyfuzji i aktywnym transporcie.
Kowalencyjnie połączona z peptydoglikanem jest lipoproteina Browna, która ma hydrofobową kotwicę, która pomaga silnie wiązać peptydoglikan z błoną zewnętrzną. Rycina 26 przedstawia położenie ściany komórkowej Gram-ujemnej. Ściana komórkowa bakterii Gram-ujemnych zawiera znacznie mniej peptydoglikanu i jest otoczona błoną zewnętrzną. Istnieje znacznie mniej wiązań poprzecznych między peptydoglikanami.
Zewnętrzna błona bakterii Gram-ujemnych jest kolejną dwuwarstwą lipidową podobną do błony cytoplazmatycznej i zawiera również białka. Stanowi barierę dla białek i zapobiega odkładaniu się uwolnionych enzymów w peryplazmie. To odpychanie pozwala tym bakteriom przetrwać w tym środowisku.
8.
Odtwarza CPM ważna rola w kompartmentalizacji i stabilizacji rybosom.
9.
CPM bierze udział w syntezie składników ściany komórkowej.
10.
CPM bierze udział w tworzeniu mezosomów (mezosomy powstają w wyniku inwazji miejsca CPM do cytoplazmy, otwierają się do przestrzeni peryplazmatycznej).
Jak działa membrana Poziom molekularny jego wiele funkcji - jedną z najpilniejsze sprawy nowoczesna biologia. Udział CPM stanowi około 10% suchej masy bakterii. Zawiera 25-40% fosfolipidy tworzące dwie warstwy, 20-75% białka i do 6% węglowodanów. Cząsteczki fosfolipidy są asymetryczne: głowy przenoszące ładunek elektryczny są hydrofilowe; ogony są neutralne i hydrofobowe. Fosfolipidy są pakowane w membranę w następujący sposób: ich polarne hydrofilowe głowy są skierowane na zewnątrz i tworzą dwie warstwy CPM – wewnętrzną i zewnętrzną, a niepolarne hydrofobowe ogony są ukryte w grubości membrany. Na wzorach dyfrakcji elektronów CPM ma postać trójwarstwowej struktury składającej się z dwóch 2 równoległe ciemne warstwy i oddzielająca je jasna warstwa. Ta warstwa jest bardziej przepuszczalna dla elektronów niż warstwy składające się z polarnych końców fosfolipidów związanych z białkami. Specyfika funkcji CPM w dużej mierze zależy od zestawu białek, które zawierają. Ich lokalizacja w CPM jest specyficzna: niektóre białka przenikają całą podwójną warstwę lipidową, pewna część białek jest związana albo tylko z wewnętrzną, albo tylko z powierzchnia zewnętrzna membrany. Wynika to z faktu, że interakcja między błoną a cytoplazmą, z jednej strony, błoną i otoczenie zewnętrzne, z drugiej strony, determinuje różne, choć wzajemnie powiązane, procesy podtrzymywania życia: ułatwioną dyfuzję, aktywny transport elementarnej błony biologicznej.
On tworzy szeroki zasięg reakcje fizjologiczne w tym wywołanie gorączki, zmiany liczby białych krwinek, wyciek z naczynia krwionośne, martwica guza i zmniejszona ciśnienie krwi prowadząc do zapaści naczyniowej i ostatecznie do wstrząsu. Błona zewnętrzna ma mniej białek i mniej unikalnych typów białek niż błona cytoplazmatyczna. Porins są szczególnie ważne komponenty ze względu na ich rolę w przepuszczalności błony zewnętrznej dla małych cząsteczek. Poryny to białka, które tworzą pory w błonie zewnętrznej wystarczająco duże, aby umożliwić przejście najmniejszym cząsteczkom hydrofilowym.
Przy nadmiernym wzroście (w porównaniu ze wzrostem ściany komórkowej) błona cytoplazmatyczna tworzy wklęsłości - wklęsłości w postaci skomplikowanych skręconych struktur błonowych, tzw. mezosomy. Mniej złożone skręcone struktury nazywane są błonami wewnątrzcytoplazmatycznymi. Rola mezosomów a błony wewnątrzcytoplazmatyczne nie zostały w pełni wyjaśnione. Sugeruje się nawet, że są one artefaktem powstałym po przygotowaniu (utrwaleniu) preparatu do mikroskopii elektronowej. Niemniej jednak uważa się, że pochodne błony cytoplazmatycznej biorą udział w podziale komórkowym, dostarczając energii do syntezy ściany komórkowej, biorą udział w wydzielaniu substancji, tworzeniu przetrwalników, czyli w procesach o dużej energochłonności.
na ryc. 28 przedstawia strukturę poriny w skali molekularnej. Wszystkie znane poryny mają podobną strukturę, z białkiem zawierającym centralny kanał, który umożliwia cząsteczkom przechodzenie. Pozwala to na transport tych cząsteczek do przestrzeni peryplazmatycznej w celu ewentualnego transportu przez cytoplazmę. błona plazmatyczna. Niektóre pory w błonie zewnętrznej są ogólne, rozróżniają proste rozmiary i ładunki, ale mają niewielką specyficzność. Inne poriny są bardziej szczegółowe.
Bardzo duże lub hydrofobowe cząsteczki nie mogą przeniknąć przez błonę zewnętrzną, więc błona zewnętrzna służy jako bariera przepuszczalności przynajmniej dla niektórych cząsteczek. Struktura molekularna poriny. Widok wewnątrz jest na zewnątrz komórki, patrząc na powierzchnię membrany. Spojrzenie to perspektywa z zewnątrz. Porin ma trzy podjednostki białkowe, a rzeczywisty por to centralny trójkątny obszar w górnym panelu utworzony przez trzy podjednostki.
Cytoplazma zajmuje większość komórki bakteryjnej i składa się z rozpuszczalnych białek, kwasów rybonukleinowych, inkluzji i licznych małych granulek - rybosomów odpowiedzialnych za syntezę (translację) białek.
Rybosomy bakterie mają rozmiar około 20 nm i współczynnik sedymentacja 70S, w przeciwieństwie do rybosomów 8OS charakterystycznych dla komórki eukariotyczne. Dlatego niektóre antybiotyki wiążą się z rybosomami bakteryjnymi i hamują syntezę białek bakteryjnych bez wpływu na syntezę białek w komórkach eukariotycznych. Rybosomy bakteryjne mogą dysocjować na dwie podjednostki, 50S i 30S. Rybosomalny RNA (rRNA.) - konserwatywne elementy bakterii ("zegar molekularny" ewolucji). rRNA 16S. jest częścią małej podjednostki rybosomów, a 23S rRNA jest częścią dużej podjednostki rybosomów. Badanie 16S rRNA. stanowi podstawę genosystematyki, pozwalając ocenić stopień pokrewieństwa organizmów.
Istnieją również inne rodzaje białek zewnętrzna męmbrana w których uczestniczą różne funkcje. Niektóre patogeny zawierają białka błony zewnętrznej, które pomagają im zneutralizować mechanizmy obronne żywiciela. Różnice między ścianami komórkowymi bakterii Gram-ujemnych i Gram-ujemnych mają ogromny wpływ na powodzenie drobnoustrojów w ich środowisku. Gruba ściana komórkowa komórek Gram-ujemnych pozwala im lepiej radzić sobie w suchych warunkach, ponieważ ogranicza utratę wody. Tabela 3 podsumowuje różnicę między ścianami komórkowymi Gram-ujemnymi i Gram-ujemnymi.
Cytoplazma zawiera różne włączenie w postaci granulek glikogen, polisacharydy, kwas beta-hydroksymasłowy i polifosforany (wolutyna). Wszystkie substancje rezerwowe występują w komórkach w postaci obojętnej chemicznie. Ten stan zapobiega naruszeniu osmostazy zawartości komórkowej. Niektóre wtrącenia po prostu leżą w cytoplazmie, inne są otoczone cienką błoną. Błona ma zwykle charakter białkowy, ale czasami może zawierać lipidy. Kumulują się wraz z nadmiarem składników pokarmowych w środowisku i służą jako substancje rezerwowe dla potrzeb żywieniowych i energetycznych.
Tabela 3 Właściwości ściany komórkowej
Podsumowanie różnic między ścianami komórkowymi Gram-ujemnymi i Gram-ujemnymi. W przypadku większości komórek bakteryjnych ściana komórkowa ma kluczowy dla przeżycia komórek, ale istnieją bakterie, które nie mają ścian komórkowych. Typowymi przykładami są gatunki Mycoplasma, a niektóre mogą być patogenami wewnątrzkomórkowymi, które rosną wewnątrz ich żywicieli. Ściany komórkowe nie są tutaj potrzebne, ponieważ komórki żyją tylko w kontrolowanym środowisku osmotycznym innych komórek.
Woliutin ma powinowactwo do barwników zasadowych i jest łatwo wykrywalny za pomocą specjalnych metod barwienia (na przykład według Neissera) w postaci metachromatycznych granulek. Błękit toluidynowy lub błękit metylenowy barwi wolutinę na czerwono-fioletowo, a cytoplazmę bakteryjną na niebiesko. Charakterystyczny układ ziarnistości wolutyny ujawnia się w prątku błonicy w postaci intensywnie wybarwionych biegunów komórki. Związane jest z metachromatycznym barwieniem wolutyny wysoka zawartość spolimeryzowany nieorganiczny polifosforan. Pod mikroskopem elektronowym wyglądają jak gęste elektronowo granulki o wielkości 0,1-1,0 µm.
Prawdopodobnie mieli zdolność tworzenia ściany komórkowej w pewnym momencie w przeszłości, ale gdy ich styl życia stał się trybem przebywania w innych komórkach, stracili zdolność tworzenia ścian. Zgodnie z tym bardzo ograniczonym sposobem życia w innych komórkach, te drobnoustroje są również bardzo małe. Nie potrzebują genów dla wszystkich rodzajów enzymów biosyntetycznych, ponieważ mogą ukraść końcowe składniki tych szlaków od gospodarza. Podobnie nie potrzebują genów, które kodują wiele różne sposoby dla różnych źródeł węgla, azotu i energii, ponieważ ich środowisko wewnątrzkomórkowe jest całkowicie przewidywalne.
nukleoid jest odpowiednikiem jądra w bakteriach. W 1956 r. odkryto „strefę jądrową” lub nukleoid wewnątrz komórek bakteryjnych, w których znajduje się chromosom bakteryjny. Bakteryjne DNA występuje w postaci kolistej i liniowej. W przypadku komórek E. coli udowodniono, że DNA istnieje w postaci kolistej cząsteczki, podczas gdy w przypadku Borrelia burgdorferi w 1989 r. wykazano, że komórki zawierają DNA w postaci liniowej. Nukleoid, w przeciwieństwie do jądra eukariotycznego, nie ma otoczka jądrowa, jąderko i główne białka (histony). Zwykle w komórka bakteryjna zawiera jeden chromosom, ale jeśli podział jest zaburzony, może zawierać 4 lub więcej kopii jednego chromosomu. U niektórych gatunków w komórkach znaleziono dwa lub trzy nieidentyczne chromosomy. Gatunki Vibrio, Leptospira interrogans mają dwa okrągłe chromosomy. Dlatego pomysł, że bakterie zawierają tylko jeden chromosom, jest uważany za przestarzały. Nukleoid jest wykrywany w mikroskopie świetlnym po barwieniu metodami specyficznymi dla DNA: według Feulgena lub Romanovsky'ego-Giemsy. Na wzorach dyfrakcji elektronów ultracienkich skrawków bakterii nukleoid ma postać jasnych stref z włóknistymi, nitkowatymi strukturami DNA związanymi z pewnymi obszarami z błoną cytoplazmatyczną lub mezosomem zaangażowanym w replikację chromosomu.
Obecność steroli w membranie przyczynia się do ich długowieczności, pomagając zwiększyć siły utrzymujące membranę razem. Bo nie byłoby ludzi, roślin, zwierząt, grzybów czy pierwotniaków. Wszystkie te organizmy są Żyjąca istota oprócz bakterii składają się z komórek eukariotycznych.
A geneza tego typu komórek nie byłaby możliwa bez symbiozy. Powstawanie komórek eukariotycznych wyjaśnia nauka tzw. teorią endosymbiontu. Zgodnie z tą teorią złożone komórki z Jądro komórkowe, oddzielone od osocza i organelli komórkowych, powstały z symbiozy prokariotów o prostszej strukturze. Tylko bakterie, które żyły jako endosymbionty w większej komórce, utworzyły organelle dzisiejszych komórek eukariotycznych.
Oprócz nukleoidu komórka bakteryjna ma pozachromosomalne czynniki dziedziczności. - plazmidy, które są kowalencyjnie zamkniętymi pierścieniami DNA, które nie kodują funkcji niezbędnych do życia komórki bakteryjnej, ale dają bakteriom przewagę w niekorzystnych warunkach bytowania.
Tworzenie mitochondriów
Ta akumulacja materii organicznej jest pożądanym źródłem składników odżywczych dla komórek heterotroficznych. Komórki zyskują rozmiar i oprócz tego, że są w stanie wchłonąć materia organiczna fagocytoza, zdolna do wchłaniania komórki organiczne a także mniejsze komórki są fagocytowane. Z reguły komórki te są w postaci pokarmu, są trawione.
Czasami jednak zdarza się, że bakterie wzięte żywcem przeżywają w środku większa komórka i wchodzić w interakcje ze swoim panem. Rozwija się symbioza, w której komórka gospodarza może efektywniej wykorzystywać produkty żywieniowe z wykorzystaniem enzymów z łańcucha oddechowego bakterii tlenowych. Z drugiej strony endosymbiont jest bardziej chroniony wewnątrz większej komórki przed zmianami środowiskowymi. Inny możliwe źródło mitochondria zostaną zaatakowane przez pasożyty bakteryjne w komórce gospodarza i wynikająca z tego symbioza.
Kapsułka, mikrokapsułka, śluz. Kapsuła- struktura śluzowa o grubości większej niż 0,2 mikrona, mocno związana ze ścianą komórkową bakterii i mająca wyraźnie określone granice zewnętrzne. Kapsułka jest rozpoznawalna w rozmazach-odciskach z materiału patologicznego. Powierzchnia kolonii komórkowych z kapsułkami wygląda na gładką, wilgotną, błyszczącą.
Komórki narządów z wieloma błonami
Nie wiadomo z absolutną pewnością, czy eukarionty faktycznie powstały w ten sposób, ale istnieje wiele faktów potwierdzających teorię endosymbiontu. Ponadto organelle nie mogą powstać z informacji genetycznej jądra, ale są mnożone przez dzielenie. Proces podziału jest podobny do procesu podziału bakterii. Rybosomy organelli komórkowych są strukturalnie podobne do rybosomów bakteryjnych, a nie eukariotycznych.
Endosymbiozy występują do dziś. Tak więc niektóre okrzemki morskie mogą najwyraźniej żyć w symbiozie z niedoborem sinic. Ponownie, symbiont jest integrowany z osoczem komórki gospodarza. Dlatego nawet dzisiaj ten rodzaj symbiozy powtarza się jako proces, który stał u początku wszelkiego wyższego życia.
Powstaje wiele bakterii mikrokapsułka- tworzenie śluzu o grubości mniejszej niż 0,2 mikrona, wykrywane tylko za pomocą mikroskopii elektronowej. Śluz należy odróżnić od kapsułki - śluzowatych egzopolisacharydów, które nie mają wyraźnych granic zewnętrznych. Slime jest rozpuszczalny w wodzie.
Egzopolisacharydy śluzowe są charakterystyczne dla śluzowatych szczepów Pseudomonas aeruginosa, często znajdowanych w plwocinie pacjentów z mukowiscydozą. Egzopolisacharydy bakteryjne biorą udział w adhezji (przyklejaniu się do podłoża); nazywane są również glikokaliksem. Oprócz syntezy egzopolisacharydów przez bakterie istnieje inny mechanizm ich powstawania: poprzez działanie pozakomórkowych enzymów bakteryjnych na disacharydy. W rezultacie powstają dekstrany i lewany.
Charakteryzuje się brakiem jądra. Ich główny podział występuje w dwóch i Archaea. W Życie codzienne Prokarionty są nam znane jako bakterie. Często słowo „bakterie” lub bakterie jest bezpośrednio utożsamiane z patogenami. Najmniej prokarionty są patologiczne - to znaczy patogenne.
Ściana komórkowa otacza błonę cytoplazmatyczną i wewnętrzna część komórki. Nie ma dekoracji wnętrz. Wszystkie prokarioty mają tę samą podstawową strukturę. Większość prokariotów ma ścianę komórkową zlokalizowaną poza błoną plazmatyczną i składającą się z pochodnej węglowodanowej. W zależności od liczby i ułożenia warstw murein rozróżnia się bakterie Gram-dodatnie i Gram-ujemne.
- Komórkę otacza błona plazmatyczna.
- Po tym typie archeony nie mają ściany komórkowej.
Torebka i śluz chronią bakterie przed uszkodzeniem i wysychaniem, ponieważ będąc hydrofilowymi, dobrze wiążą wodę i zapobiegają działaniu czynników ochronnych makroorganizmów i bakteriofagów.
wici bakterie determinują ruchliwość komórki bakteryjnej. Wici są cienkie nici, pochodzące z cytoplazmy membrany mają dłużej niż sama komórka. Wici mają grubość 12–20 nm i długość 3–15 µm. Składają się z 3 części.:
Zasadniczo komórka otoczona błoną nie jest dalej dzielona. Ogólnie rzecz biorąc, istnieje mechanizm, który pozwala ci się poruszać. Ma na błonie cytoplazmatycznej leżącej na innej warstwie komórek. Wykonany jest z muraine. Na nim leży powłoka zewnętrzna. Oblężony jednokomórkowiec wprowadził zielone algi dla subtensji. Przyczyny symbiozy drastyczne zmiany w obu komórkach. organizm jednokomórkowy przełącza się między fazą drapieżnika a fazą żywiciela z endosymbiontem, zmieniając swój bilans energetyczny z heterotroficznego na autotroficzny, tj. od dostaw do energii słonecznej.
1. - nić spiralna,
2. - hak
3. - ciało podstawowe zawierające pręcik ze specjalnymi krążkami (1 para krążków u bakterii Gram-dodatnich i 2 pary u bakterii Gram-ujemnych) oraz białkami motorycznymi (ryc.
dyski wici są przyczepione do błony cytoplazmatycznej i Ściana komórkowa. Tworzy to efekt silnika elektrycznego z prętem - wirnikiem, który obraca wić. Jako źródło energii wykorzystuje się różnicę potencjałów protonów na błonie cytoplazmatycznej. Mechanizm rotacji zapewnia syntetaza protonu ATP. Prędkość obrotu wici może osiągnąć 100 obrotów na minutę. Bakterie mają wici prawoskrętne. Jeśli bakteria ma kilka wici, zaczynają one obracać się synchronicznie, splatając się w jedną wiązkę, tworząc coś w rodzaju śmigła.
W procesie adaptacji dwóch komórek roślina wyłania się z pierwotniaków z cząsteczkami; Naukowcy nazwali je Hatena, zbyt tajemniczym dla nich Niemiecki. Ta obserwacja jest kolejnym potwierdzeniem teorii endosymbiozy, zgodnie z którą prekursory wszystkich komórek roślinnych i zwierzęcych powstały w wyniku endosymbiozy.
Jakie organelle otrzymali poprzednicy komórki roślinne według Endosombiontentoria Endosymbiosis? Wymień cztery cechy tych organelli, które potwierdzają teorię endosymbiozy! W każdym przypadku uzasadniamy, dlaczego uważasz tę cechę za dowód teorii endosymbiontu. W jakim stopniu odkrycie nowych glonów Khaten można uznać za podstawę teorii endosymbiontów? Podczas endosymbiozy z bakteriami tlenowymi pierwotne eukarionty nabyły zdolność do degradacji oksydacyjnej poprzez symbiozę z sinicami, zdolność do fotosyntezy. Nie mogą się ponownie pojawić. Sugeruje to, że pierwotnie nie powstały jako struktury komórek gospodarza. Mitochondria i chloroplasty mają własne mechanizmy syntezy białek, a ich rybosomy są zbudowane jak u prokariotów. Oba są otoczone dwiema membranami, z których wewnętrzna jest podobna do błona prokariotyczna, zewnętrzna z Błona komórkowa. Ważną cechą teorii endosymbiozy jest fakt, że nie ma form pośrednich między komórkami prokariotycznymi i eukariotycznymi, ale istnieją formy pośrednie między organizmami endosymbiotycznymi a organellami. Tą formą przejściową są obserwowane glony jednokomórkowe: po zmianie diety jednokomórkowe zależą od symbiozy z zielonymi algami, podczas gdy tylko zielone algi jest opłacalny.
- Jakie specjalne umiejętności nabyłeś?
- Mitochondria i chloroplasty są potomkami mikroorganizmów.
- Mitochondria pochodzą od bakterii, chloroplasty od sinic.
- Oba powstają tylko poprzez oddzielenie swoich rówieśników.
Wici składają się z białka flagelina(z. rozłóg- wici), która jest antygenem - tzw antygen H. Podjednostki flageliny są zwinięte.
Liczba wici w bakteriach różnego rodzaju różni się od jednego (monotryk) w vibrio cholerae do dziesięciu i setek wici rozciągających się wzdłuż obwodu bakterii (perytrycz), Na coli, protea itp. lofotryczny mieć wiązkę wici na jednym końcu komórki. amfitryki mieć jedną wici lub wiązkę wici na przeciwległych końcach komórki.
Wici są wykrywane za pomocą mikroskopii elektronowej preparatów spryskanych metalami ciężkimi lub w mikroskopie świetlnym po obróbce. specjalne metody oparte na trawieniu i adsorpcji różne substancje, prowadząc do wzrostu grubości wici (np. po srebrzenie).
kosmki lub pił(fimbriae) - nitkowate formacje, więcej cienki i krótki, niż wici. Pili wystają z powierzchni komórki i składają się z wiewiórka pilina, który ma strukturę spiralną. Architektura pilusów waha się od cienkich nitkowatych do grubych, mocnych formacji w kształcie prętów z osiowymi otworami. Mają aktywność antygenową. Wyróżnić picie, odpowiedzialny za przyczepność czyli za przyczepienie się bakterii do chorej komórki, a także za picie, odpowiedzialne za odżywianie, wymiana wodno-solna, I seksualny (P-piłem) lub pił 4. typ lub sprzężony, pił.
Zazwyczaj pił obficie- kilkaset za komórkę. Jednakże na komórkę przypada zwykle 1-3 pilusów płciowych: są one tworzone przez tak zwane „męskie” komórki dawcy zawierające przenośne plazmidy (F-, R-, Co/-plazmidy). Charakterystyczną cechą pilusów płciowych jest ich interakcja ze specjalnymi „męskimi” kulistymi bakteriofagami, które są intensywnie adsorbowane na pilusach płciowych (ryc.).
Głównym celem pilusów jest wspieranie określonych struktur mocujących (adhezyn) na ich końcach. Adhezyny są mediatorami w kontaktach bakteryjnych, w kontakcie z przedmiotami nieożywionymi, tkankami i komórkami organizmów wrażliwych. Kolonizacja tkanek gospodarza przez patogeny bakteryjne ogólnie zależy od stereochemicznego podobieństwa między architekturą adhezyny i odpowiedniego receptora komórki gospodarza (ryc.). Tym samym badanie funkcjonowania pilusów pozwoli nie tylko na głębsze zrozumienie mechanizmu kolonizacji i sygnalizacji, ale także na opracowanie nowych generacji leków przeciwdrobnoustrojowych.
spór - specyficzna forma bakterii spoczynkowych o gram-dodatnim typie budowy ściany komórkowej (ryc.).
Zarodniki powstają, gdy Nie korzystne warunki istnienie bakterii (suszenie, niedobór składników odżywczych itp.). Wewnątrz komórki bakteryjnej powstaje jeden zarodnik (endospora). Tworzenie zarodników przyczynia się do zachowania gatunku i nie jest Xia sposób rozmnażania, jak grzyby.
bakterie tworzące przetrwalniki z rodzaju bakcyl, Który wielkość zarodnika nie przekracza średnicy komórki, tzw pałeczki. bakterie tworzące przetrwalniki, które rozmiar zarodnika przekracza średnicę komórki, dlatego przybierają postać wrzeciona, tzw Clostridia, takich jak bakterie z rodzaju Clostridium(łac. Clostridium- wrzeciono). Zarodniki są kwasoodporne dlatego barwi się je metodą Aujeszky'ego lub metodą Ziehla-Nelsena na czerwono, a komórkę wegetatywną na niebiesko.
Zarodnikowanie, kształt i umiejscowienie zarodników w komórce (wegetatywne) to cecha gatunkowa bakterii, która umożliwia ich odróżnienie.
Formularz sporu może być owalny, kulisty; układ w celi :
1. terminal, tj. na końcu sztyftu (w czynniku wywołującym tężec),
2. podterminalny- bliżej końca sztyftu (dla czynników wywołujących botulizm, zgorzel gazowa)
3. centralnie e (w pałeczkach wąglika).
Proces Formacja zarodnikow (zarodnikowanie) przechodzi przez szereg etapów, podczas których część cytoplazmy i chromosom komórki wegetatywnej bakterii są oddzielane, otoczone wrośniętą błoną cytoplazmatyczną, prosperować.
Prospor otoczony dwiema błonami cytoplazmatycznymi, pomiędzy którymi tworzy się gruba zmieniona warstwa peptydoglikanu kory (kory). Od wewnątrz wchodzi w kontakt ze ścianą komórkową zarodnika, a od zewnątrz - z wewnętrzną otoczką zarodnika. Zewnętrzna powłoka zarodnika jest utworzona przez komórkę wegetatywną.
Zarodniki niektórych bakterii mają dodatkową osłonę - egzosporium.
W ten sposób powstaje wielowarstwowa słabo przepuszczalna otoczka. Zarodnikowaniu towarzyszy intensywna konsumpcja przez prosporę, a następnie wyłaniająca się otoczka zarodników kwasu dipikolinowego i jonów wapnia. Spore nabywa wytrzymałość cieplna, co jest związane z obecnością w nim dipikolinianu wapnia.
Zarodnik może przetrwać przez długi czas ze względu na obecność wielowarstwowej otoczki, dipikolinianu wapnia, niska zawartość woda i powolne procesy metaboliczne. W glebie na przykład patogeny wąglik a tężec może utrzymywać się przez dziesięciolecia.
W sprzyjających warunkach zarodniki kiełkują w trzech kolejnych stadiach (ryc.):
1. aktywacja
2. inicjacja
3 . wzrost
W tym przypadku jedna bakteria powstaje z jednego zarodnika.
Aktywacja - jest gotowy do kiełkowania. W temperaturze 60-80°C zarodnik jest aktywowany do kiełkowania.
Inicjacja kiełkowanie trwa kilka minut. Scharakteryzowano fazę wzrostu Szybki wzrost, któremu towarzyszy zniszczenie skorupy i uwolnienie sadzonki.
Pytania do samokontroli
| | | | | | | | | 10 | | | | | |
Budowę typowej komórki bakteryjnej przedstawiono na ryc. 2.3. na ryc. 2.4 przedstawia mikrografię elektronową wycinka bakterii w kształcie pałeczki. Możesz zobaczyć, jak prosta jest komórka bakteryjna, zwłaszcza w porównaniu z komórkami eukariotycznymi (ryc. 7.5 i 7.6).
Ryż. 2.3. Uogólniony schemat budowy komórki bakteryjnej w kształcie pręcika. Po prawej stronie znajdują się struktury występujące w każdej komórce, po lewej - nie występujące we wszystkich komórkach. Rozłóg czasami jeden, jak Rhizobium, lub kilka, jak Azotobacter; jest zwykle dłuższy niż klatka. Kapsuła może być śluzowa, jak u Azotobacter; jeśli kapsułka jest luźna, nazywa się to warstwa śluzowa. w kształcie rurki lub torby membrany fotosyntetyczne zawierające pigmenty są inwazjami błony plazmatycznej; w bakteriach fotosyntetyzujących, takich jak Chromatium, takie membrany są rozproszone w całej cytoplazmie. Numer pilusy, Lub pilusy, może sięgać od jednego do kilkuset, jak na przykład w Escherichia coli, Salmonella. Mezosom jest wielokrotnym wpukleniem błony plazmatycznej, jak na przykład u Bacillus subtilis. Ściana komórkowa twardy i zawiera mureinę. Rybosomy, zlokalizowany w całej cytoplazmie, jest mniejszy niż u eukariontów. Z rezerwować składniki odżywcze w komórkach bakteryjnych można wykryć lipidy, glikogen, polifosforany (granulki wolutyny). Cytoplazma nie zawiera organelli; zawiera enzymy itp.
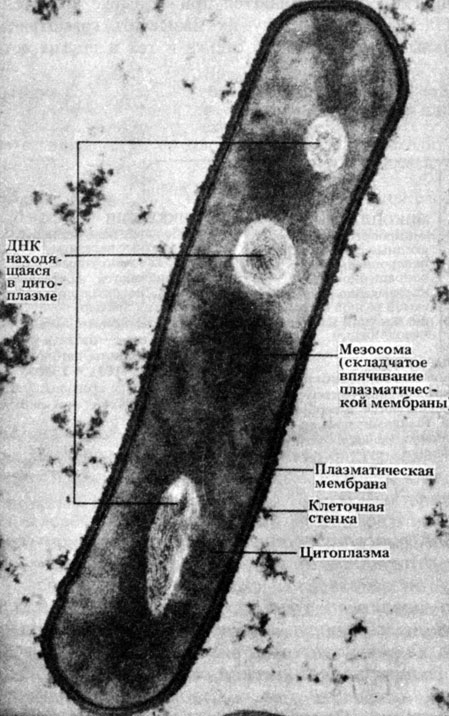
Ryż. 2.4. Mikrografia elektronowa wycinka typowej bakterii Bacillus subtilis w kształcie pręcika. Jasne obszary zawierają DNA. × 50000
Kapsułki i warstwy śluzowe
Kapsułki i warstwy śluzu to śluzowate lub lepkie wydzieliny niektórych bakterii; takie wyładowania są wyraźnie widoczne po kontrastowaniu negatywnym (gdy barwienie nie jest preparatem, ale tłem). Kapsuła jest stosunkowo gruby i zwarty, i warstwa śluzu dużo luźniejszy. W niektórych przypadkach śluz służy do tworzenia kolonii z pojedynczych komórek. Zarówno otoczka, jak i błona śluzowa stanowią dodatkową ochronę komórek. Na przykład otoczkowe szczepy pneumokoków namnażają się swobodnie w organizmie człowieka i powodują zapalenie płuc, podczas gdy szczepy nie otoczkowe są łatwo atakowane i niszczone przez fagocyty, a zatem są całkowicie nieszkodliwe.
Ściana komórkowa
Ściana komórkowa nadaje komórce określony kształt i sztywność. Jest to wyraźnie widoczne na rozcięciu (ryc. 2.4). Podobnie jak u roślin, ściana komórkowa bakterii zapobiega pęcznieniu osmotycznemu i pękaniu komórek, gdy, jak to często bywa, wchodzą one w środowisko hipotoniczne (Dodatek, Sekcja A.1.5). Woda, inne małe cząsteczki i różne jony łatwo przechodzą przez małe pory w ścianie komórkowej, ale duże cząsteczki białek i kwasów nukleinowych nie przechodzą przez nie. Ponadto ściana komórkowa ma właściwości antygenowe, które nadają jej zawarte w niej białka i polisacharydy.
Ze względu na budowę ściany komórkowej bakterie można podzielić na dwie grupy. Niektóre są barwione metodą Grama, stąd ich nazwa Gram-dodatnie, podczas gdy inne stają się bezbarwne po zmyciu barwnika (rozdz. 2.7) i dlatego nazywane są Gram-ujemne. W ścianie komórkowej obu z nich znajduje się specjalna sztywna siatka składająca się z mureina. Cząsteczka mureiny to regularna sieć równoległych łańcuchów polisacharydowych usieciowanych ze sobą krótkimi łańcuchami peptydowymi. W ten sposób każda komórka jest otoczona siatkowatym workiem złożonym tylko z jednej cząsteczki. (Polisacharydowa część mureiny jest opisana w Tabeli 5.7).
U bakterii Gram-dodatnich, takich jak Lactobacillus, inne substancje, głównie polisacharydy i białka, są osadzone w sieci mureinowej. Tworzy to stosunkowo gruby i sztywny pakiet wokół komórki. Bakterie Gram-ujemne, takie jak Escherichia coli lub Azotobacter, mają znacznie cieńszą ścianę komórkową, ale jest ona bardziej złożona. Warstwa mureinowa tych bakterii pokryta jest na zewnątrz miękką i gładką warstwą lipidów. To ich chroni przed lizozym. Lizozym znajduje się w ślinie, łzach i innych płynach ustrojowych, a także w białku kurze jajo. Katalizuje hydrolizę niektórych wiązań między resztami węglowodanowymi, a tym samym rozszczepia szkielet polisacharydowy mureiny. Ściana komórkowa pęka, a jeśli komórka znajduje się w roztworze hipotonicznym, następuje jej liza (komórka osmotycznie pęcznieje i pęka). Warstwa lipidowa nadaje komórce oporność na penicylinę. Antybiotyk ten zapobiega tworzeniu się wiązań poprzecznych w ścianie komórkowej bakterii Gram-dodatnich, co powoduje, że rosnące komórki są bardziej podatne na szok osmotyczny.
wici
Wiele bakterii jest ruchliwych, a ruchliwość ta wynika z obecności w nich jednej lub więcej wici. Wici bakteryjne są znacznie prostsze niż wici eukariotyczne (rozdział 17.6.2, tabela 2.1), a swoją budową przypominają jedną z mikrotubul wici eukariotycznej. Wici składają się z identycznych kulistych podjednostek białkowych flagelina(podobne do aktyny mięśniowej), które ułożone są spiralnie i tworzą wydrążony cylinder o średnicy około 10-20 nm. Pomimo falistego kształtu wici są dość sztywne.
Wici są napędzane przez unikalny mechanizm. Podstawa wici najwyraźniej obraca się w taki sposób, że wić wydaje się być wkręcona w pożywkę bez wykonywania przypadkowych uderzeń, a tym samym przesuwa komórkę do przodu. Jest to najwyraźniej jedyna znana w przyrodzie konstrukcja, w której zastosowano zasadę działania koła. Inny ciekawa funkcja wici to zdolność poszczególnych podjednostek flageliny do spontanicznego łączenia się w roztworze w spiralne włókna. Spontaniczny samodzielny montaż- Bardzo ważna właściwość wiele złożonych struktur biologicznych. W tym przypadku samoorganizacja jest całkowicie spowodowana sekwencją aminokwasową (strukturą pierwotną) flageliny.
Bakterie ruchliwe mogą poruszać się w odpowiedzi na określone bodźce, tj. są zdolne do taksówek. Na przykład, bakterie tlenowe mają dodatnią aerotaksję (tj. płyną tam, gdzie środowisko jest bogatsze w tlen), a ruchome bakterie fotosyntetyzujące mają dodatnią fototaksję (tj. płyną w kierunku światła).
Wici najłatwiej zobaczyć pod mikroskopem elektronowym (ryc. 2.5), stosując technikę osadzania metalu (sekcja A.2.5).
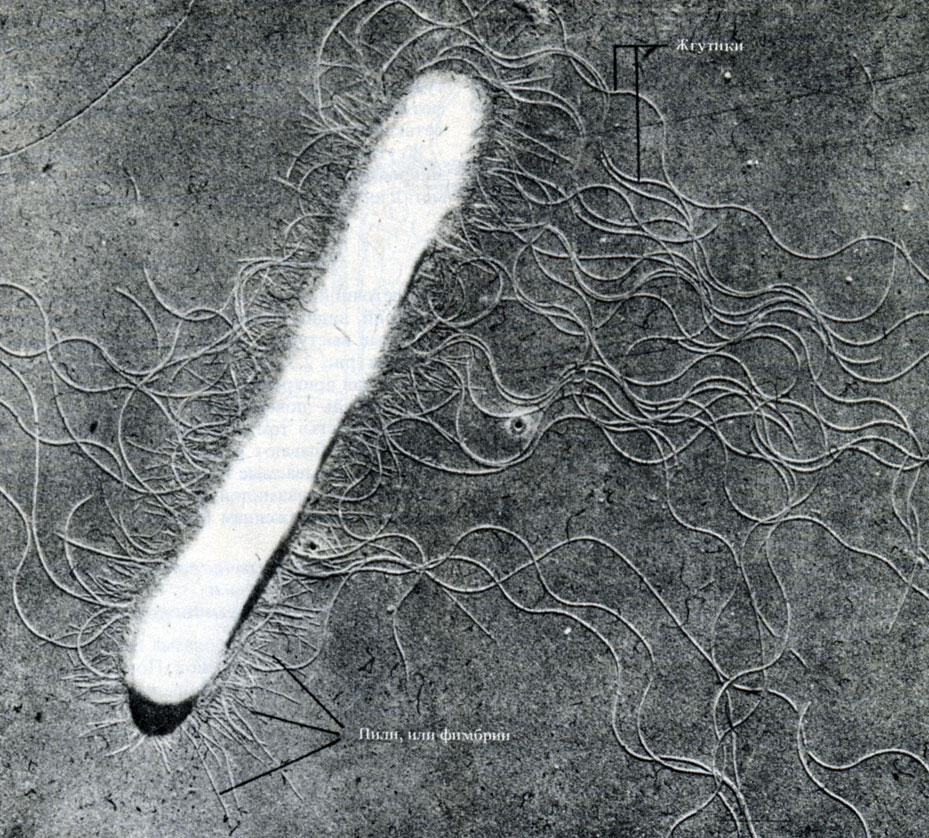
Ryż. 2.5. Mikrofotografia bakterii w kształcie pręcika uzyskana przy użyciu półprzezroczystego światła mikroskop elektronowy. Ściana komórkowa, fimbrie i długie faliste wici są wyraźnie widoczne, × 28000
Pili lub fimbrie
Na ścianie komórkowej niektórych bakterii Gram-ujemnych widoczne są cienkie wyrostki (białkowe wypustki w kształcie pręcików), które nazywane są pił Lub fimbrie(Rys. 2.5). Są krótsze i cieńsze niż wici i służą do przyczepiania komórek do siebie lub do jakiejś powierzchni, nadając specyficzną „lepkość” tym szczepom, które je posiadają. Pije się inny rodzaj. Najciekawsze są tak zwane pilusy F, które są kodowane przez specjalny plazmid (sekcja 2.2.4) i są związane z rozmnażaniem płciowym bakterii.
Błona plazmatyczna, mezosomy i błony fotosyntetyczne
Jak wszystkie komórki, protoplazma bakterii jest otoczona półprzepuszczalną błoną. Błony plazmatyczne bakterii nie różnią się strukturą i funkcją od błon komórek eukariotycznych (sekcja 7.2.1). U niektórych bakterii błona plazmatyczna wybrzusza się w komórce i tworzy mezosomy i/lub błony fotosyntetyczne.
mezosomy- pofałdowane struktury membranowe (ryc. 2.3 i 2.4), na powierzchni których znajdują się enzymy biorące udział w procesie oddychania. Dlatego mezosomy można nazwać prymitywnymi organellami. Podczas podziału komórki mezosomy wiążą się z DNA, co wydaje się ułatwiać rozdzielenie dwóch potomnych cząsteczek DNA po replikacji i promować tworzenie przegrody między komórkami potomnymi.
W bakteriach fotosyntetyzujących workowate, rurkowe lub blaszkowate wgłębienia błony komórkowej zawierają barwniki fotosyntetyczne (w tym bakteriochlorofil). Podobne formacje membranowe są również zaangażowane w wiązanie azotu.
materiał genetyczny
Bakteryjne DNA jest reprezentowane przez pojedyncze okrągłe cząsteczki o długości około 1 mm. Każda taka cząsteczka składa się z około 5×106 par zasad. Całkowita zawartość DNA (genomu) w komórce bakteryjnej jest znacznie mniejsza niż w komórce eukariotycznej, a co za tym idzie, ilość zakodowanej w niej informacji jest również mniejsza. Taki DNA zawiera średnio kilka tysięcy genów, czyli około 500 razy mniej niż komórka ludzka (patrz też Tabela 2.1 i Rycina 2.3).
Rybosomy
Zobacz tabelę. 2.1 (biosynteza białek) i ryc. 2.3.
spór
Niektóre bakterie (głównie należące do rodzaju Clostridium lub Bacillus) tworzą endospory, czyli zarodniki znajdujące się wewnątrz komórki. Endospory to grubościenne, długowieczne formacje, które są wyjątkowo odporne na ciepło i promieniowanie krótkofalowe. Są one różnie rozmieszczone wewnątrz komórki, co jest bardzo ważną cechą dla identyfikacji i systematyki takich bakterii (ryc. 2.6). Jeśli z całej komórki powstaje spoczynkowa, stabilna struktura, nazywa się to cystą. Cysty tworzą niektóre gatunki Azotobacter.
Ryż. 2.6. Różne formy bakterie na przykładzie kilku najpowszechniejszych rodzajów drobnoustrojów pożytecznych i chorobotwórczych.
A. Cocci (kulisty)
Przykładem jest Staphylococcus aureus żyjący w nosogardzieli; różne szczepy gronkowców powodują furunculosis, zapalenie płuc, zatrucia pokarmowe i inne choroby.
Przykładem jest wiele gatunków Streptococcus; jakaś przyczyna choroba zakaźna górny drogi oddechowe; na przykład S. pyogenes powoduje ból gardła i szkarlatynę; S. thermophilus nadaje jogurtowi jego pikantny smak; S. lactis - patrz rozdz. 2.3.4
Ten rodzaj obejmuje jedyny gatunek Diplococcus pneumoniae (pneumococcus), czynnik wywołujący zapalenie płuc *.
* (D. pneumoniae powoduje płatowe zapalenie płuc i liczne ciężkie ostre zakażenia ropne: zapalenie opon mózgowych, posocznicę, ropniak i zapalenie otrzewnej. - Około. tłumacz.)
B. Bacilli (w kształcie pręta)
![]()
Przykładami są Escherichia coli (pospolity symbiont jelitowy); Lactobacillus patrz rozdz. 2.3.4; Salmonella typhi jest czynnikiem sprawczym duru brzusznego.
Przykładami są Azotobacter, bakteria wiążąca azot; Bacillus anthracis jest czynnikiem sprawczym wąglika.
Bacilli z endosporami (zarodniki są w różnych pozycjach, mają różne rozmiary i kształt)
Znajduje się w centrum i nie powoduje pęcznienia komórki, na przykład u Bacillus anthracis, czynnika sprawczego wąglika.
Znajduje się na końcu komórki macierzystej, daje ją charakterystyczny kształt pałeczka do gry na bębnie, takich jak Clostridium tetani, czynnik wywołujący tężec.
Zarodnik znajduje się w pozycji subterminalnej, powodując pęcznienie komórki, np. w Clostridium botulinum (zarodniki mogą zajmować również pozycję centralną) zatrucie pokarmowe- zatrucie jadem kiełbasianym.
B. Spirilli (spirala)
Przykładem jest Spirillum; Kształt komórek krętków jest bardzo podobny, ale istnieją różnice w sposobie poruszania się, na przykład Treponema blada- czynnik sprawczy kiły.
D. Vibrios (krótkie pałeczki, zawsze zakrzywione w formie przecinka)
Przykład - Vibrio cholerae - czynnik sprawczy cholery; ma jedną wici.