Istnieją trzy hipotezy dotyczące pochodzenia komórki eukariotyczne:
- hipoteza symbiotyczna, Lub symbiogeneza,
- insynuacyjny,
- chimeryczny.
Do tej pory w świecie naukowym uznawana jest główna hipoteza pochodzenia eukariontów symbiogeneza.
Zgodnie z symbiogenezą organelle komórek eukariotycznych, takie jak chloroplasty i wici, powstały poprzez wprowadzenie jednego prokariota do innej, większej komórki prokariotycznej, która pełniła rolę komórki gospodarza.
W hipotezie symbiotycznej pojawiają się trudności w wyjaśnieniu pochodzenia jądra komórki eukariotycznej oraz w kwestii, który prokariota był przecież gospodarzem. Dane z analizy molekularnej genomu i białek eukariotycznych pokazują, że z jednej strony był to organizm zbliżony do archeonów (kiedyś należały do bakterii, potem zostały rozdzielone na osobną gałąź). Z drugiej strony u eukariontów występują białka (i geny odpowiedzialne za ich syntezę), które są charakterystyczne dla doskonałych innych grup prokariotów.
Zgodnie z hipotezą inwazji pochodzenia komórek eukariotycznych, ich organelle powstały w wyniku inwazji cytoplazmy błona plazmatyczna z późniejszym rozdzieleniem tych struktur. Powstało coś w rodzaju kulek, otoczonych błoną i zawierających wewnątrz cytoplazmy oraz uchwycone tu związki i struktury. W zależności od tego, co dostało się do środka, powstały różne organelle.
Prokarioty nie mają prawdziwych organelli, ich funkcje są wykonywane przez inwazje błonowe. Dlatego łatwo sobie wyobrazić takie sznurowanie. Na korzyść hipotezy inwazji przemawia również podobieństwo błony cytoplazmatycznej i podwójnych błon organelli.
Z punktu widzenia wgłobienia pochodzenie jądra jest łatwe do wyjaśnienia, ale niewytłumaczalne jest, dlaczego genom i rybosomy kompleksu jądrowo-cytoplazmatycznego różnią się od tych w chloroplastach i mitochondriach (przypomnijmy, że zawierają one również DNA i rybosomy). Co więcej, w tych organellach system biosyntezy białek (DNA, RNA, rybosomy) jest podobny do tego u prokariotów.
Ta różnica jest dobrze wyjaśniona już z punktu widzenia pierwszej, symbiotycznej hipotezy. Według niej, w taki czy inny sposób, tlenowy prokariota dostał się do beztlenowego prokariota. Nie został strawiony, ale wręcz przeciwnie, zaczął żywić się kosztem komórki gospodarza. Z kolei tlen wykorzystywał do pozyskiwania energii, a ta metoda utleniania jest znacznie wydajniejsza, nadmiar energii oddał gospodarzowi prokariotycznemu, co w tym przypadku również skorzystało. Powstała symbioza. Następnie wprowadzony prokariota stał się prostszy, część jego genomu migrowała do komórki gospodarza, nie mógł już istnieć samodzielnie.
W podobny sposób symbiogeneza wyjaśnia pochodzenie chloroplastów. Wprowadzono już tylko prokarionty zdolne do fotosyntezy (podobnie jak sinice).
Pierwsze eukarionty bez symbiontów fotosyntetyzujących dały początek zwierzętom, w których się pojawiły - roślinom.
Obecnie istnieją pierwotniaki (jednokomórkowe organizmy eukariotyczne), które nie mają mitochondriów ani chloroplastów. Ale zamiast nich symbionty prokariontów osiedlają się w cytoplazmie, pełniąc odpowiednie funkcje. Fakt ten, jak również podobieństwo systemu biosyntezy białek mitochondriów i plastydów z prokariotami, uważa się za dowód symbiogenezy. Dowodem jest również to, że mitochondria i chloroplasty rozmnażają się same, nigdy nie są budowane przez komórkę od podstaw.
Na korzyść trzeciej, chimerycznej hipotezy pochodzenia komórek eukariotycznych mówi duży rozmiar ich genom, który tysiąckrotnie przewyższa bakteryjny, a także różnorodność syntetyzowanych białek różne grupy prokarioty. Oczywiste jest, że podczas ewolucji eukariontów ich genom stał się bardziej skomplikowany, podwoił się, pojawiło się w nim wiele genów regulatorowych. Mimo to początkowy wzrost wielkości genomu mógł nastąpić w wyniku połączenia genomów kilku prokariotów.
Być może w starożytności pewien prokariota nabył zdolność fagocytozy i jedząc w ten sposób wchłaniał między innymi inne prokarioty, które nie zawsze były trawione. Ich genom zawierał geny przydatne dla gospodarza, a on włączył je do swojego genomu. Być może niektóre z prokariotów, które były w środku, stały się organellami, co łączy hipotezę chimeryczną z symbiogenezą.
Ponieważ wszystkie powyższe hipotezy mają silne i słabe strony, a także w dużej mierze nie wykluczają się nawzajem, to naszym zdaniem połączenie wielu czynników opisanych różnymi hipotezami może odgrywać rolę w powstawaniu struktur komórek eukariotycznych.
Należy również zauważyć, że zgodnie z hipotezą symbiotyczną pochodzenie innych organelli błonowych, takich jak wakuole, kompleks Golgiego itp., Można uznać za dalsze uproszczenie, na przykład mitochondriów.
Zauważamy również, że połączenie białek z różnych prokariotów u eukariontów może wcale nie wskazywać na chimeryzm tych pierwszych. Możliwe, że pewne funkcje enzymatyczne mogą pełnić tylko białka o określonej strukturze. A ewolucja eukariontów wielokrotnie do tego dochodziła, niezależnie od ewolucji grup prokariotów, które nie są ich przodkami. Powiedzmy, że nastąpiła konwergencja na poziomie molekularnym.
Pojawienie się eukariontów jest ważnym wydarzeniem. Zmieniła strukturę biosfery i otworzyła zasadniczo nowe możliwości dla postępowej ewolucji. Komórka eukariotyczna jest wynikiem długiej ewolucji świata prokariotów, świata, w którym różnorodne drobnoustroje przystosowywały się do siebie i szukały sposobów na efektywną współpracę.
zarys osi czasu (powtórka)

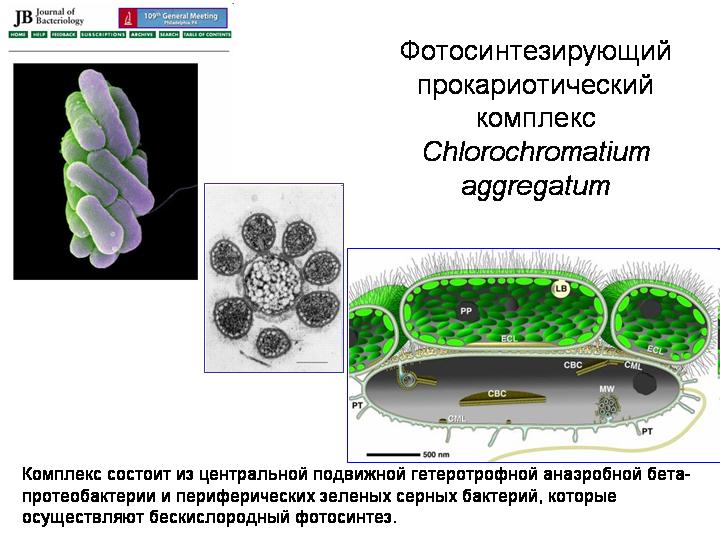
Fotosyntetyczny kompleks prokariotyczny Chlorochromatium aggregatum.
Eukarionty powstały w wyniku symbiozy kilku typów prokariotów. Ogólnie rzecz biorąc, prokarionty są dość podatne na symbiozę (patrz rozdział 3 w Narodzinach złożoności). Oto interesujący system symbiotyczny znany jako Chlorochromatium aggregatum. Żyje w głębokich jeziorach, gdzie na głębokości panują warunki beztlenowe. Centralnym składnikiem jest mobilna heterotroficzna beta-proteobakteria. Wokół niego piętrzą się od 10 do 60 fotosyntetyzujących zielonych bakterii siarkowych. Wszystkie elementy są połączone przerostami zewnętrzna męmbrana bakteria centralna. Znaczenie społeczności polega na tym, że mobilne beta-proteobakterie ciągną całe towarzystwo w miejsca sprzyjające życiu wybrednych bakterii siarkowych, a bakterie siarkowe zajmują się fotosyntezą i dostarczają pożywienia sobie i beta-proteobakteriom. Być może przodkami eukariontów były jakieś starożytne związki drobnoustrojów mniej więcej tego typu.
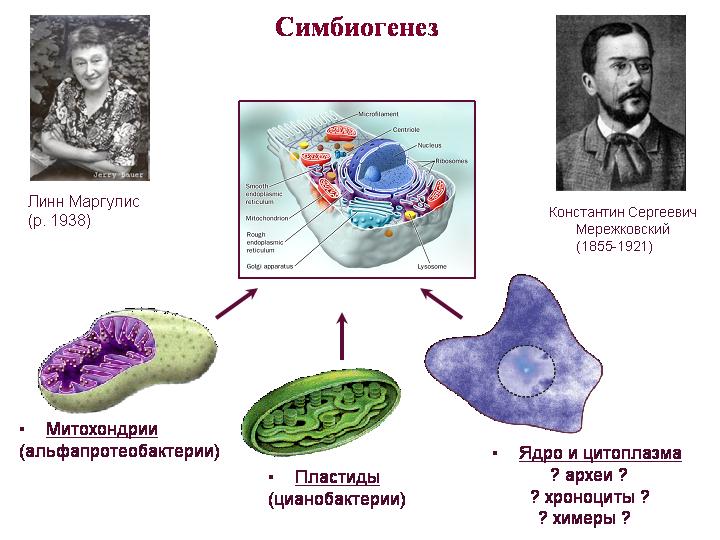
Teoria symbiogenezy. Mereżkowski, Margulis. Mitochondria są potomkami alfa-proteobakterii, plastydy są potomkami sinic. Trudniej jest zrozumieć, kto był przodkiem wszystkiego innego, to znaczy cytoplazmy i jądra. Jądro i cytoplazma eukariontów łączą cechy archeonów i bakterii, a także mają wiele unikalnych cech.
O mitochondriach. Być może to pozyskiwanie mitochondriów (a nie jądra) było kluczowym momentem w rozwoju eukariontów. Większość genów mitochondrialnych przodków została przeniesiona do jądra, gdzie znalazła się pod kontrolą jądrowych systemów regulacyjnych. Te geny jądrowe pochodzenia mitochondrialnego kodują nie tylko białka mitochondrialne, ale także wiele białek działających w cytoplazmie. Sugeruje to, że symbiont mitochondrialny odegrał ważniejszą rolę w tworzeniu komórki eukariotycznej, niż oczekiwano.
Współistnienie dwóch różnych genomów w jednej komórce wymagało opracowania efektywnego systemu ich regulacji. A żeby skutecznie zarządzać pracą dużego genomu, konieczne jest wyizolowanie genomu z cytoplazmy, w której zachodzi metabolizm i tysięcy reakcje chemiczne. Otoczka jądrowa po prostu oddziela genom od burzliwych procesów chemicznych zachodzących w cytoplazmie. Pozyskiwanie symbiontów (mitochondriów) może stać się ważnym bodźcem dla rozwoju jądra i systemów regulacji genów.
To samo dotyczy rozmnażania płciowego. Możesz żyć bez rozmnażania płciowego, o ile twój genom jest wystarczająco mały. Organizmy z dużym genomem, ale pozbawione rozmnażania płciowego, są skazane na szybkie wyginięcie, z nielicznymi wyjątkami.
![]()
Alphaproteobacteria - do tej grupy zaliczali się przodkowie mitochondriów.
Rhodospirillum to niesamowity mikroorganizm, który może żyć dzięki fotosyntezie, w tym w warunkach beztlenowych, jako tlenowy heterotrof, a nawet jako tlenowy chemoautotrof. Może na przykład rosnąć w wyniku utleniania tlenek węgla CO2 bez użycia innych źródeł energii. Oprócz tego wie również, jak wiązać azot atmosferyczny. To znaczy jest w najwyższy stopień organizm uniwersalny.
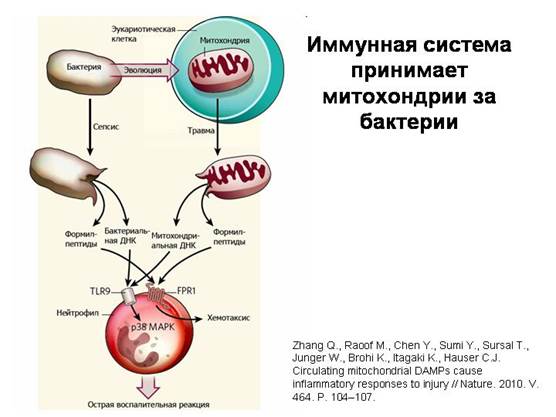
Układ odpornościowy myli mitochondria z bakteriami. Kiedy uszkodzone mitochondria dostają się do krwioobiegu podczas urazu, uwalniają charakterystyczne cząsteczki, które można znaleźć tylko w bakteriach i mitochondriach (koliste DNA i białka typu bakteryjnego, które na jednym końcu mają specjalny zmodyfikowany aminokwas formylometioninę). Wynika to z faktu, że aparat do syntezy białek w mitochondriach pozostał taki sam jak u bakterii. Komórki układ odpornościowy- neutrofile - reagują na te substancje mitochondrialne w taki sam sposób jak na bakteryjne i za pomocą tych samych receptorów. Jest to najwyraźniejsze potwierdzenie bakteryjnej natury mitochondriów.
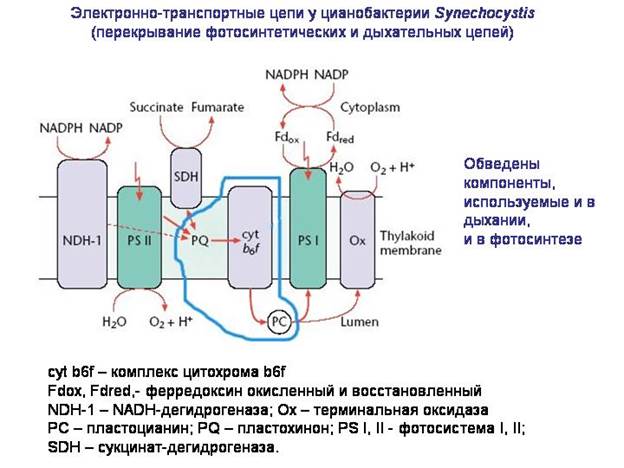
Główna funkcja mitochondria - oddychanie tlenem. Najprawdopodobniej bodźcem do asocjacji beztlenowego przodka jądra i cytoplazmy z „protomitochondriami” była potrzeba ochrony przed działanie toksyczne tlen.
Skąd bakterie, w tym alfaproteobakterie, wzięły niezbędne układy molekularne oddychanie tlenem? Wydaje się, że opierały się one na molekularnych systemach fotosyntezy. Łańcuch transportu elektronów, który powstał w bakteriach jako część aparatu fotosyntetycznego, został przystosowany do oddychania tlenem. W niektórych bakteriach części łańcuchów transportu elektronów są nadal używane jednocześnie zarówno w fotosyntezie, jak i oddychaniu. Najprawdopodobniej przodkami mitochondriów były tlenowe heterotroficzne alfa-proteobakterie, które z kolei wywodzą się od fotosyntetycznych alfa-proteobakterii, takich jak rhodospirillum.
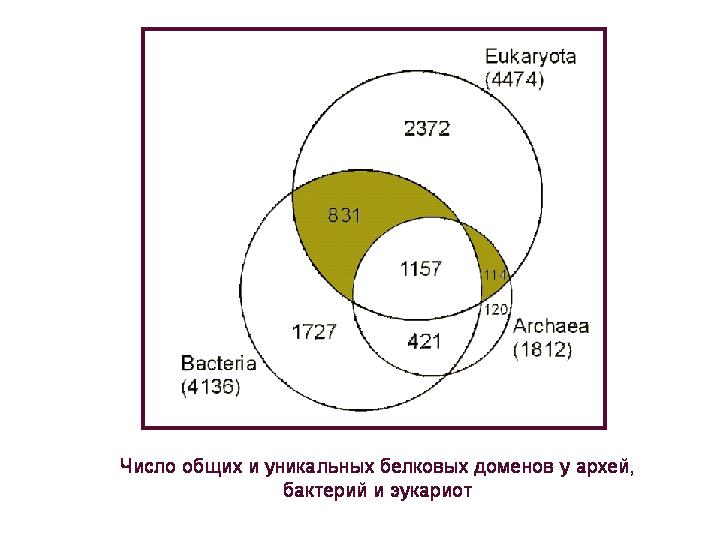
Liczba wspólnych i unikalnych domen białkowych u archeonów, bakterii i eukariontów. Domena białkowa to część cząsteczki białka, która ma określoną funkcję i charakterystyczną strukturę, czyli sekwencję aminokwasów. Każde białko z reguły zawiera jedną lub więcej takich strukturalnych i funkcjonalnych jednostek lub domen.
4,5 tys. domen białkowych, które posiadają eukarionty, można podzielić na 4 grupy: 1) dostępne tylko u eukariontów, 2) wspólne dla wszystkich trzech superkrólestw, 3) wspólne dla eukariotów i bakterii, ale nieobecne u archeonów; 4) wspólne dla eukariontów i archeonów, ale nieobecne u bakterii. Rozważymy dwa ostatnie grupy(są one zaznaczone kolorem na rysunku), ponieważ w przypadku tych białek można mówić z pewną pewnością o ich pochodzeniu: odpowiednio bakteryjnym lub archeologicznym.
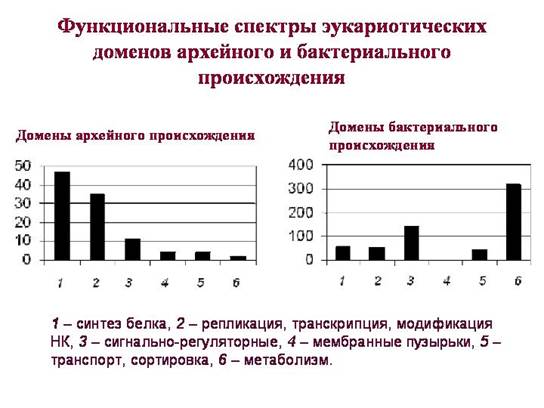
Kluczowy moment w tym, że domeny eukariotyczne rzekomo odziedziczone po bakteriach i archeonach mają znacząco różne funkcje. Domeny odziedziczone po archeonach (ich spektrum funkcjonalne pokazano na lewym wykresie) odgrywają kluczową rolę w życiu komórki eukariotycznej. Wśród nich dominują domeny związane z przechowywaniem, reprodukcją, organizacją i odczytywaniem informacji genetycznej. Większość domen „archeonów” należy do tych grup funkcyjnych, w obrębie których pozioma wymiana genów u prokariotów zachodzi najrzadziej. Najwyraźniej eukarionty otrzymały ten kompleks poprzez bezpośrednie (pionowe) dziedziczenie po archeonach.
Wśród domen pochodzenia bakteryjnego są też białka związane z procesami informacyjnymi, ale jest ich niewiele. Większość z nich działa tylko w mitochondriach lub plastydach. Eukariotyczne rybosomy cytoplazmy są pochodzenia archeologicznego, rybosomy mitochondriów i plastydów są pochodzenia bakteryjnego.
Wśród domen bakteryjnych eukariontów udział białek regulujących sygnał jest znacznie wyższy. Po bakteriach eukarionty odziedziczyły wiele białek odpowiedzialnych za mechanizmy odpowiedzi komórkowej na czynniki otoczenie zewnętrzne. A także - wiele białek związanych z metabolizmem (więcej szczegółów w rozdziale 3, „Narodziny złożoności”).
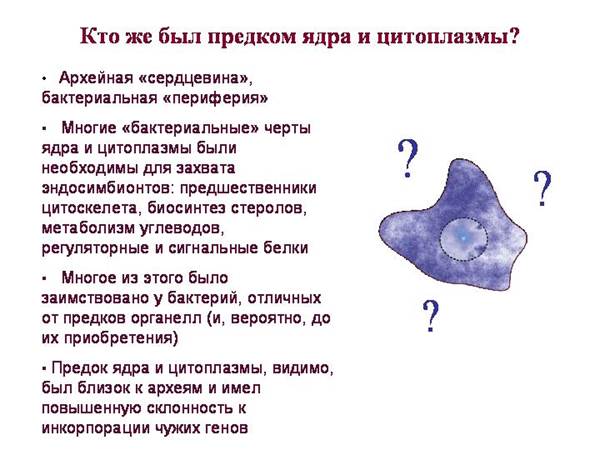
Eukarioty mają:
„Rdzeń” archeonów (mechanizmy pracy z informacją genetyczną i syntezą białek)
Bakteryjne „peryferia” (metabolizm i systemy regulujące sygnał)
Najprostszy scenariusz: ARCHEIA połknęła BAKTERIE (przodków mitochondriów i plastydów) i wszystkie ich objawy bakteryjne od nich kupiony.
· Ten scenariusz jest zbyt prosty, ponieważ eukarionty mają wiele białek bakteryjnych, których nie można było pożyczyć od mitochondrialnych lub plastydowych przodków.
Eukarionty mają wiele domen „bakteryjnych”, które nie są charakterystyczne ani dla sinic (przodków plastydów), ani alfaproteobakterii (przodków mitochondriów). Otrzymano je z innych bakterii.
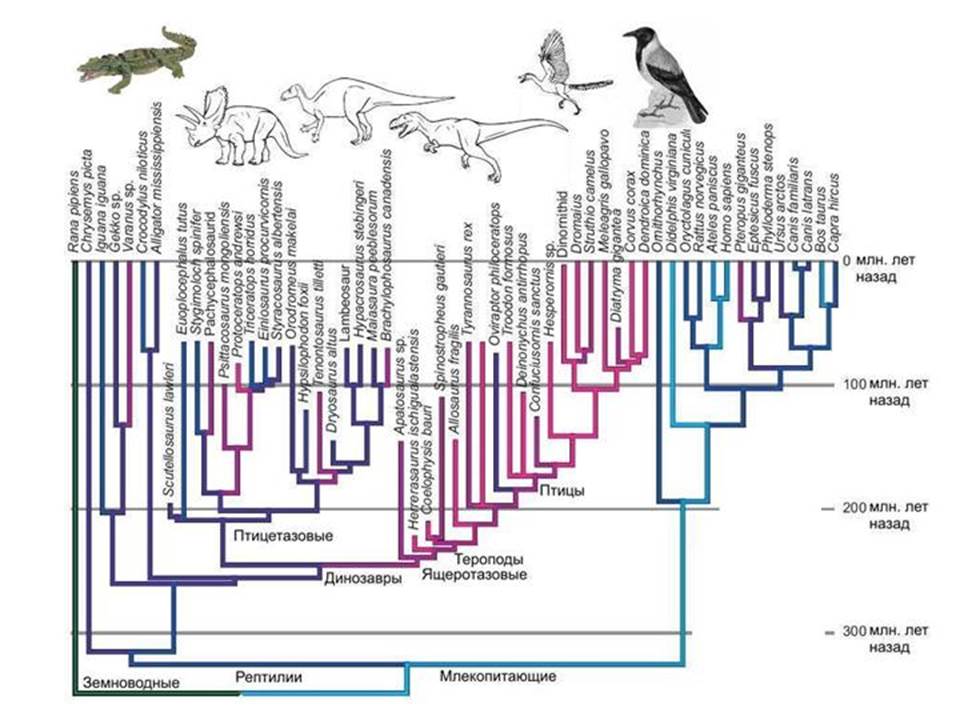
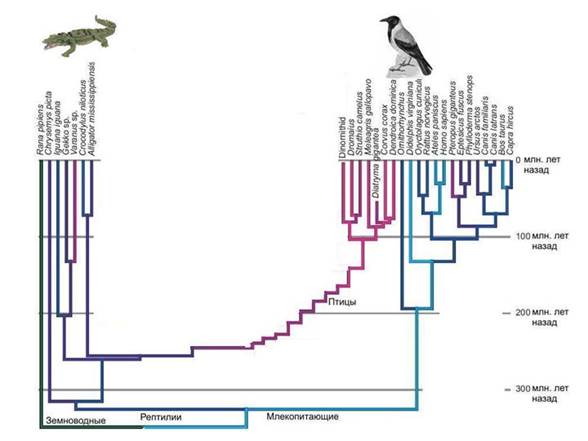
Ptaki i dinozaury. Rekonstrukcja protoeukariotów jest trudna. Oczywiste jest, że grupa starożytnych prokariotów, z której powstało jądro i cytoplazma, miała szereg unikalnych cech, których nie mają prokarioty, które przetrwały do dziś. A kiedy próbujemy zrekonstruować wygląd tego przodka, stajemy przed faktem, że pole do postawienia hipotez okazuje się zbyt duże.
Analogia. Wiadomo, że ptaki wywodzą się od dinozaurów, a nie od jakichś nieznanych dinozaurów, ale całkowicie pewna grupa- dinozaury maniraptory, które należą do teropodów, a teropody z kolei to jedna z grup dinozaurów jaszczurek. Znaleziono wiele form przejściowych między nielotnymi dinozaurami a ptakami.
Ale co moglibyśmy powiedzieć o przodkach ptaków, gdyby nie było zapisów kopalnych? W najlepszy przypadek dowiemy się, że najbliższymi krewnymi ptaków są krokodyle. Ale czy potrafilibyśmy odtworzyć wygląd bezpośrednich przodków ptaków, czyli dinozaurów? Ledwie. Ale właśnie w tej pozycji znajdujemy się, gdy próbujemy przywrócić wygląd przodka jądra i cytoplazmy. Jest jasne, że była to grupa niektórych dinozaurów prokariotycznych, wymarła grupa, która w przeciwieństwie do prawdziwych dinozaurów nie pozostawiła wyraźnych śladów w zapisie geologicznym. Współczesne archeony w stosunku do eukariontów są podobne współczesne krokodyle w kierunku ptaków. Spróbuj zrekonstruować budowę dinozaurów znając tylko ptaki i krokodyle.
Argument na rzecz tego, że w prekambrze żyło wiele drobnoustrojów, niepodobnych do obecnych. Stromatolity proterozoiczne były znacznie bardziej złożone i zróżnicowane niż współczesne. Stromatolity są produktem żywotnej aktywności społeczności drobnoustrojów. Czy to nie oznacza, że drobnoustroje proterozoiczne były również bardziej zróżnicowane niż współczesne i że wiele grup drobnoustrojów proterozoicznych po prostu nie przetrwało do dziś?

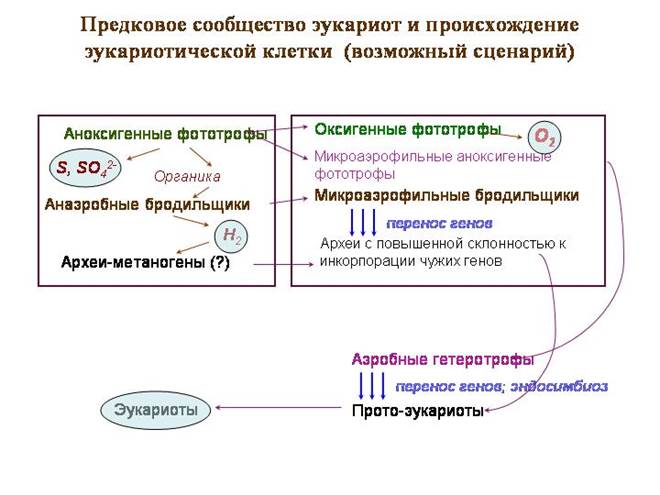
Społeczność przodków eukariontów i pochodzenie komórki eukariotycznej (możliwy scenariusz)
Hipotetyczna „społeczność przodków” to typowa mata bakteryjna, tylko w jej górnej części żyli przodkowie sinic, które nie przeszły jeszcze na fotosyntezę tlenową. Zajmowały się fotosyntezą beztlenową, donorem elektronów nie była woda, ale siarkowodór. Jak produkt uboczny wydzielała się siarka i siarczany.
Drugą warstwę zamieszkiwały fioletowe bakterie fotosyntetyzujące, w tym alfaproteobakterie, przodkowie mitochondriów. Fioletowe bakterie wykorzystują światło o dużej długości fali (czerwone i podczerwone). Fale te mają najlepszą siłę penetracji. Fioletowe bakterie nadal często żyją pod warstwą sinic. Fioletowe alfaproteobakterie również wykorzystują siarkowodór jako donor elektronów.
W trzeciej warstwie znajdowały się fermentujące bakterie przetwarzające materię organiczną; niektóre z nich emitowały wodór jako odpad. Stworzyło to bazę dla bakterii redukujących siarczany. Mogą też występować archeony metanogenne. Wśród żyjących tu archeonów byli przodkowie jądra i cytoplazmy.
Wydarzenia kryzysowe rozpoczęły się wraz z przejściem sinic do fotosyntezy tlenu. Jako donor elektronów zaczęto stosować cyjanobakterie zamiast siarkowodoru zwykła woda. To otworzyło wielkie możliwości, ale także Negatywne konsekwencje. Zamiast siarki i siarczanów podczas fotosyntezy zaczął uwalniać się tlen - substancja niezwykle toksyczna dla wszystkich starożytnych mieszkańców ziemi.
Pierwszymi, którzy zetknęli się z tą trucizną, byli jej producenci, sinice. Prawdopodobnie jako pierwsi opracowali środki ochrony przed nim. Łańcuchy transportu elektronów, które służyły do fotosyntezy, zostały zmodyfikowane i zaczęły służyć do oddychania tlenowego. Najwyraźniej pierwotnym celem nie było uzyskanie energii, a jedynie neutralizacja tlenu.
Wkrótce mieszkańcy drugiej warstwy zbiorowiska – purpurowych bakterii – musieli opracować podobne systemy obronne. Podobnie jak cyjanobakterie, rozwinęły systemy oddychania tlenowego oparte na systemach fotosyntezy. To purpurowe alfaproteobakterie rozwinęły najdoskonalszy łańcuch oddechowy, który obecnie funkcjonuje w mitochondriach eukariontów.
W trzeciej warstwie społeczności pojawienie się wolnego tlenu musiało wywołać kryzys. Wykorzystuje się metanogeny i wiele reduktorów siarczanów wodór cząsteczkowy przy pomocy enzymów hydrogenazy. Takie drobnoustroje nie mogą żyć w warunkach tlenowych, ponieważ tlen hamuje hydrogenazy. Z kolei wiele bakterii wytwarzających wodór nie rozwija się w środowisku, w którym nie ma mikroorganizmów, które go wykorzystują. Spośród fermentatorów społeczność najwyraźniej zachowała formy, które wydzielają związki o niskiej zawartości substancji organicznych (pirogronian, mleczan, octan itp.) Jako produkty końcowe. Te fermentory opracowały własne środki ochrony przed tlenem, mniej skuteczne. Wśród ocalałych były archeony - przodkowie jądra i cytoplazmy.
Być może w tym momencie kryzysu nastąpiło kluczowe wydarzenie - osłabienie izolacji genetycznej u przodków eukariontów i początek aktywnego pożyczania obcych genów. Proto-eukarioty włączyły geny różnych fermentatorów, aż same stały się mikroaerofilnymi fermentorami, fermentującymi węglowodany do pirogronianu i kwasu mlekowego.
Mieszkańcy trzeciej warstwy - przodkowie eukariontów - mieli teraz bezpośredni kontakt z nowymi mieszkańcami drugiej warstwy - tlenowymi alfaproteobakteriami, które nauczyły się wykorzystywać tlen jako źródło energii. Metabolizm protoeukariotów i alfaproteobakterii stał się komplementarny, co stworzyło warunki do symbiozy. A samo położenie alfaproteobakterii w społeczności (między górną warstwą, która uwalnia tlen, a dolną warstwą) z góry określiło ich rolę jako „obrońców” eukariotycznych przodków przed nadmiarem tlenu.
Jest prawdopodobne, że proto-eukarioty połknęły i nabyły wiele różnych bakterii jako endosymbionty. Tego rodzaju eksperymenty wciąż trwają. jednokomórkowe eukarionty, które mają ogromną różnorodność symbiontów wewnątrzkomórkowych. Spośród tych eksperymentów sojusz z alfaproteobakteriami tlenowymi okazał się najbardziej udany.
Biologia i genetyka
Zgodnie z hipotezą symbiotyczną popularnej obecnie komórki eukariotycznej organelle korpuskularne posiadające własny genom charakteryzują się niezależnym pochodzeniem i wywodzą się z komórki prokariotyczne symbionty. Początkowo ilość informacji w genomach komórki gospodarza z jednej strony i symbiontów domniemanych mitochondriów centrioli i chloroplastów z drugiej strony była najwyraźniej porównywalna. W przyszłości genomy symbiontów mogą utracić część swoich funkcji genetycznych wraz z przeniesieniem bloków genów do genomu…
Hipotezy pochodzenia komórek eukariotycznych.
1. Symbiotyczny.
Zgodnie z popularną obecnie hipotezą symbiotyczną, korpuskularne organelle komórki eukariotycznej, które posiadają własny genom, charakteryzują się niezależnym pochodzeniem i wywodzą się z prokariotycznych komórek symbiotycznych. Przypuszczalnie komórka gospodarza była beztlenowym prokariotem, zdolnym jedynie do ruchu ameboidalnego. Mitochondria i wici powstały w wyniku zmian w symbiontach komórek gospodarza, tlenowych prokariontów i bakterii, przypominających współczesne krętki. Chloroplasty komórek roślin zielonych powstały z symbiontów - komórek prokariotycznych niebiesko-zielonych alg. Szczególne znaczenie miało nabywanie przez komórkę podczas ewolucji wici o ciałach podstawnych zbliżonych do centrioli. To umożliwiło intensyfikację aktywność silnika i stworzyć mechanizm mitozy.
Kwestia pochodzenia jądra jest trudna. Początkowo ilość informacji w genomach komórki gospodarza z jednej strony i symbiontów przypuszczalnych mitochondriów, centrioli i chloroplastów z drugiej była najwyraźniej porównywalna. W przyszłości genomy symbiontów mogą utracić część swoich funkcji genetycznych wraz z przeniesieniem bloków genów do genomu komórki gospodarza. Towarzyszył temu wzrost objętości genomu jądrowego, centralizacja genetycznej regulacji funkcji komórki oraz utrata autonomii przez mitochondria i inne organelle korpuskularne. Nie można jednak wykluczyć, że jądro jest również pochodną symbiontu wewnątrzkomórkowego. Pochodzenie błon wewnątrzplazmatycznych szorstkiej i gładkiej retikulum cytoplazmatycznego, kompleksu blaszkowatego i struktur pęcherzykowo-wakuolowych tłumaczy się ewolucyjnymi przemianami błony zewnętrznej otoczka jądrowa. Hipoteza symbiotyczna nie wyjaśnia wielu faktów. Tak więc bakcylina białkowa, która tworzy rzęski i wici współczesnych prokariotów, różni się od białka tubuliny u eukariontów. Bakterie nie posiadały struktur o składzie mikrotubul „9+2” lub „9+0” typowych dla wici, rzęsek, ciałek podstawnych czy centrioli komórek eukariotycznych.
2. Inwazja.
Hipoteza inwazji dotycząca pochodzenia komórki eukariotycznej wywodzi się z faktu, że forma przodka była tlenowym prokariotem. Zawierał kilka genomów, z których każdy był dołączony Błona komórkowa. Korpuskularne organelle z DNA, jak również jądro, powstały w wyniku wgłobienia i splotu fragmentów otoczki z genomem, następnie funkcjonalnej specjalizacji w jądro, mitochondria, chloroplasty, powikłania genomu jądrowego i rozwój błon cytoplazmatycznych. Hipoteza ta w zadowalający sposób wyjaśnia obecność 2 błon w otoczce jądra, mitochondriów i chporoplasty. Napotyka trudności w wyjaśnieniu różnic w szczegółach procesu biosyntezy białek w organellach ciałka i cytoplazmie komórki eukariotycznej. W mitochondriach i chloroplastach proces ten dokładnie odpowiada procesowi we współczesnych komórkach prokariotycznych. Hipoteza symbiozy i inwazji nie wyczerpuje wszystkich punktów widzenia na pochodzenie eukariotycznego typu organizacji komórkowej.
3. Pojawienie się wielokomórkowości.
Formy kolonialne pierwotniaków heterotroficznych były przodkami organizmów wielokomórkowych.
Etap 1: w pobliżu każdej wici wyrastał z cytoplazmy kołnierz lub kieszonka, do której wbijano cząsteczki pokarmu (fagocytoza).
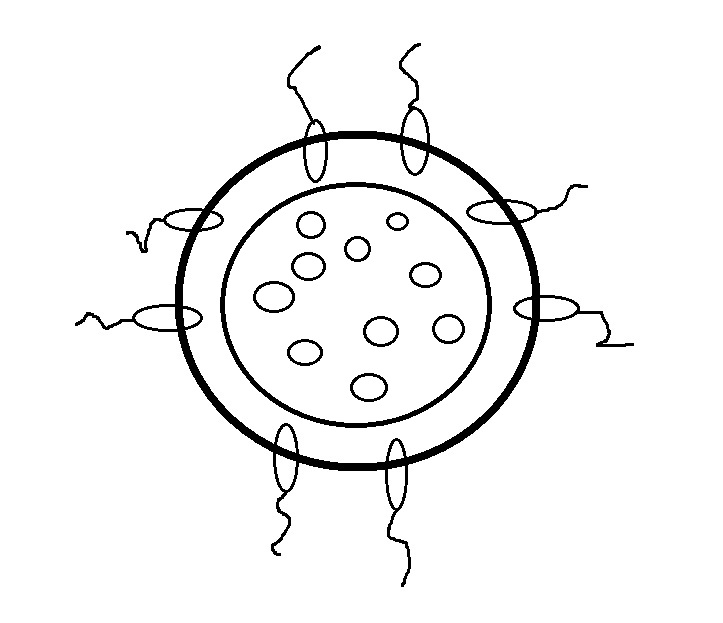
Etap 2: specjalizacja komórek fagocytów (funkcja fagocytarno-trawienna) i kinocytów (funkcja ruchowa; pozostała na obwodzie). Int. warstwa fagocytoblastu + ekst. kinocytoblast = fagocytella lub miąższ (Miecznikow).
Etap 3: na jednym biegunie tworzy się usta. dziura => dała początek gąbkom, koelenteratom itp.
Według Haeckela: kula, kolonia morska, wypełniając płyn, tworzy bąbelkowaty organizm (blastea), wgłobienie jest tworem dwuwarstwowym (gastrea); (Haeckel uważał, że pierwszym było trawienie brzuszne).
4. Cechy wielokomórkowe.
Zwierzęta wielokomórkowe tworzą największą grupę żywych organizmów na planecie, liczącą ponad 1,5 miliona gatunków. Wywodząc swoje pochodzenie od najprostszych, w procesie ewolucji uległy znaczącym przemianom związanym z komplikacjami organizacyjnymi.
Jedną z najważniejszych cech organizacji organizmów wielokomórkowych jest morfologiczna i funkcjonalna różnica między komórkami ich ciała. W toku ewolucji podobne komórki w ciele zwierząt wielokomórkowych specjalizowały się w wykonywaniu określonych funkcji, co doprowadziło do powstania tekstylia.
Połączenie różnych tkanin do organów i organów w układy narządów.Aby realizować relacje między nimi i koordynować ich pracę,systemy regulacyjnenerwowy i endokrynologiczny. Dzięki nerwowi regulacja humoralna działania wszystkich systemów organizm wielokomórkowy funkcjonuje jako integralny system biologiczny.
Dobrobyt grupy zwierząt wielokomórkowych wiąże się z komplikacją struktura anatomiczna I funkcje fizjologiczne. Tak więc wzrost wielkości ciała doprowadził do rozwoju przewód pokarmowy, co pozwalało im na spożywanie dużych ilości materiału spożywczego, dostarczającego dużej ilości energii do realizacji wszystkich procesów życiowych. Rozwinięty układ mięśniowo-szkieletowy zapewniał ruch organizmów, utrzymanie określonego kształtu ciała, ochronę i podparcie narządów. Zdolność do aktywny ruch pozwolono zwierzętom szukać pożywienia, znaleźć schronienie i osiedlić się.
Wraz ze wzrostem wielkości ciała zwierząt pojawiła się potrzeba pojawienia sięukłady krążenia wewnątrztransportowego,dostarczanie podtrzymywania życia do tkanek oddalonych od powierzchni ciała” i narządów składniki odżywcze, tlen, a także usuwanie końcowych produktów przemiany materii.
Taki krążeniowy układ transportowy stał się płynną tkanką krwi.
Intensyfikacja czynności oddechowej następowała równolegle z postępującym rozwojemsystem nerwowy i narządy zmysłów. Nastąpił ruch wydziały centralne układu nerwowego do przedniego końca ciała zwierzęcia, w wyniku czego część głowy została odizolowana. Taka budowa przedniej części ciała zwierzęcia pozwalała mu na otrzymywanie informacji o zmianach środowisko i odpowiednio na nie reagować.
Zwierzęta wielokomórkowe są niezwykle zróżnicowane pod względem budowy, cech życia, różnej wielkości, masy ciała itp.
Lekcja odpowiada programowi i podręcznikowi I.N. Ponomareva, które przewidują badanie cytologii w 11 klasie. Lekcja ma formę dyskusji naukowej i jest przewidziana na 90 minut.
Cele Lekcji: zapoznanie się z najważniejsze hipotezy o pochodzeniu komórki eukariotycznej i organizmów wielokomórkowych; rozwijanie umiejętności argumentowania i obrony własnego punktu widzenia; krzewienie kultury dyskusji publicznej.
Sprzęt: tablice przedstawiające budowę komórki i jej poszczególnych organelli, schemat powstawania komórki eukariotycznej, rysunki przedstawiające trichoplax i różne formy alg.
PODCZAS ZAJĘĆ
Nauczyciel. Wraz z rozwojem nauki o komórce – cytologii i pojawieniem się nowych wyników badań tej elementarnej układ biologiczny wybuchły gorące debaty między naukowcami, które przerodziły się w poważne dyskusje naukowe. W całej historii cytologii jako nauki niemal każda nowa hipoteza zyskała zarówno zwolenników, jak i przeciwników.
Dyskusja naukowa to publiczna dyskusja na temat pytania lub problemu. Dzisiaj na lekcji przeprowadzimy dyskusję, podczas której omówimy dwa kwestie krytyczne cytologia: problem pochodzenia pierwszych eukariontów i problem pochodzenia pierwszych organizmów wielokomórkowych.
Wiecie, że wcześniej klasa była podzielona na grupy, które będą prezentować i bronić różnych punktów widzenia na te dwa zagadnienia, które omawiamy. Przypomnę nazwy tych grup.
1. Zwolennicy autogenicznej hipotezy pochodzenia komórki eukariotycznej i jej organelli.
2. Zwolennicy symbiogenicznej hipotezy pochodzenia komórki eukariotycznej i jej organelli.
3. Zwolennicy hipotezy o kolonialnym pochodzeniu pierwszych organizmów jednokomórkowych.
4. Zwolennicy hipotezy o pochodzeniu organizmów wielokomórkowych z orzęsków poprzez komórkowanie ich komórek.
Przedstawiciele każdej grupy w krótkiej prezentacji przedstawią istotę takiego czy innego punktu widzenia na każdy z omawianych problemów, a następnie odpowiedzą na pytania, uwagi, zastrzeżenia zarówno swoich naukowych „przeciwników”, jak i wszystkich obecnych na zajęciach.
Problem pochodzenia organelle błonowe i jądra komórek eukariotycznych jest jednym z najbardziej aktualnych tematów współczesnej cytologii. Aby rozwiązać ten problem, w nauce zaproponowano kilka różnych hipotez. Przyjrzyjmy się dwóm najważniejszym z nich.
Autogeniczna hipoteza pochodzenia komórki eukariotycznej
Wystąpienie przedstawicieli pierwszej grupy studentów
Pierwsze eukarionty pojawiły się na Ziemi 1,1–1,4 miliarda lat temu (w erze proterozoicznej). Był to kolejny kamień milowy w ewolucji prekambru po pojawieniu się fotosyntezy.
Hipoteza autogeniczna, czyli sukcesyjna, głosi, że złożona komórka eukariotyczna rozwinęła się bezpośrednio z komórki prokariotycznej: w wyniku wniknięcia błony plazmatycznej, a następnie jej przegrupowania w celu pełnienia określonych funkcji, powstały najważniejsze organelle. A tak złożone organelle, jak mitochondria i plastydy, wywodzą się z wewnątrzplazmatycznych struktur błonowych struktury rurowej obecnych u prokariotów.
pytania
1. Co przemawia za tym punktem widzenia? Czy istnieją formy przejściowe, tj. komórki prokariotyczne z „podstawami” organelli eukariotycznych?
Odpowiedź. Wśród żywych organizmów nie ma takich form reliktowych. Przypomnijmy jednak obecność mezosomów w komórkach bakteryjnych, tj. struktury błonowe o kształcie rurkowatym i pęcherzykowym (kulistym), utworzone przez inwaginację błony plazmatycznej do cytoplazmy. Przyjmuje się, że mezosomy biorą udział w tworzeniu ścian komórkowych, replikacji DNA i innych procesach. Jest całkiem możliwe, że to właśnie na bazie mezosomów kompleks błonowy stopniowo powstawał i różnicował się, dając początek różnym organelli komórki eukariotycznej.
2. Wiadomo, że skład biochemiczny wielu organelli, w szczególności mitochondriów i chloroplastów, znacznie różni się od składu błony komórkowej cytoplazmy. Czy to nie wskazuje, że ich pochodzenie nie pochodzi z oryginału Błona komórkowa- błony plazmatyczne?
Odpowiedź. Rzeczywiście, skład białek mitochondriów i chloroplastów jest wyjątkowy. Ale ta oryginalność mogła zostać nabyta po raz drugi, mogła być wynikiem przystosowania się do pełnienia przez te organelle pewnych funkcji.
Nauczyciel. Posłuchajmy argumentów zwolenników symbiogenicznego pochodzenia eukariontów i ustalmy, która hipoteza jest bardziej popularna na zajęciach.
Hipoteza symbiogenna pochodzenia komórki eukariotycznej
Wystąpienie przedstawicieli drugiej grupy studentów
Hipoteza symbiogeniczna (obecnie często nazywana teorią) wywodzi się z faktu, że eukarionty są wynikiem symbiozy między różnymi prokariotami.
Już na początku XX wieku. Rosyjscy botanicy A.S. Famintsyn, B.M. Kozo-Polyansky i K.S. Mereżkowski wysunął hipotezę, że komórka roślin zielonych (eukariotów) otrzymała plastydy w wyniku symbiozy komórki wolnej od chlorofilu z komórkami sinic. Ta hipoteza o symbiogennym pochodzeniu komórki eukariotycznej wyprzedziła swoje czasy, została zapomniana i ponownie zwróciła na siebie uwagę w połowie XX wieku.
Nazwana hipoteza została opracowana przez Lynn Sagan-Margulis (1983). Zgodnie z tą hipotezą, pierwotna komórka dużej bakterii prokariotycznej, która weszła w symbiozę z komórkami niebiesko-zielonych alg, nabyła plastydy. Symbioza z heterotroficznymi komórkami prokariotycznymi doprowadziła do ich przekształcenia w mitochondria. Niektóre komórki, będąc heterotrofami, wychwytywały inne, mniejsze bakterie, którymi były nieznane przyczyny nie strawił. Wychwycone małe komórki prokariotyczne były w stanie wchłonąć tlen. Ta właściwość była korzystna dla komórki, która pochłonęła bakterię, ponieważ dała jej znacznie więcej energii zmagazynowanej w cząsteczkach ATP.
pytania
1. Jakie dowody przemawiają za symbiogennym pochodzeniem plastydów i mitochondriów?
Odpowiedź. Dowody na poparcie tego poglądu są następujące.
1. Mitochondria i chloroplasty są otoczone podwójną błoną.
2. Te organelle rozmnażają się przez podział i nie wyrastają z żadnych innych organelli błonowych.
3. Mitochondria i chloroplasty mają swój własny materiał genetyczny, który koduje tylko ich własne białka.
4. DNA tych organelli ma strukturę kolistą, jak u prokariotów, a nie liniową, jak u eukariontów.
5. Mitochondria i chloroplasty mają własne mechanizmy syntezy RNA i białek, a ich rybosomy są bardziej prokariotyczne niż eukariotyczne.
6. Niektóre białka tych organelli są podobne w swojej podstawowej strukturze do podobnych białek bakteryjnych i nie są podobne do odpowiednich białek cytoplazmatycznych.
2. Wiadomo, że nie wszystkie białka mitochondriów i chloroplastów są syntetyzowane zgodnie z własnym DNA, niektóre białka tych organelli są zakodowane w DNA jądra. Jak możesz wyjaśnić ten fakt?
Odpowiedź. Rzeczywiście, tylko część ich białek jest zakodowana we własnym DNA mitochondriów i chloroplastów, podczas gdy reszta jest zakodowana w DNA jądra komórkowego. Najwyraźniej w toku ewolucji część materiału genetycznego „przepłynęła” z genomów mitochondriów i chloroplastów do genomu jądrowego. Genom chloroplastów jest bardziej obszerny niż genom mitochondrialny, a genom mitochondrialny niższych eukariotów (np. drożdży) jest bardziej obszerny niż genom ssaków, co pośrednio potwierdza powyższe założenie.
3. Wiemy, że oprócz mitochondriów i chloroplastów niewielka ilość DNA w komórce eukariotycznej jest również zawarta w centriolach i podstawie wici. Czy one również powstały symbiogenicznie?
Odpowiedź. Tak. Symbioza z bakteriami podobnymi do krętków może prowadzić z jednej strony do pojawienia się wici, az drugiej do przekształcenia niektórych z tych komórek podobnych do krętków w centriole. Współczesna koncepcja symbiogenezy głosi, że komórka eukariotyczna powstała w wyniku kilku następujących po sobie aktów symbiogenezy.
4. Jak powstały pozostałe organelle komórki eukariotycznej (zespół Golgiego, EPS, lizosomy)?
Odpowiedź. Przypuszcza się, że retikulum endoplazmatyczne, lizosomy i aparat Golgiego mogły powstać w wyniku inwazji błony plazmatycznej.
5. Jak, zgodnie z hipotezą symbiogeniczną, mogło powstać jądro otoczone podwójną błoną?
Odpowiedź. Przypuszcza się, że jądro rozwinęło się również w wyniku izolacji DNA z cytoplazmy przez wpuklenie błony plazmatycznej i otoczenie substancji jądrowej.
6. Z Twoich odpowiedzi na dwa ostatnie pytania wynika, że powstanie komórki eukariotycznej to nie tylko efekt dziedzicznej symbiozy kilku prokariotów. Co na to powiesz?
Odpowiedź. Tak to jest. Zwolennicy hipotezy symbiogenicznej przyznają, że niektóre struktury komórki eukariotycznej powstały w wyniku jej samorozwoju i komplikacji.
7. Przyniosłeś dane biochemiczne, genetyczne, z mikroskopu elektronowego ostatnie lata na korzyść symbiogenetycznego pochodzenia komórki eukariotycznej. Ale czy istnieją relikty ewolucyjne, tak zwane żywe skamieniałości, które wskazują na powiązania między prokariotami i eukariontami?
Odpowiedź. Takie zależności obserwuje się między organizmami roślinnymi i zwierzęcymi. Na przykład wolno żyjąca ameba Pelomyxa palustris(bagno Pelomyx) nie ma mitochondriów, ale zawiera bakterie symbiotyczne, które zapewniają mu oddychanie.
Nauczyciel. Wydaje mi się, że przedstawicielom drugiej grupy zadawano wystarczająco dużo pytań. Zwolennicy jakiej hipotezy przedstawili, Pana zdaniem, bardziej przekonujące argumenty i jaka hipoteza jest bardziej rozpoznawalna w nauce?
Rzeczywiście, hipoteza symbiogeniczna znajduje wielu zwolenników. Należy jednak pamiętać, że rozważania na rzecz hipotezy symbiogenicznej nie mają potwierdzenia eksperymentalnego, dlatego nie można jej uznać za teorię. Moim zdaniem należy również pamiętać, że symbioza odegrała znaczącą, ale nie jedyną rolę w powstaniu komórki eukariotycznej. Znaczenie w powstawaniu eukariontów zachodziły również procesy samorozwoju komórek prokariotycznych.
Przejdźmy do następny problem- problem pochodzenia organizmów wielokomórkowych.
Hipotezy kolonialnego pochodzenia pierwszych metazoanów
Nauczyciel. Postępy w rozwoju cytologii i biologii molekularnej udowodniły strukturalny i biochemiczny związek organizmów jednokomórkowych i wielokomórkowych. Wszystko to potwierdziło hipotezę pochodzenia zwierząt wielokomórkowych z formy jednokomórkowe. Główne spory naukowe dotyczą kwestii, z jakich pierwotniaków wywodzą się zwierzęta wielokomórkowe i jak wyglądały pierwsze zwierzęta wielokomórkowe? Istnieje wiele hipotez dotyczących pochodzenia zwierząt wielokomórkowych od pierwotniaków. Rozważmy niektóre z nich.
Wystąpienie przedstawicieli trzeciej grupy studentów
Słusznie zauważono, że istnieje wiele hipotez dotyczących pochodzenia zwierząt wielokomórkowych od pierwotniaków. Najbardziej uzasadnioną z nich, naszym zdaniem, należy uznać za hipotezę kolonialną.
Pierwszą kolonialną hipotezę pochodzenia organizmów wielokomórkowych zaproponował Ernst Haeckel. Opracował swoją „teorię gastrei”, zgodnie z którą wspólny przodek wszystkich zwierząt wielokomórkowych jest podobny do swobodnie pływającej larwy koelenteratów - koralowców z dwiema warstwami komórek.
Rosyjski biolog Ilja Iljicz Miecznikow zaproponował inną hipotezę: zwierzęta wielokomórkowe powstały z kolonialnych pierwotniaków wiciowych zdolnych do fagocytozy, tj. wychwytywanie pokarmu przez komórki z trawieniem wewnątrzkomórkowym. Teoria ta zakłada, że komórki powierzchniowe, które przechwyciły pokarm, zatapiają się głęboko w ciele kolonii w celu trawienia, robiąc miejsce dla głodnych komórek. Rezultatem jest organizm, w którym komórki warstwy wewnętrznej trawią pokarm, a warstwa zewnętrzna, składająca się z głodnych komórek, go wychwytuje. Pełni również funkcje recepcji, ruchu i ochrony. Miecznikow nazwał proponowany organizm phagocetella (organizm składający się z komórek zaangażowanych w fagocytozę).
Wreszcie trzecią hipotezę wysunął niemiecki zoolog O. Byuchli. Według niej pierwotny organizm wielokomórkowy składał się z dwóch warstw komórek, ponadto dolną stroną pełzał po ziemi, a Górna strona pełnił funkcje ochronne i czułe. Po spotkaniu z pokarmem o dużych rozmiarach taki organizm owinął się wokół niego i strawił dolną warstwą. Bütschli nazwał ten domniemany organizm placela (zwierzę w postaci otaczającego ciasta).
pytania
1. Którą z tych trzech hipotez wolisz? Który z nich jest najbardziej rozpoznawalny w nauce?
Odpowiedź. Trudno jest preferować jakąkolwiek hipotezę, ponieważ nie ma danych paleontologicznych na temat pierwszych etapów ewolucji zwierząt wielokomórkowych. Naukowcy próbują znaleźć potwierdzenie hipotez o kolonialnym pochodzeniu zwierząt wielokomórkowych, porównując hipotetyczne organizmy z żywymi prymitywnymi formami zwierząt wielokomórkowych.
Najbardziej prymitywnym znanym obecnie zwierzęciem wielokomórkowym jest Trichoplax (ryc. 1). Opisał go Artemy Wasiljewicz Iwanow (1973). Trichoplax został znaleziony na europejskim wybrzeżu Oceanu Atlantyckiego. To zwierzę ma wygląd cienkiej płytki z dwóch warstw komórek z wici. W jamie jego ciała znajdują się oddzielne komórki trawienne. Pomiędzy dwiema warstwami komórek znajdują się komórki, które wyglądają jak ameby.
Po przestudiowaniu Trichoplax, A.V. Iwanow doszedł do wniosku, że jest bardzo podobny do hipotetycznej phagocetelli I.I. Miecznikowa i jest właściwie jego żywym modelem. Z drugiej strony Trichoplax też wygląda jak placola. Tak więc odkrycie Trichoplax, podobnie jak larwy koelenteratów, łączy hipotezy Haeckela (1866), Miecznikowa (1877) i Buchliego (1884).
2. Przez długi czas pochodzenie prymitywnych zwierząt wielokomórkowych wiązano z zielonymi wiciowcami kolonialnymi, takimi jak eudorina, volvox, pandorina. Czy nie są one obecnie uważane za formy przejściowe do wielokomórkowości?
Odpowiedź. Rzeczywiście, pochodzenie zwierząt wielokomórkowych nie jest już kojarzone z zielonymi wiciami kolonialnymi. Ten punkt widzenia opiera się na fakcie, że ich indywidualny rozwój i rozmnażanie jest zbliżone do glonów, a nie do prymitywnych zwierząt wielokomórkowych.
3. Jakie prymitywne zwierzęta wielokomórkowe dały początek hipotetycznemu organizmowi kolonialnemu? Czy były to gąbki, koelenteraty lub przedstawiciele jakiegoś innego rodzaju zwierząt?
Odpowiedź. Przyjmuje się, że kilka rodzajów zwierząt wielokomórkowych o różnym poziomie organizacji wywodzi się jednocześnie od przodków podobnych do fagocetelli: gąbki, koelenteraty i prymitywne trójwarstwowe zwierzęta zbliżone do niejelitowych planarian należących do płazińców. Gąbki i koelenteraty to ślepe zaułki ewolucji. Wąska specjalizacja tych typów, ich zdolność do przystosowania się do nieruchomego lub biernego trybu życia zamykała możliwości ich dalszego progresywnego rozwoju.
4. Ciągle mówicie o pochodzeniu pierwszych zwierząt wielokomórkowych, ale wielokomórkowość jest charakterystyczna nie tylko dla przedstawicieli królestwa zwierząt. Jak mogłoby być grzyby wielokomórkowe, rośliny?
Odpowiedź. Współczesna nauka uważa, że rośliny nie mają ostrej granicy między jednokomórkowością a wielokomórkowością. Wielokomórkowość wielokrotnie pojawiała się niezależnie od siebie w różnych działach roślin. Tak więc w złocistych, wielobiczowatych, pirofitycznych, zielonych algach obserwuje się gatunki o strukturze nitkowatej, kolonialno-wielokomórkowej i syfonowej (ciało składa się z jednej wielojądrzastej komórki olbrzymiej). Obecność takich form wskazuje naszym zdaniem na próby przebicia się na kolejny poziom organizacji – wielokomórkowość, gdzie różne komórki pełnią różne funkcje(Rys. 2).
Ryż. 2. Różne formy glony (1 - jednokomórkowe;
2 - kolonialny; 3 - wielordzeniowy; 4 - nitkowate)
Informacje dostępne dla współczesnej nauki na temat rozwoju form wielokomórkowych w prekambrze są skąpe. Dotyczy to zwłaszcza grzybów. Powodem jest słabe zachowanie szczątków tych organizmów. Podobnie jak w wielu roślinach, grzyby przechodzą przemianę ze struktury niekomórkowej (syfonowej, jak np. śluz grzyba pleśniowego) do wielokomórkowej. Tak więc, naszym zdaniem, na pewnym etapie rozwoju grzybów (około 1,3 miliarda lat temu) u niektórych z nich grzybnia syfonalna przekształciła się w formację wielokomórkową.
Nauczyciel. Podsumujmy to, co zostało powiedziane. Organizmy wielokomórkowe wg nowoczesna nauka, wcześniej pojawił się wśród roślin i grzybów. U zwierząt pierwszy formy wielokomórkowe powstał około 700 milionów lat temu. Początki zwierząt wielokomórkowych dały starożytne heterotroficzne kolonialne organizmy wiciowe.
Hipoteza o niekolonialnym pochodzeniu pierwszych metazoanów
Wystąpienie przedstawicieli czwartej grupy studentów
Przedstawiciele poprzedniej grupy twierdzą, że odległymi przodkami zwierząt wielokomórkowych były kolonie pierwotniaków. Chociaż jest to powszechny, ale nie jedyny punkt widzenia.
Przedstawiamy hipotezę sugerującą, że w procesie ewolucji pojedyncze pierwotniaki przekształciły się całkowicie w stworzenia wielokomórkowe. Pomysł ten został wysunięty przez słynnego serbskiego zoologa I. Hadji. Jego zdaniem zwierzęta wielokomórkowe wyewoluowały z orzęsków wielojądrzastych.
Orzęski mają dość złożoną strukturę. Ich cytoplazma jest reprezentowana przez dwie warstwy - obwodową i centralną, w których zachodzi trawienie wewnątrzkomórkowe. Orzęski mają komórkowe usta, gardło, kurczliwe wakuole z kanałami przywodzicieli. Wszystkie te różnie zróżnicowane części organizm jednokomórkowy- organelle - Hadji uważa za prototyp organów zwierzęcia wielokomórkowego. Tak, on tak myśli skóra stworzenia wielokomórkowe powstały z zewnętrznej warstwy cytoplazmy (ektoplazmy), a ich jelita - z wewnętrznej warstwy cytoplazmy (entoplazmy).
Przejście ze stanu jednokomórkowego do stanu wielokomórkowego mogło nastąpić w ciele orzęsków natychmiast poprzez utworzenie granic komórkowych wokół poszczególnych jąder i przylegających odcinków cytoplazmy. Ten rzekomy proces nazywa się „celularyzacją” (od łacińskiego słowa celuloza- komórka), a sama hipoteza nazywa się teorią komórkowości.
pytania
1. Czy hipoteza I. Hadji ma jakieś poparcie w embriologii niższych organizmów wielokomórkowych, jak np. hipoteza I.I. Miecznikow?
Odpowiedź. Niestety, nie ma takich danych embriologii porównawczej. Nikt nigdy nie obserwował procesu komórkowania orzęsków w przyrodzie.
2. Czy nie sądzisz, że hipoteza Hadjiego nie wytrzymuje krytyki, przede wszystkim dlatego, że sama zasada leżąca u jej podstaw jest błędna. Hipoteza komórkowości jest naszym zdaniem sprzeczna z głównymi założeniami teorii komórkowej, a mianowicie: zrównuje części pojedynczej komórki z tkankami i narządami organizmów wielokomórkowych. Co możesz o tym powiedzieć?
Odpowiedź. Nie bylibyśmy tak kategoryczni w osądach. Hipoteza Hadjiego nie utożsamia orzęsków z tkankami i narządami organizmów wielokomórkowych. Tylko ona się liczy ewentualna edukacja podobne narządy oparte na orzęskowych organellach.
3. Na czym jeszcze opiera się hipoteza Hadjiego, oprócz tego, że orzęski mają złożona struktura a ich struktury mogłyby stać się narządami zwierząt wielokomórkowych?
Odpowiedź. Pośrednim potwierdzeniem hipotezy jest pewne podobieństwo między orzęskami a turbellarianami - prymitywnymi płazińce: bliskie rozmiary, ten sam kształt ciała, lokalizacja wielu struktur itp. (Rys. 3 i 4).
Nauczyciel. Wysłuchaliśmy argumentów przemawiających za kolonialnym i niekolonialnym pochodzeniem pierwszych zwierząt wielokomórkowych, zapoznaliśmy się z poglądami na temat pochodzenia roślin wielokomórkowych i grzybów. Jaka, twoim zdaniem, hipoteza - kolonialne lub niekolonialne pochodzenie zwierząt wielokomórkowych - jest bardziej uzasadniona i udowodniona?
Z pewnością hipoteza o kolonialnym pochodzeniu pierwszych zwierząt wielokomórkowych. To właśnie ta hipoteza znajduje największe uznanie w świecie naukowym. Należy jednak pamiętać, że druga hipoteza stale ewoluuje i zmienia się.
Podsumowanie lekcji
1. Ocena za lekcję.
2. Praca domowa:
- przestudiować paragraf „Dyskusyjne problemy cytologii”;
f – zaznajomienie się w domu z jednym z omawianych w XVIII wieku zagadnień cytologii, prowadzonej między różnymi zwolennikami preformizmu; przygotować system argumentów przemawiających za różnymi punktami widzenia.