Pytanie 1. Jakie są różnice w budowie komórek eukariotycznych i prokariotycznych?
Prokarionty nie mają jądra o prawdziwym kształcie (gr. karion - jądro). Ich DNA to pojedyncza kolista cząsteczka, swobodnie zlokalizowana w cytoplazmie i nie otoczona błoną. Na komórki prokariotyczne nie ma plastydów, mitochondriów, retikulum endoplazmatycznego, aparatu Golgiego, lizosomów. Zarówno prokarionty, jak i eukarionty mają rybosomy (jądrowe mają większe). Witka komórki prokariotycznej jest cieńsza i działa na innej zasadzie niż wici eukariontów. Organizmy eukariotyczne to grzyby, rośliny, zwierzęta - jednokomórkowe i wielokomórkowe; prokariota - bakterie i niebieskozielone algi (cyjanobakterie).
Pytanie 2. Opowiedz nam o pinot - i fagocytozie. Czym różnią się te procesy?
Błona komórkowa jest ruchomą formacją zdolną do wychwytywania obiektów środowiska zewnętrznego poprzez tworzenie inwazji i wyrostków. Ten proces nazywa się endocytozą. Przyczyną endocytozy są złożone reakcje biochemiczne zachodzące w cytoplazmie i związane przede wszystkim ze zmianami trzeciorzędowej struktury białek wewnątrzkomórkowych. Jeśli komórka przechwytuje kroplę płynu - jest to pinocytoza, jeśli cząstka stała - fagocytoza. W rezultacie powstają wakuole pinocytowe lub fagocytarne (pęcherzyki błonowe). Proces odwrotny do endocytozy (uwolnienie zawartości wakuoli z komórki) nazywany jest egzocytozą.
Pytanie 3. Rozwiń związek między strukturą a funkcjami błony komórkowej.
Wiadomo, że podstawą każdej błony jest dwuwarstwowa (podwójna warstwa) fosfolipidów, w której hydrofilowe „głowy” cząsteczek (glicerolu) są skierowane na zewnątrz, a reszty hydrofobowe Kwasy tłuszczowe- wewnątrz. Cząsteczki białek są związane z dwuwarstwą lipidową, która może przylegać do błony z obu stron, zatapiać się w niej, a nawet ją przenikać. Położenie błony komórkowej na granicy komórki i środowisko określa jego główne funkcje. Mocna, elastyczna, łatwo regenerująca się dwuwarstwa stanowi barierę zapewniającą stałość środowiska wewnątrzkomórkowego i chroniącą cytoplazmę przed wnikaniem obcych substancji. Funkcja transportowa błony jest selektywna. Małe nienaładowane cząsteczki (02, N2) łatwo przenikają bezpośrednio przez dwuwarstwę. Większe i/lub naładowane cząsteczki (Na+, K+, niektóre hormony) przechodzą przez specjalne pory (kanały) białkowe lub są transportowane przez białka nośnikowe. Będąc strukturą ruchomą, błona komórkowa może również przeprowadzać transport substancji na drodze endo - i egzocytozy.
Pytanie 4. Jakie organelle komórkowe znajdują się w cytoplazmie?
Organelle znajdujące się w cytoplazmie komórek eukariotycznych można podzielić na trzy grupy: jednobłonowe, dwubłonowe i bezbłonowe. Organelle jednobłonowe obejmują retikulum endoplazmatyczne (gładkie i szorstkie), aparat Golgiego, lizosomy i wakuole. Dwa błoniaste organelle to plastydy i mitochondria; niebłonowe - rybosomy, cytoszkielet i ośrodek komórkowy.
Pytanie 5. Opisz organelle cytoplazmy i ich znaczenie w życiu komórki.
Retikulum endoplazmatyczne (ER) to zbiór wakuoli, kanałów i kanalików. Tworzy pojedynczą sieć wewnątrz cytoplazmy, połączoną z błoną zewnętrzną. otoczka jądrowa. Rozróżnij gładki i szorstki EPS. Smooth ER bierze udział w syntezie lipidów i węglowodanów, a także neutralizuje substancje toksyczne. Rybosomy znajdują się na powierzchni błon szorstkiego ER.
Aparat Golgiego to jednobłonowe organelle, które są częścią pojedynczej sieci membranowej komórki i są stosem płaskich zbiorników. To tam odbywa się ostateczne sortowanie i pakowanie produktów przemiany materii w pęcherzyki błonowe (wakuole). Między innymi aparat Golgiego tworzy lizosomy i zapewnia egzocytozę.
Lizosomy to małe błoniaste pęcherzyki zawierające enzymy trawienne składniki odżywcze. Lizosomy łączą się z wakuolą endocytarną, tworząc wakuolę trawienną. Jeśli zawartość lizosomów zostanie uwolniona wewnątrz samej komórki, następuje jej autoliza (samotrawienie komórki).
Mitochondria są klasyfikowane jako organelle dwubłonowe. Ich zewnętrzna błona jest gładka, a wewnętrzna tworzy fałdy (cristae). Mitochondria to stacje energetyczne komórki, ich główną funkcją jest synteza ATP.
Plastydy to dwubłonowe organelle komórek roślinnych. Istnieją trzy rodzaje plastydów: chloroplasty, chromoplasty i leukoplasty. Bezbarwne leukoplasty magazynują skrobię; zielone chloroplasty przeprowadzają fotosyntezę; pomarańczowe, żółte i czerwone chromoplasty zapewniają kolor owoców i kwiatów (przyciągając zapylacze i rozsiewacze nasion). Uważa się za ustalone, że w odległej przeszłości mitochondria i plastydy wywodziły się od prokariontów, „połykanych” przez komórkę eukariotyczną i wchodzących z nią w symbiozę. Mitochondria i plastydy mają koliste DNA, samodzielnie syntetyzują niektóre białka, a ich rybosomy są mniejsze niż u eukariotów.
Rybosomy to małe liczne niebłonowe organelle utworzone przez dwie podjednostki - dużą i małą. Podjednostki składają się z białka i rybosomalnego RNA. Funkcją rybosomów jest synteza białek. Niektóre rybosomy znajdują się bezpośrednio w cytoplazmie, a niektóre na błonach szorstkiego ER.
Centrum komórkowe - organoid o niebłonowej strukturze komórek zwierzęcych, grzybów i niższe rośliny. Składa się z dwóch centrioli, podobnych kształtem do cylindrów i składających się z najmniejszych rurek białkowych; uczestniczy w tworzeniu wrzeciona rozszczepienia.
Wakuola to błoniasty pęcherzyk wypełniony sokiem komórkowym. Musi być obecny w komórce roślinnej. Funkcją wakuoli jest gromadzenie wody, soli i składników odżywczych. Może również zawierać pigmenty (niebieskie, fioletowe) oraz gromadzić produkty przemiany materii.
Cytoszkielet jest organellą niebłonową, która jest przewodem białkowym znajdującym się zarówno w pobliżu błony, jak iw cytoplazmie. Ich funkcją jest utrzymanie kształtu komórki, zapewnienie wewnątrzkomórkowego transportu substancji, a także aktywnego ruchu komórki (ameba, fagocyt). Pęcherzyki błony zawierające dowolne substancje mogą poruszać się wzdłuż rurek cytoszkieletu, jak po szynach.
Jak pobrać darmowy esej? . I link do tego eseju; komórka eukariotyczna. Cytoplazma. Organelle już w Twoich zakładkach.Dodatkowe eseje na ten temat
Pytanie 1. Gdzie powstaje lizosom? Lizosomy to struktury błonowe zawierające wiele aktywnych enzymów biorących udział w rozkładzie związków wielkocząsteczkowych: białek, lipidów, węglowodanów. Lizosomy powstają w kompleksie Golgiego, do którego wchodzą i gdzie gromadzą się enzymy. Pytanie 2. Jaka jest funkcja mitochondriów? mitochondria - struktury komórkowe pokryty podwójną membraną. Na wewnętrznej błonie znajduje się wiele wypustek świetna ilość enzymy biorące udział w syntezie ATP. Stąd, główna funkcja mitochondria - dostarczanie komórkom energii poprzez syntezę ATP. Pytanie 3. Co
Pytanie 1. Jakie oznaki prymitywności prokariontów w porównaniu z eukariontami możesz wymienić? Prokarionty nie mają jądra i chromosomów. Dziedziczna informacja prokariontów jest reprezentowana przez pojedynczą cząsteczkę DNA, która jest zamknięta w formie pierścienia i znajduje się w cytoplazmie. Rybosomy u prokariotów są bardzo małe. Organelle L otoczone błonami (retikulum endoplazmatyczne, aparat Golgiego, mitochondria, plastydy), prokarioty nie mają w ogóle. W komórce eukariotycznej struktury te są przeznaczone do syntezy różnych związków. Ze względu na ich brak u prokariotów reakcja
Pytanie 1. Z czego zbudowane są ściany retikulum endoplazmatycznego i kompleksu Golgiego? Ściany retikulum endoplazmatycznego i kompleksu Golgiego są utworzone przez jednowarstwową membranę. Pytanie 2. Wymień funkcje retikulum endoplazmatycznego. Retikulum endoplazmatyczne (ER) tworzy system transportowy komórki. Na gładkim ER przeprowadzana jest synteza tłuszczów i węglowodanów. Na szorstkim (ziarnistym) ER białka są syntetyzowane dzięki pracy rybosomów przyczepionych do błon ER. Pytanie 3. Jaka jest funkcja rybosomów? Główną funkcją rybosomów jest synteza białek. Pytanie 4. Dlaczego większość rybosomów znajduje się na kanałach endoplazmatycznych
Test na temat „Porosty. Glony 1. Fotosynteza w algach zachodzi w: a) jądrze c) cytoplazmie b) chromatoforach d) chloroplastach 2. Porosty jako pojedynczy organizm sposób rozmnażania jest nieodłączny: a) płciowy c) partenogeneza b) bezpłciowy d) brak poprawnej odpowiedzi 3. Porosty nazywane są wskaźnikami czystości powietrza, ponieważ: a) oczyszczają powietrze b) zanieczyszczają powietrze c) tworzą siedlisko dla inne rośliny i zwierzęta d) rosną tylko w miejscach z czystym powietrzem 4. Czy są do tego zdolne aktywny ruch wodorost? a) wszystkie glony są w stanie się poruszać b) poruszać się
Pytanie 1. Jakie są funkcje zewnętrzna męmbrana komórki? Zewnętrzna błona komórkowa składa się z podwójnej warstwy lipidowej i cząsteczek białka, z których część znajduje się na powierzchni, a część przenika obie warstwy lipidów na wskroś. Zewnętrzna błona komórkowa pełni funkcję ochronną, oddzielając komórkę od środowiska zewnętrznego oraz zapobiega uszkodzeniu jej zawartości. Ponadto zewnętrzna błona komórkowa zapewnia transport substancji do i z komórki, umożliwia komórkom wzajemne oddziaływanie. Pytanie 2. W jaki sposób różne substancje mogą dostać się do komórki? Przez zewnętrzną
Pytanie 1. Jakie są główne etapy powstawania i rozwoju życia na Ziemi? Istnieje wiele hipotez próbujących wyjaśnić pochodzenie i rozwój życia na naszej planecie. I choć proponują różne podejścia do rozwiązania tego problemu, większość z nich zakłada istnienie trzech etapów ewolucyjnych: ewolucji chemicznej, prebiologicznej i biologicznej. Na etapie ewolucji chemicznej miała miejsce abiogeniczna synteza polimerów organicznych. W drugim etapie powstały kompleksy białko-kwas nukleinowy-lipid (naukowcy nazywali je inaczej: koacerwaty, hipercykle, probiony, progenoty i
Pytanie 1. Jakie są funkcje centrum komórkowego? Centrum komórkowe pełni funkcję tworzenia wewnętrznego szkieletu komórki (cytoszkieletu). Cytoszkielet to sieć mikrotubul penetrujących cytoplazmę, utrzymujących kształt komórki, zapewniających ruch organelli komórkowych, a także pracę wyspecjalizowanych organelli ruchowych - rzęsek i wici. Centrum komórkowe zapewnia również prawidłowy podział komórek. Centriole centrum komórki rozchodzą się do biegunów dzielącej się komórki i tworzą wrzeciono podziału, dzięki czemu z jednej komórki macierzystej powstają następnie dwie komórki potomne. Centriole składają się z cylindrów
Cytoplazma- obowiązkowa część komórki, zamknięta między błoną plazmatyczną a jądrem; Jest podzielony na hialoplazmę (główną substancję cytoplazmy), organelle (stałe składniki cytoplazmy) i inkluzje (tymczasowe składniki cytoplazmy). Skład chemiczny cytoplazmy: podstawą jest woda (60-90% całkowitej masy cytoplazmy), różne związki organiczne i nieorganiczne. Cytoplazma jest zasadowa. Cechą charakterystyczną cytoplazmy komórki eukariotycznej jest ciągły ruch ( cykloza). Jest wykrywany głównie przez ruch organelli komórkowych, takich jak chloroplasty. Jeśli ruch cytoplazmy ustanie, komórka umiera, ponieważ tylko będąc w w ciągłym ruchu, może spełniać swoje funkcje.
Hialoplazma ( cytozol) jest bezbarwnym, śluzowatym, gęstym i przezroczystym roztworem koloidalnym. To w nim zachodzą wszystkie procesy metaboliczne, zapewnia wzajemne połączenie jądra i wszystkich organelli. W zależności od przewagi części płynnej lub dużych cząsteczek w hialoplazmie wyróżnia się dwie formy hialoplazmy: sol- więcej płynnej hialoplazmy i żel- gęstsza hialoplazma. Możliwe są między nimi wzajemne przejścia: żel zamienia się w zol i odwrotnie.
Funkcje cytoplazmy:
- integracja wszystkich elementów ogniwa w jeden system,
- środowisko dla przebiegu wielu procesów biochemicznych i fizjologicznych,
- środowisko dla istnienia i funkcjonowania organelli.
Ściany komórkowe
Ściany komórkowe ograniczyć komórki eukariotyczne. W każdej błonie komórkowej można wyróżnić co najmniej dwie warstwy. Warstwa wewnętrzna przylega do cytoplazmy i jest reprezentowana przez błona plazmatyczna(synonimy - plazmalemma, błona komórkowa, błona cytoplazmatyczna), nad którymi tworzy się warstwa zewnętrzna. W klatka dla zwierząt jest cienki i nazywa się glikokaliks(tworzony przez glikoproteiny, glikolipidy, lipoproteiny), w komórce roślinnej – gruby, tzw Ściana komórkowa(utworzony przez celulozę).
Wszystko membrany biologiczne mają wspólne cechy konstrukcyjne i właściwości. Obecnie powszechnie akceptowane płynny model mozaiki struktury membrany. Podstawą błony jest dwuwarstwa lipidowa, zbudowana głównie z fosfolipidów. Fosfolipidy to triglicerydy, w których jedna reszta kwasu tłuszczowego jest zastąpiona resztą kwasu fosforowego; odcinek cząsteczki, w którym znajduje się reszta kwasu fosforowego, nazywany jest głową hydrofilową, odcinki, w których znajdują się reszty kwasu tłuszczowego, nazywane są ogonami hydrofobowymi. W błonie fosfolipidy są ułożone w ściśle uporządkowany sposób: hydrofobowe ogony cząsteczek skierowane są do siebie, a hydrofilowe głowy skierowane są na zewnątrz, w kierunku wody.
Oprócz lipidów błona zawiera białka (średnio ≈ 60%). Decydują o większości specyficznych funkcji błony (transport niektórych cząsteczek, kataliza reakcji, odbieranie i przetwarzanie sygnałów z otoczenia itp.). Rozróżnij: 1) białka obwodowe(znajduje się na zewnętrznej lub wewnętrznej powierzchni dwuwarstwy lipidowej), 2) białka półintegralne(zanurzone w dwuwarstwie lipidowej na różne głębokości), 3) białka integralne lub transbłonowe(przenikają błonę na wskroś, będąc w kontakcie zarówno z zewnętrznym, jak i wewnętrznym środowiskiem komórki). Białka integralne w niektórych przypadkach nazywane są kanałotwórczymi lub kanałowymi, ponieważ można je uznać za kanały hydrofilowe, przez które cząsteczki polarne przechodzą do komórki (lipidowy składnik błony nie przepuszcza ich).
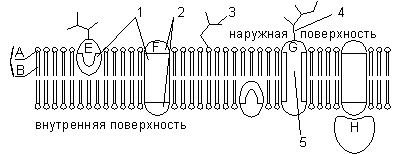
A - hydrofilowa głowa fosfolipidu; C, hydrofobowe ogony fosfolipidu; 1 - regiony hydrofobowe białek E i F; 2, hydrofilowe regiony białka F; 3 - rozgałęziony łańcuch oligosacharydowy przyłączony do lipidu w cząsteczce glikolipidu (glikolipidy występują rzadziej niż glikoproteiny); 4 - rozgałęziony łańcuch oligosacharydowy przyłączony do białka w cząsteczce glikoproteiny; 5 - kanał hydrofilowy (funkcjonuje jako por, przez który mogą przechodzić jony i niektóre cząsteczki polarne).
Błona może zawierać węglowodany (do 10%). Węglowodanowy składnik błon jest reprezentowany przez łańcuchy oligosacharydowe lub polisacharydowe związane z cząsteczkami białka (glikoproteiny) lub lipidami (glikolipidy). Zasadniczo węglowodany znajdują się na zewnętrznej powierzchni błony. Węglowodany zapewniają funkcje receptorowe błony. W komórkach zwierzęcych glikoproteiny tworzą kompleks epibłonowy, glikokaliks, o grubości kilkudziesięciu nanometrów. Znajduje się w nim wiele receptorów komórkowych, z jego pomocą dochodzi do adhezji komórek.
Cząsteczki białek, węglowodanów i lipidów są ruchome, zdolne do poruszania się w płaszczyźnie błony. Grubość błony plazmatycznej wynosi około 7,5 nm.
Funkcje membrany
Membrany pełnią następujące funkcje:
- oddzielenie zawartości komórkowej od środowiska zewnętrznego,
- regulacja metabolizmu między komórką a środowiskiem,
- podział komórki na przedziały („przedziały”),
- lokalizacja „przenośników enzymatycznych”,
- zapewnia komunikację między komórkami w tkankach Organizmy wielokomórkowe(przyczepność),
- rozpoznawanie sygnału.
Najważniejsze właściwość membrany- przepuszczalność selektywna, tj. membrany są wysoce przepuszczalne dla niektórych substancji lub cząsteczek i słabo przepuszczalne (lub całkowicie nieprzepuszczalne) dla innych. Ta właściwość leży u podstaw regulacyjnej funkcji błon, która zapewnia wymianę substancji między komórką a środowiskiem zewnętrznym. Proces, w którym substancje przechodzą przez błonę komórkową, nazywa się transport substancji. Rozróżnij: 1) transport pasywny- proces przekazywania substancji, przechodzenia bez energii; 2) transport aktywny- proces przekazywania substancji, idący kosztem energii.
Na transport pasywny substancje przemieszczają się z obszaru, w którym jest ich więcej wysokie stężenie do obszaru o niższym, tj. wzdłuż gradientu stężenia. W każdym roztworze znajdują się cząsteczki rozpuszczalnika i substancji rozpuszczonej. Proces ruchu cząsteczek substancji rozpuszczonej nazywa się dyfuzją, ruch cząsteczek rozpuszczalnika nazywa się osmozą. Jeśli cząsteczka jest naładowana, to na jej transport wpływa gradient elektryczny. Dlatego często mówi się o gradiencie elektrochemicznym, łącząc oba gradienty razem. Szybkość transportu zależy od wielkości nachylenia.
Można wyróżnić następujące typy transport bierny: 1) prosta dyfuzja- transport substancji bezpośrednio przez dwuwarstwę lipidową (tlen, dwutlenek węgla); 2) dyfuzja przez kanały membranowe- transport przez białka kanałotwórcze (Na+, K+, Ca 2+, Cl-); 3) ułatwiona dyfuzja- transport substancji za pomocą specjalnych białek transportowych, z których każde odpowiada za ruch określonych cząsteczek lub grup powiązanych cząsteczek (glukozy, aminokwasów, nukleotydów); 4) osmoza— transport cząsteczek wody (w sumie systemy biologiczne rozpuszczalnikiem jest woda).
Konieczność transport aktywny występuje, gdy konieczne jest zapewnienie przenoszenia cząsteczek przez membranę wbrew gradientowi elektrochemicznemu. Transport ten realizowany jest przez specjalne białka nośnikowe, których działanie wymaga wydatku energetycznego. Źródłem energii są cząsteczki ATP. Transport aktywny obejmuje: 1) pompę Na+/K+ (pompa sodowo-potasowa), 2) endocytozę, 3) egzocytozę.
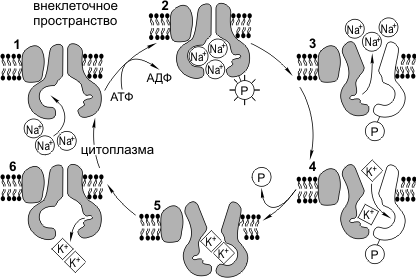
Pracuj z pompą Na + /K +. Dla normalne funkcjonowanie komórka musi utrzymywać określony stosunek jonów K + i Na + w cytoplazmie iw środowisku zewnętrznym. Stężenie K + wewnątrz komórki powinno być znacznie wyższe niż na zewnątrz, a Na + - odwrotnie. Należy zauważyć, że Na + i K + mogą swobodnie dyfundować przez pory membrany. Pompa Na+/K+ przeciwdziała wyrównywaniu się tych stężeń jonów i aktywnie pompuje Na+ z komórki, a K+ do komórki. Pompa Na+/K+ jest białkiem transbłonowym zdolnym do zmian konformacyjnych, dzięki czemu może przyłączać zarówno K+, jak i Na+. Cykl pracy pompy Na+/K+ można podzielić na następujące fazy: 1) dodanie Na+ z wewnątrz membrany, 2) fosforylacja białka pompy, 3) uwalnianie Na + w przestrzeni zewnątrzkomórkowej, 4) przyłączanie K + z poza błon komórkowych, 5) defosforylacja białka pompy, 6) uwalnianie K + w przestrzeni wewnątrzkomórkowej. Pompa sodowo-potasowa zużywa prawie jedną trzecią całej energii niezbędnej do życia komórki. Podczas jednego cyklu pracy pompa wypompowuje 3Na+ z ogniwa i pompuje 2K+.
Endocytoza- proces wchłaniania przez komórkę dużych cząstek i makrocząsteczek. Istnieją dwa rodzaje endocytozy: 1) fagocytoza- wychwytywanie i absorpcja dużych cząstek (komórek, części komórek, makrocząsteczek) oraz 2) pinocytoza- wychwytywanie i absorpcja materiału płynnego (roztwór, roztwór koloidalny, zawiesina). Zjawisko fagocytozy zostało odkryte przez I.I. Miecznikowa w 1882 r. Podczas endocytozy błona plazmatyczna tworzy zagłębienie, jej krawędzie łączą się, a struktury oddzielone od cytoplazmy pojedynczą błoną są wplecione w cytoplazmę. Wiele pierwotniaków i niektóre leukocyty są zdolne do fagocytozy. Pinocytozę obserwuje się w komórkach nabłonkowych jelita, w śródbłonku naczyń włosowatych.
Egzocytoza- proces odwrotny do endocytozy: wydalanie różne substancje z komórki. Podczas egzocytozy błona pęcherzyka łączy się z zewnętrzną błoną cytoplazmatyczną, zawartość pęcherzyka jest usuwana na zewnątrz komórki, a jego błona zostaje włączona do zewnętrznej błony cytoplazmatycznej. W ten sposób z komórek gruczołów wydzielina wewnętrzna hormony są wydzielane u pierwotniaków - niestrawione pozostałościżywność.
Iść do wykłady numer 5"Teoria komórki. Rodzaje organizacji komórkowej»
Iść do wykłady numer 7„Komórka eukariotyczna: budowa i funkcje organelli”
Treść abstrakcyjna
2.
Cytoplazma komórki eukariotycznej
4. Błony cytoplazmatyczne
4.1. Chemia i właściwości błony komórkowe
5. Membrana plazmowa
5.1. Wzrost błony plazmatycznej
6.1. Funkcje transportowe
6.2. Funkcje receptorowe błony plazmatycznej
6.3. Kontakty międzykomórkowe
6.4. Prosty kontakt. Połączenie typu blokady
6.6. kontakt pośredni
6.7. przerwa w kontakcie
Bibliografia
1. Rola naukowców w odkryciu błon komórkowych
Do końca XVIII wieku zakładano, że cele oddzielone są wspólną ścianą i nie można ich od siebie odizolować. W dalszym rozwoju tej nauki ważną rolę odegrały badania francuskiego botanika Brissota de Mirbel (1776-1854), w których po raz pierwszy wyrażono ideę badań porównawczych tkanek roślinnych. Mirbel jako pierwszy zasugerował, że sąsiadujące ze sobą komórki są oddzielone wspólną przegrodą – błoną.
Niemiecki przyrodnik Rudolphi doszedł do wniosku, że komórki są izolowane, że mają własne błony otaczające je ze wszystkich stron. Wątpliwości, czy komórki można izolować i izolować ostatecznie zniknęły po pracach botanika I. Moldengauera (1766-1827), z których wynikało, że komórka jest samodzielną strukturą pokrytą błoną. Powstał więc nowy pomysł na komórkę.
Najdobitniej sformułował to nieco później F. Meyen. Jego definicja jest następująca: „Komórka organizmu roślinnego to przestrzeń całkowicie otoczona błoną wegetatywną”. F. Meyen jasno sformułował ten pogląd na początku XIX wieku. który zastąpił pierwotny widok komórki jako czas. Ale specyficzna treść anatomii roślin nadal zasadniczo niewiele różniła się od starej. Naukowcy kontynuowali systematyczne opisywanie struktury różne ciała rośliny, a także uszczegółowić i wyjaśnić badane. Okres ten słusznie uważany jest za okres zbierania materiału, okres gromadzenia licznych informacji o najdoskonalszej budowie roślin.
Tak więc sam fakt obecności komórek w różne tkaniny nie budził wątpliwości. Ale czy całą różnorodność struktur organizmów roślinnych wyczerpują tylko komórki? Pytanie nadal nie było do końca jasne. Co więcej, większość naukowców odpowiedziała na to pytanie przecząco, wskazując obok komórek naczynia i włókna roślin jako dość osobliwe struktury. Podsumowując całą wiedzę w tej dziedzinie, Meyen bezpośrednio napisał, że tkanki roślinne składają się z komórek, włókien i naczyń krwionośnych. Pojawienie się tych ostatnich z komórek metamorfizujących ustalono ostatecznie dopiero nieco później - w latach 30. XX wieku. ostatni wiek.
2. Cytoplazma komórki eukariotycznej
Cytoplazma jest metabolicznym aparatem roboczym komórki. Koncentrują się w nim ogólne i specjalne organelle, główne procesy metaboliczne zachodzą w cytoplazmie. W ostatnich dziesięcioleciach szczególnie duży postęp dokonał się w dyskretnej analizie morfofunkcjonalnej poszczególnych organelli i układów błonowych aparatu metabolicznego cytoplazmy. Pozwala to wyobrazić sobie pracę aparatu metabolicznego jako integralnego systemu.
We współczesnych badaniach aparatu metabolicznego komórki szczególnie wyraźna jest tendencja do badania funkcjonalnego znaczenia struktur morfologicznych - do analizy funkcji poprzez strukturalną i biochemiczną organizację tych struktur. Najważniejsze sukcesy w tym zakresie osiągnięto w badaniu głównych organelli syntezy białek (rybosomów) oraz układów zapewniających metabolizm energetyczny (mitochondria, chloroplasty i błony koniugujące komórek prokariotycznych). Głównym uogólnieniem w badaniu aparatu metabolicznego cytoplazmy jest idea, że jest to trójfazowy układ zróżnicowany plastycznie, składający się z głównej cytoplazmy, błon wewnątrzkomórkowych i zawartości układu błonowego. Tak więc aparat metaboliczny cytoplazmy z jednej strony jest złożony z wyspecjalizowanych struktur pełniących określone funkcje, a z drugiej strony jest to pojedynczy integralny system, który jest w ścisłym związku zarówno z aparatem powierzchniowym, jak i jądrowym cytoplazmy. komórka.
Podejście to rozpoczęło się w latach 60. De Duve w doktrynie systemów błon egzoplazmatycznych. Następnie podejście to rozwinęło się zarówno w teorii lizosomów, jak i w współczesne idee o aparacie Golgiego i retikulum endoplazmatycznym, a także o dynamicznym oddziaływaniu układów błon wewnątrzkomórkowych. Naszym zdaniem bardzo obiecujący jest również pomysł, który ukształtował się ostatnio na temat głównej cytoplazmy - hialoplazmy. W przeciwieństwie do wcześniejszego poglądu na hialoplazmę jako jednorodny żywy układ koloidalny, interpretuje główną cytoplazmę jako zróżnicowaną, heterogeniczną fazę cytoplazmy zdolną do tworzenia złożonych struktur. Z tą właściwością hialoplazmy już częściowo zapoznaliśmy się na przykładzie układu podbłonowego aparatu powierzchniowego z jego dynamicznymi strukturami mechanochemicznymi i podtrzymującymi.
Obecnie staje się jasne, że takie otoczone błonami układy hialoplazmy powstają nie tylko na tym terenie membrany plazmatyczne, ale także w obszarze wszystkich błonowych, a czasem nawet niebłonowych (centrum komórkowe) struktur cytoplazmy. Proponuje się wyizolować tę część cytoplazmy do specjalnego podsystemu hialoplazmy, określając ją terminem „cytosol”. Naturalnie funkcje i organizacja morfobiochemiczna takiego cytozolu w obszarze retikulum endoplazmatycznego szorstkiego i gładkiego (ER), błon aparatu Golgiego, mitochondriów, centrum komórkowego, otoczki jądrowej itp. będą różne i specyficzne dla tych struktury, wokół których tworzą się odpowiednie sekcje cytozolu. .
Biorąc pod uwagę zróżnicowanie strukturalne i funkcjonalne organelli zapewniających metabolizm wewnątrzkomórkowy w cytoplazmie komórek, funkcja odcinków cytozolu odpowiadających każdemu konkretnemu organoidowi i praktycznie wchodzących w jego skład nie będzie ograniczona tylko do funkcji podporowo-kurczliwej . Struktury i enzymy tworzące cytozol są specyficzne dla każdej organelli i są bezpośrednio zaangażowane w zapewnienie procesów zachodzących w tym organoidzie. procesy biochemiczne.
Rozpatrzenie cytoplazmy warto rozpocząć od opisu problemu strukturalnej i biochemicznej organizacji rybosomów - molekularnych maszyn syntezy białek. W tym przypadku szczególnie wyraźnie widać znaczenie strukturalnej organizacji procesów biochemicznych w układach komórkowych i osiągnięto wielki sukces w jego badaniach.
Drugim obszarem biologii komórki, w którym szczegółowo zbadano również strukturalną organizację procesów biochemicznych, jest analiza tak zwanych błon koniugujących - aparatu metabolizmu energetycznego komórki. Rozważymy je przy charakteryzowaniu mitochondriów, plastydów i innych struktur cytoplazmy odpowiedzialnych za metabolizm energetyczny w komórce. Następnie omówimy wyniki dyskretnej analizy głównych membran.
Właściwe ciało komórki i jej zawartość są oddzielone od środowiska zewnętrznego lub od sąsiednich elementów w organizmach wielokomórkowych za pomocą błony plazmatycznej. Poza błoną plazmatyczną, zewnątrzkomórkową, zlokalizowaną Ściana komórkowa lub ściana, szczególnie dobrze eksprymowana w roślinach i organizmach prokariotycznych; w komórkach zwierzęcych jest nieobecny lub bardzo słabo wyrażany. Całe wnętrze komórki, z wyjątkiem jądra, nazywa się cytoplazmą. Jest to ogólny termin, który podkreśla podział komórki na dwa główne składniki: cytoplazmę i jądro.
Cytoplazma komórek eukariotycznych niejednorodny pod względem struktury i składu i obejmuje hialoplazmę, składniki błonowe i niebłonowe. Składniki błony obejmują układ wakuolowy (retikulum endoplazmatyczne, aparat Golgiego, lizosomy, wakuole roślinne), organelle błonowe (mitochondria i plastydy). Składniki niebłonowe obejmują centriole charakterystyczne dla komórek zwierzęcych oraz niebłonowe kompleksy i struktury makrocząsteczkowe, takie jak rybosomy, mikrotubule i mikrofilamenty. Taki podział cytoplazmy na odrębne składniki nie oznacza ich strukturalnej i funkcjonalnej izolacji od siebie.
3. Hialoplazma - wewnętrzne środowisko komórki
Terminy „hialoplazma” (od hialine – półprzezroczysty, przezroczysty), „plazma zasadowa” czy „macierz cytoplazmatyczna” oznaczają bardzo ważną część komórki, jej prawdziwe, wewnętrzne środowisko. W mikroskop elektronowy matryca cytoplazmatyczna ma wygląd jednorodnej lub drobnoziarnistej substancji o niskiej gęstości elektronowej. Z dawnych obserwacji właściwości fizykochemicznych cytoplazmy wywnioskowano, że hialoplazma jest złożonym układem koloidalnym, który obejmuje różne biopolimery: białka, kwasy nukleinowe, polisacharydy itp. Układ ten jest w stanie przenosić się z zolu (cieczy) Stan do żelu iz powrotem. Na przykład przy wysokich ciśnieniach hydrostatycznych cytoplazma nie skrapla się, lecz odwracalnie upłynnia. Zjawisko to tłumaczy się rozerwaniem wiązań między cząsteczkami w składzie hialoplazmy.
Dzięki metodom badań mikroskopii elektronowej i fizykochemicznej powstały idee dotyczące głównej plazmy jako zorganizowanego, uporządkowanego układu wieloskładnikowego. Oddzielne strefy hialoplazmy mogą zmieniać swój stan skupienia – w zależności od warunków lub zadania funkcjonalnego. Wiadomo zatem, że poszczególne cząsteczki białka tubuliny mogą być rozproszone w hialoplazmie, ale w pewnych momentach zaczynają się gromadzić i budować długie rurkowate struktury - mikrotubule.
Ten proces samoorganizacji mikrotubul jest odwracalny: gdy zmieniają się warunki życia komórki (wzrost ciśnienia lub zmiana przepuszczalności błon komórkowych), mikrotubule rozpadają się na monomeryczne cząsteczki tubuliny. W ten sam sposób w pozornie pozbawionej struktury hialoplazmie mogą powstawać i rozpadać się różne fibrylarne, nitkowate kompleksy cząsteczek białka. Osocze główne może brać udział w tworzeniu błon komórkowych, włókien i mikrofilamentów. W amebach zewnętrzne warstwy cytoplazmy (ektoplazmy) lub strefy cytoplazmy w obszarze pseudopodiów praktycznie składają się z jednej hialoplazmy, błony, mitochondria i inne struktury są tam rzadko spotykane. Jednak po ekspozycji na jony, które sprzyjają pęcznieniu komórek, dochodzi do szybkich i gwałtownych przegrupowań tych odcinków cytoplazmy - pojawiają się w nich błonowe małe wakuole. Dlatego główne osocze musi zawierać wszystkie elementy budulcowe (lipidy i białka), które są niezbędne do tworzenia błon.
Fibrylarne struktury cytoplazmatyczne mogą również powstać w wyniku specyficznych procesów agregacji w hialoplazmie. Jeśli duże jądra, struktury błon, a następnie rybosomy zostaną wytrącone z homogenatów komórkowych, to pozostały supernatant będzie zawierał główne składniki chemiczne hialoplazmy. Skład hialoplazmy mikrocząsteczek obejmuje głównie różne białka globularne i enzymy macierzy cytoplazmatycznej. Stanowią 20-25% treść ogólna białka w komórce eukariotycznej.
W komórki bakteryjne, ubogie w elementy błonowe, białka hialoplazmatyczne mogą stanowić około 50% wszystkich białek. Do najważniejszych enzymów macierzy należą enzymy glikolizy, enzymy metabolizmu cukrów, zasad azotowych, aminokwasów, lipidów i innych ważnych związków. Matryca zawiera enzymy do aktywacji aminokwasów podczas syntezy białek, transfer RNA. Właściwości osmotyczne i buforowe komórki są w dużej mierze zdeterminowane składem i strukturą hialoplazmy.
Najważniejszą rolą hialoplazmy jest to, że ta półpłynna pożywka łączy wszystkie struktury komórkowe i zapewnia ich wzajemne oddziaływanie chemiczne. Większość procesów transportu wewnątrzkomórkowego odbywa się przez hialoplazmę: przenoszenie aminokwasów, kwasów tłuszczowych, nukleotydów i cukrów. W hialoplazmie następuje stały przepływ jonów do iz błony plazmatycznej, do mitochondriów, jądra i wakuoli. Hialoplazma jest głównym zbiornikiem i strefą ruchu masy cząsteczek ATP. Następuje w nim odkładanie się produktów zapasowych: glikogenu, kropli tłuszczu. Strukturalnie i morfologicznie hialoplazma jest nadal słabo poznana.
4. Błony cytoplazmatyczne
Wspólną cechą wszystkich błon komórkowych, zewnętrznej błony plazmatycznej oraz wszystkich błon wewnątrzkomórkowych i organelli błonowych jest to, że są to cienkie (6-10 nm) warstwy o charakterze lipoproteinowym (lipidy w kompleksie z białkami), zamknięte w sobie. W komórce nie ma otwartych membran z wolnymi końcami. Błony komórkowe zawsze ograniczają wnęki lub obszary, zamykając je ze wszystkich stron i tym samym oddzielając zawartość takich wnęk od ich otoczenia. Tak więc błona plazmatyczna, pokrywająca całą powierzchnię komórki, która ma złożony kształt i liczne wyrostki, nie jest nigdzie przerwana, jest zamknięta. Oddziela zawartość cytoplazmy od środowiska otaczającego komórkę.
Wewnątrzkomórkowe zamknięte błony tworzą pęcherzyki - wakuole o kulistym lub spłaszczonym kształcie. W tym drugim przypadku powstają płaskie worki membranowe lub zbiorniki. Często wnęki odgraniczone membranami mają złożony kształt przypominający gąbkę lub siatkę, ale w tym przypadku takie wnęki są ograniczone membraną bez przerw. W takich wariantach błony oddzielają również dwie fazy strukturalne cytoplazmy: hialoplazmę od zawartości wakuoli i cystern. Błony mitochondriów i plastydów mają tę samą właściwość: oddzielają zawartość wewnętrzną od jam międzybłonowych i od hialoplazmy.
Błona jądrowa może być również reprezentowana jako perforowana, wydrążona, podwójna membrana o kulistym kształcie. Błony otoczki jądrowej ograniczają, oddzielają karioplazmę i chromosomy od siebie od jamy przestrzeni okołojądrowej i od hialoplazmy. Te ogólne właściwości morfologiczne błon komórkowych są określone przez ich skład chemiczny, ich lipoproteinowy charakter.
4.1. Chemia i właściwości błon komórkowych
Główny składniki chemiczne błony komórkowe to lipidy (40%) i białka (60%), ponadto w wielu błonach znajdują się węglowodany. Lipidy to duża grupa materia organiczna o słabej rozpuszczalności w wodzie (hydrofobowość) i rozpuszczalności w rozpuszczalnikach organicznych i tłuszczach (lipofilowość).
Skład lipidów w błonach komórkowych jest bardzo zróżnicowany. Charakterystycznymi przedstawicielami lipidów występujących w błonach komórkowych są fosfolipidy, sfingomieliny oraz spośród lipidów steroidowych cholesterol. Glicerolipidy to estry alkoholu trójwodorotlenowego, glicerolu z dwoma kwasami tłuszczowymi i kwasu fosforowego, które z kolei mogą być związane z różnymi grupami chemicznymi (cholina, seryna, inozytol, etanoloamina itp.).
Inną grupą lipidów błonowych są sfingomieliny, w których glicerol jest zastąpiony aminoalkoholem sfingozyną. Z lipidów związanych ze steroidami, przede wszystkim w błonach cholesterolu. W komórki roślinne cholesterolu nie znaleziono, zastępuje się go fitosterolami. Bakterie nie mają steroli. charakterystyczna cecha lipidów błonowych polega na podziale ich cząsteczek na dwie funkcjonalnie różne części: niepolarne ogony, które nie przenoszą ładunków i składają się z kwasów tłuszczowych, oraz naładowane głowy polarne. Głowy polarne przenoszą ładunki ujemne lub mogą być neutralne (jeśli mają zarówno ładunki dodatnie, jak i ujemne).
Stwierdzono, że błony komórkowe znacznie różnią się od siebie składem lipidowym. Tak więc błony plazmatyczne komórek zwierzęcych są bogate w cholesterol (do 30%), mają mało lecytyny, podczas gdy błony mitochondrialne, przeciwnie, są bogate w fosfolipidy i ubogie w cholesterol. Z całkowity zawartość lipidów lecytyny we frakcjach retikulum endoplazmatycznego wynosi 60-70% wszystkich fosfolipidów, podczas gdy w błonie komórkowej może wynosić 25-35%.
Ogólnie scharakteryzowano błonę plazmatyczną wysoka zawartość cholesterolu i sfingolipidów, a także przewagą nasyconych i jednonienasyconych kwasów tłuszczowych w składzie fosfolipidów, podczas gdy mitochondria, retikulum endoplazmatyczne i wiele innych błon cytoplazmatycznych zawiera mało cholesterolu i sfingolipidów, a stosunkowo dużo wielonienasyconych kwasów tłuszczowych. Najwyraźniej w związku z tym błony cytoplazmatyczne są mniej sztywne niż błona plazmatyczna, są bardziej topliwe.
Różna jest również ilość białek w błonach komórkowych. Tak więc w mitochondriach, w porównaniu z innymi błonami, jest najwięcej białek. Błony różnią się znacznie zestawem cząsteczek białka. Niektóre z nich są związane z głowami lipidów poprzez wiązania jonowe i dlatego są łatwo ekstrahowane z membran za pomocą roztworów soli. Inne tworzą wiązania solne z polarnymi regionami lipidów poprzez oddziaływanie z jonami Mg2+ lub Ca2+. Białka są również ekstrahowane przy użyciu związków chelatujących, takich jak wersen (EDTA). Większość białek oddziałuje z lipidami w składzie błon opartych na wiązaniach hydrofobowych.
Wiadomo, że wiele białek błonowych wydaje się składać z dwóch części: z regionów bogatych w aminokwasy polarne (przenoszące ładunek) i regionów wzbogaconych w aminokwasy niepolarne (glicyna, alanina, walion, leucyna). Takie białka w warstwach lipidowych błon są zlokalizowane w taki sposób, że ich niepolarne regiony są niejako zanurzone w „tłuszczowej” części błony, w której znajdują się hydrofobowe regiony lipidów. Polarna (hydrofilowa) część takich białek oddziałuje z główkami lipidowymi i jest skierowana w stronę fazy wodnej. Dlatego takie białka związane z lipidami przez oddziaływania hydrofobowe praktycznie nie są ekstrahowane w fazach wodnych. Można je wyizolować jedynie poprzez zniszczenie błony, ekstrakcję z niej lipidów rozpuszczalnikami organicznymi lub detergentami.
Przez rola biologiczna Białka błonowe można podzielić na trzy grupy: enzymy, białka receptorowe i białka strukturalne. Zestaw enzymów w błonach może być bardzo duży i różnorodny (na przykład w błonie plazmatycznej komórek wątroby znaleziono co najmniej 24 różne enzymy).
Różne membrany mają charakterystyczny zestaw enzymów. Innymi słowy, o chemicznej heterogeniczności błon decydują nie tylko lipidy, ale także białka. Wspólną cechą wszystkich błon komórkowych jest ich struktura lipoproteinowa; odmienność polega na ilościowych i jakościowych różnicach w lipidach i skład białek. Na przykład skład i ilość lipidów i białek będzie zupełnie inna w błonach retikulum endoplazmatycznego i wewnętrznej błonie mitochondriów.
Węglowodanowy składnik błon jest reprezentowany głównie przez glikoproteiny - cząsteczki białek kowalencyjnie (w przeciwieństwie do nukleoprotein i lipoprotein) związane z łańcuchami węglowodanowymi.
4.2. Budowa błon komórkowych
Pomysł, że błony komórkowe są oparte na podwójnej warstwie lipidowej, powstał w latach dwudziestych XX wieku. W 1925 roku opublikowano pracę, która wskazywała na tworzenie ciągłej warstwy bilipidowej błony plazmatycznej. Pomimo szeregu błędów ogólna idea istnienia warstwy bilipidowej okazała się poprawna. Okazało się, że jeśli lipidy zostaną wyekstrahowane z błony erytrocytów, a następnie umieszczone na powierzchni menisku wodnego, to można obliczyć powierzchnię zajmowaną przez powstałą monowarstwę lipidów. Okazało się, że powierzchnia ta jest dwukrotnie większa od powierzchni zajmowanej przez erytrocyty, z których ekstrahowano lipidy.
Naukowcy zasugerowali, że w błonach erytrocytów lipidy są ułożone w dwóch warstwach. Ponadto okazało się, że napięcie powierzchniowe komórki jest znacznie niższe niż napięcie powierzchniowe sztucznej warstwy lipidowej. Ponadto naukowcy odkryli, że po dodaniu białka do oleju napięcie powierzchniowe zmniejsza się do wartości charakterystycznej dla napięcia powierzchniowego komórek. Na tej podstawie stwierdzono, że błona zawiera białko.
Powstała więc hipoteza dotycząca budowy błony, według której jest to trójwarstwowa „kanapka” (białko-lipid-białko). Warstwa lipidowa w centrum jest warstwą dwucząsteczkową, w której hydrofobowe końce cząsteczek lipidowych są skierowane ku sobie, a hydrofilowe końce są skierowane ku warstwom białkowym leżącym po obu stronach warstwy lipidowej. Hipoteza ta została bezpośrednio potwierdzona, gdy stało się możliwe wykorzystanie mikroskopii elektronowej do badania błon komórkowych. Dzięki niej szczególnie dobrze widać było na błonach osłonki mielinowej nerwów, że błona ta jest naprawdę trójwarstwowa: dwie ciemne warstwy po 2,5 nm każda i szersza jasna warstwa pomiędzy nimi.
Trójwarstwową budowę błon komórkowych można pokazać w mikroskopie elektronowym metodą zamrażania-rozszczepiania, co jest szczególnie ważne, ponieważ w tym przypadku można zaobserwować określone enzymy, a nawet układy enzymatyczne. Na początku lat 60. Robertson sformułował hipotezę pojedynczej, jednolitej „elementarnej” błony biologicznej. Kwestia pochodzenia, powstawania błon w żywej komórce nie została jeszcze wystarczająco zbadana. Wiadomo, że liczba i wielkość elementów błonowych w komórkach może się zwiększać i zmniejszać, czyli można mówić o membranogenezie i niszczeniu błon.
5. Membrana plazmowa
Błona plazmatyczna, czyli plazmalemma, zajmuje szczególne miejsce wśród różnych błon komórkowych. Jest to powierzchowna struktura obwodowa, która ogranicza komórkę od zewnątrz, co warunkuje jej bezpośrednie połączenie ze środowiskiem pozakomórkowym, a co za tym idzie, ze wszystkimi substancjami i bodźcami działającymi na komórkę. Chemicznie błona plazmatyczna jest kompleksem lipoproteinowym.
Główne zasady organizacji błon zostały już omówione w poprzedniej sekcji. Ma grubość około 10 nm i jest najgrubszą z błon komórkowych. Głównymi składnikami błon plazmatycznych są lipidy (około 40%), białka (ponad 60%) i węglowodany (około 1%). Jak już wspomniano, błona plazmatyczna jest bogatsza w cholesterol w porównaniu z innymi błonami, aw jej fosfolipidach dominują nasycone kwasy tłuszczowe.
Skład białek tworzących błonę plazmatyczną jest bardzo zróżnicowany, o czym decyduje różnorodność jej obciążeń funkcjonalnych. Wśród enzymów stwierdzono 5 "- nukleotydazę, Mg-zależną ATPazę aktywowaną przez jony Na i K, fosfatazę alkaliczną i kwaśną, cyklazę adenylanową, RNazę, aktywność proteolityczną. Enzymy łańcucha oddechowego i enzymy glikolityczne są całkowicie nieobecne w błonie komórkowej komórek eukariotycznych, ale w komórkach prokariotycznych to w składzie błony plazmatycznej zlokalizowane są elementy łańcucha transportu elektronów i fosforylacji oksydacyjnej.
Należy podkreślić, że niektóre białka występujące we frakcjach błony komórkowej mogą nie być bezpośrednio związane z warstwą bilipidową. Wiele enzymów, które są aktywne we frakcjach błony komórkowej, można zlokalizować w glikokaliksie, kompleksie glikoproteinowym związanym z błoną komórkową. Skład glikokaliksu obejmuje różne węglowodany, które mogą stanowić około 1% suchej masy błon.
Węglowodany tworzą długie, rozgałęzione łańcuchy polisacharydów przyczepione do warstwy białek, która często określa kształt komórki. Woreczki błonowe mogą tu przylegać do błony plazmatycznej od wewnątrz; w tym przypadku w pobliżu powierzchni komórek znajdują się trzy warstwy błon: sama błona plazmatyczna i dwie błony pęcherzyków błonkowych.
W orzęskach buta błonka tworzy zgrubienia, ułożone w kształcie sześciokątów, w środku których wyłaniają się rzęski. Sztywność formacji błonkowych może być również związana z elementami cytoplazmy leżącej pod błoną plazmatyczną, z warstwą korową. Tak więc w grzebieniach błonki eugleny w pobliżu błony, oprócz wakuoli błonowych, znajdują się równoległe wiązki mikrotubul i mikrofilamentów. To włókniste wzmocnienie obwodowe wraz ze złożonym wielowarstwowym obrzeżem membrany tworzy sztywną strukturę błonki.
5.1. Wzrost błony plazmatycznej
Po podziale komórki następuje wzrost objętości rosnących komórek potomnych, aw związku z tym następuje wzrost powierzchni komórki, wzrost powierzchni błony plazmatycznej. Ale to nie jedyny przykład. Szybki wzrost objętość i powierzchnia. Powierzchnia szybko rosnących komórek we włóknach pręcików zbóż może wzrosnąć 65 razy w ciągu 1 godziny, tj. co minutę plazmalemma rośnie do swojej pierwotnej wartości. Tak wysokie tempo wzrostu błony plazmatycznej można wytłumaczyć jedynie szybkim wbudowywaniem gotowych prekursorów błony, lipidów i białek, w starą warstwę lipoproteinową. Podobny sposób wzrost jest całkiem możliwy, ponieważ wiadomo, że w komórce zachodzi ciągła rearanżacja błon - wymiana niektórych lipidów na inne, zmiana ich ilości, zmiana składu białek itp. Dlatego błony są bardzo nietrwałe związki.
Opisano jednak również inny przypadek powstawania nowej błony plazmatycznej. Plasmodium Myxomycete Physarum polycephalum przekłuto szklaną kapilarą, a otrzymany cylinder plazmowy, pozbawiony błony plazmatycznej, utrwalono natychmiast, a następnie po 1, 2, 3 i 6 s od rozpoczęcia eksperymentu badano pod mikroskopem elektronowym. Cytoplazma tego grzyba charakteryzuje się obecnością dużej liczby małych pęcherzyków zawierających śluz, który pokrywa całe plazmodium od zewnątrz. Można było zauważyć, że takie pęcherzyki, rozproszone w głównej plazmie, zbierały się na niezabezpieczonej krawędzi plazmy, tworząc spłaszczone wakuole, położone równolegle do zewnętrznej powierzchni naświetlanego fragmentu. Takie płaskie wakuole połączyły się i utworzyły ciągłą membranę - nową błonę plazmatyczną, która izoluje składniki plazmy od środowiska zewnętrznego.
Najwyraźniej taki proces odnowy błony plazmatycznej zachodzi nie tylko wtedy, gdy jest ona poważnie uszkodzona, ale także stale, podczas życia komórki. Tak więc, w wyniku egzocytozy, błony wakuolowe, łącząc się z błoną plazmatyczną, wydają się być w niej osadzone, zwiększając w ten sposób całkowitą powierzchnię komórki. Obserwacje procesu sekrecji (jednej z form egzocytozy) doprowadziły wielu badaczy do przekonania, że w komórce zachodzą stałe przepływy elementów błonowych: z błony plazmatycznej do cytoplazmy (endocytoza) i odwrotnie, przepływ struktur błonowych z cytoplazmy na powierzchnię komórki (egzocytoza). W tym cyklu błon wiodącą rolę odgrywa układ wakuoli błonowych aparatu Golgiego.
6. Funkcje błony plazmatycznej
Błona plazmatyczna tak cała linia najważniejsze funkcje komórkowe, z których wiodącą jest funkcja oddzielania substancji cytoplazmy od środowiska zewnętrznego oraz funkcja transportu różnych substancji zarówno do wnętrza komórki, jak i poza nią. Funkcje transportowe obejmują transport bierny wody, jonów, substancji niskocząsteczkowych i transport aktywny tych substancji wbrew gradientom stężeń, a także różne formy transportu związków i kompleksów wielkocząsteczkowych (endocytoza).
Błona plazmatyczna dodatkowo bierze udział w usuwaniu z komórek powstałych w niej produktów. Uczestniczy również w procesach zewnątrzkomórkowego rozszczepienia biopolimerów. Na powierzchni błony plazmatycznej znajdują się różne struktury receptorowe, które specyficznie oddziałują z czynnikami zewnątrzkomórkowymi i sąsiednimi komórkami. Zatem błona komórkowa bierze udział w przekazywaniu sygnałów do komórki. Błona plazmatyczna bierze udział w interakcjach międzykomórkowych w organizmach wielokomórkowych. Oddzielne sekcje błony plazmatycznej w wyspecjalizowanych komórkach zwierzęcych biorą udział w budowie specjalnych procesów komórkowych, takich jak mikrokosmki, rzęski, wyrostki receptorowe itp. Wreszcie błona plazmatyczna odgrywa ważną rolę w podziale komórki.
To krótkie wyliczenie funkcji błony plazmatycznej jest dalekie od zakończenia, ale pokazuje ogromne znaczenie tej peryferyjnej struktury każdej komórki, czy to komórki bakterii, czy organizmów eukariotycznych.
6.1. Funkcje transportowe
Błona plazmatyczna, podobnie jak inne komórkowe błony lipoproteinowe, jest półprzepuszczalna. Oznacza to, że różne cząsteczki przechodzą przez nią z różną prędkością, a im większy rozmiar cząsteczek, tym mniejsza prędkość ich przejścia przez membranę. Ta właściwość definiuje błonę plazmatyczną jako barierę osmotyczną.
Woda i rozpuszczone w niej gazy mają maksymalną zdolność penetracji, jony penetrują membranę znacznie wolniej (około 104 razy wolniej). Dlatego jeśli komórka, na przykład erytrocyt, zostanie umieszczona w środowisku, w którym stężenie soli jest niższe niż w komórce (niedociśnienie), to woda z zewnątrz napłynie do komórki, co doprowadzi do wzrostu objętości i do pęknięcia błony plazmatycznej (hipotoniczny „wstrząs”).
Wręcz przeciwnie, gdy erytrocyt zostanie umieszczony w roztworach soli o wyższym stężeniu niż w komórce, woda wypłynie z niego do otoczenie zewnętrzne. W tym samym czasie komórka będzie się marszczyć, zmniejszając objętość. Taki bierny transport wody z komórki do wnętrza komórki nadal przebiega z małą szybkością. Dzięki temu w błonie komórkowej, w jej warstwie lipoproteinowej, znajdują się specjalne pory, przez które wnika woda i jony.
6.2. Funkcje receptorowe błony plazmatycznej
Funkcje te są związane z lokalizacją na błonie plazmatycznej specjalnych struktur mających na celu specyficzne rozpoznanie czynników chemicznych lub fizycznych. Na powierzchni komórki znajduje się duży zestaw składników - receptorów, które warunkują możliwość specyficznych reakcji z różnymi czynnikami. Białka błonowe lub elementy glikokaliksu (polisacharydy, glikoproteiny) mogą pełnić rolę takich receptorów na powierzchni komórki.
Uważa się, że takie miejsca wrażliwe na poszczególne substancje mogą być rozproszone na powierzchni komórki lub gromadzone w małych strefach. Na przykład na powierzchni komórek bakteryjnych lub zwierzęcych istnieje ograniczona liczba miejsc, z którymi cząsteczki wirusa mogą się wiązać. Co ciekawe, różne wirusy zwykle wiążą się z różnymi częściami obwodu komórki, a ta sama komórka bakteryjna może mieć ich kilka różne rodzaje receptory.
Zastosowanie różnych metod immunologicznych wykazało, że antygenowo zlokalizowane na powierzchni komórki Składniki aktywne które specyficznie reagują z przeciwciałami lub komórki odpornościowe. Większość z tych antygenów powierzchniowych zawiera grupy węglowodanowe (glikolipidy, glikoproteiny). Antygenowymi składnikami ściany bakteryjnej są również najczęściej lipopolisacharydy i glikolipidy. Na powierzchni komórki, często bezpośrednio w błonie lipoproteinowej, znajdują się różne receptory, które wiążą się fizjologicznie substancje czynne jak różne hormony, neuroprzekaźniki itp.
różne komórki organizmy zwierzęce mogą mieć różne zestawy receptorów lub różną czułość tego samego receptora. Rolą wielu receptorów komórkowych jest przekazywanie sygnałów z powierzchni do wnętrza komórki. Obecnie dobrze zbadano system przekazywania bodźców do komórek za pomocą niektórych hormonów, do których należą łańcuchy peptydowe. Stwierdzono, że hormony te wiążą się ze specyficznymi receptorami na powierzchni błony plazmatycznej komórki.
Różnorodność i specyficzność zestawów receptorów na powierzchni komórek prowadzi do powstania bardzo złożonego systemu markerów, które umożliwiają odróżnienie własnych komórek (tego samego osobnika lub tego samego gatunku) od komórek innych. Podobne komórki wchodzą ze sobą w interakcje, prowadząc do adhezji powierzchni (koniugacja u pierwotniaków i bakterii, tworzenie kompleksów tkankowo-komórkowych). W tym przypadku komórki, które różnią się zestawem markerów determinant lub ich nie dostrzegają, są albo wykluczane z takiej interakcji, albo ulegają zniszczeniu u zwierząt wyższych w wyniku reakcji immunologicznych.
Błona plazmatyczna jest związana z lokalizacją specyficznych receptorów, na które reagują czynniki fizyczne. Tak więc w błonie plazmatycznej lub w jej pochodnych w bakteriach fotosyntetyzujących i niebiesko-zielonych algach zlokalizowane są białka receptorowe (chlorofile) oddziałujące z kwantami światła. W błonie plazmatycznej światłoczułych komórek zwierzęcych znajduje się specjalny system białek fotoreceptorowych (rodopsyna), za pomocą których sygnał świetlny jest przekształcany w sygnał chemiczny, co z kolei prowadzi do wytworzenia energii elektrycznej. impuls.
6.3. Kontakty międzykomórkowe
Błona plazmatyczna, jak już wspomniano, jest aktywnie zaangażowana w kontakty międzykomórkowe związane z koniugacją. Jednokomórkowe organizmy. W organizmach wielokomórkowych w wyniku interakcji międzykomórkowych powstają złożone zespoły komórkowe, których utrzymanie można przeprowadzić różne sposoby. W tkankach zarodkowych, embrionalnych, zwłaszcza na wczesne stadia rozwoju, komórki pozostają ze sobą połączone ze względu na zdolność ich powierzchni do sklejania się. Ta właściwość łączenia komórek może być określona przez właściwości ich powierzchni, które specyficznie oddziałują na siebie.
Mechanizm tych połączeń jest wciąż niedostatecznie zbadany, ale najprawdopodobniej zapewnia go interakcja między lipoproteinami a glikokaliksem błon plazmatycznych. Przy takim międzykomórkowym oddziaływaniu komórek embrionalnych między błonami plazmatycznymi zawsze pozostaje szczelina o szerokości około 20 nm, wypełniona glikokaliksem. Traktowanie tkanki enzymami naruszającymi integralność glikokaliksu (mukazy działające hydrolitycznie na mucyny, mukopolisacharydy) lub uszkadzającymi błonę komórkową (proteazy) prowadzi do izolacji komórek od siebie, do ich dysocjacji. Jednakże, jeśli czynnik dysocjacji zostanie usunięty, komórki mogą ponownie złożyć się i ponownie agregować.
Tak więc możliwe jest oddzielenie komórek gąbek o różnych kolorach - pomarańczowym i żółtym. Okazało się, że w mieszaninie tych komórek tworzą się dwa rodzaje agregatów: składające się tylko z komórek żółtych i tylko z komórek pomarańczowych. W tym przypadku mieszane zawiesiny komórek samoorganizują się, przywracając pierwotną strukturę wielokomórkową. Podobne wyniki uzyskano z rozdzielonymi zawiesinami komórkowymi zarodków płazów; w tym przypadku następuje selektywne przestrzenne oddzielenie komórek ektodermy od endodermy i od mezenchymu.
Ponadto, jeśli tkanki są używane do ponownej agregacji późne etapy rozwoju zarodków, następnie różne zespoły komórek o swoistości tkankowej i narządowej są niezależnie składane w probówce, powstają agregaty nabłonkowe podobne do kanalików nerkowych itp. Połączenia między komórkami w tkankach i narządach wielokomórkowych organizmów zwierzęcych mogą być tworzone przez złożone specjalne struktury, które wywołały właściwe kontakty międzykomórkowe. Te ustrukturyzowane kontakty międzykomórkowe są szczególnie wyraźne w tkankach granicznych powłok, w nabłonku.
Możliwe, że pierwotna izolacja warstwy komórek połączonych ze sobą za pomocą specjalnie ustrukturyzowanych kontaktów międzykomórkowych zapewniła powstawanie i rozwój tkanek i narządów w filogenezie zwierząt. Dzięki mikroskopii elektronowej zgromadzono wiele danych na temat ultrastruktury tych formacji łączących. Niestety ich skład biochemiczny i struktura molekularna nie są jeszcze dobrze poznane.
Badając połączenia komórek w warstwach nabłonka, można znaleźć następujące struktury, które łączą ze sobą komórki: kontakt prosty, połączenie typu „lock”, kontakt ścisły, kontakt pośredni lub strefa adhezji, kontakt desmosomalny , styk przypominający szczelinę.
6.4. Prosty kontakt. połączenie typu zamek
Prosty kontakt występuje wśród większości przylegających do siebie komórek różnego pochodzenia. Większość powierzchni stykających się komórek nabłonkowych jest również połączona prostym stykiem, w którym błony plazmatyczne stykających się komórek są oddzielone przestrzenią 15-20 nm. Jak już wspomniano, przestrzeń ta reprezentuje nadbłonowe składniki powierzchni komórek. Szerokość szczeliny między błonami komórkowymi może przekraczać 20 nm, tworząc wypustki, zagłębienia, ale nie mniej niż 10 nm. Od strony cytoplazmy do tej strefy błony plazmatycznej nie przylegają żadne specjalne dodatkowe struktury.
Połączenie typu „zamek” to występ błony plazmatycznej jednej komórki do wgłobienia (wypukłości) innej. Na rozcięciu ten rodzaj łączenia przypomina szew stolarski. Przestrzeń międzybłonowa i cytoplazma w strefie „zamków” mają takie same cechy jak w obszarach kontaktu prostego.
6.5. Szczelny styk zamykający
Jest to strefa, w której zewnętrzne warstwy dwóch błon plazmatycznych znajdują się tak blisko, jak to możliwe. W tym kontakcie często obserwuje się trójwarstwową membranę: dwie zewnętrzne warstwy osmofilne obu membran łączą się we wspólną warstwę o grubości 2–3 nm. Fuzja membranowa nie występuje na całym obszarze ciasnego kontaktu, ale jest szeregiem połączeń punktowych. Od strony cytoplazmy w tej strefie często spotyka się liczne fibryle o średnicy około 8 nm, położone równolegle do powierzchni plazmalemmy.
Kontakty tego typu stwierdzono między fibroblastami w hodowli tkankowej, między nabłonkiem embrionalnym a komórkami mezenchymalnymi.
6.6. Kontakt pośredni (lub strefa klejenia)
W tym miejscu odległość międzybłonowa jest nieco rozszerzona (do 25-30 nm) iw przeciwieństwie do prostego kontaktu jest wypełniona gęstą zawartością, najprawdopodobniej o charakterze białkowym.
6.7. przerwa w kontakcie
Jest to obszar o długości 0,5-3 μm, w którym błony plazmatyczne są oddzielone szczeliną 2-3 nm, co po osmacji nadaje całej strukturze siedmiowarstwowy wygląd. Od strony cytoplazmy nie znaleziono żadnych specjalnych struktur błonowych. Ten typ połączenia występuje we wszystkich rodzajach tkanek. Wydaje się, że funkcjonalną rolą złącza szczelinowego jest przenoszenie jonów i cząsteczek z komórki do komórki. Na przykład w mięśniu sercowym przenoszenie potencjału czynnościowego z komórki do komórki odbywa się poprzez rodzaj kontaktu, w którym jony mogą swobodnie przechodzić przez połączenia międzykomórkowe. Utrzymanie tego wiązania jonowego między komórkami zależy od energii uzyskanej w wyniku fosforylacji oksydacyjnej.
Bibliografia
1. Vermel E. M. Historia doktryny komórki. M.: Nauka, 1970. 259 s.
2. Gulyaev G. V., Malchenko V. V. Słownik terminów z zakresu genetyki, cytologii, hodowli, nasiennictwa i nasiennictwa. M.: Rosselchozizdat, 1983. 240 s.
3. De Duve K. Podróż do świata żywej komórki. M.: Mir, 1987. 253 s.
4. Zavarzin A. A., Kharazova A. D. Podstawy cytologii ogólnej: Podręcznik. L.: Wydawnictwo Leningrad. un-ta, 1982. 240 s.
5. Od molekuł do człowieka. / Under. wyd. Naumova NPM: Edukacja, 1973. 480 s.
6. Pausheva Z. P. Warsztaty z cytologii roślin. M.: Agropromizdat, 1988. 271 s.
7. Troshin A.S., Braun A.D., Vakhtin Yu.B., Zhilkin L.N., Sukhanova KM Cytology. Moskwa: Edukacja, 1970. 304 s.
8. Chentsov Yu. S. Cytologia ogólna. M.: MGU, 1978. 344 s.
Abstrakt na plusie
plazmalemma(ściana komórkowa) komórek zwierzęcych jest utworzona przez błonę pokrytą na zewnątrz warstwą glikokaliksu o grubości 10-20 nm. plazmalemma pełni funkcje delimitujące, barierowe, transportowe i receptorowe. Dzięki właściwości selektywnej przepuszczalności plazmalemia reguluje skład chemiczny środowiska wewnętrznego komórki. Plazmalemma zawiera cząsteczki receptorowe, które selektywnie rozpoznają pewne substancje biologicznie czynne (hormony). W warstwach i warstwach sąsiednie komórki są zatrzymywane z powodu obecności różnego rodzaju kontakty, które są reprezentowane przez sekcje plazmalemmy, które mają specjalną strukturę. Od wewnątrz warstwa korowa (korowa) przylega do błony cytoplazma Grubość 0,1-0,5 µm.
Cytoplazma. W cytoplazmie występuje szereg sformalizowanych struktur, które mają regularne cechy budowy i zachowania w różnych okresach życia komórki. Każda z tych struktur ma określoną funkcję. Stąd wzięło się ich porównanie z organami całego organizmu, w związku z czym otrzymały nazwę organelle, Lub organelle. W cytoplazmie osadzają się różne substancje - inkluzje (glikogen, krople tłuszczu, barwniki). Cytoplazma jest przesiąknięta błonami retikulum endoplazmatyczne.
Retikulum endoplazmatyczne (EMF). Retikulum endoplazmatyczne to rozgałęziona sieć kanałów i jam w cytoplazmie komórki, utworzona przez błony. Na błonach kanałów znajdują się liczne enzymy, które zapewniają żywotną aktywność komórki. Istnieją 2 rodzaje membrany EMF- gładkie i szorstkie. Na membranach retikulum endoplazmatyczne gładkie istnieją układy enzymatyczne zaangażowane w tłuszcz i metabolizm węglowodanów. główna funkcja szorstka siateczka śródplazmatyczna- synteza białek, która odbywa się w rybosomach przyczepionych do błon. Retikulum endoplazmatyczne- jest to wspólny wewnątrzkomórkowy układ krążenia, przez którego kanały substancje są transportowane wewnątrz komórki iz komórki do komórki.
Rybosomy pełnić funkcję syntezy białek. Rybosomy to kuliste cząstki o średnicy 15-35 nm, składające się z 2 podjednostek o nierównej wielkości i zawierające w przybliżeniu równą ilość białek mRNA. Rybosomy w cytoplazmie są zlokalizowane lub przyczepione do zewnętrznej powierzchni błon retikulum endoplazmatycznego. W zależności od rodzaju syntetyzowanego białka rybosomy można łączyć w kompleksy - polirybosomy. Rybosomy są obecne we wszystkich typach komórek.
kompleks Golgiego. Główny element konstrukcyjny kompleks Golgiego jest gładką membraną, która tworzy paczki spłaszczonych cystern lub dużych wakuoli lub małych pęcherzyków. Cysterny kompleksu Golgiego są połączone z kanałami retikulum endoplazmatycznego. Białka, polisacharydy, tłuszcze syntetyzowane na błonach retikulum endoplazmatycznego są transportowane do kompleksu, kondensowane wewnątrz jego struktur i „pakowane” w postaci gotowego do uwolnienia sekretu lub wykorzystywane w samej komórce podczas jej życia.
mitochondria. Ogólna dystrybucja mitochondriów u zwierząt i flora zwrócić uwagę na ważną rolę, jaką pełni mitochondria bawić się w klatce. mitochondria mają postać kulistych, owalnych i cylindrycznych ciał, mogą być nitkowate. Wielkość mitochondriów to 0,2-1 µm średnicy i do 5-7 µm długości. Długość nitkowatych form sięga 15-20 mikronów. Liczba mitochondriów w komórkach różnych tkanek nie jest taka sama, jest ich więcej tam, gdzie procesy syntezy są intensywne (wątroba) lub koszty energii są wysokie. Ściana mitochondriów składa się z 2 błon - zewnętrznej i wewnętrznej. Zewnętrzna błona jest gładka, a przegrody - grzbiety lub cristae odchodzą od wewnętrznego wnętrza organoidu. Na błonach cristae znajdują się liczne enzymy biorące udział w metabolizmie energetycznym. Główna funkcja mitochondriów - synteza ATP.
Lizosomy- małe owalne ciałka o średnicy około 0,4 mikrona, otoczone jedną trójwarstwową membraną. Lizosomy zawierają około 30 enzymów zdolnych do rozkładania białek, kwasów nukleinowych, polisacharydów, lipidów i innych substancji. Rozkład substancji przez enzymy to tzw Liza, stąd nazwa organoidu lizosom. Uważa się, że lizosomy powstają ze struktur kompleksu Golgiego lub bezpośrednio z retikulum endoplazmatycznego. Funkcje lizosomów : wewnątrzkomórkowe trawienie składników odżywczych, niszczenie struktury samej komórki podczas jej śmierci podczas rozwoju embrionalnego, kiedy tkanki embrionalne są zastępowane tkankami trwałymi iw wielu innych przypadkach.
Centriole. Centrum komórki składa się z 2 bardzo małych cylindrycznych ciał umieszczonych względem siebie pod kątem prostym. Te ciała to tzw centriole. Ściana centrioli składa się z 9 par mikrotubul. Centriole są zdolne do samoorganizacji i są samoreprodukującymi się organellami cytoplazmy. Centriole odgrywają ważną rolę w podziale komórki: rozpoczynają wzrost mikrotubul, które tworzą wrzeciono podziału.
Rdzeń. Najważniejszy jest rdzeń część komórki. Zawiera cząsteczki DNA i dlatego pełni dwie główne funkcje: 1) przechowywanie i odtwarzanie informacji genetycznej, 2) regulację procesów metabolicznych zachodzących w komórce. Komórka, która przegrała rdzeń, nie może istnieć. Jądro jest również niezdolne do niezależnego istnienia. Większość komórek ma jedno jądro, ale w jednej komórce, na przykład w komórkach wątroby, można zaobserwować 2-3 jądra. Znane komórki wielojądrzaste o liczbie jąder rzędu kilkudziesięciu. Kształt jąder zależy od kształtu komórki. Jądra są kuliste, wielopłatkowe. Jądro jest otoczone membraną składającą się z dwóch membran o zwykłej strukturze trójwarstwowej. Zewnętrzna błona jądrowa pokryta jest rybosomami, wewnętrzna błona jest gładka. Główną rolę w życiowej aktywności jądra odgrywa metabolizm między jądrem a cytoplazmą. Zawartość jądra obejmuje sok jądrowy lub karioplasm, chromatynę i jąderko. Skład soku jądrowego obejmuje różne białka, w tym większość enzymów jądrowych, wolne nukleotydy, aminokwasy, produkty jąderka i chromatynę, przemieszczające się z jądra do cytoplazmy. chromatyna zawiera DNA, białka i jest spiralną i zagęszczoną częścią chromosomów. jąderko jest gęstym zaokrąglonym ciałem znajdującym się w soku jądrowym. Liczba jąderek waha się od 1 do 5-7 lub więcej. Jąderka występują tylko w jądrach niedzielących się, podczas mitozy zanikają, a po zakończeniu podziału powstają ponownie. Jąderko nie jest samodzielną organellą komórkową, jest pozbawione błony i tworzy się wokół regionu chromosomu, w którym zakodowana jest struktura rRNA. Rybosomy powstają w jąderku, które następnie przemieszczają się do cytoplazmy. chromatyna zwane grudkami, ziarnistościami i sieciopodobnymi strukturami jądra, intensywnie wybarwionymi niektórymi barwnikami i różniącymi się kształtem od jąderka.
2)1. Teoria komórki
Teoria komórkowa jest uogólnioną ideą struktury komórek jako jednostek żywych, ich reprodukcji i roli w tworzeniu organizmów wielokomórkowych.
Powstanie i sformułowanie niektórych przepisów teoria komórki poprzedzone dość długim okresem gromadzenia obserwacji dotyczących budowy różnych organizmów jedno- i wielokomórkowych roślin i zwierząt. Okres ten wiązał się z rozwojem zastosowań i doskonaleniem różnych optycznych metod badawczych.
Robert Hooke jako pierwszy zaobserwował, używając soczewek powiększających, podział tkanki korka na „komórki” lub „komórki”. Jego opisy dały początek systematycznym badaniom anatomii roślin, które potwierdziły obserwacje Roberta Hooke'a i wykazały, że poszczególne części roślin składają się z blisko rozmieszczonych „pęcherzyków” lub „woreczków”. Później A. Leeuwenhoek odkrył świat organizmów jednokomórkowych i po raz pierwszy zobaczył komórki zwierzęce. Później komórki zwierzęce opisał F. Fontana; ale te i inne liczne badania nie doprowadziły w tamtym czasie do zrozumienia uniwersalności struktury komórkowej, do jasnego wyobrażenia o tym, czym jest komórka. Postęp w badaniach mikroanatomii i komórek związany jest z rozwojem mikroskopii w XIX wieku. Do tego czasu zmieniły się poglądy na temat struktury komórek: nie ściana komórkowa, ale jej faktyczna zawartość, protoplazma, zaczęła być uważana za główną rzecz w organizacji komórki. W protoplazmie odkryto stały składnik komórki, jądro. Wszystkie te liczne obserwacje pozwoliły T. Schwannowi w 1838 r. na dokonanie szeregu uogólnień. Pokazał, że komórki roślinne i zwierzęce są do siebie zasadniczo podobne. „Zasługą T. Schwanna nie było to, że odkrył komórki jako takie, ale to, że nauczył badaczy zrozumienia ich znaczenia”. Idee te zostały rozwinięte w pracach R. Virchowa. Stworzenie teorii komórki stało się ważnym wydarzeniem w biologii, jednym z decydujących dowodów jedności całej żywej przyrody. Teoria komórki wywarła znaczący wpływ na rozwój biologii, była głównym fundamentem rozwoju takich dyscyplin jak embriologia, histologia i fizjologia. Dało podstawy do zrozumienia życia, do wyjaśnienia pokrewieństwa organizmów, do zrozumienia indywidualnego rozwoju.
Główne założenia teorii komórki zachowały swoje znaczenie do dziś, chociaż przez ponad sto pięćdziesiąt lat zdobywano nowe informacje o budowie, czynności życiowej i rozwoju komórek. Obecnie teoria komórkowa postuluje:
1) Komórka jest podstawową jednostką życia: - poza komórką nie ma życia.
2) Komórka to pojedynczy system składający się z wielu elementów, które są ze sobą naturalnie połączone, reprezentując pewną integralną formację, składającą się ze sprzężonych jednostek funkcjonalnych - organelli lub organelli.
3) Komórki są podobne - homologiczne - pod względem struktury i podstawowych właściwości.
4) Liczba komórek zwiększa się, dzieląc pierwotną komórkę po podwojeniu jej materiału genetycznego: komórka po komórce.
5) Organizm wielokomórkowy jest nowy system, złożony zespół wielu komórek, zjednoczonych i zintegrowanych w układy tkanek i narządów, połączonych ze sobą za pomocą czynników chemicznych, humoralnych i nerwowych.
6) Komórki organizmów wielokomórkowych są totipotencjalne, tj. posiadają potencjały genetyczne wszystkich komórek danego organizmu, są równoważne pod względem informacji genetycznej, ale różnią się od siebie odmienną ekspresją różnych genów, co prowadzi do ich zróżnicowania morfologicznego i funkcjonalnego – różnicowania.
Idea komórki jako niezależnej jednostki życiowej została podana w pracach T. Schwanna. R. Virchow uważał również, że każda komórka nosi w sobie pełnię cech życia: „Komórka jest ostatnim elementem morfologicznym wszystkich żywych organizmów i nie mamy prawa szukać poza nią prawdziwej aktywności życiowej”.
Współczesna nauka w pełni potwierdziła tę tezę. W literaturze popularnej komórka często nazywana jest „atomem życia”, „kwantą życia”, podkreślając w ten sposób, że komórka jest najmniejszą jednostką żywej materii, poza którą nie ma życia.
Taka ogólna charakterystyka komórki powinna z kolei opierać się na definicji życia – czym jest życie, czym jest życie. Bardzo trudno jest podać ostateczną definicję życia, życia.
MV Volkenstein podaje następującą definicję życia: „organizmy żywe są otwartymi, samoregulującymi się i samoreprodukującymi się systemami, których najważniejszymi funkcjonującymi substancjami są białka i kwasy nukleinowe”. Istoty żywe charakteryzują się szeregiem kumulatywnych cech, takich jak zdolność do reprodukcji, wykorzystania i przekształcania energii, metabolizmu, wrażliwości i zmienności. I taką kombinację tych znaków można znaleźć na poziom komórki. Nie ma mniejszej jednostki życia niż komórka. Z komórki możemy wyizolować poszczególne składniki, a nawet molekuły i zweryfikować, czy wiele z nich ma określone cechy funkcjonalne. Zatem izolowane włókienka aktomiozyny mogą się kurczyć w odpowiedzi na dodanie ATP; poza komórką „działa” doskonale wiele enzymów biorących udział w syntezie lub rozkładzie złożonych cząsteczek bioorganicznych; izolowane rybosomy w obecności niezbędnych czynników mogą syntetyzować białka, opracowano układy pozakomórkowe do enzymatycznej syntezy kwasów nukleinowych itp. Czy wszystkie te składniki komórkowe, struktury, enzymy, cząsteczki można uznać za żywe? Czy kompleks aktomiozyny można uznać za żywy? Wydaje się, że nie, choćby dlatego, że posiada tylko część zbioru cech życia. To samo dotyczy pozostałych przykładów. Tylko komórka jako taka jest najmniejszą jednostką, która ma wszystkie właściwości razem wzięte, spełniające definicję „życia”.
3) Podstawą aparatu powierzchniowego komórek (PAC) jest na wolnym powietrzu błona komórkowa lub plazmalemma. Oprócz plazmalemmy, PAC ma kompleks nabłonkowy, podczas gdy eukarionty mają również kompleks podbłonowy. Głównymi składnikami biochemicznymi plazmalemmy (z greckiego osocza - tworzenie i lemat - skorupa, skorupa) są lipidy i białka. Ich stosunek ilościowy u większości eukariontów wynosi 1:1, a u prokariontów białka dominują w plazmalemmie. Niewielka ilość węglowodanów znajduje się w zewnętrznej błonie komórkowej oraz można znaleźć związki tłuszczopodobne (u ssaków - cholesterol, witaminy rozpuszczalne w tłuszczach). W 1925 r. E. Gorter i F. Grendel (Holandia) zasugerowali, że podstawą błony jest podwójna warstwa lipidów - warstwa bilipidowa. W 1935 roku J. Danieli i G. Dawson zaproponowali pierwszy przestrzenny model organizacji błon, nazwany modelem „kanapkowym” lub „kanapkowym”. Ich zdaniem podstawą błony jest warstwa bilipidowa, a obie powierzchnie warstwy pokryte są ciągłymi warstwami białek. Dalsze badania błon komórkowych, w tym plazmalemmy, wykazały, że prawie we wszystkich przypadkach mają one podobną budowę. W 1972 roku S. Singer i G. Nicholson (USA) sformułowali koncepcję płynna mozaika błony komórkowe (ryc.). Zgodnie z tym modelem membrany są oparte na warstwa lipidowa, ale zawarte w nim białka znajdują się w oddzielnych cząsteczkach i kompleksach, tj. mozaika (z francuskiego. mozaika - mozaika; obraz złożony z oddzielnych elementów). W szczególności cząsteczki białek integralnych (z łac. Współczesna biologia molekularna potwierdziła słuszność modelu mozaiki płynów, chociaż odkryto inne warianty błon komórkowych. W szczególności u archebakterii podstawą błony jest monowarstwa złożonego lipidu, a niektóre bakterie zawierają w cytoplazmie pęcherzyki błony, których ściany są reprezentowane przez monowarstwę białka. Kompleks nadbłonowy Aparat powierzchniowy komórek charakteryzuje się różnorodnością struktur (ryc.). U prokariotów kompleks epibłonowy jest w większości przypadków reprezentowany przez ścianę komórkową o różnej grubości, której podstawą jest złożona mureina glikoproteinowa (w archebakterii pseudomureina). U wielu eubakterii zewnętrzna część kompleksu epibłonowego składa się z innej błony o wysokiej zawartości lipopolisacharydów.U eukariotów uniwersalnym składnikiem kompleksu nadbłonowego są węglowodany - składniki glikolipidów i glikoprotein plazmalemmy. Z tego powodu pierwotnie nazywano go glikokaliksem (z gr. glycos – słodki, węglowodanowy i łac. kalum – gruba skórka, muszla). Oprócz węglowodanów glikokaliks obejmuje białka obwodowe powyżej warstwy bilipidowej. Bardziej złożone warianty kompleksu epibłonowego występują u roślin (ściana komórkowa zbudowana z celulozy), grzybów i stawonogów (powłoka zewnętrzna wykonana z chityny). podbłonowy(od łac. sub - under) kompleks jest charakterystyczny tylko dla komórek eukariotycznych. Składa się z różnorodnych białkowych struktur włóknistych: cienkich fibryli (z łac. fibril - włókno, nić), mikrofibryli (z greckiego micros - mały), szkieletowych (z gr. szkielet - wysuszony) włókienek i mikrotubul. Są one połączone ze sobą białkami i tworzą aparat mięśniowo-szkieletowy komórki. Kompleks podbłonowy oddziałuje z białkami błony komórkowej, które z kolei są związane z kompleksem nadbłonowym. W rezultacie PAH jest strukturalnie integralnym systemem. Pozwala to na wykonywanie ważnych dla komórki funkcji: izolacyjnych, transportowych, katalitycznych, sygnalizacyjnych receptorów i kontaktowych.
4) Błony zawierają również glikolipidy i cholesterol. glikolipidy to lipidy z przyłączonymi do nich węglowodanami. Podobnie jak fosfolipidy, glikolipidy Istnieją polarne głowy i niepolarne ogony. Cholesterol jest zbliżony do lipidów; jego cząsteczka ma również część polarną.
1. Twórcą teorii komórki jest:
R. Wirkchow
W I. Wernadski
T.Schwann
2. Teoria komórki powstała w:
1839
3. Grupa organelli błonowych cytoplazmy komórek eukariotycznych:
Rybosomy, mitochondria, plastydy, lizosomy
sferosomy, wakuole, plastydy
Mikrotubule, mikrofilamenty, rybosomy, centrosomy
EPS, rybosomy, lizosomy, aparat Golgiego
mitochondria, ER, aparat Golgiego
4. Organelle ogólnego przeznaczenia:
miofibryle
rybosomy, mitochondria
EPS, aparat Golgiego
wici, rybosomy
Wici, miofibryle
5. W jądrze międzyfazowym znajdują się:
Rybosomy
jąderka
kariolemma
sok jądrowy
chromatyna
6. Rodzaje chromosomów w zależności od umiejscowienia centromeru:
subcentryczny
telocentryczny
akrocentryczny
metacentryczny
nierówne ramię
7. Każdy chromosom w metafazie mitozy składa się z:
DNA, białka histonowe, białka niehistonowe
2x mikrofibryle
2x chromatydy
4x semichromatydy
chromon
8. Komórki somatyczne mają zestaw chromosomów:
poliploidalny
diploidalny
Haploidalny
tetraploidalny
9. Trwa synteza na gładkim EPS:
węglowodany
10. Chromoplasty zawierają pigment:
Hemoglobina
likopen
karoten
ksantofil
Chlorofil
11. Funkcje rybosomów:
hormony
Synteza węglowodanów
Kwasy nukleinowe
synteza białek
Synteza tłuszczów
12. Grupy integracyjne:
witamina
troficzny
minerał
wydzielniczy, wydalniczy
pigment
13. Komórki płciowe mają zestaw chromosomów:
diploidalny
poliploidalny
haploidalny
14. Do eukariontów należą:
cyjanobakteria
zwierzęta
porosty
rośliny
15. Grupa niebłonowych organelli cytoplazmy komórek eukariotycznych:
Rybosomy, wakuole, mikrotubule
rybosomy, centrosomy
ER, mitochondria, rybosomy
mikrotubule, mikrofibryle
Mikrofilamenty, plastydy, lizosomy
16. Funkcje hialoplazmy:
udział w metabolizmie
Syntetyczny
rozgraniczający
homeostatyczny
transportu
17. Funkcje jąder:
Synteza białek
tworzenie podjednostek rybosomów
synteza transportowego RNA
Tworzenie mikrofibryli
synteza rybosomalnego RNA
18. Funkcje EPS:
Chwytnik
transportu
wyznaczanie granic
Ochronny
syntetyczny
19. Funkcje lizosomów:
Synteza i akumulacja węglowodanów
niszczenie organów tymczasowych zarodków i larw
Synteza białek, tłuszczów, węglowodanów
autoliza, niszczenie cząstek obcych
trawienie wewnątrzkomórkowe
20. Vacuole to:
Organelle niebłonowe komórek zwierzęcych
Organelle błonowe komórek roślinnych
Organelle błonowe komórek bakteryjnych
Organelle błonowe zwierząt jednokomórkowych
Organelle niebłonowe komórek roślinnych
21. Funkcje wakuoli:
usunąć nadmiar wody
utrzymanie ciśnienia osmotycznego komórki
wykonują trawienie wewnątrzkomórkowe
utrzymanie turgoru w komórkach roślinnych
Przeprowadzić syntezę i akumulację tłuszczów w komórkach roślinnych
22. Chloroplasty zawierają pigment:
ksantofil
chlorofil
23. Rybosomy obejmują:
T-RNA, magnez
Białka, DNA, magnez
Tylko wiewiórki
Białka, i-RNA
białka, r-RNA, magnez
24. Skład mikrofilamentów obejmuje białko:
25. Mikrotubule zawierają białko:
tubulina
26. Mikrofibryle zawierają białko:
kolagen
keratyna
27. Jądro jest charakterystyczne dla komórek:
Wszystkie żywe organizmy
eukariont
rośliny
prokariota