STRUKTURA KOMÓRKI ROŚLINNEJ
Komórka roślinna składa się z mniej lub bardziej sztywnych Ściana komórkowa i protoplast. Ściana komórkowa to ściana komórkowa i błona cytoplazmatyczna. Termin protoplast pochodzi od słowa protoplazma, które przez długi czas odnosiło się do wszystkich żywych istot. Protoplast jest protoplazmą pojedynczej komórki.
Protoplast składa się z cytoplazmy i jądra. Cytoplazma zawiera organelle (rybosomy, mikrotubule, plastydy, mitochondria) i układy membranowe (retikulum endoplazmatyczne, dictyosomy). Cytoplazma obejmuje również macierz cytoplazmatyczną (substancję podstawową), w której zanurzone są organelle i układy błon. Cytoplazma jest oddzielona od ściany komórkowej błona plazmatyczna, która jest membraną elementarną. W przeciwieństwie do większości komórek zwierzęcych, komórki roślinne zawierają jedną lub więcej wakuoli. Są to pęcherzyki wypełnione cieczą i otoczone elementarną błoną (tonoplastem).
W żywej komórce roślinnej znajduje się główna substancja w ciągłym ruchu. Organelle biorą udział w ruchu, zwanym przepływem cytoplazmy lub cyklozą. Cykloza ułatwia przemieszczanie się substancji w komórce i ich wymianę między komórką a środowiskiem.
Membrana plazmowa. Jest to dwuwarstwowa struktura fosfolipidowa. Komórki roślinne charakteryzują się inwazjami błony plazmatycznej.
Błona plazmatyczna pełni następujące funkcje:
Uczestniczy w wymianie substancji między komórką a środowiskiem;
Koordynuje syntezę i montaż mikrofibryli celulozowych ściany komórkowej;
Przekazuje sygnały hormonalne i zewnętrzne, które kontrolują wzrost i różnicowanie komórek.
Rdzeń. Jest to najbardziej widoczna struktura w cytoplazmie komórki eukariotycznej. Jądro spełnia dwie ważne funkcje:
Kontroluje aktywność życiową komórki, określając, które białka iw jakim czasie powinny zostać zsyntetyzowane;
Przechowuje informacje genetyczne i przekazuje je komórkom potomnym podczas podziału komórki.
Jądro komórki eukariotycznej jest otoczone dwiema podstawowymi błonami, które tworzą otoczkę jądrową. Przesiąknięta jest licznymi porami o średnicy od 30 do 100 nm, widocznymi jedynie pod mikroskopem elektronowym. Pory mają złożoną strukturę. zewnętrzna męmbrana błona jądrowa w niektórych miejscach jest połączona z retikulum endoplazmatycznym. Otoczkę jądrową można postrzegać jako wyspecjalizowaną, lokalnie zróżnicowaną część retikulum endoplazmatycznego (ER).
W jądrze zabarwionym specjalnymi barwnikami można wyróżnić cienkie nici oraz grudki chromatyny i nukleoplazmy (głównej substancji jądra). Chromatyna składa się z DNA związanego ze specjalnymi białkami zwanymi histonami. W procesie podziału komórki chromatyna staje się coraz bardziej zagęszczona i składa się w chromosomy. DNA koduje informację genetyczną.
Organizmy różnią się liczbą chromosomów w komórkach somatycznych. Na przykład kapusta ma - 20 chromosomów; słonecznik - 34; pszenica - 42; ludzi - 46, a jeden z gatunków paproci Ophioglossum - 1250. Komórki płciowe (gamety) mają tylko połowę liczby chromosomów charakterystycznych dla komórek somatycznych organizmu. Liczba chromosomów w gametach nazywana jest haploidalną (pojedynczą), w komórkach somatycznych - diploidalną (podwójną). Komórki, które mają więcej niż dwa zestawy chromosomów, nazywane są poliploidalnymi.
Pod mikroskopem świetlnym można zobaczyć kuliste struktury - jąderka. Każde jądro ma jedno lub więcej jąderek, które są widoczne w jądrach niedzielących się. Rybosomalny RNA jest syntetyzowany w jąderku. Zwykle w jądrach organizmów diploidalnych występują dwa jąderka, po jednym dla każdego zbiór haploidalny chromosomy. Jąderka nie mają własnej błony. Pod względem biochemicznym jąderka są scharakteryzowane wysokie stężenie RNA, który jest tutaj związany z fosfoproteinami. Wielkość jąderka zależy od stan funkcjonalny komórki. zauważono, że w szybko rosnącej komórce, w której zachodzą intensywne procesy syntezy białek, jąderka powiększają się. W jąderkach powstają mRNA i rybosomy, które pełnią funkcję syntetyczną tylko w jądrze.
Nukleoplazma (karioplazma) jest reprezentowana przez jednorodną ciecz, w której rozpuszczają się różne białka, w tym enzymy.
Plastydy. Charakterystycznymi składnikami komórek roślinnych są wakuole, celulozowe ściany komórkowe i plastydy. Każdy plastyd ma swoją własną powłokę, składającą się z dwóch elementarnych membran. Wewnątrz plastydu wyróżnia się system membranowy i różne stopnie jednorodna substancja - zrąb. Dojrzałe plastydy są klasyfikowane na podstawie zawartych w nich pigmentów.
Chloroplasty, w których zachodzi fotosynteza, zawierają chlorofile i karotenoidy. Zwykle mają postać krążka o średnicy 4 - 5 mikronów. W jednej komórce mezofilu (środek liścia) może znajdować się 40 - 50 chloroplastów; w arkuszach mm 2 - około 500 000. w cytoplazmie chloroplasty są zwykle zlokalizowane równolegle do błony komórkowej.
Wewnętrzna struktura chloroplastu jest złożona. Zrąb jest przesiąknięty rozwiniętym systemem błon w postaci pęcherzyków - tylakoidów. Każdy tylakoid składa się z dwóch błon. Forma tylakoidów pojedynczy system. Z reguły są one zbierane w stosy - ziarna, przypominające kolumny monet. Tylakoidy poszczególnych granów są połączone tylakoidami zrębu lub tylakoidami międzyziarnistymi. Chlorofil i karotenoidy są osadzone w błonach tylakoidów. Chloroplasty w zielonych roślinach i algach często zawierają ziarna skrobi i małe kropelki lipidów (tłuszczu). Ziarna skrobi są tymczasowymi magazynami produktów fotosyntezy. Mogą zniknąć z chloroplastów rośliny, która była w ciemności tylko przez 24 godziny, i pojawić się ponownie 3-4 godziny po przeniesieniu rośliny na światło.
Chloroplasty to półautonomiczne organelle i przypominają bakterie. Na przykład rybosomy bakterii i chloroplastów mają dość duże podobieństwo. Są mniejsze niż rybosomy eukariotyczne. Synteza białek na rybosomach bakterii i chloroplastach jest hamowana przez chloramfenikol, który nie ma wpływu na komórki eukariotyczne. Ponadto zarówno bakterie, jak i chloroplasty mają podobny typ nukleoidu, zorganizowanego w podobny sposób. Pomimo faktu, że tworzenie chloroplastów i synteza zawartych w nich pigmentów jest w dużej mierze kontrolowana przez chromosomalny DNA komórki, to jednak przy braku własnego DNA chloroplasty nie tworzą się.
Chloroplasty można uznać za główne organelle komórkowe, ponieważ są pierwszymi w łańcuchu transformacji energii słonecznej, w wyniku czego ludzkość otrzymuje zarówno żywność, jak i paliwo. W chloroplastach zachodzi nie tylko fotosynteza. Biorą również udział w syntezie aminokwasów i Kwasy tłuszczowe, służą jako magazyn tymczasowych zapasów skrobi.
Chromoplasty to pigmentowane plastydy. Różnorodne w formie, nie posiadają chlorofilu, ale syntetyzują i gromadzą karotenoidy, które nadają żółty, pomarańczowy, czerwony kolor kwiatom, starym liściom, owocom i korzeniom. chromoplasty mogą rozwijać się z chloroplastów, które w tym przypadku tracą chlorofil i struktury błony wewnętrznej, akumulują karotenoidy. Dzieje się tak podczas dojrzewania wielu owoców. Chromoplasty przyciągają owady i inne zwierzęta, z którymi współewoluowały.
Leukoplasty to niepigmentowane plastydy. Niektóre z nich syntetyzują skrobię (amyloplasty), inne są zdolne do tworzenia różne substancje, w tym lipidy i białka. W świetle leukoplasty zamieniają się w chloroplasty.
Proplastydy to małe, bezbarwne lub jasnozielone, niezróżnicowane plastydy występujące w merystematycznych (dzielących się) komórkach korzeni i pędów. Są prekursorami innych, bardziej zróżnicowanych plastydów - chloroplastów, chromoplastów i aminoplastów. Jeśli rozwój protoplastów jest opóźniony z powodu braku światła, może pojawić się w nich jedno lub więcej ciałek prolamelarnych, które są półkrystalicznymi skupiskami błon kanalikowych. Plastydy zawierające ciałka prolamelarne nazywane są etioplastami. W świetle etioplasty zamieniają się w chloroplasty, podczas gdy błony ciał prolamelarnych tworzą tylakoidy. Etioplasty powstają w liściach roślin, które są w ciemności. protoplasty zarodków nasiennych najpierw przekształcają się w etioplasty, z których następnie w świetle rozwijają się chloroplasty. Plastydy charakteryzują się stosunkowo łatwymi przejściami z jednego rodzaju do drugiego. Plastydy, podobnie jak bakterie, rozmnażają się, dzieląc się na dwie części. W komórkach merystematycznych czas podziału protoplastów w przybliżeniu pokrywa się z czasem podziału komórki. Jednak w dojrzałych komórkach większość Plastyd powstaje w wyniku podziału dojrzałych plastydów.
mitochondria. Podobnie jak chloroplasty, mitochondria są otoczone dwiema podstawowymi błonami. Wewnętrzna błona tworzy liczne fałdy i wypukłości - cristae, które znacznie zwiększają wewnętrzną powierzchnię mitochondriów. Są znacznie mniejsze niż plastydy, mają średnicę około 0,5 µm oraz zmienną długość i kształt.
W mitochondriach zachodzi proces oddychania, w wyniku którego dochodzi do rozpadu cząsteczek organicznych z uwolnieniem energii i przekazaniem jej do cząsteczek ATP, głównej rezerwy energetycznej wszystkich komórek eukariotycznych. Większość komórek roślinnych zawiera setki lub tysiące mitochondriów. Ich liczba w jednej komórce jest określona przez zapotrzebowanie komórki na ATP. Mitochondria są w ciągłym ruchu, przemieszczając się z jednej części komórki do drugiej, łącząc się ze sobą i dzieląc. Mitochondria zwykle gromadzą się tam, gdzie potrzebna jest energia. Jeśli błona plazmatyczna aktywnie przenosi substancje z komórki do komórki, wówczas mitochondria znajdują się wzdłuż powierzchni błony. W ruchliwych glonach jednokomórkowych mitochondria gromadzą się u podstawy wici, dostarczając energię niezbędną do ich ruchu.
Mitochondria, podobnie jak plastydy, są półautonomicznymi organellami zawierającymi składniki niezbędne do syntezy własnych białek. Błona wewnętrzna otacza płynną matrycę, która zawiera białka, RNA, DNA, bakteryjne rybosomy i różne substancje rozpuszczone. DNA istnieje w postaci kolistych cząsteczek zlokalizowanych w jednym lub kilku nukleoidach.
Na podstawie podobieństwa bakterii do mitochondriów i chloroplastów komórek eukariotycznych można przypuszczać, że mitochondria i chloroplasty wywodzą się od bakterii, które znalazły „schronienie” w większych komórkach heterotroficznych – prekursorach eukariontów.
Mikrociała. W przeciwieństwie do plastydów i mitochondriów, które są ograniczone dwiema błonami, mikrociała są kulistymi organellami otoczonymi pojedynczą błoną. Mikrociała mają ziarnistą (granulowaną) zawartość, czasami zawierają również krystaliczne wtrącenia białkowe. Mikrociała są związane z jednym lub dwoma regionami retikulum endoplazmatycznego.
Niektóre mikrociała, zwane proksysomami, grają ważna rola w metabolizmie kwasu glikolowego, który jest bezpośrednio związany z fotooddychaniem. W zielonych liściach są one związane z mitochondriami i chloroplastami. Inne mikrociała, zwane glioksysomami, zawierają enzymy potrzebne do przekształcania tłuszczów w węglowodany. Dzieje się tak w przypadku wielu nasion podczas kiełkowania.
Wakuole to obszary komórki ograniczone błoną, wypełnione płynem - sokiem komórkowym. Otoczone są tonoplastem (błoną wakuolową).
Młoda komórka roślinna zawiera liczne małe wakuole, które w miarę starzenia się łączą się w jedną dużą wakuolę. W dojrzałej komórce do 90% jej objętości może zajmować wakuola. W tym przypadku cytoplazma jest dociskana w postaci cienkiej warstwy obwodowej do błony komórkowej. Wzrost wielkości komórek wynika głównie ze wzrostu wakuoli. W rezultacie powstaje ciśnienie turgoru i zachowana jest elastyczność tkanki. Jest to jedna z głównych funkcji wakuoli i tonoplastu.
Głównym składnikiem soku jest woda, reszta zależy od rodzaju rośliny i jej stanu fizjologicznego. Wakuole zawierają sole, cukry, rzadziej białka. Tonoplast odgrywa aktywną rolę w transporcie i gromadzeniu się niektórych jonów w wakuoli. Stężenie jonów w soku komórkowym może znacznie przekraczać jego stężenie w środowisku. Na wysoka zawartość Niektóre substancje tworzą kryształy w wakuolach. Najczęstsze kryształy szczawianu wapnia to inny kształt.
Wakuole są miejscami gromadzenia się produktów przemiany materii (metabolizmu). Mogą to być białka, kwasy, a nawet substancje toksyczne dla człowieka (alkaloidy). Często osadzają się pigmenty. Niebieski, fioletowy, purpurowy, ciemnoczerwony, karmazynowy dają komórkom roślinnym pigmenty z grupy antocyjanów. W przeciwieństwie do innych pigmentów dobrze rozpuszczają się w wodzie i znajdują się w soku komórkowym. Decydują o czerwonym i niebieskim zabarwieniu wielu warzyw (rzodkiewki, rzepa, kapusta), owoców (winogrona, śliwki, wiśnie), kwiatów (chabry, pelargonie, ostróżki, róże, piwonie). Czasami te pigmenty maskują chlorofil w liściach, na przykład w ozdobnym czerwonym klonie. Antocyjany barwią jesienne liście jasny czerwony kolor. Powstają w chłodne słoneczne dni, kiedy synteza chlorofilu zatrzymuje się w liściach. W liściach, gdy nie tworzą się antocyjany, po zniszczeniu chlorofilu zauważalne stają się żółto-pomarańczowe karotenoidy chloroplastów. Liście są najjaśniejsze w chłodnej, jasnej jesieni.
Wakuole biorą udział w niszczeniu makrocząsteczek, w krążeniu ich składników w komórce. Rybosomy, mitochondria, plastydy wpadające do wakuoli ulegają zniszczeniu. Zgodnie z tą czynnością trawienną można je porównać z lizosomami - organellami komórek zwierzęcych.
Wakuole powstają z retikulum endoplazmatycznego (retikulum)
Rybosomy. Małe cząsteczki (17 - 23nm), składające się z mniej więcej równych ilości białka i RNA. W rybosomach aminokwasy łączą się, tworząc białka. W komórkach o aktywnym metabolizmie jest ich więcej. Rybosomy znajdują się w cytoplazmie komórki swobodnie lub przyczepione do retikulum endoplazmatycznego (80S). Występują również w jądrze (80S), mitochondriach (70S), plastydach (70S).
Rybosomy mogą tworzyć kompleks, na którym zachodzi jednoczesna synteza identycznych polipeptydów, o których informacja pochodzi z jednej cząsteczki i RNA. Taki kompleks nazywa się polirybosomami (polisomami). Komórki, które syntetyzują białka w dużych ilościach, mają rozbudowany system polisomów, do których często są przyłączone powierzchnia zewnętrzna skorupy jądra.
Retikulum endoplazmatyczne. Jest to złożony trójwymiarowy system membranowy o nieokreślonym zasięgu. W kontekście ER wygląda jak dwie elementarne membrany z wąską przezroczystą przestrzenią między nimi. Kształt i długość ER zależy od rodzaju komórki, jej aktywności metabolicznej oraz stopnia zróżnicowania. W komórkach, które wydzielają lub przechowują białka, ER ma kształt płaskiego worka lub cysterny, z licznymi rybosomami przyczepionymi do jego zewnętrznej powierzchni. Ta siateczka nazywana jest szorstką retikulum endoplazmatycznym. Gładki ER ma zwykle kształt cylindryczny. W tej samej komórce może znajdować się szorstka i gładka siateczka śródplazmatyczna. Z reguły istnieje między nimi wiele połączeń.
Retikulum endoplazmatyczne działa jako system komunikacyjny komórki. Jest związany z powłoka zewnętrzna jądra. W rzeczywistości te dwie struktury tworzą jeden system membranowy. Kiedy otoczka jądrowa pęka podczas podziału komórki, jej fragmenty przypominają fragmenty ER. Retikulum endoplazmatyczne to system transportu substancji: białek, lipidów, węglowodanów do różnych części komórki. retikulum endoplazmatyczne sąsiednich komórek są połączone przez nici cytoplazmatyczne - plazmodesmy - które przechodzą przez błony komórkowe.
Retikulum endoplazmatyczne jest głównym miejscem syntezy błony komórkowe. W niektórych komórkach roślinnych tworzą się tu błony wakuoli i mikrociał, cysterny dictyosomów.
Aparat Golgiego. Termin ten jest używany w odniesieniu do wszystkich dictyosomów lub ciał Golgiego w komórce. Dictyosomy to grupy płaskich, dyskowatych pęcherzyków lub cystern, które się rozgałęziają skomplikowany system kanaliki. Dictyosomy u roślin wyższych składają się z 4 - 8 cystern, zebranych razem.
Zwykle w paczce zbiorników wyróżnia się stronę rozwijającą się i dojrzewającą. błony tworzących się cystern przypominają strukturą błony ER, a błony dojrzewających cystern przypominają błonę komórkową.
Dictyosomy biorą udział w wydzielaniu, a u większości roślin wyższych w tworzeniu błon komórkowych. Polisacharydy ściany komórkowej syntetyzowane przez dictyosomy gromadzą się w pęcherzykach, które następnie oddzielają się od dojrzewających cystern. Te pęcherzyki wydzielnicze migrują i łączą się z CMP; natomiast zawarte w nich polisacharydy wbudowują się w błonę komórkową. Niektóre substancje gromadzące się w dictyosomach powstają w innych strukturach, na przykład w ER, a następnie są transportowane do dictyosomów, gdzie są modyfikowane (modyfikowane) przed sekrecją. Na przykład glikoproteiny są ważne materiał konstrukcyjny Błona komórkowa. Część białkowa jest syntetyzowana przez polisomy szorstkiego ER, część węglowodanowa jest syntetyzowana w dictyosomach, gdzie obie części łączą się tworząc glikoproteiny.
Membrany to dynamiczne, ruchome struktury, które nieustannie zmieniają swój kształt i powierzchnię. Koncepcja układu endoplazmatycznego opiera się na ruchliwości błon. Zgodnie z tą koncepcją błony wewnętrzne cytoplazmy, z wyjątkiem błon mitochondriów i plastydów, stanowią jedną całość i wywodzą się z retikulum endoplazmatycznego. Nowe cysterny dictyosomów tworzą się z retikulum endoplazmatycznego przez etap pęcherzyków pośrednich, a pęcherzyki wydzielnicze, które oddzielają się od dictyosomów, ostatecznie przyczyniają się do tworzenia błony plazmatycznej. W ten sposób retikulum endoplazmatyczne i dictyosomy tworzą funkcjonalną całość, w której dictyosomy odgrywają rolę struktury pośrednie w procesie przekształcania błon podobnych do retikulum endoplazmatycznego w błony podobne do plazmy. W tkankach, których komórki słabo rosną i dzielą się, składniki błony są stale aktualizowane.
Mikrotubule występują w prawie wszystkich komórkach eukariotycznych. Są to cylindryczne struktury o średnicy około 24 nm. Ich długość jest różna. Każdy kanalik składa się z podjednostek białka zwanego tubuliną. Podjednostki tworzą 13 podłużnych włókien otaczających centralną jamę. Mikrotubule są strukturami dynamicznymi, są regularnie niszczone i formowane na określonych etapach cyklu komórkowego. Są montowane w specjalne miejsca zwane centrami organizacji mikrotubul. W komórkach roślinnych mają słabo wyrażoną amorficzną strukturę.
Funkcje mikrotubul: uczestniczą w tworzeniu błony komórkowej; kierują pęcherzyki dictyosomów do tworzącej się błony, jak nici wrzeciona, które tworzą się w dzielącej się komórce; odgrywają rolę w tworzeniu płytki komórkowej (początkowa granica między komórkami potomnymi). Ponadto mikrotubule ważny składnik wici i rzęski, w których ruchu odgrywają ważną rolę.
Mikrofilamenty, podobnie jak mikrotubule, znajdują się w prawie wszystkich komórkach eukariotycznych. Są to długie włókna o grubości 5–7 nm, składające się z kurczliwego białka aktyny. Pęczki mikrofilamentów znajdują się w wielu komórkach roślin wyższych. Najwyraźniej odgrywają ważną rolę w prądach cytoplazmy. Mikrofilamenty wraz z mikrotubulami tworzą elastyczną sieć zwaną cytoszkieletem.
Substancja podstawowa od dawna jest uważana za jednorodny (homogeniczny) roztwór bogaty w białko z niewielką liczbą struktur lub w ogóle ich nie ma. Jednak obecnie, przy użyciu wysokonapięciowego mikroskopu elektronowego, ustalono, że substancja podstawowa jest trójwymiarową siatką zbudowaną z cienkich (o średnicy 3–6 nm) pasm wypełniających całą komórkę. Inne składniki cytoplazmy, w tym mikrotubule i mikrofilamenty, są zawieszone w tej siatce mikrotrabekularnej.
Struktura mikrotrabekularna to sieć pasm białkowych, między którymi przestrzeń jest wypełniona wodą. Wraz z wodą siatka ma konsystencję żelu, który wygląda jak galaretowate ciała.
Organelle są przyczepione do siatki mikrotrabekularnej. Krata zapewnia połączenie między oddzielne części komórki i kieruje transportem wewnątrzkomórkowym.
Kropelki lipidów to kuliste struktury, które nadają cytoplazmie ziarnistość komórka roślinna pod mikroskopem świetlnym. Na mikrografach elektronowych wyglądają na amorficzne. Bardzo podobne, ale mniejsze kropelki znajdują się w plastydach.
Kropelki lipidów, myląc je z organellami, nazywały je sferosomami i uważały, że otoczone są jednowarstwową lub dwuwarstwową błoną. Jednak ostatnie dane pokazują, że kropelki lipidów nie mają błon, ale mogą być pokryte białkiem.
Substancje ergastyczne to „produkty pasywne” protoplastu: substancje rezerwowe lub produkty odpadowe. Mogą pojawiać się i znikać w różne okresy cykl komórkowy. Z wyjątkiem ziaren skrobi, kryształów, barwników antocyjanowych i kropelek lipidów. Należą do nich żywice, gumy, garbniki i białka. Substancje ergastyczne są częścią błony komórkowej, głównej substancji cytoplazmy i organelli, w tym wakuoli.
Wici i rzęski to cienkie, podobne do włosów struktury, które wystają z powierzchni wielu komórek eukariotycznych. Mają stałą średnicę, ale długość waha się od 2 do 150 mikronów. Konwencjonalnie dłuższe i nieliczne z nich nazywane są wiciami, a krótsze i liczniejsze nazywane są rzęskami. Nie ma wyraźnych różnic między tymi dwoma typami struktur, więc termin wici jest używany w odniesieniu do obu.
U niektórych alg i grzybów wici są narządami ruchu, za pomocą których poruszają się w wodzie. W roślinach (np. mchach, wątrobowcach, paprociach, niektórych nagonasiennych) tylko komórki rozrodcze (gamety) mają wici.
Każda wici ma określoną organizację. Zewnętrzny pierścień 9 par mikrotubul otacza dwie dodatkowe mikrotubule znajdujące się w środku wici. Zawierające enzym „uchwyty” rozciągają się od jednej mikrotubuli każdej z zewnętrznych par. Jest to podstawowy wzorzec organizacji 9+2 występujący we wszystkich wici organizmów eukariotycznych. Uważa się, że ruch wici opiera się na przesuwaniu się mikrotubul, podczas gdy zewnętrzne pary mikrotubul poruszają się jedna po drugiej bez skurczu. Przesuwanie się par mikrotubul względem siebie powoduje miejscowe wyginanie wici.
Wici „wyrastają” z cytoplazmatycznych cylindrycznych struktur zwanych ciałami podstawowymi, które również tworzą podstawową część wici. Ciała podstawowe mają Struktura wewnętrzna, przypominający strukturą wici, z wyjątkiem tego, że kanaliki zewnętrzne są zebrane w trojaczki, a nie w pary, a kanalików środkowych nie ma.
Ściana komórkowa. Ściana komórkowa ogranicza rozmiar protoplastu i zapobiega jego pęknięciu w wyniku absorpcji wody przez wakuolę.
Ściana komórkowa pełni określone funkcje, które są ważne nie tylko dla komórki i tkanki, w której komórka się znajduje, ale także dla całej rośliny. Ściany komórkowe odgrywają zasadniczą rolę we wchłanianiu, transporcie i wydalaniu substancji, a ponadto może się w nich koncentrować aktywność lizosomalna, czyli trawienna.
Składniki ściany komórkowej. Najbardziej typowym składnikiem ściany komórkowej jest celuloza, która w dużej mierze determinuje jej architekturę. Cząsteczki celulozy składają się z powtarzających się cząsteczek glukozy połączonych końcami. Długie, cienkie cząsteczki celulozy łączą się w mikrofibryle o grubości 10 - 25 nm. Mikrofibryle splatają się i tworzą cienkie nitki, które z kolei można owijać jedna wokół drugiej, jak nitki liny. Każda taka „sznurówka”, czyli makrofibryla, ma grubość około 0,5 mikrona, osiągając długość 4 mikronów. Makrofibryle są tak mocne, jak drut stalowy tej samej wielkości.
Ramka celulozowa ściany komórkowej jest wypełniona splecionymi z nią cząsteczkami matrycy celulozowej. Składa się z polisacharydów zwanych hemicelulozami oraz pektyn lub pektyn, chemicznie bardzo zbliżonych do hemiceluloz.
Inny składnik ściany komórkowej, lignina, jest najobficiej występującym polimerem w komórkach roślinnych po celulozie. Lignina zwiększa sztywność ściany i zwykle występuje w komórkach pełniących funkcję podtrzymującą lub mechaniczną.
Kutyna, suberyna, wosk - zwykle osadzają się w łuskach tkanek ochronnych roślin. Na przykład kutyna znajduje się w błonach komórkowych naskórka, a suberyna w wtórnej tkance ochronnej, korku. Obie substancje występują w połączeniu z woskami i zapobiegają nadmiernej utracie wody z rośliny.
Warstwy ściany komórkowej. Grubość ścianek komórek roślinnych jest bardzo zróżnicowana, w zależności od roli komórek w strukturze rośliny i wieku samej komórki. Pod mikroskop elektronowy w ścianie komórkowej roślin widoczne są dwie warstwy: blaszka środkowa (zwana także substancją międzykomórkową) i pierwotna ściana komórkowa. Wiele komórek kładzie kolejną warstwę - wtórną Ściana komórkowa. Płytka środkowa znajduje się między głównymi ścianami sąsiednich komórek. Ściana wtórna, jeśli występuje, jest osadzana przez protoplast komórki na wewnętrznej powierzchni pierwotnej ściany komórkowej.
Płyta środkowa. Blaszka środkowa składa się głównie z pektyn. Tam, gdzie powinna powstać ściana komórkowa, między dwiema nowo utworzonymi komórkami, najpierw zauważa się gęsty splot kanalików retikulum endoplazmatycznego i cysterny aparatu Golgiego (dictyosomy). Wtedy w tym miejscu pojawiają się bąbelki wypełnione pektyną (z polisacharydów). Pęcherzyki te są oddzielane od zbiorników aparatu Golgiego. Wczesna ściana komórkowa zawiera różne polisacharydy, z których głównymi są pektyny i hemiceluloza. Później składa się z gęstszych substancji - celulozy i ligniny.
Pierwotna ściana komórkowa. Jest to warstwa otoczki celulozowej, która jest osadzana przed lub w trakcie wzrostu komórek. Oprócz celulozy, hemicelulozy i pektyny błony pierwotne zawierają glikoproteinę. Muszle pierwotne mogą ulegać zdrewnieniu. Składnik pektynowy nadaje plastyczność, która umożliwia rozszerzanie się pierwotnej skorupy w miarę wydłużania się korzenia, łodygi lub liścia.
Komórki aktywnie dzielące się (większość dojrzałych komórek biorących udział w procesach fotosyntezy, oddychania i wydzielania) mają błony pierwotne. Takie komórki z pierwotną błoną i żywym protoplastem są zdolne do utraty charakterystyczny kształt, dzielą się i różnicują do nowego typu komórek. U roślin biorą udział w gojeniu się ran i regeneracji tkanek.
Pierwotne błony komórkowe nie mają jednolitej grubości na całej długości, ale mają cienkie sekcje, które nazywane są pierwotnymi polami porów. Nici cytoplazmy lub plazmodesmy, które łączą protoplasty sąsiednich komórek, zwykle przechodzą przez pierwotne pola porów.
wtórna ściana komórkowa. Pomimo faktu, że wiele komórek roślinnych ma tylko ścianę pierwotną, w niektórych protoplast odkłada ścianę wtórną w kierunku środka komórki. Zwykle dzieje się tak po ustaniu wzrostu komórek, a powierzchnia błony pierwotnej już się nie zwiększa. Z tego powodu wtórna powłoka różni się od podstawowej. Membrany wtórne są szczególnie potrzebne wyspecjalizowanym komórkom, które wzmacniają roślinę i przewodzą wodę. Po odłożeniu błony wtórnej protoplast tych komórek z reguły umiera. W skorupkach wtórnych jest więcej celulozy niż w skorupkach pierwotnych, a nie ma w nich substancji pektynowych i glikoprotein. Błona wtórna jest trudna do rozciągnięcia, jej macierz składa się z hemicelulozy.
W powłoce wtórnej można wyróżnić trzy warstwy - zewnętrzną, środkową i wewnętrzną (S 1, S 2, S 3). Warstwowa struktura skorup wtórnych znacznie zwiększa ich wytrzymałość. Mikrofibryle celulozy w błonie wtórnej są osadzane gęstiej niż w błonie pierwotnej. Lignina - zwykły składnik wtórne skorupy z drewna.
Pory w błonach stykających się komórek znajdują się naprzeciw siebie. Dwa przeciwległe pory i membrana porów tworzą parę porów. W komórkach z błonami wtórnymi występują dwa główne typy porów: proste i obramowane. W porach graniczących ściana wtórna wisi nad jamą porów. W prostych porach tak nie jest.
Wzrost ściany komórkowej. W miarę wzrostu komórki zwiększa się grubość i powierzchnia błony komórkowej. Rozciąganie skorupy to złożony proces. Jest kontrolowany przez protoplast i regulowany przez hormon auksyny.
W komórkach rosnących równomiernie we wszystkich kierunkach odkładanie miofibryli jest przypadkowe. Te miofibryle tworzą nieregularną sieć. Takie komórki znaleziono w rdzeniu łodygi, tkankach spichrzowych oraz podczas hodowli komórek in vitro. W wydłużających się komórkach miofibryle osłonek bocznych osadzają się pod kątem prostym do osi wydłużenia.
Substancje macierzy - pektyny, hemicelulozy i glikoproteiny są przenoszone na błonę w pęcherzykach dictyosomów. Jednocześnie pektyny są bardziej charakterystyczne dla komórek rosnących, podczas gdy hemicelulozy przeważają w komórkach nie rosnących.
Mikrofibryle celulozowe są syntetyzowane na powierzchni komórki za pomocą kompleksu enzymatycznego związanego z błoną plazmatyczną. Orientacja mikrofibryli jest kontrolowana przez mikrotubule znajdujące się w wewnętrzna powierzchnia błona plazmatyczna.
Plazmodesma. Są to cienkie nici cytoplazmy, które łączą protoplasty sąsiednich komórek. Plazmodesmy albo przechodzą przez ścianę komórkową w dowolnym miejscu, albo są skoncentrowane w pierwotnych polach porów lub w błonach między parami porów. Pod mikroskopem elektronowym plazmodesmy wyglądają jak wąskie kanały wyłożone błoną plazmatyczną. Wzdłuż osi kanału mniejsza cylindryczna rurka, desmotubula, rozciąga się od jednej komórki do drugiej, która komunikuje się z retikulum endoplazmatycznym obu sąsiednich komórek. Wiele plazmodesmy tworzy się podczas podziału komórki, kiedy rurkowa retikulum endoplazmatyczne jest pobierane przez rozwijającą się płytkę komórkową. Plazmodesmy mogą również tworzyć się w błonach komórek, które się nie dzielą. Struktury te zapewniają sprawne przenoszenie określonych substancji z komórki do komórki.
Podział komórek. Na Organizmy wielokomórkowe podział komórek wraz ze wzrostem ich wielkości jest sposobem wzrostu całego organizmu. Nowe komórki powstałe podczas podziału są podobne pod względem struktury i funkcji, zarówno do komórki macierzystej, jak i do siebie nawzajem. Proces podziału u eukariontów można podzielić na dwa częściowo nakładające się etapy: mitozę i cytokinezę.
Mitoza to tworzenie się dwóch jąder potomnych z jednego jądra, morfologicznie i genetycznie równoważnych sobie nawzajem. Cytokineza to podział cytoplazmatycznej części komórki z utworzeniem komórek potomnych.
Cykl komórkowy. Żywa komórka przechodzi przez serię następujących po sobie zdarzeń, które składają się na cykl komórkowy. Czas trwania samego cyklu różni się w zależności od rodzaju komórki i czynniki zewnętrzne, na przykład od temperatury lub bezpieczeństwa składniki odżywcze. Zwykle cykl dzieli się na interfazę i cztery fazy mitozy.
Interfaza. Okres między kolejnymi podziałami mitotycznymi.
Interfaza jest podzielona na trzy okresy, oznaczone jako G1, S, G2.
W okresie G 1, który rozpoczyna się po mitozie. W tym okresie wzrasta ilość cytoplazmy, w tym różnych organelli. Ponadto, zgodnie ze współczesną hipotezą, w okresie G 1 syntetyzowane są substancje, które stymulują lub hamują okres S i resztę cyklu, determinując w ten sposób proces podziału.
Okres S następuje po okresie G1, w którym następuje powielenie materiału genetycznego (DNA).
W okresie G2, który następuje po S, powstają struktury bezpośrednio zaangażowane w mitozę, takie jak elementy wrzeciona.
Niektóre komórki przechodzą nieograniczoną liczbę cykli komórkowych. Ten Jednokomórkowe organizmy oraz niektóre komórki stref aktywnego wzrostu (merystemy). Niektóre wyspecjalizowane komórki po dojrzewaniu tracą zdolność do reprodukcji. Trzecia grupa komórek, na przykład te tworzące tkankę rany (kalus), zachowuje zdolność do podziału tylko w szczególnych warunkach.
Mitoza, czyli rozszczepienie jądrowe. Jest to ciągły proces podzielony na cztery fazy: profaza, metafaza, anafaza, telofaza. W wyniku mitozy materiał genetyczny, który podwoił się w interfazie, jest równo dzielony między dwa jądra potomne.
Jeden z najbardziej wczesne objawy przejście komórki do podziału to pojawienie się wąskiego, pierścieniowego paska mikrotubul bezpośrednio pod błoną plazmatyczną. Ten stosunkowo gęsty pas otacza jądro w płaszczyźnie równikowej przyszłego wrzeciona mitotycznego. Ponieważ pojawia się przed profazą, nazywa się go pasem preprofazowym. Zanika po wrzecionie mitotycznym, na długo przed pojawieniem się w późnej telofazie płytki komórkowej, która rozrasta się od środka ku obrzeżom i łączy się z błoną komórki macierzystej w obszarze zajmowanym wcześniej przez pas preprofazowy.
profaza. Na początku profazy chromosomy przypominają długie nitki rozproszone wewnątrz jądra. Następnie, gdy nici skracają się i pogrubiają, można zobaczyć, że każdy chromosom składa się nie z jednej, ale z dwóch splecionych nici zwanych chromatydami. W późnej profazie dwie skrócone sparowane chromatydy każdego chromosomu leżą obok siebie równolegle, połączone wąskim odcinkiem zwanym centromerem. Ma określoną pozycję na każdym chromosomie i dzieli chromosom na dwa ramiona o różnej długości.
Mikrotubule znajdują się równolegle do powierzchni jądra wzdłuż osi wrzeciona. Jest to najwcześniejsza manifestacja zespołu wrzeciona mitotycznego.
Pod koniec profazy jąderko stopniowo traci swój wyraźny zarys i ostatecznie zanika. Wkrótce potem otoczka jądrowa również się rozpada.
Metafaza. Na początku metafazy wrzeciono, które jest strukturą trójwymiarową, najszerszą w środku i zwężającą się ku biegunom, zajmuje miejsce zajmowane wcześniej przez jądro. Włókna wrzeciona są wiązkami mikrotubul. Podczas metafazy chromosomy, z których każdy składa się z dwóch chromatyd, są ułożone w taki sposób, że ich centromery leżą w płaszczyźnie równikowej wrzeciona. Dzięki centromerowi każdy chromosom jest przyczepiony do nici wrzeciona. Jednak niektóre nici przechodzą z jednego bieguna na drugi bez przyczepiania się do chromosomów.
Kiedy wszystkie chromosomy znajdują się w płaszczyźnie równikowej, metafaza jest zakończona. Chromosomy są gotowe do podziału.
Anafaza. Chromatydy każdego chromosomu rozdzielają się. Teraz to są chromosomy potomne. Po pierwsze, centromer dzieli się, a dwa chromosomy potomne są przyciągane do przeciwnych biegunów. W tym przypadku centromery przesuwają się do przodu, a ramiona chromosomów rozciągają się do tyłu. Włókna wrzeciona przyczepione do chromosomów skracają się, przyczyniając się do rozdzielenia chromatyd i ruchu chromosomów potomnych w przeciwnych kierunkach.
telofaza. W telofazie następuje zakończenie rozdzielenia dwóch identycznych grup chromosomów, podczas gdy wokół każdej z nich tworzy się błona jądrowa. Siateczka szorstka bierze w tym czynny udział. Aparat wrzeciona znika. Podczas telofazy chromosomy tracą wyrazistość konturów, rozciągają się, zamieniając z powrotem w cienkie nitki. Jąderka są przywrócone. Kiedy chromosomy stają się niewidoczne, mitoza jest zakończona. Dwa jądra potomne wchodzą w interfazę. Są genetycznie równoważne sobie nawzajem i macierzystemu jądru. Jest to bardzo ważne, ponieważ program genetyczny, a wraz z nim wszystkie cechy, muszą zostać przeniesione na organizmy potomne.
Czas trwania mitozy jest różny różne organizmy A to zależy od rodzaju tkaniny. Jednak profaza jest najdłuższa, a anafaza najkrótsza. W komórkach wierzchołka korzenia czas trwania profazy wynosi 1–2 godziny; metafaza - 5 - 15 min; anafaza - 2 - 10 minut; telofaza - 10 - 30 min. Czas trwania interfazy wynosi od 12 do 30 godzin.
W wielu komórkach eukariotycznych centra organizacji mikrotubul odpowiedzialne za tworzenie wrzeciona mitotycznego są związane z centriolami.
Cytokineza. Jest to proces podziału cytoplazmy. W większości organizmów komórki dzielą się, cofając ścianę komórkową i tworząc bruzdę rozszczepienia, która stopniowo się pogłębia, ściskając pozostałe włókna wrzeciona mitotycznego. We wszystkich roślinach (mszaki i rośliny naczyniowe) oraz w niektórych algach komórki dzielą się w wyniku tworzenia płytki komórkowej.
We wczesnej telofazie między dwoma jądrami potomnymi tworzy się beczkowaty układ włókien zwany phragmoplastem. Włókna phragmoplastu, podobnie jak włókna wrzeciona mitotycznego, składają się z mikrotubul. Małe krople pojawiają się w płaszczyźnie równikowej fragmoplastu. Łączą się, tworząc płytkę komórkową, która rośnie, aż dotrze do ściany dzielącej się komórki. To kończy rozdzielanie dwóch komórek potomnych. Łączące się kropelki to bąbelki odrywające się od aparatu Golgiego. Zasadniczo zawierają substancje pektynowe, z których powstaje środkowa płytka. Błony pęcherzyków biorą udział w budowie błony plazmatycznej po obu stronach płytki. W tym samym czasie plazmodesmy powstają z fragmentów rurkowatej retikulum endoplazmatycznego.
Po utworzeniu płytki środkowej każdy protoplast osadza na niej pierwotną błonę. Ponadto każda komórka potomna osadza nową warstwę błony wokół całego protoplastu, która kontynuuje błonę, która powstała z płytki komórkowej. Oryginalna powłoka komórki macierzystej jest niszczona w miarę wzrostu komórek potomnych.
Różne typy mitozy eukariotycznej
Opisany powyżej podział komórkowy u roślin, a także u zwierząt, nie jest jedyną formą pośredniego podziału komórkowego. Najprostszym rodzajem mitozy jest pleuromitoza. Przypomina podział binarny komórki prokariotyczne, w którym nukleoidy po replikacji pozostają związane z błoną plazmatyczną. Błona zaczyna rosnąć między punktami wiązania DNA iw ten sposób rozprzestrzenia się chromosomy różne obszary komórki. Następnie, podczas tworzenia zwężenia komórki, każda z cząsteczek DNA znajdzie się w nowej oddzielnej komórce.
Charakterystyczne dla podziału komórek eukariotycznych jest tworzenie się wrzeciona zbudowanego z mikrotubul. Z zamkniętą pleuromitozą (nazywa się to zamkniętą, ponieważ rozbieżność chromosomów zachodzi bez przerwania otoczki jądrowej), a nie centriole, ale inne struktury zlokalizowane na wewnątrz membrana nuklearna. Są to tak zwane ciała polarne o nieokreślonej morfologii, z których wychodzą mikrotubule. Są dwa takie ciała. Oddalają się od siebie nie tracąc kontaktu otoczka jądrowa. W wyniku tego powstają dwa półwrzeciona związane z chromosomami. Cały proces tworzenia wrzeciona mitotycznego i segregacji chromosomów zachodzi w tym przypadku pod otoczką jądrową. Ten typ mitozy występuje wśród pierwotniaków i jest szeroko rozpowszechniony u grzybów (chytridia, zygomycetes, drożdże, lęgniowce, ascomycetes, myxomycetes itp.). istnieją formy półzamkniętej pleuromitozy, gdy otoczka jądrowa jest niszczona na biegunach uformowanego wrzeciona.
Następną formą mitozy jest ortomitoza. W tym przypadku COMT znajdują się w cytoplazmie, od samego początku nie tworzą się półwrzeciona, ale wrzeciono dwubiegunowe. Istnieją trzy formy ortomitozy (mitozy normalnej), półzamkniętej i zamkniętej. W półzamkniętej ortomitozie powstaje bisymetryczne wrzeciono za pomocą TsOMT znajdującego się w cytoplazmie, otoczka jądrowa jest zachowana przez całą mitozę, z wyjątkiem stref polarnych. Masy materiału ziarnistego, a nawet centriole, mogą być wykrywane jako COMT. Ta forma mitozy występuje u niektórych w zoosporach zielonych, brązowych, czerwonych alg niższe grzyby i gregaryna. Przy zamkniętej ortomitozie błona jądrowa jest całkowicie zachowana, w której powstaje prawdziwe wrzeciono. Mikrotubule powstają w karioplazmie, rzadziej wyrastają z wewnętrznego TsOMT, który nie jest związany (w przeciwieństwie do pleuromitozy) z błoną jądrową. Ten typ mitozy jest typowy dla podziału mikrojąder orzęsków, ale może również wystąpić u pierwotniaków. W otwartej ortomitozie otoczka jądrowa całkowicie się rozpada. Ten typ podziału komórek jest charakterystyczny dla organizmów zwierzęcych, niektórych pierwotniaków i komórek roślin wyższych. Z kolei ta forma mitozy jest reprezentowana przez typy astralne i anastralne.
Z krótkiej recenzji wynika, że główna cecha Ogólnie rzecz biorąc, mitoza to pojawienie się struktur wrzeciona rozszczepienia, które powstaje w związku z TsOMT, który ma zróżnicowaną budowę.
Morfologia figury mitotycznej
Aparat mitotyczny jest szczególnie dobrze wyrażany na metafazowym etapie mitozy. W metafazie, w płaszczyźnie równikowej komórki, znajdują się chromosomy, z których wychodzą w przeciwnych kierunkach tzw. nici wrzecionowate, zbiegające się na dwóch różnych biegunach figury mitotycznej. Zatem wrzeciono mitotyczne jest zbiorem chromosomów, biegunów i włókien. Włókna wrzeciona to pojedyncze mikrotubule lub ich wiązki. Mikrotubule zaczynają się od biegunów wrzeciona, a część z nich biegnie do centromerów, gdzie znajdują się kinetochory chromosomów (mikrotubule kinetochorowe), część biegnie dalej w kierunku przeciwnego bieguna, ale do niego nie dociera. Nazywa się je „mikrotubulami międzybiegunowymi”. Od biegunów odchodzi grupa promienistych mikrotubul, tworząc wokół nich strukturę przypominającą „promienną poświatę” – są to mikrotubule astralne.
Pod względem morfologii figury mitotyczne dzielą się na typy astralne i anastralne.
Typ wrzeciona astralnego lub zbieżny charakteryzuje się tym, że jego bieguny są reprezentowane przez małą strefę, do której zbiegają się (zbiegają) mikrotubule. Zwykle centrosomy zawierające centriole znajdują się na biegunach wrzecion astralnych. Znane są jednak przypadki centralnych mitoz astralnych (podczas mejozy niektórych bezkręgowców). Ponadto odnotowuje się promieniste mikrotubule promieniujące z biegunów, które nie są częścią wrzeciona, ale tworzą strefy gwiaździste - cytastry. Ten typ podziału mitotycznego przypomina hantle. Anastrialny typ figury mitotycznej nie ma cytastrów na biegunach. Regiony polarne wrzeciona są tutaj szerokie, nazywane są czapami polarnymi, zawierają centriole. W tym przypadku włókna wrzeciona nie odbiegają od jednego punktu, ale rozchodzą się szerokim frontem (rozchodzą się) z całej strefy czap polarnych. Ten typ wrzeciona jest charakterystyczny dla dzielących się komórek roślin wyższych, ale można go również znaleźć u zwierząt wyższych. We wczesnej embriogenezie ssaków mitozy centriolarne (rozbieżne) obserwuje się podczas podziału dojrzewania oocytu oraz w I i II podziale zygoty. Ale już w trzecim podziale komórkowym i we wszystkich kolejnych komórkach dzielą się z udziałem wrzecion astralnych, w biegunach których zawsze znajdują się centriole. Ogólnie rzecz biorąc, dla wszystkich form mitozy chromosomy z ich kinetoforami, ciałami polarnymi (centrosomami) i włóknami wrzeciona pozostają wspólnymi strukturami.
Centromery i kinetochory
Centromery mogą mieć inna lokalizacja wzdłuż długości chromosomów. Centromery holocentryczne występują, gdy mikrotubule są połączone wzdłuż całego chromosomu (niektóre owady, nicienie, niektóre rośliny). Centromery monocentryczne - gdy mikrotubule są połączone z chromosomami w jednym miejscu. centromery monocentryczne mogą być punktowe (na przykład u niektórych pączkujących drożdży), gdy tylko jedna mikrotubula zbliża się do kinetochoru, oraz strefowe, gdzie wiązka mikrotubul zbliża się do złożonego kinetochoru. Pomimo różnorodności stref centromerowych, wszystkie są z nimi związane złożona struktura kinetochor, który ma fundamentalne podobieństwo w strukturze i funkcji u wszystkich eukariotów. Kinetochory to wyspecjalizowane struktury białkowe przez większą część zlokalizowane w centromerach chromosomów. Są to złożone kompleksy składające się z wielu białek. morfologicznie są bardzo podobne, mają taką samą budowę, od okrzemek po ludzi. Są to struktury trójwarstwowe: wewnętrzna gęsta warstwa przylegająca do ciała chromosomu, środkowa luźna warstwa i zewnętrzna gęsta warstwa. Z warstwy zewnętrznej rozciąga się wiele włókienek, tworząc tzw. włóknistą koronę kinetochoru. W forma ogólna kinetochory mają postać płytek lub krążków leżących w strefie pierwotnego zwężenia chromosomu, w centromerze. Zwykle na chromosom lub chromatydę przypada jeden kinetochor. Przed anafazą na każdej siostrzanej chromatydzie znajdują się kinetochory, z których każda komunikuje się z własną wiązką mikrotubul. W roślinach kinetochor nie wygląda jak talerze, ale półkule. Funkcjonalną rolą kinetochorów jest wiązanie ze sobą siostrzanych chromatyd, wiązanie mikrotubul mitotycznych, regulacja separacji chromosomów i faktyczne przemieszczanie chromosomów podczas mitozy z udziałem mikrotubul. Ogólnie rzecz biorąc, struktury białkowe, kinetochory, podwajają się w okresie S, równolegle do podwajania chromosomów. Ale ich białka są obecne na chromosomach we wszystkich okresach cyklu komórkowego.
TKANKA ROŚLINNA
Tkaniny edukacyjne(merystemy)
W ciele rośliny znajdują się tkanki edukacyjne różne miejsca, dlatego dzielą się na następujące grupy (ryc. 0; 1).
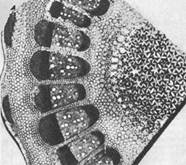
1. Merystemy wierzchołkowe (wierzchołkowe) znajdują się na wierzchołkach lub wierzchołkach narządów osiowych - łodygi, korzenia. Za pomocą tych merystemów organy wegetatywne roślin przeprowadzają wzrost długości.
2. Merystemy boczne są charakterystyczne dla narządów osiowych. Tam są umieszczone koncentrycznie, w formie sprzęgła.
3. Merystemy interkalarne lub interkalarne pochodzą z merystemów wierzchołkowych. Są to grupy komórek, które nie są jeszcze zdolne do namnażania się, ale wkroczyły na ścieżkę różnicowania. Nie ma wśród nich komórek początkowych, ale jest wiele wyspecjalizowanych.
4. Merystem rany zapewnia odbudowę uszkodzonej części ciała. Regeneracja zaczyna się od odróżnicowania, czyli odwrotnego rozwoju komórek wyspecjalizowanych do komórek merystematycznych. Zamieniają się w fellogen, który tworzy czop pokrywający powierzchnię rany. Komórki odróżnicowane, dzieląc się, mogą tworzyć luźną tkankę miąższową - kalus. Z niego, w określonych warunkach, powstają narządy roślinne.
Tkanki powłokowe
Pełnią rolę bariery granicznej, oddzielającej leżące poniżej tkanki środowisko. Powłoki pierwotne rośliny składają się wyłącznie z żywych komórek. Powłoki drugorzędowe i trzeciorzędowe - głównie z martwych z grubymi ścianami komórkowymi.
Główne funkcje tkanek powłokowych:
ochrona rośliny przed wysychaniem;
·Stopień ochrony szkodliwe mikroorganizmy;
· obrona przed oparzenie słoneczne;
· obrona przed uszkodzenie mechaniczne;
regulacja metabolizmu między rośliną a środowiskiem;
wrażenie irytacji.
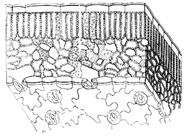
Podstawową tkanką powłokową jest naskórek, naskórek. Składa się z żywych komórek. Utworzony z merystemów wierzchołkowych. Obejmuje młode rosnące łodygi i liście.
Naskórek formował się u roślin w związku z wyjściem ze środowiska wodnego na ląd w celu zapobieżenia jego wysychaniu. Oprócz aparatów szparkowych wszystkie komórki naskórka są ze sobą ściśle połączone. Zewnętrzne ściany głównych komórek są grubsze niż pozostałe. Cała powierzchnia pokryta jest warstwą kutyny i wosków roślinnych. Ta warstwa nazywana jest naskórkiem (skórą). Nie występuje na rosnących korzeniach i podwodnych częściach roślin. Po wyschnięciu przepuszczalność łuski jest znacznie osłabiona.
Oprócz głównych komórek w naskórku znajdują się inne, w szczególności włosy lub trichomy. Są jednokomórkowe i wielokomórkowe (ryc. 2). Funkcjonalnie zwiększają powierzchnię naskórka, np. w strefie wzrostu korzenia, służą jako ochrona mechaniczna, przylegają do podpory i ograniczają utratę wody. Wiele roślin ma gruczołowe włosy, takie jak pokrzywy.
Tylko rośliny wyższe mają aparaty szparkowe w epidermie, które regulują wymianę wody i gazów. Jeśli nie ma naskórka, nie ma potrzeby stosowania aparatów szparkowych. Aparaty szparkowe to grupa komórek tworzących aparat szparkowy, który składa się z dwóch komórek spływowych oraz sąsiadujących ze sobą komórek naskórka – komórek bocznych. Różnią się od głównych komórek naskórka (ryc. 3). Komórki ochronne różnią się od otaczających je kształtem i obecnością dużej liczby chloroplastów oraz nierównomiernie pogrubionych ścian. Te, które są naprzeciw siebie, są grubsze niż pozostałe (ryc. 4). Pomiędzy komórkami ochronnymi tworzy się szczelina szparkowa, która prowadzi do przestrzeni podszparkowej zwanej jamą podszparkową. Komórki ochronne są wysoce fotosyntetyzujące. Zawierają duża liczba skrobia zapasowa i liczne mitochondria.
Liczba i rozmieszczenie aparatów szparkowych, rodzajów aparatów szparkowych różni się znacznie w różnych roślinach. U współczesnych mszaków nie ma aparatów szparkowych. Fotosynteza w nich jest przeprowadzana przez pokolenie gametofitów, a sporofity nie są zdolne do niezależnego istnienia.
Zwykle aparaty szparkowe znajdują się na spodniej stronie liścia. U roślin pływających na powierzchni wody - na górnej powierzchni. W liściach zbóż aparaty szparkowe są często równomiernie rozmieszczone po obu stronach. Takie liście są oświetlane w miarę równomiernie. Na 1 mm2 powierzchni może znajdować się od 100 do 700 aparatów szparkowych.
Wtórna tkanka powłokowa (peryderma). Ta tkanka zastępuje naskórek, gdy zielony kolor roczne pędy są zastępowane przez brązowe. Jest wielowarstwowy i składa się z centralnej warstwy komórek kambium - fellogenu. Komórki felogenu, dzieląc się, układają warstwę fellemu na zewnątrz, a felodermy do wewnątrz (ryc. 5).
Phelema lub korek. Początkowo składa się z żywych, cienkościennych komórek. Z biegiem czasu ich ściany są impregnowane suberyną i woskami roślinnymi i obumierają. Zawartość komórki jest wypełniona powietrzem.
Funkcje Fellema:
Zapobiega utracie wilgoci
chroni roślinę przed uszkodzeniami mechanicznymi;
Chroni przed patogenami
zapewnia izolację termiczną, ponieważ komórki są wypełnione powietrzem.
Komórki felogenu, zlokalizowane w samym naskórku, leżącej pod nim warstwie podnaskórkowej, rzadziej w głębokich warstwach kory pierwotnej, stanowią podstawę generacyjną kory pierwotnej.
Warstwa korka nie jest trwała. Występują w nim przerwy, które komunikują się z znajdującymi się w pobliżu przestrzeniami międzykomórkowymi. W tym samym czasie na powierzchni powstają małe guzki - przetchlinki, które komunikują przestrzenie międzykomórkowe z powietrzem atmosferycznym (ryc. 6.7).
Jesienią fallogen pod przetchlinkami układa warstwę zakorkowanych komórek, które znacznie ograniczają transpirację, ale jej całkowicie nie wykluczają. Wiosną warstwa ta zapada się od wewnątrz. Na jasnej korze brzozy soczewica jest wyraźnie widoczna w postaci ciemnych kresek.
Trzeciorzędowa tkanka powłokowa (skorupa) jest również charakterystyczna tylko dla drzewiastych form roślin.
Phellogen jest wielokrotnie odkładany w głębszych warstwach kory. Tkanki znajdujące się na zewnątrz ostatecznie obumierają, tworząc skorupę. Jego komórki są martwe i niezdolne do rozciągania. Jednak żywe komórki położone głębiej dzielą się, co prowadzi do zwiększenia poprzecznego rozmiaru łodygi. Z biegiem czasu zewnętrzna warstwa skorupy pęka. Czas wystąpienia takiej przerwy jest wartością dość stałą dla poszczególnych roślin. Na jabłoni dzieje się to w siódmym roku życia, w grabie - w pięćdziesiątym. U niektórych gatunków nie występuje w ogóle. Główną funkcją peelingu jest ochrona przed uszkodzeniami mechanicznymi i termicznymi.
Miąższ
Jest to grupa wyspecjalizowanych tkanek, które wypełniają przestrzenie wewnątrz ciała rośliny pomiędzy tkankami przewodzącymi i mechanicznymi (ryc. 8). Częściej komórki miąższu mają kształt zaokrąglony, rzadziej wydłużony. Charakterystyczna jest obecność rozwiniętych przestrzeni międzykomórkowych. Przestrzenie między komórkami tworzą razem system transportowy - apoplast. Ponadto przestrzenie międzykomórkowe tworzą „system wentylacyjny” rośliny. Poprzez aparaty szparkowe, czyli przetchlinki, są one połączone z powietrzem atmosferycznym i zapewniają optymalne warunki skład gazu wewnątrz rośliny. Rozwinięte przestrzenie międzykomórkowe są szczególnie potrzebne roślinom rosnącym na glebach podmokłych, gdzie normalna wymiana gazowa jest utrudniona. Ten miąższ nazywa się aerenchyma (ryc. 9).
Elementy miąższu, wypełniając szczeliny między innymi tkankami, pełnią również funkcję podporową. Komórki miąższu są żywe, nie mają grubych ścian komórkowych, jak sklerenchyma. Dlatego właściwości mechaniczne zapewnia turgor. Jeśli zawartość wody spada, co prowadzi do plazmolizy i więdnięcia rośliny.
Miąższ asymilacyjny tworzą cienkościenne komórki z wieloma przestrzeniami międzykomórkowymi. Komórki tej struktury zawierają wiele chloroplastów, dlatego nazywa się to chlorenchymą. Chloroplasty znajdują się wzdłuż ściany bez cieniowania się nawzajem. W miąższu asymilacyjnym zachodzą reakcje fotosyntezy, które dostarczają roślinie substancji organicznych i energii. Wynikiem procesów fotosyntezy jest możliwość istnienia wszystkich żywych organizmów na Ziemi.
Tkanki asymilacyjne występują tylko w oświetlonych częściach rośliny, oddzielone są od otoczenia przezroczystym naskórkiem. Jeśli naskórek zostanie zastąpiony nieprzezroczystymi wtórnymi tkankami powłokowymi, miąższ asymilacyjny zanika.
Miąższ spichrzowy służy jako pojemnik na substancje organiczne, które nie są czasowo wykorzystywane przez organizm roślinny. W zasadzie odkładanie materia organiczna Jak różnego rodzaju inkluzje są zdolne do każdej komórki z żywym protoplastem, ale niektóre komórki specjalizują się w tym (ryc. 10, 11). Związki bogate w energię odkładają się tylko w sezonie wegetacyjnym, zużywają w okresie spoczynku oraz w ramach przygotowań do kolejnego sezonu wegetacyjnego. Dlatego substancje rezerwowe osadzają się w organach wegetatywnych tylko u roślin wieloletnich.
Zbiornikiem wywarów mogą być zarówno narządy zwykłe (łodyga, korzeń), jak i specjalistyczne (kłącza, bulwy, cebulki). Wszystkie rośliny nasienne przechowują w nasionach cenne energetycznie substancje (liścienie, bielmo). Wiele roślin suchego klimatu magazynuje nie tylko materię organiczną, ale także wodę (ryc. 12). Na przykład aloes magazynuje wodę w mięsistych liściach, kaktusy w pędach.
tkaniny mechaniczne
Właściwości mechaniczne komórek roślinnych zapewniają:
Twarda skorupa komórki
turgor, czyli stan turgoru komórek.
Pomimo tego, że prawie wszystkie komórki tkanki mają właściwości mechaniczne, to jednak w roślinie występują tkanki, dla których właściwości mechaniczne są podstawowe. Są to collenchyma i sklerenchyma. Zwykle działają w interakcji z innymi tkankami. Wewnątrz ciała rośliny tworzą rodzaj ramy. Dlatego nazywa się je wzmocnieniem.
Nie wszystkie rośliny są równie dobrze wyrażane tkaniny mechaniczne. W znacznie mniejszym stopniu rośliny żyjące w środowisko wodne niż ziemskie. Powód jest taki, że rośliny wodne potrzebują mniej wsparcia wewnętrznego. Ich ciało jest w dużej mierze podtrzymywane przez otaczającą wodę. Powietrze na lądzie nie tworzy takiego wsparcia, ponieważ ma mniejszą gęstość niż woda. Z tego powodu obecność wyspecjalizowanych tkanek mechanicznych staje się istotna.
Udoskonalenie wewnętrznych struktur wsparcia nastąpiło w procesie ewolucji.
Collenchyma. Tworzą go tylko żywe komórki, wydłużone wzdłuż osi narządu. Ten typ tkanki mechanicznej powstaje bardzo wcześnie, w okresie pierwotnego wzrostu. Dlatego ważne jest, aby komórki pozostały żywe i zachowały zdolność do rozciągania się wraz z rozciągającymi się komórkami, które są w pobliżu.
Cechy komórek collenchyma:
Nierównomierne pogrubienie skorupy, w wyniku czego niektóre jej części pozostają cienkie, a inne pogrubiają;
muszle nie zdrewniały.
Komórki Collenchyma są rozmieszczone w różny sposób względem siebie. W sąsiednich komórkach na zwróconych do siebie rogach tworzą się zgrubienia (ryc. 13). Taka kollenchyma nazywana jest kollenchymą narożną. W innym przypadku komórki są ułożone w równoległe warstwy. Błony komórkowe zwrócone w stronę tych warstw są silnie pogrubione. To jest kolenchyma blaszkowata. Komórki mogą być ułożone luźno, z obfitymi przestrzeniami międzykomórkowymi - jest to luźna kolenchyma. Kollenchyma taka często występuje w roślinach na glebach podmokłych.
Collenchyma ma szczególne znaczenie w młodych roślinach, formach zielnych, a także w częściach roślin, w których nie występuje wzrost wtórny, takich jak liście. W tym przypadku układa się go bardzo blisko powierzchni, czasem tuż pod naskórkiem. Jeśli narząd ma twarze, wzdłuż ich grzebieni znajdują się grube warstwy kolenchymy.
Komórki Collenchyma są funkcjonalne tylko w obecności turgoru. Niedobór wody zmniejsza skuteczność kolenchymy i roślina przejściowo więdnie, np. liście ogórka zwisają w upalny dzień. Po napełnieniu komórek wodą przywracane są funkcje kolenchymy.
sklerenchyma. Drugi rodzaj tkanek mechanicznych. W przeciwieństwie do kolenchymy, gdzie wszystkie komórki są żywe, komórki sklerenchymy są martwe. Ich ściany są bardzo grube. Pełnią funkcję mechaniczną. Silne pogrubienie błony prowadzi do zakłócenia transportu substancji, w wyniku czego protoplast obumiera (ryc. 14). Lignifikacja błon komórkowych sklerenchymy następuje, gdy organ rośliny zakończył już swój wzrost. Dzięki temu nie przeszkadzają już w rozciąganiu otaczających tkanek.
W zależności od kształtu wyróżnia się dwa rodzaje komórek sklerenchymy - włókna i sklereidy.
Włókna mają silnie wydłużony kształt z bardzo grubymi ściankami i małą wnęką. Są nieco mniejsze niż włókna drzewne. Często pod naskórkiem tworzą podłużne warstwy i pasma. W łyku lub ksylemie można je znaleźć pojedynczo lub w grupach. W łyku nazywane są włóknami łykowymi (ryc. 15), aw ksylemie włóknami libriform (ryc. 16).
Sclereidy lub komórki kamieniste są reprezentowane przez zaokrąglone lub rozgałęzione komórki z potężnymi błonami. W ciele rośliny można je znaleźć pojedynczo (komórki podporowe) (ryc. 17) lub w grupach (ryc. 18). Należy zauważyć, że właściwości mechaniczne silnie zależą od lokalizacji sklereidów. Część sklereidów tworzy ciągłe warstwy, jak na przykład w łupinach orzechów lub w pestkach owoców (owoce pestkowe).
tkanki wydalnicze
Każdy żywa komórka początkowo ma funkcję usuwania substancji, podczas gdy transport może przebiegać zarówno wzdłuż gradientu stężeń, jak i pod gradient. Substancje, które zostały zsyntetyzowane w komórce, mogą zostać usunięte, a następnie wpłyną na aktywność innych komórek (fitohormony, enzymy). Ten proces nazywa się wydzielaniem. W przypadku wydalania substancji będących produktami przemiany materii komórki proces ten nazywamy wydalaniem. Pomimo faktu, że wydalanie substancji w komórkach roślinnych i zwierzęcych ma zasadnicze podobieństwo, rośliny mają szereg cech, które wynikają z fundamentalnych różnic w aktywności życiowej. Poziom reakcji metabolicznych u roślin jest znacznie niższy niż u zwierząt. W związku z tym emitowane są proporcjonalnie mniejsze ilości odpadów. Inną cechą jest to, że samodzielnie syntetyzując prawie wszystkie niezbędne związki organiczne, rośliny nigdy nie tworzą nadmiernych rezerw. Uwolnione substancje mogą służyć jako materiał wyjściowy do innych reakcji (na przykład CO 2 i H 2 O).
Jeśli u zwierząt proces wydalania toksyn wiąże się z ich usuwaniem z organizmu, rośliny mogą tego nie robić, izolując niepotrzebne substancje w żywym protoplastie (usuwanie różnych substancji do wakuoli), w martwych komórkach (większość tkanek roślina wieloletnia składa się z takich komórek), w przestrzeniach międzykomórkowych.
Uwolnione substancje można podzielić na dwie duże grupy. Pierwsza grupa to substancje organiczne syntetyzowane bezpośrednio przez komórkę (enzymy, polisacharydy, ligniny, terpeny, te drugie są pierwiastkami budulcowymi olejki eteryczne i żywica). Druga grupa – substancje, które początkowo dostają się do komórki z zewnątrz za pomocą transportu ksylemu lub łyka (woda, sole mineralne aminokwasy, cukry proste itp.). Wydalany sekret rzadko jest jednorodny i zwykle składa się z mieszaniny, w której jedna substancja ma najwyższe stężenie.
Roślinom, w przeciwieństwie do zwierząt, brakuje kompletu układ wydalniczy. Po całej roślinie rozsiane są jedynie wyspecjalizowane struktury – idioblasty. Tworzące je komórki są mniejsze niż leżące w pobliżu komórki miąższu. Mają gęstą elektronowo cytoplazmę z rozwiniętymi elementami retikulum endoplazmatycznego i kompleksu Golgiego. Najczęściej nie wyrażone centralna wakuola. Między sobą a innymi żywymi komórkami komórki te są połączone licznymi plazmodesmami.
W zależności od lokalizacji tkanki wydalnicze mogą być zewnętrzne i wewnętrzne.
Zewnętrzne tkanki wydalnicze. Włosy gruczołowe są pochodnymi naskórka. Morfologicznie są zmienne – mogą mieć wielokomórkową głowę, być wydłużone, w formie tarczy na nodze itp. (rys. 19). Należą do nich włosy pokrzywy.
Hydatody to struktury, które uwalniają nadmiar wody w warunkach niskiej transpiracji i wysokiej wilgotności – gutacji. Struktury te tworzą grupy bezbarwnych żywych komórek o cienkich ścianach - epitem. Tkanka ta sąsiaduje z wiązką przewodzącą (ryc. 20). Woda jest uwalniana przez specjalne aparaty szparkowe, które różnią się od zwykłych aparatów szparkowych bezruchem i stale otwartą szczeliną. Skład płynu gutacyjnego różni się znacznie od prawie czysta woda do bardzo złożonej mieszaniny substancji.
Nektarniki. Wiele roślin charakteryzuje się wydzielaniem cieczy zawierającej od 7 do 87% mono- i oligosacharydów. Proces ten przeprowadzają specjalne struktury - nektarniki. W zależności od lokalizacji wyróżnia się nektarniki kwiatowe lub kwiatowe (ryc. 21), a także rośliny znajdujące się na łodygach i liściach - nektarniki pozakwiatowe lub pozakwiatowe. Nektarem może być niezmieniony sok z łyka, który jest dostarczany na powierzchnię przez przestrzenie międzykomórkowe i wydalany przez aparaty szparkowe. Bardziej złożone nektary tworzą miąższ gruczołowy, pokryty naskórkiem z włoskami gruczołowymi. Nektar jest wydalany przez komórki naskórka lub włosy gruczołowe. W tym przypadku wydzielany nektar różni się od soku z łyka. Dominuje bowiem glukoza, fruktoza, sacharoza, aw soku z łyka - glukoza. Jony występują w niskich stężeniach. Jony występują w niskich stężeniach. Aby przyciągnąć zapylacze, nektar może zawierać hormony steroidowe które są niezbędne dla owadów.
Gruczoły trawienne są obecne w roślinach owadożernych (rosiczka, nepenthys itp.). W odpowiedzi na dotyk małych bezkręgowców wydzielają wydzielinę zawierającą enzymy hydrolityczne, kwas mrówkowy, masłowy i propionowy. W efekcie biopolimery rozkładają się na monomery, które roślina wykorzystuje na swoje potrzeby.
Gruczoły solne rozwijają się u roślin rosnących na glebach zasolonych. Gruczoły te znajdują się na liściach i w postaci rozpuszczonej nadmiar jonów wydostaje się na powierzchnię. Sól najpierw osadza się na naskórku, a następnie jest zmywana przez deszcz (ryc. 22). Ponadto wiele roślin ma solne włosy na liściach. Każdy włos składa się z dwóch komórek: jedna tworzy głowę, druga - łodygę. Sole gromadzą się w wakuoli komórki głowy, a gdy ich stężenie staje się wysokie, głowa odpada. W jej miejsce rośnie nowa komórka, która również gromadzi sole. Podczas wzrostu liścia głowa może spaść i uformować się kilka razy. Zaletą solonych włosów jest to, że uwalniając sole, tracą mało wody. Zjawisko to jest bardzo ważne dla roślin rosnących na glebach zasolonych, gdzie brakuje słodkiej wody.
Wewnętrzne tkanki wydalnicze. Wewnętrzne struktury wydalnicze, rozproszone po całym ciele rośliny w postaci idioblastów, nie usuwają substancji poza organizm, gromadząc je w sobie. Idioblasty to komórki roślinne różniące się kształtem, strukturą lub zawartością od innych komórek w tej samej tkance. W zależności od pochodzenia rozróżnia się schizogenne (gr. schiso - rozszczepienie) i lizogenne (gr. liza - rozpuszczenie) zbiorniki wydzielanych substancji (ryc. 23).
Naczynia schizogeniczne. Są to mniej lub bardziej rozległe przestrzenie międzykomórkowe wypełnione wydalanymi substancjami. Na przykład przejścia żywiczne z iglaków, aralii, parasoli i innych. Uważa się, że ma żywicę właściwości bakteriobójcze, odstrasza roślinożerców, czyniąc rośliny niejadalnymi dla nich.
Naczynia lizogeniczne powstają w miejscu żywych komórek, które obumierają i ulegają zniszczeniu po nagromadzeniu w nich substancji. Takie formacje można zaobserwować w skórce owoców cytrusowych.
Oprócz głównych typów istnieją formy pośrednie. Dojarki są dwojakiego rodzaju: przegubowe i nieprzegubowe. Te pierwsze powstają z wielu żywych komórek ułożonych w łańcuchy. W miejscu kontaktu muszle ulegają zniszczeniu, protoplasty łączą się, w wyniku czego powstaje pojedyncza sieć. Takie mleczne występują u Compositae. Niesegmentowane komórki mlekowe są utworzone przez olbrzymią komórkę wielojądrzastą. Powstaje we wczesnych stadiach rozwoju, wydłuża się i rozgałęzia w miarę wzrostu rośliny, tworząc złożoną sieć, na przykład u roślin wilczomlecza (ryc. 25).
Ściany komórkowe lactifers są silnie nawodnione. Nie zdrewniały i dlatego są plastyczne. Centralną część lactis zajmuje wakuola z lateksem, a protoplast zajmuje pozycję przyścienną. Nie zawsze można określić granicę między protoplastem a wakuolą. Laktyfery penetrują merystem i rosną razem z organem rośliny.
Tkanki przewodzące
Ten typ odnosi się do złożonych tkanek, składa się z różnie zróżnicowanych komórek. Oprócz właściwych elementów przewodzących tkanka zawiera elementy mechaniczne, wydalnicze i magazynujące (ryc. 26). Tkanki przewodzące łączą wszystkie narządy roślin w jeden system. Istnieją dwa rodzaje tkanek przewodzących: ksylem i łyko (gr. xylon - drzewo; phloios - kora, łyk). Mają zarówno różnice strukturalne, jak i funkcjonalne.
Elementy przewodzące ksylemu są utworzone przez martwe komórki. Przenoszą dalekosiężny transport wody i rozpuszczonych w niej substancji od korzenia do liści. Przewodzące elementy łyka utrzymują protoplast przy życiu. Przenoszą transport na duże odległości od liści fotosyntetyzujących do korzenia.
Zazwyczaj ksylem i łyko znajdują się w ciele rośliny pewien porządek, tworząc warstwy lub wiązki przewodzące. W zależności od struktury wyróżnia się kilka rodzajów wiązek przewodzących, które są charakterystyczne pewne grupy rośliny. W bocznej wiązce otwartej między ksylemem a łykiem znajduje się kambium, które zapewnia wzrost wtórny (ryc. 27-A, 28). W wiązce otwartej obubocznej łyko znajduje się po obu stronach ksylemu (ryc. 27-B, 29). Zamknięte pęczki nie zawierają kambium, a zatem nie są zdolne do wtórnego pogrubienia (ryc. 27-B, 27-D, 30.31). Można znaleźć jeszcze dwa rodzaje koncentrycznych wiązek, w których albo łyko otacza ksylem (ryc. 27-E, 32), albo ksylem otacza łyko (ryc. 27-E).
Ksylem (drewno). Rozwój ksylemu w roślinach wyższych wiąże się z zapewnieniem wymiany wody. Ponieważ woda jest stale wydalana przez naskórek, taka sama ilość wilgoci musi zostać wchłonięta przez roślinę i dodana do narządów przeprowadzających transpirację. Należy wziąć pod uwagę, że obecność żywego protoplastu w komórkach przewodzących wodę znacznie spowolniłaby transport, a martwe komórki są tutaj bardziej funkcjonalne. Jednak martwa komórka nie ma jędrności, więc membrana musi mieć właściwości mechaniczne. Uwaga: turgescencja to stan komórek, tkanek i narządów roślinnych, w którym stają się one elastyczne w wyniku nacisku zawartości komórek na ich elastyczne błony. Rzeczywiście, elementy przewodzące ksylemu składają się z martwych komórek rozciągniętych wzdłuż osi narządu z grubymi zdrewniałymi muszlami.
Początkowo ksylem tworzy się z merystemu pierwotnego – prokambium, znajdującego się na wierzchołkach narządów osiowych. Najpierw różnicuje się protoksylem, a następnie metaksylem. Znane są trzy rodzaje tworzenia ksylemów. W typie egzarchy elementy protoksylemu pojawiają się najpierw na obrzeżach wiązki prokambalnej, następnie elementy metaksylemu pojawiają się w centrum. Jeśli proces przebiega w przeciwnym kierunku (tj. od centrum do peryferii), to jest to typ endarchiczny. W typie mezarchicznym ksylem jest układany w centrum pęczka prokambalnego, po czym jest odkładany zarówno w kierunku centrum, jak i na obrzeżach.
Korzeń charakteryzuje się tworzeniem ksylemu typu egzarchowego, podczas gdy łodygi mają typ endarchiczny. W roślinach słabo zorganizowanych metody tworzenia ksylemów są bardzo zróżnicowane i mogą służyć jako charakterystyka systematyczna.
W niektórych roślinach (na przykład jednoliściennych) wszystkie komórki prokambalne różnicują się w tkanki przewodzące, które nie są zdolne do wtórnego zagęszczania. W innych formach (na przykład zdrewniałych) merystemy boczne (kambium) pozostają między ksylemem a łykiem. Komórki te są w stanie dzielić się, odnawiając ksylem i łyko. Ten proces nazywa się wzrostem wtórnym. Wiele roślin rosnących w stosunkowo stabilnych warunkach klimatycznych stale rośnie. W formach dostosowanych do sezonowych zmian klimatu – okresowo. W rezultacie powstają dobrze zaznaczone słoje wzrostu rocznego.
Główne etapy różnicowania komórek prokambalnych. Jego komórki mają cienkie błony, które nie zapobiegają ich rozciąganiu podczas wzrostu narządu. Następnie protoplast zaczyna układać błonę wtórną. Ale ten proces ma wyraźne cechy. Błona wtórna nie jest osadzana w ciągłej warstwie, która uniemożliwiałaby rozciąganie się komórki, ale w postaci pierścieni lub spirali. Wydłużanie komórek nie jest trudne. W młodych komórkach pierścienie lub zwoje helisy znajdują się blisko siebie. W dojrzałych komórkach rozchodzą się one w wyniku rozciągania komórek (ryc. 33). Pierścieniowe i spiralne zgrubienia muszli nie zapobiegają wzrostowi, jednak mechanicznie ustępują muszlom, gdzie wtórne zgrubienia tworzą ciągłą warstwę. Dlatego po ustaniu wzrostu w ksylemie powstają elementy z ciągłą zdrewniałą skorupą (metaksylem). Należy zauważyć, że wtórne zgrubienie nie jest tu pierścieniowe ani spiralne, ale punktowe, schodkowe, siatkowate (ryc. 34). Jego komórki nie są w stanie się rozciągać i umierają w ciągu kilku godzin. Proces ten w pobliskich komórkach zachodzi w sposób skoordynowany. W cytoplazmie pojawia się duża liczba lizosomów. Następnie rozkładają się, a zawarte w nich enzymy niszczą protoplast. Po zniszczeniu ścian poprzecznych komórki ułożone w łańcuch jedna nad drugą tworzą wydrążone naczynie (ryc. 35). Większość okrytozalążkowe a niektóre paprocie mają naczynia.
Komórka przewodząca, która nie tworzy się przez perforacje w jej ścianie, nazywa się tchawicą. Ruch wody przez cewki jest wolniejszy niż przez naczynia. Faktem jest, że w tracheidach pierwotna błona nie jest nigdzie przerwana. Tracheidy komunikują się ze sobą przez pory. Należy wyjaśnić, że w roślinach pory są tylko zagłębieniami w błonie wtórnej aż do błony pierwotnej i nie ma perforacji przelotowych między tracheidami.
Najczęściej są to pory obramowane (ryc. 35-1). Ich kanał, zwrócony w stronę wnęki komórki, tworzy przedłużenie - komorę porową. Pory większości roślin iglastych na pierwotnej skorupie mają zgrubienie - torus, który jest rodzajem zaworu i jest w stanie regulować intensywność transportu wody. Przesuwając się, torus blokuje przepływ wody przez pory, ale po tym nie może już powrócić do poprzedniej pozycji, wykonując jednorazową akcję.
Pory są mniej lub bardziej zaokrąglone, wydłużone prostopadle do wydłużonej osi (grupa tych porów przypomina drabinę, więc ta porowatość nazywana jest porowatością drabinkową). Transport przez pory odbywa się zarówno w kierunku wzdłużnym, jak i poprzecznym. Pory są obecne nie tylko w tracheidach, ale także w poszczególnych komórkach naczyniowych tworzących naczynie.
Z punktu widzenia teorii ewolucji tracheidy są pierwszą i główną strukturą przewodzącą wodę w ciele roślin wyższych. Uważa się, że naczynia powstały z tchawicy w wyniku lizy ścian poprzecznych między nimi (ryc. 36). Większość paproci i nagonasiennych nie ma. Poruszają wodę przez tracheidy.
W trakcie rozwoju ewolucyjnego naczynia powstały w różne grupy rośliny wielokrotnie, ale najważniejszego znaczenia funkcjonalnego nabyły u okrytonasiennych, które mają je wraz z tracheidami. Uważa się, że posiadanie bardziej zaawansowanego mechanizmu transportu pomogło im nie tylko przetrwać, ale także osiągnąć znaczną różnorodność form.
Xylem jest złożoną tkanką, oprócz elementów przewodzących wodę zawiera inne. Funkcje mechaniczne pełnią włókna libriform (łac. liber - łyk, forma - forma). Obecność dodatkowych konstrukcji mechanicznych jest o tyle ważna, że pomimo pogrubienia ścianki elementów przewodzących wodę są wciąż zbyt cienkie. Nie są w stanie się utrzymać duża masa roślina wieloletnia. Włókna rozwinęły się z cewek. Charakteryzują się mniejszymi rozmiarami, zdrewniałymi (zdrewniałymi) muszlami i wąskimi jamami. Na ścianie można znaleźć pory pozbawione obramowania. Włókna te nie mogą przewodzić wody, ich główną funkcją jest wsparcie.
Xylem zawiera również żywe komórki. Ich masa może osiągnąć 25% całkowitej objętości drewna. Ponieważ komórki te mają zaokrąglony kształt, nazywane są miąższem drzewnym. W ciele rośliny miąższ znajduje się na dwa sposoby. W pierwszym przypadku komórki układają się w pionowe sznury - jest to miąższ sznurka. W innym przypadku miąższ tworzy promienie poziome. Nazywa się je promieniami rdzenia, ponieważ łączą rdzeń i korę. Rdzeń pełni szereg funkcji, w tym magazynowanie substancji.
Łyko (łyk). Jest to złożona tkanka, ponieważ tworzą ją heterogenne komórki. Główne komórki przewodzące nazywane są elementami sitowymi (ryc. 37). Elementy przewodzące ksylemu tworzą martwe komórki, podczas gdy w łyku zachowują żywy, choć znacznie zmieniony, protoplast w okresie funkcjonowania. Łyko powoduje odpływ substancji plastycznych z organów fotosyntetyzujących. Wszystkie żywe komórki roślin posiadają zdolność przewodzenia substancji organicznych. I stąd, jeśli ksylem występuje tylko w roślinach wyższych, to transport substancji organicznych między komórkami odbywa się również w niższe rośliny.
Ksylem i łyko rozwijają się z merystemów wierzchołkowych. W pierwszym etapie protofloem tworzy się w rdzeniu prokambalnym. W miarę wzrostu otaczających tkanek rozciąga się, a po zakończeniu wzrostu zamiast protofloemu powstaje metafloem.
Na różne grupy W roślinach wyższych można spotkać dwa rodzaje elementów sitowych. U paproci i nagonasiennych jest reprezentowany przez komórki sitowe. Pola sitowe w komórkach są rozrzucone wzdłuż ścian bocznych. Protoplast zachowuje lekko zniszczone jądro.
U roślin okrytonasiennych elementy sitowe nazywane są rurkami sitowymi. Komunikują się ze sobą za pośrednictwem płyt sitowych. Dojrzałe komórki nie mają jąder. Natomiast obok rurki sitowej znajduje się komórka towarzysząca, która powstaje razem z rurką sitową w wyniku podziału mitotycznego wspólnej komórki macierzystej (ryc. 38). Komórka towarzysząca ma gęstszą cytoplazmę z dużą liczbą aktywnych mitochondriów, a także w pełni funkcjonujące jądro, świetna ilość plazmodesmy (dziesięć razy więcej niż inne komórki). Komórki towarzyszące wpływają na aktywność funkcjonalną niejądrowych komórek rurek sitowych.
Struktura dojrzałych komórek sitowych ma pewne cechy szczególne. Nie ma wakuoli, więc cytoplazma jest silnie upłynniona. Jądro może być nieobecne (u roślin okrytonasiennych) lub być w stanie pomarszczonym, funkcjonalnie nieaktywnym. Rybosomy i kompleks Golgiego są również nieobecne, ale retikulum endoplazmatyczne jest dobrze rozwinięte, które nie tylko penetruje cytoplazmę, ale także przechodzi do sąsiednich komórek przez pory pól sitowych. Dobrze rozwinięte mitochondria i plastydy występują w obfitości.
Pomiędzy komórkami transport substancji odbywa się przez otwory znajdujące się w błonach komórkowych. Takie otwory nazywane są porami, ale w przeciwieństwie do porów tchawicy są przelotowe. Przypuszcza się, że są to wysoce ekspandowane plazmodesmy, na ściankach których osadza się kalozowy polisacharyd. Pory są ułożone w grupy, tworząc pola sitowe. W formach prymitywnych pola sitowe są losowo rozrzucone na całej powierzchni skorupy, u bardziej zaawansowanych okrytonasiennych znajdują się na końcach sąsiadujących ze sobą komórek, tworząc płytkę sitową (ryc. 39). Jeśli jest na nim jedno pole sitowe, nazywa się to prostym, jeśli jest ich kilka - złożonym.
Szybkość ruchu roztworów przez elementy sitowe wynosi do 150 cm ∕ godz. To tysiąc razy więcej niż tempo swobodnej dyfuzji. Prawdopodobnie zachodzi transport aktywny, a liczne mitochondria elementów sitowych i komórki satelitarne dostarczają niezbędnego do tego ATP.
Żywotność elementów sitowych łyka zależy od obecności merystemów bocznych. Jeśli są obecne, elementy sitowe działają przez cały okres eksploatacji rośliny.
Oprócz elementów sitowych i komórek satelitarnych łyko zawiera włókna łykowe, sklereidy i miąższ.
Pytania do kontroli i powtórzenia
Oznaki roślin wyższych? Jakie pokolenie roślin wyższych nazywa się gametofitem? Sporofit?
Jakie są przyczyny, które przyczyniają się do rozwoju zróżnicowanych tkanek u roślin lądowych.
Wypisz rodzaje roślin.
Jakie znasz grupy merystemów? Gdzie występują w ciele rośliny?
Tkanki powłokowe, ich funkcje, znaczenie.
Organizacja różne rodzaje pokrywające tkanki.
Budowa aparatu szparkowego. Wymień znane ci rodzaje aparatów szparkowych. Jak działają aparaty szparkowe?
Cechy tkanek miąższowych, rodzaje miąższu.
Oznaki miąższu i sklerenchymy.
Wymień rodzaje struktur wydalniczych rośliny. Jakie jest ich znaczenie?
Metody formowania zbiorników na wydalane substancje.
Co to jest dojarzami? Na jakie typy są podzielone?
Jak zorganizowane są tkanki przewodzące?
Co to są wiązki przewodzące? Ich typy.
Kierunki przemieszczania się substancji przez ksylem? Łyko? Czy skład transportowanych substancji różni się w ksylemie i łyku?
Budowa i funkcje ksylemu, łyka.
Organizacja elementów przewodzących łyka u roślin z różnych gatunków grupy systematyczne.
W jaki sposób transportowane są substancje w łyku?
Związane z nimi dodatkowe białka, tzw. białka MAP. Białka te, stabilizując mikrotubule, przyspieszają proces polimeryzacji tubuliny. Struktura i funkcja mikrofilamentów Mikrofilamenty to bardzo cienkie i długie nitkowate struktury białkowe występujące w cytoplazmie. Pod błoną plazmatyczną mikrofilamenty tworzą ciągły splot, tworząc ...

Jak na przykład koperek, marchew, rzepa itp., w większości przypadków ma ten sam rodzaj tkanki, który właśnie zaznaczyłem w korku. W ten sposób po raz pierwszy odkryto komórkę roślinną. Ale Hooke przez długi czas nie mógł poradzić sobie ze swoim mikroskopem: pomysły innych wynalazków roiły się w jego głowie (zegary wiosenne, ulepszone kompasy itp.), I chętnie przekazał dalsze utrzymanie mikroskopijnych ...
Przynajmniej trochę pojęcia o odpowiednich metodach badań komórkowych. Mikroskopia świetlna Najstarszą i jednocześnie najpowszechniejszą metodą badania komórek jest mikroskopia. Można powiedzieć, że początek badań nad komórką położyło wynalezienie optycznego mikroskopu świetlnego. Bezbronny ludzkie oko ma rozdzielczość około 1/10 mm. Ten...
Dzięki wielu organelli komórkowych zapewnia połączenie między poszczególnymi strukturami wewnątrzkomórkowymi. Lizosomy Lizosomy (z lis- i gr. soma - ciało), struktury w komórkach organizmów zwierzęcych i roślinnych zawierające enzymy zdolne do rozkładania (tj. lizowania - stąd nazwa) białek, polisacharydów, peptydów, kwasów nukleinowych. Jest to bardzo różnorodna klasa bąbelków o wielkości 0,1-0,4 mikrona, ...
Struktura komórki roślinnej. Tkanki roślinne
STRUKTURA KOMÓRKI ROŚLINNEJ
Komórka roślinna składa się z mniej lub bardziej sztywnej ściany komórkowej i protoplastu. Ściana komórkowa to ściana komórkowa i błona cytoplazmatyczna. Termin protoplast pochodzi od słowa protoplazma, które od dawna jest używane w odniesieniu do wszystkich żywych istot. Protoplast jest protoplazmą pojedynczej komórki.
Protoplast składa się z cytoplazmy i jądra. Cytoplazma zawiera organelle (rybosomy, mikrotubule, plastydy, mitochondria) i układy membranowe (retikulum endoplazmatyczne, dictyosomy). Cytoplazma obejmuje również macierz cytoplazmatyczną (substancję podstawową), w której zanurzone są organelle i układy błon. Cytoplazma jest oddzielona od ściany komórkowej błoną plazmatyczną, która jest błoną elementarną. W przeciwieństwie do większości komórek zwierzęcych, komórki roślinne zawierają jedną lub więcej wakuoli. Są to pęcherzyki wypełnione cieczą i otoczone elementarną błoną (tonoplastem).
W żywej komórce roślinnej substancja podstawowa jest w ciągłym ruchu. Organelle biorą udział w ruchu, zwanym przepływem cytoplazmy lub cyklozą. Cykloza ułatwia przemieszczanie się substancji w komórce i ich wymianę między komórką a środowiskiem.
Membrana plazmowa. Jest to dwuwarstwowa struktura fosfolipidowa. Komórki roślinne charakteryzują się inwazjami błony plazmatycznej.
Błona plazmatyczna pełni następujące funkcje:
Uczestniczy w wymianie substancji między komórką a środowiskiem;
Koordynuje syntezę i montaż mikrofibryli celulozowych ściany komórkowej;
Przekazuje sygnały hormonalne i zewnętrzne, które kontrolują wzrost i różnicowanie komórek.
Rdzeń. Jest to najbardziej widoczna struktura w cytoplazmie komórki eukariotycznej. Jądro spełnia dwie ważne funkcje:
Kontroluje aktywność życiową komórki, określając, które białka iw jakim czasie powinny zostać zsyntetyzowane;
Przechowuje informacje genetyczne i przekazuje je komórkom potomnym podczas podziału komórki.
Jądro komórki eukariotycznej jest otoczone dwiema podstawowymi błonami, które tworzą otoczkę jądrową. Przesiąknięta jest licznymi porami o średnicy od 30 do 100 nm, widocznymi jedynie pod mikroskopem elektronowym. Pory mają złożoną strukturę. Zewnętrzna błona błony jądrowej w niektórych miejscach jest połączona z retikulum endoplazmatycznym. Otoczkę jądrową można postrzegać jako wyspecjalizowaną, lokalnie zróżnicowaną część retikulum endoplazmatycznego (ER).
W jądrze zabarwionym specjalnymi barwnikami można wyróżnić cienkie włókna i grudki chromatyny i nukleoplazmy (głównej substancji jądra). Chromatyna składa się z DNA związanego ze specjalnymi białkami zwanymi histonami. W procesie podziału komórki chromatyna staje się coraz bardziej zagęszczona i składa się w chromosomy. DNA koduje informację genetyczną.
Organizmy różnią się liczbą chromosomów w komórkach somatycznych. Na przykład kapusta ma - 20 chromosomów; słonecznik - 34; pszenica - 42; ludzi - 46, a jeden z gatunków paproci Ophioglossum - 1250. Komórki płciowe (gamety) mają tylko połowę liczby chromosomów charakterystycznych dla komórek somatycznych organizmu. Liczba chromosomów w gametach nazywana jest haploidalną (pojedynczą), w komórkach somatycznych - diploidalną (podwójną). Komórki, które mają więcej niż dwa zestawy chromosomów, nazywane są poliploidalnymi.
Pod mikroskopem świetlnym można zobaczyć kuliste struktury - jąderka. Każde jądro ma jedno lub więcej jąderek, które są widoczne w jądrach niedzielących się. Rybosomalny RNA jest syntetyzowany w jąderku. Zazwyczaj jądra organizmów diploidalnych mają dwa jąderka, po jednym dla każdego haploidalnego zestawu chromosomów. Jąderka nie mają własnej błony. Pod względem biochemicznym jąderka charakteryzują się wysokim stężeniem RNA, które jest tu związane z fosfoproteinami. Wielkość jąderka zależy od stanu funkcjonalnego komórki. zauważono, że w szybko rosnącej komórce, w której zachodzą intensywne procesy syntezy białek, jąderka powiększają się. W jąderkach powstają mRNA i rybosomy, które pełnią funkcję syntetyczną tylko w jądrze.
Nukleoplazma (karioplazma) jest reprezentowana przez jednorodną ciecz, w której rozpuszczają się różne białka, w tym enzymy.
Plastydy. Charakterystycznymi składnikami komórek roślinnych są wakuole, celulozowe ściany komórkowe i plastydy. Każdy plastyd ma swoją własną powłokę, składającą się z dwóch elementarnych membran. Wewnątrz plastydu wyróżnia się system membranowy i w różnym stopniu jednorodną substancję, zrąb. Dojrzałe plastydy są klasyfikowane na podstawie zawartych w nich pigmentów.
Chloroplasty, w których zachodzi fotosynteza, zawierają chlorofile i karotenoidy. Zwykle mają postać krążka o średnicy 4 - 5 mikronów. W jednej komórce mezofilu (środek liścia) może znajdować się 40 - 50 chloroplastów; w mm2 liścia - około 500 000. w cytoplazmie chloroplasty są zwykle zlokalizowane równolegle do błony komórkowej.
Wewnętrzna struktura chloroplastu jest złożona. Zrąb jest przesiąknięty rozwiniętym systemem błon w postaci pęcherzyków - tylakoidów. Każdy tylakoid składa się z dwóch błon. Tylakoidy tworzą jeden system. Z reguły są one zbierane w stosy - ziarna, przypominające kolumny monet. Tylakoidy poszczególnych granów są połączone tylakoidami zrębu lub tylakoidami międzyziarnistymi. Chlorofil i karotenoidy są osadzone w błonach tylakoidów. Chloroplasty w zielonych roślinach i algach często zawierają ziarna skrobi i małe kropelki lipidów (tłuszczu). Ziarna skrobi są tymczasowymi magazynami produktów fotosyntezy. Mogą zniknąć z chloroplastów rośliny, która była w ciemności tylko przez 24 godziny, i pojawić się ponownie 3-4 godziny po przeniesieniu rośliny na światło.
Chloroplasty to półautonomiczne organelle i przypominają bakterie. Na przykład rybosomy bakterii i chloroplastów mają dość duże podobieństwo. Są mniejsze niż rybosomy eukariotyczne. Synteza białek na rybosomach bakterii i chloroplastach jest hamowana przez chloramfenikol, który nie ma wpływu na komórki eukariotyczne. Ponadto zarówno bakterie, jak i chloroplasty mają podobny typ nukleoidu, zorganizowanego w podobny sposób. Pomimo faktu, że tworzenie chloroplastów i synteza zawartych w nich pigmentów jest w dużej mierze kontrolowana przez chromosomalny DNA komórki, to jednak przy braku własnego DNA chloroplasty nie tworzą się.
Chloroplasty można uznać za główne organelle komórkowe, ponieważ są pierwszymi w łańcuchu transformacji energii słonecznej, w wyniku czego ludzkość otrzymuje zarówno żywność, jak i paliwo. W chloroplastach zachodzi nie tylko fotosynteza. Biorą również udział w syntezie aminokwasów i kwasów tłuszczowych, służą jako magazyn tymczasowych rezerw skrobi.
Chromoplasty to pigmentowane plastydy. Różnorodne w formie, nie posiadają chlorofilu, ale syntetyzują i gromadzą karotenoidy, które nadają żółty, pomarańczowy, czerwony kolor kwiatom, starym liściom, owocom i korzeniom. chromoplasty mogą rozwijać się z chloroplastów, które w tym przypadku tracą chlorofil i struktury błony wewnętrznej, akumulują karotenoidy. Dzieje się tak podczas dojrzewania wielu owoców. Chromoplasty przyciągają owady i inne zwierzęta, z którymi współewoluowały.
Leukoplasty to niepigmentowane plastydy. Niektóre z nich syntetyzują skrobię (amyloplasty), inne są zdolne do tworzenia różnych substancji, w tym lipidów i białek. W świetle leukoplasty zamieniają się w chloroplasty.
Proplastydy to małe, bezbarwne lub jasnozielone, niezróżnicowane plastydy występujące w merystematycznych (dzielących się) komórkach korzeni i pędów. Są prekursorami innych, bardziej zróżnicowanych plastydów - chloroplastów, chromoplastów i aminoplastów. Jeśli rozwój protoplastów jest opóźniony z powodu braku światła, może pojawić się w nich jedno lub więcej ciałek prolamelarnych, które są półkrystalicznymi skupiskami błon kanalikowych. Plastydy zawierające ciałka prolamelarne nazywane są etioplastami. W świetle etioplasty zamieniają się w chloroplasty, podczas gdy błony ciał prolamelarnych tworzą tylakoidy. Etioplasty powstają w liściach roślin, które są w ciemności. protoplasty zarodków nasiennych najpierw przekształcają się w etioplasty, z których następnie w świetle rozwijają się chloroplasty. Plastydy charakteryzują się stosunkowo łatwymi przejściami z jednego rodzaju do drugiego. Plastydy, podobnie jak bakterie, rozmnażają się, dzieląc się na dwie części. W komórkach merystematycznych czas podziału protoplastów w przybliżeniu pokrywa się z czasem podziału komórki. Jednak w dojrzałych komórkach większość plastydów powstaje w wyniku podziału dojrzałych plastydów.
mitochondria. Podobnie jak chloroplasty, mitochondria są otoczone dwiema podstawowymi błonami. Wewnętrzna błona tworzy liczne fałdy i wypukłości - cristae, które znacznie zwiększają wewnętrzną powierzchnię mitochondriów. Są znacznie mniejsze niż plastydy, mają średnicę około 0,5 µm oraz zmienną długość i kształt.
W mitochondriach zachodzi proces oddychania, w wyniku którego dochodzi do rozpadu cząsteczek organicznych z uwolnieniem energii i przekazaniem jej do cząsteczek ATP, głównej rezerwy energetycznej wszystkich komórek eukariotycznych. Większość komórek roślinnych zawiera setki lub tysiące mitochondriów. Ich liczba w jednej komórce jest określona przez zapotrzebowanie komórki na ATP. Mitochondria są w ciągłym ruchu, przemieszczając się z jednej części komórki do drugiej, łącząc się ze sobą i dzieląc. Mitochondria zwykle gromadzą się tam, gdzie potrzebna jest energia. Jeśli błona plazmatyczna aktywnie przenosi substancje z komórki do komórki, wówczas mitochondria znajdują się wzdłuż powierzchni błony. W ruchliwych glonach jednokomórkowych mitochondria gromadzą się u podstawy wici, dostarczając energię niezbędną do ich ruchu.
Mitochondria, podobnie jak plastydy, są półautonomicznymi organellami zawierającymi składniki niezbędne do syntezy własnych białek. Błona wewnętrzna otacza płynną matrycę, która zawiera białka, RNA, DNA, bakteryjne rybosomy i różne substancje rozpuszczone. DNA istnieje w postaci kolistych cząsteczek zlokalizowanych w jednym lub kilku nukleoidach.
Na podstawie podobieństwa bakterii do mitochondriów i chloroplastów komórek eukariotycznych można przypuszczać, że mitochondria i chloroplasty wywodzą się od bakterii, które znalazły „schronienie” w większych komórkach heterotroficznych – prekursorach eukariontów.
Mikrociała. W przeciwieństwie do plastydów i mitochondriów, które są ograniczone dwiema błonami, mikrociała są kulistymi organellami otoczonymi pojedynczą błoną. Mikrociała mają ziarnistą (granulowaną) zawartość, czasami zawierają również krystaliczne wtrącenia białkowe. Mikrociała są związane z jednym lub dwoma regionami retikulum endoplazmatycznego.
Niektóre mikrociała, zwane proksysomami, odgrywają ważną rolę w metabolizmie kwasu glikolowego, który jest bezpośrednio związany z fotooddychaniem. W zielonych liściach są one związane z mitochondriami i chloroplastami. Inne mikrociała, zwane glioksysomami, zawierają enzymy potrzebne do przekształcania tłuszczów w węglowodany. Dzieje się tak w przypadku wielu nasion podczas kiełkowania.
wakuole - Są to obszary komórki ograniczone błoną, wypełnione płynem - sokiem komórkowym. Otoczone są tonoplastem (błoną wakuolową).
Młoda komórka roślinna zawiera liczne małe wakuole, które w miarę starzenia się łączą się w jedną dużą wakuolę. W dojrzałej komórce do 90% jej objętości może zajmować wakuola. W tym przypadku cytoplazma jest dociskana w postaci cienkiej warstwy obwodowej do błony komórkowej. Wzrost wielkości komórek wynika głównie ze wzrostu wakuoli. W rezultacie powstaje ciśnienie turgoru i zachowana jest elastyczność tkanki. Jest to jedna z głównych funkcji wakuoli i tonoplastu.
Głównym składnikiem soku jest woda, reszta zależy od rodzaju rośliny i jej stanu fizjologicznego. Wakuole zawierają sole, cukry, rzadziej białka. Tonoplast odgrywa aktywną rolę w transporcie i gromadzeniu się niektórych jonów w wakuoli. Stężenie jonów w soku komórkowym może znacznie przekraczać jego stężenie w środowisku. Przy wysokiej zawartości niektórych substancji w wakuolach tworzą się kryształy. Najczęściej spotykane są kryształy szczawianu wapnia, które mają inny kształt.
Wakuole są miejscami gromadzenia się produktów przemiany materii (metabolizmu). Mogą to być białka, kwasy, a nawet substancje toksyczne dla człowieka (alkaloidy). Często osadzają się pigmenty. Niebieski, fioletowy, purpurowy, ciemnoczerwony, karmazynowy dają komórkom roślinnym pigmenty z grupy antocyjanów. W przeciwieństwie do innych pigmentów dobrze rozpuszczają się w wodzie i znajdują się w soku komórkowym. Decydują o czerwonym i niebieskim zabarwieniu wielu warzyw (rzodkiewki, rzepa, kapusta), owoców (winogrona, śliwki, wiśnie), kwiatów (chabry, pelargonie, ostróżki, róże, piwonie). Czasami te pigmenty maskują chlorofil w liściach, na przykład w ozdobnym czerwonym klonie. Antocyjany barwią jesienne liście na jaskrawoczerwony. Powstają w chłodne słoneczne dni, kiedy synteza chlorofilu zatrzymuje się w liściach. W liściach, gdy nie tworzą się antocyjany, po zniszczeniu chlorofilu zauważalne stają się żółto-pomarańczowe karotenoidy chloroplastów. Liście są najjaśniejsze w chłodnej, jasnej jesieni.
Wakuole biorą udział w niszczeniu makrocząsteczek, w krążeniu ich składników w komórce. Rybosomy, mitochondria, plastydy wpadające do wakuoli ulegają zniszczeniu. Zgodnie z tą czynnością trawienną można je porównać z lizosomami - organellami komórek zwierzęcych.
Wakuole powstają z retikulum endoplazmatycznego (retikulum)
Rybosomy. Małe cząsteczki (17 - 23nm), składające się z mniej więcej równych ilości białka i RNA. W rybosomach aminokwasy łączą się, tworząc białka. W komórkach o aktywnym metabolizmie jest ich więcej. Rybosomy znajdują się w cytoplazmie komórki swobodnie lub przyczepione do retikulum endoplazmatycznego (80S). Występują również w jądrze (80S), mitochondriach (70S), plastydach (70S).
Rybosomy mogą tworzyć kompleks, na którym zachodzi jednoczesna synteza identycznych polipeptydów, o których informacja pochodzi z jednej cząsteczki i RNA. Taki kompleks nazywa się polirybosomami (polisomami). Komórki, które syntetyzują białka w dużych ilościach, mają rozbudowany system polisomów, które często są przyczepione do zewnętrznej powierzchni otoczki jądrowej.
Retikulum endoplazmatyczne. Jest to złożony trójwymiarowy system membranowy o nieokreślonym zasięgu. W kontekście ER wygląda jak dwie elementarne membrany z wąską przezroczystą przestrzenią między nimi. Kształt i długość ER zależy od rodzaju komórki, jej aktywności metabolicznej oraz stopnia zróżnicowania. W komórkach, które wydzielają lub przechowują białka, ER ma kształt płaskiego worka lub cysterny, z licznymi rybosomami przyczepionymi do jego zewnętrznej powierzchni. Ta siateczka nazywana jest szorstką retikulum endoplazmatycznym. Gładki ER ma zwykle kształt cylindryczny. W tej samej komórce może znajdować się szorstka i gładka siateczka śródplazmatyczna. Z reguły istnieje między nimi wiele połączeń.
Retikulum endoplazmatyczne działa jako system komunikacyjny komórki. Jest związany z zewnętrzną powłoką jądra. W rzeczywistości te dwie struktury tworzą jeden system membranowy. Kiedy otoczka jądrowa pęka podczas podziału komórki, jej fragmenty przypominają fragmenty ER. Retikulum endoplazmatyczne to system transportu substancji: białek, lipidów, węglowodanów do różnych części komórki. retikulum endoplazmatyczne sąsiednich komórek są połączone przez nici cytoplazmatyczne - plazmodesmy - które przechodzą przez błony komórkowe.
Retikulum endoplazmatyczne jest głównym miejscem syntezy błon komórkowych. W niektórych komórkach roślinnych tworzą się tu błony wakuoli i mikrociał, cysterny dictyosomów.
Aparat Golgiego. Termin ten jest używany w odniesieniu do wszystkich dictyosomów lub ciał Golgiego w komórce. Dictyosomy to grupy płaskich pęcherzyków w kształcie dysku lub cystern, które rozgałęziają się wzdłuż krawędzi w złożony system kanalików. Dictyosomy u roślin wyższych składają się z 4 - 8 cystern, zebranych razem.
Zwykle w paczce zbiorników wyróżnia się stronę rozwijającą się i dojrzewającą. błony tworzących się cystern przypominają strukturą błony ER, a błony dojrzewających cystern przypominają błonę komórkową.
Dictyosomy biorą udział w wydzielaniu, a u większości roślin wyższych w tworzeniu błon komórkowych. Polisacharydy ściany komórkowej syntetyzowane przez dictyosomy gromadzą się w pęcherzykach, które następnie oddzielają się od dojrzewających cystern. Te pęcherzyki wydzielnicze migrują i łączą się z CMP; natomiast zawarte w nich polisacharydy wbudowują się w błonę komórkową. Niektóre substancje gromadzące się w dictyosomach powstają w innych strukturach, na przykład w ER, a następnie są transportowane do dictyosomów, gdzie są modyfikowane (modyfikowane) przed sekrecją. Na przykład glikoproteiny są ważnym materiałem budulcowym błony komórkowej. Część białkowa jest syntetyzowana przez polisomy szorstkiego ER, część węglowodanowa jest syntetyzowana w dictyosomach, gdzie obie części łączą się tworząc glikoproteiny.
Membrany to dynamiczne, ruchome struktury, które nieustannie zmieniają swój kształt i powierzchnię. Koncepcja układu endoplazmatycznego opiera się na ruchliwości błon. Zgodnie z tą koncepcją błony wewnętrzne cytoplazmy, z wyjątkiem błon mitochondriów i plastydów, stanowią jedną całość i wywodzą się z retikulum endoplazmatycznego. Nowe cysterny dictyosomów tworzą się z retikulum endoplazmatycznego przez etap pęcherzyków pośrednich, a pęcherzyki wydzielnicze, które oddzielają się od dictyosomów, ostatecznie przyczyniają się do tworzenia błony plazmatycznej. Zatem retikulum endoplazmatyczne i dictyosomy tworzą jednostkę funkcjonalną, w której dictyosomy pełnią rolę struktur pośrednich w procesie przekształcania błon, takich jak retikulum endoplazmatyczne, w błony takie jak plazma. W tkankach, których komórki słabo rosną i dzielą się, składniki błony są stale aktualizowane.
mikrotubule występuje w prawie wszystkich komórkach eukariotycznych. Są to cylindryczne struktury o średnicy około 24 nm. Ich długość jest różna. Każdy kanalik składa się z podjednostek białka zwanego tubuliną. Podjednostki tworzą 13 podłużnych włókien otaczających centralną jamę. Mikrotubule są strukturami dynamicznymi, są regularnie niszczone i formowane na określonych etapach cyklu komórkowego. Ich gromadzenie odbywa się w specjalnych miejscach, które nazywane są centrami organizacji mikrotubul. W komórkach roślinnych mają słabo wyrażoną amorficzną strukturę.
Funkcje mikrotubul: uczestniczą w tworzeniu błony komórkowej; kierują pęcherzyki dictyosomów do tworzącej się błony, jak nici wrzeciona, które tworzą się w dzielącej się komórce; odgrywają rolę w tworzeniu płytki komórkowej (początkowa granica między komórkami potomnymi). Ponadto mikrotubule są ważnym składnikiem wici i rzęsek, w których ruchu odgrywają ważną rolę.
mikrofilamenty, podobnie jak mikrotubule, znajdują się w prawie wszystkich komórkach eukariotycznych. Są to długie włókna o grubości 5–7 nm, składające się z kurczliwego białka aktyny. Pęczki mikrofilamentów znajdują się w wielu komórkach roślin wyższych. Najwyraźniej odgrywają ważną rolę w prądach cytoplazmy. Mikrofilamenty wraz z mikrotubulami tworzą elastyczną sieć zwaną cytoszkieletem.
Substancja podstawowa przez długi czas był uważany za jednorodny (homogeniczny) bogaty w białko roztwór o niewielkiej liczbie struktur lub nawet bez struktury. Jednak obecnie, przy użyciu wysokonapięciowego mikroskopu elektronowego, ustalono, że substancja podstawowa jest trójwymiarową siatką zbudowaną z cienkich (o średnicy 3–6 nm) pasm wypełniających całą komórkę. Inne składniki cytoplazmy, w tym mikrotubule i mikrofilamenty, są zawieszone w tej siatce mikrotrabekularnej.
Struktura mikrotrabekularna to sieć pasm białkowych, między którymi przestrzeń jest wypełniona wodą. Wraz z wodą siatka ma konsystencję żelu, który wygląda jak galaretowate ciała.
Organelle są przyczepione do siatki mikrotrabekularnej. Sieć zapewnia komunikację pomiędzy poszczególnymi częściami komórki oraz kieruje transportem wewnątrzkomórkowym.
Krople lipidowe- struktury o kulistym kształcie, nadające ziarnistość cytoplazmie komórki roślinnej pod mikroskopem świetlnym. Na mikrografach elektronowych wyglądają na amorficzne. Bardzo podobne, ale mniejsze kropelki znajdują się w plastydach.
Kropelki lipidów, myląc je z organellami, nazywały je sferosomami i uważały, że otoczone są jednowarstwową lub dwuwarstwową błoną. Jednak ostatnie dane pokazują, że kropelki lipidów nie mają błon, ale mogą być pokryte białkiem.
Substancje ergastyczne - są to „produkty pasywne” protoplastu: substancje rezerwowe lub produkty odpadowe. Mogą pojawiać się i znikać w różnych okresach cyklu komórkowego. Z wyjątkiem ziaren skrobi, kryształów, barwników antocyjanowych i kropelek lipidów. Należą do nich żywice, gumy, garbniki i białka. Substancje ergastyczne są częścią błony komórkowej, głównej substancji cytoplazmy i organelli, w tym wakuoli.
Wici i rzęski - są to cienkie, przypominające włosy struktury, które wystają z powierzchni wielu komórek eukariotycznych. Mają stałą średnicę, ale długość waha się od 2 do 150 mikronów. Konwencjonalnie dłuższe i nieliczne z nich nazywane są wiciami, a krótsze i liczniejsze nazywane są rzęskami. Nie ma wyraźnych różnic między tymi dwoma typami struktur, więc termin wici jest używany w odniesieniu do obu.
U niektórych alg i grzybów wici są narządami ruchu, za pomocą których poruszają się w wodzie. W roślinach (np. mchach, wątrobowcach, paprociach, niektórych nagonasiennych) tylko komórki rozrodcze (gamety) mają wici.
Każda wici ma określoną organizację. Zewnętrzny pierścień 9 par mikrotubul otacza dwie dodatkowe mikrotubule znajdujące się w środku wici. Zawierające enzym „uchwyty” rozciągają się od jednej mikrotubuli każdej z zewnętrznych par. Jest to podstawowy wzorzec organizacji 9+2 występujący we wszystkich wici organizmów eukariotycznych. Uważa się, że ruch wici opiera się na przesuwaniu się mikrotubul, podczas gdy zewnętrzne pary mikrotubul poruszają się jedna po drugiej bez skurczu. Przesuwanie się par mikrotubul względem siebie powoduje miejscowe wyginanie wici.
Wici „wyrastają” z cytoplazmatycznych cylindrycznych struktur zwanych ciałami podstawowymi, które również tworzą podstawową część wici. Ciała podstawowe mają wewnętrzną strukturę przypominającą wici, z wyjątkiem tego, że kanaliki zewnętrzne są ułożone raczej w trojaczki niż w pary, a kanalików środkowych nie ma.
Ściana komórkowa.Ściana komórkowa ogranicza rozmiar protoplastu i zapobiega jego pęknięciu w wyniku absorpcji wody przez wakuolę.
Ściana komórkowa pełni określone funkcje, które są ważne nie tylko dla komórki i tkanki, w której komórka się znajduje, ale także dla całej rośliny. Ściany komórkowe odgrywają zasadniczą rolę we wchłanianiu, transporcie i wydalaniu substancji, a ponadto może się w nich koncentrować aktywność lizosomalna, czyli trawienna.
Składniki ściany komórkowej. Najbardziej typowym składnikiem ściany komórkowej jest celuloza, która w dużej mierze determinuje jej architekturę. Cząsteczki celulozy składają się z powtarzających się cząsteczek glukozy połączonych końcami. Długie, cienkie cząsteczki celulozy łączą się w mikrofibryle o grubości 10 - 25 nm. Mikrofibryle splatają się i tworzą cienkie nitki, które z kolei można owijać jedna wokół drugiej, jak nitki liny. Każda taka „sznurówka”, czyli makrofibryla, ma grubość około 0,5 mikrona, osiągając długość 4 mikronów. Makrofibryle są tak mocne, jak drut stalowy tej samej wielkości.
Ramka celulozowa ściany komórkowej jest wypełniona splecionymi z nią cząsteczkami matrycy celulozowej. Składa się z polisacharydów zwanych hemicelulozami oraz pektyn lub pektyn, chemicznie bardzo zbliżonych do hemiceluloz.
Inny składnik ściany komórkowej, lignina, jest najobficiej występującym polimerem w komórkach roślinnych po celulozie. Lignina zwiększa sztywność ściany i zwykle występuje w komórkach pełniących funkcję podtrzymującą lub mechaniczną.
Kutyna, suberyna, wosk - zwykle osadzają się w łuskach tkanek ochronnych roślin. Na przykład kutyna znajduje się w błonach komórkowych naskórka, a suberyna w wtórnej tkance ochronnej, korku. Obie substancje występują w połączeniu z woskami i zapobiegają nadmiernej utracie wody z rośliny.
Warstwy ściany komórkowej. Grubość ścianek komórek roślinnych jest bardzo zróżnicowana, w zależności od roli komórek w strukturze rośliny i wieku samej komórki. Pod mikroskopem elektronowym w ścianie komórkowej rośliny widoczne są dwie warstwy: blaszka środkowa (zwana także substancją międzykomórkową) i pierwotna ściana komórkowa. Wiele komórek tworzy kolejną warstwę, wtórną ścianę komórkową. Płytka środkowa znajduje się między głównymi ścianami sąsiednich komórek. Ściana wtórna, jeśli występuje, jest osadzana przez protoplast komórki na wewnętrznej powierzchni pierwotnej ściany komórkowej.
Płyta środkowa. Blaszka środkowa składa się głównie z pektyn. Tam, gdzie powinna powstać ściana komórkowa, między dwiema nowo utworzonymi komórkami, najpierw zauważa się gęsty splot kanalików retikulum endoplazmatycznego i cysterny aparatu Golgiego (dictyosomy). Wtedy w tym miejscu pojawiają się bąbelki wypełnione pektyną (z polisacharydów). Pęcherzyki te są oddzielane od zbiorników aparatu Golgiego. Wczesna ściana komórkowa zawiera różne polisacharydy, z których głównymi są pektyny i hemiceluloza. Później składa się z gęstszych substancji - celulozy i ligniny.
Pierwotna ściana komórkowa. Jest to warstwa otoczki celulozowej, która jest osadzana przed lub w trakcie wzrostu komórek. Oprócz celulozy, hemicelulozy i pektyny błony pierwotne zawierają glikoproteinę. Muszle pierwotne mogą ulegać zdrewnieniu. Składnik pektynowy nadaje plastyczność, która umożliwia rozszerzanie się pierwotnej skorupy w miarę wydłużania się korzenia, łodygi lub liścia.
Komórki aktywnie dzielące się (większość dojrzałych komórek biorących udział w procesach fotosyntezy, oddychania i wydzielania) mają błony pierwotne. Takie komórki z pierwotną błoną i żywym protoplastem są w stanie utracić swój charakterystyczny kształt, dzielić się i różnicować w nowy typ komórek. U roślin biorą udział w gojeniu się ran i regeneracji tkanek.
Pierwotne błony komórkowe nie mają jednolitej grubości na całej długości, ale mają cienkie sekcje, które nazywane są pierwotnymi polami porów. Nici cytoplazmy lub plazmodesmy, które łączą protoplasty sąsiednich komórek, zwykle przechodzą przez pierwotne pola porów.
wtórna ściana komórkowa. Pomimo faktu, że wiele komórek roślinnych ma tylko ścianę pierwotną, w niektórych protoplast odkłada ścianę wtórną w kierunku środka komórki. Zwykle dzieje się tak po ustaniu wzrostu komórek, a powierzchnia błony pierwotnej już się nie zwiększa. Z tego powodu wtórna powłoka różni się od podstawowej. Membrany wtórne są szczególnie potrzebne wyspecjalizowanym komórkom, które wzmacniają roślinę i przewodzą wodę. Po odłożeniu błony wtórnej protoplast tych komórek z reguły umiera. W skorupkach wtórnych jest więcej celulozy niż w skorupkach pierwotnych, a nie ma w nich substancji pektynowych i glikoprotein. Błona wtórna jest trudna do rozciągnięcia, jej macierz składa się z hemicelulozy.
W powłoce wtórnej można wyróżnić trzy warstwy - zewnętrzną, środkową i wewnętrzną (S1, S2, S3). Warstwowa struktura skorup wtórnych znacznie zwiększa ich wytrzymałość. Mikrofibryle celulozy w błonie wtórnej są osadzane gęstiej niż w błonie pierwotnej. Lignina jest powszechnym składnikiem skór wtórnych drewna.
Pory w błonach stykających się komórek znajdują się naprzeciw siebie. Dwa przeciwległe pory i membrana porów tworzą parę porów. W komórkach z błonami wtórnymi występują dwa główne typy porów: proste i obramowane. W porach graniczących ściana wtórna wisi nad jamą porów. W prostych porach tak nie jest.
Wzrost ściany komórkowej. W miarę wzrostu komórki zwiększa się grubość i powierzchnia błony komórkowej. Rozciąganie skorupy to złożony proces. Jest kontrolowany przez protoplast i regulowany przez hormon auksyny.
W komórkach rosnących równomiernie we wszystkich kierunkach odkładanie miofibryli jest przypadkowe. Te miofibryle tworzą nieregularną sieć. Takie komórki znaleziono w rdzeniu łodygi, tkankach spichrzowych oraz podczas hodowli komórek in vitro. W wydłużających się komórkach miofibryle osłonek bocznych osadzają się pod kątem prostym do osi wydłużenia.
Substancje macierzy - pektyny, hemicelulozy i glikoproteiny są przenoszone na błonę w pęcherzykach dictyosomów. Jednocześnie pektyny są bardziej charakterystyczne dla komórek rosnących, podczas gdy hemicelulozy przeważają w komórkach nie rosnących.
Mikrofibryle celulozowe są syntetyzowane na powierzchni komórki za pomocą kompleksu enzymatycznego związanego z błoną plazmatyczną. Orientacja mikrofibryli jest kontrolowana przez mikrotubule znajdujące się w pobliżu wewnętrznej powierzchni błony plazmatycznej.
Plazmodesma. Są to cienkie nici cytoplazmy, które łączą protoplasty sąsiednich komórek. Plazmodesmy albo przechodzą przez ścianę komórkową w dowolnym miejscu, albo są skoncentrowane w pierwotnych polach porów lub w błonach między parami porów. Pod mikroskopem elektronowym plazmodesmy wyglądają jak wąskie kanały wyłożone błoną plazmatyczną. Wzdłuż osi kanału mniejsza cylindryczna rurka, desmotubula, rozciąga się od jednej komórki do drugiej, która komunikuje się z retikulum endoplazmatycznym obu sąsiednich komórek. Wiele plazmodesmy tworzy się podczas podziału komórki, kiedy rurkowa retikulum endoplazmatyczne jest pobierane przez rozwijającą się płytkę komórkową. Plazmodesmy mogą również tworzyć się w błonach komórek, które się nie dzielą. Struktury te zapewniają sprawne przenoszenie określonych substancji z komórki do komórki.
Podział komórek. W organizmach wielokomórkowych podział komórek wraz ze wzrostem ich wielkości jest sposobem wzrostu całego organizmu. Nowe komórki powstałe podczas podziału są podobne pod względem struktury i funkcji, zarówno do komórki macierzystej, jak i do siebie nawzajem. Proces podziału u eukariontów można podzielić na dwa częściowo nakładające się etapy: mitozę i cytokinezę.
Mitoza to tworzenie się dwóch jąder potomnych z jednego jądra, morfologicznie i genetycznie równoważnych sobie nawzajem. Cytokineza to podział cytoplazmatycznej części komórki z utworzeniem komórek potomnych.
Cykl komórkowy.Żywa komórka przechodzi przez serię następujących po sobie zdarzeń, które składają się na cykl komórkowy. Czas trwania samego cyklu różni się w zależności od rodzaju komórki oraz czynników zewnętrznych, takich jak temperatura czy dostępność składników odżywczych. Zwykle cykl dzieli się na interfazę i cztery fazy mitozy.
Interfaza. Okres między kolejnymi podziałami mitotycznymi.
Interfaza jest podzielona na trzy okresy, oznaczone jako G1, S, G2.
W okresie G1, który rozpoczyna się po mitozie. W tym okresie wzrasta ilość cytoplazmy, w tym różnych organelli. Ponadto, zgodnie z obecną hipotezą, w okresie G1 syntetyzowane są substancje, które stymulują lub hamują okres S i resztę cyklu, determinując w ten sposób proces podziału.
Okres S następuje po okresie G1, w którym następuje duplikacja materiału genetycznego (DNA).
W okresie G2, który następuje po S, powstają struktury bezpośrednio zaangażowane w mitozę, takie jak elementy wrzeciona.
Niektóre komórki przechodzą nieograniczoną liczbę cykli komórkowych. Są to organizmy jednokomórkowe oraz niektóre komórki stref aktywnego wzrostu (merystemy). Niektóre wyspecjalizowane komórki po dojrzewaniu tracą zdolność do reprodukcji. Trzecia grupa komórek, na przykład te tworzące tkankę rany (kalus), zachowuje zdolność do podziału tylko w szczególnych warunkach.
Mitoza, lub rozszczepienie jądrowe. Jest to ciągły proces podzielony na cztery fazy: profaza, metafaza, anafaza, telofaza. W wyniku mitozy materiał genetyczny, który podwoił się w interfazie, jest równo dzielony między dwa jądra potomne.
Jedną z najwcześniejszych oznak przejścia komórki do podziału jest pojawienie się wąskiego, pierścieniowego pasma mikrotubul tuż pod błoną plazmatyczną. Ten stosunkowo gęsty pas otacza jądro w płaszczyźnie równikowej przyszłego wrzeciona mitotycznego. Ponieważ pojawia się przed profazą, nazywa się go pasem preprofazowym. Zanika po wrzecionie mitotycznym, na długo przed pojawieniem się w późnej telofazie płytki komórkowej, która rozrasta się od środka ku obrzeżom i łączy się z błoną komórki macierzystej w obszarze zajmowanym wcześniej przez pas preprofazowy.
profaza. Na początku profazy chromosomy przypominają długie nitki rozproszone wewnątrz jądra. Następnie, gdy nici skracają się i pogrubiają, można zobaczyć, że każdy chromosom składa się nie z jednej, ale z dwóch splecionych nici zwanych chromatydami. W późnej profazie dwie skrócone sparowane chromatydy każdego chromosomu leżą obok siebie równolegle, połączone wąskim odcinkiem zwanym centromerem. Ma określoną pozycję na każdym chromosomie i dzieli chromosom na dwa ramiona o różnej długości.
Mikrotubule znajdują się równolegle do powierzchni jądra wzdłuż osi wrzeciona. Jest to najwcześniejsza manifestacja zespołu wrzeciona mitotycznego.
Pod koniec profazy jąderko stopniowo traci swój wyraźny zarys i ostatecznie zanika. Wkrótce potem otoczka jądrowa również się rozpada.
Metafaza. Na początku metafazy wrzeciono, które jest strukturą trójwymiarową, najszerszą w środku i zwężającą się ku biegunom, zajmuje miejsce zajmowane wcześniej przez jądro. Włókna wrzeciona są wiązkami mikrotubul. Podczas metafazy chromosomy, z których każdy składa się z dwóch chromatyd, są ułożone w taki sposób, że ich centromery leżą w płaszczyźnie równikowej wrzeciona. Dzięki centromerowi każdy chromosom jest przyczepiony do nici wrzeciona. Jednak niektóre nici przechodzą z jednego bieguna na drugi bez przyczepiania się do chromosomów.
Kiedy wszystkie chromosomy znajdują się w płaszczyźnie równikowej, metafaza jest zakończona. Chromosomy są gotowe do podziału.
Anafaza. Chromatydy każdego chromosomu rozdzielają się. Teraz to są chromosomy potomne. Po pierwsze, centromer dzieli się, a dwa chromosomy potomne są przyciągane do przeciwnych biegunów. W tym przypadku centromery przesuwają się do przodu, a ramiona chromosomów rozciągają się do tyłu. Włókna wrzeciona przyczepione do chromosomów skracają się, przyczyniając się do rozdzielenia chromatyd i ruchu chromosomów potomnych w przeciwnych kierunkach.
STRUKTURA KOMÓRKI ROŚLINNEJ
Komórka roślinna składa się z mniej lub bardziej sztywnej ściany komórkowej i protoplastu. Ściana komórkowa to ściana komórkowa i błona cytoplazmatyczna. Termin protoplast pochodzi od słowa protoplazma, które od dawna jest używane w odniesieniu do wszystkich żywych istot. Protoplast jest protoplazmą pojedynczej komórki.
Protoplast składa się z cytoplazmy i jądra. Cytoplazma zawiera organelle (rybosomy, mikrotubule, plastydy, mitochondria) i układy membranowe (retikulum endoplazmatyczne, dictyosomy). Cytoplazma obejmuje również macierz cytoplazmatyczną (substancję podstawową), w której zanurzone są organelle i układy błon. Cytoplazma jest oddzielona od ściany komórkowej błoną plazmatyczną, która jest błoną elementarną. W przeciwieństwie do większości komórek zwierzęcych, komórki roślinne zawierają jedną lub więcej wakuoli. Są to pęcherzyki wypełnione cieczą i otoczone elementarną błoną (tonoplastem).
W żywej komórce roślinnej substancja podstawowa jest w ciągłym ruchu. Organelle biorą udział w ruchu, zwanym przepływem cytoplazmy lub cyklozą. Cykloza ułatwia przemieszczanie się substancji w komórce i ich wymianę między komórką a środowiskiem.
Membrana plazmowa. Jest to dwuwarstwowa struktura fosfolipidowa. Komórki roślinne charakteryzują się inwazjami błony plazmatycznej.
Błona plazmatyczna pełni następujące funkcje:
Uczestniczy w wymianie substancji między komórką a środowiskiem;
Koordynuje syntezę i montaż mikrofibryli celulozowych ściany komórkowej;
Przekazuje sygnały hormonalne i zewnętrzne, które kontrolują wzrost i różnicowanie komórek.
Rdzeń. Jest to najbardziej widoczna struktura w cytoplazmie komórki eukariotycznej. Jądro spełnia dwie ważne funkcje:
Kontroluje aktywność życiową komórki, określając, które białka iw jakim czasie powinny zostać zsyntetyzowane;
Przechowuje informacje genetyczne i przekazuje je komórkom potomnym podczas podziału komórki.
Jądro komórki eukariotycznej jest otoczone dwiema podstawowymi błonami, które tworzą otoczkę jądrową. Przesiąknięta jest licznymi porami o średnicy od 30 do 100 nm, widocznymi jedynie pod mikroskopem elektronowym. Pory mają złożoną strukturę. Zewnętrzna błona błony jądrowej w niektórych miejscach jest połączona z retikulum endoplazmatycznym. Otoczkę jądrową można postrzegać jako wyspecjalizowaną, lokalnie zróżnicowaną część retikulum endoplazmatycznego (ER).
W jądrze zabarwionym specjalnymi barwnikami można wyróżnić cienkie włókna i grudki chromatyny i nukleoplazmy (głównej substancji jądra). Chromatyna składa się z DNA związanego ze specjalnymi białkami zwanymi histonami. W procesie podziału komórki chromatyna staje się coraz bardziej zagęszczona i składa się w chromosomy. DNA koduje informację genetyczną.
Organizmy różnią się liczbą chromosomów w komórkach somatycznych. Na przykład kapusta ma - 20 chromosomów; słonecznik - 34; pszenica - 42; ludzi - 46, a jeden z gatunków paproci Ophioglossum - 1250. Komórki płciowe (gamety) mają tylko połowę liczby chromosomów charakterystycznych dla komórek somatycznych organizmu. Liczba chromosomów w gametach nazywana jest haploidalną (pojedynczą), w komórkach somatycznych - diploidalną (podwójną). Komórki, które mają więcej niż dwa zestawy chromosomów, nazywane są poliploidalnymi.
Pod mikroskopem świetlnym można zobaczyć kuliste struktury - jąderka. Każde jądro ma jedno lub więcej jąderek, które są widoczne w jądrach niedzielących się. Rybosomalny RNA jest syntetyzowany w jąderku. Zazwyczaj jądra organizmów diploidalnych mają dwa jąderka, po jednym dla każdego haploidalnego zestawu chromosomów. Jąderka nie mają własnej błony. Pod względem biochemicznym jąderka charakteryzują się wysokim stężeniem RNA, które jest tu związane z fosfoproteinami. Wielkość jąderka zależy od stanu funkcjonalnego komórki. zauważono, że w szybko rosnącej komórce, w której zachodzą intensywne procesy syntezy białek, jąderka powiększają się. W jąderkach powstają mRNA i rybosomy, które pełnią funkcję syntetyczną tylko w jądrze.
Nukleoplazma (karioplazma) jest reprezentowana przez jednorodną ciecz, w której rozpuszczają się różne białka, w tym enzymy.
Plastydy. Charakterystycznymi składnikami komórek roślinnych są wakuole, celulozowe ściany komórkowe i plastydy. Każdy plastyd ma swoją własną powłokę, składającą się z dwóch elementarnych membran. Wewnątrz plastydu wyróżnia się system membranowy i w różnym stopniu jednorodną substancję, zrąb. Dojrzałe plastydy są klasyfikowane na podstawie zawartych w nich pigmentów.
Chloroplasty, w których zachodzi fotosynteza, zawierają chlorofile i karotenoidy. Zwykle mają postać krążka o średnicy 4 - 5 mikronów. W jednej komórce mezofilu (środek liścia) może znajdować się 40 - 50 chloroplastów; w mm2 liścia - około 500 000. w cytoplazmie chloroplasty są zwykle zlokalizowane równolegle do błony komórkowej.
Wewnętrzna struktura chloroplastu jest złożona. Zrąb jest przesiąknięty rozwiniętym systemem błon w postaci pęcherzyków - tylakoidów. Każdy tylakoid składa się z dwóch błon. Tylakoidy tworzą jeden system. Z reguły są one zbierane w stosy - ziarna, przypominające kolumny monet. Tylakoidy poszczególnych granów są połączone tylakoidami zrębu lub tylakoidami międzyziarnistymi. Chlorofil i karotenoidy są osadzone w błonach tylakoidów. Chloroplasty w zielonych roślinach i algach często zawierają ziarna skrobi i małe kropelki lipidów (tłuszczu). Ziarna skrobi są tymczasowymi magazynami produktów fotosyntezy. Mogą zniknąć z chloroplastów rośliny, która była w ciemności tylko przez 24 godziny, i pojawić się ponownie 3-4 godziny po przeniesieniu rośliny na światło.
Chloroplasty to półautonomiczne organelle i przypominają bakterie. Na przykład rybosomy bakterii i chloroplastów mają dość duże podobieństwo. Są mniejsze niż rybosomy eukariotyczne. Synteza białek na rybosomach bakterii i chloroplastach jest hamowana przez chloramfenikol, który nie ma wpływu na komórki eukariotyczne. Ponadto zarówno bakterie, jak i chloroplasty mają podobny typ nukleoidu, zorganizowanego w podobny sposób. Pomimo faktu, że tworzenie chloroplastów i synteza zawartych w nich pigmentów jest w dużej mierze kontrolowana przez chromosomalny DNA komórki, to jednak przy braku własnego DNA chloroplasty nie tworzą się.
Chloroplasty można uznać za główne organelle komórkowe, ponieważ są pierwszymi w łańcuchu transformacji energii słonecznej, w wyniku czego ludzkość otrzymuje zarówno żywność, jak i paliwo. W chloroplastach zachodzi nie tylko fotosynteza. Biorą również udział w syntezie aminokwasów i kwasów tłuszczowych, służą jako magazyn tymczasowych rezerw skrobi.
Chromoplasty to pigmentowane plastydy. Różnorodne w formie, nie posiadają chlorofilu, ale syntetyzują i gromadzą karotenoidy, które nadają żółty, pomarańczowy, czerwony kolor kwiatom, starym liściom, owocom i korzeniom. chromoplasty mogą rozwijać się z chloroplastów, które w tym przypadku tracą chlorofil i struktury błony wewnętrznej, akumulują karotenoidy. Dzieje się tak podczas dojrzewania wielu owoców. Chromoplasty przyciągają owady i inne zwierzęta, z którymi współewoluowały.
Leukoplasty to niepigmentowane plastydy. Niektóre z nich syntetyzują skrobię (amyloplasty), inne są zdolne do tworzenia różnych substancji, w tym lipidów i białek. W świetle leukoplasty zamieniają się w chloroplasty.
Proplastydy to małe, bezbarwne lub jasnozielone, niezróżnicowane plastydy występujące w merystematycznych (dzielących się) komórkach korzeni i pędów. Są prekursorami innych, bardziej zróżnicowanych plastydów - chloroplastów, chromoplastów i aminoplastów. Jeśli rozwój protoplastów jest opóźniony z powodu braku światła, może pojawić się w nich jedno lub więcej ciałek prolamelarnych, które są półkrystalicznymi skupiskami błon kanalikowych. Plastydy zawierające ciałka prolamelarne nazywane są etioplastami. W świetle etioplasty zamieniają się w chloroplasty, podczas gdy błony ciał prolamelarnych tworzą tylakoidy. Etioplasty powstają w liściach roślin, które są w ciemności. protoplasty zarodków nasiennych najpierw przekształcają się w etioplasty, z których następnie w świetle rozwijają się chloroplasty. Plastydy charakteryzują się stosunkowo łatwymi przejściami z jednego rodzaju do drugiego. Plastydy, podobnie jak bakterie, rozmnażają się, dzieląc się na dwie części. W komórkach merystematycznych czas podziału protoplastów w przybliżeniu pokrywa się z czasem podziału komórki. Jednak w dojrzałych komórkach większość plastydów powstaje w wyniku podziału dojrzałych plastydów.
mitochondria. Podobnie jak chloroplasty, mitochondria są otoczone dwiema podstawowymi błonami. Wewnętrzna błona tworzy liczne fałdy i wypukłości - cristae, które znacznie zwiększają wewnętrzną powierzchnię mitochondriów. Są znacznie mniejsze niż plastydy, mają średnicę około 0,5 µm oraz zmienną długość i kształt.
W mitochondriach zachodzi proces oddychania, w wyniku którego dochodzi do rozpadu cząsteczek organicznych z uwolnieniem energii i przekazaniem jej do cząsteczek ATP, głównej rezerwy energetycznej wszystkich komórek eukariotycznych. Większość komórek roślinnych zawiera setki lub tysiące mitochondriów. Ich liczba w jednej komórce jest określona przez zapotrzebowanie komórki na ATP. Mitochondria są w ciągłym ruchu, przemieszczając się z jednej części komórki do drugiej, łącząc się ze sobą i dzieląc. Mitochondria zwykle gromadzą się tam, gdzie potrzebna jest energia. Jeśli błona plazmatyczna aktywnie przenosi substancje z komórki do komórki, wówczas mitochondria znajdują się wzdłuż powierzchni błony. W ruchliwych glonach jednokomórkowych mitochondria gromadzą się u podstawy wici, dostarczając energię niezbędną do ich ruchu.
Mitochondria, podobnie jak plastydy, są półautonomicznymi organellami zawierającymi składniki niezbędne do syntezy własnych białek. Błona wewnętrzna otacza płynną matrycę, która zawiera białka, RNA, DNA, bakteryjne rybosomy i różne substancje rozpuszczone. DNA istnieje w postaci kolistych cząsteczek zlokalizowanych w jednym lub kilku nukleoidach.
Na podstawie podobieństwa bakterii do mitochondriów i chloroplastów komórek eukariotycznych można przypuszczać, że mitochondria i chloroplasty wywodzą się od bakterii, które znalazły „schronienie” w większych komórkach heterotroficznych – prekursorach eukariontów.
Mikrociała. W przeciwieństwie do plastydów i mitochondriów, które są ograniczone dwiema błonami, mikrociała są kulistymi organellami otoczonymi pojedynczą błoną. Mikrociała mają ziarnistą (granulowaną) zawartość, czasami zawierają również krystaliczne wtrącenia białkowe. Mikrociała są związane z jednym lub dwoma regionami retikulum endoplazmatycznego.
Niektóre mikrociała, zwane proksysomami, odgrywają ważną rolę w metabolizmie kwasu glikolowego, który jest bezpośrednio związany z fotooddychaniem. W zielonych liściach są one związane z mitochondriami i chloroplastami. Inne mikrociała, zwane glioksysomami, zawierają enzymy potrzebne do przekształcania tłuszczów w węglowodany. Dzieje się tak w przypadku wielu nasion podczas kiełkowania.
wakuole - Są to obszary komórki ograniczone błoną, wypełnione płynem - sokiem komórkowym. Otoczone są tonoplastem (błoną wakuolową).
Młoda komórka roślinna zawiera liczne małe wakuole, które w miarę starzenia się łączą się w jedną dużą wakuolę. W dojrzałej komórce do 90% jej objętości może zajmować wakuola. W tym przypadku cytoplazma jest dociskana w postaci cienkiej warstwy obwodowej do błony komórkowej. Wzrost wielkości komórek wynika głównie ze wzrostu wakuoli. W rezultacie powstaje ciśnienie turgoru i zachowana jest elastyczność tkanki. Jest to jedna z głównych funkcji wakuoli i tonoplastu.
Głównym składnikiem soku jest woda, reszta zależy od rodzaju rośliny i jej stanu fizjologicznego. Wakuole zawierają sole, cukry, rzadziej białka. Tonoplast odgrywa aktywną rolę w transporcie i gromadzeniu się niektórych jonów w wakuoli. Stężenie jonów w soku komórkowym może znacznie przekraczać jego stężenie w środowisku. Przy wysokiej zawartości niektórych substancji w wakuolach tworzą się kryształy. Najczęściej spotykane są kryształy szczawianu wapnia, które mają inny kształt.
Wakuole są miejscami gromadzenia się produktów przemiany materii (metabolizmu). Mogą to być białka, kwasy, a nawet substancje toksyczne dla człowieka (alkaloidy). Często osadzają się pigmenty. Niebieski, fioletowy, purpurowy, ciemnoczerwony, karmazynowy dają komórkom roślinnym pigmenty z grupy antocyjanów. W przeciwieństwie do innych pigmentów dobrze rozpuszczają się w wodzie i znajdują się w soku komórkowym. Decydują o czerwonym i niebieskim zabarwieniu wielu warzyw (rzodkiewki, rzepa, kapusta), owoców (winogrona, śliwki, wiśnie), kwiatów (chabry, pelargonie, ostróżki, róże, piwonie). Czasami te pigmenty maskują chlorofil w liściach, na przykład w ozdobnym czerwonym klonie. Antocyjany barwią jesienne liście na jaskrawoczerwony. Powstają w chłodne słoneczne dni, kiedy synteza chlorofilu zatrzymuje się w liściach. W liściach, gdy nie tworzą się antocyjany, po zniszczeniu chlorofilu zauważalne stają się żółto-pomarańczowe karotenoidy chloroplastów. Liście są najjaśniejsze w chłodnej, jasnej jesieni.
Wakuole biorą udział w niszczeniu makrocząsteczek, w krążeniu ich składników w komórce. Rybosomy, mitochondria, plastydy wpadające do wakuoli ulegają zniszczeniu. Zgodnie z tą czynnością trawienną można je porównać z lizosomami - organellami komórek zwierzęcych.
Wakuole powstają z retikulum endoplazmatycznego (retikulum)
Rybosomy. Małe cząsteczki (17 - 23nm), składające się z mniej więcej równych ilości białka i RNA. W rybosomach aminokwasy łączą się, tworząc białka. W komórkach o aktywnym metabolizmie jest ich więcej. Rybosomy znajdują się w cytoplazmie komórki swobodnie lub przyczepione do retikulum endoplazmatycznego (80S). Występują również w jądrze (80S), mitochondriach (70S), plastydach (70S).
Rybosomy mogą tworzyć kompleks, na którym zachodzi jednoczesna synteza identycznych polipeptydów, o których informacja pochodzi z jednej cząsteczki i RNA. Taki kompleks nazywa się polirybosomami (polisomami). Komórki, które syntetyzują białka w dużych ilościach, mają rozbudowany system polisomów, które często są przyczepione do zewnętrznej powierzchni otoczki jądrowej.
Retikulum endoplazmatyczne. Jest to złożony trójwymiarowy system membranowy o nieokreślonym zasięgu. W kontekście ER wygląda jak dwie elementarne membrany z wąską przezroczystą przestrzenią między nimi. Kształt i długość ER zależy od rodzaju komórki, jej aktywności metabolicznej oraz stopnia zróżnicowania. W komórkach, które wydzielają lub przechowują białka, ER ma kształt płaskiego worka lub cysterny, z licznymi rybosomami przyczepionymi do jego zewnętrznej powierzchni. Ta siateczka nazywana jest szorstką retikulum endoplazmatycznym. Gładki ER ma zwykle kształt cylindryczny. W tej samej komórce może znajdować się szorstka i gładka siateczka śródplazmatyczna. Z reguły istnieje między nimi wiele połączeń.
Retikulum endoplazmatyczne działa jako system komunikacyjny komórki. Jest związany z zewnętrzną powłoką jądra. W rzeczywistości te dwie struktury tworzą jeden system membranowy. Kiedy otoczka jądrowa pęka podczas podziału komórki, jej fragmenty przypominają fragmenty ER. Retikulum endoplazmatyczne to system transportu substancji: białek, lipidów, węglowodanów do różnych części komórki. retikulum endoplazmatyczne sąsiednich komórek są połączone przez nici cytoplazmatyczne - plazmodesmy - które przechodzą przez błony komórkowe.
Retikulum endoplazmatyczne jest głównym miejscem syntezy błon komórkowych. W niektórych komórkach roślinnych tworzą się tu błony wakuoli i mikrociał, cysterny dictyosomów.
Aparat Golgiego. Termin ten jest używany w odniesieniu do wszystkich dictyosomów lub ciał Golgiego w komórce. Dictyosomy to grupy płaskich pęcherzyków w kształcie dysku lub cystern, które rozgałęziają się wzdłuż krawędzi w złożony system kanalików. Dictyosomy u roślin wyższych składają się z 4 - 8 cystern, zebranych razem.
Zwykle w paczce zbiorników wyróżnia się stronę rozwijającą się i dojrzewającą. błony tworzących się cystern przypominają strukturą błony ER, a błony dojrzewających cystern przypominają błonę komórkową.
Dictyosomy biorą udział w wydzielaniu, a u większości roślin wyższych w tworzeniu błon komórkowych. Polisacharydy ściany komórkowej syntetyzowane przez dictyosomy gromadzą się w pęcherzykach, które następnie oddzielają się od dojrzewających cystern. Te pęcherzyki wydzielnicze migrują i łączą się z CMP; natomiast zawarte w nich polisacharydy wbudowują się w błonę komórkową. Niektóre substancje gromadzące się w dictyosomach powstają w innych strukturach, na przykład w ER, a następnie są transportowane do dictyosomów, gdzie są modyfikowane (modyfikowane) przed sekrecją. Na przykład glikoproteiny są ważnym materiałem budulcowym błony komórkowej. Część białkowa jest syntetyzowana przez polisomy szorstkiego ER, część węglowodanowa jest syntetyzowana w dictyosomach, gdzie obie części łączą się tworząc glikoproteiny.
Membrany to dynamiczne, ruchome struktury, które nieustannie zmieniają swój kształt i powierzchnię. Koncepcja układu endoplazmatycznego opiera się na ruchliwości błon. Zgodnie z tą koncepcją błony wewnętrzne cytoplazmy, z wyjątkiem błon mitochondriów i plastydów, stanowią jedną całość i wywodzą się z retikulum endoplazmatycznego. Nowe cysterny dictyosomów tworzą się z retikulum endoplazmatycznego przez etap pęcherzyków pośrednich, a pęcherzyki wydzielnicze, które oddzielają się od dictyosomów, ostatecznie przyczyniają się do tworzenia błony plazmatycznej. Zatem retikulum endoplazmatyczne i dictyosomy tworzą jednostkę funkcjonalną, w której dictyosomy pełnią rolę struktur pośrednich w procesie przekształcania błon, takich jak retikulum endoplazmatyczne, w błony takie jak plazma. W tkankach, których komórki słabo rosną i dzielą się, składniki błony są stale aktualizowane.
mikrotubule występuje w prawie wszystkich komórkach eukariotycznych. Są to cylindryczne struktury o średnicy około 24 nm. Ich długość jest różna. Każdy kanalik składa się z podjednostek białka zwanego tubuliną. Podjednostki tworzą 13 podłużnych włókien otaczających centralną jamę. Mikrotubule są strukturami dynamicznymi, są regularnie niszczone i formowane na określonych etapach cyklu komórkowego. Ich gromadzenie odbywa się w specjalnych miejscach, które nazywane są centrami organizacji mikrotubul. W komórkach roślinnych mają słabo wyrażoną amorficzną strukturę.
Funkcje mikrotubul: uczestniczą w tworzeniu błony komórkowej; kierują pęcherzyki dictyosomów do tworzącej się błony, jak nici wrzeciona, które tworzą się w dzielącej się komórce; odgrywają rolę w tworzeniu płytki komórkowej (początkowa granica między komórkami potomnymi). Ponadto mikrotubule są ważnym składnikiem wici i rzęsek, w których ruchu odgrywają ważną rolę.
mikrofilamenty, podobnie jak mikrotubule, znajdują się w prawie wszystkich komórkach eukariotycznych. Są to długie włókna o grubości 5–7 nm, składające się z kurczliwego białka aktyny. Pęczki mikrofilamentów znajdują się w wielu komórkach roślin wyższych. Najwyraźniej odgrywają ważną rolę w prądach cytoplazmy. Mikrofilamenty wraz z mikrotubulami tworzą elastyczną sieć zwaną cytoszkieletem.
Substancja podstawowa przez długi czas był uważany za jednorodny (homogeniczny) bogaty w białko roztwór o niewielkiej liczbie struktur lub nawet bez struktury. Jednak obecnie, przy użyciu wysokonapięciowego mikroskopu elektronowego, ustalono, że substancja podstawowa jest trójwymiarową siatką zbudowaną z cienkich (o średnicy 3–6 nm) pasm wypełniających całą komórkę. Inne składniki cytoplazmy, w tym mikrotubule i mikrofilamenty, są zawieszone w tej siatce mikrotrabekularnej.
Struktura mikrotrabekularna to sieć pasm białkowych, między którymi przestrzeń jest wypełniona wodą. Wraz z wodą siatka ma konsystencję żelu, który wygląda jak galaretowate ciała.
Organelle są przyczepione do siatki mikrotrabekularnej. Sieć zapewnia komunikację pomiędzy poszczególnymi częściami komórki oraz kieruje transportem wewnątrzkomórkowym.
Krople lipidowe- struktury o kulistym kształcie, nadające ziarnistość cytoplazmie komórki roślinnej pod mikroskopem świetlnym. Na mikrografach elektronowych wyglądają na amorficzne. Bardzo podobne, ale mniejsze kropelki znajdują się w plastydach.
Kropelki lipidów, myląc je z organellami, nazywały je sferosomami i uważały, że otoczone są jednowarstwową lub dwuwarstwową błoną. Jednak ostatnie dane pokazują, że kropelki lipidów nie mają błon, ale mogą być pokryte białkiem.
Substancje ergastyczne - są to „produkty pasywne” protoplastu: substancje rezerwowe lub produkty odpadowe. Mogą pojawiać się i znikać w różnych okresach cyklu komórkowego. Z wyjątkiem ziaren skrobi, kryształów, barwników antocyjanowych i kropelek lipidów. Należą do nich żywice, gumy, garbniki i białka. Substancje ergastyczne są częścią błony komórkowej, głównej substancji cytoplazmy i organelli, w tym wakuoli.
Wici i rzęski - są to cienkie, przypominające włosy struktury, które wystają z powierzchni wielu komórek eukariotycznych. Mają stałą średnicę, ale długość waha się od 2 do 150 mikronów. Konwencjonalnie dłuższe i nieliczne z nich nazywane są wiciami, a krótsze i liczniejsze nazywane są rzęskami. Nie ma wyraźnych różnic między tymi dwoma typami struktur, więc termin wici jest używany w odniesieniu do obu.
U niektórych alg i grzybów wici są narządami ruchu, za pomocą których poruszają się w wodzie. W roślinach (np. mchach, wątrobowcach, paprociach, niektórych nagonasiennych) tylko komórki rozrodcze (gamety) mają wici.
Każda wici ma określoną organizację. Zewnętrzny pierścień 9 par mikrotubul otacza dwie dodatkowe mikrotubule znajdujące się w środku wici. Zawierające enzym „uchwyty” rozciągają się od jednej mikrotubuli każdej z zewnętrznych par. Jest to podstawowy wzorzec organizacji 9+2 występujący we wszystkich wici organizmów eukariotycznych. Uważa się, że ruch wici opiera się na przesuwaniu się mikrotubul, podczas gdy zewnętrzne pary mikrotubul poruszają się jedna po drugiej bez skurczu. Przesuwanie się par mikrotubul względem siebie powoduje miejscowe wyginanie wici.
Wici „wyrastają” z cytoplazmatycznych cylindrycznych struktur zwanych ciałami podstawowymi, które również tworzą podstawową część wici. Ciała podstawowe mają wewnętrzną strukturę przypominającą wici, z wyjątkiem tego, że kanaliki zewnętrzne są ułożone raczej w trojaczki niż w pary, a kanalików środkowych nie ma.
Ściana komórkowa.Ściana komórkowa ogranicza rozmiar protoplastu i zapobiega jego pęknięciu w wyniku absorpcji wody przez wakuolę.
Ściana komórkowa pełni określone funkcje, które są ważne nie tylko dla komórki i tkanki, w której komórka się znajduje, ale także dla całej rośliny. Ściany komórkowe odgrywają zasadniczą rolę we wchłanianiu, transporcie i wydalaniu substancji, a ponadto może się w nich koncentrować aktywność lizosomalna, czyli trawienna.
Składniki ściany komórkowej. Najbardziej typowym składnikiem ściany komórkowej jest celuloza, która w dużej mierze determinuje jej architekturę. Cząsteczki celulozy składają się z powtarzających się cząsteczek glukozy połączonych końcami. Długie, cienkie cząsteczki celulozy łączą się w mikrofibryle o grubości 10 - 25 nm. Mikrofibryle splatają się i tworzą cienkie nitki, które z kolei można owijać jedna wokół drugiej, jak nitki liny. Każda taka „sznurówka”, czyli makrofibryla, ma grubość około 0,5 mikrona, osiągając długość 4 mikronów. Makrofibryle są tak mocne, jak drut stalowy tej samej wielkości.
Ramka celulozowa ściany komórkowej jest wypełniona splecionymi z nią cząsteczkami matrycy celulozowej. Składa się z polisacharydów zwanych hemicelulozami oraz pektyn lub pektyn, chemicznie bardzo zbliżonych do hemiceluloz.
Inny składnik ściany komórkowej, lignina, jest najobficiej występującym polimerem w komórkach roślinnych po celulozie. Lignina zwiększa sztywność ściany i zwykle występuje w komórkach pełniących funkcję podtrzymującą lub mechaniczną.
Kutyna, suberyna, wosk - zwykle osadzają się w łuskach tkanek ochronnych roślin. Na przykład kutyna znajduje się w błonach komórkowych naskórka, a suberyna w wtórnej tkance ochronnej, korku. Obie substancje występują w połączeniu z woskami i zapobiegają nadmiernej utracie wody z rośliny.
Warstwy ściany komórkowej. Grubość ścianek komórek roślinnych jest bardzo zróżnicowana, w zależności od roli komórek w strukturze rośliny i wieku samej komórki. Pod mikroskopem elektronowym w ścianie komórkowej rośliny widoczne są dwie warstwy: blaszka środkowa (zwana także substancją międzykomórkową) i pierwotna ściana komórkowa. Wiele komórek tworzy kolejną warstwę, wtórną ścianę komórkową. Płytka środkowa znajduje się między głównymi ścianami sąsiednich komórek. Ściana wtórna, jeśli występuje, jest osadzana przez protoplast komórki na wewnętrznej powierzchni pierwotnej ściany komórkowej.
Płyta środkowa. Blaszka środkowa składa się głównie z pektyn. Tam, gdzie powinna powstać ściana komórkowa, między dwiema nowo utworzonymi komórkami, najpierw zauważa się gęsty splot kanalików retikulum endoplazmatycznego i cysterny aparatu Golgiego (dictyosomy). Wtedy w tym miejscu pojawiają się bąbelki wypełnione pektyną (z polisacharydów). Pęcherzyki te są oddzielane od zbiorników aparatu Golgiego. Wczesna ściana komórkowa zawiera różne polisacharydy, z których głównymi są pektyny i hemiceluloza. Później składa się z gęstszych substancji - celulozy i ligniny.
Pierwotna ściana komórkowa. Jest to warstwa otoczki celulozowej, która jest osadzana przed lub w trakcie wzrostu komórek. Oprócz celulozy, hemicelulozy i pektyny błony pierwotne zawierają glikoproteinę. Muszle pierwotne mogą ulegać zdrewnieniu. Składnik pektynowy nadaje plastyczność, która umożliwia rozszerzanie się pierwotnej skorupy w miarę wydłużania się korzenia, łodygi lub liścia.
Komórki aktywnie dzielące się (większość dojrzałych komórek biorących udział w procesach fotosyntezy, oddychania i wydzielania) mają błony pierwotne. Takie komórki z pierwotną błoną i żywym protoplastem są w stanie utracić swój charakterystyczny kształt, dzielić się i różnicować w nowy typ komórek. U roślin biorą udział w gojeniu się ran i regeneracji tkanek.
Pierwotne błony komórkowe nie mają jednolitej grubości na całej długości, ale mają cienkie sekcje, które nazywane są pierwotnymi polami porów. Nici cytoplazmy lub plazmodesmy, które łączą protoplasty sąsiednich komórek, zwykle przechodzą przez pierwotne pola porów.
wtórna ściana komórkowa. Pomimo faktu, że wiele komórek roślinnych ma tylko ścianę pierwotną, w niektórych protoplast odkłada ścianę wtórną w kierunku środka komórki. Zwykle dzieje się tak po ustaniu wzrostu komórek, a powierzchnia błony pierwotnej już się nie zwiększa. Z tego powodu wtórna powłoka różni się od podstawowej. Membrany wtórne są szczególnie potrzebne wyspecjalizowanym komórkom, które wzmacniają roślinę i przewodzą wodę. Po odłożeniu błony wtórnej protoplast tych komórek z reguły umiera. W skorupkach wtórnych jest więcej celulozy niż w skorupkach pierwotnych, a nie ma w nich substancji pektynowych i glikoprotein. Błona wtórna jest trudna do rozciągnięcia, jej macierz składa się z hemicelulozy.
W powłoce wtórnej można wyróżnić trzy warstwy - zewnętrzną, środkową i wewnętrzną (S1, S2, S3). Warstwowa struktura skorup wtórnych znacznie zwiększa ich wytrzymałość. Mikrofibryle celulozy w błonie wtórnej są osadzane gęstiej niż w błonie pierwotnej. Lignina jest powszechnym składnikiem skór wtórnych drewna.
Pory w błonach stykających się komórek znajdują się naprzeciw siebie. Dwa przeciwległe pory i membrana porów tworzą parę porów. W komórkach z błonami wtórnymi występują dwa główne typy porów: proste i obramowane. W porach graniczących ściana wtórna wisi nad jamą porów. W prostych porach tak nie jest.
Wzrost ściany komórkowej. W miarę wzrostu komórki zwiększa się grubość i powierzchnia błony komórkowej. Rozciąganie skorupy to złożony proces. Jest kontrolowany przez protoplast i regulowany przez hormon auksyny.
W komórkach rosnących równomiernie we wszystkich kierunkach odkładanie miofibryli jest przypadkowe. Te miofibryle tworzą nieregularną sieć. Takie komórki znaleziono w rdzeniu łodygi, tkankach spichrzowych oraz podczas hodowli komórek in vitro. W wydłużających się komórkach miofibryle osłonek bocznych osadzają się pod kątem prostym do osi wydłużenia.
Substancje macierzy - pektyny, hemicelulozy i glikoproteiny są przenoszone na błonę w pęcherzykach dictyosomów. Jednocześnie pektyny są bardziej charakterystyczne dla komórek rosnących, podczas gdy hemicelulozy przeważają w komórkach nie rosnących.
Mikrofibryle celulozowe są syntetyzowane na powierzchni komórki za pomocą kompleksu enzymatycznego związanego z błoną plazmatyczną. Orientacja mikrofibryli jest kontrolowana przez mikrotubule znajdujące się w pobliżu wewnętrznej powierzchni błony plazmatycznej.
Plazmodesma. Są to cienkie nici cytoplazmy, które łączą protoplasty sąsiednich komórek. Plazmodesmy albo przechodzą przez ścianę komórkową w dowolnym miejscu, albo są skoncentrowane w pierwotnych polach porów lub w błonach między parami porów. Pod mikroskopem elektronowym plazmodesmy wyglądają jak wąskie kanały wyłożone błoną plazmatyczną. Wzdłuż osi kanału mniejsza cylindryczna rurka, desmotubula, rozciąga się od jednej komórki do drugiej, która komunikuje się z retikulum endoplazmatycznym obu sąsiednich komórek. Wiele plazmodesmy tworzy się podczas podziału komórki, kiedy rurkowa retikulum endoplazmatyczne jest pobierane przez rozwijającą się płytkę komórkową. Plazmodesmy mogą również tworzyć się w błonach komórek, które się nie dzielą. Struktury te zapewniają sprawne przenoszenie określonych substancji z komórki do komórki.
Podział komórek. W organizmach wielokomórkowych podział komórek wraz ze wzrostem ich wielkości jest sposobem wzrostu całego organizmu. Nowe komórki powstałe podczas podziału są podobne pod względem struktury i funkcji, zarówno do komórki macierzystej, jak i do siebie nawzajem. Proces podziału u eukariontów można podzielić na dwa częściowo nakładające się etapy: mitozę i cytokinezę.
Mitoza to tworzenie się dwóch jąder potomnych z jednego jądra, morfologicznie i genetycznie równoważnych sobie nawzajem. Cytokineza to podział cytoplazmatycznej części komórki z utworzeniem komórek potomnych.
Cykl komórkowy.Żywa komórka przechodzi przez serię następujących po sobie zdarzeń, które składają się na cykl komórkowy. Czas trwania samego cyklu różni się w zależności od rodzaju komórki oraz czynników zewnętrznych, takich jak temperatura czy dostępność składników odżywczych. Zwykle cykl dzieli się na interfazę i cztery fazy mitozy.
Interfaza. Okres między kolejnymi podziałami mitotycznymi.
Interfaza jest podzielona na trzy okresy, oznaczone jako G1, S, G2.
W okresie G1, który rozpoczyna się po mitozie. W tym okresie wzrasta ilość cytoplazmy, w tym różnych organelli. Ponadto, zgodnie z obecną hipotezą, w okresie G1 syntetyzowane są substancje, które stymulują lub hamują okres S i resztę cyklu, determinując w ten sposób proces podziału.
Okres S następuje po okresie G1, w którym następuje duplikacja materiału genetycznego (DNA).
W okresie G2, który następuje po S, powstają struktury bezpośrednio zaangażowane w mitozę, takie jak elementy wrzeciona.
Niektóre komórki przechodzą nieograniczoną liczbę cykli komórkowych. Są to organizmy jednokomórkowe oraz niektóre komórki stref aktywnego wzrostu (merystemy). Niektóre wyspecjalizowane komórki po dojrzewaniu tracą zdolność do reprodukcji. Trzecia grupa komórek, na przykład te tworzące tkankę rany (kalus), zachowuje zdolność do podziału tylko w szczególnych warunkach.
Mitoza, lub rozszczepienie jądrowe. Jest to ciągły proces podzielony na cztery fazy: profaza, metafaza, anafaza, telofaza. W wyniku mitozy materiał genetyczny, który podwoił się w interfazie, jest równo dzielony między dwa jądra potomne.
Jedną z najwcześniejszych oznak przejścia komórki do podziału jest pojawienie się wąskiego, pierścieniowego pasma mikrotubul tuż pod błoną plazmatyczną. Ten stosunkowo gęsty pas otacza jądro w płaszczyźnie równikowej przyszłego wrzeciona mitotycznego. Ponieważ pojawia się przed profazą, nazywa się go pasem preprofazowym. Zanika po wrzecionie mitotycznym, na długo przed pojawieniem się w późnej telofazie płytki komórkowej, która rozrasta się od środka ku obrzeżom i łączy się z błoną komórki macierzystej w obszarze zajmowanym wcześniej przez pas preprofazowy.
profaza. Na początku profazy chromosomy przypominają długie nitki rozproszone wewnątrz jądra. Następnie, gdy nici skracają się i pogrubiają, można zobaczyć, że każdy chromosom składa się nie z jednej, ale z dwóch splecionych nici zwanych chromatydami. W późnej profazie dwie skrócone sparowane chromatydy każdego chromosomu leżą obok siebie równolegle, połączone wąskim odcinkiem zwanym centromerem. Ma określoną pozycję na każdym chromosomie i dzieli chromosom na dwa ramiona o różnej długości.
Mikrotubule znajdują się równolegle do powierzchni jądra wzdłuż osi wrzeciona. Jest to najwcześniejsza manifestacja zespołu wrzeciona mitotycznego.
Pod koniec profazy jąderko stopniowo traci swój wyraźny zarys i ostatecznie zanika. Wkrótce potem otoczka jądrowa również się rozpada.
Metafaza. Na początku metafazy wrzeciono, które jest strukturą trójwymiarową, najszerszą w środku i zwężającą się ku biegunom, zajmuje miejsce zajmowane wcześniej przez jądro. Włókna wrzeciona są wiązkami mikrotubul. Podczas metafazy chromosomy, z których każdy składa się z dwóch chromatyd, są ułożone w taki sposób, że ich centromery leżą w płaszczyźnie równikowej wrzeciona. Dzięki centromerowi każdy chromosom jest przyczepiony do nici wrzeciona. Jednak niektóre nici przechodzą z jednego bieguna na drugi bez przyczepiania się do chromosomów.
Kiedy wszystkie chromosomy znajdują się w płaszczyźnie równikowej, metafaza jest zakończona. Chromosomy są gotowe do podziału.
Anafaza. Chromatydy każdego chromosomu rozdzielają się. Teraz to są chromosomy potomne. Po pierwsze, centromer dzieli się, a dwa chromosomy potomne są przyciągane do przeciwnych biegunów. W tym przypadku centromery przesuwają się do przodu, a ramiona chromosomów rozciągają się do tyłu. Włókna wrzeciona przyczepione do chromosomów skracają się, przyczyniając się do rozdzielenia chromatyd i ruchu chromosomów potomnych w przeciwnych kierunkach.
telofaza. W telofazie następuje zakończenie rozdzielenia dwóch identycznych grup chromosomów, podczas gdy wokół każdej z nich tworzy się błona jądrowa. Siateczka szorstka bierze w tym czynny udział. Aparat wrzeciona znika. Podczas telofazy chromosomy tracą wyrazistość konturów, rozciągają się, zamieniając z powrotem w cienkie nitki. Jąderka są przywrócone. Kiedy chromosomy stają się niewidoczne, mitoza jest zakończona. Dwa jądra potomne wchodzą w interfazę. Są genetycznie równoważne sobie nawzajem i macierzystemu jądru. Jest to bardzo ważne, ponieważ program genetyczny, a wraz z nim wszystkie cechy, muszą zostać przeniesione na organizmy potomne.
Czas trwania mitozy jest różny u różnych organizmów i zależy od rodzaju tkanki. Jednak profaza jest najdłuższa, a anafaza najkrótsza. W komórkach wierzchołka korzenia czas trwania profazy wynosi 1–2 godziny; metafaza - 5 - 15 min; anafaza - 2 - 10 minut; telofaza - 10 - 30 min. Czas trwania interfazy wynosi od 12 do 30 godzin.
W wielu komórkach eukariotycznych centra organizacji mikrotubul odpowiedzialne za tworzenie wrzeciona mitotycznego są związane z centriolami.
Cytokineza. Jest to proces podziału cytoplazmy. U większości organizmów komórki dzielą się poprzez cofnięcie błony komórkowej i utworzenie bruzdy rozszczepienia, która stopniowo się pogłębia, ściskając pozostałe włókna wrzeciona mitotycznego.U wszystkich roślin (mszaków i naczyniowych) oraz u niektórych glonów komórki dzielą się w wyniku tworzenia się płytka komórkowa.
We wczesnej telofazie między dwoma jądrami potomnymi tworzy się beczkowaty układ włókien zwany phragmoplastem. Włókna phragmoplastu, podobnie jak włókna wrzeciona mitotycznego, składają się z mikrotubul. Małe krople pojawiają się w płaszczyźnie równikowej fragmoplastu. Łączą się, tworząc płytkę komórkową, która rośnie, aż dotrze do ściany dzielącej się komórki. To kończy rozdzielanie dwóch komórek potomnych. Łączące się kropelki to bąbelki odrywające się od aparatu Golgiego. Zasadniczo zawierają substancje pektynowe, z których powstaje środkowa płytka. Błony pęcherzyków biorą udział w budowie błony plazmatycznej po obu stronach płytki. W tym samym czasie plazmodesmy powstają z fragmentów rurkowatej retikulum endoplazmatycznego.
Po utworzeniu płytki środkowej każdy protoplast osadza na niej pierwotną błonę. Ponadto każda komórka potomna osadza nową warstwę błony wokół całego protoplastu, która kontynuuje błonę, która powstała z płytki komórkowej. Oryginalna powłoka komórki macierzystej jest niszczona w miarę wzrostu komórek potomnych.
Różne typy mitozy eukariotycznej
Opisany powyżej podział komórkowy u roślin, a także u zwierząt, nie jest jedyną formą pośredniego podziału komórkowego. Najprostszym rodzajem mitozy jest pleuromitoza. Przypomina binarne rozszczepienie komórek prokariotycznych, w którym nukleoidy po replikacji pozostają związane z błoną plazmatyczną. Błona zaczyna rosnąć między punktami wiązania DNA, a tym samym rozprzestrzenia chromosomy do różnych części komórki. Następnie, podczas tworzenia zwężenia komórki, każda z cząsteczek DNA znajdzie się w nowej oddzielnej komórce.
Charakterystyczne dla podziału komórek eukariotycznych jest tworzenie się wrzeciona zbudowanego z mikrotubul. Na zamknięta pleuromitoza(nazywa się to zamkniętym, ponieważ dywergencja chromosomów zachodzi bez naruszenia otoczki jądrowej) nie centriole, ale inne struktury znajdujące się po wewnętrznej stronie błony jądrowej uczestniczą jako centra organizacji mikrotubul (MCMT). Są to tak zwane ciała polarne o nieokreślonej morfologii, z których wychodzą mikrotubule. Są dwa takie ciała. Oddalają się od siebie nie tracąc kontaktu z otoczką jądrową. W wyniku tego powstają dwa półwrzeciona związane z chromosomami. Cały proces tworzenia wrzeciona mitotycznego i segregacji chromosomów zachodzi w tym przypadku pod otoczką jądrową. Ten typ mitozy występuje wśród pierwotniaków i jest szeroko rozpowszechniony u grzybów (chytridia, zygomycetes, drożdże, lęgniowce, ascomycetes, myxomycetes itp.). spotkać formy półzamknięta pleuromitoza gdy otoczka jądrowa zostaje zniszczona na biegunach uformowanego wrzeciona.
Następną formą mitozy jest ortomitoza. W tym przypadku COMT znajdują się w cytoplazmie, od samego początku nie tworzą się półwrzeciona, ale wrzeciono dwubiegunowe. Istnieją trzy formy ortomitoza ( normalna mitoza) półzamknięte I Zamknięte. W półzamkniętej ortomitozie powstaje bisymetryczne wrzeciono za pomocą TsOMT znajdującego się w cytoplazmie, otoczka jądrowa jest zachowana przez całą mitozę, z wyjątkiem stref polarnych. Masy materiału ziarnistego, a nawet centriole, mogą być wykrywane jako COMT. Ta forma mitozy występuje w zoosporach zielonych, brunatnych i czerwonych alg, w niektórych niższych grzybach i gregarynach. Przy zamkniętej ortomitozie błona jądrowa jest całkowicie zachowana, w której powstaje prawdziwe wrzeciono. Mikrotubule powstają w karioplazmie, rzadziej wyrastają z wewnętrznego TsOMT, który nie jest związany (w przeciwieństwie do pleuromitozy) z błoną jądrową. Ten typ mitozy jest typowy dla podziału mikrojąder orzęsków, ale może również wystąpić u pierwotniaków. W otwartej ortomitozie otoczka jądrowa całkowicie się rozpada. Ten typ podziału komórek jest charakterystyczny dla organizmów zwierzęcych, niektórych pierwotniaków i komórek roślin wyższych. Z kolei ta forma mitozy jest reprezentowana przez typy astralne i anastralne.
Z pokrótce przejrzanego materiału można zauważyć, że główną cechą mitozy w ogóle jest pojawianie się struktur wrzeciona rozszczepienia, które powstają w połączeniu z CMT, które różnią się budową.
Morfologia figury mitotycznej
Aparat mitotyczny jest szczególnie dobrze wyrażany na metafazowym etapie mitozy. W metafazie, w płaszczyźnie równikowej komórki, znajdują się chromosomy, z których wychodzą w przeciwnych kierunkach tzw. nici wrzecionowate, zbiegające się na dwóch różnych biegunach figury mitotycznej. Zatem wrzeciono mitotyczne jest zbiorem chromosomów, biegunów i włókien. Włókna wrzeciona to pojedyncze mikrotubule lub ich wiązki. Mikrotubule zaczynają się od biegunów wrzeciona, a niektóre z nich odchodzą do centromerów gdzie się znajdują kinetochory chromosomy (mikrotubule kinetochoru), niektóre przechodzą dalej w kierunku przeciwnego bieguna, ale go nie docierają. Nazywa się je „mikrotubulami międzybiegunowymi”. Od biegunów odchodzi grupa promienistych mikrotubul, tworząc wokół nich strukturę przypominającą „promienną poświatę” – są to mikrotubule astralne.
Pod względem morfologii figury mitotyczne dzielą się na typy astralne i anastralne.
Typ wrzeciona astralnego lub zbieżny charakteryzuje się tym, że jego bieguny są reprezentowane przez małą strefę, do której zbiegają się (zbiegają) mikrotubule. Zwykle centrosomy zawierające centriole znajdują się na biegunach wrzecion astralnych. Znane są jednak przypadki centralnych mitoz astralnych (podczas mejozy niektórych bezkręgowców). Ponadto odnotowuje się promieniste mikrotubule promieniujące z biegunów, które nie są częścią wrzeciona, ale tworzą strefy gwiaździste - cytastry. Ten typ podziału mitotycznego przypomina hantle. Anastrialny typ figury mitotycznej nie ma cytastrów na biegunach. Regiony polarne wrzeciona są tutaj szerokie, nazywane są czapami polarnymi, zawierają centriole. W tym przypadku włókna wrzeciona nie odbiegają od jednego punktu, ale rozchodzą się szerokim frontem (rozchodzą się) z całej strefy czap polarnych. Ten typ wrzeciona jest charakterystyczny dla dzielących się komórek roślin wyższych, ale można go również znaleźć u zwierząt wyższych. We wczesnej embriogenezie ssaków mitozy centriolarne (rozbieżne) obserwuje się podczas podziału dojrzewania oocytu oraz w I i II podziale zygoty. Ale już w trzecim podziale komórkowym i we wszystkich kolejnych komórkach dzielą się z udziałem wrzecion astralnych, w biegunach których zawsze znajdują się centriole. Ogólnie rzecz biorąc, dla wszystkich form mitozy chromosomy z ich kinetoforami, ciałami polarnymi (centrosomami) i włóknami wrzeciona pozostają wspólnymi strukturami.
Centromery i kinetochory
Centromery mogą mieć różną lokalizację wzdłuż długości chromosomów. Centromery holocentryczne występują, gdy mikrotubule są połączone wzdłuż całego chromosomu (niektóre owady, nicienie, niektóre rośliny). Centromery monocentryczne - gdy mikrotubule są połączone z chromosomami w jednym miejscu. centromery monocentryczne mogą być punktowe (na przykład u niektórych pączkujących drożdży), gdy tylko jedna mikrotubula zbliża się do kinetochoru, oraz strefowe, gdzie wiązka mikrotubul zbliża się do złożonego kinetochoru. Pomimo różnorodności stref centromerowych, wszystkie z nich są związane ze złożoną strukturą kinetochoru, który ma fundamentalne podobieństwo w budowie i funkcjach u wszystkich eukariontów. Kinetochory to specjalne struktury białkowe, zlokalizowane głównie w strefach centromerowych chromosomów. Są to złożone kompleksy składające się z wielu białek. morfologicznie są bardzo podobne, mają taką samą budowę, od okrzemek po ludzi. Są to struktury trójwarstwowe: wewnętrzna gęsta warstwa przylegająca do ciała chromosomu, środkowa luźna warstwa i zewnętrzna gęsta warstwa. Z warstwy zewnętrznej rozciąga się wiele włókienek, tworząc tzw. włóknistą koronę kinetochoru. W ogólnej postaci kinetochory wyglądają jak płytki lub dyski leżące w strefie pierwotnego zwężenia chromosomu, w centromerze. Zwykle na chromosom lub chromatydę przypada jeden kinetochor. Przed anafazą na każdej siostrzanej chromatydzie znajdują się kinetochory, z których każda komunikuje się z własną wiązką mikrotubul. W roślinach kinetochor nie wygląda jak talerze, ale półkule. Funkcjonalną rolą kinetochorów jest wiązanie ze sobą siostrzanych chromatyd, wiązanie mikrotubul mitotycznych, regulacja separacji chromosomów i faktyczne przemieszczanie chromosomów podczas mitozy z udziałem mikrotubul. Ogólnie rzecz biorąc, struktury białkowe, kinetochory, podwajają się w okresie S, równolegle do podwajania chromosomów. Ale ich białka są obecne na chromosomach we wszystkich okresach cyklu komórkowego.
TKANKA ROŚLINNA
Tkanki edukacyjne (merystemy)
Tkanki edukacyjne w ciele roślin znajdują się w różnych miejscach, dlatego dzieli się je na następujące grupy (ryc. 0;1).
Merystemy wierzchołkowe (wierzchołkowe) znajdują się na wierzchołkach lub wierzchołkach narządów osiowych - łodygi, korzenia. Za pomocą tych merystemów organy wegetatywne roślin przeprowadzają wzrost długości.
Merystemy boczne są charakterystyczne dla narządów osiowych. Tam są umieszczone koncentrycznie, w formie sprzęgła.
Merystemy interkalarne lub interkalarne pochodzą z merystemów wierzchołkowych. Są to grupy komórek, które nie są jeszcze zdolne do namnażania się, ale wkroczyły na ścieżkę różnicowania. Nie ma wśród nich komórek początkowych, ale jest wiele wyspecjalizowanych.
Merystemy ran zapewniają odbudowę uszkodzonej części ciała. Regeneracja zaczyna się od odróżnicowania, czyli odwrotnego rozwoju komórek wyspecjalizowanych do komórek merystematycznych. Zamieniają się w fellogen, który tworzy czop pokrywający powierzchnię rany. Komórki odróżnicowane, dzieląc się, mogą tworzyć luźną tkankę miąższową - kalus. Z niego, w określonych warunkach, powstają narządy roślinne.
Tkanki powłokowe
Pełnią rolę bariery granicznej, oddzielającej leżące poniżej tkanki od środowiska. Powłoki pierwotne rośliny składają się wyłącznie z żywych komórek. Powłoki drugorzędowe i trzeciorzędowe - głównie z martwych z grubymi ścianami komórkowymi.
Główne funkcje tkanek powłokowych:
ochrona rośliny przed wysychaniem;
ochrona przed wnikaniem szkodliwych mikroorganizmów;
ochrona przed oparzeniami słonecznymi;
ochrona przed uszkodzeniami mechanicznymi;
regulacja metabolizmu między rośliną a środowiskiem;
wrażenie irytacji.
Podstawową tkanką powłokową jest naskórek, naskórek. Składa się z żywych komórek. Utworzony z merystemów wierzchołkowych. Obejmuje młode rosnące łodygi i liście.
Naskórek formował się u roślin w związku z wyjściem ze środowiska wodnego na ląd w celu zapobieżenia jego wysychaniu. Oprócz aparatów szparkowych wszystkie komórki naskórka są ze sobą ściśle połączone. Zewnętrzne ściany głównych komórek są grubsze niż pozostałe. Cała powierzchnia pokryta jest warstwą kutyny i wosków roślinnych. Ta warstwa nazywana jest naskórkiem (skórą). Nie występuje na rosnących korzeniach i podwodnych częściach roślin. Po wyschnięciu przepuszczalność łuski jest znacznie osłabiona.
Oprócz głównych komórek w naskórku znajdują się inne, w szczególności włosy lub trichomy. Są jednokomórkowe i wielokomórkowe (Rys. 2). Funkcjonalnie zwiększają powierzchnię naskórka, np. w strefie wzrostu korzenia, służą jako ochrona mechaniczna, przylegają do podpory i ograniczają utratę wody. Wiele roślin ma gruczołowe włosy, takie jak pokrzywy.
Tylko rośliny wyższe mają aparaty szparkowe w epidermie, które regulują wymianę wody i gazów. Jeśli nie ma naskórka, nie ma potrzeby stosowania aparatów szparkowych. Aparaty szparkowe to grupa komórek tworzących aparat szparkowy, który składa się z dwóch komórek spływowych oraz sąsiadujących ze sobą komórek naskórka – komórek bocznych. Różnią się od głównych komórek naskórka (ryc.3). Komórki ochronne różnią się od otaczających je kształtem i obecnością dużej liczby chloroplastów oraz nierównomiernie pogrubionych ścian. Te, które są naprzeciw siebie, są grubsze niż pozostałe (rys. 4). Pomiędzy komórkami ochronnymi tworzy się szczelina szparkowa, która prowadzi do przestrzeni podszparkowej zwanej jamą podszparkową. Komórki ochronne są wysoce fotosyntetyzujące. Zawierają dużą ilość skrobi rezerwowej oraz liczne mitochondria.
Liczba i rozmieszczenie aparatów szparkowych, rodzajów aparatów szparkowych różni się znacznie w różnych roślinach. U współczesnych mszaków nie ma aparatów szparkowych. Fotosynteza w nich jest przeprowadzana przez pokolenie gametofitów, a sporofity nie są zdolne do niezależnego istnienia.
Zwykle aparaty szparkowe znajdują się na spodniej stronie liścia. U roślin pływających na powierzchni wody - na górnej powierzchni. W liściach zbóż aparaty szparkowe są często równomiernie rozmieszczone po obu stronach. Takie liście są oświetlane w miarę równomiernie. Na 1 mm2 powierzchni może znajdować się od 100 do 700 aparatów szparkowych.
Wtórna tkanka powłokowa (peryderma). Tkanka ta zastępuje naskórek, gdy zielony kolor jednorocznych pędów zmienia się na brązowy. Jest wielowarstwowy i składa się z centralnej warstwy komórek kambium - fellogenu. Komórki felogenu, dzieląc się, układają warstwę fellemu na zewnątrz, a felodermy od wewnątrz. (Rys.5).
Phelema lub korek. Początkowo składa się z żywych, cienkościennych komórek. Z biegiem czasu ich ściany są impregnowane suberyną i woskami roślinnymi i obumierają. Zawartość komórki jest wypełniona powietrzem.
Funkcje Fellema:
zapobiega utracie wilgoci;
chroni roślinę przed uszkodzeniami mechanicznymi;
chroni przed patogenami;
zapewnia izolację termiczną, ponieważ komórki są wypełnione powietrzem.
Komórki felogenu, zlokalizowane w samym naskórku, leżącej pod nim warstwie podnaskórkowej, rzadziej w głębokich warstwach kory pierwotnej, stanowią podstawę generacyjną kory pierwotnej.
Warstwa korka nie jest trwała. Występują w nim przerwy, które komunikują się z znajdującymi się w pobliżu przestrzeniami międzykomórkowymi. W tym samym czasie na powierzchni powstają małe guzki - soczewica, które komunikują przestrzenie międzykomórkowe z powietrzem atmosferycznym. (ryc.6,7).
Jesienią fallogen pod przetchlinkami układa warstwę zakorkowanych komórek, które znacznie ograniczają transpirację, ale jej całkowicie nie wykluczają. Wiosną warstwa ta zapada się od wewnątrz. Na jasnej korze brzozy soczewica jest wyraźnie widoczna w postaci ciemnych kresek.
Trzeciorzędowa tkanka powłokowa (skorupa) jest również charakterystyczna tylko dla drzewiastych form roślin.
Phellogen jest wielokrotnie odkładany w głębszych warstwach kory. Tkanki znajdujące się na zewnątrz ostatecznie obumierają, tworząc skorupę. Jego komórki są martwe i niezdolne do rozciągania. Jednak żywe komórki położone głębiej dzielą się, co prowadzi do zwiększenia poprzecznego rozmiaru łodygi. Z biegiem czasu zewnętrzna warstwa skorupy pęka. Czas wystąpienia takiej przerwy jest wartością dość stałą dla poszczególnych roślin. Na jabłoni dzieje się to w siódmym roku życia, w grabie - w pięćdziesiątym. U niektórych gatunków nie występuje w ogóle. Główną funkcją peelingu jest ochrona przed uszkodzeniami mechanicznymi i termicznymi.
Miąższ
Jest to grupa wyspecjalizowanych tkanek, które wypełniają przestrzenie wewnątrz ciała rośliny pomiędzy tkankami przewodzącymi i mechanicznymi. (Rys. 8). Częściej komórki miąższu mają kształt zaokrąglony, rzadziej wydłużony. Charakterystyczna jest obecność rozwiniętych przestrzeni międzykomórkowych. Przestrzenie między komórkami tworzą razem system transportowy - apoplast. Ponadto przestrzenie międzykomórkowe tworzą „system wentylacyjny” rośliny. Poprzez aparaty szparkowe lub przetchlinki łączą się z powietrzem atmosferycznym i zapewniają optymalny skład gazów wewnątrz rośliny. Rozwinięte przestrzenie międzykomórkowe są szczególnie potrzebne roślinom rosnącym na glebach podmokłych, gdzie normalna wymiana gazowa jest utrudniona. Ten miąższ nazywa się aerenchymą. (Rys. 9).
Elementy miąższu, wypełniając szczeliny między innymi tkankami, pełnią również funkcję podporową. Komórki miąższu są żywe, nie mają grubych ścian komórkowych, jak sklerenchyma. Dlatego właściwości mechaniczne zapewnia turgor. Jeśli zawartość wody spada, co prowadzi do plazmolizy i więdnięcia rośliny.
Miąższ asymilacyjny tworzą cienkościenne komórki z wieloma przestrzeniami międzykomórkowymi. Komórki tej struktury zawierają wiele chloroplastów, dlatego nazywa się to chlorenchymą. Chloroplasty znajdują się wzdłuż ściany bez cieniowania się nawzajem. W miąższu asymilacyjnym zachodzą reakcje fotosyntezy, które dostarczają roślinie substancji organicznych i energii. Wynikiem procesów fotosyntezy jest możliwość istnienia wszystkich żywych organizmów na Ziemi.
Tkanki asymilacyjne występują tylko w oświetlonych częściach rośliny, oddzielone są od otoczenia przezroczystym naskórkiem. Jeśli naskórek zostanie zastąpiony nieprzezroczystymi wtórnymi tkankami powłokowymi, miąższ asymilacyjny zanika.
Miąższ spichrzowy służy jako pojemnik na substancje organiczne, które nie są czasowo wykorzystywane przez organizm roślinny. W zasadzie każda komórka z żywym protoplastem jest zdolna do odkładania materii organicznej w postaci różnego rodzaju inkluzji, ale niektóre komórki się w tym specjalizują. (Rysunek 10, 11). Związki bogate w energię odkładają się tylko w sezonie wegetacyjnym, zużywają w okresie spoczynku oraz w ramach przygotowań do kolejnego sezonu wegetacyjnego. Dlatego substancje rezerwowe osadzają się w organach wegetatywnych tylko u roślin wieloletnich.
Zbiornikiem wywarów mogą być zarówno narządy zwykłe (łodyga, korzeń), jak i specjalistyczne (kłącza, bulwy, cebulki). Wszystkie rośliny nasienne przechowują w nasionach cenne energetycznie substancje (liścienie, bielmo). Wiele roślin w suchym klimacie magazynuje nie tylko materię organiczną, ale także wodę. (Rys. 12). Na przykład aloes magazynuje wodę w mięsistych liściach, kaktusy w pędach.
tkaniny mechaniczne
Właściwości mechaniczne komórek roślinnych zapewniają:
twarda skorupa komórki;
turgor, czyli stan turgoru komórek.
Pomimo tego, że prawie wszystkie komórki tkanki mają właściwości mechaniczne, to jednak w roślinie występują tkanki, dla których właściwości mechaniczne są podstawowe. Są to collenchyma i sklerenchyma. Zwykle działają w interakcji z innymi tkankami. Wewnątrz ciała rośliny tworzą rodzaj ramy. Dlatego nazywa się je wzmocnieniem.
Nie wszystkie rośliny mają równie dobrze zdefiniowane tkanki mechaniczne. W znacznie mniejszym stopniu rośliny żyjące w środowisku wodnym potrzebują wewnętrznego wsparcia niż rośliny lądowe. Powodem jest to, że rośliny wodne potrzebują mniej wsparcia wewnętrznego. Ich ciało jest w dużej mierze podtrzymywane przez otaczającą wodę. Powietrze na lądzie nie tworzy takiego wsparcia, ponieważ ma mniejszą gęstość niż woda. Z tego powodu obecność wyspecjalizowanych tkanek mechanicznych staje się istotna.
Udoskonalenie wewnętrznych struktur wsparcia nastąpiło w procesie ewolucji.
Collenchyma. Tworzą go tylko żywe komórki, wydłużone wzdłuż osi narządu. Ten typ tkanki mechanicznej powstaje bardzo wcześnie, w okresie pierwotnego wzrostu. Dlatego ważne jest, aby komórki pozostały żywe i zachowały zdolność do rozciągania się wraz z rozciągającymi się komórkami, które są w pobliżu.
Cechy komórek collenchyma:
nierównomierne pogrubienie skorupy, w wyniku czego niektóre jej części pozostają cienkie, a inne pogrubiają;
muszle nie zdrewniały.
Komórki Collenchyma są rozmieszczone w różny sposób względem siebie. W sąsiednich komórkach na rogach skierowanych do siebie tworzą się zgrubienia ( Ryż. 13). Taka kollenchyma nazywana jest kollenchymą narożną. W innym przypadku komórki są ułożone w równoległe warstwy. Błony komórkowe zwrócone w stronę tych warstw są silnie pogrubione. To jest kolenchyma blaszkowata. Komórki mogą być ułożone luźno, z obfitymi przestrzeniami międzykomórkowymi - jest to luźna kolenchyma. Kollenchyma taka często występuje w roślinach na glebach podmokłych.
Collenchyma ma szczególne znaczenie w młodych roślinach, formach zielnych, a także w częściach roślin, w których nie występuje wzrost wtórny, takich jak liście. W tym przypadku układa się go bardzo blisko powierzchni, czasem tuż pod naskórkiem. Jeśli narząd ma twarze, wzdłuż ich grzebieni znajdują się grube warstwy kolenchymy.
Komórki Collenchyma są funkcjonalne tylko w obecności turgoru. Niedobór wody zmniejsza skuteczność kolenchymy i roślina przejściowo więdnie, np. liście ogórka zwisają w upalny dzień. Po napełnieniu komórek wodą przywracane są funkcje kolenchymy.
sklerenchyma. Drugi rodzaj tkanek mechanicznych. W przeciwieństwie do kolenchymy, gdzie wszystkie komórki są żywe, komórki sklerenchymy są martwe. Ich ściany są bardzo grube. Pełnią funkcję mechaniczną. Silne pogrubienie błony prowadzi do naruszenia transportu substancji, w wyniku czego protoplast umiera ( Ryż. 14). Lignifikacja błon komórkowych sklerenchymy następuje, gdy organ rośliny zakończył już swój wzrost. Dzięki temu nie przeszkadzają już w rozciąganiu otaczających tkanek.
W zależności od kształtu wyróżnia się dwa rodzaje komórek sklerenchymy - włókna i sklereidy.
Włókna mają silnie wydłużony kształt z bardzo grubymi ściankami i małą wnęką. Są nieco mniejsze niż włókna drzewne. Często pod naskórkiem tworzą podłużne warstwy i pasma. W łyku lub ksylemie można je znaleźć pojedynczo lub w grupach. W łyku nazywane są włóknami łykowymi ( Ryż. 15), aw ksylemie - włókna libriform ( Ryż. 16).
Sclereidy lub komórki kamieniste są reprezentowane przez zaokrąglone lub rozgałęzione komórki z potężnymi błonami. W ciele rośliny można je znaleźć pojedynczo (komórki podporowe) ( rys.17) lub grupy (ryc. 18). Należy zauważyć, że właściwości mechaniczne silnie zależą od lokalizacji sklereidów. Część sklereidów tworzy ciągłe warstwy, jak na przykład w łupinach orzechów lub w pestkach owoców (owoce pestkowe).
tkanki wydalnicze
Każda żywa komórka początkowo ma funkcję usuwania substancji, podczas gdy transport może przebiegać zarówno wzdłuż gradientu stężeń, jak i pod gradient. Substancje, które zostały zsyntetyzowane w komórce, mogą zostać usunięte, a następnie wpłyną na aktywność innych komórek (fitohormony, enzymy). Ten proces nazywa się wydzielaniem. W przypadku wydalania substancji będących produktami przemiany materii komórki proces ten nazywamy wydalaniem. Pomimo faktu, że wydalanie substancji w komórkach roślinnych i zwierzęcych ma zasadnicze podobieństwo, rośliny mają szereg cech, które wynikają z fundamentalnych różnic w aktywności życiowej. Poziom reakcji metabolicznych u roślin jest znacznie niższy niż u zwierząt. W związku z tym emitowane są proporcjonalnie mniejsze ilości odpadów. Inną cechą jest to, że samodzielnie syntetyzując prawie wszystkie niezbędne związki organiczne, rośliny nigdy nie tworzą ich nadmiernych zapasów. Uwolnione substancje mogą służyć jako materiał wyjściowy do innych reakcji (na przykład CO2 i H2O).
Jeśli u zwierząt proces wydalania toksyn wiąże się z ich usuwaniem z organizmu, rośliny mogą tego nie robić, izolując zbędne substancje w żywym protoplastie (usuwanie różnych substancji do wakuoli), w komórkach martwych (większość tkanek rośliny wieloletniej rośliny składają się z takich komórek), w przestrzeniach międzykomórkowych.
Emitowane substancje można podzielić na dwie duże grupy. Pierwsza grupa to substancje organiczne syntetyzowane bezpośrednio przez komórkę (enzymy, polisacharydy, ligniny, terpeny, te drugie są składnikami olejków eterycznych i żywic). Druga grupa - substancje, które początkowo dostają się do komórki z zewnątrz za pomocą transportu ksylemu lub łyka (woda, sole mineralne, aminokwasy, monosacharydy itp.). Wydalany sekret rzadko jest jednorodny i zwykle składa się z mieszaniny, w której jedna substancja ma najwyższe stężenie.
Rośliny, w przeciwieństwie do zwierząt, nie mają kompletnego systemu wydalniczego. Po całej roślinie rozsiane są jedynie wyspecjalizowane struktury – idioblasty. Tworzące je komórki są mniejsze niż leżące w pobliżu komórki miąższu. Mają gęstą elektronowo cytoplazmę z rozwiniętymi elementami retikulum endoplazmatycznego i kompleksu Golgiego. Najczęściej nie wyrażona, jest centralna wakuola. Między sobą a innymi żywymi komórkami komórki te są połączone licznymi plazmodesmami.
W zależności od lokalizacji tkanki wydalnicze mogą być zewnętrzne i wewnętrzne.
Zewnętrzne tkanki wydalnicze. Włosy gruczołowe są pochodnymi naskórka. Morfologicznie są zmienne – mogą mieć wielokomórkową głowę, być wydłużone, w formie tarczy na nodze itp. (rys. 19). Należą do nich włosy pokrzywy.
Hydatody to struktury, które uwalniają nadmiar wody w warunkach niskiej transpiracji i wysokiej wilgotności – gutacji. Struktury te tworzą grupy bezbarwnych żywych komórek o cienkich ścianach - epitem. Tkanka ta sąsiaduje z wiązką przewodzącą (ryc. 20). Woda jest uwalniana przez specjalne aparaty szparkowe, które różnią się od zwykłych aparatów szparkowych bezruchem i stale otwartą szczeliną. Skład płynu gutacyjnego jest bardzo zróżnicowany, od prawie czystej wody po bardzo złożoną mieszaninę substancji.
Nektarniki. Wiele roślin charakteryzuje się wydzielaniem cieczy zawierającej od 7 do 87% mono- i oligosacharydów. Proces ten przeprowadzają specjalne struktury - nektarniki. W zależności od lokalizacji wyróżnia się nektarniki kwiatowe lub kwiatowe (ryc. 21), a także rośliny znajdujące się na łodygach i liściach - nektarniki pozakwiatowe lub pozakwiatowe. Nektarem może być niezmieniony sok z łyka, który jest dostarczany na powierzchnię przez przestrzenie międzykomórkowe i wydalany przez aparaty szparkowe. Bardziej złożone nektary tworzą miąższ gruczołowy, pokryty naskórkiem z włoskami gruczołowymi. Nektar jest wydalany przez komórki naskórka lub włosy gruczołowe. W tym przypadku wydzielany nektar różni się od soku z łyka. Dominuje bowiem glukoza, fruktoza, sacharoza, aw soku z łyka - glukoza. Jony występują w niskich stężeniach. Jony występują w niskich stężeniach. Aby przyciągnąć owady zapylające, nektar może zawierać hormony steroidowe, które są niezbędne owadom.
Gruczoły trawienne są obecne w roślinach owadożernych (rosiczka, nepenthys itp.). W odpowiedzi na dotyk małych bezkręgowców wydzielają wydzielinę zawierającą enzymy hydrolityczne, kwas mrówkowy, masłowy i propionowy. W efekcie biopolimery rozkładają się na monomery, które roślina wykorzystuje na swoje potrzeby.
Gruczoły solne rozwijają się u roślin rosnących na glebach zasolonych. Gruczoły te znajdują się na liściach i w postaci rozpuszczonej nadmiar jonów wydostaje się na powierzchnię. Sól najpierw osadza się na naskórku, a następnie jest zmywana przez deszcz (ryc. 22). Ponadto wiele roślin ma solne włosy na liściach. Każdy włos składa się z dwóch komórek: jedna tworzy głowę, druga - łodygę. Sole gromadzą się w wakuoli komórki głowy, a gdy ich stężenie staje się wysokie, głowa odpada. W jej miejsce rośnie nowa komórka, która również gromadzi sole. Podczas wzrostu liścia głowa może spaść i uformować się kilka razy. Zaletą solonych włosów jest to, że uwalniając sole, tracą mało wody. Zjawisko to jest bardzo ważne dla roślin rosnących na glebach zasolonych, gdzie brakuje słodkiej wody.
Wewnętrzne tkanki wydalnicze. Wewnętrzne struktury wydalnicze, rozproszone po całym ciele rośliny w postaci idioblastów, nie usuwają substancji poza organizm, gromadząc je w sobie. Idioblasty to komórki roślinne różniące się kształtem, strukturą lub zawartością od innych komórek w tej samej tkance. W zależności od pochodzenia rozróżnia się schizogenne (gr. schiso - rozszczepienie) i lizogenne (gr. liza - rozpuszczenie) zbiorniki wydzielanych substancji (ryc. 23).
Naczynia schizogeniczne. Są to mniej lub bardziej rozległe przestrzenie międzykomórkowe wypełnione wydalanymi substancjami. Na przykład przejścia żywiczne z iglaków, aralii, parasoli i innych. Uważa się, że żywica ma właściwości bakteriobójcze, odstrasza roślinożerców, czyniąc rośliny niejadalnymi dla nich.
Naczynia lizogeniczne powstają w miejscu żywych komórek, które obumierają i ulegają zniszczeniu po nagromadzeniu w nich substancji. Takie formacje można zaobserwować w skórce
STRUKTURA KOMÓRKI ROŚLINNEJ
Komórka roślinna składa się z mniej lub bardziej sztywnych Ściana komórkowa I prototyp.Ściana komórkowa to ściana komórkowa i błona cytoplazmatyczna. Termin protoplast pochodzi od słowa protoplazma, które od dawna jest używane w odniesieniu do wszystkich żywych istot. Protoplast jest protoplazmą pojedynczej komórki.
Protoplast zbudowany jest z cytoplazma I jądra. Cytoplazma zawiera organelle (rybosomy, mikrotubule, plastydy, mitochondria) i układy membranowe (retikulum endoplazmatyczne, dictyosomy). Cytoplazma obejmuje również macierz cytoplazmatyczną ( substancja podstawowa), w których zanurzone są organelle i układy membranowe. Cytoplazma jest oddzielona od ściany komórkowej błona plazmatyczna, która jest membraną elementarną. W przeciwieństwie do większości komórek zwierzęcych, komórki roślinne zawierają jeden lub więcej wakuole. Są to wypełnione płynem pęcherzyki otoczone elementarną błoną ( tonoplast).
W żywej komórce roślinnej substancja podstawowa jest w ciągłym ruchu. W ruchu zwanym prądem cytoplazmatycznym lub cykloza, biorą udział organelle. Cykloza ułatwia przemieszczanie się substancji w komórce i ich wymianę między komórką a środowiskiem.
Membrana plazmowa. Jest to dwuwarstwowa struktura fosfolipidowa. Komórki roślinne charakteryzują się inwazjami błony plazmatycznej.
Błona plazmatyczna pełni następujące funkcje:
Uczestniczy w wymianie substancji między komórką a środowiskiem;
Koordynuje syntezę i montaż mikrofibryli celulozowych ściany komórkowej;
Przekazuje sygnały hormonalne i zewnętrzne, które kontrolują wzrost i różnicowanie komórek.
Rdzeń. Jest to najbardziej widoczna struktura w cytoplazmie komórki eukariotycznej. Jądro spełnia dwie ważne funkcje:
Kontroluje aktywność życiową komórki, określając, które białka iw jakim czasie powinny zostać zsyntetyzowane;
Przechowuje informacje genetyczne i przekazuje je komórkom potomnym podczas podziału komórki.
Jądro komórki eukariotycznej jest otoczone przez dwie elementarne błony, które się tworzą otoczka jądrowa. Przesiąknięta jest licznymi porami o średnicy od 30 do 100 nm, widocznymi jedynie pod mikroskopem elektronowym. Pory mają złożoną strukturę. Zewnętrzna błona błony jądrowej w niektórych miejscach jest połączona z retikulum endoplazmatycznym. Otoczkę jądrową można postrzegać jako wyspecjalizowaną, lokalnie zróżnicowaną część retikulum endoplazmatycznego (ER).
W rdzeniu barwionym specjalnymi barwnikami można wyróżnić cienkie nitki i grudki chromatyna I nukleoplazma(główna substancja jądra). Chromatyna składa się z DNA związanego ze specjalnymi białkami zwanymi histonami. W procesie podziału komórki chromatyna staje się coraz bardziej gęsta i gromadzi się chromosomy. DNA koduje informację genetyczną.
Organizmy różnią się liczbą chromosomów w komórkach somatycznych. Na przykład kapusta ma - 20 chromosomów; słonecznik - 34; pszenica - 42; ludzie - 46, a jeden z gatunków paproci Ophioglossum – 1250. Komórki płciowe (gamety) mają tylko połowę liczby chromosomów charakterystycznych dla komórek somatycznych organizmu. Nazywa się liczbę chromosomów w gametach haploidalny(pojedynczy), w komórkach somatycznych - diploidalny(podwójnie). Komórki posiadające więcej niż dwa zestawy chromosomów to tzw poliploidalny.
Pod mikroskopem świetlnym można zobaczyć kuliste struktury - jąderka. Każde jądro ma jedno lub więcej jąderek, które są widoczne w jądrach niedzielących się. Rybosomalny RNA jest syntetyzowany w jąderku. Zazwyczaj jądra organizmów diploidalnych mają dwa jąderka, po jednym dla każdego haploidalnego zestawu chromosomów. Jąderka nie mają własnej błony. Pod względem biochemicznym jąderka charakteryzują się wysokim stężeniem RNA, które jest tu związane z fosfoproteinami. Wielkość jąderka zależy od stanu funkcjonalnego komórki. zauważono, że w szybko rosnącej komórce, w której zachodzą intensywne procesy syntezy białek, jąderka powiększają się. W jąderkach powstają mRNA i rybosomy, które pełnią funkcję syntetyczną tylko w jądrze.
Nukleoplazma (karioplazma) jest reprezentowana przez jednorodną ciecz, w której rozpuszczają się różne białka, w tym enzymy.
Plastydy. Vacuole, celulozowa ściana komórkowa i plastydy są charakterystycznymi składnikami komórek roślinnych. Każdy plastyd ma swoją własną powłokę, składającą się z dwóch elementarnych membran. Wewnątrz plastydu wyróżnia się układ membranowy i różny stopień jednorodności substancji - zrąb. Dojrzałe plastydy są klasyfikowane na podstawie zawartych w nich pigmentów.
chloroplasty, w których zachodzi fotosynteza, zawierają chlorofile i karotenoidy. Zwykle mają postać krążka o średnicy 4 - 5 mikronów. W jednej komórce mezofilu (środek liścia) może znajdować się 40 - 50 chloroplastów; w arkuszach mm 2 - około 500 000. w cytoplazmie chloroplasty są zwykle zlokalizowane równolegle do błony komórkowej.
Wewnętrzna struktura chloroplastu jest złożona. Podścielisko jest przesiąknięte rozwiniętym systemem błon w postaci pęcherzyków - tylakoidy. Każdy tylakoid składa się z dwóch błon. Tylakoidy tworzą jeden system. Z reguły są one gromadzone w stosach - ziarna przypominające kolumny monet. Tylakoidy poszczególnych granów są połączone tylakoidami zrębu lub tylakoidami międzyziarnistymi. Chlorofil i karotenoidy są osadzone w błonach tylakoidów. Chloroplasty w zielonych roślinach i algach często zawierają ziarna skrobi i małe kropelki lipidów (tłuszczu). Ziarna skrobi są tymczasowymi magazynami produktów fotosyntezy. Mogą zniknąć z chloroplastów rośliny, która była w ciemności tylko przez 24 godziny, i pojawić się ponownie 3-4 godziny po przeniesieniu rośliny na światło.
Chloroplasty to półautonomiczne organelle i przypominają bakterie. Na przykład rybosomy bakterii i chloroplastów mają dość duże podobieństwo. Są mniejsze niż rybosomy eukariotyczne. Synteza białek na rybosomach bakterii i chloroplastach jest hamowana przez chloramfenikol, który nie ma wpływu na komórki eukariotyczne. Ponadto zarówno bakterie, jak i chloroplasty mają podobny typ nukleoidu, zorganizowanego w podobny sposób. Pomimo faktu, że tworzenie chloroplastów i synteza zawartych w nich pigmentów jest w dużej mierze kontrolowana przez chromosomalny DNA komórki, to jednak przy braku własnego DNA chloroplasty nie tworzą się.
Chloroplasty można uznać za główne organelle komórkowe, ponieważ są pierwszymi w łańcuchu transformacji energii słonecznej, w wyniku czego ludzkość otrzymuje zarówno żywność, jak i paliwo. W chloroplastach zachodzi nie tylko fotosynteza. Biorą również udział w syntezie aminokwasów i kwasów tłuszczowych, służą jako magazyn tymczasowych rezerw skrobi.
Chromoplasty - plastydy pigmentowe. Różnorodne w formie, nie posiadają chlorofilu, ale syntetyzują i gromadzą karotenoidy, które nadają żółty, pomarańczowy, czerwony kolor kwiatom, starym liściom, owocom i korzeniom. chromoplasty mogą rozwijać się z chloroplastów, które w tym przypadku tracą chlorofil i struktury błony wewnętrznej, akumulują karotenoidy. Dzieje się tak podczas dojrzewania wielu owoców. Chromoplasty przyciągają owady i inne zwierzęta, z którymi współewoluowały.
Leukoplasty - niepigmentowane plastydy. Niektóre z nich syntetyzują skrobię ( amyloplastów), inne są zdolne do tworzenia różnych substancji, w tym lipidów i białek. W świetle leukoplasty zamieniają się w chloroplasty.
proplastydy - małe, bezbarwne lub bladozielone niezróżnicowane plastydy, które znajdują się w merystematycznych (dzielących się) komórkach korzeni i pędów. Są prekursorami innych, bardziej zróżnicowanych plastydów - chloroplastów, chromoplastów i aminoplastów. Jeśli rozwój protoplastów jest opóźniony z powodu braku światła, jeden lub więcej ciałka prolamelarne, które są półkrystalicznymi nagromadzeniami rurkowatych membran. Plastydy zawierające ciałka prolamelarne to tzw etioplasty. W świetle etioplasty zamieniają się w chloroplasty, podczas gdy błony ciał prolamelarnych tworzą tylakoidy. Etioplasty powstają w liściach roślin, które są w ciemności. protoplasty zarodków nasiennych najpierw przekształcają się w etioplasty, z których następnie w świetle rozwijają się chloroplasty. Plastydy charakteryzują się stosunkowo łatwymi przejściami z jednego rodzaju do drugiego. Plastydy, podobnie jak bakterie, rozmnażają się, dzieląc się na dwie części. W komórkach merystematycznych czas podziału protoplastów w przybliżeniu pokrywa się z czasem podziału komórki. Jednak w dojrzałych komórkach większość plastydów powstaje w wyniku podziału dojrzałych plastydów.
mitochondria. Podobnie jak chloroplasty, mitochondria są otoczone dwiema podstawowymi błonami. Błona wewnętrzna tworzy wiele fałd i wypukłości - Chrystus, które znacznie zwiększają wewnętrzną powierzchnię mitochondriów. Są znacznie mniejsze niż plastydy, mają średnicę około 0,5 µm oraz zmienną długość i kształt.
W mitochondriach zachodzi proces oddychania, w wyniku którego dochodzi do rozpadu cząsteczek organicznych z uwolnieniem energii i przekazaniem jej do cząsteczek ATP, głównej rezerwy energetycznej wszystkich komórek eukariotycznych. Większość komórek roślinnych zawiera setki lub tysiące mitochondriów. Ich liczba w jednej komórce jest określona przez zapotrzebowanie komórki na ATP. Mitochondria są w ciągłym ruchu, przemieszczając się z jednej części komórki do drugiej, łącząc się ze sobą i dzieląc. Mitochondria zwykle gromadzą się tam, gdzie potrzebna jest energia. Jeśli błona plazmatyczna aktywnie przenosi substancje z komórki do komórki, wówczas mitochondria znajdują się wzdłuż powierzchni błony. W ruchliwych glonach jednokomórkowych mitochondria gromadzą się u podstawy wici, dostarczając energię niezbędną do ich ruchu.
Mitochondria, podobnie jak plastydy, są półautonomicznymi organellami zawierającymi składniki niezbędne do syntezy własnych białek. Błona wewnętrzna otacza płynną matrycę, która zawiera białka, RNA, DNA, bakteryjne rybosomy i różne substancje rozpuszczone. DNA istnieje w postaci kolistych cząsteczek zlokalizowanych w jednym lub kilku nukleoidach.
Na podstawie podobieństwa bakterii do mitochondriów i chloroplastów komórek eukariotycznych można przypuszczać, że mitochondria i chloroplasty wywodzą się od bakterii, które znalazły „schronienie” w większych komórkach heterotroficznych – prekursorach eukariontów.
Mikrociała. W przeciwieństwie do plastydów i mitochondriów, które są ograniczone dwiema błonami, mikrociało to kuliste organelle otoczone pojedynczą błoną. Mikrociała mają ziarnistą (granulowaną) zawartość, czasami zawierają również krystaliczne wtrącenia białkowe. Mikrociała są związane z jednym lub dwoma regionami retikulum endoplazmatycznego.
Niektóre mikrociała tzw proksysomy, odgrywają ważną rolę w metabolizmie kwasu glikolowego, który jest bezpośrednio związany z fotooddychaniem. W zielonych liściach są one związane z mitochondriami i chloroplastami. Inne mikroorganizmy, tzw glioksysomy, zawierają enzymy potrzebne do przekształcania tłuszczów w węglowodany. Dzieje się tak w przypadku wielu nasion podczas kiełkowania.
wakuole - Są to obszary komórki ograniczone błoną, wypełnione płynem - sok komórkowy. Są otoczeni tonoplast(membrana wakuolowa).
Młoda komórka roślinna zawiera liczne małe wakuole, które w miarę starzenia się łączą się w jedną dużą wakuolę. W dojrzałej komórce do 90% jej objętości może zajmować wakuola. W tym przypadku cytoplazma jest dociskana w postaci cienkiej warstwy obwodowej do błony komórkowej. Wzrost wielkości komórek wynika głównie ze wzrostu wakuoli. W rezultacie powstaje ciśnienie turgoru i zachowana jest elastyczność tkanki. Jest to jedna z głównych funkcji wakuoli i tonoplastu.
Głównym składnikiem soku jest woda, reszta zależy od rodzaju rośliny i jej stanu fizjologicznego. Wakuole zawierają sole, cukry, rzadziej białka. Tonoplast odgrywa aktywną rolę w transporcie i gromadzeniu się niektórych jonów w wakuoli. Stężenie jonów w soku komórkowym może znacznie przekraczać jego stężenie w środowisku. Przy wysokiej zawartości niektórych substancji w wakuolach tworzą się kryształy. Najczęściej spotykane są kryształy szczawianu wapnia, które mają inny kształt.
Wakuole są miejscami gromadzenia się produktów przemiany materii (metabolizmu). Mogą to być białka, kwasy, a nawet substancje toksyczne dla człowieka (alkaloidy). Często osadzają się pigmenty. Niebieski, fioletowy, purpurowy, ciemnoczerwony, karmazynowy dają komórkom roślinnym pigmenty z grupy antocyjanów. W przeciwieństwie do innych pigmentów dobrze rozpuszczają się w wodzie i znajdują się w soku komórkowym. Decydują o czerwonym i niebieskim zabarwieniu wielu warzyw (rzodkiewki, rzepa, kapusta), owoców (winogrona, śliwki, wiśnie), kwiatów (chabry, pelargonie, ostróżki, róże, piwonie). Czasami te pigmenty maskują chlorofil w liściach, na przykład w ozdobnym czerwonym klonie. Antocyjany barwią jesienne liście na jaskrawoczerwony. Powstają w chłodne słoneczne dni, kiedy synteza chlorofilu zatrzymuje się w liściach. W liściach, gdy nie tworzą się antocyjany, po zniszczeniu chlorofilu zauważalne stają się żółto-pomarańczowe karotenoidy chloroplastów. Liście są najjaśniejsze w chłodnej, jasnej jesieni.
Wakuole biorą udział w niszczeniu makrocząsteczek, w krążeniu ich składników w komórce. Rybosomy, mitochondria, plastydy wpadające do wakuoli ulegają zniszczeniu. Dzięki tej czynności trawiennej można je porównać lizosomy- organelle komórek zwierzęcych.
Wakuole powstają z retikulum endoplazmatycznego (retikulum)
Rybosomy. Małe cząsteczki (17 - 23nm), składające się z mniej więcej równych ilości białka i RNA. W rybosomach aminokwasy łączą się, tworząc białka. W komórkach o aktywnym metabolizmie jest ich więcej. Rybosomy znajdują się w cytoplazmie komórki swobodnie lub przyczepione do retikulum endoplazmatycznego (80S). Występują również w jądrze (80S), mitochondriach (70S), plastydach (70S).
Rybosomy mogą tworzyć kompleks, na którym zachodzi jednoczesna synteza identycznych polipeptydów, o których informacja pochodzi z jednej cząsteczki i RNA. Taki kompleks nazywa się polirybosomy (polisomy). Komórki, które syntetyzują białka w dużych ilościach, mają rozbudowany system polisomów, które często są przyczepione do zewnętrznej powierzchni otoczki jądrowej.
Retikulum endoplazmatyczne. Jest to złożony trójwymiarowy system membranowy o nieokreślonym zasięgu. W kontekście ER wygląda jak dwie elementarne membrany z wąską przezroczystą przestrzenią między nimi. Kształt i długość ER zależy od rodzaju komórki, jej aktywności metabolicznej oraz stopnia zróżnicowania. W komórkach, które wydzielają lub przechowują białka, ER ma postać płaskich worków lub cysterny, z licznymi rybosomami przyczepionymi do jego zewnętrznej powierzchni. To siateczka nazywa się szorstka siateczka śródplazmatyczna. Gładki ER ma zwykle kształt cylindryczny. W tej samej komórce może znajdować się szorstka i gładka siateczka śródplazmatyczna. Z reguły istnieje między nimi wiele połączeń.
Retikulum endoplazmatyczne działa jako system komunikacyjny komórki. Jest związany z zewnętrzną powłoką jądra. W rzeczywistości te dwie struktury tworzą jeden system membranowy. Kiedy otoczka jądrowa pęka podczas podziału komórki, jej fragmenty przypominają fragmenty ER. Retikulum endoplazmatyczne to system transportu substancji: białek, lipidów, węglowodanów do różnych części komórki. retikulum endoplazmatyczne sąsiednich komórek połączone są nićmi cytoplazmatycznymi plazmodesmy - które przechodzą przez ściany komórkowe.
Retikulum endoplazmatyczne jest głównym miejscem syntezy błon komórkowych. W niektórych komórkach roślinnych błony wakuoli i mikrociał, cysterny dictyosomy.
Aparat Golgiego. Termin ten jest używany w odniesieniu do wszystkich dictyosom, Lub ciałko Golgiego, w klatce. Dictyosomy to grupy płaskich pęcherzyków w kształcie dysku lub cystern, które rozgałęziają się wzdłuż krawędzi w złożony system kanalików. Dictyosomy u roślin wyższych składają się z 4 - 8 cystern, zebranych razem.
Zwykle w paczce zbiorników wyróżnia się stronę rozwijającą się i dojrzewającą. błony tworzących się cystern przypominają strukturą błony ER, a błony dojrzewających cystern przypominają błonę komórkową.
Dictyosomy biorą udział w wydzielaniu, a u większości roślin wyższych w tworzeniu błon komórkowych. Polisacharydy ściany komórkowej syntetyzowane przez dictyosomy gromadzą się w pęcherzykach, które następnie oddzielają się od dojrzewających cystern. Te pęcherzyki wydzielnicze migrują i łączą się z CMP; natomiast zawarte w nich polisacharydy wbudowują się w błonę komórkową. Niektóre substancje gromadzące się w dictyosomach powstają w innych strukturach, na przykład w ER, a następnie są transportowane do dictyosomów, gdzie są modyfikowane (modyfikowane) przed sekrecją. Na przykład glikoproteiny są ważnym materiałem budulcowym błony komórkowej. Część białkowa jest syntetyzowana przez polisomy szorstkiego ER, część węglowodanowa jest syntetyzowana w dictyosomach, gdzie obie części łączą się tworząc glikoproteiny.
Membrany to dynamiczne, ruchome struktury, które nieustannie zmieniają swój kształt i powierzchnię. Koncepcja opiera się na ruchliwości błony układ endoplazmatyczny. Zgodnie z tą koncepcją błony wewnętrzne cytoplazmy, z wyjątkiem błon mitochondriów i plastydów, stanowią jedną całość i wywodzą się z retikulum endoplazmatycznego. Nowe cysterny dictyosomów tworzą się z retikulum endoplazmatycznego przez etap pęcherzyków pośrednich, a pęcherzyki wydzielnicze, które oddzielają się od dictyosomów, ostatecznie przyczyniają się do tworzenia błony plazmatycznej. Zatem retikulum endoplazmatyczne i dictyosomy tworzą jednostkę funkcjonalną, w której dictyosomy pełnią rolę struktur pośrednich w procesie przekształcania błon, takich jak retikulum endoplazmatyczne, w błony takie jak plazma. W tkankach, których komórki słabo rosną i dzielą się, składniki błony są stale aktualizowane.
mikrotubule występuje w prawie wszystkich komórkach eukariotycznych. Są to cylindryczne struktury o średnicy około 24 nm. Ich długość jest różna. Każdy kanalik składa się z podjednostek białka o nazwie tubulina. Podjednostki tworzą 13 podłużnych włókien otaczających centralną jamę. Mikrotubule są strukturami dynamicznymi, są regularnie niszczone i formowane na określonych etapach cyklu komórkowego. Ich gromadzenie odbywa się w specjalnych miejscach, które nazywane są centrami organizacji mikrotubul. W komórkach roślinnych mają słabo wyrażoną amorficzną strukturę.
Funkcje mikrotubul: uczestniczą w tworzeniu błony komórkowej; kierują pęcherzyki dictyosomów do tworzącej się błony, jak nici wrzeciona, które tworzą się w dzielącej się komórce; odgrywają rolę w tworzeniu płytki komórkowej (początkowa granica między komórkami potomnymi). Ponadto mikrotubule są ważnym składnikiem wici i rzęsek, w których ruchu odgrywają ważną rolę.
mikrofilamenty, podobnie jak mikrotubule, znajdują się w prawie wszystkich komórkach eukariotycznych. Są to długie włókna o grubości 5–7 nm, składające się z kurczliwego białka aktyny. Pęczki mikrofilamentów znajdują się w wielu komórkach roślin wyższych. Najwyraźniej odgrywają ważną rolę w prądach cytoplazmy. Mikrofilamenty wraz z mikrotubulami tworzą elastyczną sieć tzw cytoszkielet.
Substancja podstawowa przez długi czas był uważany za jednorodny (homogeniczny) bogaty w białko roztwór o niewielkiej liczbie struktur lub nawet bez struktury. Jednak obecnie, przy użyciu wysokonapięciowego mikroskopu elektronowego, ustalono, że substancja podstawowa jest trójwymiarową siatką zbudowaną z cienkich (o średnicy 3–6 nm) pasm wypełniających całą komórkę. Z tego zawieszone są inne składniki cytoplazmy, w tym mikrotubule i mikrofilamenty siatka mikrotrabekularna.
Struktura mikrotrabekularna to sieć pasm białkowych, między którymi przestrzeń jest wypełniona wodą. Wraz z wodą siatka ma konsystencję żelu, który wygląda jak galaretowate ciała.
Organelle są przyczepione do siatki mikrotrabekularnej. Sieć zapewnia komunikację pomiędzy poszczególnymi częściami komórki oraz kieruje transportem wewnątrzkomórkowym.
Krople lipidowe- struktury o kulistym kształcie, nadające ziarnistość cytoplazmie komórki roślinnej pod mikroskopem świetlnym. Na mikrografach elektronowych wyglądają na amorficzne. Bardzo podobne, ale mniejsze kropelki znajdują się w plastydach.
Kropelki lipidów, myląc je z organellami, nazywały je sferosomami i uważały, że otoczone są jednowarstwową lub dwuwarstwową błoną. Jednak ostatnie dane pokazują, że kropelki lipidów nie mają błon, ale mogą być pokryte białkiem.
Substancje ergastyczne - są to „produkty pasywne” protoplastu: substancje rezerwowe lub produkty odpadowe. Mogą pojawiać się i znikać w różnych okresach cyklu komórkowego. Z wyjątkiem ziaren skrobi, kryształów, barwników antocyjanowych i kropelek lipidów. Należą do nich żywice, gumy, garbniki i białka. Substancje ergastyczne są częścią błony komórkowej, głównej substancji cytoplazmy i organelli, w tym wakuoli.
Wici i rzęski - są to cienkie, przypominające włosy struktury, które wystają z powierzchni wielu komórek eukariotycznych. Mają stałą średnicę, ale długość waha się od 2 do 150 mikronów. Konwencjonalnie dłuższe i nieliczne z nich nazywane są wiciami, a krótsze i liczniejsze nazywane są rzęskami. Nie ma wyraźnych różnic między tymi dwoma typami struktur, więc termin ten jest używany w odniesieniu do obu. rozłóg.
U niektórych alg i grzybów wici są narządami ruchu, za pomocą których poruszają się w wodzie. W roślinach (np. mchach, wątrobowcach, paprociach, niektórych nagonasiennych) tylko komórki rozrodcze (gamety) mają wici.
Każda wici ma określoną organizację. Zewnętrzny pierścień 9 par mikrotubul otacza dwie dodatkowe mikrotubule znajdujące się w środku wici. Zawierające enzym „uchwyty” rozciągają się od jednej mikrotubuli każdej z zewnętrznych par. Jest to podstawowy wzorzec organizacji 9+2 występujący we wszystkich wici organizmów eukariotycznych. Uważa się, że ruch wici opiera się na przesuwaniu się mikrotubul, podczas gdy zewnętrzne pary mikrotubul poruszają się jedna po drugiej bez skurczu. Przesuwanie się par mikrotubul względem siebie powoduje miejscowe wyginanie wici.
Wici „wyrastają” z cytoplazmy cylindrycznych struktur tzw ciała podstawowe, uformowana i podstawna część wici. Ciała podstawowe mają wewnętrzną strukturę przypominającą wici, z wyjątkiem tego, że kanaliki zewnętrzne są ułożone raczej w trojaczki niż w pary, a kanalików środkowych nie ma.
Ściana komórkowa.Ściana komórkowa ogranicza rozmiar protoplastu i zapobiega jego pęknięciu w wyniku absorpcji wody przez wakuolę.
Ściana komórkowa pełni określone funkcje, które są ważne nie tylko dla komórki i tkanki, w której komórka się znajduje, ale także dla całej rośliny. Ściany komórkowe odgrywają zasadniczą rolę we wchłanianiu, transporcie i wydalaniu substancji, a ponadto może się w nich koncentrować aktywność lizosomalna, czyli trawienna.
Składniki ściany komórkowej. Najbardziej typowym składnikiem ściany komórkowej jest celuloza, co w dużej mierze determinuje jego architekturę. Cząsteczki celulozy składają się z powtarzających się cząsteczek glukozy połączonych końcami. Długie, cienkie cząsteczki celulozy są łączone w mikrofibryle grubość 10 - 25 nm. Mikrofibryle splatają się i tworzą cienkie nitki, które z kolei można owijać jedna wokół drugiej, jak nitki liny. Każda taka „sznurówka”, czyli makrofibryla, ma grubość około 0,5 mikrona, osiągając długość 4 mikronów. Makrofibryle są tak mocne, jak drut stalowy tej samej wielkości.
Ramka celulozowa ściany komórkowej jest wypełniona splecionymi z nią cząsteczkami matrycy celulozowej. Zawiera polisacharydy tzw hemicelulozy, I pektyny, Lub pektyny, chemicznie bardzo zbliżone do hemiceluloz.
Kolejnym składnikiem ściany komórkowej jest lignina - jest najobficiej występującym polimerem w komórkach roślinnych po celulozie. Lignina zwiększa sztywność ściany i zwykle występuje w komórkach pełniących funkcję podtrzymującą lub mechaniczną.
Kutyna, suberyna, wosk- osadzają się zwykle w łuskach tkanek ochronnych roślin. Na przykład kutyna znajduje się w błonach komórkowych naskórka, a suberyna w wtórnej tkance ochronnej, korku. Obie substancje występują w połączeniu z woskami i zapobiegają nadmiernej utracie wody z rośliny.
Warstwy ściany komórkowej. Grubość ścianek komórek roślinnych jest bardzo zróżnicowana, w zależności od roli komórek w strukturze rośliny i wieku samej komórki. Pod mikroskopem elektronowym w ścianie komórkowej rośliny widoczne są dwie warstwy: płyta środkowa(zwana także substancją międzykomórkową) i pierwotna ściana komórkowa. Wiele komórek kładzie kolejną warstwę - wtórna ściana komórkowa. Płytka środkowa znajduje się między głównymi ścianami sąsiednich komórek. Ściana wtórna, jeśli występuje, jest osadzana przez protoplast komórki na wewnętrznej powierzchni pierwotnej ściany komórkowej.
Płyta środkowa. Blaszka środkowa składa się głównie z pektyn. Tam, gdzie powinna powstać ściana komórkowa, między dwiema nowo utworzonymi komórkami, najpierw zauważa się gęsty splot kanalików retikulum endoplazmatycznego i cysterny aparatu Golgiego (dictyosomy). Wtedy w tym miejscu pojawiają się bąbelki wypełnione pektyną (z polisacharydów). Pęcherzyki te są oddzielane od zbiorników aparatu Golgiego. Wczesna ściana komórkowa zawiera różne polisacharydy, z których głównymi są pektyny i hemiceluloza. Później składa się z gęstszych substancji - celulozy i ligniny.
Pierwotna ściana komórkowa. Jest to warstwa otoczki celulozowej, która jest osadzana przed lub w trakcie wzrostu komórek. Oprócz celulozy, hemicelulozy i pektyny błony pierwotne zawierają glikoproteinę. Muszle pierwotne mogą ulegać zdrewnieniu. Składnik pektynowy nadaje plastyczność, która umożliwia rozszerzanie się pierwotnej skorupy w miarę wydłużania się korzenia, łodygi lub liścia.
Komórki aktywnie dzielące się (większość dojrzałych komórek biorących udział w procesach fotosyntezy, oddychania i wydzielania) mają błony pierwotne. Takie komórki z pierwotną błoną i żywym protoplastem są w stanie utracić swój charakterystyczny kształt, dzielić się i różnicować w nowy typ komórek. U roślin biorą udział w gojeniu się ran i regeneracji tkanek.
Pierwotne błony komórkowe nie mają jednolitej grubości na całej swojej długości, ale mają cienkie odcinki tzw pierwotne pola porów. Nici cytoplazmy lub plazmodesmy, które łączą protoplasty sąsiednich komórek, zwykle przechodzą przez pierwotne pola porów.
wtórna ściana komórkowa. Pomimo faktu, że wiele komórek roślinnych ma tylko ścianę pierwotną, w niektórych protoplast odkłada ścianę wtórną w kierunku środka komórki. Zwykle dzieje się tak po ustaniu wzrostu komórek, a powierzchnia błony pierwotnej już się nie zwiększa. Z tego powodu wtórna powłoka różni się od podstawowej. Membrany wtórne są szczególnie potrzebne wyspecjalizowanym komórkom, które wzmacniają roślinę i przewodzą wodę. Po odłożeniu błony wtórnej protoplast tych komórek z reguły umiera. W skorupkach wtórnych jest więcej celulozy niż w skorupkach pierwotnych, a nie ma w nich substancji pektynowych i glikoprotein. Błona wtórna jest trudna do rozciągnięcia, jej macierz składa się z hemicelulozy.
W powłoce wtórnej można wyróżnić trzy warstwy - zewnętrzną, środkową i wewnętrzną (S 1, S 2, S 3). Warstwowa struktura skorup wtórnych znacznie zwiększa ich wytrzymałość. Mikrofibryle celulozy w błonie wtórnej są osadzane gęstiej niż w błonie pierwotnej. Lignina jest powszechnym składnikiem skór wtórnych drewna.
Pory w błonach stykających się komórek znajdują się naprzeciw siebie. Tworzą się dwa przeciwstawne pory i membrana porów kilka razy. W komórkach z błonami wtórnymi występują dwa główne typy porów: proste i obramowane. W graniczących porach membrana wtórna zwisa jama porów. W prostych porach tak nie jest.
Wzrost ściany komórkowej. W miarę wzrostu komórki zwiększa się grubość i powierzchnia błony komórkowej. Rozciąganie skorupy to złożony proces. Jest kontrolowany przez protoplast i regulowany przez hormon hormon.
W komórkach rosnących równomiernie we wszystkich kierunkach odkładanie miofibryli jest przypadkowe. Te miofibryle tworzą nieregularną sieć. Takie komórki znajdują się w rdzeniu łodygi, tkankach spichrzowych oraz podczas hodowli komórek in vitro. W wydłużających się komórkach miofibryle osłonek bocznych osadzają się pod kątem prostym do osi wydłużenia.
Substancje macierzy - pektyny, hemicelulozy i glikoproteiny są przenoszone na błonę w pęcherzykach dictyosomów. Jednocześnie pektyny są bardziej charakterystyczne dla komórek rosnących, podczas gdy hemicelulozy przeważają w komórkach nie rosnących.
Mikrofibryle celulozowe są syntetyzowane na powierzchni komórki za pomocą kompleksu enzymatycznego związanego z błoną plazmatyczną. Orientacja mikrofibryli jest kontrolowana przez mikrotubule znajdujące się w pobliżu wewnętrznej powierzchni błony plazmatycznej.
Plazmodesma. Są to cienkie nici cytoplazmy, które łączą protoplasty sąsiednich komórek. Plazmodesmy albo przechodzą przez ścianę komórkową w dowolnym miejscu, albo są skoncentrowane w pierwotnych polach porów lub w błonach między parami porów. Pod mikroskopem elektronowym plazmodesmy wyglądają jak wąskie kanały wyłożone błoną plazmatyczną. Mniejsza cylindryczna rurka rozciąga się wzdłuż osi kanału od jednej komórki do drugiej - rura desmotyczna, który komunikuje się z retikulum endoplazmatycznym obu sąsiadujących komórek. Wiele plazmodesmy tworzy się podczas podziału komórki, kiedy rurkowa retikulum endoplazmatyczne jest pobierane przez rozwijającą się płytkę komórkową. Plazmodesmy mogą również tworzyć się w błonach komórek, które się nie dzielą. Struktury te zapewniają sprawne przenoszenie określonych substancji z komórki do komórki.
Podział komórek. W organizmach wielokomórkowych podział komórek wraz ze wzrostem ich wielkości jest sposobem wzrostu całego organizmu. Nowe komórki powstałe podczas podziału są podobne pod względem struktury i funkcji, zarówno do komórki macierzystej, jak i do siebie nawzajem. Proces podziału u eukariontów można podzielić na dwa częściowo nakładające się etapy: mitoza I cytokineza.
Mitoza- jest to tworzenie dwóch jąder potomnych z jednego jądra, morfologicznie i genetycznie równoważnych sobie nawzajem. Cytokineza - Jest to podział cytoplazmatycznej części komórki z utworzeniem komórek potomnych.
Cykl komórkowy.Żywa komórka przechodzi przez serię następujących po sobie zdarzeń, które składają się na cykl komórkowy. Czas trwania samego cyklu różni się w zależności od rodzaju komórki oraz czynników zewnętrznych, takich jak temperatura czy dostępność składników odżywczych. Zwykle cykl dzieli się na interfaza i cztery fazy mitoza.
Interfaza. Okres między kolejnymi podziałami mitotycznymi.
Interfaza jest podzielona na trzy okresy, oznaczone jako G 1 , S , G 2 .
W okresie G 1, który rozpoczyna się po mitozie. W tym okresie wzrasta ilość cytoplazmy, w tym różnych organelli. Ponadto, zgodnie ze współczesną hipotezą, w okresie G 1 syntetyzowane są substancje, które stymulują lub hamują okres S i resztę cyklu, determinując w ten sposób proces podziału.
Okres S następuje po okresie G1, w którym następuje powielenie materiału genetycznego (DNA).
W okresie G2, który następuje po S, powstają struktury bezpośrednio zaangażowane w mitozę, na przykład elementy wrzeciona.
Niektóre komórki przechodzą nieograniczoną liczbę cykli komórkowych. Są to organizmy jednokomórkowe oraz niektóre komórki stref aktywnego wzrostu (merystemy). Niektóre wyspecjalizowane komórki po dojrzewaniu tracą zdolność do reprodukcji. Trzecia grupa komórek, na przykład te tworzące tkankę rany (kalus), zachowuje zdolność do podziału tylko w szczególnych warunkach.
Mitoza, lub rozszczepienie jądrowe. Jest to ciągły proces podzielony na cztery fazy: profaza, metafaza, anafaza, telofaza. W wyniku mitozy materiał genetyczny, który podwoił się w interfazie, jest równo dzielony między dwa jądra potomne.
Jedną z najwcześniejszych oznak przejścia komórki do podziału jest pojawienie się wąskiego, pierścieniowego pasma mikrotubul tuż pod błoną plazmatyczną. Ten stosunkowo gęsty pas otacza jądro w płaszczyźnie równikowej przyszłego wrzeciona mitotycznego. Ponieważ pojawia się przed profazą, nazywa się pas preprofazowy. Zanika po wrzecionie mitotycznym, na długo przed pojawieniem się w późnej telofazie płytki komórkowej, która rozrasta się od środka ku obrzeżom i łączy się z błoną komórki macierzystej w obszarze zajmowanym wcześniej przez pas preprofazowy.
profaza. Na początku profazy chromosomy przypominają długie nitki rozproszone wewnątrz jądra. Następnie, gdy nici skracają się i pogrubiają, można zauważyć, że każdy chromosom składa się nie z jednej, ale z dwóch splecionych nici, zwanych chromatydy. W późnej profazie dwie skrócone sparowane chromatydy każdego chromosomu leżą obok siebie równolegle, połączone wąskim odcinkiem zwanym centromer. Ma określoną pozycję na każdym chromosomie i dzieli chromosom na dwa ramiona o różnej długości.
Mikrotubule znajdują się równolegle do powierzchni jądra wzdłuż osi wrzeciona. Jest to najwcześniejsza manifestacja zespołu wrzeciona mitotycznego.
Pod koniec profazy jąderko stopniowo traci swój wyraźny zarys i ostatecznie zanika. Wkrótce potem otoczka jądrowa również się rozpada.
Metafaza. Najpierw wrzeciono metafazowe, który reprezentuje trójwymiarową strukturę, najszerszą w środku i zwężającą się ku biegunom, zajmuje miejsce poprzednio zajmowane przez jądro. Włókna wrzeciona są wiązkami mikrotubul. Podczas metafazy chromosomy, z których każdy składa się z dwóch chromatyd, są ułożone w taki sposób, że ich centromery leżą w płaszczyźnie równikowej wrzeciona. Dzięki centromerowi każdy chromosom jest przyczepiony do nici wrzeciona. Jednak niektóre nici przechodzą z jednego bieguna na drugi bez przyczepiania się do chromosomów.
Kiedy wszystkie chromosomy znajdują się w płaszczyźnie równikowej, metafaza jest zakończona. Chromosomy są gotowe do podziału.
Anafaza. Chromatydy każdego chromosomu rozdzielają się. teraz to dziecko chromosomy. Po pierwsze, centromer dzieli się, a dwa chromosomy potomne są przyciągane do przeciwnych biegunów. W tym przypadku centromery przesuwają się do przodu, a ramiona chromosomów rozciągają się do tyłu. Włókna wrzeciona przyczepione do chromosomów skracają się, przyczyniając się do rozdzielenia chromatyd i ruchu chromosomów potomnych w przeciwnych kierunkach.
telofaza. W telofazie następuje zakończenie rozdzielenia dwóch identycznych grup chromosomów, podczas gdy wokół każdej z nich tworzy się błona jądrowa. Siateczka szorstka bierze w tym czynny udział. Aparat wrzeciona znika. Podczas telofazy chromosomy tracą wyrazistość konturów, rozciągają się, zamieniając z powrotem w cienkie nitki. Jąderka są przywrócone. Kiedy chromosomy stają się niewidoczne, mitoza jest zakończona. Dwa jądra potomne wchodzą w interfazę. Są genetycznie równoważne sobie nawzajem i macierzystemu jądru. Jest to bardzo ważne, ponieważ program genetyczny, a wraz z nim wszystkie cechy, muszą zostać przeniesione na organizmy potomne.
Czas trwania mitozy jest różny u różnych organizmów i zależy od rodzaju tkanki. Jednak profaza jest najdłuższa, a anafaza najkrótsza. W komórkach wierzchołka korzenia czas trwania profazy wynosi 1–2 godziny; metafaza - 5 - 15 min; anafaza - 2 - 10 minut; telofaza - 10 - 30 min. Czas trwania interfazy wynosi od 12 do 30 godzin.
W wielu komórkach eukariotycznych centra organizujące mikrotubule odpowiedzialne za tworzenie wrzeciona mitotycznego są związane z centriole.
Cytokineza. Jest to proces podziału cytoplazmy. U większości organizmów komórki dzielą się poprzez cofnięcie błony komórkowej i utworzenie bruzdy rozszczepialnej, która stopniowo się pogłębia, ściskając pozostałe włókna wrzeciona mitotycznego.U wszystkich roślin (mszaków i naczyniowych) oraz u niektórych glonów komórki dzielą się w wyniku powstania płytka komórkowa.
We wczesnej telofazie między dwoma jądrami potomnymi tworzy się beczkowaty układ włókien, tzw fragmoplast. Włókna phragmoplastu, podobnie jak włókna wrzeciona mitotycznego, składają się z mikrotubul. Małe krople pojawiają się w płaszczyźnie równikowej fragmoplastu. Łączą się, tworząc płytkę komórkową, która rośnie, aż dotrze do ściany dzielącej się komórki. To kończy rozdzielanie dwóch komórek potomnych. Łączące się kropelki to bąbelki odrywające się od aparatu Golgiego. Zasadniczo zawierają substancje pektynowe, z których powstaje środkowa płytka. Błony pęcherzyków biorą udział w budowie błony plazmatycznej po obu stronach płytki. W tym samym czasie plazmodesmy powstają z fragmentów rurkowatej retikulum endoplazmatycznego.
Po utworzeniu płytki środkowej każdy protoplast osadza na niej pierwotną błonę. Ponadto każda komórka potomna osadza nową warstwę błony wokół całego protoplastu, która kontynuuje błonę, która powstała z płytki komórkowej. Oryginalna powłoka komórki macierzystej jest niszczona w miarę wzrostu komórek potomnych.
Różne typy mitozy eukariotycznej
Opisany powyżej podział komórkowy u roślin, a także u zwierząt, nie jest jedyną formą pośredniego podziału komórkowego. Najprostszym rodzajem mitozy jest pleuromitoza. Przypomina podział binarny komórki prokariotyczne, w którym nukleoidy po replikacji pozostają związane z błoną plazmatyczną. Błona zaczyna rosnąć między punktami wiązania DNA, a tym samym rozprzestrzenia chromosomy do różnych części komórki. Następnie, podczas tworzenia zwężenia komórki, każda z cząsteczek DNA znajdzie się w nowej oddzielnej komórce.
Charakterystyczne dla podziału komórki eukariotyczne jest tworzenie się wrzeciona zbudowanego z mikrotubul. Na zamknięta pleuromitoza(nazywa się to zamkniętym, ponieważ dywergencja chromosomów zachodzi bez naruszenia otoczki jądrowej) nie centriole, ale inne struktury znajdujące się po wewnętrznej stronie błony jądrowej uczestniczą jako centra organizacji mikrotubul (MCMT). Są to tak zwane ciała polarne o nieokreślonej morfologii, z których wychodzą mikrotubule. Są dwa takie ciała. Oddalają się od siebie nie tracąc kontaktu z otoczką jądrową. W wyniku tego powstają dwa półwrzeciona związane z chromosomami. Cały proces tworzenia wrzeciona mitotycznego i segregacji chromosomów zachodzi w tym przypadku pod otoczką jądrową. Ten typ mitozy występuje wśród pierwotniaków i jest szeroko rozpowszechniony u grzybów (chytridia, zygomycetes, drożdże, lęgniowce, ascomycetes, myxomycetes itp.). spotkać formy półzamknięta pleuromitoza gdy otoczka jądrowa zostaje zniszczona na biegunach uformowanego wrzeciona.
Następną formą mitozy jest ortomitoza. W tym przypadku COMT znajdują się w cytoplazmie, od samego początku nie tworzą się półwrzeciona, ale wrzeciono dwubiegunowe. Istnieją trzy formy ortomitoza ( normalna mitoza) półzamknięte I Zamknięte. W półzamkniętej ortomitozie powstaje bisymetryczne wrzeciono za pomocą TsOMT znajdującego się w cytoplazmie, otoczka jądrowa jest zachowana przez całą mitozę, z wyjątkiem stref polarnych. Masy materiału ziarnistego, a nawet centriole, mogą być wykrywane jako COMT. Ta forma mitozy występuje w zoosporach zielonych, brunatnych i czerwonych alg, w niektórych niższych grzybach i gregarynach. Przy zamkniętej ortomitozie błona jądrowa jest całkowicie zachowana, w której powstaje prawdziwe wrzeciono. Mikrotubule powstają w karioplazmie, rzadziej wyrastają z wewnętrznego TsOMT, który nie jest związany (w przeciwieństwie do pleuromitozy) z błoną jądrową. Ten typ mitozy jest typowy dla podziału mikrojąder orzęsków, ale może również wystąpić u pierwotniaków. W otwartej ortomitozie otoczka jądrowa całkowicie się rozpada. Ten typ podziału komórek jest charakterystyczny dla organizmów zwierzęcych, niektórych pierwotniaków i komórek roślin wyższych. Z kolei ta forma mitozy jest reprezentowana przez typy astralne i anastralne.
Z pokrótce przejrzanego materiału można zauważyć, że główną cechą mitozy w ogóle jest pojawianie się struktur wrzeciona rozszczepienia, które powstają w połączeniu z CMT, które różnią się budową.
Morfologia figury mitotycznej
Aparat mitotyczny jest szczególnie dobrze wyrażany na metafazowym etapie mitozy. W metafazie, w płaszczyźnie równikowej komórki, zlokalizowane są chromosomy, z których tzw gwinty wrzecion, zbiegające się na dwóch różnych biegunach figury mitotycznej. Zatem wrzeciono mitotyczne jest zbiorem chromosomów, biegunów i włókien. Włókna wrzeciona to pojedyncze mikrotubule lub ich wiązki. Mikrotubule zaczynają się od biegunów wrzeciona, a niektóre z nich odchodzą do centromerów gdzie się znajdują kinetochory chromosomy (mikrotubule kinetochoru), niektóre przechodzą dalej w kierunku przeciwnego bieguna, ale go nie docierają. Nazywa się je „mikrotubulami międzybiegunowymi”. Od biegunów odchodzi grupa promienistych mikrotubul, tworząc wokół nich strukturę przypominającą „promienną poświatę” – są to mikrotubule astralne.
Pod względem morfologii figury mitotyczne dzielą się na typy astralne i anastralne.
Typ wrzeciona astralnego lub zbieżny charakteryzuje się tym, że jego bieguny są reprezentowane przez małą strefę, do której zbiegają się (zbiegają) mikrotubule. Zwykle centrosomy zawierające centriole znajdują się na biegunach wrzecion astralnych. Znane są jednak przypadki centralnych mitoz astralnych (podczas mejozy niektórych bezkręgowców). Ponadto odnotowuje się promieniste mikrotubule promieniujące z biegunów, które nie są częścią wrzeciona, ale tworzą strefy gwiaździste - cytastry. Ten typ podziału mitotycznego przypomina hantle. Anastrialny typ figury mitotycznej nie ma cytastrów na biegunach. Regiony polarne wrzeciona są tutaj szerokie, nazywane są czapami polarnymi, zawierają centriole. W tym przypadku włókna wrzeciona nie odbiegają od jednego punktu, ale rozchodzą się szerokim frontem (rozchodzą się) z całej strefy czap polarnych. Ten typ wrzeciona jest charakterystyczny dla dzielących się komórek roślin wyższych, ale można go również znaleźć u zwierząt wyższych. We wczesnej embriogenezie ssaków mitozy centriolarne (rozbieżne) obserwuje się podczas podziału dojrzewania oocytu oraz w I i II podziale zygoty. Ale już w trzecim podziale komórkowym i we wszystkich kolejnych komórkach dzielą się z udziałem wrzecion astralnych, w biegunach których zawsze znajdują się centriole. Ogólnie rzecz biorąc, dla wszystkich form mitozy chromosomy z ich kinetoforami, ciałami polarnymi (centrosomami) i włóknami wrzeciona pozostają wspólnymi strukturami.
Centromery i kinetochory
Centromery mogą mieć różną lokalizację wzdłuż długości chromosomów. holocentryczny centromery występują, gdy mikrotubule są połączone wzdłuż całego chromosomu (niektóre owady, nicienie, niektóre rośliny). Monocentryczny centromery - gdy mikrotubule są związane z chromosomami w jednym obszarze. mogą być centromery monocentryczne sprecyzować(na przykład u niektórych pączkujących drożdży), gdy tylko jedna mikrotubula zbliża się do kinetochoru i strefowy, gdzie wiązka mikrotubul zbliża się do złożonego kinetochoru. Pomimo różnorodności stref centromerowych, wszystkie są związane ze złożoną strukturą. kinetochor, który ma fundamentalne podobieństwo w strukturze i funkcji u wszystkich eukariontów. Kinetochory to specjalne struktury białkowe, zlokalizowane głównie w strefach centromerowych chromosomów. Są to złożone kompleksy składające się z wielu białek. morfologicznie są bardzo podobne, mają taką samą budowę, od okrzemek po ludzi. Są to struktury trójwarstwowe: wewnętrzna gęsta warstwa przylegająca do ciała chromosomu, środkowa luźna warstwa i zewnętrzna gęsta warstwa. Z warstwy zewnętrznej rozciąga się wiele włókienek, tworząc tzw. włóknistą koronę kinetochoru. W ogólnej postaci kinetochory wyglądają jak płytki lub dyski leżące w strefie pierwotnego zwężenia chromosomu, w centromerze. Zwykle na chromosom lub chromatydę przypada jeden kinetochor. Przed anafazą na każdej siostrzanej chromatydzie znajdują się kinetochory, z których każda komunikuje się z własną wiązką mikrotubul. W roślinach kinetochor nie wygląda jak talerze, ale półkule. Funkcjonalną rolą kinetochorów jest wiązanie ze sobą siostrzanych chromatyd, wiązanie mikrotubul mitotycznych, regulacja separacji chromosomów i faktyczne przemieszczanie chromosomów podczas mitozy z udziałem mikrotubul. Ogólnie rzecz biorąc, struktury białkowe, kinetochory, podwajają się w okresie S, równolegle do podwajania chromosomów. Ale ich białka są obecne na chromosomach we wszystkich okresach cyklu komórkowego.
TKANKA ROŚLINNA
Tkanki edukacyjne (merystemy)
Tkanki edukacyjne w ciele roślin znajdują się w różnych miejscach, dlatego dzieli się je na następujące grupy (ryc. 0;1).
Merystemy wierzchołkowe (wierzchołkowe). znajduje się na wierzchołkach lub wierzchołkach narządów osiowych - łodygi, korzenia. Za pomocą tych merystemów organy wegetatywne roślin przeprowadzają wzrost długości.
Merystemy boczne charakterystyczne dla narządów osiowych. Tam są umieszczone koncentrycznie, w formie sprzęgła.
Przestępny, Lub interkalarne, merystemy wywodzą się z merystemów wierzchołkowych. Są to grupy komórek, które nie są jeszcze zdolne do namnażania się, ale wkroczyły na ścieżkę różnicowania. Nie ma wśród nich komórek początkowych, ale jest wiele wyspecjalizowanych.
merystemy rany zapewnić odbudowę uszkodzonej części ciała. Regeneracja zaczyna się od odróżnicowania, czyli odwrotnego rozwoju komórek wyspecjalizowanych do komórek merystematycznych. Zamieniają się w fellogen, który tworzy korek pokrywając powierzchnię rany. Komórki odróżnicowane, dzieląc się, mogą tworzyć luźną tkankę miąższową - kostnina. Z niego, w określonych warunkach, powstają narządy roślinne.
Tkanki powłokowe
Pełnią rolę bariery granicznej, oddzielającej leżące poniżej tkanki od środowiska. Powłoki pierwotne rośliny składają się wyłącznie z żywych komórek. Powłoki drugorzędowe i trzeciorzędowe - głównie z martwych z grubymi ścianami komórkowymi.
Główne funkcje tkanek powłokowych:
ochrona rośliny przed wysychaniem;
ochrona przed wnikaniem szkodliwych mikroorganizmów;
ochrona przed oparzeniami słonecznymi;
ochrona przed uszkodzeniami mechanicznymi;
regulacja metabolizmu między rośliną a środowiskiem;
wrażenie irytacji.
Pierwotna tkanka powłokowa - naskórek, naskórek . Składa się z żywych komórek. Utworzony z merystemów wierzchołkowych. Obejmuje młode rosnące łodygi i liście.
Naskórek formował się u roślin w związku z wyjściem ze środowiska wodnego na ląd w celu zapobieżenia jego wysychaniu. Oprócz aparatów szparkowych wszystkie komórki naskórka są ze sobą ściśle połączone. Zewnętrzne ściany głównych komórek są grubsze niż pozostałe. Cała powierzchnia pokryta jest warstwą kutyny i wosków roślinnych. Ta warstwa nazywa się naskórek(skóra). Nie występuje na rosnących korzeniach i podwodnych częściach roślin. Po wyschnięciu przepuszczalność łuski jest znacznie osłabiona.
Z wyjątkiem podstawowe komórki, w szczególności w naskórku są inne owłosienie, Lub trichomy. Są jednokomórkowe i wielokomórkowe (Rys. 2). Funkcjonalnie zwiększają powierzchnię naskórka, np. w strefie wzrostu korzenia, służą jako ochrona mechaniczna, przylegają do podpory i ograniczają utratę wody. Wiele roślin ma włosy gruczołowe np. pokrzywa.
Tylko w wyższych roślinach występują w epidermie szparki które regulują wymianę wody i gazów. Jeśli nie ma naskórka, nie ma potrzeby stosowania aparatów szparkowych. Aparaty szparkowe to grupa komórek, które się tworzą aparat szparkowy, który składa się z dwóch komórki strażnicze i przylegające komórki naskórka komórki boczne. Różnią się od głównych komórek naskórka (ryc.3 ). Komórki ochronne różnią się od otaczających je kształtem i obecnością dużej liczby chloroplastów oraz nierównomiernie pogrubionych ścian. Te, które są naprzeciw siebie, są grubsze niż pozostałe (rys. 4) . Pomiędzy komórkami ochronnymi powstaje otwór szparkowy, który prowadzi do przestrzeń podszparkowa zwany jama podszparkowa. Komórki ochronne są wysoce fotosyntetyzujące. Zawierają dużą ilość skrobi rezerwowej oraz liczne mitochondria.
Liczba i rozmieszczenie aparatów szparkowych, rodzajów aparatów szparkowych różni się znacznie w różnych roślinach. U współczesnych mszaków nie ma aparatów szparkowych. Fotosynteza w nich jest przeprowadzana przez pokolenie gametofitów, a sporofity nie są zdolne do niezależnego istnienia.
Zwykle aparaty szparkowe znajdują się na spodniej stronie liścia. U roślin pływających na powierzchni wody - na górnej powierzchni. W liściach zbóż aparaty szparkowe są często równomiernie rozmieszczone po obu stronach. Takie liście są oświetlane w miarę równomiernie. Na 1 mm2 powierzchni może znajdować się od 100 do 700 aparatów szparkowych.
Wtórna tkanka powłokowa (peryderma). Tkanka ta zastępuje naskórek, gdy zielony kolor jednorocznych pędów zmienia się na brązowy. Jest wielowarstwowa i składa się z centralnej warstwy komórek kambium - felogen. Komórki phellogenu, dzieląc się, układają warstwę na zewnątrz faceci, i w środku - koroderma(Rys.5).
Fellema, Lub korek. Początkowo składa się z żywych, cienkościennych komórek. Z biegiem czasu ich ściany są impregnowane suberyną i woskami roślinnymi i obumierają. Zawartość komórki jest wypełniona powietrzem.
Funkcje Fellema:
zapobiega utracie wilgoci;
chroni roślinę przed uszkodzeniami mechanicznymi;
chroni przed patogenami;
zapewnia izolację termiczną, ponieważ komórki są wypełnione powietrzem.
Komórki felogenu, zlokalizowane w samym naskórku, leżącej pod nim warstwie podnaskórkowej, rzadziej w głębokich warstwach kory pierwotnej, stanowią podstawę generacyjną kory pierwotnej.
Warstwa korka nie jest trwała. Występują w nim przerwy, które komunikują się z znajdującymi się w pobliżu przestrzeniami międzykomórkowymi. W tym samym czasie na powierzchni tworzą się małe guzki - soczewica, które komunikują przestrzenie międzykomórkowe z powietrzem atmosferycznym (ryc.6,7).
Jesienią fallogen pod przetchlinkami układa warstwę zakorkowanych komórek, które znacznie ograniczają transpirację, ale jej całkowicie nie wykluczają. Wiosną warstwa ta zapada się od wewnątrz. Na jasnej korze brzozy soczewica jest wyraźnie widoczna w postaci ciemnych kresek.
Trzeciorzędowa tkanka powłokowa (skorupa), jest również charakterystyczny tylko dla drzewiastych form roślin.
Phellogen jest wielokrotnie odkładany w głębszych warstwach kory. Tkanki znajdujące się na zewnątrz ostatecznie obumierają, tworząc skorupę. Jego komórki są martwe i niezdolne do rozciągania. Jednak żywe komórki położone głębiej dzielą się, co prowadzi do zwiększenia poprzecznego rozmiaru łodygi. Z biegiem czasu zewnętrzna warstwa skorupy pęka. Czas wystąpienia takiej przerwy jest wartością dość stałą dla poszczególnych roślin. Na jabłoni dzieje się to w siódmym roku życia, w grabie - w pięćdziesiątym. U niektórych gatunków nie występuje w ogóle. Główną funkcją peelingu jest ochrona przed uszkodzeniami mechanicznymi i termicznymi.
Miąższ
Jest to grupa wyspecjalizowanych tkanek, które wypełniają przestrzenie wewnątrz ciała rośliny pomiędzy tkankami przewodzącymi i mechanicznymi. (Rys. 8). Częściej komórki miąższu mają kształt zaokrąglony, rzadziej wydłużony. Charakterystyczna jest obecność rozwiniętych przestrzeni międzykomórkowych. Przestrzenie między komórkami razem tworzą system transportowy - apoplast Ponadto przestrzenie międzykomórkowe tworzą „system wentylacyjny” rośliny. Poprzez aparaty szparkowe lub przetchlinki łączą się z powietrzem atmosferycznym i zapewniają optymalny skład gazów wewnątrz rośliny. Rozwinięte przestrzenie międzykomórkowe są szczególnie potrzebne roślinom rosnącym na glebach podmokłych, gdzie normalna wymiana gazowa jest utrudniona. Ten miąższ nazywa się aerenchyma(Rys. 9).
Elementy miąższu, wypełniając szczeliny między innymi tkankami, pełnią również funkcję podporową. Komórki miąższu są żywe, nie mają grubych ścian komórkowych, jak sklerenchyma. Dlatego właściwości mechaniczne zapewnia turgor. Jeśli zawartość wody spada, co prowadzi do plazmolizy i więdnięcia rośliny.
Miąższ asymilacyjny tworzony przez cienkościenne komórki z wieloma przestrzeniami międzykomórkowymi. Komórki tej struktury zawierają wiele chloroplastów, tak to się nazywa chlorenchyma. Chloroplasty znajdują się wzdłuż ściany bez cieniowania się nawzajem. W miąższu asymilacyjnym zachodzą reakcje fotosyntezy, które dostarczają roślinie substancji organicznych i energii. Wynikiem procesów fotosyntezy jest możliwość istnienia wszystkich żywych organizmów na Ziemi.
Tkanki asymilacyjne występują tylko w oświetlonych częściach rośliny, oddzielone są od otoczenia przezroczystym naskórkiem. Jeśli naskórek zostanie zastąpiony nieprzezroczystymi wtórnymi tkankami powłokowymi, miąższ asymilacyjny zanika.
Miąższ spichrzowy służy jako rezerwuar substancji organicznych, które nie są czasowo wykorzystywane przez organizm roślinny. W zasadzie każda komórka z żywym protoplastem jest zdolna do odkładania materii organicznej w postaci różnego rodzaju inkluzji, ale niektóre komórki się w tym specjalizują. (Rysunek 10, 11). Związki bogate w energię odkładają się tylko w sezonie wegetacyjnym, zużywają w okresie spoczynku oraz w ramach przygotowań do kolejnego sezonu wegetacyjnego. Dlatego substancje rezerwowe osadzają się w organach wegetatywnych tylko u roślin wieloletnich.
Zbiornikiem wywarów mogą być zarówno narządy zwykłe (łodyga, korzeń), jak i specjalistyczne (kłącza, bulwy, cebulki). Wszystkie rośliny nasienne przechowują w nasionach cenne energetycznie substancje (liścienie, bielmo). Wiele roślin w suchym klimacie magazynuje nie tylko materię organiczną, ale także wodę. (Rys. 12). Na przykład aloes magazynuje wodę w mięsistych liściach, kaktusy w pędach.
tkaniny mechaniczne
Właściwości mechaniczne komórek roślinnych zapewniają:
twarda skorupa komórki;
turgor, czyli stan turgoru komórek.
Pomimo tego, że prawie wszystkie komórki tkanki mają właściwości mechaniczne, to jednak w roślinie występują tkanki, dla których właściwości mechaniczne są podstawowe. Ten kolenchyma I sklerenchyma. Zwykle działają w interakcji z innymi tkankami. Wewnątrz ciała rośliny tworzą rodzaj ramy. Dlatego są nazywane podporowy.
Nie wszystkie rośliny mają równie dobrze zdefiniowane tkanki mechaniczne. W znacznie mniejszym stopniu rośliny żyjące w środowisku wodnym potrzebują wewnętrznego wsparcia niż rośliny lądowe. Powodem jest to, że rośliny wodne potrzebują mniej wsparcia wewnętrznego. Ich ciało jest w dużej mierze podtrzymywane przez otaczającą wodę. Powietrze na lądzie nie tworzy takiego wsparcia, ponieważ ma mniejszą gęstość niż woda. Z tego powodu obecność wyspecjalizowanych tkanek mechanicznych staje się istotna.
Udoskonalenie wewnętrznych struktur wsparcia nastąpiło w procesie ewolucji.
Collenchyma. Tworzą go tylko żywe komórki, wydłużone wzdłuż osi narządu. Ten typ tkanki mechanicznej powstaje bardzo wcześnie, w okresie pierwotnego wzrostu. Dlatego ważne jest, aby komórki pozostały żywe i zachowały zdolność do rozciągania się wraz z rozciągającymi się komórkami, które są w pobliżu.
Cechy komórek collenchyma:
nierównomierne pogrubienie skorupy, w wyniku czego niektóre jej części pozostają cienkie, a inne pogrubiają;
muszle nie zdrewniały.
Komórki Collenchyma są rozmieszczone w różny sposób względem siebie. W sąsiednich komórkach na rogach skierowanych do siebie tworzą się zgrubienia ( Ryż. 13 ). Ta kollenchyma nazywa się narożnik. W innym przypadku komórki są ułożone w równoległe warstwy. Błony komórkowe zwrócone w stronę tych warstw są silnie pogrubione. Ten kollenchyma blaszkowata. Komórki mogą być ułożone luźno, z licznymi przestrzeniami międzykomórkowymi jest luźną kolenchymą. Kollenchyma taka często występuje w roślinach na glebach podmokłych.
Collenchyma ma szczególne znaczenie w młodych roślinach, formach zielnych, a także w częściach roślin, w których nie występuje wzrost wtórny, takich jak liście. W tym przypadku układa się go bardzo blisko powierzchni, czasem tuż pod naskórkiem. Jeśli narząd ma twarze, wzdłuż ich grzebieni znajdują się grube warstwy kolenchymy.
Komórki Collenchyma są funkcjonalne tylko w obecności turgoru. Niedobór wody zmniejsza skuteczność kolenchymy i roślina przejściowo więdnie, np. liście ogórka zwisają w upalny dzień. Po napełnieniu komórek wodą przywracane są funkcje kolenchymy.
sklerenchyma. Drugi rodzaj tkanek mechanicznych. W przeciwieństwie do kolenchymy, gdzie wszystkie komórki są żywe, komórki sklerenchymy są martwe. Ich ściany są bardzo grube. Pełnią funkcję mechaniczną. Silne pogrubienie błony prowadzi do naruszenia transportu substancji, w wyniku czego protoplast umiera ( Ryż. 14 ). Lignifikacja błon komórkowych sklerenchymy następuje, gdy organ rośliny zakończył już swój wzrost. Dzięki temu nie przeszkadzają już w rozciąganiu otaczających tkanek.
W zależności od kształtu wyróżnia się dwa rodzaje komórek sklerenchymy - włókna i sklereidy.
włókna mają silnie wydłużony kształt z bardzo grubymi ścianami i małą wnęką. Są nieco mniejsze niż włókna drzewne. Często pod naskórkiem tworzą podłużne warstwy i pasma. W łyku lub ksylemie można je znaleźć pojedynczo lub w grupach. W floemie nazywają się włókna łykowe(Ryż. 15 ) oraz w ksylemie włókna libriform(Ryż. 16 ).
sklereidy, Lub kamienne komórki, są reprezentowane przez zaokrąglone lub rozgałęzione komórki z potężnymi muszlami. W ciele rośliny można je znaleźć pojedynczo (komórki podporowe) ( rys.17 ) lub grupy (ryc. 18). Należy zauważyć, że właściwości mechaniczne silnie zależą od lokalizacji sklereidów. Część sklereidów tworzy ciągłe warstwy, jak na przykład w łupinach orzechów lub w pestkach owoców (owoce pestkowe).
tkanki wydalnicze
Każda żywa komórka początkowo ma funkcję usuwania substancji, podczas gdy transport może przebiegać zarówno wzdłuż gradientu stężeń, jak i pod gradient. Substancje, które zostały zsyntetyzowane w komórce, mogą zostać usunięte, a następnie wpłyną na aktywność innych komórek (fitohormony, enzymy). Taki proces nazywa się wydzielanie. W przypadku wydalania substancji będących produktami przemiany materii komórki proces ten nazywa się wydalanie. Pomimo faktu, że wydalanie substancji w komórkach roślinnych i zwierzęcych ma zasadnicze podobieństwo, rośliny mają szereg cech, które wynikają z fundamentalnych różnic w aktywności życiowej. Poziom reakcji metabolicznych u roślin jest znacznie niższy niż u zwierząt. W związku z tym emitowane są proporcjonalnie mniejsze ilości odpadów. Inną cechą jest to, że samodzielnie syntetyzując prawie wszystkie niezbędne związki organiczne, rośliny nigdy nie tworzą ich nadmiernych zapasów. Uwolnione substancje mogą służyć jako materiał wyjściowy do innych reakcji (na przykład CO 2 i H 2 O).
Jeśli u zwierząt proces wydalania toksyn wiąże się z ich usuwaniem z organizmu, rośliny mogą tego nie robić, izolując zbędne substancje w żywym protoplastie (usuwanie różnych substancji do wakuoli), w komórkach martwych (większość tkanek rośliny wieloletniej rośliny składają się z takich komórek), w przestrzeniach międzykomórkowych.
Emitowane substancje można podzielić na dwie duże grupy. Pierwsza grupa to substancje organiczne syntetyzowane bezpośrednio przez komórkę (enzymy, polisacharydy, ligniny, terpeny, te drugie są składnikami olejków eterycznych i żywic). Druga grupa - substancje, które początkowo dostają się do komórki z zewnątrz za pomocą transportu ksylemu lub łyka (woda, sole mineralne, aminokwasy, monosacharydy itp.). Wydalany sekret rzadko jest jednorodny i zwykle składa się z mieszaniny, w której jedna substancja ma najwyższe stężenie.
Rośliny, w przeciwieństwie do zwierząt, nie mają kompletnego systemu wydalniczego. Istnieją tylko wyspecjalizowane struktury rozsiane po całej roślinie - idioblasty. Tworzące je komórki są mniejsze niż leżące w pobliżu komórki miąższu. Mają gęstą elektronowo cytoplazmę z rozwiniętymi elementami retikulum endoplazmatycznego i kompleksu Golgiego. Najczęściej nie wyrażona, jest centralna wakuola. Między sobą a innymi żywymi komórkami komórki te są połączone licznymi plazmodesmami.
W zależności od lokalizacji mogą występować tkanki wydalnicze zewnętrzny i wewnętrzny.
Zewnętrzne tkanki wydalnicze.włosy gruczołowe są instrumentami pochodnymi naskórek. Morfologicznie są zmienne – mogą mieć wielokomórkową głowę, być wydłużone, w formie tarczy na nodze itp. (rys. 19). Należą do nich włosy pokrzywy.
Hydatody- są to konstrukcje uwalniające nadmiar wody w warunkach niskiej transpiracji i wysokiej wilgotności - patroszenie. Struktury te tworzą grupy bezbarwnych żywych komórek o cienkich ścianach - epitet. Tkanka ta sąsiaduje z wiązką przewodzącą (ryc. 20). Woda jest uwalniana przez specjalne aparaty szparkowe, które różnią się od zwykłych aparatów szparkowych bezruchem i stale otwartą szczeliną. Skład płynu gutacyjnego jest bardzo zróżnicowany, od prawie czystej wody po bardzo złożoną mieszaninę substancji.
Nektarniki. Wiele roślin charakteryzuje się wydzielaniem cieczy zawierającej od 7 do 87% mono- i oligosacharydów. Proces ten przeprowadzają specjalne struktury - nektarniki. W zależności od lokalizacji są kwiat, Lub kwiatowy, nektarniki (ryc. 21), a także rośliny znajdujące się na łodygach, liściach - ekstraflora, Lub nektarniki pozakwiatowe. Nektar może reprezentować Jest to niezmieniony sok z łyka, który jest dostarczany na powierzchnię przez przestrzenie międzykomórkowe i wydalany przez aparaty szparkowe. Bardziej złożone nektary tworzą miąższ gruczołowy, pokryty naskórkiem z włoskami gruczołowymi. Nektar jest wydalany przez komórki naskórka lub włosy gruczołowe. W tym przypadku wydzielany nektar różni się od soku z łyka. Dominuje bowiem glukoza, fruktoza, sacharoza, aw soku z łyka - glukoza. Jony występują w niskich stężeniach. Jony występują w niskich stężeniach. Aby przyciągnąć owady zapylające, nektar może zawierać hormony steroidowe, które są niezbędne owadom.
Gruczoły trawienne obecne w roślinach owadożernych (rosa, nepentis itp.). W odpowiedzi na dotyk małych bezkręgowców wydzielają wydzielinę zawierającą enzymy hydrolityczne, kwas mrówkowy, masłowy i propionowy. W efekcie biopolimery rozkładają się na monomery, które roślina wykorzystuje na swoje potrzeby.
Gruczoły solne rozwijają się u roślin rosnących na glebach zasolonych. Gruczoły te znajdują się na liściach i w postaci rozpuszczonej nadmiar jonów wydostaje się na powierzchnię. Sól najpierw osadza się na naskórku, a następnie jest zmywana przez deszcz (ryc. 22). Ponadto wiele roślin ma liście solne włosy. Każdy włos składa się z dwóch komórek: jedna tworzy głowę, druga - łodygę. Sole gromadzą się w wakuoli komórki głowy, a gdy ich stężenie staje się wysokie, głowa odpada. W jej miejsce rośnie nowa komórka, która również gromadzi sole. Podczas wzrostu liścia głowa może spaść i uformować się kilka razy. Zaletą solonych włosów jest to, że uwalniając sole, tracą mało wody. Zjawisko to jest bardzo ważne dla roślin rosnących na glebach zasolonych, gdzie brakuje słodkiej wody.
Wewnętrzne tkanki wydalnicze. Wewnętrzne struktury wydalnicze, rozproszone po całym ciele rośliny w postaci idioblastów, nie usuwają substancji poza organizm, gromadząc je w sobie. Idioblasty to komórki roślinne różniące się kształtem, strukturą lub zawartością od innych komórek w tej samej tkance. W zależności od pochodzenia rozróżnia się schizogenne (gr. schiso - rozszczepienie) i lizogenne (gr. liza - rozpuszczenie) zbiorniki wydzielanych substancji (ryc. 23).
Naczynia schizogeniczne. Są to mniej lub bardziej rozległe przestrzenie międzykomórkowe wypełnione wydalanymi substancjami. Na przykład przejścia żywiczne z iglaków, aralii, parasoli i innych. Uważa się, że żywica ma właściwości bakteriobójcze, odstrasza roślinożerców, czyniąc rośliny niejadalnymi dla nich.
Pojemniki lizogeniczne powstają w miejscu żywych komórek, które obumierają i ulegają zniszczeniu po nagromadzeniu w nich substancji. Takie formacje można zaobserwować w skórce owoców cytrusowych.
Oprócz głównych typów istnieją formy pośrednie. mleczny- są dwa rodzaje: łączone I niesegmentowane. Te pierwsze powstają z wielu żywych komórek ułożonych w łańcuchy. W miejscu kontaktu muszle ulegają zniszczeniu, protoplasty łączą się, w wyniku czego powstaje pojedyncza sieć. Takie mleczne występują u Compositae. niesegmentowane lactifers są utworzone przez olbrzymią komórkę wielojądrzastą. Powstaje we wczesnych stadiach rozwoju, wydłuża się i rozgałęzia w miarę wzrostu rośliny, tworząc złożoną sieć, na przykład u roślin wilczomlecza (ryc. 25).
Ściany komórkowe lactifers są silnie nawodnione. Nie zdrewniały i dlatego są plastyczne. Centralną część lactis zajmuje wakuola z lateksem, a protoplast zajmuje pozycję przyścienną. Nie zawsze można określić granicę między protoplastem a wakuolą. Laktyfery penetrują merystem i rosną razem z organem rośliny.
Tkanki przewodzące
Ten typ należy do trudny tkanek, składa się z różnie zróżnicowanych komórek. Oprócz właściwych elementów przewodzących tkanka zawiera elementy mechaniczne, wydalnicze i magazynujące (ryc. 26). Tkanki przewodzące łączą wszystkie narządy roślin w jeden system. Istnieją dwa rodzaje tkanek przewodzących: ksylem I łyko(Grecki xylon - drzewo; phloios - kora, łyk). Mają zarówno różnice strukturalne, jak i funkcjonalne.
Elementy przewodzące ksylemu są utworzone przez martwe komórki. Przenoszą dalekosiężny transport wody i rozpuszczonych w niej substancji od korzenia do liści. Przewodzące elementy łyka utrzymują protoplast przy życiu. Przenoszą transport na duże odległości od liści fotosyntetyzujących do korzenia.
Zwykle ksylem i łyko znajdują się w ciele rośliny w określonej kolejności, tworząc warstwy lub wiązki przewodzące. W zależności od budowy wyróżnia się kilka rodzajów wiązek przewodzących, które są charakterystyczne dla niektórych grup roślin. W otwartym pakiecie zabezpieczeń między ksylemem a łykiem znajduje się kambium, które zapewnia wzrost wtórny (ryc. 27-A, 28). W dwustronnym pakiecie otwartymłyko jest usytuowane względem ksylemu po obu stronach (ryc. 27-B, 29). wiązki zamknięte nie zawierają kambium, a zatem nie są zdolne do wtórnego zgrubienia (ryc. 27-B, 27-D, 30.31). Są jeszcze dwa rodzaje wiązki koncentryczne, gdzie albo łyko otacza ksylem (ryc. 27-E, 32), albo ksylem otacza łyko (ryc. 27-E).
Ksylem (drewno). Rozwój ksylemu w roślinach wyższych wiąże się z zapewnieniem wymiany wody. Ponieważ woda jest stale wydalana przez naskórek, taka sama ilość wilgoci musi zostać wchłonięta przez roślinę i dodana do narządów przeprowadzających transpirację. Należy wziąć pod uwagę, że obecność żywego protoplastu w komórkach przewodzących wodę znacznie spowolniłaby transport, a martwe komórki są tutaj bardziej funkcjonalne. Jednak martwej komórki nie ma jędrność , dlatego powłoka musi mieć właściwości mechaniczne. Notatka: turgescencja - stan komórek, tkanek i narządów roślinnych, w którym uelastyczniają się pod wpływem nacisku zawartości komórek na ich sprężyste błony. Rzeczywiście, elementy przewodzące ksylemu składają się z martwych komórek rozciągniętych wzdłuż osi narządu z grubymi zdrewniałymi muszlami.
Początkowo ksylem tworzy się z merystemu pierwotnego – prokambium, znajdującego się na wierzchołkach narządów osiowych. Najpierw różnicuje protoksylem, Następnie metaksylem. Znane są trzy rodzaje tworzenia ksylemów. Na egzarcha typu, elementy protoksylemowe pojawiają się najpierw na obrzeżach pęczka prokambalnego, następnie elementy metaksylemowe pojawiają się w środku. Jeśli proces przebiega w przeciwnym kierunku (tj. od centrum do peryferii), to tak endarchiczny typ. Na typu mesarch ksylem jest układany w środku pęczka prokambalnego, po czym jest odkładany zarówno w kierunku centrum, jak i na obrzeżach.
Korzeń charakteryzuje się tworzeniem ksylemu typu egzarchowego, podczas gdy łodygi mają typ endarchiczny. W roślinach słabo zorganizowanych metody tworzenia ksylemów są bardzo zróżnicowane i mogą służyć jako charakterystyka systematyczna.
W niektórych roślinach (na przykład jednoliściennych) wszystkie komórki prokambalne różnicują się w tkanki przewodzące, które nie są zdolne do wtórnego zagęszczania. W innych formach (na przykład zdrewniałych) merystemy boczne (kambium) pozostają między ksylemem a łykiem. Komórki te są w stanie dzielić się, odnawiając ksylem i łyko. Taki proces nazywa się wzrost wtórny. Wiele roślin rosnących w stosunkowo stabilnych warunkach klimatycznych stale rośnie. W formach dostosowanych do sezonowych zmian klimatu – okresowo. W rezultacie powstają dobrze zaznaczone słoje wzrostu rocznego.
Główne etapy różnicowania komórek prokambalnych. Jego komórki mają cienkie błony, które nie zapobiegają ich rozciąganiu podczas wzrostu narządu. Następnie protoplast zaczyna układać błonę wtórną. Ale ten proces ma wyraźne cechy. Błona wtórna nie jest osadzana w ciągłej warstwie, która uniemożliwiałaby rozciąganie się komórki, ale w postaci pierścieni lub spirali. Wydłużanie komórek nie jest trudne. W młodych komórkach pierścienie lub zwoje helisy znajdują się blisko siebie. W dojrzałych komórkach rozchodzą się one w wyniku rozciągania komórek (ryc. 33). Pierścieniowe i spiralne zgrubienia muszli nie zapobiegają wzrostowi, jednak mechanicznie ustępują muszlom, gdzie wtórne zgrubienia tworzą ciągłą warstwę. Dlatego po ustaniu wzrostu w ksylemie powstają elementy z ciągłą zdrewniałą skorupą ( metaksylem). Należy zauważyć, że wtórne zgrubienie nie jest tu pierścieniowe ani spiralne, ale punktowe, schodkowe, siatkowate (ryc. 34). Jego komórki nie są w stanie się rozciągać i umierają w ciągu kilku godzin. Proces ten w pobliskich komórkach zachodzi w sposób skoordynowany. W cytoplazmie pojawia się duża liczba lizosomów. Następnie rozkładają się, a zawarte w nich enzymy niszczą protoplast. Po zniszczeniu ścian poprzecznych komórki ułożone w łańcuch jedna nad drugą tworzą wydrążone naczynie (ryc. 35). Większość roślin okrytonasiennych i niektóre paprocie mają naczynia.
Nazywa się komórkę przewodzącą, która nie tworzy się przez perforacje w jej ścianie tchawica. Ruch wody przez cewki jest wolniejszy niż przez naczynia. Faktem jest, że w tracheidach pierwotna błona nie jest nigdzie przerwana. Tracheidy komunikują się ze sobą poprzez od. Należy wyjaśnić, że w roślinach pory są tylko zagłębieniami w błonie wtórnej aż do błony pierwotnej i nie ma perforacji przelotowych między tracheidami.
Najczęściej są to pory obramowane (ryc. 35-1). Ich kanał, skierowany w stronę jamy komórki, tworzy przedłużenie - komora porowa. Pory większości roślin iglastych na pierwotnej skorupie mają pogrubienie - torus, który jest rodzajem zaworu i jest w stanie regulować intensywność transportu wody. Przesuwając się, torus blokuje przepływ wody przez pory, ale po tym nie może już powrócić do poprzedniej pozycji, wykonując jednorazową akcję.
Pory są mniej lub bardziej zaokrąglone, wydłużone prostopadle do wydłużonej osi (grupa tych porów przypomina drabinę, więc ta porowatość nazywana jest porowatością drabinkową). Transport przez pory odbywa się zarówno w kierunku wzdłużnym, jak i poprzecznym. Pory są obecne nie tylko w tracheidach, ale także w poszczególnych komórkach naczyniowych tworzących naczynie.
Z punktu widzenia teorii ewolucji tracheidy są pierwszą i główną strukturą przewodzącą wodę w ciele roślin wyższych. Uważa się, że naczynia powstały z tchawicy w wyniku lizy ścian poprzecznych między nimi (ryc. 36). Większość paproci i nagonasiennych nie ma. Poruszają wodę przez tracheidy.
W procesie rozwoju ewolucyjnego naczynia powstawały wielokrotnie w różnych grupach roślin, ale najważniejszego znaczenia funkcjonalnego nabrały u roślin okrytonasiennych, które mają je wraz z tracheidami. Uważa się, że posiadanie bardziej zaawansowanego mechanizmu transportu pomogło im nie tylko przetrwać, ale także osiągnąć znaczną różnorodność form.
Xylem jest złożoną tkanką, oprócz elementów przewodzących wodę zawiera inne. Funkcje mechaniczne są wykonywane przez włókna libriform ( łac. liber – lub, forma – forma). Obecność dodatkowych konstrukcji mechanicznych jest o tyle ważna, że pomimo pogrubienia ścianki elementów przewodzących wodę są wciąż zbyt cienkie. Nie są w stanie samodzielnie utrzymać dużej masy rośliny wieloletniej. Włókna rozwinęły się z cewek. Charakteryzują się mniejszymi rozmiarami, zdrewniałymi (zdrewniałymi) muszlami i wąskimi jamami. Na ścianie można znaleźć pory pozbawione obramowania. Włókna te nie mogą przewodzić wody, ich główną funkcją jest wsparcie.
Xylem zawiera również żywe komórki. Ich masa może osiągnąć 25% całkowitej objętości drewna. Ponieważ komórki te mają zaokrąglony kształt, nazywane są miąższem drzewnym. W ciele rośliny miąższ znajduje się na dwa sposoby. W pierwszym przypadku komórki są ułożone w postaci pionowych pasm - tak jest miąższ nici. W innym przypadku miąższ tworzy promienie poziome. Nazywają się promienie rdzenia, ponieważ łączą rdzeń i korę. Rdzeń pełni szereg funkcji, w tym magazynowanie substancji.
Łyko (łyk). Jest to złożona tkanka, ponieważ tworzą ją heterogenne komórki. Nazywa się główne komórki przewodzące elementy sitowe(Rys. 37). Elementy przewodzące ksylemu tworzą martwe komórki, podczas gdy w łyku zachowują żywy, choć znacznie zmieniony, protoplast w okresie funkcjonowania. Łyko powoduje odpływ substancji plastycznych z organów fotosyntetyzujących. Wszystkie żywe komórki roślin posiadają zdolność przewodzenia substancji organicznych. I stąd, jeśli ksylem występuje tylko w roślinach wyższych, to transport substancji organicznych między komórkami odbywa się również w roślinach niższych.
Ksylem i łyko rozwijają się z merystemów wierzchołkowych. W pierwszym etapie w rdzeniu prokambalnym a protofloem. W miarę wzrostu otaczających tkanek rozciąga się, a po zakończeniu wzrostu tworzy się zamiast protofloemu metafloem.
W różnych grupach roślin wyższych można spotkać dwa rodzaje elementów sitowych. Jest reprezentowany w paprociach i nagonasiennych komórki sitowe. Pola sitowe w komórkach są rozrzucone wzdłuż ścian bocznych. Protoplast zachowuje lekko zniszczone jądro.
W roślinach okrytonasiennych nazywane są elementy sitowe rurki sitowe. Komunikują się ze sobą za pośrednictwem płyt sitowych. Dojrzałe komórki nie mają jąder. Jednak obok znajduje się rura sitowa komórka satelitarna, która powstaje wraz z rurką sitową w wyniku podziału mitotycznego wspólnej komórki macierzystej (ryc. 38). Komórka towarzysząca ma gęstszą cytoplazmę z dużą liczbą aktywnych mitochondriów, a także w pełni funkcjonujące jądro, ogromną liczbę plazmodesmy (dziesięć razy więcej niż inne komórki). Komórki towarzyszące wpływają na aktywność funkcjonalną niejądrowych komórek rurek sitowych.
Struktura dojrzałych komórek sitowych ma pewne cechy szczególne. Nie ma wakuoli, więc cytoplazma jest silnie upłynniona. Jądro może być nieobecne (u roślin okrytonasiennych) lub być w stanie pomarszczonym, funkcjonalnie nieaktywnym. Rybosomy i kompleks Golgiego są również nieobecne, ale retikulum endoplazmatyczne jest dobrze rozwinięte, które nie tylko penetruje cytoplazmę, ale także przechodzi do sąsiednich komórek przez pory pól sitowych. Dobrze rozwinięte mitochondria i plastydy występują w obfitości.
Pomiędzy komórkami transport substancji odbywa się przez otwory znajdujące się w błonach komórkowych. Takie otwory nazywane są porami, ale w przeciwieństwie do porów tchawicy są przelotowe. Przypuszcza się, że są to wysoce ekspandowane plazmodesmy, na ściankach których osadza się kalozowy polisacharyd. Pory są ułożone w grupy, tworząc pola sitowe. W formach prymitywnych pola sitowe są losowo rozrzucone na całej powierzchni skorupy, u bardziej zaawansowanych okrytozalążkowych znajdują się na końcach sąsiadujących ze sobą komórek, tworząc płyta sitowa(Rys. 39). Jeśli jest na nim jedno pole sitowe, nazywa się to prostym, jeśli jest ich kilka - złożonym.
Szybkość ruchu roztworów przez elementy sitowe wynosi do 150 cm ∕ godz. To tysiąc razy więcej niż tempo swobodnej dyfuzji. Prawdopodobnie zachodzi transport aktywny, a liczne mitochondria elementów sitowych i komórki satelitarne dostarczają niezbędnego do tego ATP.
Żywotność elementów sitowych łyka zależy od obecności merystemów bocznych. Jeśli są obecne, elementy sitowe działają przez cały okres eksploatacji rośliny.
Oprócz elementów sitowych i komórek satelitarnych łyko zawiera włókna łykowe, sklereidy i miąższ.
Pytania do kontroli i powtórzenia
Oznaki roślin wyższych? Jakie pokolenie roślin wyższych nazywa się gametofitem? Sporofit?
Jakie są przyczyny, które przyczyniają się do rozwoju zróżnicowanych tkanek u roślin lądowych.
Wypisz rodzaje roślin.
Jakie znasz grupy merystemów? Gdzie występują w ciele rośliny?
Tkanki powłokowe, ich funkcje, znaczenie.
Organizacja różnych typów tkanek powłokowych.
Budowa aparatu szparkowego. Wymień znane ci rodzaje aparatów szparkowych. Jak działają aparaty szparkowe?
Cechy tkanek miąższowych, rodzaje miąższu.
Oznaki miąższu i sklerenchymy.
Wymień rodzaje struktur wydalniczych rośliny. Jakie jest ich znaczenie?
Metody formowania zbiorników na wydalane substancje.
Co to jest dojarzami? Na jakie typy są podzielone?
Jak zorganizowane są tkanki przewodzące?
Co to są wiązki przewodzące? Ich typy.
Kierunki przemieszczania się substancji przez ksylem? Łyko? Czy skład transportowanych substancji różni się w ksylemie i łyku?
Budowa i funkcje ksylemu, łyka.
Organizacja elementów przewodzących łyka w roślinach z różnych grup systematycznych.
Wyspecjalizowane i bakteryjne, mają zewnętrzną ścianę komórkową. W komórkach wyższych Budowa i funkcje organelli komórki. Prawa pracy G. MendelControl >>
Typ tekstylia. Nasi najbliżsi krewni, inne ssaki, Struktura i... , składające się z jednego komórki. Rośliny składają się również z różnych… mikrotubul, centrioli ośrodek komórkowy. W warzywo komórki wydzielane są również chloroplasty, w których ...
Kolor czerwony. W tkanki rośliny pochodne antracenu można znaleźć ... pochodne antracenu jak w medycynie warzywo surowce i ... są otoczone 2-3, rzadziej 4 komórki naskórek. Włosy jednokomórkowe, ... związki, na bazie Budynki z czego struktura antracenu...