W jądrach pierwotniaków, a także w jądrach organizmów wielokomórkowych, jest otoczka, sok jądrowy (kariolimfa), chromatyna (chromosomy) i jąderka. Jednak pod względem wielkości i struktury jądra różne pierwotniaki są bardzo zróżnicowane (ryc.). Różnice te wynikają z proporcji Elementy konstrukcyjne jądra: ilość soku jądrowego, liczba i wielkość jąderek (jąderek), stopień zachowania struktury chromosomów w jądrze międzyfazowym itp.
Ryż. Jądra pierwotniaków.
A - mikrojądro orzęsków Paramecium aurelia;
B - wiciowiec Trypanosoma brucei,
B - ameba Ameba sphaeronuclei;
G - wiciowiec pancerny Geratium fuscus;
E - radiolarian Aulacantha scolymantha;
E - makrogameta kokcydiów Aggregata eberthi;
G - makrojądro orzęsków Epidinium ecaudatum
Większość pierwotniaków ma jedno jądro. Istnieją jednak również wielojądrowe gatunki pierwotniaków.
U niektórych pierwotniaków, mianowicie infuzorii i kilku kłączy - otwornic, obserwuje się ciekawe zjawisko dualizmu (dwoistości) aparatu jądrowego. Sprowadza się to do tego, że w organizmie najprostszym znajdują się dwa jądra dwóch kategorii, różniących się zarówno budową, jak i fizjologiczną rolą w komórce. Na przykład Infusoria mają dwa rodzaje jąder: duże jądro bogate w chromatynę - makrojądro i małe jądro - mikrojądro. Pierwsza związana jest z pełnieniem funkcji wegetatywnych w komórce, druga z procesem seksualnym.
Najprostsze, jak wszystkie organizmy, mają tendencję do rozmnażania się. Istnieją dwie główne formy rozmnażania się pierwotniaków: bezpłciowe i płciowe. Oba opierają się na procesie podziału komórki.
Przy rozmnażaniu bezpłciowym liczba osobników wzrasta w wyniku podziału. Na przykład ameba podczas rozmnażania bezpłciowego jest podzielona na dwie ameby przez zwężenie ciała. Proces ten rozpoczyna się od jądra, a następnie obejmuje cytoplazmę. Czasami rozmnażanie bezpłciowe nabiera charakteru wielokrotnego podziału. W tym przypadku jądro jest wstępnie podzielone kilka razy, a najprostsze staje się wielojądrowe. Następnie cytoplazma rozpada się na szereg przedziałów odpowiadających liczbie jąder.
W rezultacie organizm najprostszych natychmiast rodzi znacząca liczba małe osobniki. W ten sposób zachodzi na przykład rozmnażanie bezpłciowe plazmodium malarii, czynnika sprawczego ludzkiej malarii.
rozmnażanie płciowe pierwotniaki charakteryzują się tym, że właściwy rozród (wzrost liczby osobników) poprzedzony jest procesem płciowym, piętno czyli fuzja dwóch komórek płciowych (gamet) lub dwóch jąder płciowych, prowadząca do powstania jednej komórki – zygoty, dająca początek nowej generacji.
Formy procesu płciowego i rozmnażania płciowego u pierwotniaków są niezwykle różnorodne.
W przypadku rozmnażania pierwotniaków charakterystyczne są dwa rodzaje procesów. Jeden z nich - rozmnażanie agamiczne lub bezpłciowe - sprowadza się do wzrostu liczby osobników danego gatunku i może przebiegać na różne sposoby. Osobniki powstałe w wyniku takiej reprodukcji mogą mieć także inny los. Mogą bezpośrednio wyrosnąć na organizm podobny do matki, przejść metamorfozę (jak włóczęga suctoria lub peritrich), całkowicie przekształcić się w gametę lub gametocyt (jak sporozoity gregaryny).
Ponadto proces seksualny jest włączony w cykl życiowy wielu protistów. Wiąże się to z powstawaniem specjalnych stadiów - gamet, które mogą się uformować różne sposoby. Procesowi płciowemu może w ogóle nie towarzyszyć reprodukcja (koniugacja, autogamia, partenogeneza). Najważniejszym momentem procesu płciowego jest przebudowa jądra i innych składników komórki, która przebiega przez szereg faz i prowadzi do powstania nowego pokolenia, które różni się żywotnością i właściwościami dziedzicznymi od poprzednich pokoleń. Jak już wspomniano, nie wszystkie rodzaje procesu płciowego prowadzą do wzrostu liczby osobników, dlatego w odniesieniu do pierwotniaków zwykle mówi się, że z jednej strony mają rozmnażanie bezpłciowe, az drugiej proces płciowy.
^
Rozmnażanie bezpłciowe
Prosty podział na pół lub monotomia. Ten rodzaj rozmnażania występuje najczęściej w pierwotniakach i komórkach tkanek wielokomórkowych. W typowym przypadku jest to prawidłowy, równomierny podział organizmu najprostszego na dwa identyczne osobniki potomne, poprzedzony okresem odżywiania i wzrostu. Klasyczny przykład reprodukcja ameby. U ameby podział nie ma określonej orientacji. Ciało ameby jest rozciągnięte, na równiku zarysowuje się poprzeczne zwężenie, które stopniowo się pogłębia, aż wreszcie komórka dzieli się na dwie połowy, które natychmiast zaczynają się poruszać i żywić samodzielnie. Podziałowi cytoplazmy towarzyszy podział mitotyczny jądra. Inni organelli komórkowych albo dzielą się same, albo są po prostu rozdzielone między osobniki potomne, a brakujące formują się na nowo. Ten sam niezorientowany podział obserwuje się u części słoneczników i radiolarianów.
Obecność szkieletu oczywiście nieco komplikuje ten proces. W amebach testatowych - testacey (Arcella) i gromiid (Euglypha) przed podziałem podstawy płyt krzemionkowych są układane w specjalnych wakuolach. Następnie rozpoczyna się podział, którego płaszczyzna jest prostopadła do osi podłużnej skorupy. Jądro dzieli się i około połowa cytoplazmy wraz z jądrem potomnym i płytkami krzemionkowymi wychodzi z ujścia na zewnątrz i jest otoczona nową otoczką. W radiolarianach, które mają szkielet złożony z dwóch zastawek, położenie płaszczyzny rozszczepienia jest określone przez położenie zastawek. Podczas podziału ulotki rozpraszają się wraz z połową cytoplazmy, a brakująca ulotka zostaje uzupełniona.
U wiciowców i orzęsków pozycja płaszczyzny podziału jest bardziej stała i stabilna. U wiciowców występuje zwykle w kierunku podłużnym, u orzęsków - w kierunku poprzecznym. U wiciowców podział rozpoczyna się na przednim końcu ciała i trwa w kierunku tylnego końca. W tym przypadku zdarza się, że oddzielające połówki komórki są wygięte w przeciwnych kierunkach, co stwarza wrażenie poprzecznego podziału. U wiciowców płaszczyzna podziału jest odchylona od podłużnej – podział następuje w kierunku ukośnym. Również w ukośnym kierunku jest płaszczyzna podziału opali.
W złożonych mastigoforach podział jest związany ze zjawiskami reorganizacji. W tym samym czasie niektóre organelle są resorbowane i ponownie formowane u osobników potomnych. Podobne zjawiska mogą wystąpić w ziarnistościach podstawnych wici, aksostylu i wici polimastiginowej.
U orzęsków proces podziału prowadzi do jeszcze bardziej złożonej reorganizacji niż u wiciowców. Tak więc u hipotrychów, u osobników potomnych przednich, stare błony adoralne stopniowo zmniejszają się, a jednocześnie u ich podstawy pojawiają się zaczątki nowej błony, która rośnie w miarę wchłaniania starych. Podobnie wymieniane są cirri brzuszne, dzięki czemu obie córki otrzymują zupełnie nowy zestaw organelli ruchowych.
Specjalny przypadek reorganizacja orzęsków - stomatogeneza. Stomatogeneza to proces morfogenetyczny, który można zaobserwować podczas podziału, regeneracji orzęsków, a także podczas przekształcania form mikrostomicznych w makrostomiczne. Pierwszym etapem stomatogenezy jest tworzenie nowych kinetosomów, które tworzą zalążek rzęsek okołoustnych. Ten etap o godz różne grupy orzęski przebiegają na różne sposoby, co stanowi podstawę konstrukcji filogenetycznych i systematycznych.
Początkowa postać stomatogenezy jest uważana za telokinetalną - tworzenie podstaw orzęsków, w których kinetosomy pochodzą z przednich segmentów kinet somatycznych lub sąsiednich fragmentów kinet. Jest to typowe dla przedstawicieli klasy Kinetofragminophora.
Wiele hymenostoma, spirotrichus i peritrichus ma stomatogenezę policzkowo-kinetyczną - obok kinetosomów starego aparatu jamy ustnej powstają nowe kinetosomy. W której ciała podstawowe wyspecjalizowane sekcje perystomu dają początek różnym składnikom nowej jamy ustnej. Oddzielenie dwóch nowo utworzonych kompleksów rzęsek następuje poprzez serię złożonych ruchów, takich jak przesunięcia i obroty.
Stomatogeneza parakinetalna występuje u niektórych błonkówek i krętków. Wraz z nim stare struktury jamy ustnej rozpuszczają się, a nowe powstają z powodu (lub w pobliżu) somatycznych rzędów rzęsek po brzusznej stronie ciała.
Jeśli nie można ustalić związku między nowymi kinetosomami a dawnymi strukturami jamy ustnej, mówi się o stomatogenezie apokinetycznej.
Natychmiast po utworzeniu ciał podstawowych już się pokazują różne systemy mikrotubule, takie jak nicienie, mikrotubule postrzęskowe i poprzeczne. Następnie kinetosomy migrują do plazmalemmy, gdzie zaczynają rosnąć rzęski.
NA Następny krok stomatogenezie, kinetosomy ustawiają się w szczątkach pewien porządek. Proces ten przebiega w określonym kierunku iw ściśle określonej kolejności. Pod koniec stomatogenezy rozwija się aparat do wychwytywania pokarmu - strefa, z której wyplatają się wakuole pokarmowe.
palintomia. Palintomia nazywa się podziałem na pół, powtarzanym wielokrotnie, bez pośredniego etapu odżywiania i wzrostu. Cechami charakterystycznymi palintomii są: 1. Wcześniejszy wzrost przerostowy osobnika matki, który umożliwia jej wielokrotne podziały. 2. Palintomia bardzo często występuje w okresach cyklu życia, które poprzedzone są wzmożonym odżywieniem organizmu. 3. Palintomia charakteryzuje się zwykle tymczasowym odróżnicowaniem produktów rozszczepienia, ponieważ produkty rozszczepienia nie mają czasu na przejście przez proces reorganizacji. Dlatego palintomia często odbywa się na etapie torbieli. Palintomię dzieli się na liniową, gdy komórki potomne układają się w łańcuch (bruzdnice, orzęski-apostomata) i tabelaryczną, która powstaje w wyniku naprzemiennych podziałów podłużnych w dwóch wzajemnie prostopadłych płaszczyznach, w wyniku czego produkty rozszczepienia są ułożone równolegle do siebie w płaskiej płytce. Taka palintomia jest charakterystyczna dla fitomonad, niektórych wiciowców i euglenidów. Palintomia jest często obserwowana podczas tworzenia gametocytów i gamet.
początkujący. Pączkowanie wywodzi się z nierównego podziału. Jednocześnie wyróżnia się morfologicznie osobnik ze strony matki (trofont) i córka (włóczęga, telotroch, nerka, tomit). Istnieją monotomiczne pączkowanie, gdy tworzy się jeden włóczęga, i pączkowanie wielokrotne. Ponadto różne grupy orzęsków mają pączkowanie wewnętrzne, pączkowanie zewnętrzne i niektóre przejściowe formy pączkowania. Pączkowanie jest charakterystyczne dla orzęsków siedzących - suctorium, peritrich, chonotrich, rhynchodid, występuje w słonecznikach. charakterystyczna cecha pączkowanie to brak reorganizacji u osobników matczynych - z reguły zachowują one wszystko niezmienione, w tym organelle ruchowe lub ustne. Wśród mastigoforów pączkowanie jest niezwykle rzadkie; w szczególności proste pączkowanie zewnętrzne jest znane u Haplozoon i niektórych chryzomonad. W słonecznikach znane jest wielokrotne pączkowanie zewnętrzne. W niektórych myxosporidium obserwuje się separację i separację części ciała stadiów plazmatycznych.
^ Synthomia lub podział wielokrotny . Przy tego rodzaju reprodukcji zwierzę na pewnym etapie koło życia(zwykle po okresie odżywiania i wzrostu) przechodzi szereg powtarzających się podziałów jądrowych i staje się przejściowo wielojądrowy, po czym natychmiast rozpada się na odpowiednią liczbę komórek. W zależności od tego, czy w wyniku procesu powstają osobniki wegetatywne, czy komórki rozrodcze, stadium rozmnażające się przez syntomię nazywa się schizontem lub gamontem, a gdy podział wielokrotny następuje bezpośrednio po procesie płciowym, sporontem. Produkty schizogonii nazywane są odpowiednio merozoitami, gamogonią - gametami i sporogonią - sporozoitami dla sporozoan i zoosporami dla innych grup. Syntomia jest szeroko rozpowszechniona wśród sporozoan, rzadziej u mastigoforów (bruzdnicowatych - Noctiluca) i nieobecna u orzęsków.
Plazmatomia. Plazmatomia jest rodzajem rozmnażania bezpłciowego, w którym ciało pierwotniaka wielojądrzastego dzieli się na dwie lub więcej wielojądrowe osobniki potomne. W tym przypadku podział komórki zachodzi niezgodnie z podziałami jądrowymi i niezależnie od nich. Plastomia obejmuje podział plazmodiów ślimaków, myxosporidium, ameby - pelomyksyny itp. Niektórzy autorzy kwestionują przypisanie tej metody rozmnażania do specjalnego typu.
^ Rozmnażanie kolonii. Inny rodzaj rozmnażania bezpłciowego. Istnieją trzy sposoby rozmnażania kolonii. Pierwszym jest podzielenie całej kolonii na dwie części. W ten sposób rozmnażają się kolonialne radiolarie i chrysomonady. Drugim sposobem jest oddzielenie osobników (włóczęgów) od kolonii, które tworzą nowe kolonie. Metoda ta występuje u siedzących wiciowców i peritrichów (Zoothamnium arbuscula). Trzecia metoda jest podobna do drugiej, ale tutaj nie powstają włóczęgi, a osobniki założycielskie pozostają w starej kolonii i dają początek kolonii potomnej. Jest to charakterystyczne dla kolonialnych volvocinów.
Należy zauważyć, że kolonialność jest również uważana za wynik rozmnażania bezpłciowego, które nie zostało zakończone, a ponieważ kolonialność jest uważana za ewolucyjną podstawę wielokomórkowości, ta ostatnia może również w pewnym stopniu wywodzić się z rozmnażania bezpłciowego.
Główną formą rozmnażania pierwotniaków jest monotomia- prosty podział na dwie części. Pomiędzy dwoma podziałami zwierzę nabywa wszystkie struktury utracone w procesie podziału, aktywnie odżywia się, rośnie, a na początku nowego podziału ma czas, aby osiągnąć rozmiar osobnika rodzicielskiego (ameba, euglena, ceratium itp.) .
specjalny kształt reprodukcja pierwotniaków jest palintomia lub redystrybucja. Podział powtarza się wielokrotnie bez kolejnego okresu odżywiania i wzrostu, a produkty pośrednie palintomii pozostają niezróżnicowane, a ich wielkość zmniejsza się z każdym podziałem. Tylko najnowsza generacja osiąga pełny rozwój, odżywia się i rośnie. Podział palintomiczny zwykle przebiega pod osłoną błon ochronnych (podział oocysty w zarodźcu malarii, zygoty w volvox itp.).
U pierwotniaków powszechny jest również wielokrotny podział - syntomia(schizogonia). W tym przypadku najpierw następuje wielokrotny podział jądra bez podziału cytoplazmy, w wyniku czego komórka przejściowo staje się polienergetyczna, następnie cytoplazma natychmiast rozpada się na wiele (w zależności od liczby jąder) komórek potomnych (trypanosomów , zarodźca malarii, otwornice).
Proces płciowy u pierwotniaków
Proces płciowy może zachodzić u pierwotniaków w formie kopulacja I koniugacje. Na kopulacja następuje całkowite połączenie dwóch osobników płciowych - gamet. Proces, w którym gamety nie różnią się od siebie, nazywa się izogamią. W innych przypadkach powstają duże makrogamety (żeńskie) i małe mikrogamety (męskie); występuje anizogamia. Proces, w którym makrogamety stają się nieruchome i gromadzą zapasowe składniki odżywcze nazywa się oogamią.
Koniugacja jest bardziej złożoną formą procesu seksualnego i występuje tylko w orzęskach. W tym przypadku następuje tylko tymczasowe połączenie dwóch osobników i wymiana części mikrojąder. Prawdziwe gamety nie powstają w tym przypadku, ale migrujące jądro jest ubrane w cienką warstwę cytoplazmy i odpowiada mikrogamecie, a reszta ciała rzęskowego z nieruchomym jądrem to makrogameta. Migrujące (męskie) jądro każdego koniugatu przechodzi do ciała partnera i kopuluje z jego nieruchomym (żeńskim) jądrem - następuje zapłodnienie krzyżowe. Makrojądra nie biorą udziału w tym procesie.
cykle jądrowe
Cykl jądrowy jest związany z procesem seksualnym. Wspólne dla wszystkich cykli jądrowych jest przemiana pokoleń haploidalnych i diploidalnych. Gamety są zawsze haploidalne; po ich połączeniu powstaje diploidalna zygota; na pewnym etapie cyklu życiowego następuje podział redukcyjny (mejoza), który przywraca jądra do stanu haploidalnego.
Istnieją trzy rodzaje cykli jądrowych, z których każdy charakteryzuje się tym, jakie miejsce w tym cyklu zajmuje rozszczepienie redukujące (ryc. 3).
1) Cykle z redukcją zygotyczną. Większość cykl życiowy przypada na pokolenie haploidalne. Pokolenie diploidalne jest reprezentowane przez jedną zygotę, która dzieli się w sposób redukujący, podczas gdy haploidalność zostaje przywrócona. Takie cykle są charakterystyczne dla Phytomonadida, Sporozoa, części Polymastigida, części Hypermastlgida.
2) Cykle z redukcją gamet. Większość cyklu przypada na pokolenie diploidalne. Pokolenie haploidalne jest reprezentowane tylko przez gamety, podział redukcyjny następuje tuż przed ich powstaniem. Takie cykle są charakterystyczne dla pozostałych Polymastigida i Hypermaatlgida, Heliozoa, Ciliophora, Opalinida, Cnidosporidia i prawie wszystkich Metazoa.
3) Cykle z redukcją pośrednią. Pokolenia haploidalne i diploidalne zajmują w przybliżeniu równe części cyklu: zygota dzieli się mitotycznie. Pojawiają się bardziej rozwinięte stadia pokolenia diploidalnego (agamont). W pewnym momencie następuje podział redukcyjny z utworzeniem jednojądrowych haploidalnych agametów - rozmnażanie agamiczne (bezpłciowe). Agamety rozwijają się w bardziej zaawansowane stadia generacji haploidalnej. Następnie to dorosłe pokolenie haploidalne (gamont) rozpada się na gamety, które kopulują - zachodzi proces płciowy. Wraz z utworzeniem zygoty cykl się zamyka. Taki cykl występuje tylko u otwornic i jako rzadki wyjątek u niektórych wrotków.
Cykle życia
Cykle życiowe pierwotniaków są bardzo zróżnicowane. Istnieją pierwotniaki, które mają cykl bez procesu płciowego, tj. bez cyklu jądrowego (na przykład u trypanoz, w których naprzemiennie występują różne stadia wegetatywne).
W najbardziej zróżnicowanym cyklu życia wyróżnia się cztery okresy:
1) progamiczny, kończący się powstaniem gamet;
2) syngamiczny, w tym kopulacja gamet i tworzenie zygoty;
3) metagamiczne, obejmujące kilka pokoleń sporozoitów;
4) wegetatywny, podczas którego następuje wzrost, wzrost masy osobników.
W pewnym momencie cyklu życia następuje redukcja liczby chromosomów.
Dla wygody porównania cykli życia zestawimy tabelę, którą podzielimy pionowo na 4 części, odpowiadające wskazanym czterem okresom. W tej tabeli umieściliśmy kilka cykli najprostszych i Cykl organizm wielokomórkowy, umieszczając okresy ich cykli w odpowiednich kolumnach (ryc. 4). Wybieramy te najprostsze z najprostszymi i najbardziej rozczłonkowanymi cyklami.
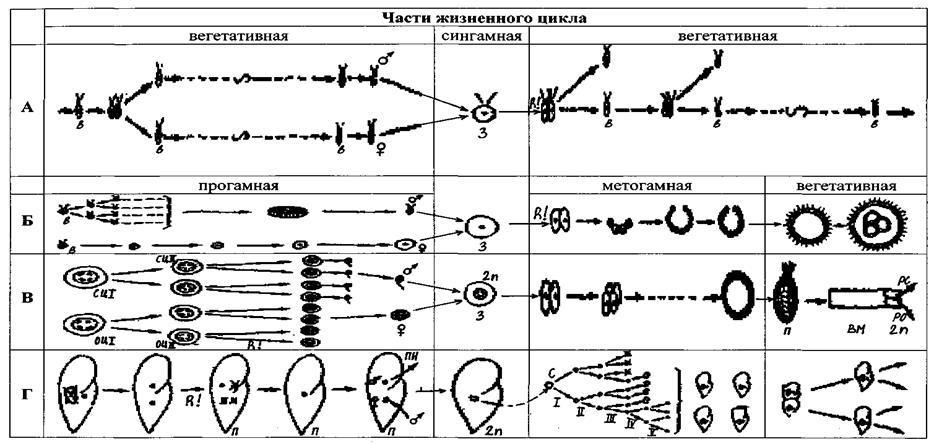
Ryc. 4. Porównanie cykli życiowych różnych pierwotniaków i organizmów wielokomórkowych (z K.V. Beklemishev, 1979):
A – nieskończenie zwielokrotniony cykl niższych fitomonad z fakultatywnym procesem płciowym; B - cykl Volvox z podziałem cyklu na części wegetatywną, progamiczną i metagamiczną; C - cykl organizmów wielokomórkowych ze zwiększonymi różnicami między częściami cyklu; D - cykl orzęsków (w metagamicznej części cyklu pokazany jest tylko podział jądrowy, nie pokazano konturów komórek). 3 - zygota, C - synkaryon, R! - podział redukcyjny, PN - przedjądrza, P - miąższ, rs - rozmnażanie i wzrost sporogonii, rho - rozmnażanie i wzrost oogonii, VM - dorosłe wielokomórkowe, sI i ssP - spermatocyty I i II rzędu, otsI i otsP - oocyty I i II zamówienia
Ze wszystkich różnorodnych cykli życiowych pierwotniaków z procesem seksualnym monoenergetyczne wiciowce, na przykład Chlamydomonas, mają najprostszy cykl. U niektórych gatunków tego rodzaju występuje wiele pokoleń bezpłciowych, które powstają w wyniku monotomii, proces płciowy następuje, gdy pogarszają się warunki bytowania. Gamety morfologicznie niewiele różnią się od osobników pokoleń bezpłciowych. Po kopulacji zygota jest ubrana w muszlę, pod osłoną której przez podział palintomii powstają 4–32 osobniki, które wyłaniają się z muszli i rozpoczynają rozmnażanie bezpłciowe. Redukcja liczby chromosomów następuje podczas pierwszych podziałów zygoty. W tym cyklu wszystkie stadia są haploidalne, jedynym etapem diploidalnym jest zygota. To jest cykl redukcji zygoty. U innych gatunków z rodzaju Chlamydomonas rozmnażanie bezpłciowe zachodzi przez palintomię, gamety również powstają w wyniku palintomii.
Najbardziej przeanalizowany cykl życiowy występuje w „wielokomórkowym” pierwotniaku – Volvox. W toku zoologii niewiele uwagi poświęca się temu obiektowi, ponieważ. ma autotroficzny rodzaj odżywiania. Jednak Volvox bierze ważne miejsce w anatomii porównawczej bezkręgowców, ponieważ jego cykl życiowy jest najbliższy cyklowi organizmów wielokomórkowych. Cykl życiowy Volvox charakteryzuje się ściśle ustalonym miejscem procesu płciowego, który poprzedzony jest progamiczną częścią cyklu, w trakcie której zachodzi homogonia, tj. przekształcenie specjalnych komórek generatywnych w gamety: makrogamont rośnie, gromadzi w sobie składniki odżywcze i zamienia się w dużą nieruchomą makrogametę, mikrogamont dzieli się palintomicznie, dając początek duża liczba mikrogamety. Następnie następuje fuzja gamet (syngamiczna część cyklu) i podział redukcyjny. Dalej - część metagamiczna - zygota, dzieli się palintomicznie, prowadząc do powstania kolonii przypominającej blastulę, bardzo przypominającej stadium blastuli organizmów wielokomórkowych; ta kolonia rośnie, tworzą się w niej nowe kolonie potomne - po metagamii pojawia się wegetatywna część cyklu. W cyklu życiowym Volvox należy zwrócić uwagę na fakt, że rozwój kolonii potomnej zygoty następuje najpierw w stanie zakrzywienia (tj. wydrążona kula) i dopiero przed wyjściem z utworzonej kolonii potomnej następuje wypukłość od matki - wywinięcie, w wyniku którego komórki przybierają ostateczny układ z wici na zewnątrz. Podział redukcyjny u Volvox, podobnie jak u sporozoanów, następuje po utworzeniu zygoty – cyklu z redukcją zygoty. Cykl orzęsków jest bardzo osobliwy. Podręczniki zoologii podają opis cyklu orzęsków bez uwzględnienia cyklu jądrowego. Dlatego należy zwrócić uwagę na fakt, że w części programowej uczestniczy tylko mikrojądro, które uległo podziałowi redukcyjnemu, z czterech utworzonych jąder haploidalnych trzy ulegają redukcji, a pozostaje jedno, które dzieli się z utworzeniem dwóch przedjąder. Żeńskie przedjądrze zamienia się w jądro nieruchome, pozostaje w orzęskach, a zatem całość odpowiada makrogamecie, a męskie przedjądrze zamienia się w jądro migrujące odpowiadające mikrogamecie i przechodzi do partnera, łącząc się z jej nieruchomym jądrem ( syngamiczna część cyklu). W metagamicznej części cyklu orzęsków nie obserwuje się palintomii; następuje podział synkarionu, który po serii przegrupowań prowadzi do przywrócenia dwujądrowej natury każdego osobnika i przywrócenia poliploidalności makrojądra. W wegetatywnej części cyklu orzęski rosną i rozmnażają się. bezpłciowo. Jest to cykl z redukcją gamet.
W cyklu życiowym każdego organizmu wielokomórkowego, podobnie jak u Volvox, występują części progamiczne, syngamiczne, metagamiczne i wegetatywne. Ale część programowa staje się bardziej skomplikowana ze względu na fakt, że organizmy wielokomórkowe przechodzą w tym okresie złożoną restrukturyzację aparatu jądrowego: tworzenie tetraploidalnych oo- i spermatocytów pierwszego rzędu, z których diploidalne oo- i spermatocyty drugiego rzędu i wreszcie podział redukcyjny, w wyniku którego powstają haploidalne gamety.
Metagamiczna palintomiczna część cyklu metazoan jest bardzo podobna do tej u fitomonad. Jednak wegetatywna część cyklu jest inna: jest scharakteryzowana wysoki stopień integracji ciała i dużej różnorodności komórek.
Z tabeli wynika, że u rozważanych pierwotniaków cykl Volvox jest najbliższy cyklowi życiowemu organizmów wielokomórkowych. Jego część metagamiczna kończy się utworzeniem kulistego stadium przypominającego blastulę; podobnie jak Metazoa ma część wegetatywną, podczas której organizm (lub kolonia) rośnie, jego komórki różnicują się, a tylko niewielka ich część specjalizuje się w generatywnych . Główną różnicą między cyklem życia Volvox a cyklem wielokomórkowym jest miejsce dział redukcji. Tak szczegółowe porównanie cykli różnych pierwotniaków i wielokomórkowców jest niezbędne do zrozumienia pochodzenia zwierząt wielokomórkowych. Należy jednak jasno zrozumieć, że Volvox w żadnym wypadku nie powinien być uważany za bezpośredniego przodka organizmów wielokomórkowych. Obiekt ten służy jedynie jako model dogodny do zrozumienia procesów, jakie mogły zachodzić w ewolucji kolonialnych heterotroficznych wiciowców, najprawdopodobniej bliskich współczesnym Choanoflagellata, które obecnie uważane są za przodków wielokomórkowców. Wśród współczesnych wiciowców obrożnych nie ma gatunków o procesie płciowym, ale uważa się, że to właśnie takie kolonialne Choanoflagellata, które miały kształt kuli i cykl życiowy typu Volvox, ale z redukcją gamet, były przodkami wiciowców Organizmy wielokomórkowe.
Bezpłciowy - przez mitozę jądra i podział komórki na dwie części (u ameby, eugleny, orzęsków), a także przez schizogonia- podział wielokrotny (u sporozoa).
Seksualny - kopulacja. Komórka pierwotniaka staje się funkcjonalną gametą; W wyniku fuzji gamet powstaje zygota.
W przypadku orzęsków charakterystyczny jest proces seksualny - koniugacja. Polega ona na tym, że komórki wymieniają informację genetyczną, ale nie następuje wzrost liczby osobników.
Wiele pierwotniaków może istnieć w dwóch formach - trofozoit(forma wegetatywna zdolna do aktywne odżywianie i ruch) i cysty, która powstaje w niesprzyjających warunkach. Komórka jest unieruchomiona, odwodniona, okryta gęsta skorupa metabolizm drastycznie zwalnia. W tej formie pierwotniaki są łatwo przenoszone na duże odległości przez zwierzęta, przez wiatr i są rozpraszane. Kiedy trafiłeś korzystne warunki siedliska następuje ekscystacja, komórka zaczyna funkcjonować w stanie trofozoitu. Zatem otorbienie nie jest sposobem rozmnażania, ale pomaga komórce przetrwać niekorzystne warunkiśrodowisko.
Dla wielu przedstawicieli tego typu pierwotniaki charakteryzuje się obecnością koło życia, polegający na regularnej przemianie formy życia. Z reguły następuje zmiana pokoleń z rozmnażaniem bezpłciowym i płciowym. Tworzenie cyst nie jest częścią regularnego cyklu życia.
Czas Pokolenie dla pierwotniaków jest to 6-24 h. Oznacza to, że już w organizmie żywiciela komórki zaczynają się namnażać wykładniczo i teoretycznie mogą doprowadzić do jego śmierci. Tak się jednak nie dzieje, ponieważ zaczynają działać mechanizmy ochronne organizmu gospodarza.
Znaczenie medyczne mają przedstawiciele pierwotniaków należących do klas sarkodów, wiciowców, orzęsków i sporozoanów.
Typ Protozoa obejmuje organizmy, których ciało składa się z pojedynczej komórki, która funkcjonuje jednak jako cały organizm. Komórki pierwotniaków są zdolne do wyżywienie we własnym zakresie, ruch, ochrona przed wrogami i doświadczanie niesprzyjających warunków.
Odżywianie pierwotniaków odbywa się za pomocą wakuoli trawiennych, przeprowadzanych przez faga - lub pinocytozę. Pozostaje niestrawione jedzenie wyrzucony. Niektóre pierwotniaki zawierają chloroplasty i są w stanie żywić się fotosyntezą.
Większość pierwotniaków ma organelle do poruszania się: wici, rzęski i pseudopodia (tymczasowe ruchome wyrostki cytoplazmy). Formy organelli ruchu leżą u podstaw taksonomii pierwotniaków.
Zwykle przeprowadza się rozmnażanie pierwotniaków Różne formy podziały są odmianami mitozy. Charakterystyczny jest również proces płciowy: w postaci fuzji komórek - kopulacji - lub wymiany części materiału dziedzicznego - koniugacji.
Większość pierwotniaków ma jedno jądro, ale są też formy wielojądrowe.
Znaczenie medyczne mają pierwotniaki należące do klas Sarcodaceae, wiciowce, orzęski i zarodniki.