Edinstven na svoj način zagotavlja njihovo raznolikost in značilnosti življenja.
Sodobna mikrobiologija za preučevanje in razvrščanje uporablja naslednje značilne značilnosti bakterije:
- Morfološka celična zgradba:
- sferični "koki";
- paličasti "bacili";
- spiralne "spirohete";
- zvita oblika "vibrios".
- Dih:
- dihajte kisik "aerobno";
- anoksični "anaerobni".
- Način hranjenja:
- avtotrofni "neodvisni" - sposobni sintetizirati organska snov iz anorganske s pomočjo energije kemične reakcije ali fotosinteza (na primer modrozelene alge);
- heterotrofni - pretvarjajo in predelujejo že pripravljene organske snovi, pridobljene kot posledica vitalne aktivnosti drugih organizmov, to vključuje patogene (patogene) in simbiotske (koristne) mikroorganizme, pa tudi bakterije, ki živijo zaradi procesov fermentacije, razpada itd. .
- Habitat:
- zunanje okolje (tla, voda itd.);
- znotraj živih organizmov;
- odmrle organske snovi (trupla, usedline, odpadki).
- Način porazdelitve in preživetja (ali se tvorijo celične spore, ovoji, sluz).
- Reakcija na Gramov test (ločevanje glede na sestavo in strukturo sten, ključna klasifikacija v kontekstu teme strukture celične stene):
- gram-pozitiven - obarvan med postopkom obarvanja po Gramu (lupina je prepustna za anilinsko barvilo (kristalno vijolično, metil vijolično itd.));
- po Gramu negativni - med postopkom barvanja po Gramu ne barvajte (celična stena je neprepustna za barvilo).
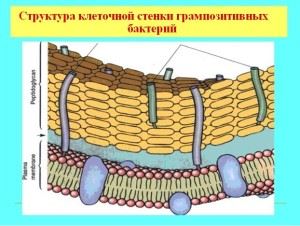
Značilnosti celične stene gram-pozitivnih bakterij
Bistvo testa (ki ga je leta 1884 predlagal G. K. Gram) je postopek obdelave vzorca bakterijske kulture z anilinskim barvilom (kristalno vijolično, kristalno vijolično). Po naknadnem pranju z alkoholom Gram (+) vrste mikroorganizmov ohranijo modro barvo, druge pa se obarvajo. Sposobnost celične stene za fiksiranje barvila je določena s sestavo in zgradbo celične stene. Razvrstitev enoceličnih organizmov glede na vrsto strukture celične stene, ki jo je mogoče obarvati ali ne z metodo po Gramu, je ena ključnih. V praksi rezultati gram testa pomagajo pri diagnosticiranju nalezljive bolezništudija biokemijske lastnosti mikroorganizmi.
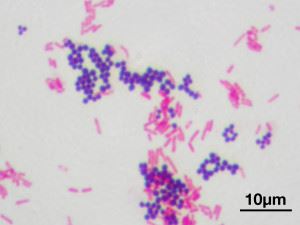
Barvanje po Gramu. Koki (sferični) - gram-pozitivni in bacili (palčke) - gram-negativni
Večina gram-pozitivnih bakterij ima masivno, enoslojno membrano, ki je prepustna za anilinsko barvilo. Sestava stene vključuje kompleksne organske molekule, osnovo mehanske togosti dajejo murein (heteropolimer), glikopeptidi, mukopeptidi. Struktura stene določa obliko mikroorganizma in je nekaj podobnega zunanjemu skeletu, s porozno strukturo, debelo približno 40 glikopeptidnih molekul. Poleg peptidoglikanske baze lupina vsebuje še teihojske kisline in polisaharide.
Kljub splošna reakcija na barvilu lahko med gram-pozitivnimi organizmi srečamo organizme, ki pripadajo različnim podvrstam, različne oblike, velikost, življenjski prostor in način prehranjevanja.
Večina patogenih bakterij za ljudi je gra-pozitivnih, med katerimi so na primer povzročitelji takšnih nalezljivih bolezni:
- tetanus (Clostridium tetani),
- botulizem (Clostridium botulinum),
- antraks (),
- streptokoki,
- stafilokoki (Staphylococcus aureus), ki so zelo odporni na temperaturo in delovanje zdravila.
Kislinsko odporni, zaradi posebne večplastne strukture lupine (vključuje voske, polisaharide in beljakovine), praviloma niso obarvani z metodo po Gramu. Zanje se uporablja posebna metoda Ziel-Nelsen. Toda nekateri dajejo pozitiven rezultat po metodi po Gramu v primeru zelo visoke koncentracije barvila oz povišana temperatura med postopkom.
Kisloodporni enocelični organizmi preživijo v kislih in alkalna okolja, čeprav je normalno okolje za njihov razvoj in rast nevtralno. običajno razvrščeni kot po Gramu pozitivni. Predstavniki te skupine, nevarni za ljudi in živali, so povzročitelji bolezni, kot sta tuberkuloza in gobavost. Na kisline so odporne tudi nekatere talne bakterije, ki lahko vežejo atmosferski dušik in so rastlinski simbionti.
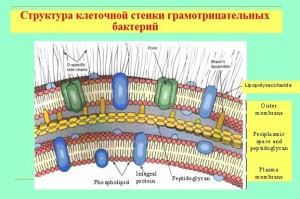
Značilnosti lupine gram-negativnih bakterij
Čeprav je debelina njihove mukopeptidne stene precej tanjša, je stena neprepustna za kristalno vijolično barvilo. Gre za bistveno drugačno strukturo in kemično sestavo celice. Lupina ima večplastno strukturo in je sestavljena iz:
- zunanja lupina (membans), njena glavna sestava so liposaharidi in beljakovine;
- periplazemski prostor;
- notranja plast (murein peptidoglikan), je po sestavi podobna stenam gram-pozitivnih bakterij, vendar praviloma več kot dvakrat tanjša.
Hvala za več kompleksna struktura nekateri predstavniki te skupine imajo povečano odpornost na protitelesa in antibiotike.
Z zapletom celične stene so se pojavile nove funkcije in možnosti. Nekatere vrste bakterij, ki živijo naprej trdih površinah, pridobil sposobnost spreminjanja oblike celice (stene "okostja" so postale plastične). To je pomembno pri drsenju in premikanju po neravninah trdnega habitata.
Periplazmatski prostor je dobil funkcijo izoliranega skladišča nekaterih hidroliznih encimov, potrebnih za življenje celice, hkrati pa sposobni razgraditi polimerne molekule same bakterije. Zaradi hidrolize polimernih molekul, ki vstopajo od zunaj živa celica razširi nabor snovi, primernih za prehrano, hkrati pa notranja membrana, ki je neprepustna za encime, preprečuje "samoprebavo" bakterijske celice.
Struktura zunanje membrane vključuje beljakovine, ki lahko tvorijo hidrofilne pore, skozi katere lahko nekatere makromolekule (sladkorji in aminokisline), potrebne za bakterijsko prehrano, preidejo v celico.
Ta razred bakterij vključuje številne bakterije, ki se lahko spreminjajo kemična sestavaživljenjski prostor (bakterije ocetne in alkoholne fermentacije); simbiotski - Escherichia coli (E coli), številni patogeni (povzročitelji bolezni), kot je meningokok (Neisseria meningitidis); legionela (Legionella pneumophila), in slavna Helicobacter pylori izzivalen peptični ulkus in visoka kislinska odpornost.

Človeška uporaba strukturnih značilnosti bakterij
Prvi mikroorganizmi, ki jih je človek uporabil za svoje potrebe, so bile bakterije mlečnokislinskega in alkoholnega vrenja. Prav oni so nam kuhali in še kuhajo sir, kruh in vino. Poleg tega so ljudje začeli uporabljati produkt svojega dela že dolgo pred odkritjem bakterij in začetkom proučevanja strukture in sestave njihovih presnovnih produktov.
Trenutno so na voljo ne samo biološke metode za čiščenje odpadnih in fekalnih voda komunalna podjetja ampak tudi lastnikom zasebnih hiš. Mirujoče kulture so del sodobna zdravila za greznice in lokalno kanalizacijo. Kmetije pogosto uporabljena kislinsko odporna za hitro predelavo komposta in živalskih odpadkov v gnojila in za številne druge namene.
Prav tako je veliko gensko spremenjenih bakterij vključenih v proizvodnjo zdravil, sintezo novih vrst polimerov in drugih materialov z edinstvenimi lastnostmi. kemijske lastnosti in kompleksno strukturo.
Kemične sestavine celične stene običajno delimo na bazične, ovojne in odložene na njeni površini (adkrustne). Na glavno kemične komponente vključujejo celulozo, hemicelulozo, pektin, beljakovine. Komponenti, ki obdajata celično steno, sta lignin in suberin. Kutin in vosek se lahko odlagata na površini celične stene. Poleg tega celične stene vsebujejo silikate in kalcijeve karbonate.
Najpogostejše sestavine obnovljivih rastlinskih surovin so celuloza, lignin in hemiceluloze (polioze), ki jih je 40 oz. 30 in 26 % tega totalna teža. Celuloza je ojačitveni material celičnih sten, ki tvori urejene nitaste strukture (fibrile). Lignin in hemiceluloze so amorfni biopolimeri z mrežasto ali razvejano strukturo in imajo vlogo veziva (polnila).
Struktura teh snovi, njihova medmolekularna interakcija določajo lastnosti rastlinskih substratov kot celote in njihovo obnašanje med predelavo.
Celuloza
Celuloza je najpogostejša snov na Zemlji, je skelet celične stene rastlin. Struktura celuloze bombažnih vlaken in vlaken, ki jih sintetizirajo nekatere bakterije, alge in mehkužci, je najbolje raziskana, saj je v njih celuloza v najčistejši obliki. V lesu se pojavlja le v obliki kompozita z drugimi biopolimeri, od koder ga je težko ekstrahirati brez delnega uničenja.
Kemično je celuloza polisaharid, katerega makromolekule so zgrajene iz ostankov ß-D-glukoze in predstavljajo linearno verigo anhidroglukopiranoznih enot, povezanih z 1,4-glukozidnimi vezmi (slika 9).
Slika 9. Kemična struktura celuloza
Je stereopravilen polimer, v verigi katerega ostanek celobioze služi kot stereoponavljajoča se povezava. Celotna formula celuloze ima obliko - (C 6 H 10 O 5) n ali [C 6 H 7 O 2 (OH) 3] n. Monomerna enota celuloze vsebuje tri alkoholne hidroksilne skupine, od katerih je ena primarna - CH 2 OH, dve pri C 2 in C 3 pa sta sekundarni - CHOH. Končni členi se razlikujejo od preostalih členov verige. Ena končna povezava (pri C 4) ima dodaten, prosti sekundarni alkoholni hidroksil. Končna povezava (pri C 1) vsebuje prosti glikozidni (semiacetal) hidroksil in lahko obstaja v dveh tavtomernih oblikah - ciklični (semiacetal) in odprti (aldehid). Končna aldehidna skupina daje celulozi redukcijsko (obnovitveno) sposobnost. Sposobnost celuloze, da se podvrže redukcijski reakciji, lahko uporabimo za karakterizacijo dolžine njenih verig, ki je značilna za te vrste rastline.
V ostanku glukoze je običajno razlikovati med hidrofilnimi in hidrofobnimi ravninami, odvisno od tega, koliko OH in CH skupin mu ustreza. To je pomembno pri proučevanju interakcije ostanka glukoze z ostanki aromatskih aminokislin med encimsko cepitvijo celulozne molekule v aktivnem središču encima.
Zaradi dejstva, da se hidroksilne skupine pri prvem in četrtem atomu glukoze razlikujejo glede na ravnino obroča, se izkaže, da so sosednji ostanki glukoze v polimerni verigi zasukani za 180 0 glede na drugega. Polimerne verige so stabilizirane z vodikovimi vezmi, ki nastanejo med ostanki glukoze med interakcijo hidroksilnih skupin pri 6,2,3 atomih ogljika in kisika v obroču. Linearne polimerne verige so med seboj povezane z medmolekularnimi vodikovimi vezmi, ki nastanejo med kisikovimi atomi ene verige in hidroksilnimi skupinami pri C 6 glukoze druge verige (slika 10). Kot rezultat takšnih interakcij nastanejo močna vlakna. Fleksibilnost celulozne molekule je določena z možnostjo rotacije anhidroglukoznih ostankov v celobiozni povezavi glede na glikozidno vez. Stabilnost linearne konformacije celuloze je določena zlasti z minimalnim odbojem C1-H in C4-H.
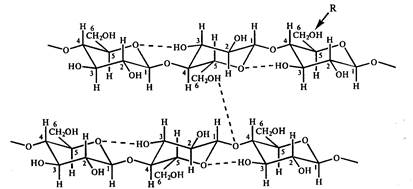
riž. 10. Shematski prikaz sistema vodikovih vezi med glukoznimi ostanki v celulozi. R je središče možne kemične modifikacije celuloze. Torej, v metilcelulozi, acetilcelulozi in karboksimetilcelulozi R \u003d CH 3, COCH 3, CH 2 COOHa
Molekula glukoze je predstavljena v konformaciji stola, v kateri atomi C2, C3, C5 in O5 ležijo v isti ravnini, atoma C1 in C4 pa ležita na nasprotnih straneh le-te. Glukozni ostanek lahko zavzame tudi energijsko manj ugodne konformacije, ki so glede na stopnjo stabilnosti razvrščene v naslednjem vrstnem redu: poševna konformacija, konformacija čolna (ali kopeli) in najmanj ugodna konformacija polstola, v kateri je 5 atomov. 6-členskega piranoznega obroča padejo naenkrat v eno ravnino. Sposobnost anhidroglukozne enote, da v molekuli celuloze poleg najbolj stabilnih sprejme tudi manj stabilne konformacije (na primer konformacijo čolna ali konformacijo polstola). pomembnost v nasprotju z urejeno supramolekularno strukturo celuloze, pa tudi pri encimski hidrolizi glikozidnih vezi v njeni molekuli.
Osnovna vlakna (micele) celuloze so sestavljena iz 100-200 polimernih verig, povezanih z vodikovimi mostovi. Ta vlakna v celični steni so med seboj vzporedna in združena tako, da nastanejo snopi ovalne oblike. – mikrofibrile . Približno 10 micel tvori mikrofibrilo s premerom 0,025 nm (slika 11).
Vsak snop ima osrednji kristalni del – jedro, v katerem so verige zložene tako tesno, da vanje z veliko težavo prodrejo tudi molekule vode. Okoli jedra je parakristalni korteks z manj urejeno strukturo. Te manj urejene regije, imenovane tudi amorfne regije, običajno obsegajo približno 15 % mikrostrukture celuloze. Mikrofibrile se lahko združijo v posamezne skupine ali plasti – makrofibrile - debeline 0,4-0,5 mikronov, vidne v svetlobnem mikroskopu. Premer in stopnja kristaliničnosti mikrofibril se zelo razlikujeta glede na vrsto rastline in fiziološko stanje celice.
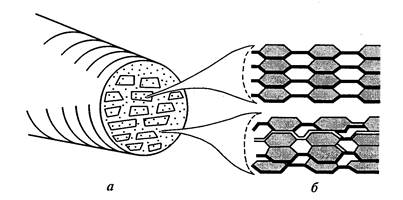
Slika 11. Model zgradbe mikrofibrila: a) prerez mikrofibrila z miceli in medmicelarnimi prostori; b) postavitev celuloze
Za lesno celulozo z indeksom urejenosti 50-70 % prečne dimenzije kristalita ne presegajo 4x6 nm. Glede na elektronsko mikroskopijo so lahko povprečne dimenzije v premeru 1,5-2x2,5-3,5 nm. Od 40 do 80% molekul s takšno velikostjo se nahaja na površini kristalita, kjer je možna njihova večja hidracija, kršitev daljnosežnega reda v strukturi vodikovih vezi in tudi močnejši vpliv kemičnih vplivov.
Naravna celuloza je polidisperzna, število glukoznih ostankov v celulozni molekuli in posledično dolžina samih molekul se lahko zelo razlikujeta. celične stene razne rastline. Torej, za celulozo iz bombažnih las je stopnja polimerizacije primarne celične stene 2-6 tisoč, sekundarne - 13-14 tisoč, za celulozo Acetobacter xylinum - 2-6 tisoč, za les - 8-10 tisoč. Največja eksperimentalno ugotovljena molekulska masa lanene celuloze je 6 000 000. Ker je stopnja polimerizacije povprečni indikator, je najbolj natančna značilnost krivulja porazdelitve celuloznih molekul po dolžini (porazdelitev molekulske mase). Vendar pa je to značilnost težko doseči, zato se uporabljajo povprečne značilnosti: številčna povprečna (DP n) in masna povprečna (DP w) stopnja polimerizacije. Številčna povprečna stopnja polimerizacije je enaka skupnemu številu anhidroglukoznih ostankov v vzorcu celuloze, deljeno s skupno število polimerne molekule. Da bi ga določili, redukcijske konce titriramo v vzorcu polisaharida. Masno povprečno stopnjo polimerizacije celuloze določimo z raztapljanjem vzorca celuloze v alkalnem kompleksu bakra, cinka, kobalta ali kadmija z etilendiaminom ali v kompleksu LiCl z N,N'-dimetilacetamidom. razmerje DP w/DP n enaka enoti samo za monodisperzne polimere.
Za lesno celulozo smreke, bora, breze in trepetlike je utežna povprečna stopnja polimerizacije 12000, medtem ko dolžina takšne molekule v razširjenem stanju doseže skoraj 6 mikronov, kar je 6000-krat več od njenega premera. Tako dolga in pravilno zgrajena molekula je zložena na določen način. Metoda zlaganja določa supramolekularno strukturo biopolimera, ki temelji na intra- in medmolekularnih vodikovih vezeh, ki jih tvorijo celobiozne enote. Preučevanje metode zlaganja in vrst vodikovih vezi v makromolekuli celuloze poteka s pomočjo konformacijske analize, difrakcije (malokotno sipanje rentgenskih žarkov, sipanje elektronov), IR spektroskopije in NMR. Kljub pomembnemu napredku na tem področju vprašanje, kako zložiti celulozne molekule, ni dokončno rešeno.
Celuloza je netopna v vodi in organskih topilih, kemično inertna, mehansko zelo močna. Voda med tesno razmaknjenimi, vendar ne neposredno medsebojno delujočimi površinami kristalitov je strukturirana. Odstranitev hidratacijske vode, na primer kot posledica sušenja, spodbuja konvergenco prvotno nekontaktnih celuloznih molekul in nastanek različnih vrst vodikovih vezi med njimi, ki se med rehidracijo ne uničijo. Raziskovalci pripisujejo povečanje stopnje kristaliničnosti med vlaženjem prodiranju vode iz manj urejenih v bolj urejena področja. Posledično opazimo zmanjšanje absolutnih velikosti kristalitov in povečanje stopnje splošne urejenosti. Domneva se, da voda v tem primeru deluje po principu »molekularnega mazanja«, ki neurejenim fragmentom molekul olajša dokončanje urejene strukture s kooperativno interakcijo s kristalnim »semenom«. Zamenjava polarnih skupin celuloze, na primer s karboksimetilom, vodi do kršitve molekularne pravilnosti, spodbuja tvorbo številnih vodikovih vezi z vodo in naredi celulozo topno.
Odpornost celuloze proti cepljenju, tako v naravnih kot laboratorijskih pogojih, ni posledica značilnosti b-1,4-glukozidne vezi, ampak kristalne strukture celuloze in posebnega "pakiranja" njenih molekul v biološke strukture.
Celulolitični encimi ne morejo prodreti v notranje strukture celuloznih vlaken. Delujejo na njihovi površini. Število glikozidnih vezi, ki so na voljo za delovanje encimov, je v veliki meri odvisno od stopnje nabrekanja celuloze. Povečanje stopnje nabrekanja je mogoče doseči s predhodno obdelavo - parjenje, mletje, ultrazvočna obdelava itd. Nabrekanje celotnega vlakna povečajo mineralne kisline in alkalije v visoke koncentracije, saj lahko pretrgajo vodikove vezi in prodrejo v kristalna področja. Na amorfne dele celuloze razmeroma zlahka vplivajo kisline in alkalije.
Pod vplivom koncentriranih alkalijskih raztopin na celulozo se spremenijo njene fizikalno-kemijske in strukturne lastnosti: opazimo močno otekanje in spremeni se stopnja kristaliničnosti. Frakcija celuloze z visoko molekulsko maso, netopna v 17,5 % NaOH, se imenuje a-celuloza. Bombaž je sestavljen pretežno iz a-celuloze, medtem ko druge rastline in les vsebujejo pretežno b-celulozo, topno v koncentrirani raztopini alkalije.
Mikrofibrile so potopljene v amorfni lupinasti matriks, ki je kompleksna mešanica polimerov, med katerimi prevladujejo polisaharidi različnih molekulskih mas. Predpostavlja se, da je na površini vseh mikrofibril plast molekul ksiloglukana, sorbiranih z vodikovimi vezmi. Lignin in hemiceluloza zapolnjujeta prostor med osnovnimi celuloznimi vlakni. Tako je lignificirana snov polimerna struktura, sestavljena iz celulozne ojačitve, potopljene v lignocelulozno matrico.
Za celulozo in hemicelulozo je značilna različna kinetika tvorbe, kar očitno kaže različni kraji njihovo sintezo. Zdi se, da sinteza celuloze poteka zunaj celice na površini citoplazmatsko membrano s sodelovanjem GDF ali UDF. Donatorji glikozilnih skupin so sladkorni nukleozidni difosfati, ki nastanejo iz glukoze in saharoze. različne poti. Na primer iz nukleozid trifosfatov in sladkor-1-fosfatov s sodelovanjem pirofosforilaze. Glikozilni ostanki so pritrjeni na ostanke sladkorja na nereduciranih koncih rastoče verige. Med prvim ogljikom "dodanega" ostanka in hidroksilno skupino četrtega atoma ogljika končnega ostanka nastane glikozidna vez s sodelovanjem sintetaze.
N (nukleozid difosfat sladkor) + (sladkor) n sintetaza n (nukleozid difosfat) + (sladkor) n +1
V celični steni so našli encime za sintezo celuloze in zrnca s premerom 8 nm, ki so očitno multiencimski kompleksi, ki se nahajajo na zunanjo površino plazmalema in v matriksu stene. Celulozna sintetaza (E) v plazmalemi je povezana z aktivacijskim proteinom P, ki je sposoben vezati GTP in preiti v aktivno obliko s tvorbo kompleksa P-E-GTP.
V procesu sinteze celuloze ne nastajajo le posamezne celulozne verige, temveč tudi celulozne mikrofibrile. Takšna sinteza se verjetno izvaja s sodelovanjem številnih molekul celuloznih sintetaz, koncentriranih na ločenih mestih plazmaleme, celulozne sintetaze pa so sestavljene iz številnih podenot, od katerih je vsaka odgovorna za sintezo ene glukanske verige. Vklopljeno znotraj membrane, so bile najdene "rozete" šestih beljakovinskih globul. Rozete se izločajo v plazmalemo preko endomembranskega sistema veziklov. Na podlagi značilnosti strukturne povezave rozet, končnih kompleksov in nastalih celuloznih mikrofibril se je pojavila domneva o mehanizmu nastajanja mikrofibril v komplementarni transmembranski strukturi: rozete - terminalni kompleks - rastoči konec mikrofibrila. S koncem novo sintetiziranega mikrofibrila je povezan terminalni kompleks, rozete pa so mesto vezave prekurzorjev celuloze in izločanja »prenašalcev« ostankov glukoze.
Posamezne rozete, skupaj z ustreznimi končnimi kompleksi, so odgovorne za sintezo naključno usmerjenih mikrofibril primarne celične stene (slika 12), usmerjen niz do 175 rozet v ločenih delih plazmaleme pa določa nastanek vzporednih celic. mikrofibrile sekundarne celične stene (slika 13).
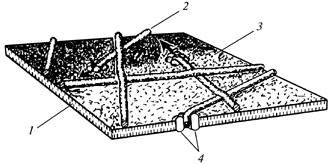
Sl. 12. Celulozna mikrofibrila med nastajanjem
primarna celična stena: 1
- lipidna dvojna plast; 2
- mikrofibril; 3
- zunanja površina plazmaleme; 4
- vtičnica
Vsaka rozeta tvori mikrofibrilo s premerom 5 nm. Razdalja med vrstami rozet v plazemski membrani ustreza razdalji med mikrofibrili v celični steni. Tanke mikrofibrile vsebujejo približno 50 poliglukanskih verig. Ti mikrofibrili tvorijo fibrile sekundarne celične stene, ki imajo lahko premer do 28 nm. Celuloza je za razliko od škroba težko presnovljiva spojina. Njegova prebavljivost je odvisna od izvora, vsebnosti v prehrani, vrste predobdelave in se v povprečju giblje od 6 do 23%. Da, prebavljivost celuloze pšenični otrobi približno 15 %.
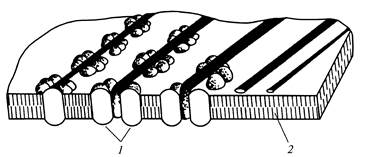
riž. 13.Mikrofibrile celuloze med nastajanjem
sekundarna celična stena: 1
- vtičnica; 2
- lipidna dvojna plast
IN prehranske vlaknine celuloza predstavlja približno eno tretjino. Njegova učinkovita razgradnja je posledica simbiotske interakcije različnih mikroorganizmov debelega črevesa in njihovih celulolitičnih encimov. IN prebavni trakt celuloza spodbuja aktivnost črevesja, povečuje njegovo peristaltiko, normalizira aktivnost črevesna mikroflora, absorbira sterole, preprečuje njihovo absorpcijo in spodbuja sproščanje holesterola. Mikrokristalna celuloza (MCC), nevlaknasta praškasta modifikacija naravne celuloze, je našla široko uporabo kot prehransko dopolnilo. Ne samo da zmanjša vsebnost kalorij v hrani, ampak je tudi njen zgoščevalec in dispergator, kar izboljša kakovost hrane.
Gram-negativne evbakterije imajo veliko bolj zapleteno strukturo celične stene kot Gram-pozitivne. Vsebuje veliko večje število makromolekul različnih kemična vrsta. Peptidoglikan tvori samo notranjo plast celične stene, ohlapno ob CPM. Za različni tipi Gramnegativne eubakterije, vsebnost tega heteropolimera se zelo razlikuje. Pri večini vrst tvori eno- ali dvoslojno strukturo, za katero so značilne zelo redke navzkrižne povezave med heteropolimernimi verigami.
Nekatere drsne bakterije (myxobacteria, flexibacteria) lahko občasno spreminjajo obliko celic v procesu premikanja vzdolž trdnega substrata, na primer z upogibanjem, kar kaže na elastičnost njihove celične stene in predvsem njene peptidoglikanske plasti. Elektronski mikroskopski pregled pa je razkril, da imajo celično steno, značilno za gramnegativne evbakterije. Najverjetnejša razlaga prožnosti celične stene teh bakterij je izjemno nizka navzkrižna povezava njene peptidoglikanske komponente.
Po Gramu negativni prokarionti imajo zunanja membrana, ki vključuje lipide (22%), beljakovine, polisaharide, lipoproteine.
Lipopolisaharidi (LPS) so heteropolimeri s kompleksno strukturo, ki imajo različne biološke aktivnosti. Lipoidni kompleks povzroča toksičnost (vnetne reakcije, vročina, endotoksinski šok), polisaharidna komponenta je odgovorna za specifičnost O-antigena. LPS inducira sintezo protiteles Jg M in se v imunologiji uporablja kot adjuvans in poliklonski aktivator celic B.
Celična stena v bakterijah opravlja predvsem oblikovne in zaščitne funkcije, zagotavlja togost, tvori kapsulo in določa sposobnost celice za adsorpcijo fagov.
№15 Filtrirane, involutivne oblike bakterij, protoplasti, sferoplasti, L-oblike.
protoplasti, sferoplasti, L-oblike so oblike bakterij brez KS
L-oblike se lahko pojavijo v naravnih pogojih v človekovem organizmu kot posledica dolgotrajnega zdravljenja z nekaterimi antibiotiki, najpogosteje s penicilinom.
Obstajajo nestabilne in stabilne L-oblike bakterij. Prvi se lahko vrnejo v prvotno obliko, ko je odpravljen vzrok, ki je povzročil njihov nastanek. Obnovijo sposobnost sinteze peptidoglikana CS. Slednji praviloma niso sposobni reverzije. L-oblike različnih bakterij igrajo bistveno vlogo v patogenezi številnih nalezljivih bolezni.
Glavne lastnosti L-oblik bakterij:
stalna transformacija iz grampol-s v grammatrits-e.
sprememba antigenskih lastnosti
zmanjšanje virulence
sposobnost dolgotrajnega vztrajanja
možnost vrnitve v prvotno obliko v primeru nepopolne izgube sineta COP.
№16 Tinktorialne lastnosti bakterij. Sofisticirane metode obarvanja. Princip barvanja bakterij po Gramu.
Tehnika barvanja po Gramu
Na razmaz položimo filtrirni papir in za 1-2 minuti prelijemo karbolno raztopino encijan vijoleta.
Odstranite papir, odcedite barvo in brez izpiranja razmaza z vodo za 1 minuto prelijte Lugolovo raztopino.
Lugolovo raztopino odlijemo in pripravek razbarvamo v 96 0 alkoholu 30 sekund.
Oprano z vodo.
Barvanje 1-2 min vodna raztopina magenta.
Načelo obkroženosti Po Gramu
Po Gramu pozitivno bakterije zadržijo encijan vijolico v kombinaciji z jodom - vijolično barvanje bakterije;
po Gramu negativno bakterije po izpostavljenosti alkoholu izgubijo barvo, se razbarvajo in se ob obdelavi s fuksinom obarvajo Rdeča barva.
Kompleksne metode Barve se uporabljajo za proučevanje celične strukture in diferenciacije mikroorganizmov. Obarvane brise mikroskopiramo v imerzijskem sistemu. V pripravek dosledno uporabljajte določena barvila, ki se razlikujejo po kemični sestavi in barvi, jedkala, alkohole, kisline itd.
tinktorialne lastnosti bakterije (lat. tinctura, iz tingo - barvam) sposobnost barvanja: dovzetnost za barvo, kislinsko-alkoholno-alkalna odpornost, barvna enakomernost, metakromatičnost, odnos do barve po Gramovi metodi.
№17 Značilnosti kemične sestave celične stene kislinsko odpornih bakterij. Princip barvanja po metodi Ziehl-Neelsena.
kislinsko odporne bakterije. Celična stena nekaterih bakterij vsebuje veliko količino lipidov in voskov, zaradi česar so odporne na naknadno razbarvanje po obarvanju s kislinami, alkalijami ali etanolom (na primer vrste Mycobacterium ali Nocardia). Takšne bakterije imenujemo kislinsko odporne in jih je težko obarvati po Gramu (čeprav kislinsko odporne bakterije veljajo za grampozitivne). Za njihovo barvanje se uporablja metoda Ziehl-Neelsena.
Za mikobakterije in nokardije je značilna zapletena zgradba celične stene. Tako kot po Gramu pozitivne bakterije temeljijo na mureinskem ogrodju, vendar je slednji povezan s polisaharidi in lipidi. Lipide predstavljajo mikolne kisline, ki dajejo celični površini hidrofobnost. Hidrofobnost po eni strani naredi celico odporno na delovanje različnih kemikalij (takšne bakterije imenujemo kislinsko odporne), po drugi strani pa zavira izmenjavo celice z okoljem in upočasnjuje njeno rast. Zato hranilnim medijem za gojenje Mycobacterium tuberculosis dodajamo površinsko aktivne snovi. Kislinska odpornost mikobakterij je pomembna diagnostična lastnost, za njeno določanje se uporablja barvanje po Ziehl-Neelsenu.
Tehnika barvanja za kislinsko odporne bakterije
po metodi Ziehl-Neelsen
Na fiksiran razmaz položimo filtrirni papir in nalijemo Ziehlov karbolni fuksin ter rahlo segrevamo na gorilniku, dokler se ne pojavijo hlapi. Operacija se ponovi 2-3 krat.
Ko se zdravilo ohladi, odstranimo filtrirni papir, odlijemo barvilo in zdravilo speremo z vodo.
Zdravilo se 2-3 krat potopi v kozarec s 5% žveplovo kislino za 1-2 sekundi.
Preparat temeljito speremo z vodo in obarvamo z alkalnim metilen modrim 3-5 minut.
Oprano z vodo in posušeno.
Kislinsko odporne bakterije se z žveplovo kislino ne razbarvajo in ohranijo svojo rdečo barvo, kislinsko neodporne bakterije izgubijo barvilo in se z metilen modrim obarvajo modro.
№18 Vključki bakterij, njihova sestava in biološka vloga. V citoplazmi so različni vključki v obliki glikogenskih granul, polisaharidov, beta-hidroksimaslene kisline in polifosfatov (volutin). So rezervne snovi za prehrano in energijske potrebe bakterij. Volyutin ima afiniteto do osnovnih barvil in ga zlahka zaznamo z uporabo posebne metode obarvanost (npr. po Neisserju) v obliki metakromatskih granul. Značilna razporeditev volutinskih zrnc se pri difteričnem bacilu kaže v obliki intenzivno obarvanih polov celice. (po Neisserju je obarvan temno modro)
№19 Spore bakterij, njihova struktura Nekatere bakterije ob koncu obdobja aktivne rasti lahko tvorijo spore. Pred tem je osiromašenje okolja s hranili, sprememba njegovega pH in kopičenje strupenih presnovnih produktov. Praviloma ena bakterijska celica tvori eno sporo - lokalizacija spor je različna (centralna, terminalna, subterminalna)
Če velikost trosov ne presega prečne velikosti paličaste bakterije, se slednja imenuje bacil (povzročitelj). antraks). Pri večjem premeru trosov so bakterije vretenaste oblike in se imenujejo klostridije (povzročitelji anaerobne okužbe). Tetanus Clostridia ima okrogle spore in spominja na bobnične palčke. Za Clostridia botulinum so značilne velike ovalne spore, ki jim dajejo videz teniškega loparja.
Glede na kemično sestavo je razlika med sporami in vegetativnimi celicami le v količinski vsebnosti kemičnih spojin. Spore vsebujejo manj vode in več lipidov.
Nastanek spor je povezan s stiskanjem in izolacijo določenega območja citoplazme vegetativne celice, čemur sledi tvorba okroglega ali ovalnega telesa znotraj bakterije, prekritega z gosto večplastno membrano, ki je nasičena z velika količina lipidov, kalcija in dipikolinske kisline (slika 12).
Ko so v telesu ljudi in živali, spore kalijo v vegetativne celice. Proces kalitve spor vključuje tri stopnje: aktivacijo, začetni fazi in stopnje rasti. Aktivatorji, ki motijo stanje mirovanja, so povišana temperatura, kisla reakcija okolja, mehanske poškodbe in drugi Spora začne absorbirati vodo, se sprosti iz kalcijevega dipikolata in s pomočjo hidrolitičnih encimov uniči številne lastne strukturne komponente. Po uničenju zunanjih plasti se začne obdobje nastajanja vegetativne celice z aktivacijo biosinteze, ki se konča z delitvijo celice.
Metoda obarvanja spor po Ozheshku
Spore trdno držijo karbolni fuksin in so obarvane rdeče, citoplazma bakterij se razbarva s 5% žveplovo kislino in po obarvanju z metilen modrim postane modra.
№20 Kapsula in mikrokapsula bakterij Kapsula je sluzna plast celične stene bakterij, sestavljena iz polisaharidov (pnevmokok) ali polipeptidov (antraksni bacil). Mikrokapsulo (manj kot 0,2 µm debelo) lahko tvori večina bakterij, dobro definirano makrokapsulo (več kot 0,2 µm debelo) tvorijo pnevmokok, Klebsiella, antraks in nekatere druge. Pri patogenih bakterijah se kapsula tvori v makroorganizmu, na umetnih hranilnih medijih pa se običajno izgubi (z izjemo Klebsiella).
Pri ljudeh in živalih kapsula ščiti patogene bakterije od bakteriofaga, fagocitoze in humoralnih imunskih dejavnikov, določa antigensko specifičnost mikroorganizmov.
Kapsule, ki imajo konsistenco gela, slabo zadržijo barvilo, zato se za njihovo odkrivanje najpogosteje uporabljajo metode negativnega kontrasta.
Metoda odkrivanja kapsul Burri-Gins
Bakterije so obarvane rdeče, neobarvane kapsule so v kontrastu s temnim ozadjem zdravila.
№21 Bakterijske bičke Flagele delujejo kot gibalni organ, ki omogoča bakterijam, da se premikajo s hitrostjo 20-60 mikronov / s. Bakterije imajo lahko eno (monotrično) ali več flagel, ki se nahajajo po celotni površini telesa (peritrične) ali zbrane v snope (lofotrične).
Peritrična lokacija flagel je značilna za enterobakterije, povzročitelje anaerobnih okužb, tetanusa, botulizma; Vibrio cholerae je monotrik, Pseudomonas pa lofotrik. Nekatere vrste spiril imajo amfitrično razporeditev bičkov. Debelina flagele je v povprečju 10-30 nm, dolžina pa doseže 10-20 mikronov.
Osnova bička je dolg spiralni filament (fibril), ki na površini celične stene preide v odebeljeno ukrivljeno strukturo - kavelj in je pritrjen na bazalno zrnce, vgrajeno v celično steno in CPM (slika 10) .
Bazalne granule imajo premer približno 40 nm in so sestavljene iz več obročev (en par pri gram-pozitivnih bakterijah, štirje pri gram-negativnih prokariontih). Odstranitev peptidoglikanske plasti celične stene vodi do izgube sposobnosti gibanja bakterij, čeprav bički ostanejo nedotaknjeni.
Bički so skoraj v celoti sestavljeni iz beljakovine flagelin, z nekaj vsebnosti ogljikovih hidratov in RNK.
Pod mikroskopom lahko bičke vidimo šele po posebnih metodah jedkanja in impregnacije s srebrovimi in živosrebrovimi solmi, ki jim sledi barvanje z metilen modrim (metoda po Lefflerju). Upoštevati je treba, da so flagele zelo občutljive na različne mehanske vplive. Prisotnost bičkov lahko posredno ocenimo po smeri gibanja v "viseči" in "zdrobljeni" kapljici v temnopoljskih in faznokontrastnih mikroskopih ali pri svetlopoljski mikroskopiji s spuščenim kondenzorjem in delno pokrito diafragmo mikroskopa. pokrito.
Barvanje bičkov po Lefflerjevi metodi
Pod mikroskopom končan izdelek flagele so vidne kot tanke nitaste strukture.
#22 PiliPovršina enterobakterij in več drugih mikroorganizmov je prekrita veliko število(od 10 do nekaj tisoč) resice - nitaste tvorbe beljakovinske narave. Tako kot flagele so zgrajene iz ene vrste beljakovin – pilina, katerega podenote so v notranjosti organizirane v obliki votle niti in izhajajo iz CPM. So krajši in tanjši od bičkov, njihova širina je 10–12 nm, njihova dolžina pa do 12 μm.
Resice so polifunkcionalne: zagotavljajo prenosljiv prenos genov (konjugacijo), so fagni receptorji, organ za pritrditev bakterij na hranilni substrat (adhezija) in sodelujejo pri transportu metabolitov.
Streptokoki imajo zunanjo plast beljakovinskih dlak (fimbrije), ki jih imenujemo protein M (M-protein). Ta veverica se igra pomembno vlogo v procesih interakcije bakterij z makroorganizmom.
№23 Aktinomicete
Predstavljajo svojevrstno skupino bakterij, ki izgledajo kot majhne ali dolge nerazcepljene razvejane nitke. Kopičenje hif se imenuje micelij. Podobnost z glivami je izključno zunanja, saj imajo aktinomicete prokariontski tip celic s celično steno, ki ne vsebuje hitina in celuloze. Aktinomicete so gram-pozitivne, številne oblike so kislinsko odporne, nekatere aktinomicete imajo kapsulo okoli filamentov.
Substratni micelij nastane kot posledica vraščanja micelija v hranilni medij in zraka, ki raste na površini gojišča (slika 19). V prizadetih tkivih (tkivna oblika) lahko aktinomicete tvorijo drusenske granule iz gosto prepletenih niti v obliki žarkov, ki segajo od središča in se končajo z zgoščevanjem v obliki bučke.
Aktinomicete se množijo aseksualno, ki tvorijo konidije ali sporofore s sporangiji na koncih zračnega micelija. Nosilci spor so lahko ravni, valoviti, spiralni. Spore - ovalne, okrogle, cilindrične, z gladko površino ali konicami, včasih mobilne zaradi flagel (zoospore). Spore se uporabljajo za razmnoževanje aktinomicet, niso toplotno odporne, lahko pa prenesejo sušenje. Poleg tega je možno brstenje in drobljenje micelija v paličaste ali kokalne oblike.
Aktinomicete so v naravi zelo razširjene, živijo v vodi, zemlji, bogati s humusom. Sodelujejo v kroženju snovi v naravi. Ločene vrste aktinomicete se uporabljajo kot proizvajalci antibiotikov, vitaminov, lipidov, proteaz, aminokislin, steroidov.
Aktinomicete spadajo v red Actinomycetales, ki vključuje družine: Actinomycetaceae, Nocardiaceae, Streptomycetaceae, Mycobacteriaceae.
Za človeka patogene vrste najdemo med predstavniki družin Actinomycetaceae in Nocardiaceae. Prvi izgledajo kot dolge ali kratke razvejane palice, ki ne tvorijo zračnega micelija. So povzročitelji humane aktinomikoze in tvorijo druze v prizadetih tkivih.
Predstavniki družine Nocardiaceae spominjajo na mikobakterije, imajo nitaste celice in tvorijo zračni in substratni micelij na hranilnih medijih. Hife so razdrobljene na kokoidne in paličaste celice. Patogena nokardija povzroča nokardiozo.
Raziskovalne metode. obarvano po Gramu in Ziehl-Neelsenu. Druzo odstranimo iz patološkega materiala z zanko, damo v kapljico vode na predmetnem stekelcu, rahlo pritisnemo navzdol s pokrovnim stekelcem, nato pa pod steklo vnesemo kapljico alkalne raztopine metilen modrega in mikroskopiramo, fazno kontrastno je lahko uporabljen.
№24 Spirohete
spiralno zavite bakterije z aktivno gibljivostjo. Dimenzije spirohet se razlikujejo po debelini od 0,1-0,3 mikronov, po dolžini od 7-500 mikronov. Različni gibi - od vijačnih do upogibnih. Elektronski mikroskopski pregled je pri spirohetah omogočil razlikovanje protoplazmatskega valja (celičnega telesa), aksialne (podporne) niti in troslojne zunanje lupine. Aksialni filament se nahaja v periplazemskem prostoru med zunanjo lupino in protoplazmatskim valjem in je sestavljen iz posameznih fibril (endoflagel), katerih število se razlikuje pri različnih vrstah: v treponemi in leptospiri - 3-4; v boreliji - do 30. Vsaka od fibril (endoflagela) je pritrjena na območju pritrdilnih diskov na koncih protoplazemskega valja in se razteza do nasprotnega konca, se ovija okoli njega in se prosto konča. Kemična sestava fibril je podobna bičkom (slika 20).
Protoplazmatski valj vsebuje: nukleoid, ribosome, mezosome, vključke. Zunanja lupina (celična stena) vsebuje tanko plast peptidoglikana, je elastična in nima togosti. Te bakterije ne tvorijo endospor, kapsul in eksflagelatov, so gramnegativne, v razmazu so razporejene naključno.
Spirohete pripadajo redu Spirochaetales, družini Spirochaetaceae, ki vključuje tri rodove:
Borrelia - ima 3-10 neenakomernih poševnih kodrov, koniči so koničasti, dolgi 10-30 mikronov. Gibanje je sunkovito, po Romanovsky-Giemsa so pobarvane v modri barvi vijolična(predstavnik Borreliarecurrentis - povzroča epidemično povratno mrzlico; Borreliaburgdorferi - povzroča boreliozo).
Treponema - ima 8-14 tesno zvitih kodrov enake amplitude, dolžine 5-15 mikronov. Gibanje je gladko, počasno z vrtenjem okoli vzdolžne osi, po Romanovsky-Giemsa so naslikane bledo roza barva(predstavnik Treponemapallidum - povzročitelja sifilisa).
Leptospira - ima do dva ducata majhnih pogostih kodrov, ki se končajo s kavljem z gumbastim zgoščevanjem, dolgim 5-15 mikronov. Gibanje je zelo aktivno, translacijsko gibanje naprej, upogibanje in vrtenje okoli osi. Po Romanovsky-Giemsa se rahlo obarvajo v rožnato-lila barvi (predstavnik Leptospirainterrogans je povzročitelj leptospiroze).
Raziskovalne metode. V živem stanju spirohete proučujemo s faznokontrastnim mikroskopom in mikroskopom s temnim poljem, pri čemer opazujemo aktivno značilno gibanje spirohete, značilnosti njihove oblike.
Pripravki so pripravljeni po Burriju (lahki nagubani filamenti spirohet postanejo vidni na temnem ozadju pripravka), obarvani po Romanovsky-Giemsi po metodi Morozov.
#25 Rikecije
razred" Alfaproteobakterije"
naročilo Rickettsiales
družina Rickettsiaceae
Rod Rickettsia
družina Ehrlichiaceae
Rikecije so raznolike po obliki in jih delimo na naslednje vrste:
kokoidni enozrnati (do 0,5 mikronov);
paličasti dvozrnati (1-1,5 mikronov);
bacilarna tri-štiri-zrnata (3-4 mikrona);
filiformno večzrnato (10-40 mikronov).
Zrna (nukleoproteine) odkrijemo z barvanjem po Romanovsky-Giemsi. Vse oblike so vzajemne. Strukturno imajo vse sestavine bakterijske celice: celično steno, lipoidno kapsulo, citoplazmo, nukleoid, ribosome, pili. Rikecije vsebujejo tako DNK kot RNK, imajo visoka vsebnost fosfolipidi, nizka vsebnost ogljikovih hidratov.
V večini primerov (razen vrste Rochalimaeaguintana) rikecije ne rastejo na umetnih hranilnih gojiščih. Življenjski cikel rikecije je odvisen od vitalne aktivnosti gostiteljske celice in je sestavljen iz dveh stopenj: vegetativnega in mirovanja (elementarna telesa). Rikecije, ki so v vegetativni fazi (slika 23), se aktivno razmnožujejo z binarno cepitvijo in imajo aktivno mobilnost, očitno zaradi bičkovih struktur. Rikecije v fazi mirovanja (elementarna telesa) so sferične in neaktivne.
Raziskovalne metode. Rikecije so dobro obarvane po Romanovsky-Giemsa v lila barvi, po Morozovu (po metodi srebrovanja) v črni barvi. Za razlikovanje rikecij se uporablja metoda obarvanja, ki jo je predlagal P.F. Zdrodovski:
Rickettsia obarva rubinasto rdeče in se zlahka zazna na ozadju modre citoplazme in modrega celičnega jedra.
№26 Klamidija
razred Klamidije
naročilo Chlamydiales
Familia Chlamydiaceae
Rod: klamidija
1). Elementarna telesca so majhne (0,2-0,5 μm) sferične strukture z elektronsko gostoto, brez presnovne aktivnosti, s kompaktnim nukleoidom in togo celično steno, ki se filtrirajo skozi bakterijske filtre. So kužni začetek klamidij in zagotavljajo njihovo preživetje v zunajceličnem okolju ter okužbo novih celic.
2). Retikularna telesa so večja (0,8-1,5 mikronov), sferične tvorbe z mrežasto strukturo s tanko celično steno in fibrilarnim nukleoidom. Rastejo iz osnovnih teles znotraj celic, so brez nalezljivosti in z delitvijo zagotavljajo razmnoževanje klamidije. Od tod drugo, zgodovinsko prvo ime retikularnih teles - "začetno telo". Retikularna telesca so vegetativna oblika klamidije.
3). Vmesna telesca so vmesna stopnja med osnovnimi in retikularnimi telesci.
Življenjski cikel klamidije se začne z dejstvom, da gostiteljska celica fagocitira osnovna telesa, nato pa se v nekaj urah reorganizirajo, povečajo in spremenijo v retikularne oblike, ki se razmnožujejo s prečno delitvijo. Življenjski cikel se konča, ko se nastajajoče vmesne oblike stisnejo, zmanjšajo in spremenijo v osnovna telesa. Pri razmnoževanju v citoplazemskih vakuolah klamidija tvori mikrokolonije (vključke), obdane z membrano. V sestavi mikrokolonij najdemo vse tri stopnje razvoja klamidije. Po pretrganju stene vakuole (mehurčkov) in membrane gostiteljske celice se novonastale klamidije sprostijo in osnovna telesca, ki okužijo druge celice, ponovijo cikel razvoja. V optimalnih pogojih rasti v evkariontskih celicah življenski krog klamidije je 17-40 ur (slika 24).
Posebnost klamidij se kaže tudi v zgradbi njihove celične stene. Je brez peptidoglikana in je dvoslojna membrana, katere togost določajo peptidi, zamreženi z disulfidnimi mostovi. Sicer pa klamidije spominjajo na gramnegativne bakterije, saj vsebujejo glikolipide, podobne lipopolisaharidom.
Red Chlamydiales vključuje eno družino Chlamydiaceae z enim samim rodom Chlamydia. Za človeka so patogene vrste C.trachomatis, C.psittaci, C.pneumoniae. Klamidija povzroča okužbe oči, dihal in urogenitalnega sistema in se združiti pod pogosto ime"klamidija".
Raziskovalne metode. Za mikroskopsko odkrivanje inkluzijskih teles (mikrokolonije) klamidije v okuženih celicah (tkivih) se uporabljajo različne metode barvanja: Romanovsky-Giemsa, Macchiavello in druge. Pri barvanju po Romanovsky-Giemsi pridobijo modro ali vijolično barvo. Poleg tega so klamidije jasno vidne v neobarvanem stanju pri mikroskopiranju mokrih preparatov pod steklom z uporabo faznokontrastnega optičnega sistema. IN Zadnje čase najpogosteje uporabljena direktna imunofluorescenčna reakcija, barvanje z akridinsko oranžno barvo.
№27 Mikoplazme razred Mollicutes
naročilo Mycoplasmatales
družina Mycoplasmataceae
Rod Mycoplasma
Rod Ureaplasma
Za človeka je patogenih pet vrst: M . pneumoniae , M . hominis , M . genitalij , M . incognitus in U . urealyticum .
mikoplazme- najmanjši prokarioti (125-150 nm), ki se lahko samostojno razmnožujejo. Menijo, da so mikoplazme najbližji potomci prvotnih prokariontskih celic. Genom mikoplazem je za celico minimalen, petkrat manjši od genoma coli in je 0,45 MD. Glavna značilnost mikoplazme je odsotnost celične stene. Obdaja jih kapsula podobna plast, pod katero je le tanka troslojna membrana debeline 7,5-10 nm, ki vsebuje precejšnjo količino holesterola. Posledično so mikoplazme izolirane v poseben oddelek Tenericutes, razred Mollicutes ("nežna koža"), red Mycoplasmatales.
Zaradi odsotnosti celične stene so mikoplazme (slika 21) osmotsko občutljive in imajo različne oblike:
a) majhne sferične ali jajčaste celice velikosti 0,2 µm (elementarna telesca), ki se filtrirajo skozi bakterijske filtre;
b) večje sferične, velikosti do 1,5 mikrona;
c) nitaste, razvejane celice velikosti do 150 mikronov.
Mikoplazme ne tvorijo spor, flagel, nekatere vrste imajo drsno mobilnost.
Razmnožujejo se z binarno cepitvijo sferičnih in nitastih celic, brstenjem in sproščanjem številnih elementarnih teles, oblikovanih v filamentih.
Kar zadeva energijo, jo mikoplazme pridobivajo na običajen način za fakultativne anaerobe, s fermentacijo ogljikovih hidratov ali aminokislin. Zaradi majhnega genoma mikoplazme imajo omejene biosintetske sposobnosti, zato jih je treba gojiti na hranilnih gojiščih, obogatenih z lipidi, beljakovinami, prekurzorji nukleinskih kislin. Rastejo počasi, kolonije z gostim središčem rastejo v gojišče in spominjajo na "pečeno jajce" (temna sredina in svetlejši odprti rob). Velikost kolonij je majhna, ne presega 600 mikronov.
V človeški patologiji ima največjo vlogo več predstavnikov rodu Mycoplasma: M.pneumoniae, M.hominis, M.anthritidis in edina vrsta rodu Ureaplasma–U.urealyticum (tako imenovana zaradi aktivnosti ureaze). Patogene mikoplazme povzročajo bolezni (mikoplazmoze) dihal, urogenitalnega trakta in sklepov z različnimi kliničnimi manifestacijami. Pri zdravljenju teh bolezni je treba upoštevati, da mikoplazme niso občutljive na beta-laktamske antibiotike in druga zdravila, ki zavirajo sintezo celične stene (zaradi njene odsotnosti v patogenu).
Raziskovalne metode. V svetlobnem mikroskopu najdemo samo največje oblike mikoplazm. V živem stanju jih proučujemo s temnopoljskim in faznokontrastnim mikroskopom, ultrastrukturne komponente zaznavamo z elektronsko mikroskopijo.
№28 Gobe
Razredi (7): · Basidiomycetes(gobe s klobukom). · Zigomicete (rod Mucor - mukormikoza ljudi in živali) Askomicete (vrečarji, rodovi Aspergillus , Penicillium , kvasovke) Deuteromycetes - nepopolne glive, se spolno ne razmnožujejo ( Candida )
Glive so velika in uspešno razvijajoča se skupina organizmov, ki vključuje približno 80.000 identificiranih vrst. Njihove velikosti segajo od enoceličnih kvasovk do velikih ponirkov, puhlic in rogov. Gobe zasedajo različne habitate – tako v vodi kot na kopnem. Poleg tega so pomembne v povezavi z vlogo, ki jo imajo v biosferi, in v povezavi z dejstvom, da jih človek uporablja v medicini in gospodarstvu.
Raziskovalne metode. Za mikroskopsko preiskavo pripravimo tako nativne (neobarvane) kot obarvane preparate.
Študija neobarvanih preparatov
Da bi jasneje razlikovali elemente glive, se pripravek razjasni, za to se patološki material (skorje, koščki nohtov, lasje, ostružki iz sluznice, vsebina granulomatoznih žarišč) položi na urno steklo ali petrijevko, kamor vlijemo 10-15% raztopino jedkega natrija ali kalija in postavimo v termostat pri 37 0 C za 20-30 minut. Nato material odstranimo in ga damo v kapljico 50 % raztopine glicerola na predmetno stekelce in pokrijemo s pokrovnim stekelcem, mikroskopsko v faznokontrastnem ali svetlobnem mikroskopu. Lahko uporabimo tudi drugo metodo: kapljico glicerina z dodatkom 10% jedkega kalija nanesemo na patološki material in ga pregledamo po 4-5 minutah, pokrijemo s pokrovnim stekelcem.
Gnoj iz abscesov, vsebine razjed, sputuma razredčimo s fiziološko raztopino ali vodno-alkoholom (1: 1) ali 50% vodno raztopino glicerola, pripravimo "zdrobljeno kapljico" in pregledamo pri povečavi x 200, x 400 s faznim kontrastom.
Študija obarvanih preparatov
Iz gnoja, krvi, cerebrospinalne tekočine, sedimenta bronhialnih izpiranj in urina pripravimo tanke brise, ki jih fiksiramo v mešanici Nikiforova, Carnoya, alkohola in formola, posušimo in obarvamo:
Barvanje laktofuksin ki vsebuje kisli fuksin - 0,1 g, mlečno kislino - 100 ml. Obarvano 3-5 minut. Ozadje zdravila je rožnato, micelij je opalescenten v modri barvi. Gobe se dobro obarvajo z mukormikozo in aspergilozo.
№29 Kvasovke in kvasovkam podobne celice kvasovk imajo zaobljeno, pialno ali podolgovato obliko, velikosti 8-10 mikronov, z dvokrožno membrano. V citoplazmi opazimo vključke v obliki glikogenskih granul, volutina in lipidov. Razmnoževanje poteka z brstenjem in askosporami. Kvasovkam podobne kvasovke so podobne pravim kvasovkam, razlika je v odsotnosti askospor in sposobnosti tvorbe psevdomicelija. Z nastankom psevdomicelija se celice podaljšajo in se dotikajo z ozko bazo. Povzročajo kandidozo, ki se razvije pri bolnih ljudeh z močnim zmanjšanjem odpornosti telesa in dolgotrajno uporabo antibiotikov. Metode za preučevanje kvasovk
Za sodobno obdobje proučevanja biološke raznovrstnosti je značilen intenziven razvoj filogenetske sistematike, ki je usmerjena v rekonstrukcijo specifičnih poti zgodovinskega razvoja organizmov. V mikrobiologiji je filogenetska sistematika dobila močan zagon za razvoj šele ob koncu 20. stoletja. v povezavi s primerjalno študijo ohranjenih nukleotidnih zaporedij v rRNA. Pri kvasovkah ta taksonomija temelji predvsem na preučevanju dveh regij rDNA z dolžino približno 600 nukleotidnih parov: domena D1/D2 na 5' koncu gena, ki kodira 26S rRNA, in interna transkribirana distančna regija (ITS) , ki vključuje gen 5.8S rRNA. Menijo, da so zaradi konzervativnosti teh regij razlike med njimi neposredno sorazmerne s filogenetsko razdaljo, stopnjo evolucijske povezanosti. Sekvenciranje nekleotidnih zaporedij rDNA se je izkazalo za močno orodje za izdelavo filogenetske klasifikacije kvasovk in določanje njihovega mesta v splošnem sistemu gliv.
Do danes so bila nukleotidna zaporedja rRNA dešifrirana in postavljena v računalniške podatkovne banke, ki so na voljo na internetu, v predstavnikih vseh znane vrste kvas. To omogoča gradnjo filogenetskih dreves, ki odražajo razvoj njihovih ribosomskih genov. Izkazalo se je, da razvrščanje kvasovk na podlagi podobnosti nukleotidnih zaporedij rRNA v mnogih primerih ne sovpada z razvrščanjem po fenotipskih lastnostih. Številne tradicionalne lastnosti, ki se uporabljajo pri klasifikaciji kvasovk, kot so značilnosti vegetativnega razmnoževanja, oblika askospor, fermentabilnost in asimilacija sladkorja, se štejejo za nezanesljive in neprimerne za določanje filogenetskih odnosov. Sekvenciranje rRNA (rDNA) se zdaj šteje za bistveno pri opisovanju novih vrst kvasovk.
Posebej močan vpliv na preučevanje kvasovk, kot tudi večine drugih skupin mikroorganizmov, je imel hiter razvoj ob koncu 20. stoletja. molekularna biologija. V sodobni taksonomiji kvasovk se široko uporabljajo metode genske sistematike, ki temeljijo na neposredni primerjavi genomov in sekvenciranju nukleotidnih zaporedij. Uporaba poenotenih molekularno bioloških metod je omogočila še zbližanje pristopov k taksonomiji kvasovk in nitastih gliv, ugotavljanje povezav med kvasovkami anamorfi in nitastimi teleomorfi ter razvoj novih kriterijev za oblikovanje enotnega filogenetskega sistema za celotno kraljestvo Mycota. . Hkrati je novo znanje rodilo novo znanstveni problemi zlasti problem korelacije med najnovejšimi molekularnimi metodami in tradicionalnimi metodami, ki temeljijo na morfoloških in fizioloških pristopih k preučevanju kvasovk. Genom Saccharomyces cerevisiae je skoraj v celoti dešifriran, kar odpira velike možnosti za genomiko kvasovk in nova obzorja za njihovo biotehnološko uporabo. Tako se znanost o kvasu, ki je prepotovala več kot stoletje in pol, še naprej hitro razvija v 21. stoletju.
Za mikroskopsko preiskavo pripravimo tako nativne (neobarvane) kot obarvane preparate. Za preučevanje gliv v tkivih se opravi patohistološki pregled.
celični micelij trosov kvasovk
30. Morfologija in raziskovalne metode nitastih gliv. Pri glivah obstajata dve vrsti rasti: hifalna rast (hyphomycetes) in rast kvasovk (blastomycetes). Običajno je vegetativno telo nitastih gliv sestavljeno iz filamentov, debelih približno 5 µm, ki so zelo razvejani in se imenujejo hife. Hife nimajo prečnih pretin (pri nižjih glivah) ali pa so s pregradami (septami) ločene v celice (pri višje gobe). Celična stena je lahko različne debeline, obvod je pogosto dobro viden, med vključki v citoplazmi so najbolj značilna zrnca volutina, glikogena, pigmenta melanina. Zrele stare celice gliv so bogate z lipidi. Jedro vsebuje nukleolus in kromatinsko mrežo; celice so lahko večjedrne. Zbirka hif tvori micelij (micelij). Micelij je lahko substrat, ki nastane kot posledica vraščanja hif v hranilni medij, in zrak, ki raste na površini medija. Micelij predstavlja razvejane cevi, razvejanje izvajajo stranski izrastki hif. Micealni filamenti so včasih razporejeni v vzporednih vrstah, tesno drug ob drugem, ki spominjajo na "stenj", od tod tudi ime "koremija" (pri dermatofitih).
31. Osnovna načela razvrščanja in morfologije praživali. Metode njihovega raziskovanja in medicinski pomen.
Praživali so enocelični evkarionti, po strukturi podobni celicam kompleksno organiziranih živali. Oblika je lahko hruškasta (Trichomonas, Giardia), jajčasta (Balantidia), vretenasta (Trypanosoma, Leishmania), lahko prevzame najbolj bizarno konfiguracijo (ameba)
Večina protozojev je mobilnih in gibanje se izvaja s pomočjo psevdopodije (ameba, malarijski plazmodij), bičkov (giardia, leishmania), cilij (balantidij).
Psevdopodiji- začasne izbokline citoplazme, ob sproščanju katerih praživali ves čas spreminjajo obliko telesa.
Najenostavnejši pripadajo kraljestvu Praživali (protos - prvi, zoa - živali). Zdravstvenega pomena so:
Phylum Sarcomastigophora, subphylum Sarcodina (Sarcodidae). Njihovo telo je brez ovoja, premikajo se s pomočjo psevdopodijev. Ta razred vključuje različne vrste ameba, vključno z dizenterično amebo (Entamoeba histolytica).
Raziskovalne metode. Za preučevanje praživali se pripravijo začasni in trajni (obarvani) preparati. Začasne pripravke pripravljamo po metodi "zdrobljene kapljice" ali "viseče kapljice" z dodatkom tople fiziološka raztopina ali vitalna intravitalna barvila. Iz krvnih pripravkov pripravimo "gosto kapljico". Da bi to naredili, prst, obdelan z etrom, obrnemo s punkcijo navzdol in stekleno stekelce pripeljemo do štrlečih kapljic, na katere vzamemo 2-3 kapljice krvi, nato pa kri porazdelimo z iglo ali vogalom drugo stekelce, da dobimo oval približno 1 cm, da pospešimo sušenje zdravila, ga lahko postavimo v termostat na 35-37 0 C.
Za odkrivanje cist se uporablja močna Lugolova raztopina, ki obarva strukture cist v temno rjavo barvo.
Obarvano z železovim hematoksilinom po metodi Heidenhain
Brise po fiksaciji damo v 2,5% raztopino železovega galuna za 1 uro.
Po trikratnem izpiranju v vodi obarvamo z barvilom (0,5 g hematoksilina, 10 ml 96 0 alkohola in po raztapljanju dodamo 90 ml destilirane vode) 5-10 minut.
Oprano z vodo in posušeno.
Barvanje je nepogrešljivo v primerih, ko je treba razkriti najmanjše podrobnosti strukture jedra in citoplazme protozojev.
Celične stene rastlin imajo izredno trdnost, v procesu rasti pa rastline spreminjajo svojo strukturo in sestavo. Glavne sestavine rastlinskih celičnih sten so polisaharidi, med katerimi prevladuje celuloza, ki v veliki meri določa arhitekturo stene.
Celuloza. Ta homopolisaharid je najpogostejši ogljikov hidrat na Zemlji (rastline tvorijo do 1011 ton celuloze na leto). Celulozni monomeri so ostanki glukoze, povezani v dolge verige (do 10.000 ostankov glukoze v vsaki) z b (1 → 4) -glikozidnimi vezmi (slika 5.3). V takšni molekuli ni popolne svobode vrtenja okoli 1C-O- in O-4C-vezi, polimer pa pridobi konformacijo, ugodno za tvorbo medverižnih vodikovih vezi, ko so verige razporejene antiparalelno. Posledično se molekule celuloze združijo v mikrofibrile približno 10 do 25 nm debeline. Mikrofibrile se prepletajo in oblikujejo tanke niti, ki jih je mogoče oviti enega okoli drugega, kot pramene v vrvi, ki tvorijo makrofibrile. Vsaka makrofibrila je debela približno 0,5 µm in lahko doseže dolžino 6-8 µm. Trdnost makrofibril je primerljiva z trdnostjo jeklene žice enake debeline. Poleg tega ločeni odseki mikrofibrili imajo urejeno strukturo in dajejo celični steni kristalne lastnosti. Tako lahko opazimo kompleksnost in visoko urejenost celuloze v sestavi celičnih sten, kar ni naključje: ta polimer opravlja zaščitne in podporne funkcije v rastlini.
V tej obliki so polisaharidi nedostopni za delovanje lastnih encimov, celuloze pa rastlina ne more uporabiti kot rezervno snov. Le nekaj organizmov (nekatere bakterije, glive, praživali in redke živali) ima encimske sisteme, ki lahko razgradijo celulozo.
Mikro- in makrofibrile celuloze v celični steni rastlin so potopljene v matrica, ki je prav tako sestavljen pretežno iz polisaharidov in med rastjo rastlin spreminja svojo strukturo. Vklopljeno zgodnje faze razvojna matrica je sestavljena iz pektinske snovi, in se pozneje pojavijo v njem ksilani in različni nevtralni polisaharidi ("hemiceluloza"). Pektinske snovi so polimeri a-galakturonske kisline, v katerih so nekateri atomi vodika nadomeščeni z metilnimi skupinami (-CH 3) (slika 5.3). Ksilani so polimeri ksiloze (slika 5.3).
Na poznejših stopnjah razvoja, ko pride do lignifikacije celičnih sten, lignin- vsebuje kemično odporen polimer velika številka aromatični obroči. Poleg tega se v sestavi rastlinskih celičnih sten nahajajo majhne količine glikoproteinov, netopnih lipidnih polimerov. drugačna struktura in voski.
Celične stene nekaterih rastlin vsebujejo redke polisaharide z nenavadno strukturo. Na primer v stenah in medceličnini
Snov morskih rdečih alg vsebuje kompleksen heteropolisaharidni agar, ki je mešanica sulfatiranih polisaharidov - agaroze in agaropektina. Agaroza je zgrajena iz izmenjujočih se ostankov D-galaktoze in 3,6-anhidro-L-laktoze, povezanih izmenično z b(1→4)- in a(1→3)-vezmi. Agaropektin ima več kompleksna struktura: Vsebuje D-galaktozo, 3,6-anhidrogalaktozo, uronske kisline in sulfat. Agar se uporablja kot najpogostejši kompaktor za trdna gojišča, nepogrešljiv v mikrobiologiji, pa tudi v Prehrambena industrija za želirne izdelke. Treba je opozoriti, da velika večina mikroorganizmov ne more razgraditi agarja, in to je ena njegovih glavnih prednosti pred drugim kompaktorjem hranilnih medijev - želatino. Agarozne najdbe široka uporaba v biokemijskih raziskavah: ona vodno okolje tvori gel z velikimi porami, katerih velikost je določena z njegovo koncentracijo. Agarozni geli se uporabljajo za frakcioniranje proteinov in nukleinskih kislin ter za imobilizacijo celic.
- 20,44 KbCelična stena, struktura, kemična sestava. Spremembe celične stene.
Prisotnost močne lupine značilnost rastlinska celica, ki jo razlikuje od živalske celice. Lupina daje celici določeno obliko in trdnost ter ščiti živo vsebino - protoplast in predvsem plazmalemo, ki je z notranje strani tesno stisnjena na lupino. Gojene na posebnih hranilnih medijih imajo celice višjih rastlin, v katerih je membrana odstranjena z encimsko metodo, vedno sferično obliko. Brez lupine je obstoj rastlinske celice v normalnih pogojih nemogoč, saj bi njen turgorski tlak, uravnotežen s protitlakom lupine, neizogibno povzročil raztrganje plazmaleme in uničenje protoplasta. Skupek celičnih sten v sestavi telesa kopenske rastline, ki se dviga nad površino tal in nima notranjega skeleta, je nekakšen nosilni okvir, ki daje rastlini mehansko moč.
Lupina je praviloma brezbarvna in prozorna, zlahka prenaša sončno svetlobo. Po njej se lahko premikajo voda in raztopljene snovi z nizko molekulsko maso. Vsaka celica ima svojo lupino; lupine sosednjih celic so tako rekoč cementirane z medceličnimi snovmi, ki tvorijo tako imenovano srednjo lamino. Posledično so sosednje celice med seboj ločene s steno, ki jo tvorita dve lupini in srednja plošča, zaradi česar lupino imenujemo tudi celična stena.
Lupino gradi protoplast celice in zato lahko raste le v stiku z njim. Zelo pogosto se lupina obdrži dlje kot protoplast (ko celica predčasno odmre), vendar to običajno ne vodi do prenehanja obstoja celice kot strukturne enote, ker se oblika celice ne spremeni zaradi trdnost lupine. Zato lahko rastlinske celice tudi po smrti opravljajo pomembne funkcije transporta raztopin ali mehanske podpore. Številne vrste rastlinskih celic - vlakna, traheidi, žilni segmenti, celice plute - so v odraslem stanju ena celična membrana. Večinoma je les sestavljen iz lupin odmrlih celic.
Celične membrane so tako raznolike kot celice same. Sestavo in strukturo membrane lahko pogosto uporabimo za presojo izvora in delovanja celic različnih vrst. Tako zgradbo fosilnih rastlin proučujemo predvsem s preučevanjem membran njihovih celic.
Sprva se primarna celična stena razvije navzven od plazmaleme. Sestavljen je iz polisaharidov - pektina in celuloze. Primarne celične stene sosednjih celic povezuje protopektinska mediana lamina. V celični steni so linearne, zelo dolge molekule celuloze, sestavljene iz glukoze, sestavljene v snope - micele, ki se nato združijo v fibrile - najtanjša vlakna nedoločene dolžine. Celuloza tvori večdimenzionalen skelet, ki je potopljen v amorfno močno razvodnjeno matrico neceluloznih ogljikovih hidratov: pektinov, hemiceluloz itd. Celuloza je tista, ki določa trdnost celične stene. Mikrofibrile so elastične in imajo podobno natezno trdnost kot jeklo. Matrični polisaharidi določajo lastnosti stene, kot so visoka prepustnost za vodo, raztopljene majhne molekule in ione ter močno nabrekanje. Zahvaljujoč matrici se lahko voda in snovi premikajo iz celice v celico vzdolž sten, ki mejijo ena na drugo. Nekatere hemiceluloze se lahko odlagajo v celične stene semen kot rezervne snovi.
Material lupine, vključno s celulozo, se pogosto uporablja v industriji. Lastnosti lesa, papirja, tekstilnih materialov so v veliki meri odvisne od molekularne zgradbe in kemične sestave lupine, poznavanje le-teh pa pomaga izboljšati proizvodno tehnologijo in kakovost izdelkov.
Kemična sestava in molekularna organizacija lupine. Celična membrana je zgrajena pretežno iz polisaharidov, katerih monomeri – sladkorji so med seboj povezani z glikozidno vezjo (-O-) v obliki verige. Polisaharide poimenujemo glede na sestavo monomerov, ki jih tvorijo, z dodatkom končnice "an". Na primer, polisaharid ksilan je sestavljen iz ostankov sladkorja ksiloze, manan - manoza, glukan - glukoza itd.
Poleg polisaharidov lahko sestava lupine vključuje beljakovine, mineralne soli, lignin, pigmente, lipide. Ponavadi so lupine impregnirane z vodo. Polisaharide lupine glede na njihovo vlogo lahko razdelimo na skeletne snovi in matrične snovi. Skeletna snov lupine višje rastline je celuloza (vlaknina), ki kemično predstavlja β-1, 4-D glukan. Število glukoznih ostankov v molekuli in posledično dolžina samih molekul se lahko v celičnih stenah različnih rastlin močno razlikuje, kar vpliva na lastnosti celuloze. Njegove nitaste molekule v lupini so razporejene vzporedno druga z drugo in so združene po več deset tako, da se pojavi podolgovata tridimenzionalna mreža, ki je značilna za kristale. Te kristalne skupine, pri nastanku katerih sodelujejo kovalentne in vodikove vezi, tvorijo osnovo mikrofibril - najtanjših vlaken, ki ustvarjajo strukturni okvir lupine, njeno "ojačitev".
Kristalno stanje celuloznih molekul v mikrofibrilih določa dvolomnost celične stene, ki jo opazujemo v polarizacijskem mikroskopu. V mikrofibrilih obstajajo parakristalna področja, tj. območja z ohlapnejšo vzporedno, vendar brez tridimenzionalne orientacije, razporeditvijo celuloznih molekul. Stopnja kristaliničnosti mikrofibril, razmerje med kristalnimi in parakristaliničnimi conami se precej razlikujejo glede na vrsto rastline, stanje celice in vrsto lupine, kar vpliva na mehanske in fizikalne celuloze kot surovine. Premer mikrofibril se razlikuje tudi v različnih celicah (običajno znotraj 10-30 nm), dolžina lahko doseže več mikrometrov. Mikrofibrile lahko združimo v ločene skupine ali plasti - makrofibrile z debelino 0,4-0,5 mikronov, vidne v svetlobnem mikroskopu.
Slika 25
Med življenjem celic, zlasti glede na izvajanje katere koli specializirane funkcije, pride do sekundarnih sprememb v celični steni. Pogosto so povezani z radikalnimi spremembami v njegovi kemični sestavi, strukturi in fizikalno-kemijskih lastnostih.
Drvarjenje. Celična stena je obložena (impregnirana) s posebno snovjo - ligninom, ki poveča trdoto, kalorično vsebnost, gostoto celične stene ter zmanjša njeno plastičnost in sposobnost rasti. Lignificirane celične stene ne izgubijo sposobnosti prehajanja vode in zraka. Njihov protoplast lahko ostane živ, čeprav običajno odmre. Sečnja je v naravi zelo razširjena. Daje trdnost deblom in vejam dreves. Les iglavcev in listavcev vsebuje do 50% celuloze in 20...30% lignina. Lignificirane celične stene in številna zelišča, zlasti proti koncu rastne sezone.
Sondiranje ali suberinizacija. Kot posledica obilnega nanosa v celično steno kemično odporne snovi - suberina, nastane njen suberin. Po svoji kemični naravi je suberin blizu maščobam. Je amorfna hidrofobna spojina, ki je sestavljena iz visoko polimernih nasičenih maščobnih kislin in kompleksnih hidroksi kislin. Suberin se nalaga predvsem v celičnih stenah sekundarnega pokrivnega tkiva – plute, ki je odličen vodni in toplotni izolator. V majhnih količinah se kopiči v celičnih stenah različnih tkiv. S popolnim zamašitvijo protoplast celice umre, saj suberinske lamele preprečujejo prodiranje vode in zraka, z delnim - protoplast se ohrani dolgo časa.
Mineralizacija. V procesu življenja lahko stene površinskih celic poganjkov kopičijo mineralne snovi, zlasti silicijev dioksid in kalcijev karbonat.Nekatere skupine rastlin, na primer številne vrste šašev, trave, preslice, imajo vedno celice pokrovnega tkiva , povrhnjica, močno prekrita s solmi. Poleg tega ni lahko inkrustrirana samo celična stena, ampak tudi različni izrastki povrhnjice - trihomi. Poleg tega se mineralne snovi odlagajo na površini celic pokrivnega tkiva - povrhnjice. Zaradi mineralizacije pridobijo aksialni organi večjo mehansko trdnost.
Hujšanje. Preoblikovanje celuloze in pektina v sluz in bližnje gume, ki so polimerni ogljikovi hidrati, ki se odlikujejo po svoji sposobnosti, da močno nabreknejo v stiku z vodo. Sluz opazimo v celičnih celicah semenskega ovoja, na primer pri lanu. Tvorba sluzi je velikega prilagoditvenega pomena. Ko semena kalijo, jih sluz fiksira na določenem mestu, zlahka absorbira in zadržuje vlago, ščiti semena pred izsušitvijo in izboljša vodni režim sadik. Sluz celičnih sten koreninskega vloska zagotavlja njihovo močno vez z delci zemlje.
Maceracija je raztapljanje medcelične snovi, kar vodi do ločitve celic. Naravna maceracija poteka v zrelih plodovih. Umetno se izvaja, na primer, ko je lan lobiran, da se sprostijo predilne surovine - skupine celic ličnih vlaken.
Opis dela
Prisotnost močne lupine je značilna lastnost rastlinska celica ki ga razlikuje od živalske celice. Lupina daje celici določeno obliko in trdnost ter ščiti živo vsebino - protoplast in predvsem plazmalemo, ki je z notranje strani tesno stisnjena na lupino. Gojene na posebnih hranilnih medijih imajo celice višjih rastlin, v katerih je membrana odstranjena z encimsko metodo, vedno sferično obliko.