Visi ląstelių formos gyvybę žemėje galima suskirstyti į dvi karalystes pagal jas sudarančių ląstelių struktūrą – prokariotus (ikibranduolinius) ir eukariotus (branduolinius). Prokariotinės ląstelės yra paprastesnės struktūros, matyt, jos atsirado anksčiau evoliucijos procese. Eukariotinės ląstelės – sudėtingesnės, atsirado vėliau. Ląstelės, sudarančios žmogaus kūną, yra eukariotinės.
Nepaisant formų įvairovės, visų gyvų organizmų ląstelių organizavimui taikomi vienodi struktūriniai principai.
prokariotinė ląstelė
eukariotinė ląstelė
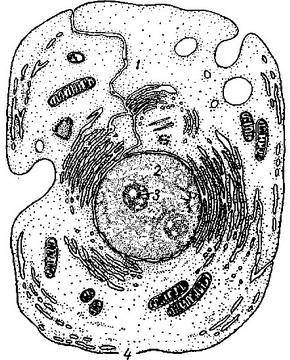
Eukariotinės ląstelės sandara
paviršiaus kompleksas gyvūnų ląstelė
Apima glikokaliksas, plazmolema ir apatinis žievės citoplazmos sluoksnis. Plazminė membrana taip pat vadinama plazmolema ląstelės membrana. Tai maždaug 10 nanometrų storio biologinė membrana. Visų pirma atlieka ribinę funkciją, susijusią su išorine ląstelės aplinka. Be to, atlieka transportavimo funkciją. Ląstelė neeikvoja energijos savo membranos vientisumui palaikyti: molekulės laikomos pagal tą patį principą, pagal kurį laikomos kartu riebalų molekulės – termodinamiškai naudingiau, jei hidrofobinės molekulių dalys yra arti vienas kitą. Glikokaliksas susideda iš oligosacharidų, polisacharidų, glikoproteinų ir glikolipidų molekulių, „įtvirtintų“ plazmalemoje. Glikokaliksas atlieka receptorių ir žymenų funkcijas. Gyvūnų ląstelių plazmos membraną daugiausia sudaro fosfolipidai ir lipoproteinai, susimaišę su baltymų molekulėmis, ypač paviršiaus antigenais ir receptoriais. Žievėje (greta plazmos membrana) citoplazmos sluoksnyje yra specifiniai citoskeleto elementai – tam tikru būdu sutvarkyti aktino mikrofilamentai. Pagrindinė ir svarbiausia žievės sluoksnio (žievės) funkcija yra pseudopodinės reakcijos: pseudopodijų išstūmimas, prisitvirtinimas ir sumažinimas. Tokiu atveju mikrofilamentai pertvarkomi, pailginami arba trumpinami. Ląstelės forma (pavyzdžiui, mikrovilliukų buvimas) taip pat priklauso nuo žievės sluoksnio citoskeleto struktūros.
Citoplazmos struktūra
Skystas citoplazmos komponentas taip pat vadinamas citozoliu. Šviesos mikroskopu atrodė, kad ląstelė užpildyta kažkuo panašaus į skystą plazmą ar solą, kuriame „plaukioja“ branduolys ir kitos organelės. Iš tikrųjų taip nėra. Vidinė eukariotinės ląstelės erdvė yra griežtai sutvarkyta. Organelių judėjimas koordinuojamas pasitelkiant specializuotas transporto sistemas, vadinamuosius mikrotubulus, kurie tarnauja kaip tarpląsteliniai „keliai“, ir specialius baltymus dyneinus ir kinezinus, kurie atlieka „variklio“ vaidmenį. Atskiros baltymų molekulės taip pat laisvai neišsisklinda po visą viduląstelinę erdvę, o nukreipiamos į reikiamus skyrius, naudojant specialius signalus ant jų paviršiaus, atpažįstamus ląstelės transportavimo sistemų.
Endoplazminis Tinklelis
Eukariotų ląstelėje yra membraninių skyrių, einančių vienas į kitą (vamzdeliai ir rezervuarai), sistema, vadinama endoplazminiu tinkleliu (arba endoplazminiu tinkleliu, EPR arba EPS). Ta ER dalis, prie kurios membranos yra prijungtos ribosomos, vadinama granuliuotas(arba šiurkštus) į endoplazminį tinklą, jo membranose vyksta baltymų sintezė. Tie skyriai, kurių sienelėse nėra ribosomų, klasifikuojami kaip sklandžiai(arba agranuliuotas) EPR, kuris dalyvauja lipidų sintezėje. Lygios ir granuliuotos ER vidinės erdvės nėra izoliuotos, o pereina viena į kitą ir susisiekia su branduolinės membranos spindžiu.
Goldžio kompleksas
Šerdis
citoskeletas
Centrioliai
Mitochondrijos
Pro- ir eukariotinių ląstelių palyginimas
Svarbiausias skirtumas tarp eukariotų ir prokariotų ilgam laikui buvo laikomas susiformavusio branduolio buvimas ir membranos organelės. Tačiau iki 1970 ir 1980 m tapo aišku, kad tai tik gilesnių citoskeleto organizavimo skirtumų pasekmė. Kurį laiką buvo manoma, kad citoskeletas būdingas tik eukariotams, tačiau 1990 m. bakterijose taip pat buvo rasta baltymų, homologiškų pagrindiniams eukariotų citoskeleto baltymams.
Būtent specialiai išdėstyto citoskeleto buvimas leidžia eukariotams sukurti judrių vidinių membraninių organelių sistemą. Be to, citoskeletas leidžia atlikti endo- ir egzocitozę (manoma, kad būtent dėl endocitozės eukariotinėse ląstelėse atsirado tarpląsteliniai simbiontai, įskaitant mitochondrijas ir plastidus). Kita svarbi eukariotų citoskeleto funkcija – užtikrinti eukariotinės ląstelės branduolio (mitozės ir mejozės) ir kūno (citotomijos) dalijimąsi (prokariotinių ląstelių dalijimasis organizuojamas paprasčiau). Citoskeleto struktūros skirtumai paaiškina ir kitus skirtumus tarp pro- ir eukariotų – pavyzdžiui, formų pastovumą ir paprastumą. prokariotinės ląstelės ir reikšminga formos įvairovė ir galimybė ją keisti eukariotuose, taip pat santykinai dideli dydžiai pastarasis. Taigi prokariotinių ląstelių dydis yra vidutiniškai 0,5–5 mikronai, eukariotinių ląstelių dydžiai - vidutiniškai nuo 10 iki 50 mikronų. Be to, tik tarp eukariotų yra tikrai milžiniškų ląstelių, tokių kaip masyvūs ryklių ar stručių kiaušinėliai ( paukščio kiaušinis visas trynys yra viena didžiulė kiaušinių ląstelė), stambių žinduolių neuronai, kurių procesai, sustiprinti citoskeleto, gali siekti keliasdešimt centimetrų.
Anaplazija
Ląstelių struktūros sunaikinimas (pavyzdžiui, esant piktybiniams navikams) vadinamas anaplazija.
Ląstelių atradimo istorija
Pirmasis žmogus, pamatęs ląsteles, buvo anglų mokslininkas Robertas Hukas (mums žinomas Huko dėsnio dėka). Tais metais, bandydamas suprasti, kodėl kamštienos medis taip gerai plaukia, Hooke'as savo patobulintu mikroskopu pradėjo tirti plonas kamštienos dalis. Jis išsiaiškino, kad kamštiena buvo padalinta į daugybę mažyčių celių, kurios jam priminė vienuolines ląsteles, ir pavadino šias ląsteles (anglų kalba cell reiškia „ląstelė, ląstelė, ląstelė“). Tais metais olandų meistras Antony van Leeuwenhoek (Anton van Leeuwenhoek, -) mikroskopu pirmą kartą vandens laše pamatė „gyvūnus“ – judančius gyvus organizmus. Taigi iki XVIII amžiaus pradžios mokslininkai žinojo, kad esant dideliam padidinimui augalai turi ląstelinę struktūrą, ir pamatė kai kuriuos organizmus, kurie vėliau buvo pavadinti vienaląsčiais. Tačiau ląstelinė organizmų sandaros teorija susiformavo tik XIX amžiaus viduryje, pasirodžius galingesniems mikroskopams ir sukūrus ląstelių fiksavimo bei dažymo metodus. Vienas iš jos įkūrėjų buvo Rudolfas Virchovas, tačiau jo idėjose buvo nemažai klaidų: pavyzdžiui, jis manė, kad ląstelės yra silpnai sujungtos viena su kita ir kiekviena egzistuoja „savaime“. Tik vėliau pavyko įrodyti ląstelių sistemos vientisumą.
Planas:
- Įvadas
- 1 Ląstelių struktūra
- 1.1 prokariotinė ląstelė
- 1.2
eukariotinė ląstelė
- 1.2.1
Eukariotinės ląstelės sandara
- 1.2.1.1 Gyvūnų ląstelių paviršiaus kompleksas
- 1.2.1.2 Citoplazmos struktūra
- 1.2.1.3 Endoplazminis Tinklelis
- 1.2.1.4 Golgi aparatas
- 1.2.1.5 Branduolys
- 1.2.1.6 Lizosomos
- 1.2.1.7 Citoskeletas
- 1.2.1.8 Centrioliai
- 1.2.1.9 Mitochondrijos
- 1.2.1
Eukariotinės ląstelės sandara
- 1.3 Pro- ir eukariotinių ląstelių palyginimas
- 1.4 Anaplazija
- 2 Ląstelių atradimo istorija
- 3 Cheminė sudėtis ląstelės
Įvadas
Ląstelė- elementarus visų gyvų organizmų (išskyrus virusus, kurie dažnai vadinami neląstelinėmis gyvybės formomis) sandaros ir gyvybinės veiklos vienetas, turintis savo medžiagų apykaitą, galintis savarankiškai egzistuoti, savaime daugintis ir vystytis. Visi gyvi organizmai, kaip ir daugialąsčiai gyvūnai, augalai ir grybai, susideda iš daugelio ląstelių arba, kaip ir daugelis pirmuonių ir bakterijų, yra vienaląsčiai organizmai. Biologijos šaka, tirianti ląstelių sandarą ir aktyvumą, vadinama citologija. IN Pastaruoju metu taip pat įprasta kalbėti apie ląstelės biologiją arba ląstelių biologija(Anglų) ląstelių biologija).
Nuotraukose žalias fluorescencinis baltymas rodo įvairių ląstelės dalių vietą.
1. Ląstelių sandara
Visos ląstelinės gyvybės formos žemėje gali būti suskirstytos į dvi karalystes, remiantis jas sudarančių ląstelių struktūra:
- prokariotai (ikibranduoliniai) - paprastesnės struktūros, matyt, jie atsirado anksčiau evoliucijos procese;
- eukariotai (branduoliniai) – sudėtingesni, atsirado vėliau. Ląstelės, sudarančios žmogaus kūną, yra eukariotinės.
Nepaisant formų įvairovės, visų gyvų organizmų ląstelių organizavimui taikomi vienodi struktūriniai principai.
Ląstelės turinys nuo aplinkos atskiriamas plazmine membrana arba plazmelema. Ląstelės viduje yra citoplazma, kurioje yra įvairios organelės Ir ląstelių inkliuzai, taip pat genetinė medžiaga DNR molekulės pavidalu. Kiekviena ląstelės organelė atlieka savo ypatingą funkciją ir visos kartu lemia visos ląstelės gyvybinę veiklą.
1.1. prokariotinė ląstelė
Tipiškos prokariotinės ląstelės struktūra: kapsulė, ląstelių sienelės, plazmalema, citoplazma, ribosomos, plazmidė, pilis, žvyneliai, nukleoidas.
prokariotai(iš lat. pro- prieš, prieš ir graikiškai. κάρῠον - branduolys, riešutas) - organizmai, kurie, skirtingai nei eukariotai, neturi susiformavusio ląstelės branduolio ir kitų vidinių membranų organelių (išskyrus plokščias cisternas fotosintetinėse rūšyse, pavyzdžiui, melsvadumblių). Vienintelė didelė žiedinė (kai kuriose rūšyse – linijinė) dvigrandė DNR molekulė, kurioje yra pagrindinė ląstelės genetinės medžiagos dalis (vadinamasis nukleoidas), nesudaro komplekso su histono baltymais (vadinamuoju chromatinu). Prokariotai apima bakterijas, įskaitant melsvadumblius (melsvadumblius) ir archajas. Organelės yra prokariotinių ląstelių palikuonys. eukariotinės ląstelės- mitochondrijos ir plastidai.
1.2. eukariotinė ląstelė
eukariotų(eukariotai) (iš graikų k. ευ - gerai, visiškai κάρῠον - branduolys, riešutas) - organizmai, kurie, skirtingai nei prokariotai, turi suformuotą ląstelės branduolį, atskirtą nuo citoplazmos branduoline membrana. Genetinė medžiaga yra uždaryta keliose linijinėse dvigrandėse DNR molekulėse (priklausomai nuo organizmų tipo, jų skaičius viename branduolyje gali svyruoti nuo dviejų iki kelių šimtų), pritvirtintomis iš vidaus prie membranos. ląstelės branduolys ir didžiojoje daugumoje (išskyrus dinoflagellatus) sudaro kompleksą su histono baltymais, vadinamą chromatinu. Eukariotinės ląstelės turi vidinių membranų sistemą, kuri, be branduolio, sudaro daugybę kitų organelių (endoplazminis tinklas, Golgi aparatas ir kt.). Be to, didžioji dauguma turi nuolatinius viduląstelinius simbiontus-prokariotus – mitochondrijas, o dumbliai ir augalai taip pat turi plastidžių.
1.2.1. Eukariotinės ląstelės sandara
Scheminis gyvūno ląstelės vaizdas. (Spustelėdami bet kurį iš pavadinimų sudedamosios dalys langelius, būsite nukreipti į atitinkamą straipsnį.)
1.2.1.1. Gyvūnų ląstelių paviršiaus kompleksas
Jį sudaro glikokaliksas, plazmalema ir po juo esantis citoplazmos žievės sluoksnis. Plazminė membrana taip pat vadinama plazmelema, išorine ląstelės membrana. Tai biologinė membrana, apie 10 nanometrų storio. Visų pirma atlieka ribinę funkciją, susijusią su išorine ląstelės aplinka. Be to, atlieka transportavimo funkciją. Ląstelė neeikvoja energijos savo membranos vientisumui palaikyti: molekulės laikomos pagal tą patį principą, pagal kurį laikomos kartu riebalų molekulės – termodinamiškai naudingiau, jei hidrofobinės molekulių dalys yra arti vienas kitą. Glikokaliksas yra plazmalema pritvirtintas oligosacharidas, polisacharidas, glikoproteinas ir glikolipido molekulė. Glikokaliksas atlieka receptorių ir žymenų funkcijas. Gyvūnų ląstelių plazmos membraną daugiausia sudaro fosfolipidai ir lipoproteinai, susimaišę su baltymų molekulėmis, ypač paviršiaus antigenais ir receptoriais. Žieviniame (greta plazminės membranos) citoplazmos sluoksnyje yra specifiniai citoskeleto elementai – tam tikru būdu sutvarkyti aktino mikrofilamentai. Pagrindinė ir svarbiausia žievės sluoksnio (žievės) funkcija yra pseudopodinės reakcijos: pseudopodijų išstūmimas, prisitvirtinimas ir susitraukimas. Tokiu atveju mikrofilamentai pertvarkomi, pailginami arba trumpinami. Ląstelės forma (pavyzdžiui, mikrovilliukų buvimas) taip pat priklauso nuo žievės sluoksnio citoskeleto struktūros.
1.2.1.2. Citoplazmos struktūra
Skystas citoplazmos komponentas taip pat vadinamas citozoliu. Šviesos mikroskopu atrodė, kad ląstelė užpildyta kažkuo panašaus į skystą plazmą ar solą, kuriame „plūduriavo“ branduolys ir kitos organelės. Iš tikrųjų taip nėra. Vidinė eukariotinės ląstelės erdvė yra griežtai sutvarkyta. Organelių judėjimas koordinuojamas pasitelkiant specializuotas transporto sistemas, vadinamuosius mikrotubulus, kurie tarnauja kaip tarpląsteliniai „keliai“, ir specialius baltymus dyneinus ir kinezinus, kurie atlieka „variklio“ vaidmenį. Atskiros baltymų molekulės taip pat laisvai neišsisklinda po visą viduląstelinę erdvę, o nukreipiamos į reikiamus skyrius, naudojant specialius signalus ant jų paviršiaus, atpažįstamus ląstelės transportavimo sistemų.
1.2.1.3. Endoplazminis Tinklelis
Eukariotų ląstelėje yra membraninių skyrių, einančių vienas į kitą (vamzdeliai ir rezervuarai), sistema, vadinama endoplazminiu tinkleliu (arba endoplazminiu tinkleliu, EPR arba EPS). Ta ER dalis, prie kurios membranos yra prijungtos ribosomos, vadinama granuliuotas(arba šiurkštus) į endoplazminį tinklą, jo membranose vyksta baltymų sintezė. Tie skyriai, kurių sienelėse nėra ribosomų, klasifikuojami kaip sklandžiai(arba agranuliuotas) EPR, kuris dalyvauja lipidų sintezėje. Lygios ir granuliuotos ER vidinės erdvės nėra izoliuotos, o pereina viena į kitą ir bendrauja su spindžiu branduolinis apvalkalas.
1.2.1.4. Goldžio kompleksas
Golgi aparatas yra plokščių membraninių cisternų krūva, šiek tiek išsiplėtusi arčiau kraštų. Golgi aparato rezervuaruose bręsta kai kurie baltymai, susintetinti ant granuliuoto ER membranų ir skirti sekrecijai arba lizosomų formavimui. Golgi aparatas yra asimetriškas - cisternos yra arčiau ląstelės branduolio ( cis-Golgi) turi mažiausiai subrendusių baltymų, prie šių rezervuarų nuolatos prisitvirtina membraninės pūslelės, pūslelės, išaugančios iš endoplazminio tinklo. Matyt, tų pačių pūslelių pagalba vyksta tolesnis bręstančių baltymų judėjimas iš vienos talpos į kitą. Galiausiai iš priešingo organelės galo ( transas-Golgi) pūslelės, kuriose yra visiškai subrendusių baltymų, išsiskleidžia.
1.2.1.5. Šerdis
Ląstelės branduolyje yra DNR molekulės, kuriose yra genetinė organizmo informacija. Branduolyje vyksta replikacija – DNR molekulių dubliavimasis, taip pat transkripcija – RNR molekulių sintezė ant DNR matricos. Branduolyje susintetintos RNR molekulės patiria tam tikrų modifikacijų (pavyzdžiui, susijungimo procese nereikšmingos, beprasmės atkarpos pašalinamos iš pasiuntinio RNR molekulių), po to patenka į citoplazmą. Ribosomų surinkimas taip pat vyksta branduolyje. Specialusis ugdymas vadinami branduoliais. Branduolio skyrius - karioteka - susidaro išplečiant ir sujungiant vienas su kitu endoplazminio tinklo rezervuarus taip, kad branduolys turėtų dvigubas sieneles dėl jį supančių siaurų branduolio membranos skyrių. Branduolinio apvalkalo ertmė vadinama liumenas arba perinuklearinė erdvė. Vidinis paviršius Branduolinė membrana yra padengta branduolio sluoksniu, standžia baltymo struktūra, kurią sudaro laminų baltymai, prie kurių yra prijungtos chromosomų DNR grandinės. Kai kuriose vietose vidinė ir išorinė branduolio apvalkalo membranos susilieja ir suformuoja vadinamąsias branduolio poras, per kurias vyksta medžiagų mainai tarp branduolio ir citoplazmos.
1.2.1.6. Lizosomos
Lizosoma yra mažas kūnas, kurį riboja viena citoplazmos membrana. Jame yra lizinių fermentų, kurie gali suskaidyti visus biopolimerus. Pagrindinė funkcija yra autolizė - tai yra atskirų organelių, ląstelės citoplazmos dalių suskaidymas.
1.2.1.7. citoskeletas
Citoskeleto elementai apima baltymines fibrilines struktūras, esančias ląstelės citoplazmoje: mikrovamzdelius, aktiną ir tarpinius siūlus. Mikrovamzdeliai dalyvauja organelių transporte, yra žvynelių dalis, o mitozinis velenas yra sudarytas iš mikrovamzdelių. Aktino gijos yra būtinos ląstelių formai palaikyti, pseudopodialinėms reakcijoms. Atrodo, kad tarpinių gijų vaidmuo taip pat yra išlaikyti ląstelės struktūrą. Citoskeleto baltymai sudaro kelias dešimtis procentų ląstelės baltymo masės.
1.2.1.8. Centrioliai
Centrioliai yra cilindrinės baltymų struktūros, esančios šalia gyvūnų ląstelių branduolio (augalai centriolių neturi). Centrolė yra cilindras, kurio šoninį paviršių sudaro devyni mikrovamzdelių rinkiniai. Mikrotubulių skaičius rinkinyje gali svyruoti skirtingi organizmai nuo 1 iki 3.
Aplink centrioles yra vadinamasis citoskeleto organizavimo centras – sritis, kurioje sugrupuoti ląstelės mikrotubulių minusiniai galai.
Prieš dalijimąsi ląstelėje yra du centrioliai, išdėstyti stačiu kampu vienas kito atžvilgiu. Mitozės metu jie nukrypsta į skirtingus ląstelės galus, sudarydami dalijimosi veleno polius. Po citokinezės kiekviena dukterinė ląstelė gauna vieną centriolę, kuri padvigubėja kitam dalijimuisi. Centrolių padvigubėjimas vyksta ne dalijantis, o sintezuojant naują struktūrą, statmeną esamai.
Centrioliai atrodo homologiški baziniai kūnaižvyneliai ir blakstienos.
1.2.1.9. Mitochondrijos
Mitochondrijos yra specialios ląstelių organelės, kurių pagrindinė funkcija yra ATP – universalaus energijos nešiklio – sintezė. kvėpavimas (deguonies paėmimas ir išleidimas anglies dioksidas) taip pat atsiranda dėl mitochondrijų fermentinių sistemų.
Vidinis mitochondrijų spindis vadinamas matrica nuo citoplazmos atskirtas dviem membranomis lauke Ir vidinis, tarp kurių yra tarpmembraninė erdvė. Vidinė mitochondrijų membrana formuoja raukšles, vadinamąsias cristae. Matricoje yra įvairių fermentų, dalyvaujančių kvėpavime ir ATP sintezėje. Vidinės mitochondrijų membranos vandenilio potencialas yra labai svarbus ATP sintezei.
Mitochondrijos turi savo DNR genomą ir prokariotines ribosomas, o tai neabejotinai rodo šių organelių simbiotinę kilmę. Ne visi mitochondrijų baltymai yra užkoduoti mitochondrijų DNR, dauguma mitochondrijų baltymų genai išsidėstę branduoliniame genome, o atitinkami jų produktai sintetinami citoplazmoje ir vėliau transportuojami į mitochondrijas. Mitochondrijų genomai skiriasi dydžiu: pavyzdžiui, žmogaus mitochondrijų genome yra tik 13 genų. Dauguma didelis skaičius mitochondrijų genai (97) iš tirtų organizmų turi paprasčiausią Amerikos reclinomonas.
1.3. Pro- ir eukariotinių ląstelių palyginimas
Ilgą laiką svarbiausias skirtumas tarp eukariotų ir prokariotų buvo gerai suformuoto branduolio ir membraninių organelių buvimas. Tačiau iki 1970 ir 1980 m tapo aišku, kad tai tik gilesnių citoskeleto organizavimo skirtumų pasekmė. Kurį laiką buvo manoma, kad citoskeletas būdingas tik eukariotams, tačiau 1990 m. bakterijose taip pat buvo rasta baltymų, homologiškų pagrindiniams eukariotų citoskeleto baltymams.
Būtent specialiai išdėstyto citoskeleto buvimas leidžia eukariotams sukurti judrių vidinių membraninių organelių sistemą. Be to, citoskeletas leidžia atlikti endo- ir egzocitozę (manoma, kad būtent dėl endocitozės eukariotinėse ląstelėse atsirado tarpląsteliniai simbiontai, įskaitant mitochondrijas ir plastidus). Kita svarbi eukariotų citoskeleto funkcija – užtikrinti eukariotinės ląstelės branduolio (mitozės ir mejozės) ir kūno (citotomijos) dalijimąsi (prokariotinių ląstelių dalijimasis organizuojamas paprasčiau). Citoskeleto struktūros skirtumai paaiškina ir kitus skirtumus tarp pro- ir eukariotų – pavyzdžiui, prokariotinių ląstelių formų pastovumą ir paprastumą bei didelę eukariotų formų įvairovę ir gebėjimą ją keisti, taip pat palyginti didelis pastarųjų dydis. Taigi prokariotinių ląstelių dydis yra vidutiniškai 0,5-5 mikronai, eukariotinių ląstelių matmenys yra vidutiniškai nuo 10 iki 50 mikronų. Be to, tik tarp eukariotų yra tikrai gigantiškų ląstelių, tokių kaip masyvūs ryklių ar stručių kiaušinėliai (paukščio kiaušinyje visas trynys yra vienas didžiulis kiaušinis), stambių žinduolių neuronai, kurių procesus sustiprina citoskeletas, gali siekti keliasdešimt centimetrų ilgio.
1.4. Anaplazija
Ląstelių struktūros sunaikinimas (pavyzdžiui, su piktybiniai navikai) vadinama anaplazija.
2. Ląstelių atradimo istorija
Pirmasis žmogus, pamatęs ląsteles, buvo anglų mokslininkas Robertas Hukas (mums žinomas Huko dėsnio dėka). 1665 m., bandydamas suprasti, kodėl kamštienos medis taip gerai plūduriuoja, Hukas pradėjo tirti plonas kamštienos dalis naudodamas savo patobulintą mikroskopą. Jis išsiaiškino, kad kamštiena buvo padalinta į daugybę mažyčių celių, kurios jam priminė vienuolines ląsteles, ir pavadino šias ląsteles (anglų kalba cell reiškia „ląstelė, ląstelė, ląstelė“). 1675 metais italų gydytojas M. Malpighi, o 1682 metais anglų botanikas N. Gru patvirtino augalų ląstelinę sandarą. Jie pradėjo kalbėti apie ląstelę kaip apie „maistingų sulčių pripildytą burbulą“. 1674 metais olandų meistras Antony van Leeuwenhoek (Anton van Leeuwenhoek, 1632-1723), naudodamas mikroskopą, pirmą kartą vandens laše pamatė „gyvūnus“ – judančius gyvus organizmus (blakstienas, amebas, bakterijas). Leeuwenhoekas taip pat pirmą kartą stebėjo gyvūnų ląsteles – eritrocitus ir spermatozoidus. Taigi iki XVIII amžiaus pradžios mokslininkai žinojo, kad esant dideliam padidinimui augalai turi ląstelinę struktūrą, ir pamatė kai kuriuos organizmus, kurie vėliau buvo pavadinti vienaląsčiais. 1802–1808 m. prancūzų tyrinėtojas Charlesas-Francois Mirbelis nustatė, kad visi augalai susideda iš audinių, formuojasi ląstelės. J. B. Lamarkas 1809 metais paskleidė Mirbelio idėją ląstelių struktūra ir ant gyvūnų organizmų. 1825 metais čekų mokslininkė J. Purkynė atrado paukščių kiaušinėlio ląstelės branduolį, o 1839 metais įvedė terminą „protoplazma“. 1831 metais anglų botanikas R. Brownas pirmą kartą aprašė augalo ląstelės branduolį, o 1833 metais nustatė, kad branduolys yra esminė augalo ląstelės organelė. Nuo tada pagrindinis dalykas organizuojant ląsteles yra ne membrana, o turinys.
Ląstelinę organizmų sandaros teoriją 1839 m. sukūrė vokiečių zoologai T. Schwann ir M. Schleiden ir apėmė tris nuostatas. 1858 m. Rudolfas Virchovas ją papildė dar viena nuostata, tačiau jo idėjose buvo nemažai klaidų: pavyzdžiui, jis manė, kad ląstelės yra silpnai sujungtos viena su kita ir kiekviena egzistuoja „savaime“. Tik vėliau pavyko įrodyti ląstelių sistemos vientisumą.
1878 m. rusų mokslininkas ID Chistyakovas atrado mitozę augalų ląstelėse; 1878 metais W. Flemmingas ir P. I. Peremezhko atrado mitozę gyvūnams. 1882 metais W. Flemmingas stebėjo mejozę gyvūnų ląstelėse, o 1888 metais E. Strasburgeris – augalų ląstelėse.
3. Ląstelės cheminė sudėtis
1 grupė (iki 98%) (makroelementai)
- Anglies
- Vandenilis
- Deguonis
2 grupė (1,5–2 proc.)
- Kalis
- Natrio
- Kalcis
- Magnis
- Fosforas
- Geležis
3 grupė (>0,01 %) (mikroelementai)
- Kobaltas
- Molibdenas
4 grupė (>0,00001 %) (ultra mikroelementai)
- Radis
- Auksas
Ši santrauka parengta remiantis straipsniu iš rusiškos Vikipedijos. Sinchronizavimas baigtas 07/09/11 23:16:18
Ląstelė – struktūrinė ir funkcinis vienetas kuria grindžiama visų gyvų organizmų struktūra, vystymasis ir gyvenimas.
Ląstelės egzistuoja kaip savarankiški organizmai – vienaląsčiai gyvūnai ir augalai – arba kaip elementarios dalys daugialąstelis organizmas- audinių ląstelės.
Vienų ląstelių forma, pavyzdžiui, amebos arba, yra kintama, kitų forma yra daugiau ar mažiau pastovi ir būdinga kiekvienam ląstelių tipui – blakstienoms, nervinėms ląstelėms, daugumai. augalų ląstelės ir kt.
Ląstelių dydžiai labai įvairūs, dauguma jų matomi tik pro mikroskopą. Mažiausių ląstelių skersmuo yra apie 4 mikronai.
Ląstelę nuo aplinkos riboja plona ląstelės membrana, kurios storis ir struktūra žaidžia svarbus vaidmuo reguliuojant ląstelių metabolizmą. Membrana dažniausiai yra trijų sluoksnių, susideda iš baltymų ir lipoidinių medžiagų.
Kiekvienos ląstelės gyvavimo ciklas susideda iš dviejų periodų: tarpfazės (laikotarpis tarp dviejų dalijimosi) ir paties dalijimosi (žr. Mitozė, Mejozė).
Trukmė gyvenimo ciklas ląstelės skirtingi tipai labai skiriasi. Pavyzdžiui, tarpfazė nervų ląstelės trunka visą organizmo gyvenimą, o epitelio ląstelės dalijasi labai greitai.
Ląstelė susideda iš dviejų pagrindinių dalių (1 ir 2 pav.) – citoplazmos (žr.) ir branduolio (žr. Ląstelės branduolys), kurios negali egzistuoti atskirai, nes yra nuolatinėje sąveikoje ir tarpusavio priklausomybėje.
Ryžiai. 1. Ląstelės sandaros schema (mikroskopija): 1 - branduolys; 2 - šerdis; 3 -; 4 - vakuolė; 5 - citoplazma; 6 - centrosoma.
Ryžiai. 2. Ląstelės sandaros schema (): 1 - citoplazma; 2 - šerdis; 3 - branduolys; 4 - ląstelės membrana.
Ląstelių intarpai priklauso nuo funkcinė būklė ląstelės ir skiriasi tiek savo cheminė prigimtis(baltymai, riebalai, angliavandeniai ir kt.), o pagal tankį (granulės ir intarpai su skystu kiekiu – vakuolės).
Branduolys yra antrasis pagrindinis ląstelės komponentas. Per gyvavimo ciklą branduolys patiria daugybę sudėtingų pokyčių. Branduolio chromatino komponentuose yra, kurio citoplazmoje nėra. Branduolys turi reguliuojančią įtaką ląstelės vystymuisi ir yra jos paveldimų savybių nešėjas.
Augalinės ląstelės nuo gyvūnų ląstelių skiriasi gana storu apvalkalu, kuris aiškiai matomas šviesos mikroskopu. Daugelio augalų ląstelių citoplazmoje yra specialių organelių – plastidžių: chloroplastų, kurie vykdo fotosintezę; chromoplastai, turintys geltonų ir raudonų pigmentų (karotinoidų); bespalviai leukoplastai ir apvalūs, stipriai šviesą laužantys kūnai – sferosomos. Augalų ląstelių citoplazmoje yra daugiau ar mažiau išsivysčiusi vakuolių sistema.
kaip ląstelė gyvoji sistema geba išlaikyti ir išlaikyti savo specifinę struktūrą dėl nuolatinio energijos ir iš aplinkos gaunamų medžiagų vartojimo. Metaboliniai galutiniai produktai išsiskiria į aplinką. Kiekviena ląstelė, pasiekusi tam tikrą vystymosi stadiją, dalijasi į dvi dukterines ląsteles. Dalijimasis vyksta mitoze, rečiau – amitoze, pirmuoju atveju įvyksta kompleksinis branduolio ir citoplazmos persitvarkymas.
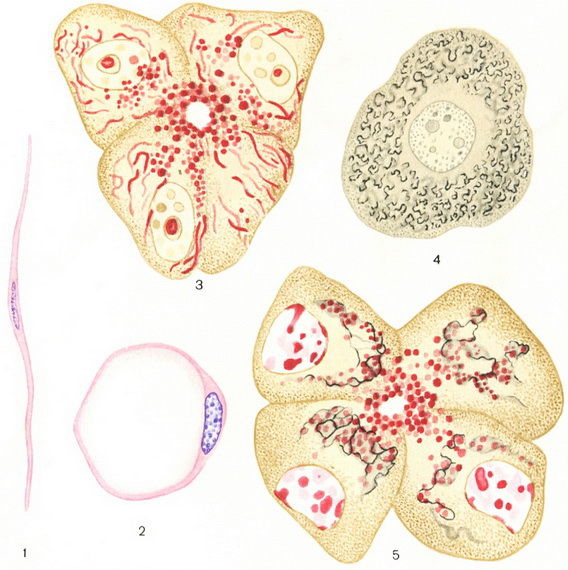
Ryžiai. 1. Lygus raumenų ląstelėžarnynas. Ryžiai. 2. Riebalų ląstelė iš poodinis audinysžiurkės. Ryžiai. 3. Kasos ląstelių mitochondrijos ir sekrecinės granulės. Ryžiai. 4. Golgi aparatas stuburo ganglijos jutiminėje ląstelėje. Ryžiai. 5. Golgi aparatas ir sekrecinės granulės kasos ląstelėse. 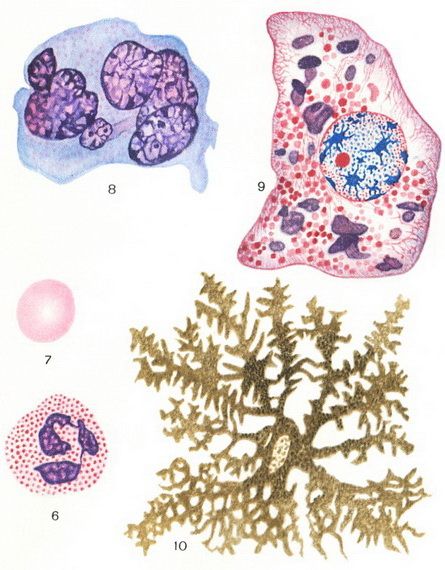
Ryžiai. 6. Neutrofilinis leukocitas. Ryžiai. 7. Žmogaus eritrocitas. Ryžiai. 8. Megakariocitas iš kaulų čiulpai asmuo. Ryžiai. 9. Aksolotlio kepenų ląstelė. Citoplazmoje - raudonos chondriosomos ir purpuriniai baltymų intarpai; branduolyje – raudonas oksifilinis branduolys ir bazofiliniai chromatino gumulėliai (stambesni gumulėliai – kariosomos). Ryžiai. 10. Axolotl chromatoforas, užpildytas pigmento granulėmis.