^ plazmos membrana , arba plazmolema, užima ypatingą vietą tarp įvairių ląstelių membranų. Tai paviršinė periferinė struktūra, ribojanti ląstelę iš išorės, kuri lemia tiesioginį jos ryšį su ekstraląsteline aplinka, taigi ir su visomis ląstelę veikiančiomis medžiagomis ir dirgikliais. Todėl plazminės membranos vaidmuo yra būti barjeru, barjeru tarp sudėtingai organizuoto tarpląstelinio turinio ir išorinės aplinkos. Šiuo atveju plazmolema atlieka ne tik mechaninio barjero vaidmenį, bet, svarbiausia, riboja laisvą mažos ir didelės molekulinės masės medžiagų srautą abiem kryptimis per membraną. Be to, plazmolema veikia kaip struktūra, kuri „atpažįsta“, receptorius, įvairias chemines medžiagas ir selektyviai reguliuoja šių medžiagų transportavimą į ląstelę ir iš jos. Kitaip tariant, plazminė membrana atlieka funkcijas, susijusias su reguliuojamu selektyviu transmembraniniu medžiagų transportavimu, ir veikia kaip pirminis ląstelių analizatorius. Šiuo atžvilgiu galima apsvarstyti plazmalemą ląstelės organoidasįtrauktas į ląstelės vakuolinę sistemą. Kaip ir kitos šios sistemos membranos (lizosomų, endosomų membranos, Golgi aparatas ir kt.), Ji atsiranda ir atnaujinama dėl endoplazminio tinklo sintetinio aktyvumo ir yra panašios sudėties. Kaip bebūtų keista, plazminę membraną galima prilyginti intraląstelinės vakuolės membranai, bet apversti iš vidaus: ji nėra apsupta hialoplazmos, o ją supa.
Plazmalemmos barjerinis transportavimo vaidmuo
Ląstelę iš visų pusių supanti plazminė membrana veikia kaip mechaninis barjeras. Norint jį pradurti mikroadatomis ar mikropipetėmis, reikia įdėti nemažai pastangų. Paspaudus ant jos mikroadatą, ji iš pradžių stipriai išsilenkia, o tik tada prasilaužia. Dirbtinės lipidinės membranos yra mažiau stabilios. Šį mechaninį plazminės membranos stabilumą gali lemti papildomi komponentai, tokie kaip glikokaliksas ir citoplazmos žievės sluoksnis (127 pav.).
Glikokaliksas yra lipoproteininės membranos išorinis sluoksnis, kuriame yra membranos vientisų baltymų – glikoproteinų – polisacharidinės grandinės. Šiose grandinėse yra tokių angliavandenių kaip manozė, gliukozė, N-acetilgliukozaminas, sialo rūgštis ir kt. Tokie angliavandenių heteropolimerai sudaro išsišakojusias grandines, tarp kurių gali išsidėstyti iš ląstelės išskirti glikolipidai ir proteoglikanai. Glikokalikso sluoksnis yra stipriai laistomas, turi želė konsistenciją, kuri žymiai sumažina difuzijos greitį šioje zonoje. įvairių medžiagų. Čia taip pat gali „užstrigti“ ląstelės išskiriami hidroliziniai fermentai, kurie dalyvauja ekstraląsteliniame polimerų skilime (ekstraląsteliniame virškinime) iki monomerų molekulių, kurios vėliau per plazmos membraną transportuojamos į citoplazmą.
IN elektroninis mikroskopas, ypač kai specialius metodus kontrastingi polisacharidai, glikokaliksas yra laisvo pluoštinio 3-4 nm storio sluoksnio, dengiančio visą ląstelės paviršių. Glikokaliksas ypač gerai išreikštas sugeriamojo žarnyno epitelio ląstelių (enterocitų) šepetėlio pakraštyje, tačiau randamas beveik visose gyvūnų ląstelėse, tačiau jos sunkumo laipsnis skiriasi (128 pav.).
Be to, plazminės membranos mechaninį stabilumą užtikrina šalia jos esančio žievės sluoksnio struktūra iš citoplazmos pusės ir tarpląstelinės fibrilinės struktūros. žievės(nuo žodžio - žievė - žievė, žievelė) sluoksnis citoplazma, glaudžiai susijusi su lipoproteinu išorinė membrana, turi daugybę funkcijų. Čia, 0,1-0,5 mikrono storio, nėra ribosomų ir membraninių pūslelių, tačiau dideliais kiekiais yra fibriliniai citoplazmos elementai – mikrofilamentai ir dažnai mikrovamzdeliai. Pagrindinis fibrilinis žievės sluoksnio komponentas yra aktino mikrofibrilių tinklas. Čia taip pat yra nemažai pagalbinių baltymų, reikalingų citoplazmos sekcijų judėjimui (daugiau informacijos apie ląstelių skeleto-motorinę sistemą žr. toliau). Šių su aktinu susijusių baltymų vaidmuo yra labai svarbus, nes tai paaiškina jų dalyvavimą plazmos membranos integralinių baltymų jungtyje, „inkaravime“.
Daugelio pirmuonių, ypač blakstienų, formavime dalyvauja plazminė membrana granulės, standus sluoksnis, kuris dažnai lemia ląstelės formą. Membraniniai maišeliai čia gali priglusti prie plazminės membranos iš vidaus; šiuo atveju šalia ląstelių paviršiaus yra trys membranos sluoksniai: pati plazminė membrana ir dvi pelikulinių alveolių membranos. Bato blakstienose sruogelė formuoja sustorėjimus, išsidėsčiusius šešiakampių pavidalu, kurių centre iškyla blakstienos (129 pav.). Pelikulinių formacijų standumas taip pat gali būti siejamas su citoplazmos elementais, esančiais po plazmos membrana, su žievės sluoksniu. Taigi prie membranos esančiose eugleno pelikulo keterose, be membraninių vakuolių, randama lygiagrečių mikrovamzdelių ir mikrofilamentų pluoštų. Šis fibrilinis periferinis sutvirtinimas kartu su sulankstyta daugiasluoksne membranos periferija sukuria standžią granulės struktūrą.
Plasmalemmos barjerinis vaidmuo taip pat yra laisvos medžiagų difuzijos ribojimas. Modeliniai eksperimentai su dirbtinėmis lipidinėmis membranomis parodė, kad jos yra laidžios vandeniui, dujoms, mažoms nepolinėms riebaluose tirpių medžiagų molekulėms, tačiau visiškai nepralaidžios įkrautoms molekulėms (jonams) ir didelėms neįkrautoms (cukrui) (130 pav.).
Natūralios membranos taip pat riboja mažos molekulinės masės junginių įsiskverbimo į ląstelę greitį.
Transmembraninis jonų ir mažos molekulinės masės junginių pernešimas
Plazminė membrana, kaip ir kitos lipoproteininės ląstelių membranos, yra pusiau laidi. Tai reiškia, kad skirtingos molekulės per ją praeina skirtingu greičiu ir didesnio dydžio molekulių, tuo lėčiau jos praeina per membraną. Ši savybė apibrėžia plazmos membraną kaip osmosinį barjerą. Didžiausia prasiskverbimo galia pasižymi vanduo ir jame ištirpusios dujos, jonai į membraną prasiskverbia daug lėčiau (apie 10 4 kartus lėčiau). Todėl, jei ląstelė, pavyzdžiui, eritrocitas, patalpinama į aplinką, kurioje druskų koncentracija mažesnė nei ląstelėje (hipotenzija), vanduo iš išorės pateks į ląstelę, o tai padidins ląstelės tūris ir plazminės membranos plyšimas ("hipotoninis šokas"). Priešingai, kai eritrocitas dedamas į druskos tirpalus, daugiau didelė koncentracija nei ląstelėje, bus vandens išėjimas iš ląstelės į išorinę aplinką. Tuo pačiu metu ląstelė susiraukšlės, sumažės tūris.
Toks pasyvus vandens transportavimas iš ląstelės ir į ląstelę vis dar vyksta nedideliu greičiu. Vandens prasiskverbimo pro membraną greitis yra apie 10 -4 cm/s, tai yra 100 000 kartų mažesnis nei vandens molekulių difuzijos greitis per 7,5 nm storio vandeninį sluoksnį. Buvo padaryta išvada, kad m ląstelės membrana, jo lipoproteininiame sluoksnyje yra specialios „poros“, skirtos vandeniui ir jonams prasiskverbti. Jų skaičius nėra toks didelis: bendras plotas, kurio vienos „poros“ dydis yra apie 0,3–0,8 nm, turėtų sudaryti tik 0,06% viso ląstelės paviršiaus.
Skirtingai nuo dirbtinių dvisluoksnių lipidų membranų, natūralios membranos ir pirmiausia plazminė membrana vis dar gali pernešti jonus ir daug monomerų, tokių kaip cukrus, aminorūgštys ir kt. Jonų pralaidumas yra mažas, o skirtingų jonų pralaidumas yra mažesnis. ne tas pats. Didesnis pralaidumas katijonams (K +, Na +) ir daug mažesnis anijonams (Cl -).
Jonų pernešimas per plazmalemą vyksta dėl dalyvavimo šiame membranos transportavimo baltymų – permeazių – procese. Šie baltymai gali transportuoti vieną medžiagą viena kryptimi (uniport) arba kelias medžiagas vienu metu (symport), arba kartu su vienos medžiagos importu pašalinti iš ląstelės kitą (antiport). Taigi, pavyzdžiui, gliukozė gali patekti į ląsteles kartu su Na + jonu.
Gali vykti jonų pernešimas išilgai koncentracijos gradiento - pasyviai be papildomų energijos sąnaudų. Pavyzdžiui, Na + jonas į ląstelę patenka iš išorinė aplinka kur jo koncentracija didesnė nei citoplazmoje. Pasyviojo pernešimo atveju kai kurie membraniniai transportavimo baltymai sudaro molekulinius kompleksus, kanalai per kurią ištirpusių medžiagų molekulės praeina pro membraną paprastos difuzijos būdu pagal koncentracijos gradientą. Kai kurie iš šių kanalų yra atviri visam laikui, o kita dalis gali užsidaryti arba atsidaryti reaguodama į prisijungimą prie signalinių molekulių arba į ląstelėje esančių jonų koncentracijos pokyčius. Kitais atvejais specialūs membraniniai baltymai - vežėjai selektyviai jungiasi prie vieno ar kito jono ir perneša jį per membraną (palengvinta difuzija) (131 pav.).
Atrodytų, kad tokių baltymų transportavimo kanalų ir nešėjų buvimas turėtų lemti jonų ir mažos molekulinės masės medžiagų koncentracijų pusiausvyrą abiejose membranos pusėse. Tiesą sakant, taip nėra: jonų koncentracija ląstelių citoplazmoje smarkiai skiriasi ne tik nuo išorinės aplinkos, bet net nuo kraujo plazmos, kuri maudosi ląsteles gyvūno kūne. Ant stalo 14 parodyta jonų koncentracija ląstelės viduje ir išorėje.
14 lentelė
| Ir jis | Intraląstelinė koncentracija, mM | Ekstraląstelinė koncentracija, mM |
| Na+ | 5-15 | 145 |
| K+ | 140 | 5 |
| Mg2+ | 30 | 1-2 |
| *Ca2+ | 1-2 | 2,5-5 |
| Cl- | 4 | 110 |
*Ca 2+ koncentracija laisvoje būsenoje eukariotinių ląstelių citozolyje yra 10 -7 M, o išorėje - 10 -3 M.
Kaip matyti, šiuo atveju bendra vienavalenčių katijonų koncentracija tiek ląstelių viduje, tiek išorėje yra beveik vienoda (150 mM), izotoninis. Bet pasirodo, kad citoplazmoje K + koncentracija yra beveik 50 kartų didesnė, o Na + mažesnė nei kraujo plazmoje. Be to, šis skirtumas išlaikomas tik gyvoje ląstelėje: jei ląstelė žūsta arba joje vykstantys medžiagų apykaitos procesai slopinami, tai po kurio laiko joniniai skirtumai abiejose plazminės membranos pusėse išnyks. Galite tiesiog atvėsinti ląsteles iki +2 0 C, o po kurio laiko K + ir Na + koncentracija abiejose membranos pusėse taps vienoda. Kai ląstelės pašildomos, šis skirtumas atsistato. Šis reiškinys atsiranda dėl to, kad ląstelėse yra membraninių baltymų nešėjų, kurie veikia prieš koncentracijos gradientą, o energiją eikvoja dėl ATP hidrolizės. Toks darbas vadinamas aktyvus transportas, ir tai atliekama baltymų pagalba jonų siurbliai. Plazminėje membranoje yra dviejų subvienetų molekulės (K + + Na +) siurblys, kuris taip pat yra ATPazė. Veikimo metu šis siurblys per vieną ciklą išpumpuoja 3 Na + jonus ir į ląstelę pumpuoja 2 K + jonus prieš koncentracijos gradientą. Šiuo atveju išeikvojama viena ATP molekulė, kuri pereina į ATPazės fosforilinimą, ko pasekoje Na + pernešamas per membraną iš ląstelės, o K + gauna galimybę prisijungti prie baltymo molekulės ir tada perkeliama į ląstelė (132 pav.). Dėl aktyvaus transportavimo membraninių siurblių pagalba ląstelėje reguliuojama ir dvivalenčių katijonų Mg 2+ ir Ca 2+ koncentracija, taip pat vartojant ATP.
Toks Darbas visu etatu permeazė ir siurbliai sukuria pastovią jonų ir mažos molekulinės masės medžiagų koncentraciją ląstelėje, sukuria vadinamąją. homeostazė, koncentracijų pastovumas osmosiškai veikliosios medžiagos. Reikėtų pažymėti, kad maždaug 80% viso ląstelės ATP išleidžiama homeostazei palaikyti.
Kartu su aktyviu jonų pernešimu per plazmos membraną pernešami įvairūs cukrūs, nukleotidai ir aminorūgštys.
Taigi nuo (K + + Na +) siurblio aktyvumo priklausys aktyvus gliukozės transportavimas, kuris simportiškai (vienu metu) patenka į ląstelę kartu su pasyviai pernešamo Na + jono srautu. Jei šis (K + -Na +) siurblys bus užblokuotas, netrukus Na + koncentracijos skirtumas abiejose membranos pusėse išnyks, o Na + difuzija į ląstelę sumažės, o tuo pačiu gliukozės srautas į ląstelę sustos. Kai tik atstatomas (K + -Na +)-ATPazės darbas ir susidaro jonų koncentracijos skirtumas, iš karto padidėja difuzinis Na + srautas ir tuo pačiu gliukozės pernešimas. Panašiai per membraną ir aminorūgščių srautą, kurias perneša specialūs baltymai-nešėjai, kurie veikia kaip simpporto sistemos, kartu pernešdamos jonus.
Aktyvus cukrų ir aminorūgščių pernešimas bakterijų ląstelėse vyksta dėl vandenilio jonų gradiento.
Pats savaime specialių membraninių baltymų, dalyvaujančių pasyviame arba aktyviame mažos molekulinės masės junginių pernešime, dalyvavimas rodo didelį šio proceso specifiškumą. Net ir pasyviojo jonų transportavimo atveju baltymai „atpažįsta“ duotą joną, sąveikauja su juo, specifiškai jungiasi, keičia savo konformaciją ir funkcionuoja. Todėl jau transporto pavyzdžiu paprastos medžiagos membranos veikia kaip analizatoriai, kaip receptoriai. Šis receptoriaus vaidmuo ypač pasireiškia, kai ląstelė absorbuoja biopolimerus.
Vezikulinis transportas: endocitozė ir egzocitozė
Makromolekulės, tokios kaip baltymai, nukleorūgštys, polisacharidai, lipoproteinų kompleksai ir kt., neprasiskverbia pro ląstelių membranas, priešingai nei pernešami jonai ir monomerai. Mikromolekulių, jų kompleksų, dalelių pernešimas į ląstelę ir iš jos vyksta visai kitaip – vezikuliniu pernešimu. Šis terminas reiškia, kad įvairios makromolekulės, biopolimerai ar jų kompleksai negali patekti į ląstelę per plazminę membraną. Ir ne tik per jį: jokios ląstelių membranos nepajėgios pernešti biopolimerų per membraną, išskyrus membranas, turinčias specialių baltymų komplekso nešėjų – porinų (mitochondrijų, plastidų, peroksisomų membranos). Makromolekulės patenka į ląstelę arba iš vienos membranos skyriaus į kitą, uždarytos vakuolėse ar pūslelėse. Toks vezikulinis pernešimas galima suskirstyti į du tipus: egzocitozė- stambiamolekulinių produktų pašalinimas iš ląstelės ir endocitozė- makromolekulių absorbcija ląstelėje (133 pav.).
Endocitozės metu tam tikra plazmalemos dalis tarsi apgaubia ekstraląstelinę medžiagą, uždaro ją į membranos vakuolę, atsiradusią dėl plazminės membranos invaginacijos. Tokioje pirminėje vakuolėje arba in endosomas, gali patekti bet kokie biopolimerai, stambiamolekuliniai kompleksai, ląstelių dalys ar net visos ląstelės, kur vėliau suyra, depolimerizuojasi iki monomerų, kurie transmembraniniu būdu patenka į hialoplazmą. Pagrindinė endocitozės biologinė reikšmė yra statybinių blokų įsigijimas per intracelulinis virškinimas, kuri atliekama antroje endocitozės stadijoje po pirminės endosomos suliejimo su lizosoma, vakuole, kurioje yra hidrolizinių fermentų rinkinys (žr. toliau).
Endocitozė formaliai skirstoma į pinocitozė Ir fagocitozė(134 pav.). Fagocitozę - didelių dalelių (kartais net ląstelių ar jų dalių) gaudymą ir absorbciją - pirmą kartą aprašė I. I. Mechnikovas. Fagocitozė, gebėjimas užfiksuoti dideles daleles ląstele, randama tarp gyvūnų ląstelių, tiek vienaląsčių (pavyzdžiui, amebų, kai kurių plėšriųjų blakstienų), tiek specializuotose daugialąsčių gyvūnų ląstelėse. Specializuotos ląstelės – fagocitai – būdingos tiek bestuburiams (kraujo ar ertmės skysčio amebocitams), tiek stuburiniams gyvūnams (neutrofilams ir makrofagams). Pinocitozė iš pradžių buvo apibrėžta kaip vandens absorbcija arba vandeniniai tirpalai skirtingos medžiagos. Dabar žinoma, kad ir fagocitozė, ir pinocitozė vyksta labai panašiai, todėl šių terminų vartojimas gali atspindėti tik absorbuotų medžiagų tūrio ir masės skirtumus. Šie procesai turi bendrą tai, kad absorbuotos medžiagos plazminės membranos paviršiuje yra apsuptos vakuolės pavidalo membrana – endosoma, kuri juda ląstelės viduje.
Endocitozė, įskaitant pinocitozę ir fagocitozę, gali būti nespecifinė arba konstitucinė, nuolatinė ir specifinė, tarpininkaujama receptorių (receptorių). Nespecifinis endocitas h (pinocitozė ir fagocitozė), taip vadinama, nes ji vyksta tarsi automatiškai ir dažnai gali lemti ląstelei visiškai svetimų arba abejingų medžiagų, pavyzdžiui, suodžių ar dažų dalelių, paėmimą ir absorbciją.
Nespecifinę endocitozę dažnai lydi pradinė sulaikančios medžiagos sorbcija plazmos membranos glikokaliksu. Glikokaliksas dėl savo polisacharidų rūgščių grupių turi neigiamą krūvį ir gerai jungiasi su įvairiomis teigiamai įkrautomis baltymų grupėmis. Esant tokiai adsorbcijai, absorbuojama nespecifinė endocitozė, makromolekulės ir mažos dalelės (rūgštiniai baltymai, feritinas, antikūnai, virionai, koloidinės dalelės). Skystos fazės pinocitozė sukelia tirpių molekulių, kurios nesijungia su plazmalema, absorbciją kartu su skysta terpe.
Įjungta Kitas žingsnis pasikeičia ląstelės paviršiaus morfologija: tai arba mažų plazminės membranos invaginacijų atsiradimas, invaginacija, arba ataugų, raukšlių ar „maivumo“ (rafl - anglų k.) atsiradimas ant ląstelės paviršiaus. , kurie tarsi persidengia, susilanksto, atskirdami nedidelius skystos terpės tūrius (135, 136 pav.). Pirmasis pinocitinės pūslelės, pinosomų, atsiradimo tipas būdingas žarnyno epitelio, endotelio, amebų ląstelėms, antrasis - fagocitams ir fibroblastams. Šie procesai priklauso nuo energijos tiekimo: kvėpavimo inhibitoriai blokuoja šiuos procesus.
Po šio paviršiaus pertvarkymo seka besiliečiančių membranų sukibimo ir susiliejimo procesas, dėl kurio susidaro penicitinė pūslelė (pinosoma), kuri atsiskiria nuo ląstelės paviršiaus ir patenka gilyn į citoplazmą. Tiek nespecifinė, tiek receptorinė endocitozė, sukelianti membraninių pūslelių skilimą, atsiranda specializuotuose plazmos membranos regionuose. Tai vadinamieji ribojamos duobės. Taip jie vadinami todėl, kad iš citoplazmos pusės plazminė membrana yra padengta, aptraukta plonu (apie 20 nm) pluoštiniu sluoksniu, kuris ant itin plonų pjūvių tarsi ribojasi, dengia nedidelius išsikišimus, duobutes (1 pav.). 137). Beveik visos gyvūnų ląstelės turi šias duobutes; jos užima apie 2% ląstelės paviršiaus. Aplinkinis sluoksnis daugiausia sudarytas iš baltymų klatrinas susijęs su daugybe papildomų baltymų. Trys klatrino molekulės kartu su trimis mažos molekulinės masės baltymo molekulėmis sudaro triskeliono struktūrą, panašią į trijų spindulių svastiką (138 pav.). Plazminės membranos duobučių vidinio paviršiaus klatrino triskelionai sudaro laisvą tinklą, susidedantį iš penkiakampių ir šešiakampių, paprastai primenančių krepšį. Klatrino sluoksnis dengia visą besiskiriančių pirminių endocitinių vakuolių, besiribojančių su pūslelėmis, perimetrą.
Klatrinas priklauso vienai iš vadinamųjų rūšių. „tvarsčių“ baltymai (COP – coated proteins). Šie baltymai jungiasi prie integruotų receptorių baltymų iš citoplazmos pusės ir aplink besiformuojančios pinosomos perimetrą sudaro tvarsliavą, pirminę endosominę pūslelę - „ribinę“ pūslelę. pirminės endosomos atskyrime dalyvauja ir baltymai - dinaminai, kurie polimerizuojasi aplink besiskiriančios pūslelės kaklelį (139 pav.).
Apribotai pūslelei atsiskyrus nuo plazmolemos ir pradėjus gilintis į citoplazmą, klatrino sluoksnis suyra, disocijuoja, endosomų membrana (pinosomos) įgauna įprastą formą. Praradus klatrino sluoksnį, endosomos pradeda jungtis viena su kita.
Nustatyta, kad ribojamų duobių membranose yra palyginti mažai cholesterolio, kuris gali lemti membranos standumo sumažėjimą ir prisidėti prie burbuliukų susidarymo. Biologinė klatrino „sluoksnio“ atsiradimo išilgai pūslelių periferijos prasmė gali būti ta, kad jis užtikrina besiribojančių pūslelių sukibimą su citoskeleto elementais ir jų tolesnį transportavimą į ląstelę ir neleidžia joms susilieti. .
Skystos fazės nespecifinės pinocitozės intensyvumas gali būti labai didelis. Taigi epitelio ląstelė plonoji žarna per sekundę sudaro iki 1000 pinosomų, o makrofagai – apie 125 pinosomas per minutę. Pinosomų dydis yra mažas, apatinė riba yra 60–130 nm, tačiau jų gausa lemia tai, kad endocitozės metu plazmolema greitai pakeičiama, tarsi „išeikvojama“ daugybei mažų vakuolių susidarymo. Taigi makrofaguose visa plazminė membrana pakeičiama per 30 minučių, fibroblastuose – per dvi valandas.
Tolesnis endosomų likimas gali būti kitoks, dalis jų gali grįžti į ląstelės paviršių ir su juo susilieti, bet dauguma patenka į tarpląstelinio virškinimo procesą. Pirminėse endosomose daugiausia yra svetimų molekulių, įstrigusių skystoje terpėje, ir jose nėra hidrolizinių fermentų. endosomos gali susilieti viena su kita, didindamos dydį. Tada jie susilieja su pirminėmis lizosomomis (žr. toliau), kurios į endosomų ertmę įveda fermentus, kurie hidrolizuoja įvairius biopolimerus. Šių lizosomų hidrolazių veikimas sukelia tarpląstelinį virškinimą – polimerų suskaidymą iki monomerų.
Kaip jau minėta, fagocitozės ir pinocitozės metu ląstelės praranda didelį plazmolemos plotą (žr. makrofagus), tačiau membranos perdirbimo metu greitai atsistato dėl vakuolių grįžimo ir jų įtraukimo į plazmolemą. Taip yra dėl to, kad mažos pūslelės gali atsiskirti nuo endosomų ar vakuolių, taip pat nuo lizosomų, kurios vėl susilieja su plazmine membrana. Su tokiu perdirbimu įvyksta savotiškas membranų pernešimas: plazmolema - pinosoma - vakuolė - plazmolemma. Tai veda prie pradinės plazmos membranos srities atkūrimo. Nustatyta, kad su tokiu grįžimu, membranos perdirbimu, visa absorbuota medžiaga išlieka likusioje endosomoje.
Specifinis arba tarpininkauja receptoriams endocitozė turi daug skirtumų nuo nespecifinės. Svarbiausia, kad būtų absorbuojamos molekulės, kurioms plazmos membranoje yra specifiniai receptoriai, susiję tik su tokio tipo molekulėmis. Dažnai vadinamos tokios molekulės, kurios jungiasi prie receptorių baltymų ląstelių paviršiuje ligandai.
Receptorių sukelta endocitozė pirmą kartą buvo aprašyta baltymų kaupimuisi paukščių oocituose. Jame sintetinami trynio granulių baltymai, vitelogeninai įvairių audinių, bet tada su kraujotaka patenka į kiaušides, kur prisijungia prie specialių membraninių kiaušialąsčių receptorių ir tada endocitozės pagalba patenka į ląstelę, kur vyksta trynio granulių nusėdimas.
Kitas selektyvios endocitozės pavyzdys yra cholesterolio pernešimas į ląstelę. Šis lipidas sintetinamas kepenyse ir kartu su kitais fosfolipidais bei baltymo molekule sudaro vadinamąjį. mažo tankio lipoproteinų (MTL), kuriuos išskiria kepenys ir kraujotakos sistema plinta visame kūne (140 pav.). Specialūs plazmos membranos receptoriai, difuziškai išsidėstę paviršiuje įvairios ląstelės, atpažįsta MTL baltyminį komponentą ir sudaro specifinį receptoriaus-ligando kompleksą. Po to toks kompleksas persikelia į ribojamų duobių zoną ir yra internalizuojamas – apsuptas membrana ir panardinamas giliai į citoplazmą. Įrodyta, kad mutantiniai receptoriai gali surišti MTL, bet nesikaupia ribojamų duobių srityje. Be MTL receptorių, buvo rasta daugiau nei dvi dešimtys kitų medžiagų, dalyvaujančių įvairių medžiagų receptorių endocitozėje, ir visos jos naudoja tą patį internalizacijos kelią per apribotas duobes. Tikėtina, kad jų vaidmuo yra receptorių kaupimas: vienoje ir toje pačioje apvaduotoje duobėje galima surinkti apie 1000 skirtingų klasių receptorių. Tačiau fibroblastuose MTL receptorių klasteriai yra ribojamų duobių zonoje, net jei terpėje nėra ligando.
Tolesnis absorbuotos MTL dalelės likimas yra tas, kad kompozicijoje jos suyra antrinė lizosoma. Panardinus į ribojančios pūslelės, pakrautos MTL, citoplazmą, greitai prarandamas klatrino sluoksnis, membranos pūslelės pradeda jungtis viena su kita, sudarydamos endosomą – vakuolę, kurioje yra absorbuotų MTL dalelių, vis dar susijusių su receptoriais membranos paviršiuje. . Tada įvyksta ligando-receptoriaus komplekso disociacija, nuo endosomos atsiskiria mažos vakuolės, kurių membranose yra laisvųjų receptorių. Šios pūslelės yra perdirbamos, įtraukiamos į plazmos membraną, todėl receptoriai grįžta į ląstelės paviršių. MTL likimas yra tas, kad po susiliejimo su lizosomomis jie hidrolizuojasi iki laisvo cholesterolio, kuris gali būti įtrauktas į ląstelių membranas.
Endosomoms būdinga žemesnė pH vertė (pH 4-5), rūgštesnė aplinka nei kitos ląstelės vakuolės. Taip yra dėl to, kad jų membranose yra baltymų. protonų siurblys, siurbiant vandenilio jonus kartu sunaudodama ATP (nuo H + priklausomą ATPazę). Rūgšti aplinka endosomose vaidina lemiamą vaidmenį disociuojant receptorius ir ligandus. Be to, rūgštinė aplinka yra optimalus lizosomų sudėtyje esančių hidrolizinių fermentų aktyvavimui, kurie aktyvuojami susiliejus lizosomoms su endosomomis ir veda prie jų susidarymo. endolizosomos, kuriame vyksta absorbuotų biopolimerų skilimas.
Kai kuriais atvejais disocijuotų ligandų likimas nėra susijęs su lizosomų hidrolize. Taigi kai kuriose ląstelėse, plazmolemos receptoriams prisijungus prie tam tikrų baltymų, klatrinu padengtos vakuolės nugrimzta į citoplazmą ir perkeliamos į kitą ląstelės sritį, kur vėl susilieja su plazmos membrana, o surišti baltymai atsiskiria nuo receptoriai. Taip vyksta kai kurių baltymų pernešimas, transcitozė per endotelio ląstelės sienelę iš kraujo plazmos į tarpląstelinę aplinką (141 pav.). Kitas transcitozės pavyzdys yra antikūnų pernešimas. Taigi žinduoliams motinos antikūnai gali būti perduodami jaunikliui per pieną. Šiuo atveju receptoriaus-antikūno kompleksas endosomoje išlieka nepakitęs.
Fagocitozė
Kaip jau minėta, fagocitozė yra endocitozės atmaina ir yra susijusi su didelių makromolekulių agregatų absorbcija ląstelėje iki gyvų ar negyvų ląstelių. Taip pat kaip pinocitozė, fagocitozė gali būti nespecifinė (pavyzdžiui, koloidinio aukso ar dekstrano polimero dalelių absorbcija fibroblastais ar makrofagais) ir specifinė, tarpininkaujama receptorių fagocitinių ląstelių plazminės membranos paviršiuje. Fagocitozės metu susidaro didelės endocitinės vakuolės - fagosoma, kurios vėliau susilieja su lizosomomis ir susidaro fagolizosomos.
Ląstelių, galinčių fagocituoti, paviršiuje (žinduoliams tai yra neutrofilai ir makrofagai) yra receptorių, kurie sąveikauja su ligandų baltymais, rinkinys. Taigi at bakterinės infekcijos prie paviršių jungiasi antikūnai prieš bakterijų baltymus bakterijų ląstelės, sudarydamas sluoksnį, kuriame antikūnų F c sritis žvelgia į išorę. Šį sluoksnį atpažįsta specifiniai makrofagų ir neutrofilų paviršiuje esantys receptoriai, o jų surišimo vietose prasideda bakterijos absorbcija, apgaubiant ją ląstelės plazmine membrana (142 pav.).
Egzocitozė
Plazminė membrana dalyvauja pašalinant medžiagas iš ląstelės, padedant egzocitozė- atvirkštinis endocitozės procesas (žr. 133 pav.).
Egzocitozės atveju intraląsteliniai produktai, uždengti vakuolėmis arba pūslelėmis ir atskirti nuo hialoplazmos membrana, artėja prie plazminės membranos. Jų sąlyčio taškuose plazminė membrana ir vakuolės membrana susilieja, o burbulas ištuštinamas į aplinką. Egzocitozės pagalba vyksta endocitozėje dalyvaujančių membranų perdirbimo procesas.
Egzocitozė yra susijusi su įvairių ląstelėje susintetintų medžiagų išsiskyrimu. Išskirdamos, išskirdamos į aplinką medžiagas, ląstelės gali gaminti ir išskirti mažos molekulinės masės junginius (acetilcholiną, biogeninius aminus ir kt.), taip pat dažniausiai makromolekules (peptidus, baltymus, lipoproteinus, peptidoglikanus ir kt.). Egzocitozė arba sekrecija daugeliu atvejų atsiranda reaguojant į išorinį signalą (nervinį impulsą, hormonus, mediatorius ir kt.). Nors kai kuriais atvejais egzocitozė vyksta nuolat (fibronektino ir kolageno sekrecija fibroblastais). Panašiai kai kurie polisacharidai (hemiceliuliozės), dalyvaujantys formuojant ląstelių sieneles, pašalinami iš augalų ląstelių citoplazmos.
Daugumą išskiriamų medžiagų panaudoja kitos daugialąsčių organizmų ląstelės (pieno, virškinimo sulčių, hormonų ir kt. sekrecija). Tačiau dažnai ląstelės išskiria medžiagas savo reikmėms. Pavyzdžiui, plazminės membranos augimas atliekamas įterpiant membranos dalis kaip egzocitinių vakuolių dalį, kai kuriuos glikokalikso elementus ląstelė išskiria glikoproteinų molekulių pavidalu ir kt.
Hidroliziniai fermentai, išskirti iš ląstelių egzocitozės būdu, gali būti sorbuoti glikokalikso sluoksnyje ir užtikrinti su membrana susietą tarpląstelinį įvairių biopolimerų ir organinių molekulių skilimą. Membraninis neląstelinis virškinimas yra labai svarbus gyvūnams. Buvo nustatyta, kad m žarnyno epitelisžinduolių siurbiamo epitelio vadinamojo šepetėlio krašto srityje, ypač gausu glikokalikso puiki sumaįvairių fermentų. Dalis šių fermentų yra kasos kilmės (amilazės, lipazės, įvairios proteinazės ir kt.), o kai kuriuos išskiria pačios epitelio ląstelės (egzohidrolazės, kurios, susidarant pernešamiems produktams, skaido daugiausia oligomerus ir dimerus).
Plazmalemmos receptorių vaidmuo
Su šia plazminės membranos savybe jau susidūrėme susipažinę su jos transportavimo funkcijomis. Nešiklio baltymai ir siurbliai taip pat yra receptoriai, atpažįstantys tam tikrus jonus ir su jais sąveikaujantys. Receptoriniai baltymai jungiasi prie ligandų ir dalyvauja atrenkant į ląsteles patenkančias molekules.
Membraniniai baltymai arba glikokalikso elementai – glikoproteinai gali veikti kaip tokie receptoriai ląstelės paviršiuje. Tokios atskiroms medžiagoms jautrios vietos gali būti išsklaidytos po ląstelės paviršių arba surinktos mažose zonose.
Skirtingos gyvūnų organizmų ląstelės gali turėti skirtingus receptorių rinkinius arba skirtingą to paties receptoriaus jautrumą.
Daugelio ląstelių receptorių vaidmuo yra ne tik specifinių medžiagų surišimas ar gebėjimas reaguoti fiziniai veiksniai bet ir perduodant tarpląstelinius signalus iš paviršiaus į ląstelę. Šiuo metu yra gerai ištirta signalų perdavimo į ląsteles sistema tam tikrų hormonų, įskaitant peptidines grandines, pagalba. Nustatyta, kad šie hormonai jungiasi prie specifinių receptorių ląstelės plazminės membranos paviršiuje. Receptoriai, prisijungę prie hormono, aktyvuoja kitą baltymą, kuris jau yra plazmos membranos citoplazminėje dalyje – adenilato ciklazę. Šis fermentas iš ATP sintetina ciklinę AMP molekulę. Ciklinio AMP (cAMP) vaidmuo yra tas, kad jis yra antrinis pasiuntinys – fermentų – kinazių, kurios sukelia kitų fermentų baltymų modifikacijas, aktyvatorius. Taigi, kai kasos hormonas gliukagonas, kurį gamina Langerhanso salelių A ląstelės, veikia kepenų ląsteles, hormonas prisijungia prie specifinio receptoriaus, kuris skatina adenilato ciklazės aktyvavimą. Susintetintas cAMP aktyvuoja baltymų kinazę A, kuri savo ruožtu suaktyvina fermentų kaskadą, kuri galiausiai suskaido glikogeną (gyvūnų saugojimo polisacharidą) į gliukozę. Insulino veikimas yra priešingas – jis skatina gliukozės patekimą į kepenų ląsteles ir jos nusėdimą glikogeno pavidalu.
Apskritai įvykių grandinė klostosi taip: hormonas specifiškai sąveikauja su šios sistemos receptorine dalimi ir, neįsiskverbdamas į ląstelę, aktyvuoja adenilatciklazę, kuri sintetina cAMP, kuri aktyvuoja arba slopina tarpląstelinį fermentą ar fermentų grupę. . Taigi komanda, signalas iš plazminės membranos perduodamas ląstelės viduje. Šios adenilato ciklazės sistemos efektyvumas yra labai didelis. Taigi, vienos ar kelių hormonų molekulių sąveika dėl daugelio cAMP molekulių sintezės gali sukelti signalo stiprinimą tūkstančius kartų. Šiuo atveju adenilato ciklazės sistema tarnauja kaip išorinių signalų keitiklis.
Yra ir kitas būdas, kuriuo naudojami kiti antriniai pasiuntiniai – tai yra vadinamasis. fosfatidilinozitolio kelias. Veikiant atitinkamam signalui (kai kuriems nerviniams mediatoriams ir baltymams), suaktyvinamas fermentas fosfolipazė C, kuris skaido fosfatidilinozitolio difosfato fosfolipidą, kuris yra plazminės membranos dalis. Šio lipido hidrolizės produktai, viena vertus, suaktyvina proteinkinazę C, kuri suaktyvina kinazės kaskadą, kuri sukelia tam tikras ląstelių reakcijas, kita vertus, skatina kalcio jonų išsiskyrimą, kuris reguliuoja visa linija ląstelių procesai.
Kitas receptorių aktyvumo pavyzdys yra acetilcholino, svarbaus neurotransmiterio, receptoriai. Acetilcholinas išsiskiria iš nervų galūnė, jungiasi prie receptorių ant raumenų skaidulos, sukelia impulsyvų Na + srautą į ląstelę (membranos depoliarizacija), iš karto atverdama apie 2000 jonų kanalų neuroraumeninio galo zonoje.
Dėl ląstelių paviršiuje esančių receptorių rinkinių įvairovės ir specifiškumo susidaro labai sudėtinga žymenų sistema, leidžianti atskirti savo ląsteles (to paties individo ar tos pačios rūšies) nuo kitų. Panašios ląstelės sąveikauja viena su kita, todėl paviršiai sukimba (konjugacija pirmuoniuose ir bakterijose, susidaro audinių ląstelių kompleksai). Tokiu atveju ląstelės, kurios skiriasi determinantinių žymenų rinkiniu arba jų nesuvokia, yra pašalinamos iš tokios sąveikos arba sunaikinamos aukštesniuose gyvūnuose dėl imunologinių reakcijų (žr. toliau).
Plazminė membrana yra susijusi su specifinių receptorių, reaguojančių į fizinius veiksnius, lokalizacija. Taigi plazmos membranoje arba jos dariniuose fotosintetinėse bakterijose ir melsvadumbliuose yra lokalizuoti receptorių baltymai (chlorofilai), sąveikaujantys su šviesos kvantais. Šviesai jautrių gyvūnų ląstelių plazminėje membranoje yra speciali fotoreceptorių baltymų (rodopsino) sistema, kurios pagalba šviesos signalas paverčiamas cheminiu, o tai savo ruožtu sukelia elektrinio impulso generavimą.
Tarpląstelinis atpažinimas
Daugialąsteliuose organizmuose dėl tarpląstelinės sąveikos susidaro sudėtingi ląstelių ansambliai, kurių priežiūra gali būti vykdoma. Skirtingi keliai. Gimdos, embriono audiniuose, ypač ant ankstyvosios stadijos vystymosi, ląstelės lieka sujungtos viena su kita dėl jų paviršių gebėjimo sulipti. Šis turtas Sukibimas(sujungimas, sukibimas) ląstelių gali būti nulemtos pagal jų paviršiaus savybes, kurios specifiškai sąveikauja viena su kita. Šių jungčių mechanizmas yra gerai ištirtas, jį užtikrina plazmos membranų glikoproteinų sąveika. Esant tokiai tarpląstelinei ląstelių sąveikai tarp plazmos membranų, visada lieka apie 20 nm pločio tarpas, užpildytas glikokaliksu. Audinių apdorojimas fermentais, pažeidžiančiais glikokalikso vientisumą (gleivių, kurios hidroliziškai veikia mucinus, mukopolisacharidus) arba pažeidžiantys plazmos membraną (proteazes), lemia ląstelių izoliaciją viena nuo kitos, jų disociaciją. Tačiau pašalinus disociacijos faktorių ląstelės gali vėl susiburti ir vėl agreguotis. Taigi galima atskirti skirtingų spalvų, oranžinės ir geltonos spalvos kempinių ląsteles. Paaiškėjo, kad šių ląstelių mišinyje susidaro dviejų tipų agregatai: susidedantys tik iš geltonų ir tik iš oranžinių ląstelių. Tokiu atveju mišrios ląstelių suspensijos savaime susitvarko, atkuria pirminę daugialąstę struktūrą. Panašūs rezultatai gauti su atskirtomis varliagyvių embrionų ląstelių suspensijomis; šiuo atveju vyksta selektyvus erdvinis ektodermos ląstelių atskyrimas nuo endodermos ir nuo mezenchimo. Be to, jei audiniai naudojami reagregacijai vėlyvieji etapai vystosi embrionai, tada mėgintuvėlyje savarankiškai susirenka įvairūs audinių ir organų specifiškumo ląstelių ansambliai, susidaro epitelio agregatai, panašūs į inkstų kanalėlių ir kt.
Nustatyta, kad transmembraniniai glikoproteinai yra atsakingi už vienalyčių ląstelių agregaciją. Tiesiogiai už ryšį, sukibimą ląstelės yra atsakingos už vadinamųjų molekulių. CAM baltymai (ląstelių adhezijos molekulės). Vieni jų ląsteles tarpusavyje jungia tarpusavyje dėl tarpmolekulinės sąveikos, kiti formuoja specialius tarpląstelinius ryšius ar kontaktus.
Sąveika tarp lipnių baltymų gali būti homofiliškas kai kaimyninės ląstelės jungiasi viena su kita vienarūšių molekulių pagalba, heterofilinis kai adhezijoje dalyvauja įvairių rūšių CAM ant kaimyninių ląstelių. Tarpląstelinis surišimas vyksta per papildomas jungties molekules.
Yra keletas CAM baltymų klasių. Tai kadherinai, į imunoglobulinus panašūs N-CAM (nervinių ląstelių adhezijos molekulės), selektinai, integrinai.
Kadherinai yra integralūs fibrilinės membranos baltymai, kurie sudaro lygiagrečius homodimerus. Atskiri šių baltymų domenai yra susieti su Ca 2+ jonais, o tai suteikia jiems tam tikrą standumą. Yra daugiau nei 40 kadherinų rūšių. Taigi E-kadherinas būdingas iš anksto implantuotų embrionų ląstelėms ir suaugusių organizmų epitelio ląstelėms. P-kadherinas būdingas trofoblastų, placentos ir epidermio ląstelėms; N-kadherinas yra nervų ląstelių paviršiuje, lęšio ląstelėse, širdies ir skeleto raumenyse.
^ Nervų ląstelių adhezijos molekulės (N-CAM) priklauso imunoglobulinų superšeimai, jie sudaro ryšius tarp nervinių ląstelių. Kai kurios N-CAM yra susijusios su sinapsių ryšiu ir ląstelių adhezija. Imuninė sistema.
selektinų taip pat integraliniai plazmos membranos baltymai dalyvauja endotelio ląstelių sukibime, trombocitų, leukocitų surišime.
Integrinai yra heterodimerai, turintys ir grandines. Integrinai pirmiausia jungia ląsteles su tarpląsteliniais substratais, tačiau jie taip pat gali dalyvauti ląstelių adhezijoje vienas su kitu.
Svetimų baltymų atpažinimas
Kaip jau minėta, į organizmą patekusias svetimas makromolekules (antigenus) vystosi sudėtinga sudėtinga reakcija - Imuninis atsakas. Jo esmė slypi tame, kad kai kurie limfocitai gamina specialius baltymus – antikūnus, kurie specifiškai jungiasi su antigenais. Pavyzdžiui, makrofagai atpažįsta antigeno-antikūno kompleksus su savo paviršiaus receptoriais ir juos sugeria (pavyzdžiui, bakterijų absorbcija fagocitozės metu).
Be to, visų stuburinių gyvūnų organizme yra svetimų ar savų ląstelių priėmimo sistema, tačiau su pakitusiais plazmos membranos baltymais, pvz. virusinės infekcijos arba su mutacijomis, dažnai susijusiomis su ląstelių naviko degeneracija.
Baltymai yra visų stuburinių ląstelių paviršiuje, vadinamieji. pagrindinis histo suderinamumo kompleksas(pagrindinis histokompatibilumo kompleksas – MHC). Tai integraliniai baltymai, glikoproteinai, heterodimerai. Labai svarbu atsiminti, kad kiekvienas žmogus turi skirtingą šių MHC baltymų rinkinį. Taip yra dėl to, kad jie yra labai polimorfiški, nes kiekvienas individas turi daug kintančių to paties geno formų (daugiau nei 100), be to, yra 7-8 lokusai, koduojantys MHC molekules. Dėl to susidaro kiekviena ląstelė duotas organizmas, turintis MHC baltymų rinkinį, skirsis nuo tos pačios rūšies individo ląstelių. ypatinga forma limfocitai, T-limfocitai, atpažįsta savo kūno MHC, bet menkiausias pokytis MHC struktūroje (pavyzdžiui, susiejimas su virusu arba atskirų ląstelių mutacijos rezultatas), lemia tai, kad T-limfocitai atpažįsta tokias pakitusias ląsteles ir jas sunaikina, bet ne fagocitozės būdu. Jie iš sekrecinių vakuolių išskiria specifinius perforino baltymus, kurie yra įterpti į pakitusios ląstelės citoplazminę membraną, formuoja joje transmembraninius kanalus, todėl plazminė membrana tampa pralaidi, todėl pakitusios ląstelės žūva (143, 144 pav.).
Specialūs tarpląsteliniai ryšiai
Be šių gana paprastų lipnių (bet specifinių) jungčių (145 pav.), yra nemažai specialių tarpląstelinių struktūrų, kontaktų ar jungčių, kurios atlieka tam tikras funkcijas. Tai yra fiksavimo, inkaravimo ir ryšių jungtys (146 pav.).
Užrakinimas arba sandarus ryšys būdingas viensluoksniam epiteliui. Tai yra zona, kurioje dviejų plazminių membranų išoriniai sluoksniai yra kuo arčiau. Šiame kontakte dažnai matoma trijų sluoksnių membrana: abu išoriniai osmofiliniai abiejų membranų sluoksniai tarsi susilieja į vieną bendrą 2–3 nm storio sluoksnį. Membranų susiliejimas nevyksta visoje glaudaus kontakto srityje, o yra taškinės membranų konvergencijos serija (147a, 148 pav.).
Plazminių membranų lūžių plokštuminiuose preparatuose sandaraus kontakto zonoje, taikant užšaldymo ir smulkinimo metodą, nustatyta, kad membranų sąlyčio taškai yra rutuliukų eilės. Tai baltymai okludinas ir klaudinas, specialūs integraliniai plazmos membranos baltymai, pastatyti eilėmis. Tokios rutuliukų ar juostelių eilės gali susikirsti taip, kad skilimo paviršiuje susidaro tarsi gardelė arba tinklas. Ši struktūra labai būdinga epiteliams, ypač liaukams ir žarnynui. Pastaruoju atveju glaudus kontaktas sudaro ištisinę plazmos membranų susiliejimo zoną, apjuosiančią ląstelę jos viršūninėje (viršutinėje, žiūrint į žarnyno spindį) dalyje (148 pav.). Taigi kiekviena sluoksnio ląstelė yra tarsi apsupta šio kontakto juosta. Tokias struktūras galima pamatyti ir specialiais dėmėmis šviesos mikroskopu. Pavadinimą jie gavo iš morfologų galinės plokštės. Paaiškėjo, kad šiuo atveju uždaro sandaraus kontakto vaidmuo yra ne tik mechaninis ląstelių sujungimas tarpusavyje. Ši kontaktinė sritis yra prastai pralaidi makromolekulėms ir jonams, todėl užrakina, blokuoja tarpląstelines ertmes, izoliuodama jas (o kartu ir vidinę organizmo aplinką) nuo išorinės aplinkos (šiuo atveju žarnyno spindžio).
Tai galima įrodyti naudojant elektronų tankius kontrastininkus, tokius kaip lantano hidroksido tirpalas. Jei žarnyno arba kurios nors liaukos latako spindis užpildytas lantano hidroksido tirpalu, elektroniniu mikroskopu esančiose pjūviuose zonos, kuriose yra ši medžiaga, turi didelį elektronų tankį ir bus tamsios. Paaiškėjo, kad nei glaudaus kontakto zona, nei po ja esančios tarpląstelinės erdvės netamsėja. Jei sandarios jungtys yra pažeistos (lengvai apdorojant fermentiniu būdu arba pašalinus Ca ++ jonus), lantanas taip pat prasiskverbia į tarpląstelines sritis. Panašiai įrodyta, kad inkstų kanalėliuose esančios sandarios jungtys yra nepralaidžios hemoglobinui ir feritinui.
Taigi sandarios jungtys yra kliūtys ne tik makromolekulėms, bet ir nepralaidžios skysčiams bei jonams.
Uždaromas arba sandarus kontaktas atsiranda tarp visų tipų vienasluoksnio epitelio (endotelio, mezotelio, ependimos).
Inkaravimas arba blokuojančių jungčių arba kontaktai taip vadinami, nes jungia ne tik gretimų ląstelių plazmines membranas, bet ir jungiasi su fibriliniais citoskeleto elementais (149 pav.). Šio tipo junginiams būdingas dviejų tipų baltymų buvimas. Vienas iš jų yra transmembraniniai jungiamieji (surišantys) baltymai, dalyvaujantys arba faktiniame tarpląsteliniame jungime, arba plazmolemos sujungime su ekstraląstelinės matricos komponentais (bazine epitelio membrana, tarpląsteliniais jungiamojo audinio struktūriniais baltymais).
Antrasis – tarpląsteliniai baltymai, jungiantys arba įtvirtinantys citoskeleto citoplazmines fibriles už tokio kontakto membraninių elementų.
Inkaravimo jungtys apima tarpląstelines tvirtinimo taškų jungtis, tvirtinimo juostas, židinio jungtis arba inkaravimo plokšteles, kurios visos ląstelėje jungiasi su aktino mikrofilamentais.
Kita inkaravimo tarpląstelinių jungčių grupė, desmosomos ir hemidesmosomos, jungiasi su kitais citoskeleto elementais, būtent prie tarpinių gijų.
Daugelyje ne epitelinių audinių buvo rasta tarpląstelinių smailių jungčių, tačiau struktūra buvo aiškesnė. specialios (lipnios) juostos vienasluoksniame epitelyje (150 pav.). Ši struktūra juosia visą epitelio ląstelės perimetrą, panašiai kaip ir esant sandariai susijungimui. Dažniausiai toks diržas ar juosta guli po sandaria jungtimi (žr. 146 pav.). Šioje vietoje plazminės membranos nėra suartintos, o net šiek tiek pasislinkusios 25–30 nm atstumu, tarp jų matoma padidėjusio tankio zona. Tai ne kas kita, kaip transmembraninių glikoproteinų sąveikos vietos, kurios specifiškai prilimpa viena prie kitos ir užtikrina mechaninį ryšį tarp dviejų gretimų ląstelių membranų. Šie jungiamieji baltymai priklauso E-kadherinams, baltymams, kurie specifiškai atpažįsta vienalytes membranas ląstelėse. Šio glikoproteinų sluoksnio sunaikinimas lemia atskirų ląstelių izoliaciją ir epitelio sluoksnio sunaikinimą. Citoplazminėje pusėje prie membranos matosi tam tikros tankios medžiagos sankaupa, prie kurios prigludęs plonų (6-7 nm) gijų sluoksnis, išsidėstęs išilgai plazmos membranos ryšulio pavidalu, einantis per visą perimetrą. ląstelė. Plonos gijos yra aktino fibrilės; jie jungiasi prie plazmos membranos per katenino baltymą, kuris šalia membranos sudaro tankų sluoksnį.
Tokios juostelės jungties funkcinė reikšmė slypi tik mechaniniame ląstelių sukibime viena su kita: juostelėje susitraukus aktino gijų, ląstelės forma gali keistis. Manoma, kad dėl kooperatyvinio aktino fibrilių susitraukimo visose epitelio sluoksnio ląstelėse gali pasikeisti jo geometrija, pavyzdžiui, susilankstyti į vamzdelį, panašiai kaip stuburinių embrionų nervinio vamzdelio formavimosi metu.
^ židinio kontaktai arba sukibimo plokštelės atsiranda daugelyje ląstelių ir yra ypač gerai ištirtos fibroblastuose. Jie pastatyti pagal bendrasis planas su lipniomis juostomis, bet išreiškiami nedideliais ploteliais – apnašomis ant plazmalemos. Šiuo atveju transmembraniniai jungiamieji integrino baltymai specifiškai jungiasi prie ekstraląstelinės matricos baltymų (pavyzdžiui, fibronektino) (151 pav.). Iš citoplazmos pusės tie patys glikoproteinai yra susiję su membranos baltymais, į kuriuos įeina ir vinkulinas, kuris savo ruožtu yra susijęs su aktino gijų pluoštu. Funkcinė židinio kontaktų reikšmė yra tiek pritvirtinant ląstelę prie tarpląstelinių struktūrų, tiek kuriant mechanizmą, leidžiantį ląstelėms judėti.
Desmosomos, plokštelių ar mygtukų pavidalo struktūros taip pat jungia ląsteles viena su kita (152, 153a pav.). Tarpląstelinėje erdvėje čia taip pat matomas tankus sluoksnis, kurį vaizduoja sąveikaujantys vientisai membraniniai kadherinai – desmogleinai, jungiantys ląsteles viena su kita. Citoplazminėje pusėje prie plazmolemos yra desmoplakino baltymo sluoksnis, su kuriuo yra susiję tarpiniai citoskeleto siūlai. Desmosomos dažniausiai randamos epitelyje, tokiu atveju tarpinėse gijose yra keratino. Širdies raumenų ląstelėse, kardiomiocituose, yra desmino fibrilių, kurie yra desmosomų dalis. Kraujagyslių endotelyje desmosomose yra tarpinių vimentino gijų.
Hemidesmosomos- iš esmės jie savo struktūra yra panašūs į desmosomą, tačiau tai yra ląstelių jungtis su tarpląstelinėmis struktūromis. Taigi epitelyje desmosomų jungiamieji glikoproteinai (integrinai) sąveikauja su vadinamųjų baltymų baltymais. bazinė membrana, kurią sudaro kolagenas, lamininas, proteoglikanai ir kt.
Funkcinis desmosomų ir hemidesmosomų vaidmuo yra grynai mechaninis – jos tvirtai prilimpa ląsteles viena prie kitos ir prie pagrindinės tarpląstelinės matricos, todėl epitelio sluoksniai gali atlaikyti dideles mechanines apkrovas. Panašiai desmosomos glaudžiai suriša širdies raumens ląsteles viena su kita, o tai leidžia joms atlikti didžiulę mechaninę apkrovą, išlikdamos surištos į vieną susitraukiančią struktūrą.
Skirtingai nuo glaudaus kontakto, visų tipų jungiamieji kontaktai yra pralaidūs vandeniniams tirpalams ir neriboja difuzijos.
^ Spragų kontaktai yra laikomi ląstelių ryšio ryšiais; Tai yra struktūros, kurios yra tiesiogiai perduodamos cheminių medžiagų iš ląstelės į ląstelę, kuri gali atlikti didelį fiziologinį vaidmenį ne tik specializuotų ląstelių funkcionavime, bet ir užtikrinti tarpląstelinę sąveiką organizmo vystymosi metu, jo ląstelių diferenciacijos metu. Šio tipo kontaktams būdinga dviejų gretimų ląstelių plazminių membranų konvergencija 2-3 nm atstumu (147b, 153b pav.). Būtent ši aplinkybė ilgam laikui neleido atskirti ant itin plonų pjūvių ši rūšis kontaktas nuo sandaraus izoliuojančio (uždarančio) kontakto. Naudojant lantano hidroksidą, pastebėta, kad kai kurie sandarūs kontaktai nuteka kontrastas. Šiuo atveju lantanas užpildė ploną maždaug 3 nm pločio tarpą tarp gretimų gretimų ląstelių plazminių membranų. Iš čia kilo terminas – tarpo kontaktas. Tolesnė pažanga iššifruojant jo struktūrą buvo pasiekta naudojant užšaldymo-čipavimo metodą. Paaiškėjo, kad tarpo jungties zonos (nuo 0,5 iki 5 µm) ant membranų plyšių yra išmargintos 7–8 nm skersmens dalelėmis, šešiakampėmis išsidėsčiusiomis 8–10 nm periodu, turinčioje maždaug 2 nm pločio kanalą. centras. Šios dalelės vadinamos jungtys(154 pav.). Tarpo kontakto zonose gali būti nuo 10-20 iki kelių tūkstančių jungčių, priklausomai nuo funkcinės savybės ląstelės. Konneksonai buvo išskirti preparatiškai, susideda iš šešių subvienetų konnekino, baltymo, kurio molekulinė masė apie 30 tūkst.. Jungdamiesi tarpusavyje, jungiasi suformuoja cilindrinį agregatą – konneksoną, kurio centre yra kanalas. Atskiros jungtys yra įterptos į plazmos membraną taip, kad prasiskverbia pro ją. Vienai ląstelės plazminės membranos jungtis tiksliai priešinasi gretimos ląstelės plazminės membranos jungtis, todėl dviejų jungčių kanalai sudaro vieną vienetą. Jungtys atlieka tiesioginių tarpląstelinių kanalų, per kuriuos jonai ir mažos molekulinės masės medžiagos gali sklisti iš ląstelės į ląstelę, vaidmenį. Nustatyta, kad jungtys gali užsidaryti, pakeisdamos vidinio kanalo skersmenį ir taip dalyvauti reguliuojant molekulių transportavimą tarp ląstelių.
Tarpų jungčių funkcinė reikšmė buvo suprasta tiriant milžiniškas ląsteles seilių liaukos Diptera. Dėl savo dydžio mikroelektrodai gali būti lengvai įvedami į tokias ląsteles, kad būtų galima ištirti jų membranų elektrinį laidumą. Paaiškėjo, kad jei elektrodai įkišti į dvi gretimas ląsteles, jų plazminės membranos pasižymi maža elektrine varža, o srovė teka tarp ląstelių. Be to, paaiškėjo, kad kai į vieną ląstelę įšvirkščiamas fluorescencinis dažiklis, žymė greitai aptinkama kaimyninėse ląstelėse. Naudojant skirtingus fluorochromus, ant žinduolių audinių kultūros ląstelių buvo nustatyta, kad medžiagos, kurių molekulinė masė ne didesnė kaip 1-1,5 tūkst., o dydis ne didesnis kaip 1,5 nm, gali būti pernešamos per tarpines jungtis (vabzdžiuose – medžiagos, kurių molekulinė masė sveriančių iki 2 tūkst.). Tarp šių medžiagų buvo įvairių jonų, aminorūgščių, nukleotidų, cukrų, vitaminų, steroidų, hormonų, cAMP. Nei baltymai, nei nukleorūgštys negali praeiti per tarpų jungtis.
Ši tarpų jungčių galimybė tarnauti kaip mažos molekulinės masės junginių transportavimo vieta yra naudojama ląstelių sistemos, kur reikalingas greitas elektrinio impulso (sužadinimo bangos) perdavimas iš ląstelės į ląstelę, nedalyvaujant nerviniam tarpininkui. Taigi visos širdies miokardo raumenų ląstelės yra sujungtos naudojant tarpines jungtis (be to, ląstelės ten taip pat yra sujungtos lipniomis jungtimis) (147b pav.). Tai sukuria sąlygas sinchroniškai sumažinti daugybę ląstelių. Augant embrioninių širdies raumens ląstelių (miokardiocitų) kultūrai, kai kurios sluoksnio ląstelės pradeda spontaniškai trauktis nepriklausomai viena nuo kitos. skirtingas dažnis, ir tik tarp jų susidarius tarpinėms jungtims, jos pradeda sinchroniškai plakti kaip vienas susitraukiantis ląstelių sluoksnis. Tokiu pat būdu užtikrinamas gimdos sienelės lygiųjų raumenų ląstelių susitraukimas.
Tarpų jungtys gali būti naudojamos metaboliniam bendradarbiavimui tarp ląstelių, keičiantis įvairiomis molekulėmis, hormonais, cAMP ar metabolitais. To pavyzdys yra timidinkinazės mutantinių ląstelių auginimas kartu su normaliomis ląstelėmis: kai tarp šių ląstelių tipų atsiranda tarpinių jungčių, mutantinės ląstelės per tarpines jungtis gauna timidino trifosfatą iš normalių ląstelių ir gali dalyvauti DNR sintezėje.
Ankstyvuosiuose stuburinių embrionuose, pradedant nuo 8 ląstelių stadijos, dauguma ląstelių yra sujungtos viena su kita tarpų jungtimis. Kai embrionas diferencijuojasi, tarpų jungtys tarp visų ląstelių išnyksta ir išlieka tik tarp specializuotų ląstelių grupių. Pavyzdžiui, formuojantis nerviniam vamzdeliui šios struktūros ląstelių ryšys su likusia epidermio dalimi nutrūksta, atsijungia.
Tarpų jungčių vientisumas ir funkcionavimas labai priklauso nuo Ca 2+ jonų lygio ląstelės viduje. Paprastai kalcio koncentracija citoplazmoje yra labai maža. Jei Ca 2+ įšvirkščiama į vieną iš audinių kultūros sluoksnio ląstelių, tai kaimyninėse ląstelėse Ca 2+ kiekis citoplazmoje nepadidėja; ląstelės tarsi yra atjungtos nuo kaimynų, nustoja veikti elektros ir dažikliai. Po kurio laiko, įvestą kalcį sukaupus mitochondrijoms, atsistato tarpinių jungčių struktūra ir funkcijos. Ši savybė yra labai svarbi norint išlaikyti viso ląstelių sluoksnio vientisumą ir veikimą, nes vienos iš jų pažeidimai neperduodama kaimyniniam per tarpines jungtis, kurios nustoja veikti kaip tarpląsteliniai difuzijos kanalai.
^ sinapsinis kontaktas (sinapsės). Šio tipo kontaktai būdingi nervinis audinys ir atsiranda tiek tarp dviejų neuronų, tiek tarp neurono ir kokio nors kito elemento – receptoriaus arba efektoriaus (pavyzdžiui, neuroraumeninės galūnės). Sinapsės – tai dviejų ląstelių, kurios specializuojasi sužadinimo ar slopinimo vienpusiam perdavimui iš vieno elemento į kitą, sąlyčio sritys (155 pav.). Iš esmės šios rūšies funkcinė apkrova, impulsų perdavimas gali būti vykdomas ir kitų tipų kontaktais (pavyzdžiui, tarpo kontaktas širdies raumenyje), tačiau sinapsiniame ryšyje didelis efektyvumas realizuojant nervinį impulsą. Nervinių ląstelių procesuose susidaro sinapsės - tai galinės dendritų ir aksonų dalys. Tarpneuroninės sinapsės dažniausiai turi kriaušės formos išplėtimus, plokšteles nervinės ląstelės proceso pabaigoje. Toks vienos nervinės ląstelės proceso galinis išplėtimas gali liestis ir sudaryti sinapsinį ryšį tiek su kitos nervinės ląstelės kūnu, tiek su jo procesais. Nervinių ląstelių (aksonų) periferiniai procesai sudaro specifinius kontaktus su efektorinėmis arba receptorinėmis ląstelėmis. Todėl sinapsė yra struktūra, kuri susidaro tarp dviejų ląstelių sričių (taip pat ir desmosomos). Šių ląstelių membranas skiria tarpląstelinė erdvė – apie 20-30 nm pločio sinapsinis plyšys. Neretai šio plyšio spindyje matosi membranoms statmena smulkaus pluošto medžiaga. Vienos ląstelės sinapsinio kontakto srityje esanti membrana vadinama presinaptine, kita, suvokianti impulsą, vadinama postsinaptine. Elektroniniame mikroskope abi membranos atrodo tankios ir storos. Netoli presinapsinės membranos atsiskleidžia daugybė mažų vakuolių, sinapsinių pūslelių, užpildytų neurotransmiteriais. Sinapsinės pūslelės, praeinant nerviniam impulsui, išmeta savo turinį į sinapsinį plyšį. Postsinapsinė membrana dažnai atrodo storesnė nei įprastos membranos, nes aplink ją iš citoplazmos pusės susikaupia daug plonų fibrilių.
Plazmodesmata. Šio tipo tarpląstelinis ryšys randamas augaluose. Plazmodesmos yra ploni vamzdiniai citoplazminiai kanalai, jungiantys dvi gretimas ląsteles. Šių kanalų skersmuo paprastai yra 20-40 nm. Šiuos kanalus ribojanti membrana tiesiogiai patenka į gretimų ląstelių plazmines membranas. Plazmodesmos praeina per ląstelės sienelę, kuri skiria ląsteles (156, 157 pav.). Taigi kai kuriose augalų ląstelėse plazmodesmos jungia gretimų ląstelių hialoplazmą, todėl formaliai nėra visiško skirtumo, vienos ląstelės kūno atskyrimo nuo kitos, tai veikiau sincitija: daugelio ląstelių teritorijų susijungimas citoplazmos pagalba. tiltai. Membraniniai vamzdiniai elementai gali prasiskverbti į plazmodesmatos vidų, sujungdami kaimyninių ląstelių endoplazminio tinklo cisternas. Plazmodezmos susidaro ląstelių dalijimosi metu, kai susidaro pirminė ląstelės sienelė (žr. toliau). Naujai padalintose ląstelėse plazmodesmų skaičius gali būti labai didelis (iki 1000 ląstelėje), ląstelėms senstant jų skaičius mažėja dėl plyšimų didėjant storiui. ląstelių sienelės.
Funkcinis plazmodesmos vaidmuo yra labai didelis: su jų pagalba tarpląstelinė tirpalų cirkuliacija maistinių medžiagų, jonai ir kiti junginiai. Lipidų lašeliai gali judėti išilgai plazmodesmų.Ląstelės užkrečiamos augalų virusais per plazmodesmatas. Tačiau eksperimentai rodo, kad laisvas pernešimas per plazmodesmatas apsiriboja dalelėmis, kurių masė ne didesnė kaip 800 daltonų.
> Plazminės membranos funkcijos. Medžiagų transportavimo per plazmalemą mechanizmai. Plazlemos receptorių funkcija
Plazminė membrana atlieka keletą svarbių funkcijų:
1) barjeras. Plazminės membranos barjerinė funkcija yra apriboti laisvą medžiagų difuziją iš ląstelės į ląstelę, užkirsti kelią vandenyje tirpaus ląstelės turinio nutekėjimui. Bet kadangi ląstelė turi gauti reikiamas maistines medžiagas, išskirti galutinius medžiagų apykaitos produktus, reguliuoti tarpląstelines jonų koncentracijas, joje susiformavo specialūs medžiagų pernešimo per ląstelės membraną mechanizmai.
2) Transportas. Transporto funkcija yra Įvairių medžiagų patekimo į ląstelę ir iš jos išėjimo užtikrinimas. Svarbi nuosavybė membranos - selektyvus pralaidumas, arba pusiau pralaidumas. Jis lengvai praleidžia vandenį ir vandenyje tirpias dujas bei atstumia polines molekules, tokias kaip gliukozė ar aminorūgštys.
Yra keletas medžiagų transportavimo per membraną mechanizmų:
pasyvus transportas;
aktyvus transportas;
transportavimas membraninėje pakuotėje.
Pasyvus transportas.difuzija - Tai yra terpės dalelių judėjimas, dėl kurio medžiaga perkeliama iš vietos, kurioje jos koncentracija didelė, į mažos koncentracijos sritį. Difuzijos transportavimo metu membrana veikia kaip osmosinis barjeras. Difuzijos greitis priklauso nuo molekulių dydžio ir santykinio jų tirpumo riebaluose. Kaip mažesni dydžiai molekulių ir kuo daugiau jos tirpsta riebaluose (lipofilinės), tuo greičiau judės per lipidų dvisluoksnį sluoksnį. Difuzija gali būti neutralus(neįkrautų molekulių perkėlimas) ir lengvas(specialių nešančiųjų baltymų pagalba). Supaprastinta difuzija yra greitesnė nei neutrali difuzija. Vanduo turi didžiausią prasiskverbimo galią, nes jo molekulės yra mažos ir neįkrautos. Vandens difuzija per ląstelės membraną vadinama osmosas. Daroma prielaida, kad ląstelės membranoje yra specialios „poros“, skirtos vandeniui ir kai kuriems jonams prasiskverbti. Jų skaičius mažas, o skersmuo apie 0,3-0,8 nm. Lengvai tirpios lipidų dvisluoksnio molekulės, tokios kaip O, ir neįkrautos mažo skersmens polinės molekulės (CO, karbamidas) greičiausiai difunduoja per membraną.
Polinių molekulių (cukrų, aminorūgščių) perkėlimas, atliekamas naudojant specialius membraninius transportavimo baltymus, vadinamas palengvinta difuzija. Tokie baltymai randami visų tipų biologinėse membranose, o kiekvienas specifinis baltymas yra skirtas tam tikros klasės molekulėms pernešti. Transporto baltymai yra transmembraniniai, jų polipeptidinė grandinė kelis kartus kerta lipidų dvisluoksnį sluoksnį, susidaranti per jame esančius praėjimus. Tai užtikrina specifinių medžiagų pernešimą per membraną be tiesioginio kontakto su ja. Yra dvi pagrindinės transporto baltymų klasės: nešantys baltymai (transporteriai) Ir kanalo formavimas baltymai (baltymų kanalai). Nešėjai baltymai perneša molekules per membraną, pirmiausia pakeisdami jų konfigūraciją. Kanalą formuojantys baltymai membranoje sudaro vandens užpildytas poras. Kai poros atsivėrusios, pro jas praeina specifinių medžiagų molekulės (dažniausiai tinkamo dydžio ir krūvio neorganiniai jonai). Jei gabenamos medžiagos molekulė neturi krūvio, tada transportavimo kryptį lemia koncentracijos gradientas. Jei molekulė yra įkrauta, tai jos pernešimui, be koncentracijos gradiento, įtakos turi ir membranos elektrinis krūvis (membranos potencialas). Vidinė pusė Plazlema paprastai yra neigiamai įkrauta, palyginti su išorine. Membranos potencialas palengvina teigiamo krūvio jonų prasiskverbimą į ląstelę ir neleidžia prasiskverbti neigiamo krūvio jonams.
aktyvus transportas. Aktyvus transportavimas yra medžiagų judėjimas prieš elektrocheminį gradientą. Jį visada vykdo baltymai transporteriai ir yra glaudžiai susiję su energijos šaltiniu. Nešantys baltymai turi prisijungimo vietas su transportuojama medžiaga. Kuo daugiau tokių vietų susieta su medžiaga, tuo didesnis transportavimo greitis. Atrankinis vienos medžiagos perkėlimas vadinamas uniportas. Atliekamas kelių medžiagų perkėlimas bendro transporto sistemos. Jei perkėlimas vyksta viena kryptimi, tai yra simportas, jei priešingai antiportas. Pavyzdžiui, gliukozė iš ekstraląstelinio skysčio į ląstelę pernešama vienaip. Gliukozės ir Na 4 perkėlimas iš žarnyno ertmės arba atitinkamai inkstų kanalėlių į žarnyno ar kraujo ląsteles atliekamas paprastai, o C1 ~ ir HCO pernešimas yra antiportas.
Baltymų nešiklio, kuris naudoja ATP hidrolizės metu išsiskiriančią energiją medžiagoms transportuoti, pavyzdys Na + -Į + siurblys, randama visų ląstelių plazminėje membranoje. Na + -K siurblys veikia antiport principu, pumpuoja Na "iš ląstelės ir K t į ląstelę prieš jų elektrocheminius gradientus. Na + gradientas sukuria osmosinį slėgį, palaiko ląstelės tūrį ir užtikrina cukrų bei aminorūgščių transportavimą Trečdalis visos energijos, reikalingos ląstelių gyvybinei veiklai, sunaudojama šiam siurbliui.Tiriant Na + -K + siurblio veikimo mechanizmą, buvo nustatyta, kad tai yra ATPazės fermentas ir transmembraninis vientisas baltymas. Na + ir ATP buvimas, veikiant ATPazei, galinis fosfatas yra atskiriamas nuo ATP ir prijungiamas prie liekanos asparto rūgštis ant ATPazės molekulės. ATPazės molekulė fosforilinama, keičia savo konfigūraciją ir Na + išsiskiria iš ląstelės. Pašalinus Na iš ląstelės, K "visada pernešamas į ląstelę. Tam, dalyvaujant K, iš ATPazės atskaldomas anksčiau prijungtas fosfatas. Fermentas defosforilinamas, atkuria jo konfigūraciją, o K 1 yra " pumpuojamas“ į ląstelę.
ATPazę sudaro du subvienetai - didelis ir mažas. Didelis subvienetas susideda iš tūkstančių aminorūgščių liekanų, kurios kelis kartus kerta dvisluoksnį. Jis turi katalizinį aktyvumą ir gali būti grįžtamai fosforilinamas ir defosforilinamas. Didelis subvienetas citoplazminėje pusėje turi Na + ir ATP surišimo vietas ir toliau lauke-K + ir ouabaino surišimo vietos. Mažas subvienetas yra glikoproteinas ir jo funkcija dar nežinoma.
Na + -K siurblys turi elektrogeninį poveikį. Jis pašalina iš ląstelės tris teigiamai įkrautus Na f jonus ir įveda į ją du K jonus. Dėl to per membraną teka srovė, suformuodama neigiamą elektrinį potencialą vidinėje ląstelės dalyje, palyginti su jos išoriniu paviršiumi. . Na "-K + siurblys reguliuoja ląstelės tūrį, kontroliuoja medžiagų koncentraciją ląstelės viduje, palaiko osmosinį slėgį ir dalyvauja kuriant membranos potencialą.
Transportavimas membraninėje pakuotėje. Makromolekulių (baltymų, nukleino rūgščių, polisacharidų, lipoproteinų) ir kitų dalelių pernešimas per membraną vyksta nuosekliai formuojant ir susiliejant pūsleles (pūsleles), apsuptas membranos. Vezikulinio transportavimo procesas vyksta dviem etapais. Iš pradžių pūslelių membrana ir plazmolema sulimpa, o vėliau susilieja. 2 etapo eigoje būtina, kad vandens molekulės būtų išstumtos sąveikaujančiais lipidų dvisluoksniais sluoksniais, kurie artėtų vienas prie kito 1-5 nm atstumu. Manoma, kad šį procesą suaktyvina specialūs sulieti baltymai(jie iki šiol buvo išskirti tik virusuose). Vezikulinis transportas turi svarbi savybė- absorbuotos arba išskiriamos makromolekulės pūslelėse dažniausiai nesimaišo su kitomis ląstelės makromolekulėmis ar organelėmis. Burbulai gali susilieti su specifinėmis membranomis, o tai užtikrina makromolekulių mainus tarp tarpląstelinės erdvės ir ląstelės turinio. Panašiai makromolekulės perkeliamos iš vieno ląstelės skyriaus į kitą.
Makromolekulių ir dalelių pernešimas į ląstelę vadinamas endocitozė. Tokiu atveju pernešamos medžiagos yra apgaubiamos plazminės membranos dalimi, susidaro burbulas (vakuolė), kuris juda ląstelės viduje. Priklausomai nuo susidariusių pūslelių dydžio, išskiriami du endocitozės tipai - pinocitozė ir fagocitozė.
pinocitozė užtikrina skystų ir ištirpusių medžiagų absorbciją mažų burbuliukų pavidalu (d=150 nm). fagocitozė - tai didelių dalelių, mikroorganizmų ar organelių, ląstelių fragmentų absorbcija. Tokiu atveju susidaro didelės pūslelės, fagosomos arba vakuolės (d-250 nm ir daugiau). Pirmuonių fagocitinė funkcija yra mitybos forma. Žinduolių fagocitinę funkciją atlieka makrofagai ir neutrofilai, kurie apsaugo organizmą nuo infekcijos, apimdami invazinius mikrobus. Makrofagai taip pat dalyvauja šalinant senas ar pažeistas ląsteles ir jų fragmentus (žmogaus organizme makrofagai kasdien sugeria daugiau nei 100 senų raudonųjų kraujo kūnelių). Fagocitozė prasideda tik tada, kai absorbuota dalelė prisijungia prie fagocito paviršiaus ir aktyvuoja specializuotas receptorines ląsteles. Dalelių prisijungimas prie specifinių membranos receptorių sukelia pseudopodijų susidarymą, kurie apgaubia dalelę ir, susilieję kraštuose, sudaro burbulą - fagosoma. Fagosomos susidarymas ir tinkama fagocitozė įvyksta tik tuo atveju, jei gaubimo proceso metu dalelė nuolat liečiasi su plazmalemos receptoriais, tarsi „užtrauktuku“.
Didelė dalis medžiagos, kurią ląstelė absorbuoja endocitozės būdu, patenka į lizosomas. Į jį įtrauktos didelės dalelės fagosomos kurios vėliau susilieja su lizosomomis ir susidaro fagolizosomos. Pinocitozės metu paimtos skysčiai ir makromolekulės iš pradžių perkeliamos į endosomas, kurios taip pat susilieja su lizosomomis, sudarydamos endolizosomas. Lizosomose esantys įvairūs hidroliziniai fermentai greitai sunaikina makromolekules. Hidrolizės produktai (aminorūgštys, cukrūs, nukleotidai) iš lizosomų pernešami į citozolį, kur juos panaudoja ląstelė. Dauguma endocitinių pūslelių membraninių komponentų iš fagosomų ir endosomų eksocitozės būdu grąžinami į plazmos membraną ir ten pakartotinai panaudojami. Pagrindinis biologinė reikšmė Endocitozė yra statybinių blokų įsigijimas per ląstelinį makromolekulių virškinimą lizosomose.
Medžiagų absorbcija eukariotinės ląstelės prasideda specializuotuose plazminės membranos regionuose, vadinamuosiuose ribojamos duobės. Elektroninėse mikrografijose duobutės atrodo kaip plazminės membranos invaginacijos, kurių citoplazminė pusė padengta pluoštiniu sluoksniu. Sluoksnis tarsi ribojasi su mažomis plazmalemos duobutėmis. Duobės užima apie 2% viso eukariotinės ląstelės membranos paviršiaus. Per minutę duobutės išauga, įsiskverbia vis giliau ir giliau, įsitraukia į ląstelę, o po to, susiaurėjusios prie pagrindo, atsiskiria, suformuodamos ribojančias pūsleles. Nustatyta, kad maždaug ketvirtadalis membranos ribojamų pūslelių pavidalu atsiskiria nuo fibroblastų plazminės membranos per vieną minutę. Pūslelės greitai praranda savo ribą ir įgyja galimybę susilieti su lizosoma.
Gali būti endocitozė nespecifinis(konstitucinis) ir specifinis(receptorius). At nespecifinė endocitozė ląstelė sulaiko ir sugeria jai visiškai svetimas medžiagas, pavyzdžiui, suodžių daleles, dažus. Iš pradžių dalelės nusėda ant plazmalemos glikokalikso. Teigiamai įkrautos baltymų grupės yra ypač gerai nusodinamos (adsorbuojamos), nes glikokaliksas turi neigiamą krūvį. Tada pasikeičia ląstelės membranos morfologija. Jis gali arba nuskęsti, sudarydamas invaginacijas (invaginacijas), arba, atvirkščiai, formuoti ataugas, kurios tarsi susilanksto, atskirdamos nedidelius skystos terpės tūrius. Invaginacijų formavimasis labiau būdingas žarnyno epitelio ląstelėms, ameboms, o ataugoms - fagocitams ir fibroblastams. Šiuos procesus gali blokuoti kvėpavimo inhibitoriai. Susidariusios pūslelės – pirminės endosomos – gali susilieti viena su kita, didėja jų dydis. Vėliau jie susijungia su lizosomomis ir virsta endolizosoma – virškinimo vakuole. Skystos fazės nespecifinės pinocitozės intensyvumas yra gana didelis. Makrofagai susidaro iki 125, o plonosios žarnos epitelio ląstelės – iki tūkstančio pinosomų per minutę. Pinosomų gausa lemia tai, kad plazmolema greitai išleidžiama daugybei mažų vakuolių susidarymo. Membranos atstatymas vyksta gana greitai per reciklą eksocitozės metu dėl vakuolių sugrįžimo ir jų įtraukimo į plazmalemą. Makrofaguose visa plazminė membrana pakeičiama per 30 minučių, o fibroblastuose – per 2 valandas.
Daugiau efektyvus būdas specifinių makromolekulių absorbcija iš ekstraląstelinio skysčio yra specifinė endocitozė(tarpininkauja receptoriai). Tokiu atveju makromolekulės jungiasi prie papildančių receptorių ląstelės paviršiuje, kaupiasi ribojančioje duobėje, o tada, suformuodamos endosomą, panardinamos į citozolį. Receptorių endocitozė užtikrina specifinių makromolekulių kaupimąsi prie jo receptoriaus. Molekulės, kurios jungiasi prie plazmalemos paviršiuje esančio receptoriaus, vadinamos ligandai. Daugelio gyvūnų ląstelių receptorių endocitozės pagalba cholesterolis absorbuojamas iš tarpląstelinės aplinkos.
Plazminė membrana dalyvauja pašalinant medžiagas iš ląstelės (egzocitozė). Tokiu atveju vakuolės artėja prie plazmalemos. Sąlyčio taškuose plazmolema ir vakuolės membrana susilieja ir vakuolės turinys patenka į aplinką. Kai kurių pirmuonių ląstelės membranoje yra iš anksto nustatytos egzocitozės vietos. Taigi kai kurių blakstienų plazmos membranoje yra tam tikrų sričių, kuriose yra teisingas didelių integruotų baltymų rutuliukų išdėstymas. Visiškai pasirengusių sekrecijai blakstienų mukocistos ir trichocistos turi integruotų baltymų rutuliukų aureolę viršutinėje plazmalemos dalyje. Šios mukocistų ir trichocistų membranos dalys liečiasi su ląstelės paviršiumi. Neutrofiluose stebima savita egzocitozė. Tam tikromis sąlygomis jie gali išleisti savo lizosomas į aplinką. Kai kuriais atvejais susidaro nedidelės plazmalemos ataugos, kuriose yra lizosomų, kurios vėliau nutrūksta ir patenka į aplinką. Kitais atvejais plazmalema įsiskverbia giliai į ląstelę ir užfiksuoja lizosomas, esančias toli nuo ląstelės paviršiaus.
Endocitozės ir egzocitozės procesai atliekami dalyvaujant citoplazmos fibrilinių komponentų sistemai, susijusiai su plazmolema.
Plazlemos receptorių funkcija. Tai viena iš pagrindinių, universalių visoms ląstelėms, yra plazmalemos receptorių funkcija. Tai lemia ląstelių sąveiką tarpusavyje ir su išorine aplinka.
Visa informacinių tarpląstelinių sąveikų įvairovė gali būti schematiškai pavaizduota kaip nuoseklių reakcijų signalas-receptorius-antrinis pasiuntinys-atsakymas grandinė (signalo-atsakymo koncepcija). Informacijos perdavimas iš ląstelės į ląstelę vykdomas signalinėmis molekulėmis, kurios gaminamos vienose ląstelėse ir specifiškai veikia kitas, jautrias signalui (taikinių ląsteles). Signalo molekulė - pirminis tarpininkas jungiasi prie receptorių, esančių tikslinėse ląstelėse, kurie reaguoja tik į tam tikrus signalus. Signalų molekulės - ligandai - prieiti prie jų receptorių kaip prie spynos rakto. Membranų receptorių (plazmalemos receptorių) ligandai yra hidrofilinės molekulės, peptidiniai hormonai, neurotransmiteriai, citokinai, antikūnai, o branduoliniams receptoriams - riebaluose tirpios molekulės, steroidiniai ir skydliaukės hormonai, vitaminas D. Membranos baltymai arba glikokalikso elementai gali veikti kaip receptoriai ant ląstelės paviršius – polisacharidai ir glikoproteinai. Manoma, kad atskiroms medžiagoms jautrios sritys yra išsklaidytos po ląstelės paviršių arba surenkamos mažose zonose. Taigi, prokariotinių ląstelių ir gyvūnų ląstelių paviršiuje yra ribotas vietų, su kuriomis gali prisijungti virusinės dalelės, skaičius. Membraniniai baltymai (nešikliai ir kanalai) atpažįsta, sąveikauja ir perneša tik tam tikras medžiagas. Ląstelių receptoriai dalyvauja perduodant signalus iš ląstelės paviršiaus į ją. Dėl ląstelės paviršiaus receptorių rinkinių įvairovės ir specifiškumo susidaro labai sudėtinga žymenų sistema, leidžianti atskirti savo ląsteles nuo kitų. Panašios ląstelės sąveikauja viena su kita, jų paviršiai gali sulipti (konjugacija pirmuoniuose, audinių formavimasis daugialąsteliuose). Ląstelės, kurios nesuvokia žymenų, taip pat tos, kurios skiriasi determinantinių žymenų rinkiniu, yra sunaikinamos arba atmetamos. Susidarius receptoriaus-ligando kompleksui, aktyvuojami transmembraniniai baltymai: konverterinis baltymas, stiprintuvo baltymas. Dėl to receptorius keičia savo konformaciją ir sąveikauja su antrojo pasiuntinio pirmtaku, esančiu ląstelėje - pasiuntinys. Pasiuntiniai gali būti jonizuotas kalcis, fosfolipazė C, adenilato ciklazė, guanilato ciklazė. Pasiuntinio įtakoje suaktyvinami sintezėje dalyvaujantys fermentai cikliniai monofosfatai – AMP arba HMF. Pastarieji keičia dviejų tipų baltymų kinazės fermentų aktyvumą ląstelės citoplazmoje, todėl fosforilinami daugybė intraląstelinių baltymų.
Dažniausias cAMP susidarymas, kuriam veikiant padidėja daugelio hormonų - tiroksino, kortizono, progesterono - sekrecija, glikogeno skilimas kepenyse ir raumenyse, širdies susitraukimų dažnis ir stiprumas, osteodestrukcija ir atvirkštinis. padidėja vandens absorbcija nefrono kanalėliuose.
Adenilato ciklazės sistemos aktyvumas yra labai didelis – cAMP sintezė lemia dešimties tūkstantųjų signalo padidėjimą.
Veikiant cGMP, padidėja insulino sekrecija kasoje, histamino – putliųjų ląstelių, serotonino – trombocitų, sumažėja lygiųjų raumenų audinys.
Daugeliu atvejų, formuojantis receptoriaus-ligando kompleksui, pasikeičia membranos potencialas, o tai savo ruožtu lemia plazmalemos pralaidumą ir pokyčius. medžiagų apykaitos procesai narve.
Plazminėje membranoje yra specifinių receptorių, kurie reaguoja į fizinius veiksnius. Taigi fotosintetinėse bakterijose ląstelės paviršiuje yra chlorofilai, kurie reaguoja į šviesą. Šviesai jautrių gyvūnų plazminėje membranoje yra visa sistema fogoreceptorių baltymai-rodopsinai, kurių pagalba šviesos dirgiklis paverčiamas cheminiu signalu, o vėliau – elektriniu impulsu.
Ląstelių (plazmos) membrana yra pusiau pralaidi barjera, atskirianti ląstelių citoplazmą nuo aplinkos. Šiuo metu biologijoje remiamasi skystos mozaikos membranos modeliu, kurį aštuntajame dešimtmetyje pasiūlė J. Singeris ir G. Nicholsonas.
Šis modelis yra pagrįstas keliais pagrindiniais principais:
1. Membrana susideda iš dvigubo lipidų molekulių sluoksnio. Hidrofilinės, polinės molekulių dalys (galvutės) yra membranos išorėje, hidrofobinės, nepolinės dalys (uodega) – viduje.
2. Membraniniai baltymai yra mozaikiškai įterpti į lipidų dvisluoksnį sluoksnį. Kai kurie iš jų praeina pro membraną (jie vadinami integraliniais), kiti yra išoriniame arba vidiniame membranos paviršiuje (jie vadinami periferiniais).
3. Membranos lipidinė bazė turi skysčio savybių (pvz skystas aliejus) ir gali keisti jo tankį. Membranos klampumas priklauso nuo lipidų sudėties ir temperatūros. Šiuo atžvilgiu patys membraniniai baltymai ir lipidai gali laisvai judėti išilgai membranos ir jos viduje. Tai labai svarbi membranos savybė, užtikrinanti, kad membrana atliktų daug funkcijų (žr. toliau)
4. Daugumos tarpląstelinės membranos membranos organelės iš esmės yra panašūs į plazminę membraną.
5. Nepaisant bendros visų ląstelių membranų struktūros, baltymų ir lipidų sudėtis kiekvieno tipo ląstelėse ir ląstelės viduje skiriasi. Taip pat skiriasi išorinio ir vidinio lipidų sluoksnių sudėtis.
Plazminės membranos funkcijos
Membrana ląstelėje atlieka daugybę funkcijų, susijusių su ląstelės vientisumo palaikymu, medžiagų ir energijos mainų užtikrinimu tarp ląstelės ir aplinką ir sąveika su kitomis ląstelėmis.
transportavimo funkcija
Plazminė membrana pasižymi pusiau pralaidžio barjero savybėmis, kurios leidžia ir nepraleidžia tik tam tikroms medžiagoms ir molekulėms. Yra keli transportavimo per membraną tipai: pasyvus transportavimas, aktyvusis transportavimas, jonų pernešimas, transportavimas membraninėje pakuotėje (endo ir egzocitozė).
Pasyvus transportas yra medžiagų pernešimas per membraną iš didelės koncentracijos zonos į mažos koncentracijos sritį. Jis vykdomas dviem formomis: paprastos difuzijos ir „palengvintos“ difuzijos forma. Abu šie procesai nereikalauja energijos, yra gana lėti ir sustoja, kai medžiagų koncentracija abiejose membranos pusėse susilygina. Difuzijos greitis ir pati galimybė pernešti medžiagas per membraną priklauso (be koncentracijos) nuo daugelio veiksnių: temperatūros, molekulių dydžio ir gebėjimo ištirpti lipiduose. Riebaluose tirpios medžiagos per lipidų sluoksnius prasiskverbia lengvai, vandenyje – sunkiai. Membranoje yra specialios lipidų ir baltymų „poros“, pro kurias praeina difuzija. Paprasta difuzija yra gana lėtas procesas, o gamta pritaikė specialius membraninius nešiklius, kad pagreitintų transportavimą. Jie susijungia su transportuojama medžiaga ir perkelia ją iš vienos membranos pusės į kitą. Kiekvienai medžiagų grupei membrana turi turėti savo nešiklius. Toks procesas vadinamas „palengvinta difuzija“, vykstantis dešimt kartų greičiau nei paprasta difuzija.
Jonų pernešimas yra tam tikras pasyvus įkrautų jonų pernešimas, tačiau jis turi savo ypatybes. Ląstelės citoplazmoje esantys baltymai savo paviršiuje dažniausiai turi neigiamą krūvį, todėl ląstelėje susidaro tam tikras elektrinis fonas. Jei teigiamų (katijonų) arba neigiamų (anijonų) jonų srautai nukreipiami į ląstelę, tada teigiami jonai pateks daugiau, nes dalis katijonų bus surišti su citoplazmos baltymais, neutralizuojami ir sukurs papildomą skirtingai įkrautų jonų koncentracijų skirtumą ląstelės viduje ir išorėje. Jonams pernešti membranoje yra specialios jonų poros arba nešikliai.
aktyvus transportas- dauguma svarbi forma transportavimas ląstelei. Difuzija, jei ji tęsiasi pakankamai ilgai, lemia tai, kad pusiausvyra nusistovi abiejose membranos pusėse, o tai ląstelei yra nepriimtina - tai yra mirtis. Citoplazmos sudėtis ir intersticinis skystis labai skiriasi. Todėl ląstelėje yra „aktyvaus transporto“ sistema. Šiuo atveju molekulių pernešimas atliekamas prieš koncentracijos gradientą (iš mažos koncentracijos zonos į didelės koncentracijos zoną). Tam yra specialūs baltymų membranų kompleksai (joniniai ir molekuliniai kanalai), kurie veikia su energijos sąnaudomis. Iki 40% visos elemento pagaminamos energijos tenka transportavimo išlaidoms!
Transportavimas membraninėje pakuotėje (endo ir egzocitozė). Daugeliu atvejų ląstelė per membraną turi transportuoti ne atskiras molekules ar jonus, o ištisus molekulinius kompleksus ir net daleles. Jie neprasiskverbia pro membranos poras. Šiuo atžvilgiu yra specialus transportavimo būdas ląstelėje - transportavimas membraninėje pakuotėje. Kai kuriais atvejais atsiranda išorinės plazminės membranos išsikišimas, uždengiantis svetimą dalelę (pavyzdžiui: maisto dalelę ar bakteriją), uždaranti ją į membranos žiedą ir giliai panardinanti į ląstelės citoplazmą. Tokį procesą atrado garsus rusų mokslininkas, Nobelio premijos laureatas I.I.Mechnikovas ir pavadino fagocitoze. Šis reiškinys buvo atrastas studijuojant apsaugines savybes kraujo ląstelių, kurios sulaiko bakterijas ir jas neutralizuoja. Ląstelė sugauna smulkesnes skysčio daleles ir lašelius, suformuodama membranos invaginacijas – šis procesas vadinamas pinocitoze (gr. pinos – skystis).
Tiesą sakant, fagocitozė ir pinocitozė yra dažno reiškinio, būdingo daugumai ląstelių – endocitozės – atmainos. Atvirkštinis endocitozės procesas – bet kokių medžiagų ir produktų pašalinimas iš ląstelių vadinamas egzocitoze. Membraninio transportavimo pagrindu yra sukurtas visas ląstelių paslapčių ir hormonų išskyrimo ir įsisavinimo procesas. Svarbu pabrėžti, kad endo- ir egzocitozė yra energiją vartojantys procesai, todėl jie yra aktyvaus transporto rūšys.
Membraninių receptorių funkcija
Membraninių receptorių veikimo principai.
Ląstelė nuolat gauna signalus iš išorinės aplinkos apie įvairių signalinių molekulių buvimą ir turi adekvačiai reaguoti į šiuos signalus, perduodama informaciją iš jų į ląstelę. Tam į plazmos membraną įmontuoti specialūs receptorių kompleksai. Paprastai tai yra sudėtingi kelių baltymų molekulių dariniai (receptorių sudėtyje taip pat gali būti membraninių lipidų ir angliavandenių).
Visi įvairūs ląstelių membranų receptoriai turi keletą bendrų bruožų:
1. Receptoriai yra specifiniai – t.y. jungiasi tik su tam tikromis medžiagomis. Receptoriaus specifiškumą lemia jo molekulėje esančios „aktyvios vietos“ struktūra ir kitų molekulių gebėjimas prisijungti prie šios aktyvios vietos.
2. Signalo priėmimo ir perdavimo į membraną arba giliai į ląstelę procesas vyksta su didelėmis energijos sąnaudomis.
3. Pagal veikimo principą visus receptorius galima suskirstyti į tris grupes: receptorius tiesioginis veiksmas; netiesioginiai receptoriai ir kataliziniai receptoriai. Pirmuoju atveju molekulė prisijungia prie komplekso receptoriaus dalies ir perduoda signalą tiesiai į jonų kanalą. Antrojoje versijoje komplekso receptorių dalis perduoda signalą į jonų kanalą membranoje arba giliai į ląstelę per pagalbinių baltymų sistemą, vadinamą „antraisiais pasiuntiniais“. Trečiame variante komplekso receptorinė dalis, sąveikaudama su signaline molekule, yra aktyvuojama ir atlieka fermento funkcijas, taip paveikdama ląstelės funkcionavimą.
Izoliacinė membranos funkcija. Tarpląsteliniai kontaktai
IN daugialąstelis organizmas ląstelės nuolat kontaktuoja viena su kita, o tai kartais įgauna artimos sąveikos bruožų. Tai ypač akivaizdu audiniuose, kurie ribojasi su išorine aplinka (skerdenos, žarnos, Kvėpavimo takai) arba kai reikalingas glaudus ląstelių kontaktas koordinuotam darbui (lygieji ir širdies raumenys, nervų ląstelės ir pluoštai).
Yra keturių tipų tarpląstelinės jungtys: izoliacinės jungtys (tigth junction), sandarios jungtys (tarpo jungtis), desmosomos (desmosoma) ir lipniosios jungtys (adhezijos jungtis).
Kontaktų izoliavimas atitinka jų vardą - baltymų kompleksai gretimų ląstelių membranose yra sujungtos taip, kad per šį kontaktą nepraeitų jokios molekulės ir jonai. Tvirtos jungtys leidžia mažoms molekulėms ir jonams prasiskverbti pro juos, o per jas gali vykti mainai tarp ląstelių. Visų pirma įtemptas sandūras pritaiko nervingi ir raumenų ląstelės elektros signalams perduoti. Desmosomos yra skirtos laikyti ląstelių grupę viena su kita arba prie kito substrato.
Jie gali greitai išnykti ir vėl atsirasti. Sukibimo kontaktai taip pat skirti pritvirtinti ląsteles viena prie kitos. Jie yra labiau labilūs ir gali greitai susiformuoti ir prireikus išnykti.
Branduolys yra atsakingas už genetinės medžiagos, įrašytos DNR, saugojimą, taip pat kontroliuoja visus ląstelės procesus. Citoplazmoje yra organelių, kurių kiekviena atlieka savo funkcijas, tokias kaip, pavyzdžiui, sintezė. organinės medžiagos, virškinimas ir tt Ir apie paskutinį komponentą mes kalbėsime išsamiau šiame straipsnyje.
biologijoje?
kalbantis paprasta kalba, tai apvalkalas. Tačiau jis ne visada yra visiškai nepralaidus. Beveik visada leidžiama pernešti tam tikras medžiagas per membraną.
Citologijoje membranas galima suskirstyti į du pagrindinius tipus. Pirmasis yra plazminė membrana, dengianti ląstelę. Antrasis yra organelių membranos. Yra organelių, turinčių vieną ar dvi membranas. Vienos membranos ląstelės apima endoplazminį tinklą, vakuoles ir lizosomas. Plastidės ir mitochondrijos priklauso dviejų membranų.
Be to, membranos gali būti organelių viduje. Paprastai tai yra dviejų membranų organelių vidinės membranos dariniai.
Kaip išdėstytos dviejų membranų organelių membranos?
Plastidės ir mitochondrijos turi dvi membranas. Abiejų organelių išorinė membrana yra lygi, tačiau vidinė formuoja struktūras, būtinas organoido funkcionavimui.
Taigi, mitochondrijų apvalkalas turi išsikišimus į vidų - kriaušes arba keteras. Ant jų vyksta cheminių reakcijų ciklas, būtinas ląstelių kvėpavimui.
Chloroplastų vidinės membranos dariniai yra disko formos maišeliai – tilakoidai. Jie surenkami į krūvas – grūdus. Atskiros granos sujungiamos viena su kita lamelių pagalba – ilgos struktūros taip pat susidaro iš membranų.
Vienos membranos organelių membranų sandara
Šios organelės turi tik vieną membraną. Paprastai tai yra lygi membrana, sudaryta iš lipidų ir baltymų.
Ląstelės plazminės membranos struktūros ypatumai
Membrana sudaryta iš medžiagų, tokių kaip lipidai ir baltymai. Plazminės membranos struktūra numato 7-11 nanometrų storį. Didžiąją membranos dalį sudaro lipidai. 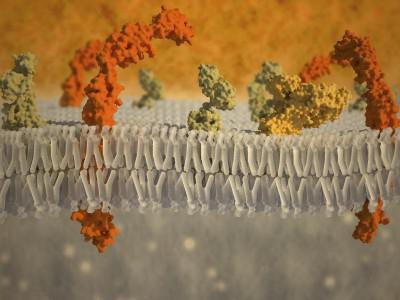
Plazminės membranos struktūra numato, kad joje yra du sluoksniai. Pirmasis yra dvigubas fosfolipidų sluoksnis, o antrasis - baltymų sluoksnis.
Plazmos membranos lipidai
Lipidai, sudarantys plazmos membraną, skirstomi į tris grupes: steroidus, sfingofosfolipidus ir glicerofosfolipidus. Pastarosios molekulės sudėtyje yra trihidroksilo alkoholio glicerolio liekana, kurioje dviejų hidroksilo grupių vandenilio atomai pakeisti riebalų rūgščių grandinėmis, o trečiosios hidroksilo grupės vandenilio atomas yra pakeistas fosforo rūgšties liekana. , prie kurio, savo ruožtu, yra prijungtas vienos iš azotinių bazių likutis.
Glicerofosfolipido molekulę galima suskirstyti į dvi dalis: galvą ir uodegas. Galva yra hidrofilinė (tai yra, ji ištirpsta vandenyje), o uodegos yra hidrofobinės (jos atstumia vandenį, bet ištirpsta organiniuose tirpikliuose). Dėl šios struktūros glicerofosfolipidų molekulė gali būti vadinama amfifiline, tai yra, tuo pačiu metu ir hidrofobine, ir hidrofiline.
Sfingofosfolipidai yra panašūs cheminė struktūra glicerofosfolipidams. Tačiau jie skiriasi nuo aukščiau paminėtų tuo, kad jų sudėtyje vietoj glicerolio liekanos yra sfingozino alkoholio liekanos. Jų molekulės taip pat turi galvas ir uodegas.
Žemiau pateiktame paveikslėlyje aiškiai parodyta plazminės membranos struktūra.
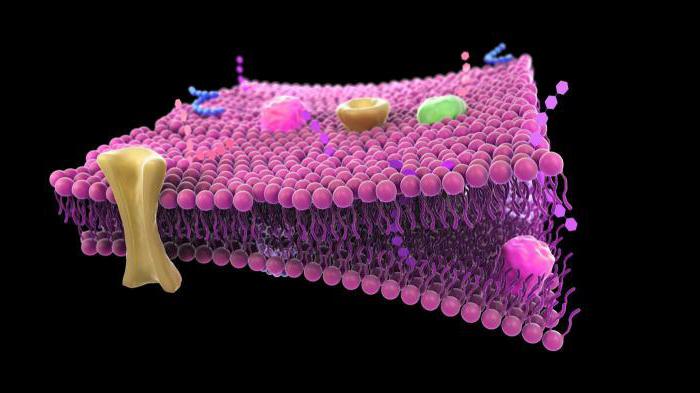
Plazmos membranos baltymai
Kalbant apie baltymus, sudarančius plazmos membranos struktūrą, tai daugiausia yra glikoproteinai.
Priklausomai nuo jų vietos apvalkale, jas galima suskirstyti į dvi grupes: periferines ir vientisas. Pirmieji yra tie, kurie yra membranos paviršiuje, o antrieji yra tie, kurie prasiskverbia per visą membranos storį ir yra lipidų sluoksnio viduje.
Priklausomai nuo baltymų atliekamų funkcijų, juos galima suskirstyti į keturias grupes: fermentus, struktūrinius, transportavimo ir receptorių.
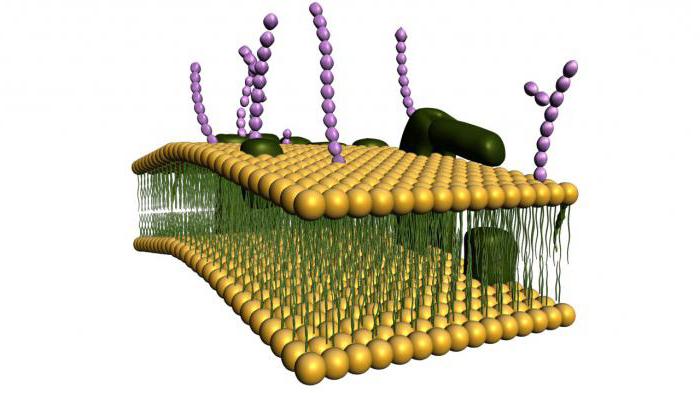
Visi baltymai, esantys plazmos membranos struktūroje, nėra chemiškai susiję su fosfolipidais. Todėl jie gali laisvai judėti pagrindiniame membranos sluoksnyje, burtis į grupes ir pan.. Štai kodėl ląstelės plazminės membranos struktūra negali būti vadinama statine. Ji yra dinamiška, nes nuolat keičiasi.
Koks yra ląstelės membranos vaidmuo?
Plazminės membranos struktūra leidžia jai susidoroti su penkiomis funkcijomis.
Pirmasis ir pagrindinis yra citoplazmos apribojimas. Dėl to ląstelė turi nuolatinė forma ir dydis. Šią funkciją užtikrina tai, kad plazminė membrana yra tvirta ir elastinga.
Antrasis vaidmuo – aprūpinimas Dėl savo elastingumo plazminės membranos gali susidaryti ataugų ir raukšlių sandūroje.
Kita funkcija ląstelių sienelės- transportas. Jį suteikia specialūs baltymai. Ačiū jiems tinkamų medžiagų galima vežtis į narvą, o iš jo išmesti nereikalingus.
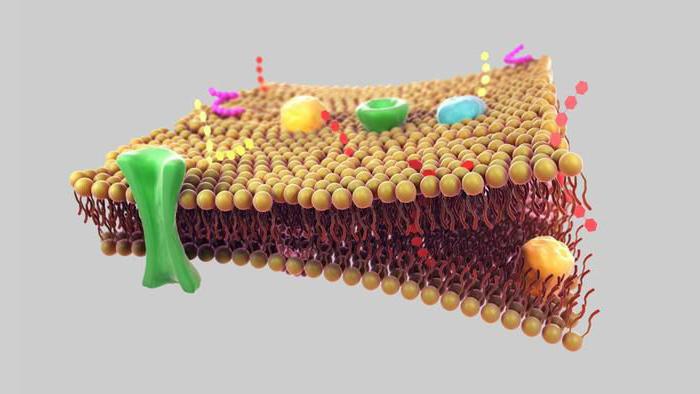
Be to, plazminė membrana atlieka fermentinę funkciją. Tai taip pat atliekama baltymų dėka.
Ir paskutinė funkcija yra signalizacija. Dėl to, kad baltymai, veikiami tam tikrų sąlygų, gali pakeisti savo erdvinę struktūrą, plazminė membrana gali siųsti signalus į ląsteles.
Dabar žinote viską apie membranas: kas yra membrana biologijoje, kas tai yra, kaip yra išdėstyta plazminė membrana ir organoidinės membranos, kokias funkcijas jos atlieka.
4 paskaita.
Valandų skaičius: 2
plazmos membrana
1.
2.
3. Tarpląsteliniai kontaktai.
1. Plazminės membranos struktūra
Plazminė membrana arba plazmolema, yra paviršiaus periferinė struktūra, kuri ribojaląstelę iš išorės ir užtikrina jos ryšį su kitomis ląstelėmis bei tarpląsteline aplinka. Jis turi storįapie 10 nm. Tarp kitų ląstelių membranų plazmolema yra storiausia. Chemiškai plazminė membrana yra lipoproteinų kompleksas. Pagrindiniai komponentai yra lipidai (apie 40%), baltymai (daugiau nei 60%) ir angliavandeniai (apie 2-10%).
Lipidai yra didelė grupė organinės medžiagos, kurios blogai tirpsta vandenyje (hidrofobiškumas) ir gerai tirpsta organiniuose tirpikliuose ir riebaluose (lipofiliškumas).Tipiški lipidai, randami plazmos membranoje, yra fosfolipidai, sfingomielinai ir cholesterolis. IN augalų ląstelės cholesterolis pakeičiamas fitosteroliu. Autorius biologinis vaidmuo plazmos membranos baltymus galima suskirstyti į fermentiniai baltymai, receptoriai ir struktūriniai baltymai. Plazmalemmos angliavandeniai yra surištos būsenos plazmalemos dalis (glikolipidai ir glikoproteinai).
Šiuo metu tai visuotinai priimta skysčio mozaikinis biologinės membranos struktūros modelis. Pagal šį modelį membranos struktūrinį pagrindą sudaro dvigubas fosfolipidų sluoksnis, inkrustuotas baltymais. Molekulių uodegos susiduria viena su kita dvigubu sluoksniu, o poliarinės galvutės lieka išorėje ir sudaro hidrofilinius paviršius. Baltymų molekulės nesudaro ištisinio sluoksnio, jos yra lipidiniame sluoksnyje, pasinerdamos į skirtingą gylį (yra periferiniai baltymai, dalis baltymų prasiskverbia pro membraną, dalis yra panardinta į lipidų sluoksnį). Dauguma baltymų nėra susiję su membranos lipidais; jie tarsi plūduriuoja „lipidiniame ežere“. Todėl baltymų molekulės gali judėti išilgai membranos, burtis į grupes arba, atvirkščiai, išsisklaidyti membranos paviršiuje. Tai rodo, kad plazminė membrana nėra statinis, sustingęs darinys.
Už plazmalemos yra epimembraninis sluoksnis - glikokaliksas. Šio sluoksnio storis apie 3-4 nm. Glikokaliksas randamas beveik visose gyvūnų ląstelėse. Jis yra susijęs su plazmine membrana glikoproteinų kompleksas. Angliavandeniai sudaro ilgas, išsišakojusias polisacharidų grandines, susijusias su plazmos membranos baltymais ir lipidais. Glikokalikse gali būti fermentų baltymų, dalyvaujančių tarpląsteliniame įvairių medžiagų skaidyme. Fermentinio aktyvumo produktai (aminorūgštys, nukleotidai, riebalų rūgštis ir tt) yra pernešami per plazmos membraną ir absorbuojami ląstelių.
Plazminė membrana nuolat atnaujinama. Tai atsitinka, kai maži burbuliukai nuo jos paviršiaus į ląstelę patenka, o vakuoles iš ląstelės vidaus įterpiant į membraną. Taigi ląstelėje vyksta nuolatinis membraninių elementų srautas: iš plazminės membranos į citoplazmą (endocitozė) ir membraninių struktūrų tekėjimą iš citoplazmos į ląstelės paviršių (egzocitozė). Membranų cirkuliacijoje pagrindinis vaidmuo skiriamas Golgi komplekso membraninių vakuolių sistemai.
4. Plazminės membranos funkcijos. Medžiagų transportavimo per plazmalemą mechanizmai. Plazlemos receptorių funkcija
Plazminė membrana atlieka keletą svarbių funkcijų:
1) Barjeras.Plazminės membranos barjerinė funkcija yraribojant laisvą medžiagų difuziją iš ląstelės į ląstelę, užkertant keliąvandenyje tirpaus ląstelės turinio sukimosi nutekėjimas. Bet kadangikaip ląstelė turi gauti reikiamų maistinių medžiagų, tudalyti galutinius medžiagų apykaitos produktus, reguliuoti tarpląstelinįjonų koncentracijos, tada ji suformavo specialius medžiagų pernešimo per ląstelės membraną mechanizmus.
2) Transportas.Transporto funkcija yra Įvairių medžiagų patekimo į ląstelę ir iš jos išėjimo užtikrinimas. Svarbi membranos savybė yra selektyvus pralaidumas, arba pusiau pralaidumas. Jis lengvai praleidžia vandenį ir tirpsta vandenyjedujas ir atbaido polines molekules, tokias kaip gliukozė arba amino rūgštys.
Yra keletas medžiagų transportavimo per membraną mechanizmų:
pasyvus transportas;
aktyvus transportas;
transportavimas membraninėje pakuotėje.
Pasyvus transportas. difuzija -tai terpės dalelių judėjimas, vedantis į pernešimąmedžiagos iš vietovės, kurioje jo koncentracija yra didelė, į vietą, kurioje koncentracija mažasijos. Difuzijos transportavimo metu membrana veikia kaip osmosinis barjeras. Sklaidos greitis priklauso nuo vertėsmolekulės ir jų santykinis tirpumas riebaluose. Kuo mažiau kartųmolekulių matai ir kuo daugiau jos tirpsta riebaluose (lipofilinės), tuo greičiau judės per lipidų dvisluoksnį sluoksnį.Difuzija gali būti neutralus(neapmokestinto pervedimasmolekulės) ir lengvas(specialių baltymų pagalbanosis). Supaprastinta difuzija yra greitesnė nei neutrali difuzija.Maksimalus prasiskverbimasvanduo turi savybękaip jo molekulės yra mažos ir neįkrautos. Vandens difuzija per ląstelesmembrana vadinama osmo šamasDaroma prielaida, kad ląstelėjemembrana prasiskverbimuivandens ir kai kurių jonųyra specialios „poros“. Jų skaičiusmažas, o skersmuo yraapie 0,3-0,8 nm. Greičiausiai pasklinda per membraną gerai, lengvai tirpsta lipiduose dvisluoksnis molekulės sluoksnis, pavyzdžiui, O ir neįkrautos polinės molekulėsly mažo skersmens (SO, mo chevin).
Polinių molekulių transportavimas (sucukrus, aminorūgštys), įgyvendintapristatomas naudojant specialų membraninį transportąbaltymai vadinami palengvinta difuzija. Tokie baltymai yrarandami visų tipų biologinėse membranose ir kiekviena specifinė ny baltymas skirtas perduoti tam tikros klasės molekules sa. Transporto baltymai yra transmembraniniai, jų polipeptidinė grandinė kelis kartus kerta lipidų dvisluoksnį, susidarant jis turi perėjimus. Taip užtikrinamas specifinių perdavimasmedžiagos per membraną be tiesioginio kontakto su ja.Yra dvi pagrindinės transporto baltymų klasės: voverės - vežėjai (vežėjai) Ir kanalo formavimas baltymai (baltiki kanalus). Nešėjai baltymai perneša molekules per membraną, pirmiausia pakeisdami jų konfigūraciją. Kanalą formuojantys baltymai sudaro užpildytas membranas porų vanduo. Kai poros yra atviros, tam tikrų medžiagų molekulės(dažniausiai per juos praeina tinkamo dydžio ir krūvio neorganiniai jonai). Jei gabenamos medžiagos molekulė neturi krūvio, tada transportavimo kryptį lemia koncentracijos gradientas. Jei molekulė yra įkrauta, tada ją transportuojant, be gradiento, kon koncentracija, veikia ir membranos elektros krūvis (membranapotencialus). Vidinė plazmolemos pusė dažniausiai įkraunama iš neigiamas išorės atžvilgiu. Membranos potencialas palengvina teigiamo krūvio jonų prasiskverbimą į ląstelę ir neleidžia prasiskverbti neigiamo krūvio jonams.
aktyvus transportas. Aktyvus transportavimas yra medžiagų judėjimas prieš elektrocheminį gradientą. Jį visada vykdo trans baltymai.nešikai ir glaudžiai susiję zan su energijos šaltiniuva. Baltymų pernešime chiki turi sklypus surišimas su transportutinkama medžiaga. Kuo daugiau tokių tkov bendrauja su daiktaiskuo didesnis greitistransporto augimas. Atrankinis vienos medžiagos perkėlimas vadinamas uniportas. Atliekamas kelių medžiagų perkėlimas kotran sporto sistemos. Jei perkėlimas vyksta viena kryptimi -Tai simportas, jei priešingai antiportas. Taigi,pavyzdžiui, gliukozė iš ekstraląstelinio skysčio į ląstelę pernešama vienportu būdu. Gliukozės pernešimas ir Na 4 iš žarnyno trakto arbaInkstų kanalėliai, atitinkamai, į žarnyno ląsteles arba kraują vyksta simporiškai, o C1 ~ ir HCO pernešimas yra antiportas. Daroma prielaida, kad perdavimo metu grįžtama konformacinė keičiasi konvejeris, leidžiantis judėti su juo prijungtoms medžiagoms.
Transportui naudojamo baltymo nešiklio pavyzdysmedžiagų, ATP hidrolizės metu išsiskirianti energija yraNa + -K + siurblys, randama visų ląstelių plazminėje membranoje. Na+-K Siurblys veikia anti-port, siurbimo principu wai Na "iš ląstelės ir į ląstelės vidų prieš jų elektrocheminį poveikį gradientai. Gradientas Na+ sukuria osmosinį slėgį, palaiko ląstelių tūrį ir užtikrina cukrų bei aminorūgščių transportavimąnorūgštys. Šio siurblio veikimui sunaudojama trečdalis visos gyvybinei ląstelių veiklai reikalingos energijos.Tiriant veikimo mechanizmą Na + - K + buvo sumontuotas siurblysyra žinoma, kad tai yra ATPazės fermentas ir transmembraninis fermentas tegralis baltymas. Esant Na+ ir ATP veikiant ATP-galinis fosfatas atskiriamas nuo ATP ir prijungiamas prie liekanosasparto rūgštis ant ATPazės molekulės. Phos ATPazės molekulėforiluojamas, keičia savo konfigūraciją ir Na+ yra kilęs iš ląstelės. Po pasitraukimo Na K" visada pernešamas iš ląstelės į ląstelę. Tam, dalyvaujant K, iš ATPazės atskaldomas anksčiau prijungtas fosfatas. Fermentas defosforilinamas, atkuria jo konfigūraciją, o K 1 "siurbiamas" į ląstelę.
ATPazę sudaro du subvienetai - didelis ir mažas.Didelis subvienetas susideda iš tūkstančių aminorūgščių liekanų,kelis kartus kertant dvisluoksnį. Jis turi katalizatorių aktyvumą ir gali būti grįžtamai fosforilinti ir defosforaižyti. Didelis subvienetas citoplazminėje pusėjeneturi rišamųjų vietų Na+ ir ATP, o išorėje -K + ir ouabaino surišimo vietos. Mažasis subvienetas yraglikoproteinas ir jo funkcija dar nežinoma.
Na+-K siurblys turi elektrogeninį poveikį. Jis pašalina tristeigiamai įkrautas jonas Na f iš kameros ir įveda dujonas K Dėl to per membraną teka srovė, suformuodama elektronąric potencialas su neigiama verte vidinėje ląstelės dalyje, palyginti su jos išoriniu paviršiumi. Na "-K + siurblys reguliuoja ląstelių tūrį, kontroliuoja medžiagų koncentracijąląstelės viduje, palaiko osmosinį slėgį, dalyvauja kuriant membraninį potencialą.
Transportavimas membraninėje pakuotėje. Makromolekulių (baltymų, nukleorūgščių) pernešimas per membranąpartija, polisacharidai, lipoproteinai) ir kitos dalelės yra atliekamos nuosekliai formuojant ir susiliejantmembranos surištos pūslelės (pūslelės). Vezikulinis transportavimo procesasji praeina per du etapus. Pradžiojepūslelių membrana ir plazmolemasulipti ir tada sujungti.Norint pereiti 2 etapą, būtinadimo, kad vandens molekulės būtų jūsyra perpildyti sąveikaujančių lipidų dvigubų sluoksnių, kurie artėja vienas prie kito 1-5 nm atstumu. Mano kad šis procesas suaktyvintasypatingas sulieti baltymai(Jie kol kas izoliuoti tik nuo virusų). Vezikulinis transportas turisvarbi savybė – absorbuojamos arba išskiriamos makromolekulės,yra pūslelėse, dažniausiai nemaišoma su kitais makromoliaisculae arba ląstelių organelės. Pu pūslelės gali susijungti su specifinėmis membranos, kurios suteikiachivaet keitimasis makromolekulėmis tarpdu tarpląstelinės erdvės irląstelės turinys. Panašiaimakromolekulės perkeliamos iš vieno ląstelės skyriaus į kitą.
Makromolekulių ir dalelių pernešimas į ląstelę vadinamas endo citozė.Tokiu atveju gabenamos medžiagos yra apgaubtos arbataplazmos membrana susidaro pūslelė (vakuolė), kurikuris juda į ląstelę. Priklausomai nuo vaizdo dydžioburbuliukai, yra dviejų tipų endocitozė - pinocitozė ir fagocitozė.
pinocitozėužtikrina skysčio ir ištirpusio absorbcijąmedžiagos mažų burbuliukų pavidalu ( d =150 nm). fagocitozė -yra didelių dalelių, mikroorganizmų absorbcijašauksmas ar organelių, ląstelių fragmentai. Tuo pačiu metu jie formuojasiXia didelės pūslelės, fagosomos arba vakuolės ( d –250 nm ar daugiau). At pirmuonių fagocitinė funkcija – mitybos forma. Žinduolių fagocitinę funkciją atlieka makrofagai ir nutasprofiliai, apsaugantys organizmą nuo infekcijos, sugerdami įsiveržusius mikrobus. Makrofagai taip pat dalyvauja panaudojimesenų ar pažeistų ląstelių ir jų fragmentų (kūnožmogaus makrofagai kasdien suryja daugiau nei 100 senų eritritųRocitai). Fagocitozė prasideda tik prarijus dalelęjungiasi prie fagocito paviršiaus ir aktyvuoja specializuotąnye receptorių ląstelės. Dalelių susiejimas su specifine remembraniniai receptoriai sukelia pseudopodijų susidarymą, kurisrugiai apgaubia dalelę ir, susiliedami kraštuose, sudaro burbulą -fagosoma.Tinkamas fagosomų susidarymas ir fagocitozėvaikšto tik tada, kai apgaubia dalelęnuolat kontaktuoja su plazmalemos receptoriais, tarsi „sustabdęs mirksi žaibas“.
Nemaža dalis medžiagos, kurią ląstelė sugeria endocitozė, patenka į lizosomas. Didelės dalelės, įskaitanttikiuosi fagosomos kurios vėliau susilieja su lizosomomis ir susidaro fagolizosomos. Skystis ir makromolekulės absorbuojamos perpinocitozė, iš pradžių perkeliama į endosomas, kuriossusilieja su lizosomomis, kad susidarytų endolizosomos. Esu esantis įvairių hidrolizinių fermentų, esančių lizosomosero sunaikinti makromolekules. hidrolizės produktai (aminorūgštispartijos, cukrūs, nukleotidai) pernešami iš lizosomų į citozolį, kur juos naudoja ląstelė. Dauguma membranos komponentų endocitinės pūslelės iš fagosomų ir endosomų eksocitozės būdu grįžta į plazmos membraną ir ten vėl išnykstalyse. Pagrindinė endocitozės biologinė reikšmė yra yra statybinių blokų gavimas dėl tarpląstelinio makromolekulių virškinimas lizosomose.
Medžiagų absorbcija eukariotinėse ląstelėse prasidedacializuoti plazminės membranos regionai, vadinamiejimes esame X ribojamos duobės. Ant elektroninių mikrografijųduobutės atrodo kaip plazminės membranos, citoplazmos invaginacijoskurio matinė pusė padengta pluoštiniu sluoksniu. sluoksnis panašusribotųsi mažomis duobėmis plaz malemmas. Duobės užima apie 2 procvirš ląstelės membranos paviršiausmes eukariotai. Per minutę duobės auga vis giliau Xia, yra įtraukiami į narvą, o tada susiaurėjimas prie pagrindo, atskilęs,formuojasi kutais pūslelės.Nustatyta, kad nuomatinės membranos fibroblastaitov per vieną minutęapie ketvirtadalįmembranos ribojamos pu formos zyrkovas. Burbulai greitai praranda savo sieną ir įgyti keliągebėjimas susilieti su lizosoma.
Gali būti endocitozė nespecifinis(konstitucinis)Ir specifinis(receptorius).At nespecifinė endocitozė ląstelė perimasugeria jai visiškai svetimas medžiagas, pavyzdžiui, suodžių daleles,dažikliai. Pirma, dalelės nusėda ant glikokalikso plazmolema. Ypač gerai nusodinamas (adsorbuojamas). teigiamai įkrautas baltymų grupes, nes glikokaliksas neša neigiamas krūvis. Tada pasikeičia ląstelių morfologijamembranos. Jis gali nuskęsti, sudarydamas invaginijas(invaginacija) arba, priešingai, formuoti ataugas, kurios tarsi sumuojasi, atskirdamos mažus tūrius skysta aplinka. Būdingesnis invaginacijų formavimasisžarnyno epitelio ląstelėms, ameboms ir ataugoms - fagocitams ir fibroblastai. Šiuos procesus gali blokuoti inhibitoriaikvėpavimas. Susidariusios pūslelės yra pirminės endosomos, kurios gali susijungti vienas kitą, didėja dydis. Ateityje jie prisijungs susimaišo su lizosomomis, virsta endolizosoma – virškintuvu nauja vakuolė. Skystos fazės nespecifinės pinocitozės intensyvumas ikipašėlusiai aukštai. Makrofagai susidaro iki 125, o epitelio ląstelės smulkiaižarnyne iki tūkstančio pinosomų per minutę. Pinosomų gausa lemia tai, kad plazmalemma greitai išleidžiama formuojant kelismažų vakuolių buvimas. Membranos atsistatymas vyksta gana greitai.tro perdirbimo metu egzocitozės procese dėl grįžimo wakuoles ir jų įtraukimas į plazmalemą. Makrofaguose visa plazmaKlasikinė membrana pakeičiama per 30 minučių, o fibroblastuose – per 2 valandas.
Veiksmingesnis būdas pasisavinti iš tarpląstelinio skysčiokaulams būdingos makromolekulės yra specifinis en docitozė(tarpininkauja receptoriai). Makromolekulės šiamejungiasi prie paviršiuje esančių papildomų receptoriųląstelės kaupiasi apribotoje duobėje, o tada, suformuodamos endosomą, panardinamos į citozolį. Receptorių endocitozė užtikrina specifinių makromolekulių kaupimąsi prie jo receptoriaus.Molekulės, kurios plazmalemos paviršiuje jungiasi su receptoriumiTorus vadinami ligandai. Receptoriaus pagalba endocitozė daugelyje gyvūnų ląstelių yra absorbcijaekstraląstelinis cholesterolis aplinką.
Plazminė membrana dalyvauja pašalinant medžiagas iš ląstelės (egzocitozė). Tokiu atveju vakuolės artėja prie plazmalemos. Sąlyčio taškuose plazmolema ir vakuolės membrana susilieja ir vakuolės turinys patenka į aplinką.Kai kurių pirmuonių ląstelės membranoje yra iš anksto nustatytos egzocitozės vietos. Taigi, plazmos membranoje kai kurie ciliariniai blakstienos turi tam tikras sritis, kuriose yra teisingai išsidėstę dideli vientisų baltymų rutuliukai. Atblakstienų mukocistos ir trichocistos visiškai paruoštos sekrecijai, viršutinėje plazmalemos dalyje yra vientisų rutuliukų vainikasbaltymai. Šios soprikos mukocistų ir trichocistų membranos skyriaiprilipti prie ląstelės paviršiaus.Neutrofiluose stebima savita egzocitozė. Jie spotam tikromis sąlygomis gali patekti į aplinkąišpūskite savo lizosomas. Kai kuriais atvejais susidaro nedidelės plazmalemos ataugos, kuriose yra lizosomų, kurios vėliau nutrūksta ir patenka į aplinką. Kitais atvejais plazmalema įsiskverbia giliai į ląstelę ir užfiksuoja lizosomas, esančias nyh toli nuo ląstelės paviršiaus.
Endocitozės ir egzocitozės procesai atliekami dalyvaujant citoplazmos fibrilinių komponentų sistemai, susijusiai su plazmolema.
Plazlemos receptorių funkcija. Šitas Pagrindinis, universalus visoms ląstelėms, yra replazmolemos receptorių funkcija. Tai apibrėžia sąveikąląstelės tarpusavyje ir su aplinka.
Visą informacinių tarpląstelinių sąveikų įvairovę galima schematiškai pavaizduoti kaip nuoseklią grandinęsignalo-receptoriaus-antrojo pasiuntinio-atsako reakcijos (koncepcija signalas-atsakymas).Signalai perduoda informaciją iš ląstelės į ląstelęnye molekulių, kurios gaminamos kai kuriose ląstelėse ir specialiosfiziškai paveikti kitus, jautrius signalui (ląsteles-mi sheni). Signalo molekulė - pirminis tarpininkas privalomas etsya su receptoriais, esančiais tikslinėse ląstelėse, reaguoja tik tam tikriems signalams. Signalų molekulės - ligandai - prieiti prie jų receptorių kaip prie spynos rakto. ligandas-mi membraniniams receptoriams (plazmalemos receptoriams) yrahidrofilinės molekulės, peptidiniai hormonai, neuromedija tori, citokinai, antikūnai, o branduoliniams receptoriams – riebalai tirpios molekulės, steroidiniai ir skydliaukės hormonai, vitaminas DKaip receptoriai viršujeląstelės gali veikti kaip baltymaimembranos arba glikokalitiniai elementaica – polisacharidai ir glikoproteinai.Manoma, kad jautrusatskirti medžiagų sritis, išsklaidytisan išilgai ląstelės paviršiaus arbabranes į mažas juosteles. Taip, įjungtaprokariotinių ląstelių paviršiusir gyvūnų ląstelėms yra ribosfiksuotas skaičius vietų, kuriose jie galisurišti virusines daleles. memasvoverės (nešioklės ir kanosly) mokytis, bendrauti ir perduotidėvėti tik tam tikras medžiagas.Ląstelių receptoriai dalyvaujasignalų perdavimas iš ląstelės paviršiaus į vidų.Įvairovė ir specifiškumasreceptorių griovelis ląstelės paviršiujeveda prie labai sudėtingos sistemosesame žymekliai, skirti atskirtisavo ląsteles iš nepažįstamų žmonių. Panašios ląstelėssąveikauja vienas su kitu, jų paviršiai gali sulipti (konjugacija atpirmuonys, audinių formavimasis daugialąsčiuose organizmuose). Ląstelės nesuvokiabendri žymekliai, taip pat tie, kurie skiriasideterminantinių žymenų borasyra suplėšyti arba atmesti.Susidarius receptoriaus-ligando kompleksui,transmembraniniai baltymai: baltymų konverteris, baltymų stipriklis.Dėl to receptorius keičia savo konformaciją ir sąveikąne su antrojo pasiuntinio ląsteliniu pirmtaku ka - pasiuntinys.Pasiuntiniai gali būti jonizuotas kalcis, fosfolipasC – adenilato ciklazė, guanilato ciklazė. Pasiuntinio paveiktassintezėje dalyvaujančių fermentų aktyvinimas cikliniai monofosfatai – AMP arba HMF. Pastarieji keičia turtąDviejų tipų baltymų kinazės fermentų buvimas ląstelės citoplazmoje, dėl kurio fosforilinami daugybė intraląstelinių baltymų.
Dažniausias cAMP susidarymas, veikiantpadidėja daugelio hormonų - tiroksino, kortizono, progesterono - sekrecija, padidėja glikogeno skilimas kepenyse ir raumenyse,širdies ritmas ir jėga, osteodestrukcija, atvirkštinė vandens absorbcija nefrono kanalėliuose.
Adenilato ciklazės sistemos aktyvumas yra labai didelis – cAMP sintezė lemia dešimties tūkstantųjų signalo padidėjimą.
Veikiant cGMP, insulino sekrecija iš kasos, histamino – putliųjų ląstelių, serotonino.bocitų, sumažėja lygiųjų raumenų audinys.
Daugeliu atvejų susiformavus receptoriaus-ligando kompleksuipasikeičia membranos potencialas, o tai savo ruožtu lemia plazmos lemos ir medžiagų apykaitos pralaidumą.kai kurie procesai ląstelėje.
Plazminėje membranoje yra specifinių receptorių tori reaguoja į fizinius veiksnius. Taigi fotosintetinėse bakterijose chlorofilai yra ląstelės paviršiuje,reaguoja į šviesą. Šviesai jautriems gyvūnams plazmojeKaustinėje membranoje yra visa fogoreceptorių baltymų sistema -rodopsinų, kurių pagalba transformuojasi šviesos dirgiklis paverčiamas cheminiu signalu, o vėliau – elektriniu impulsu.
3. Tarpląsteliniai kontaktai
Daugialąsčių gyvūnų formavime dalyvauja plazmolema tarpląsteliniai ryšiai užtikrina tarpląstelinę sąveiką. Yra keletas tokių konstrukcijų tipų.
§ Paprastas kontaktas.Paprastas kontaktas randamas tarp daugumos įvairios kilmės greta esančių ląstelių. Rodo gretimų ląstelių plazminių membranų konvergenciją 15-20 nm atstumu. Tokiu atveju atsiranda gretimų ląstelių glikokalikso sluoksnių sąveika.
§ Tvirtas (uždarantis) kontaktas. Su tokia jungtimi abiejų plazminių membranų išoriniai sluoksniai yra kuo arčiau. Suartėjimas yra toks tankus, kad vyksta savotiškas dviejų gretimų ląstelių plazminių membranų dalių susiliejimas. Membranų susiliejimas nevyksta visoje sandaraus kontakto srityje, o yra taškinio membranų konvergencijos serija. Glaudaus kontakto vaidmuo yra mechaniškai sujungti ląsteles viena su kita. Ši sritis yra nepralaidi makromolekulėms ir jonams, todėl užrakina, atriboja tarpląstelinius tarpus (o kartu su jais ir vidinę kūno aplinką) nuo išorinės aplinkos.
§ Sukibimo lopinėlis arba desmosoma. Desmosoma yra mažas plotas iki 0,5 µm skersmens. Desmosomos zonoje citoplazmos pusėje yra plonų fibrilių sritis. Funkcinis desmosomų vaidmuo daugiausia susijęs su mechaniniu ryšiu tarp ląstelių.
§ Kontakto tarpas arba ryšys. Esant tokio tipo kontaktui, gretimų ląstelių plazminės membranos yra atskiriamos 2–3 nm tarpu 0,5–3 µm atstumu. Plazmolemų struktūroje yra specialūs baltymų kompleksai (konneksonai). Vienai ląstelės plazminės membranos jungčiai tiksliai priešinasi gretimos ląstelės plazminės membranos jungtis. Dėl to kanalas susidaro iš vienos ląstelės į kitą. Konneksonai gali susitraukti, keisdami vidinio kanalo skersmenį ir taip dalyvauti reguliuojant molekulių transportavimą tarp ląstelių. Šio tipo ryšys randamas visose audinių grupėse. Tarpo jungties funkcinis vaidmuo yra pernešti jonus ir mažas molekules iš ląstelės į ląstelę. Taigi širdies raumenyje sužadinimas, pagrįstas jonų pralaidumo keitimo procesu, per ryšį perduodamas iš ląstelės į ląstelę.
§ Sinapsinis kontaktas arba sinapsė. Sinapsės yra dviejų ląstelių kontakto sritys, kurios specializuojasi vienpusiam sužadinimo ar slopinimo perdavimui iš vieno elemento į kitą. Šio tipo ryšys būdingas nerviniam audiniui ir atsiranda tiek tarp dviejų neuronų, tiek tarp neurono ir kokio nors kito elemento. Šių ląstelių membranas skiria tarpląstelinė erdvė – apie 20-30 nm pločio sinapsinis plyšys. Vienos ląstelės sinapsinio kontakto srityje esanti membrana vadinama presinaptine, kitos - postsinaptine. Netoli presinapsinės membranos atsiskleidžia daugybė mažų vakuolių (sinapsinių pūslelių), kuriose yra neurotransmiteris. Kai praeina nervinis impulsas, sinapsinės pūslelės išstumia neuromediatorių į sinapsinį plyšį. Mediatorius sąveikauja su postsinapsinės membranos receptorių vietomis, o tai galiausiai lemia nervinio impulso perdavimą. Be nervinio impulso perdavimo, sinapsės užtikrina standų ryšį tarp dviejų sąveikaujančių ląstelių paviršių.
§ Plazmodezma.Šio tipo tarpląstelinis ryšys randamas augaluose. Plazmodesmata yra ploni vamzdiniai kanalai, jungiantys dvi gretimas ląsteles. Šių kanalų skersmuo paprastai yra 40-50 nm. Plazmodesmos praeina per ląstelės sienelę, kuri skiria ląsteles. Jaunose ląstelėse plazmodesmų skaičius gali būti labai didelis (iki 1000 vienoje ląstelėje). Senstant ląstelėms, jų skaičius mažėja dėl plyšimų, padidėjus ląstelės sienelės storiui. Funkcinis plazmodesmos vaidmuo yra užtikrinti tarpląstelinę maistinių medžiagų, jonų ir kitų junginių turinčių tirpalų cirkuliaciją. Plazmodesmos užkrečia ląsteles augalų virusais.
Specializuotos plazminės membranos struktūros
Daugelio gyvūnų ląstelių plazmalema formuoja įvairių struktūrų (mikrovilų, blakstienų, žvynelių) ataugas. Dažniausiai randama daugelio gyvūnų ląstelių paviršiuje mikrovileliai.Šios citoplazmos ataugos, apribotos plazmalemos, yra cilindro formos su apvalia viršūne. Mikrovileliai būdingi epitelio ląstelėms, bet randami ir kitų audinių ląstelėse. Mikrovielių skersmuo yra apie 100 nm. Jų skaičius ir ilgis skiriasi skirtingi tipai ląstelės. Mikrovillių reikšmė slypi reikšmingame ląstelės paviršiaus ploto padidėjime. Tai ypač svarbu ląstelėms, kurios dalyvauja absorbcijoje. Taigi žarnyno epitelyje 1 mm 2 paviršiaus yra iki 2x10 8 mikrovielių.
[Ankstesnė paskaita] [Turinys] [Kita paskaita]