Unikalūs savaip suteikia jų įvairovę ir gyvenimo bruožus.
Šiuolaikinė mikrobiologija tyrimams ir klasifikavimui naudoja šiuos dalykus skiriamieji bruožai bakterijos:
- Morfologinė ląstelių struktūra:
- sferiniai "kokai";
- lazdelės formos „bacilos“;
- spiralinės "spirochetos";
- vingiuota forma "vibrios".
- Kvėpavimas:
- kvėpuoti deguonimi "aerobinis";
- anoksinis „anaerobinis“.
- Maitinimo būdas:
- autotrofinis „nepriklausomas“ – galintis sintetinti organinės medžiagos iš neorganinių energijos pagalba cheminės reakcijos arba fotosintezė (pavyzdžiui, melsvadumbliai);
- heterotrofiniai - paverčia ir apdoroja paruoštas organines medžiagas, gautas dėl kitų organizmų gyvybinės veiklos, įskaitant patogeninius (patogeninius) ir simbiotinius (naudingus) mikroorganizmus, taip pat bakterijas, gyvenančias dėl fermentacijos, skilimo ir kt. .
- Buveinė:
- išorinė aplinka (dirvožemis, vanduo ir kt.);
- gyvų organizmų viduje;
- negyvos organinės medžiagos (lavonai, nuosėdos, atliekos).
- Pasiskirstymo ir išgyvenimo būdas (ar susiformuoja ląstelių sporos, apvalkalai, gleivės).
- Reakcija į Gramo testą (atskyrimas pagal sienų sudėtį ir struktūrą, pagrindinė klasifikacija konstrukcijos temos kontekste ląstelių sienelės):
- gramteigiamas – nudažytas Gramo dažymo procedūros metu (apvalkalas pralaidus anilino dažams (kristalinis violetinis, metilvioletas ir kt.));
- gramneigiamas – nedažyti Gram dažymo procedūros metu (ląstelės sienelė nepralaidi dažams).
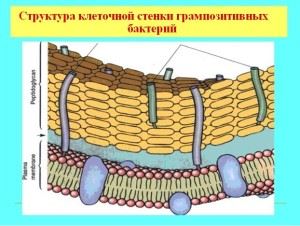
Gramteigiamų bakterijų ląstelės sienelės ypatybės
Tyrimo (1884 m. pasiūlyto G.K. Gramo) esmė – bakterijų kultūros mėginio apdorojimo anilino dažais (kristaliniu violetiniu, krištoliniu violetiniu) procedūra. Vėliau nuplovus alkoholiu, Gram (+) rūšių mikroorganizmai išlaiko mėlyną spalvą, o kiti pasikeičia. Ląstelės sienelės gebėjimą fiksuoti dažus lemia ląstelės sienelės sudėtis ir struktūra. Vienaląsčių organizmų klasifikavimas pagal ląstelių sienelės struktūros tipą, kurį galima nudažyti arba nedažyti Gramo metodu, yra vienas iš pagrindinių. Praktiškai diagnozuoti padeda gramo tyrimo rezultatai užkrečiamos ligos studijuoti biochemines savybes mikroorganizmai.
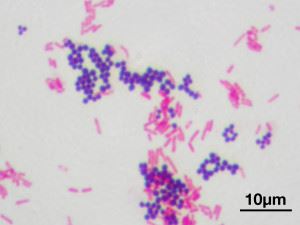
Gramo dėmė. Cocci (sferiniai) - gramteigiami ir bacilos (lazdelės) - gramneigiami
Dauguma gramteigiamų bakterijų turi masyvią, vieno sluoksnio membraną, pralaidžią anilino dažams. Sienos sudėtis apima sudėtingas organines molekules, mechaninio standumo pagrindą suteikia mureinas (heteropolimeras), glikopeptidai, mukopeptidai. Sienelės struktūra lemia mikroorganizmo formą ir yra kažkas panašaus į išorinį skeletą, porėtos struktūros, apie 40 glikopeptidų molekulių storio. Be peptidoglikano bazės, apvalkale yra techo rūgščių ir polisacharidų.
Nepaisant bendra reakcija ant dažiklio tarp gramteigiamų organizmų galima sutikti organizmų, priklausančių įvairiems porūšiams, skirtingos formos, dydis, buveinė ir šėrimo būdas.
Dauguma patogeninių bakterijų žmonėms yra gra-teigiamos, tarp jų, pavyzdžiui, tokių infekcinių ligų sukėlėjai:
- stabligė (Clostridium tetani),
- botulizmas (Clostridium botulinum),
- juodligė (),
- streptokokai,
- stafilokokai (Staphylococcus aureus), kurie yra labai atsparūs temperatūrai ir veikimui vaistai.
Atsparus rūgštims, dėl ypatingos daugiasluoksnės apvalkalo struktūros (į ją įeina vaškai, polisacharidus ir baltymai), kaip taisyklė, Gramo metodu jie nedažomi. Jiems naudojamas specialus Ziel-Nelsen metodas. Bet kai kurie duoda teigiamas rezultatas pagal Gramo metodą esant labai didelei dažų koncentracijai arba pakilusi temperatūra procedūros metu.
Rūgštims atsparūs vienaląsčiai organizmai išgyvena rūgštinėse ir šarminė aplinka, nors jų vystymuisi ir augimui normali aplinka yra neutrali. paprastai klasifikuojami kaip gramteigiami. Šios žmonėms ir gyvūnams pavojingos grupės atstovai yra ligų, tokių kaip tuberkuliozė ir raupsai, sukėlėjai. Be to, rūgštims atsparios yra kai kurios dirvožemio bakterijos, galinčios fiksuoti atmosferos azotą ir būti augalų simbiontais.
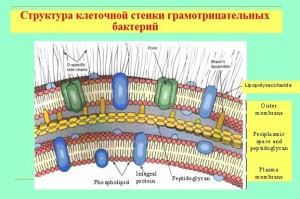
Gramneigiamų bakterijų apvalkalo ypatybės
Nors jų mukopeptidinės sienelės storis daug plonesnis, sienelė yra nepralaidi krištolo violetiniams dažams. Viskas apie iš esmės skirtingą ląstelės struktūrą ir cheminę sudėtį. Korpusas yra daugiasluoksnės struktūros ir susideda iš:
- išorinis apvalkalas (membanai), jo pagrindinė sudėtis yra liposacharidai ir baltymai;
- periplazminė erdvė;
- vidinis sluoksnis (mureino peptidoglikanas), jo sudėtis yra panaši į gramteigiamų bakterijų sienelių sudėtį, tačiau, kaip taisyklė, yra daugiau nei du kartus plonesnė.
Ačiū daugiau sudėtinga struktūra kai kuriems šios grupės atstovams padidėjęs atsparumas antikūnams ir antibiotikams.
Komplikavus ląstelės sienelę, atsirado naujų funkcijų ir galimybių. Kai kurios bakterijų rūšys, kurios gyvena toliau kietus paviršius, įgijo galimybę keisti ląstelės formą ("skeleto" sienelės tapo plastikinės). Tai svarbu slystant ir judant per kietos buveinės nelygumus.
Periplazminė erdvė įgijo izoliuotos kai kurių hidrolizuojančių fermentų, reikalingų ląstelės gyvavimui, saugojimo vietos funkciją, tačiau tuo pačiu galinti suskaidyti pačios bakterijos polimerines molekules. Dėl polimero molekulių, patenkančių iš išorės, hidrolizės gyva ląstelė praplečia mitybai tinkamų medžiagų asortimentą, tuo pačiu fermentams nepralaidi vidinė membrana neleidžia „savaiminiam virškinimui“ bakterijų ląstelei.
Išorinės membranos struktūroje yra baltymų, kurie gali sudaryti hidrofilines poras, per kurias į ląstelę gali patekti kai kurios bakterijų mitybai reikalingos makromolekulės (cukrus ir aminorūgštys).
Šiai bakterijų klasei priklauso daug bakterijų, kurios gali keistis cheminė sudėtis buveinė (acto ir alkoholinės fermentacijos bakterijos); simbiotinė – Escherichia coli (E coli), daug patogeninių (ligą sukeliančių) organizmų, tokių kaip meningokokas (Neisseria meningitidis); legionelės (Legionella pneumophila), ir garsiosios Helicobacter pylori iššaukiantis pepsinė opa ir didelis atsparumas rūgštims.

Bakterijų struktūrinių savybių naudojimas žmonėms
Pirmieji mikroorganizmai, kuriuos žmogus panaudojo savo reikmėms, buvo pieno ir alkoholinės fermentacijos bakterijos. Tai jie mums gamino ir iki šiol verda sūrį, duoną ir vyną. Be to, žmonės pradėjo naudoti savo darbo produktą dar gerokai anksčiau nei atrado bakterijas ir pradėjo tyrinėti jų medžiagų apykaitos produktų struktūrą ir sudėtį.
Šiuo metu biologiniai nuotekų ir išmatų valymo metodai tapo prieinami ne tik savivaldybės įmonės bet ir privačių namų savininkams. Neveikiančios kultūros yra dalis šiuolaikiniai vaistai vandens telkiniams ir vietinei kanalizacijai. Ūkiai dažnai naudojamas atsparus rūgštims komposto ir gyvulinių atliekų perdirbimui į trąšas ir daugeliui kitų tikslų.
Taip pat daug genetiškai modifikuotų bakterijų dalyvauja vaistų gamyboje, naujų tipų polimerų ir kitų unikalių savybių turinčių medžiagų sintezėje. cheminės savybės ir sudėtinga struktūra.
Cheminiai ląstelės sienelės komponentai paprastai skirstomi į pagrindinius, inkrustuojančius ir nusėdančius ant jos paviršiaus (adkrustacija). Į pagrindinį cheminiai komponentai apima celiuliozę, hemiceliuliozę, pektiną, baltymus. Ląstelės sienelę inkrustuojantys komponentai yra ligninas ir suberinas. Cutinas ir vaškas gali nusodinti ant ląstelės sienelės paviršiaus. Be to, ląstelių sienelėse yra silikatų ir kalcio karbonatų.
Dažniausi atsinaujinančių augalinių medžiagų komponentai yra celiuliozė, ligninas ir hemiceliuliozės (poliozės), kurių atitinkamai yra 40; 30 ir 26 proc Bendras svoris. Celiuliozė yra ląstelių sieneles stiprinanti medžiaga, formuojanti tvarkingas siūlines struktūras (fibriles). Ligninas ir hemiceliuliozės yra amorfiniai biopolimerai, turintys tinklinę arba šakotą struktūrą ir atlieka rišiklių (užpildo) vaidmenį.
Šių medžiagų struktūra, jų tarpmolekulinė sąveika lemia augalų substratų kaip visumos savybes ir jų elgseną perdirbimo metu.
Celiuliozė
Celiuliozė yra labiausiai paplitusi medžiaga Žemėje, tai yra augalų ląstelės sienelės skeletas. Geriausiai ištirta medvilnės pluošto ir kai kurių bakterijų, dumblių ir moliuskų sintezuojamų pluoštų celiuliozės struktūra, nes juose celiuliozė randama gryniausia forma. Medienoje jis būna tik kompozito pavidalu su kitais biopolimerais, iš kurių sunku išgauti be dalinio sunaikinimo.
Chemiškai celiuliozė yra polisacharidas, kurio makromolekulės yra sudarytos iš ß-D-gliukozės liekanų ir sudaro linijinę anhidrogliukopiranozės grandinę, sujungtą 1,4-gliukozidiniais ryšiais (9 pav.).
9 pav. Cheminė struktūra celiuliozė
Tai stereoreguliarus polimeras, kurio grandinėje celobiozės likučiai atlieka stereokartojimo grandį. Bendra celiuliozės formulė yra (C 6 H 10 O 5) n arba [C 6 H 7 O 2 (OH) 3] n. Celiuliozės monomero vienete yra trys alkoholio hidroksilo grupės, iš kurių viena yra pirminė - CH 2 OH, o dvi C 2 ir C 3 yra antrinės - CHOH. Galinės jungtys skiriasi nuo likusių grandinės grandžių. Viena galinė jungtis (C 4) turi papildomą laisvą antrinį alkoholio hidroksilą. Galinėje jungtyje (C 1) yra laisvo glikozidinio (pusacetalio) hidroksilo ir gali būti dvi tautomerinės formos – ciklinė (pusacetalinė) ir atvira (aldehidas). Galinė aldehido grupė suteikia celiuliozei redukcinį (atkuriamąjį) gebėjimą. Celiuliozės gebėjimas atlikti redukcijos reakciją gali būti naudojamas apibūdinti jos grandinių ilgį, kuris yra būdingas ši rūšis augalai.
Gliukozės liekanoje įprasta skirti hidrofilines ir hidrofobines plokštumas, priklausomai nuo to, kiek OH ir CH grupių ją atitinka. Tai svarbu tiriant gliukozės likučio sąveiką su aromatinių aminorūgščių liekanomis celiuliozės molekulės fermentinio skilimo metu aktyviame fermento centre.
Dėl to, kad pirmojo ir ketvirtojo gliukozės atomų hidroksilo grupės skiriasi žiedo plokštumos atžvilgiu, gretimos gliukozės liekanos polimero grandinėje yra pasuktos 180 0 viena kitos atžvilgiu. Polimerų grandines stabilizuoja vandeniliniai ryšiai, susidarantys tarp gliukozės likučių sąveikaujant 6,2,3 anglies atomų turinčioms hidroksilo grupėms ir žiedo deguoniui. Linijinės polimerų grandinės tarpusavyje sujungiamos naudojant tarpmolekulinius vandenilio ryšius, atsirandančius tarp vienos grandinės deguonies atomų ir kitos grandinės gliukozės C 6 hidroksilo grupių (10 pav.). Dėl tokios sąveikos susidaro stiprūs pluoštai. Celiuliozės molekulės lankstumą lemia galimybė celiuliozės jungtyje esantiems anhidrogliukozės likučiams suktis glikozidinės jungties atžvilgiu. Linijinės celiuliozės konformacijos stabilumą visų pirma lemia minimalus C1-H ir C4-H atstūmimas.
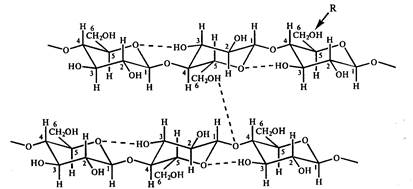
Ryžiai. 10. Scheminis vandenilinių ryšių tarp gliukozės likučių celiuliozėje sistemos pavaizdavimas. R yra galimo cheminio celiuliozės modifikavimo centras. Taigi metilceliuliozėje, acetilceliuliozėje ir karboksimetilceliuliozėje atitinkamai R \u003d CH 3, COCH 3, CH 2 COOHa
Gliukozės molekulė pateikiama kėdės konformacijoje, kurioje C2, C3, C5 ir O5 atomai yra vienoje plokštumoje, o C1 ir C4 atomai yra priešingose jos pusėse. Gliukozės liekana gali turėti ir energetiškai mažiau palankias konformacijas, kurios pagal stabilumo laipsnį išsidėsto tokia tvarka: įstrižinė konformacija, valties (arba vonios) konformacija ir nepalankiausia puskėdės konformacija, kurioje 5 atomai. iš 6 narių piranozės žiedo iš karto patenka į vieną plokštumą. Anhidrogliukozės vieneto gebėjimas priimti celiuliozės molekulę, be stabiliausių, mažiau stabilių konformacijų (pavyzdžiui, valties ar pusės kėdės konformacijos), turi svarbą pažeidžiant tvarkingą supramolekulinę celiuliozės struktūrą, taip pat jos molekulėje vykstant glikozidinių jungčių fermentinei hidrolizei.
Elementarus celiuliozės pluoštas (micelės) susideda iš 100-200 polimerų grandinių, sujungtų vandenilio tilteliais. Šios ląstelės sienelės skaidulos yra lygiagrečios viena kitai ir sugrupuotos taip, kad susidaro ovalo formos ryšuliai. – mikrofibrilės . Apie 10 micelių sudaro 0,025 nm skersmens mikrofibrilę (11 pav.).
Kiekvienas ryšulėlis turi centrinę kristalinę dalį – šerdį, kurioje grandinės suspaustos taip stipriai, kad net vandens molekulės į jas labai sunkiai prasiskverbia. Aplink branduolį yra mažiau tvarkingos struktūros parakristalinė žievė. Šios mažiau tvarkingos sritys, dar vadinamos amorfinėmis sritimis, paprastai sudaro apie 15% celiuliozės mikrostruktūros. Mikrofibrilės gali susijungti atskiros grupės arba sluoksniai – makrofibrilės – 0,4-0,5 mikrono storio, matomos šviesos mikroskopu. Mikrofibrilių skersmuo ir kristališkumo laipsnis gana stipriai skiriasi priklausomai nuo augalo rūšies ir ląstelės fiziologinės būklės.
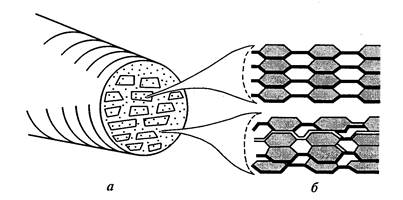
11 pav. Mikrofibrilės struktūros modelis: a) mikrofibrilės skerspjūvis su micelėmis ir tarpląsteliniais tarpais; b) celiuliozės išdėstymas
Medienos plaušienos, kurios užsakymo indeksas yra 50-70%, kristalito skersiniai matmenys neviršija 4x6 nm. Pagal elektroninę mikroskopiją vidutiniai skersmens matmenys gali būti 1,5-2x2,5-3,5 nm. Nuo 40 iki 80% tokio dydžio molekulių yra kristalito paviršiuje, kur galimas didesnis jų drėkinimas, vandenilio jungčių struktūros ilgalaikės tvarkos pažeidimas, taip pat stipresnė cheminių poveikių įtaka.
Natūrali celiuliozė yra polidispersinė, todėl gliukozės likučių skaičius celiuliozės molekulėje ir, atitinkamai, pačių molekulių ilgis gali labai skirtis ląstelių sienelėsįvairūs augalai. Taigi, medvilnės plaukų celiuliozės pirminės ląstelės sienelės polimerizacijos laipsnis yra 2-6 tūkst., antrinės - 13-14 tūkst., Acetobacter xylinum celiuliozės - 2-6 tūkst., medienos - 8-10 tūkst. Didžiausia eksperimentiškai nustatyta linų celiuliozės molekulinė masė yra 6 000 000. Dėl to, kad polimerizacijos laipsnis yra vidutinis rodiklis, tiksliausia charakteristika yra celiuliozės molekulių pasiskirstymo išilgai kreivė (molekulinės masės pasiskirstymas). Tačiau šią charakteristiką sunku gauti, todėl naudojamos vidutinės charakteristikos: vidutinis skaičius (DP n) ir vidutinis masės (DP w) polimerizacijos laipsniai. Vidutinis polimerizacijos laipsnis yra lygus bendram anhidrogliukozės likučių skaičiui celiuliozės mėginyje, padalytam iš iš viso polimerų molekulės. Norėdami jį nustatyti, polisacharido mėginyje titruojami redukuojantys galai. Vidutinis masės celiuliozės polimerizacijos laipsnis nustatomas ištirpinant celiuliozės mėginį šarminiame vario, cinko, kobalto arba kadmio komplekse su etilendiaminu arba LiCl komplekse su N,N'-dimetilacetamidu. DP santykis w/DP n lygus vienybei tik monodispersiniams polimerams.
Eglės, pušies, beržo ir drebulės medienos masės vidutinis polimerizacijos laipsnis yra 12 000, o tokios molekulės ilgis išsiplėtusioje būsenoje siekia beveik 6 mikronus, o tai yra 6000 kartų didesnis už jos skersmenį. Tokia ilga ir reguliariai pastatyta molekulė yra tam tikru būdu sukrauta. Krovimo metodas nustato supramolekulinę biopolimero struktūrą, kuri yra pagrįsta vidinėmis ir tarpmolekulinėmis vandenilinėmis jungtimis, kurias sudaro celobiozės vienetai. Celiuliozės makromolekulėje esančių vandenilinių jungčių krovimo metodo ir tipų tyrimas atliekamas naudojant konformacinę analizę, difrakciją (mažo kampo rentgeno sklaidą, elektronų sklaidą), IR spektroskopiją ir BMR. Nepaisant didelės pažangos šioje srityje, klausimas, kaip sukrauti celiuliozės molekules, nebuvo galutinai išspręstas.
Celiuliozė netirpi vandenyje ir organiniuose tirpikliuose, chemiškai inertiška, mechaniškai labai stipri. Vanduo tarp glaudžiai išdėstytų, bet ne tiesiogiai sąveikaujančių kristalitų paviršių yra struktūrizuotas. Hidratacijos vandens pašalinimas, pavyzdžiui, dėl džiovinimo, skatina iš pradžių nekontaktuojančių celiuliozės molekulių konvergenciją ir įvairių tipų vandenilinių jungčių susidarymą tarp jų, kurios nesunaikinamos rehidratacijos metu. Tyrėjai priskiria kristališkumo laipsnio padidėjimą drėkinimo metu su vandens prasiskverbimu iš mažiau tvarkingų regionų į labiau tvarkingus regionus. Dėl to pastebimas kristalitų absoliučių dydžių mažėjimas ir bendrojo sutvarkymo laipsnio padidėjimas. Manoma, kad vanduo šiuo atveju veikia pagal „molekulinio tepimo“ principą, todėl netvarkingiems molekulių fragmentams lengviau užbaigti sutvarkytą struktūrą bendradarbiaujant su kristaline „sėkla“. Polinių celiuliozės grupių pakeitimas, pavyzdžiui, karboksimetilu, pažeidžia molekulinį dėsningumą, skatina daugelio vandenilio jungčių susidarymą su vandeniu ir daro celiuliozę tirpią.
Celiuliozės atsparumą skilimui tiek natūraliomis, tiek laboratorinėmis sąlygomis lemia ne tiek b-1,4-gliukozidinės jungties savybės, kiek kristalinė celiuliozės struktūra ir ypatingas jos molekulių „pakavimas“ biologines struktūras.
Celiuliozės fermentai negali prasiskverbti į vidines celiuliozės fibrilių struktūras. Jie veikia savo paviršiuje. Glikozidinių jungčių, skirtų fermentams veikti, skaičius labai priklauso nuo celiuliozės išbrinkimo laipsnio. Padidinti patinimo laipsnį galima naudojant išankstinį apdorojimą - garinant, malant, apdorojant ultragarsu ir kt. Viso pluošto pabrinkimą didina jame esančios mineralinės rūgštys ir šarmai didelės koncentracijos, nes jie gali nutraukti vandenilio ryšius ir prasiskverbti į kristalines sritis. Amorfines celiuliozės dalis gana lengvai paveikia rūgštys ir šarmai.
Koncentruotiems šarmų tirpalams veikiant celiuliozę, pasikeičia jos fizikinės ir cheminės bei struktūrinės savybės: pastebimas intensyvus patinimas, kinta kristališkumo laipsnis. Didelės molekulinės masės celiuliozės frakcija, netirpi 17,5 % NaOH, vadinama a-celiulioze. Medvilnę daugiausia sudaro a-celiuliozė, o kituose augaluose ir medienoje daugiausia yra b-celiuliozės, tirpios koncentruotame šarminiame tirpale.
Mikrofibrilės panardinamos į amorfinę apvalkalo matricą, kuri yra sudėtingas polimerų mišinys, kuriame vyrauja įvairios molekulinės masės polisacharidai. Daroma prielaida, kad visų mikrofibrilių paviršiuje yra ksilogliukano molekulių sluoksnis, sorbuotas vandeniliniais ryšiais. Ligninas ir hemiceliuliozės užpildo erdvę tarp elementariųjų celiuliozės fibrilių. Taigi lignifikuota medžiaga yra polimerinė struktūra, sudaryta iš celiuliozės armatūros, panardintos į lignoceliuliozės matricą.
Celiuliozės ir hemiceliuliozės pasižymi skirtinga formavimosi kinetika, kuri, matyt, rodo skirtingos vietos jų sintezė. Atrodo, kad celiuliozės sintezė vyksta ne ląstelės paviršiuje citoplazminė membrana dalyvaujant GDF arba UDF. Glikozilo grupių donorai yra cukraus nukleozidų difosfatai, kurie susidaro iš gliukozės ir sacharozės. Skirtingi keliai. Pavyzdžiui, iš nukleozidų trifosfatų ir cukraus-1-fosfatų, dalyvaujant pirofosforilazei. Glikozilo likučiai yra prijungti prie cukraus likučių neredukuojančiuose augimo grandinės galuose. Tarp pirmosios „pridėtos“ liekanos anglies ir galinės liekanos ketvirtojo anglies atomo hidroksilo grupės susidaro glikozidinė jungtis, dalyvaujant sintetazei.
N (nukleozidifosfatas cukrus) + (cukrus) n sintetazė n (nukleozidifosfatas) + (cukrus) n +1
Ląstelės sienelėje aptikti celiuliozės sintezės fermentai ir 8 nm skersmens granulės, kurios, matyt, yra daugiafermentiniai kompleksai, esantys išorinis paviršius plazmalemma ir sienelės matricoje. Celiuliozės sintetazė (E) plazmalemoje yra susijusi su aktyvuojančiu baltymu P, kuris gali surišti GTP ir pereiti į aktyvią formą, susidarant P-E-GTP kompleksui.
Celiuliozės sintezės procese susidaro ne tik pavienės celiuliozės grandinės, bet ir celiuliozės mikrofibrilės. Tokia sintezė tikriausiai vykdoma dalyvaujant daugybei celiuliozės sintetazių molekulių, sutelktų atskirose plazmalemos vietose, o celiuliozės sintetazės susideda iš daugybės subvienetų, kurių kiekvienas yra atsakingas už vienos gliukano grandinės sintezę. Įjungta viduje membranų, rastos šešių baltymų rutuliukų „rozetės“. Rozetės išskiriamos į plazmalemą per endomembraninę pūslelių sistemą. Remiantis rozečių, galinių kompleksų ir susidarančių celiuliozės mikrofibrilių struktūrinio ryšio ypatumais, buvo daroma prielaida apie mikrofibrilių susidarymo mechanizmą komplementarinėje transmembraninėje struktūroje: rozetės – galinis kompleksas – augantis mikrofibrilės galas. Prie naujai susintetintos mikrofibrilės galo prijungtas terminalinis kompleksas, o rozetės yra celiuliozės pirmtakų surišimo ir gliukozės likučių „nešiklių“ išskyrimo vieta.
Pavienės rozetės kartu su atitinkamais galiniais kompleksais yra atsakingos už atsitiktinai orientuotų pirminės ląstelės sienelės mikrofibrilių sintezę (12 pav.), o orientuotas iki 175 rozečių rinkinys atskirose plazmalemos dalyse lemia lygiagrečių formavimąsi. antrinės ląstelės sienelės mikrofibrilės (13 pav.).
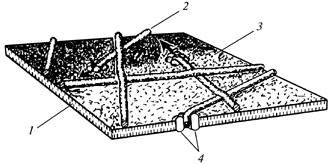
12 pav. Celiuliozės mikrofibrilės formavimosi metu
pirminė ląstelės sienelė: 1
- lipidų dvigubas sluoksnis; 2
- mikrofibrilė; 3
- išorinis plazmalemos paviršius; 4
- lizdas
Kiekviena rozetė sudaro 5 nm skersmens mikrofibrilę. Atstumas tarp rozečių eilių plazmos membranoje atitinka atstumą tarp mikrofibrilių ląstelės sienelėje. Plonose mikrofibrilėse yra apie 50 poligliukano grandinių. Šios mikrofibrilės sudaro antrinės ląstelės sienelės fibriles, kurios gali būti iki 28 nm skersmens. Celiuliozė, skirtingai nei krakmolas, yra sunkiai metabolizuojamas junginys. Jo virškinamumas priklauso nuo kilmės, kiekio racione, paruošiamojo apdorojimo pobūdžio ir vidutiniškai svyruoja nuo 6 iki 23 %. Taip, celiuliozės virškinamumas kviečių sėlenos apie 15 proc.
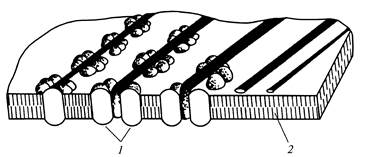
Ryžiai. 13.Celiuliozės mikrofibrilės formavimosi metu
antrinė ląstelės sienelė: 1
- lizdas; 2
- lipidų dvigubas sluoksnis
IN maistinė lasteliena celiuliozė sudaro apie trečdalį. Jo efektyvus skilimas vyksta dėl įvairių storosios žarnos mikroorganizmų ir jų celiuliolizinių fermentų simbiotinės sąveikos. IN Virškinimo traktas celiuliozė skatina žarnyno veiklą, didina jo peristaltiką, normalizuoja veiklą žarnyno mikroflora, sugeria sterolius, neleidžia jiems pasisavinti ir skatina cholesterolio išsiskyrimą. Mikrokristalinė celiuliozė (MCC), nepluoštinė natūralios celiuliozės miltelių modifikacija, plačiai naudojama kaip maisto papildas. Tai ne tik sumažina maisto kaloringumą, bet ir yra jo tirštiklis bei dispergentas, gerinantis maisto kokybę.
Gramneigiamos eubakterijos turi daug sudėtingesnę ląstelių sienelių struktūrą nei gramteigiamos. Jame yra daug daugiau skirtingų makromolekulių cheminis tipas. Peptidoglikanas sudaro tik vidinį ląstelės sienelės sluoksnį, laisvai besiribojantį su CPM. Dėl skirtingi tipai Gramneigiamos eubakterijos, šio heteropolimero kiekis labai skiriasi. Daugumoje rūšių jis sudaro vieno arba dviejų sluoksnių struktūrą, kuriai būdingi labai reti kryžminiai ryšiai tarp heteropolimerų grandinių.
Kai kurios sklandančios bakterijos (miksobakterijos, fleksibakterijos) gali periodiškai keisti ląstelių formą, judėdamos išilgai kieto substrato, pavyzdžiui, lenkdamos, o tai rodo jų ląstelės sienelės, o pirmiausia jos peptidoglikano sluoksnio, elastingumą. Tačiau elektronų mikroskopinis tyrimas atskleidė, kad jie turi gramneigiamoms eubakterijoms būdingą ląstelės sienelę. Labiausiai tikėtinas šių bakterijų ląstelių sienelės lankstumo paaiškinimas yra itin mažas peptidoglikano komponento kryžminis ryšys.
Gramneigiami prokariotai turi išorinė membrana, kuri apima lipidus (22%), baltymus, polisacharidus, lipoproteinus.
Lipopolisacharidai (LPS) yra sudėtingos struktūros heteropolimerai, pasižymintys įvairiu biologiniu aktyvumu. Lipoidų kompleksas sukelia toksiškumą (uždegiminės reakcijos, karščiavimas, endotoksinis šokas), polisacharido komponentas atsakingas už O-antigeno specifiškumą. LPS skatina Jg M antikūnų sintezę ir yra naudojamas imunologijoje kaip adjuvantas ir polikloninis B ląstelių aktyvatorius.
Bakterijų ląstelės sienelė daugiausia atlieka formavimo ir apsaugines funkcijas, suteikia standumo, formuoja kapsulę ir lemia ląstelių gebėjimą adsorbuoti fagus.
№15 Filtruojamos, involiucinės bakterijų formos, protoplastai, sferoplastai, L formos.
protoplastai, sferoplastai, L formos yra bakterijų formos be KS
L formos gali atsirasti natūraliomis sąlygomis žmogaus organizme dėl ilgalaikio gydymo tam tikrais antibiotikais, dažniausiai penicilinu.
Yra nestabilios ir stabilios L formos bakterijos. Pirmieji gali grįžti į pradinę formą, kai pašalinama priežastis, sukėlusi jų susidarymą. Jie atkuria gebėjimą sintetinti peptidoglikaną CS. Pastarieji, kaip taisyklė, negali grįžti. Įvairių bakterijų L formos vaidina esminį vaidmenį daugelio infekcinių ligų patogenezėje.
Pagrindinės bakterijų L formų savybės:
nuolatinis transformavimas iš grampol-s į gramatrits-e.
antigeninių savybių pasikeitimas
virulentiškumo sumažėjimas
gebėjimas ilgai ištverti
gebėjimas grįžti į pradinę formą, jei COP sinetza ne visiškai prarasta.
№16 Bakterijų tinktūrinės savybės. Sudėtingi dažymo metodai. Bakterijų dažymo gramais principas.
Gramo beicavimo technika
Ant tepinėlio uždedamas filtravimo popierius ir 1-2 minutes pilamas gencijonų violetinės karbolinis tirpalas.
Nuimkite popierių, nusausinkite dažus ir, neplovę tepinėlio vandeniu, 1 minutę užpilkite Lugolio tirpalu.
Lugolio tirpalas nusausinamas ir preparatas 30 sek. balinamas 96 0 spirite.
Nuplauti vandeniu.
Dažyti 1-2 min vandeninis tirpalas rausvai raudona.
Apsupimo principas Pagal gramą
Gram teigiamas bakterijos išlaiko gencijonų violetinę spalvą kartu su jodu - violetinė spalva bakterijos;
Gram neigiamas Po alkoholio poveikio bakterijos netenka dažų, pakinta spalva ir, apdorojus fuksinu, nusidažo raudona spalva.
Sudėtingi metodai dėmės naudojamos ląstelių struktūrai ir mikroorganizmų diferenciacijai tirti. Dažyti tepinėliai mikroskopuojami imersinėje sistemoje. Preparatui nuosekliai naudokite tam tikrus dažus, kurie skiriasi chemine sudėtimi ir spalva, kandiklius, alkoholius, rūgštis ir kt.
tintoriaus savybės bakterijos (lot. tinctura, iš tingo - I spalva) gebėjimas spalvinti: jautrumas spalvai, atsparumas rūgštims-alkoholiui-šarmui, spalvos vienodumas, metachromatiškumas, santykis su spalva Gramo metodu.
№17 Rūgščiai atsparių bakterijų ląstelės sienelės cheminės sudėties ypatybės. Dažymo principas pagal Ziehl-Neelsen metodą.
rūgštims atsparios bakterijos. Kai kurių bakterijų ląstelių sienelėse yra daug lipidų ir vaškų, todėl jos yra atsparios vėlesniam spalvos pakitimui po dažymo rūgštimis, šarmais ar etanoliu (pavyzdžiui, Mycobacterium arba Nocardia rūšys). Tokios bakterijos vadinamos atsparios rūgščiai ir sunkiai nudažomos gramais (nors rūgštims atsparios bakterijos laikomos gramteigiamos). Jų dažymui naudojamas Ziehl-Neelsen metodas.
Mikobakterijoms ir nokardijai būdinga sudėtinga ląstelės sienelės struktūra. Jos, kaip ir gramteigiamos bakterijos, yra pagrįstos mureino karkasu, tačiau pastarasis siejamas su polisacharidais ir lipidais. Lipidus atstovauja mikolio rūgštys, kurios suteikia ląstelės paviršiaus hidrofobiškumą. Hidrofobiškumas, viena vertus, daro ląstelę atsparią įvairių cheminių medžiagų veikimui (tokios bakterijos vadinamos atspariomis rūgštims), kita vertus, slopina ląstelės mainus su aplinka ir lėtina jos augimą. Todėl paviršinio aktyvumo medžiagos pridedamos prie maistinių medžiagų, skirtų Mycobacterium tuberculosis auginti. Mikobakterijų atsparumas rūgštims yra svarbus diagnostinis požymis, jo nustatymui naudojamas Ziehl-Neelsen dėmė.
Rūgščiai atsparių bakterijų dažymo technika
pagal Ziehl-Neelsen metodą
Ant fiksuoto tepinėlio uždedamas filtravimo popierius, pilamas Ziehl karbolinis fuksinas ir švelniai kaitinamas ant degiklio, kol atsiras garų. Operacija kartojama 2-3 kartus.
Vaistui atvėsus, nuimkite filtravimo popierių, nusausinkite dažus ir nuplaukite vaistą vandeniu.
Vaistas 2-3 kartus panardinamas į stiklinę su 5% sieros rūgštimi 1-2 sekundėms.
Preparatas kruopščiai nuplaunamas vandeniu ir 3-5 minutes dažomas šarminiu metileno mėlynu.
Nuplauti vandeniu ir išdžiovinti.
Rūgštims atsparios bakterijos neblunka su sieros rūgštimi ir išlaiko raudoną spalvą, rūgštims neatsparios bakterijos praranda dažus ir metileno mėlynu nusidažo mėlyna spalva.
№18 Bakterijų inkliuzai, jų sudėtis ir biologinis vaidmuo. Citoplazmoje yra įvairių intarpų glikogeno granulių, polisacharidų, beta-hidroksisviesto rūgšties ir polifosfatų (volutino) pavidalu. Jie yra rezervinės medžiagos bakterijų mitybos ir energijos poreikiams tenkinti. Volyutin turi afinitetą pagrindiniams dažams ir yra lengvai aptinkamas naudojant specialius metodus dažymas (pavyzdžiui, pagal Neisser) metachromatinių granulių pavidalu. Būdingas volutino granulių išsidėstymas difterijos baciloje atsiskleidžia intensyviai dažytų ląstelės polių pavidalu. (pagal Neisser jis yra tamsiai mėlynos spalvos)
№19 Bakterijų sporos, jų sandara Kai kurios bakterijos aktyvaus augimo periodo pabaigoje sugeba formuoti sporas. Prieš tai aplinkos išeikvojimas maistinėmis medžiagomis, pakinta jos pH, kaupiasi toksiški medžiagų apykaitos produktai. Paprastai viena bakterijos ląstelė sudaro vieną sporą - sporų lokalizacija yra skirtinga (centrinė, galinė, subterminalinė)
Jei sporų dydis neviršija lazdelės formos bakterijos skersinio dydžio, tai pastaroji vadinama bacila (sukėlėjas juodligė). Kai sporos skersmuo didesnis, bakterijos būna verpstės formos ir vadinamos klostridijomis (anaerobinės infekcijos sukėlėjai). Stabligės klostridijos turi apvalias sporas ir primena blauzdeles. Clostridia botulinum pasižymi didelėmis ovaliomis sporomis, kurios suteikia joms teniso raketės išvaizdą.
Pagal cheminę sudėtį sporos ir vegetatyvinės ląstelės skiriasi tik kiekybiniu cheminių junginių kiekiu. Sporose yra mažiau vandens ir daugiau lipidų.
Sporų susidarymas yra susijęs su tam tikros vegetatyvinės ląstelės citoplazmos srities sutankinimu ir išskyrimu, o po to bakterijos viduje susidaro apvalus arba ovalus kūnas, padengtas tankia daugiasluoksne membrana, kuri yra prisotinta didelis kiekis lipidų, kalcio ir dipikolino rūgšties (12 pav.).
Patekusios į žmonių ir gyvūnų organizmą, sporos sudygsta į vegetatyvines ląsteles. Sporų dygimo procesas susideda iš trijų etapų: aktyvacijos, Pradinis etapas ir augimo stadijos. Ramybės būseną trikdančios aktyvuojančios medžiagos yra padidėjusi temperatūra, rūgštinė aplinkos reakcija, mechaniniai pažeidimai ir kt.. Sporos pradeda sugerti vandenį, išsiskiria iš kalcio dipikolato ir hidrolizinių fermentų pagalba sunaikina daugelį savo struktūrinių komponentų. Sunaikinus išorinius sluoksnius, prasideda vegetatyvinės ląstelės formavimosi laikotarpis su biosintezės aktyvavimu, baigiasi ląstelių dalijimusi.
Sporų dažymo metodas pagal Ožeško
Sporos tvirtai sulaiko karbolinį fuksiną ir nusidažo raudonai, bakterijų citoplazma nuspalvinama 5% sieros rūgštimi ir, nudažius metileno mėlynu, tampa mėlyna.
№20 Bakterijų kapsulė ir mikrokapsulė Kapsulė – tai gleivinis bakterijų ląstelės sienelės sluoksnis, susidedantis iš polisacharidų (pneumokokų) arba polipeptidų (juodligės bacilų). Mikrokapsulę (mažiau nei 0,2 µm storio) gali sudaryti dauguma bakterijų, tiksliai apibrėžtą makrokapsulę (daugiau nei 0,2 µm storio) sudaro pneumokokas, Klebsiella, juodligė ir kai kurios kitos. Patogeninėse bakterijose kapsulė susidaro makroorganizme, dirbtinėse maistinėse terpėse ji dažniausiai prarandama (išskyrus Klebsiella).
Žmonėms ir gyvūnams kapsulė apsaugo patogeninių bakterijų nuo bakteriofago, fagocitozės ir humoralinio imuniteto faktorių, lemia mikroorganizmų antigeninį specifiškumą.
Kapsulės, turinčios gelio konsistenciją, blogai išlaiko dažus, joms nustatyti dažniausiai naudojami neigiamo kontrasto metodai.
Burri-Gins kapsulės aptikimo metodas
Bakterijos nusidažė raudonai, tamsiame vaisto fone kontrastuoja nedažytos kapsulės.
№21 Bakterinės žiogelės Vėželės veikia kaip judėjimo organas, leidžiantis bakterijoms judėti 20-60 mikronų per sekundę greičiu. Bakterijos gali turėti vieną (monotrichinę) arba kelias žiuželes, išsidėsčiusias visame kūno paviršiuje (peritrichous), arba susikaupusios į ryšulius (lopotrichas).
Peritrichinė žvynelių vieta būdinga enterobakterijoms, anaerobinių infekcijų sukėlėjams, stabligei, botulizmui; Vibrio cholerae yra monotrichas, o Pseudomonas yra lopotrichas. Kai kurios spirilių rūšys turi amfitricinį žvynelių išsidėstymą. Žvynelių storis vidutiniškai siekia 10-30 nm, o ilgis siekia 10-20 mikronų.
Žiedyno pagrindas yra ilgas spiralinis siūlas (fibrilė), kuris ląstelės sienelės paviršiuje pereina į sustorėjusią lenktą struktūrą – kabliuką ir yra pritvirtintas prie bazinės granulės, įterptos į ląstelės sienelę ir CPM (10 pav.) .
Bazinės granulės yra apie 40 nm skersmens ir susideda iš kelių žiedų (viena pora gramteigiamų bakterijų, keturios gramneigiamų prokariotų). Pašalinus peptidoglikano sluoksnį iš ląstelės sienelės, prarandamas bakterijų gebėjimas judėti, nors žvyneliai lieka nepažeisti.
Žvyneliai beveik visiškai sudaryti iš baltymo flagellino, kuriame yra šiek tiek angliavandenių ir RNR.
Mikroskopu žvyneliai gali būti matomi tik po specialių ėsdinimo ir impregnavimo sidabro ir gyvsidabrio druskomis metodų, vėliau dažymo metileno mėlynu (Lefflerio metodas). Reikėtų nepamiršti, kad žvyneliai yra labai jautrūs įvairiems mechaniniams poveikiams. Žvynelių buvimas gali būti netiesiogiai vertinamas pagal judėjimo kryptingą pobūdį „kabančio“ ir „susmulkinto“ lašo metu tamsaus lauko ir fazinio kontrasto mikroskopuose arba šviesaus lauko mikroskopijoje, kai kondensatorius nuleistas ir mikroskopo diafragma iš dalies. uždengtas.
Flagellum dažymas Lefflerio metodu
Mikroskopu Galutinis produktasžvyneliai matomi kaip ploni siūlai panašūs dariniai.
#22 PiliPadengtas enterobakterijų ir kelių kitų mikroorganizmų paviršius didelis skaičius(nuo 10 iki kelių tūkstančių) gaureliai – gijiniai baltyminio pobūdžio dariniai. Kaip ir žvyneliai, jie yra pagaminti iš vieno tipo baltymų - pilino, kurio subvienetai yra suskirstyti į tuščiavidurį siūlą viduje ir kilę iš CPM. Jos trumpesnės ir plonesnės už žvynelius, plotis 10–12 nm, ilgis iki 12 μm.
Villi yra polifunkciniai: užtikrina perduodamą genų perdavimą (konjugaciją), yra fagų receptoriai, organas, skirtas bakterijoms prisijungti prie maistinių medžiagų substrato (adhezija), dalyvauja metabolitų pernešime.
Streptokokai turi išorinį baltymų plaukelių sluoksnį (fimbria), kurie vadinami baltymu M (M baltymu). Ši voverė žaidžia svarbus vaidmuo bakterijų sąveikos su makroorganizmu procesuose.
№23 Actinomycetes
Jie atstovauja savotiškai bakterijų grupei, kuri atrodo kaip maži arba ilgi nesišakojantys siūlai. Hifų sankaupa vadinama grybiena. Panašumas į grybus yra grynai išorinis, nes aktinomicetai turi prokariotinių ląstelių tipą su ląstelės sienele, kurioje nėra chitino ir celiuliozės. Aktinomicetai yra gramteigiami, daugelis formų yra atsparios rūgštims, kai kurie aktinomicetai turi kapsulę aplink siūlus.
Substrato grybiena susidaro dėl grybienos įaugimo į maistinę terpę ir orą, augant terpės paviršiuje (19 pav.). Pažeistuose audiniuose (audinių forma) aktinomicetai gali sudaryti drūzų granules iš tankiai susipynusių siūlų spindulių pavidalu, besitęsiančių iš centro ir baigiant kolbos formos sustorėjimais.
Aktinomicetai dauginasi aseksualiai, suformuojantys konidijas arba sporoforus su sporangijomis oro grybienos galuose. Sporų nešėjai gali būti tiesūs, banguoti, spiraliniai. Sporos – ovalios, apvalios, cilindrinės, lygiu paviršiumi arba spygliais, kartais paslankios dėl žvynelių (zoosporų). Sporos naudojamos aktinomicetams daugintis, jos neatsparios karščiui, bet atlaiko džiūvimą. Be to, galimas grybienos pumpurų atsiradimas ir suskaidymas į lazdelės arba kokos formas.
Aktinomicetai plačiai paplitę gamtoje, gyvena vandenyje, dirvožemyje, kuriame gausu humuso. Jie dalyvauja medžiagų cikle gamtoje. Atskiros rūšys aktinomicetai naudojami kaip antibiotikų, vitaminų, lipidų, proteazių, aminorūgščių, steroidų gamintojai.
Aktinomicetai priklauso Actinomycetales būriui, kuriai priklauso šeimos: Actinomycetaceae, Nocardiaceae, Streptomycetaceae, Mycobacteriaceae.
Žmonėms patogeniškos rūšys aptinkamos tarp Actinomycetaceae ir Nocardiaceae šeimų atstovų. Pirmieji atrodo kaip ilgos arba trumpos šakotos lazdelės, kurios nesudaro oro grybienos. Jie yra žmogaus aktinomikozės sukėlėjai ir formuoja drūzus paveiktuose audiniuose.
Nocardiaceae šeimos atstovai primena mikobakterijas, turi gijines ląsteles ir ant maistinių medžiagų terpėje sudaro oro ir substrato grybieną. Hifai suskaidomi į kokoidines ir lazdelės formos ląsteles. Patogeninė nokardija sukelia nokardiozę.
Tyrimo metodai. nudažyti pagal Gram ir Ziehl-Neelsen. Drūza kilpa pašalinama iš patologinės medžiagos, dedama į vandens lašą ant stiklelio, lengvai prispaudžiama dengiamuoju stikleliu, tada po stiklu įlašinamas lašas šarminio metileno mėlynojo tirpalo ir mikroskopuojamas, fazinis kontrastas. Gali būti naudojamas.
№24 Spirochetes
spirališkai susisukusios bakterijos, turinčios aktyvų judrumą. Spirochetų matmenys svyruoja nuo 0,1-0,3 mikronų storio, 7-500 mikronų ilgio. Įvairūs judesiai – nuo sraigtinio iki lenkimo. Elektroninis mikroskopinis tyrimas leido spirochetose atskirti protoplazminį cilindrą (ląstelės korpusą), ašinį (atraminį) siūlą ir trijų sluoksnių išorinį apvalkalą. Ašinis siūlas yra periplazminėje erdvėje tarp išorinio apvalkalo ir protoplazminio cilindro ir susideda iš atskirų fibrilių (endoflagelių), kurių skaičius įvairiose rūšyse skiriasi: sergant treponema ir leptospira – 3-4; borelijoje - iki 30. Kiekviena iš fibrilių (endoflagella) yra pritvirtinta prie protoplazminio cilindro galų esančių tvirtinimo diskų srityje ir tęsiasi iki priešingo galo, apsivyniodama aplink jį ir baigdama laisvai. Fibrilių cheminė sudėtis panaši į žvynelių (20 pav.).
Protoplazminiame cilindre yra: nukleoidas, ribosomos, mezosomos, inkliuzai. Išorinis apvalkalas (ląstelės sienelė) turi ploną peptidoglikano sluoksnį, yra elastingas ir neturi standumo. Šios bakterijos nesudaro endosporų, kapsulių ir žvynelių, yra gramneigiamos, tepinėlyje išsidėsčiusios atsitiktinai.
Spirochaetai priklauso Spirochaetales būriui, Spirochaetaceae šeimai, kurią sudaro trys gentys:
Borelijos - turi 3-10 nelygių pasvirusių garbanų, galai smailūs, 10-30 mikronų ilgio. Judėjimas trūkčiojantis, pasak Romanovskio-Giemsos, jie nudažyti mėlyna spalva violetinė(atstovas Borreliarecurrentis – sukelia epideminę recidyvuojančią karštligę; Borreliaburgdorferi – sukelia Laimo ligą).
Treponema - turi 8-14 vienodos amplitudės sandariai susuktų garbanų, kurių ilgis 5-15 mikronų. Judėjimas yra sklandus, lėtas, sukasi aplink išilginę ašį, pasak Romanovskio-Giemsos, jie nudažyti blyškiai rožinės spalvos(Treponemapallidum - sifilio sukėlėjo - atstovas).
Leptospira - turi iki dviejų dešimčių mažų dažnų garbanų, kurios baigiasi kabliuku su sagos pavidalo sustorėjimu, 5-15 mikronų ilgio. Judėjimas yra labai aktyvus, judesys į priekį, lenkimas ir sukimasis aplink ašį. Pasak Romanovskio-Giemsos, jie nusidažo silpnai rausvai alyvine spalva (leptospirainterrogans atstovas yra leptospirozės sukėlėjas).
Tyrimo metodai. Gyvoje būsenoje spirochetos tiriamos fazinio kontrasto mikroskopu ir tamsaus lauko mikroskopu, stebint aktyvų būdingas judėjimas spirochetos, jų formos ypatybės.
Preparatai ruošiami pagal Burri (tamsiame preparato fone matomi šviesūs gofruoti spirochetų siūleliai), dažomi pagal Romanovsky-Giemsa, Morozovo metodu.
# 25 Rickettsia
klasė " Alfaproteobakterijos"
Įsakymas Rickettsiales
Šeima Rickettsiaceae
Rickettsia gentis
Šeima Ehrlichiaceae
Riketsijos yra įvairios formos ir skirstomos į šias rūšis:
kokoidas vienagrūdis (iki 0,5 mikrono);
lazdelės formos dvigrūdės (1-1,5 mikrono);
bacilinis trijų-keturių granulių (3-4 mikronai);
siūliškas daugiagrūdis (10-40 mikronų).
Grūdai (nukleoproteinai) aptinkami dažant pagal Romanovsky-Giemsa. Visos formos yra abipusės. Struktūriškai jie turi visus bakterinės ląstelės komponentus: ląstelės sienelę, lipoidinę kapsulę, citoplazmą, nukleoidą, ribosomas, pili. Riketsijose yra ir DNR, ir RNR didelis kiekis fosfolipidai, mažai angliavandenių.
Daugeliu atvejų (išskyrus Rochalimaeaguintana rūšį) riketsijos neauga ant dirbtinių maistinių medžiagų. Riketsijos gyvenimo ciklas priklauso nuo ląstelės-šeimininkės gyvybinės veiklos ir susideda iš dviejų etapų: vegetatyvinio ir ramybės (elementariniai kūnai). Vegetacinės stadijos riketijos (23 pav.) aktyviai dauginasi dvejetainiu dalijimusi ir turi aktyvų judrumą, matyt, dėl žvynelių struktūrų. Ramybės stadijos riketsijos (elementarieji kūnai) yra sferinės ir neaktyvios.
Tyrimo metodai. Riketsijos gerai nudažytos pagal Romanovsky-Giemsa alyvine spalva, pagal Morozovą (sidabravimo metodu) juodai. Norint atskirti riketsiją, dažymo metodas, kurį pasiūlė P.F. Zdrodovskis:
Riketsijos nusidažo rubino raudonai ir lengvai aptinkamos mėlynos citoplazmos ir mėlynos spalvos ląstelės branduolio fone.
№26 Chlamidijos
klasė Chlamidijos
Įsakymas Chlamidijos
Šeima Chlamidijos
Gentis: Chlamidijos
1). Elementarieji kūnai yra mažos (0,2-0,5 μm) elektronų tankios sferinės struktūros, neturinčios metabolito aktyvumo, turinčios kompaktišką nukleoidą ir standžią ląstelės sienelę, kurios filtruojamos per bakterinius filtrus. Jie yra infekcinė chlamidijų pradžia ir užtikrina jų išlikimą tarpląstelinėje aplinkoje bei naujų ląstelių užkrėtimą.
2). Tinkliniai kūnai yra didesni (0,8-1,5 mikrono), sferiniai dariniai, turintys tinklinę struktūrą su plona ląstelės sienele ir fibriliniu nukleoidu. Jie auga iš elementarių kūnų ląstelių viduje, nėra užkrečiami ir, vykstant dalijimuisi, užtikrina chlamidijų dauginimąsi. Iš čia ir kitas, istoriškai pirmasis tinklinių kūnų pavadinimas – „pradinis kūnas“. Tinkliniai kūnai yra vegetatyvinė chlamidijos forma.
3). Tarpiniai kūnai yra tarpinė stadija tarp elementarių ir tinklinių kūnų.
Chlamidijų gyvavimo ciklas prasideda tuo, kad elementarius kūnus fagocituoja ląstelė-šeimininkė, o po to per kelias valandas jie persitvarko, padidėja ir virsta tinklinėmis formomis, kurios dauginasi skersinio dalijimosi būdu. Gyvavimo ciklas baigiasi, kai atsirandančios tarpinės formos sutankinamos, sumažinamos ir virsta elementariais kūnais. Daugindamosi citoplazminių vakuolių viduje, chlamidijos sudaro mikrokolonijas (inkliuzus), apsuptas membrana. Visi trys chlamidijų vystymosi etapai randami mikrokolonijų sudėtyje. Plyšus vakuolės sienelei (pūslelėms) ir ląstelės-šeimininkės membranai, atsipalaiduoja naujai susidariusios chlamidijos, o elementarieji kūnai, užkrėsdami kitas ląsteles, kartoja vystymosi ciklą. Optimaliomis augimo sąlygomis eukariotinėse ląstelėse gyvenimo ciklas chlamidija yra 17-40 valandų (24 pav.).
Chlamidijų ypatumas pasireiškia ir jų ląstelės sienelės sandaroje. Jame nėra peptidoglikano ir yra dviejų sluoksnių membrana, kurios standumą lemia peptidai, sujungti disulfidiniais tilteliais. Priešingu atveju chlamidijos primena gramneigiamas bakterijas, nes jose yra glikolipidų, panašių į lipopolisacharidus.
Chlamydiales būriui priklauso viena šeima Chlamydiaceae ir viena gentis Chlamydia. Žmonėms patogeninės rūšys yra C.trachomatis, C.psittaci, C.pneumoniae. Chlamidijos sukelia akių, kvėpavimo takų ir urogenitalinės sistemos ir susivienyti Dažnas vardas"chlamidija".
Tyrimo metodai. Chlamidijų inkliuzinių kūnų (mikrokolonijų) užkrėstose ląstelėse (audinuose) aptikimui mikroskopiškai naudojami įvairūs dažymo metodai: Romanovsky-Giemsa, Macchiavello ir kt. Beicuojant pagal Romanovsky-Giemsa, jie įgauna mėlyną arba violetinę spalvą. Be to, mikroskopuojant šlapius preparatus po stiklu, naudojant fazinio kontrasto optinę sistemą, chlamidijos yra aiškiai matomos nedažytos. IN Pastaruoju metu dažniausiai naudojama tiesioginė imunofluorescencinė reakcija – akridino oranžinė dėmė.
№27 Mikoplazmos klasė Mollicutes
Įsakymas Mikoplazmatinės
Šeima Mycoplasmataceae
Mycoplasma gentis
Ureaplasma gentis
Penkios rūšys yra patogeniškos žmonėms: M . pneumoniae , M . hominis , M . genitalijų , M . incognitus Ir U . urealyticum .
Mikoplazmos- mažiausi prokariotai (125-150 nm), galintys daugintis savarankiškai. Manoma, kad mikoplazmos yra artimiausi pirminių prokariotinių ląstelių palikuonys. Mikoplazmų genomas ląstelei yra minimalus, jis yra penkis kartus mažesnis už genomą coli ir yra 0,45 MD. Pagrindinis mikoplazmų požymis yra ląstelės sienelės nebuvimas. Juos gaubia į kapsulę panašus sluoksnis, po kuriuo yra tik plona 7,5-10 nm storio trijų sluoksnių membrana, kurioje yra nemažas kiekis cholesterolio. Dėl to mikoplazmos yra išskiriamos į specialų Tenericutes, Mollicutes klasės ("gležni oda") skyrių, užsakymą Mycoplasmatales.
Dėl to, kad nėra ląstelės sienelės, mikoplazmos (21 pav.) yra osmosiškai jautrios ir įvairių formų:
a) mažos 0,2 µm dydžio sferinės arba kiaušiniškos ląstelės (elementarieji kūnai), kurios filtruojamos per bakterinius filtrus;
b) didesnis sferinis, iki 1,5 mikrono dydžio;
c) siūlinės, išsišakojusios ląstelės iki 150 mikronų dydžio.
Mikoplazmos nesudaro sporų, žvynelių, kai kurios rūšys turi sklandų judrumą.
Jie dauginasi dvejetainiu sferinių ir siūlinių ląstelių dalijimusi, pumpurų atsiradimu ir daugelio elementarių kūnų, susidariusių gijose, išsiskyrimu.
Kalbant apie energiją, mikoplazmos ją gauna įprastu fakultatyviniams anaerobams būdu, fermentuodamos angliavandenius arba aminorūgštis. Dėl nedidelio mikoplazmų genomo jų biosintezės galimybės yra ribotos, todėl jas tenka auginti maistinėse terpėse, praturtintose lipidais, baltymais, nukleorūgščių pirmtakais. Jie auga lėtai, kolonijos su tankiu centru auga į terpę, panašios į "keptą kiaušinį" (tamsus centras ir šviesesnė ažūrinė periferija). Kolonijų dydis yra mažas, ne didesnis kaip 600 mikronų.
Žmogaus patologijoje didžiausią vaidmenį atlieka keli Mycoplasma genties atstovai: M.pneumoniae, M.hominis, M.anthritidis ir vienintelė Ureaplasma–U.urealyticum genties rūšis (taip pavadinta dėl ureazės aktyvumo). Patogeninės mikoplazmos sukelia įvairias klinikines apraiškas kvėpavimo takų, urogenitalinių takų ir sąnarių ligas (mikoplazmozę). Gydant šias ligas, reikia atminti, kad mikoplazmos nėra jautrios beta laktaminiams antibiotikams ir kitiems vaistams, slopinantiems ląstelės sienelės sintezę (dėl to, kad jos nėra patogene).
Tyrimo metodai. Šviesos mikroskopu aptinkamos tik didžiausios mikoplazmų formos. Gyvoje būsenoje jie tiriami tamsaus lauko ir fazinio kontrasto mikroskopu, ultrastruktūriniai komponentai aptinkami naudojant elektroninę mikroskopiją.
№28 Grybai
Klasės (7): · Bazidiomicetai(kepurių grybai). · Zygomycetes (gentis Mucor - žmonių ir gyvūnų mukormikozė) Ascomycetes (žudikai, gentys Aspergilas , Penicillium , mielių grybai) Deuteromicetai - netobuli grybai, nesidaugina lytiškai ( Candida )
Grybai yra didelė ir sėkmingai besivystanti organizmų grupė, apimanti apie 80 000 nustatytų rūšių. Jų dydžiai svyruoja nuo vienaląsčių mielių iki didelių žiobrių, pūkelių ir ragų. Grybai užima įvairias buveines – tiek vandenyje, tiek sausumoje. Be to, jie svarbūs dėl vaidmens, kurį jie atlieka biosferoje, ir dėl to, kad žmonės juos naudoja medicinoje ir ekonomikoje.
Tyrimo metodai. Mikroskopiniam tyrimui ruošiami ir natūralūs (nedažyti), ir beicuoti preparatai.
Nedažytų preparatų tyrimas
Siekiant aiškiau atskirti grybelio elementus, preparatas yra nuskaidrintas, tam uždedama patologinė medžiaga (pluta, nago gabalėliai, plaukai, gleivinės įbrėžimai, granuliomatinių židinių turinys). laikrodžio stiklas arba Petri lėkštelė, kur pilamas 10-15% kaustinės natrio arba kalio tirpalas ir 20-30 minučių įstatomas į 37 0 C temperatūros termostatą. Tada medžiaga pašalinama ir įlašinama į 50% glicerolio tirpalo lašą ant stiklelio ir uždengiama dengiamuoju stikleliu, mikroskopiškai fazinio kontrasto arba šviesos mikroskopu. Galima naudoti ir kitą būdą: ant patologinės medžiagos užlašinamas lašas glicerino, pridedant 10 % kaustinio kalio, ir po 4-5 minučių tiriama, uždengiama dengiamuoju stikleliu.
Pūliniai pūliniai, opų turinys, skrepliai skiedžiami fiziologiniu fiziologiniu tirpalu arba vandens-alkoholiu (1:1) arba 50% vandeniniu glicerolio tirpalu, paruošiamas preparatas „smulkintas lašas“ ir tiriamas x200, x400 didinimu naudojant fazinį kontrastą.
Beicuotų preparatų tyrimas
Iš pūlių, kraujo, smegenų skysčio, bronchų plovimo nuosėdų ir šlapimo paruošiami ploni tepinėliai, kurie fiksuojami Nikiforovo, Carnoy, alkoholio-formolio mišinyje, išdžiovinami ir nudažomi:
Dažymas laktofuksinas kurių sudėtyje yra rūgštaus fuksino - 0,1 g, pieno rūgšties - 100 ml. Dažyti 3-5 minutes. Vaisto fonas yra rausvas, grybiena opalizuoja mėlyna spalva. Grybai gerai nusidažo mukormikoze ir aspergilioze.
№29 Mielės ir į mieles panašios Mielių ląstelės yra suapvalintos, pailgos arba pailgos formos, 8–10 mikronų dydžio, su dviejų grandinių membrana. Citoplazmoje pastebimi intarpai glikogeno granulių, volutino ir lipidų pavidalu. Dauginasi pumpurais ir askosporomis. Į mieles panašios mielės panašios į tikras mieles, skiriasi askosporų nebuvimas ir gebėjimas formuoti pseudomicelį. Susidarius pseudomiceliui, ląstelės pailgėja ir liečiasi siauru pagrindu. Jie sukelia kandidozę, kuri išsivysto sergantiems žmonėms, smarkiai sumažėjus organizmo atsparumui ir ilgai vartojant antibiotikus. Mielių tyrimo metodai
Šiuolaikiniam biologinės įvairovės tyrimo laikotarpiui būdinga intensyvi filogenetinės sistematikos plėtra, kuria siekiama rekonstruoti specifinius organizmų istorinės raidos kelius. Mikrobiologijoje filogenetinė sistematika galingą impulsą vystytis gavo tik pačioje XX amžiaus pabaigoje. ryšium su lyginamuoju konservuotų nukleotidų sekų tyrimu rRNR. Mielėse ši taksonomija daugiausia pagrįsta dviejų rDNR regionų, kurių ilgis yra apie 600 nukleotidų porų, tyrimu: D1/D2 domeno, esančio geno, koduojančio 26S rRNR, 5' gale ir vidinio transkribuoto tarpinio regiono (ITS) , kuri apima 5.8S rRNR geną Manoma, kad dėl šių sričių konservatyvumo skirtumai tarp jų yra tiesiogiai proporcingi filogenetiniam atstumui, evoliucinio ryšio laipsniui.Nekleotidinių rDNR sekų sekų nustatymas pasirodė esąs galingas įrankis, leidžiantis konstruoti filogenetinę mieliagrybių klasifikaciją ir nustatyti jų vietą bendroje grybų sistemoje.
Iki šiol rRNR nukleotidų sekos buvo iššifruotos ir patalpintos į kompiuterinius duomenų bankus, prieinamus internete visų atstovų. žinomos rūšys mielės. Tai leidžia kurti filogenetinius medžius, atspindinčius jų ribosomų genų evoliuciją. Paaiškėjo, kad mielių grupavimas pagal rRNR nukleotidų sekų panašumą daugeliu atvejų nesutampa su grupavimu pagal fenotipinius požymius. Daugelis mielių klasifikacijoje naudojamų tradicinių požymių, tokių kaip vegetatyvinio dauginimosi ypatybės, askosporų forma, fermentacija ir cukraus asimiliacija, buvo laikomi nepatikimais, netinkamais filogenetiniams ryšiams nustatyti. rRNR (rDNR) sekos nustatymas dabar laikomas esminiu aprašant naujas mielių rūšis.
Ypač didelę įtaką mielių, kaip ir daugumos kitų mikroorganizmų grupių, tyrimams sparčiai vystėsi XX amžiaus pabaigoje. molekulinė biologija. Šiuolaikinėje mielių taksonomijoje plačiai naudojami genų sistematikos metodai, pagrįsti tiesioginiu genomų palyginimu ir nukleotidų sekų nustatymu. Vieningų molekulinės biologijos metodų naudojimas leido dar labiau suartinti mielių ir siūlinių grybų taksonomiją, nustatyti ryšius tarp mielių anamorfų ir siūlinių teleomorfų bei sukurti naujus kriterijus, kaip sukurti vieningą filogenetinę sistemą visai Mikotos karalystei. . Tuo pačiu metu naujos žinios sukėlė naujų mokslines problemas ypač koreliacijos tarp naujausių molekulinių metodų ir tradicinių metodų, pagrįstų morfologiniais ir fiziologiniais mielių tyrimo metodais, problema. Saccharomyces cerevisiae genomas buvo beveik visiškai iššifruotas, o tai atveria puikias mielių genomikos perspektyvas ir naujus jų biotechnologinio panaudojimo horizontus. Taigi mielių mokslas, nukeliavęs daugiau nei pusantro šimtmečio, ir toliau sparčiai vystosi XXI amžiuje.
Mikroskopiniam tyrimui ruošiami ir natūralūs (nedažyti), ir beicuoti preparatai. Norint ištirti grybelius audiniuose, atliekamas patohistologinis tyrimas.
ląstelių mielių sporų grybiena
30. Gijinių grybų morfologija ir tyrimo metodai. Yra 2 grybų augimo tipai: hifinis augimas (hifomicetai) ir mielių augimas (blastomicetai). Paprastai siūlinių grybų vegetatyvinį kūną sudaro maždaug 5 µm storio gijos, labai išsišakojusios ir vadinamos hifais. Hifai arba neturi skersinių pertvarų (apatiniuose grybuose) arba pertvaromis (septomis) yra atskirtos į ląsteles aukštesni grybai). Ląstelės sienelė gali būti įvairaus storio, dažnai aiškiai matomas aplinkkelis, tarp citoplazmoje esančių intarpų būdingiausi volutino, glikogeno, melanino pigmento grūdeliai. Subrendusiose senose grybelių ląstelėse gausu lipidų. Branduolys turi branduolį ir chromatino tinklą; ląstelės gali būti daugiabranduolės. Hifų rinkinys sudaro grybieną (micelį). Grybiena gali būti substratas, susidaręs dėl hifų įaugimo į maistinę terpę, ir oras, augantis terpės paviršiuje. Grybiena žymi išsišakojusius vamzdelius, išsišakojimą atlieka šoninės hifų ataugos. Grybų gijos kartais išsidėsčiusios lygiagrečiomis eilėmis, glaudžiai greta viena kitos, panašios į „dagtį“, taigi ir pavadinimas „koremija“ (dermatofituose).
31. Pagrindiniai pirmuonių klasifikavimo ir morfologijos principai. Jų tyrimo metodai ir medicininė reikšmė.
Pirmuonys yra vienaląsčiai eukariotai, savo struktūra panašūs į sudėtingai organizuotų gyvūnų ląsteles. Forma gali būti kriaušės (Trichomonas, Giardia), kiaušiniška (Balantidia), verpstės formos (Trypanosoma, Leishmania), gali įgauti pačią keisčiausią konfigūraciją (ameba)
Dauguma pirmuonių yra judrūs, o judėjimas vyksta pseudopodijų (amebų, maliarijos plazmodijus), žvynelių (giardijos, leišmanijos), blakstienų (balantidijų) pagalba.
Pseudopodia- laikini citoplazmos išsikišimai, kuriuos išleisdami pirmuonys visą laiką keičia kūno formą.
Paprasčiausi priklauso pirmuonių karalystei (protos – pirmieji, zoa – gyvūnai). Medicinos svarbos yra:
Phylum Sarcomastigophora, pofilis Sarcodina (Sarcodidae). Jų kūne nėra dėmių, jie juda pseudopodijų pagalba. Ši klasė apima Skirtingos rūšys ameba, įskaitant dizenterinę amebą (Entamoeba histolytica).
Tyrimo metodai. Pirmuonių tyrimui ruošiami laikini ir nuolatiniai (dažyti) preparatai. Laikini preparatai ruošiami „susmulkinto lašo“ arba „pakabinamo lašo“ metodu, pridedant šilto. fiziologinis tirpalas arba gyvybiškai svarbūs intravitaliniai dažai. Iš kraujo preparatų ruošiamas „storas lašas“. Norėdami tai padaryti, eteriu apdorotas pirštas praduriamas žemyn ir prie išsikišusių lašų atnešamas stiklelis, ant kurio paimami 2-3 kraujo lašai, o tada kraujas paskirstomas adata arba kampu. kitą stiklinę, kad gautumėte apie 1 cm ovalą, kad pagreitintumėte vaisto džiūvimą, jį galima įdėti į 35-37 0 C temperatūros termostatą.
Cistoms aptikti naudojamas stiprus Lugolio tirpalas, kuris nudažo cistų struktūras tamsiai ruda spalva.
Dažyta geležies hematoksilinu pagal Heidenhaino metodą
Tepinėliai po fiksavimo dedami į 2,5% geležies alūno tirpalą 1 valandai.
Tris kartus perplovus vandenyje, beicuoti dažais (0,5 g hematoksilino, 10 ml 96 0 alkoholio, o ištirpinus įpilti 90 ml distiliuoto vandens) 5-10 min.
Nuplauti vandeniu ir išdžiovinti.
Dažymas yra būtinas tais atvejais, kai reikia atskleisti smulkiausias pirmuonių branduolio ir citoplazmos struktūros detales.
Augalų ląstelių sienelės turi nepaprastą stiprumą, o augimo procese augalai keičia savo struktūrą ir sudėtį. Pagrindiniai augalų ląstelių sienelių komponentai yra polisacharidai, tarp kurių vyrauja celiuliozė, kuri daugiausia lemia sienelės architektūrą.
Celiuliozė. Šis homopolisacharidas yra labiausiai paplitęs angliavandenis Žemėje (augalai per metus suformuoja iki 1011 tonų celiuliozės). Celiuliozės monomerai yra gliukozės likučiai, sujungti ilgomis grandinėmis (kiekvienoje iki 10 000 gliukozės likučių), naudojant b (1 → 4) -glikozidinius ryšius (5.3 pav.). Tokioje molekulėje nėra visiškos sukimosi laisvės aplink 1C-O- ir O-4C-jungtis, o polimeras įgyja palankią konformaciją tarpgrandiniams vandeniliniams ryšiams susidaryti, kai grandinės yra išsidėsčiusios antilygiagrečiai. Dėl to celiuliozės molekulės sujungiamos į mikrofibrilės apie 10–25 nm storio. Mikrofibrilės susipina ir susidaro ploni siūlai, kurios, savo ruožtu, gali būti apvyniotos viena aplink kitą, kaip sruogos virve, formuojant makrofibrilės. Kiekviena makrofibrilė yra apie 0,5 µm storio ir gali siekti 6–8 µm ilgį. Makrofibrilių stiprumas yra panašus į plieninės vielos stiprumą, vienodo storio. Be to, atskiri skyriai mikrofibrilės turi tvarkingą struktūrą ir suteikia ląstelės sienelei kristalinių savybių. Taigi galima pastebėti celiuliozės sudėtingumą ir aukštą tvarkingumą ląstelių sienelių sudėtyje, o tai nėra atsitiktinumas: šis polimeras atlieka apsaugines ir palaikomąsias funkcijas augale.
Šioje formoje polisacharidai yra nepasiekiami savo fermentų veikimui, o celiuliozės augalas negali naudoti kaip atsarginės medžiagos. Tik keli organizmai (kai kurios bakterijos, grybai, pirmuonys ir reti gyvūnai) turi fermentų sistemas, galinčias skaidyti celiuliozę.
Augalų ląstelės sienelėje yra celiuliozės mikro- ir makrofibrilės matrica, kuris taip pat daugiausia susideda iš polisacharidų ir keičia savo struktūrą augalų augimo metu. Įjungta ankstyvosios stadijos raidos matrica susideda iš pektino medžiagos, o vėliau jame pasirodys ksilanai ir įvairūs neutralūs polisacharidai („hemiceliuliozė“). Pektino medžiagos yra a-galakturono rūgšties polimerai, kuriuose kai kurie vandenilio atomai pakeisti metilo grupėmis (-CH 3) (5.3 pav.). Ksilanai yra ksilozės polimerai (5.3 pav.).
Vėlesniuose vystymosi etapuose, kai vyksta ląstelių sienelių lignifikacija, lignino- chemiškai atsparaus polimero, kurio sudėtyje yra didelis skaičius aromatiniai žiedai. Be to, augalų ląstelių sienelių sudėtyje yra nedidelis kiekis glikoproteinų, netirpių lipidų polimerų. skirtinga struktūra ir vaškai.
Kai kurių augalų ląstelių sienelėse yra retų neįprastos struktūros polisacharidų. Pavyzdžiui, sienose ir tarpląsteliniame
Jūros raudonųjų dumblių medžiagoje yra sudėtingas heteropolisacharidinis agaras, kuris yra sulfatuotų polisacharidų - agarozės ir agaropektino mišinys. Agarozė sudaryta iš kintančių D-galaktozės ir 3,6-anhidro-L-laktozės liekanų, pakaitomis sujungtų b(1→4)- ir a(1→3)-ryšiais. Agaropektino turi daugiau sudėtinga struktūra: Sudėtyje yra D-galaktozės, 3,6-anhidrogalaktozės, urono rūgščių ir sulfato. Agaras naudojamas kaip labiausiai paplitęs tankintuvas kietoms terpėms, nepakeičiamas mikrobiologijoje, taip pat Maisto pramone gaminiams želuoti. Reikia pažymėti, kad didžioji dauguma mikroorganizmų nesugeba suskaidyti agaro, ir tai yra vienas pagrindinių jo pranašumų prieš kitą maistinės terpės tankintuvą – želatiną. Agarozės radiniai platus pritaikymas biocheminiuose tyrimuose: ji vandens aplinka sudaro gelį su didelėmis poromis, kurių dydį lemia jo koncentracija. Agarozės geliai naudojami baltymų ir nukleorūgščių frakcionavimui, taip pat ląstelių imobilizavimui.
- 20,44 KbLąstelės sienelė, struktūra, cheminė sudėtis. Ląstelės sienelės pokyčiai.
Stipraus apvalkalo buvimas charakteristika augalo ląstelė, kuri ją skiria nuo gyvūninės ląstelės. Apvalkalas suteikia ląstelei tam tikrą formą ir stiprumą bei apsaugo gyvąjį turinį – protoplastą ir, svarbiausia, plazmalemą, tvirtai prispaustą prie apvalkalo iš vidaus. Specialiose maistinėse terpėse auginamos aukštesniųjų augalų ląstelės, kuriose membrana pašalinama fermentiniu būdu, visada įgauna sferinę formą. Be apvalkalo augalo ląstelės egzistavimas normaliomis sąlygomis yra neįmanomas, nes jai būdingas turgorinis slėgis, subalansuotas apvalkalo priešslėgio, neišvengiamai sukeltų plazmalemos plyšimą ir protoplasto sunaikinimą. Ląstelių sienelių visuma sausumos augalo kūno sudėtyje, kuri iškyla virš dirvos paviršiaus ir neturi vidinio skeleto, yra tam tikras atraminis karkasas, suteikiantis augalui mechaninį stiprumą.
Korpusas, kaip taisyklė, yra bespalvis ir skaidrus, lengvai praleidžia saulės šviesą. Juo gali judėti vanduo ir ištirpusios mažos molekulinės masės medžiagos. Kiekviena ląstelė turi savo apvalkalą; gretimų ląstelių apvalkalai yra tarsi sucementuoti tarpląstelinėmis medžiagomis, kurios sudaro vadinamąją medianinę sluoksnį. Dėl to kaimyninės ląstelės yra atskirtos viena nuo kitos sienele, kurią sudaro du apvalkalai ir vidurinė plokštė, todėl apvalkalą galima vadinti ir ląstelės sienele.
Apvalkalą sudaro ląstelės protoplastas, todėl gali augti tik kontaktuodamas su juo. Labai dažnai apvalkalas išsilaiko ilgiau nei protoplastas (kai ląstelė anksti miršta), tačiau tai dažniausiai nenutraukia ląstelės, kaip struktūrinio vieneto, egzistavimo, nes ląstelės forma nesikeičia dėl apvalkalo stiprumas. Todėl net ir po mirties augalų ląstelės gali atlikti svarbias tirpalų transportavimo ar mechaninės paramos funkcijas. Daugelio rūšių augalų ląstelės – skaidulos, tracheidės, kraujagyslių segmentai, kamštienos ląstelės – suaugusiųjų būsenoje yra viena ląstelės membrana. Dažniausiai medieną sudaro negyvų ląstelių apvalkalai.
Ląstelių membranos yra tokios pat įvairios, kaip ir pačios ląstelės. Iš membranos sudėties ir struktūros dažnai galima spręsti apie įvairių tipų ląstelių kilmę ir funkciją. Taigi iškastinių augalų struktūra daugiausia tiriama tiriant jų ląstelių membranas.
Iš pradžių pirminė ląstelės sienelė kyla iš plazmalemos. Jį sudaro polisacharidai - pektinas ir celiuliozė. Kaimyninių ląstelių pirminės ląstelių sienelės yra sujungtos protopektino mediana. Ląstelės sienelėje linijinės, labai ilgos celiuliozės molekulės, susidedančios iš gliukozės, surenkamos į ryšulius – miceles, kurios, savo ruožtu, susijungia į fibriles – ploniausias neriboto ilgio skaidulas. Celiuliozė sudaro daugiamatį karkasą, kuris yra panardintas į amorfinę labai drėkinamą neceliuliozės angliavandenių matricą: pektinus, hemiceliuliozes ir kt. Būtent celiuliozė lemia ląstelės sienelės tvirtumą. Mikrofibrilės yra elastingos ir turi panašų tempimo stiprumą kaip plieno. Matriciniai polisacharidai lemia tokias sienelės savybes kaip didelis pralaidumas vandeniui, ištirpusios mažos molekulės ir jonai, stiprus brinkimas. Matricos dėka vanduo ir medžiagos gali judėti iš ląstelės į ląstelę išilgai greta esančių sienelių. Kai kurios hemiceliuliozės gali būti nusėdusios sėklų ląstelių sienelėse kaip atsarginės medžiagos.
Apvalkalo medžiaga, įskaitant celiuliozę, plačiai naudojama pramonėje. Medienos, popieriaus, tekstilės medžiagų savybės didele dalimi priklauso nuo apvalkalo molekulinės struktūros ir cheminės sudėties, o jų išmanymas padeda tobulinti gamybos technologijas ir gaminių kokybę.
Cheminė apvalkalo sudėtis ir molekulinė struktūra. Ląstelės membrana daugiausia sudaryta iš polisacharidų, kurių monomerai - cukrūs - yra tarpusavyje sujungti glikozidine jungtimi (-O-) grandinės pavidalu. Polisacharidai pavadinti pagal juos sudarančių monomerų sudėtį, pridedant galūnę „an“. Pavyzdžiui, polisacharidas ksilanas susideda iš ksilozės cukraus likučių, manano – manozės, gliukano – gliukozės ir kt.
Be polisacharidų, apvalkalo sudėtyje gali būti baltymų, mineralinių druskų, lignino, pigmentų, lipidų. Paprastai kriauklės yra impregnuojamos vandeniu. Lukšto polisacharidai pagal savo vaidmenį gali būti suskirstyti į skeleto medžiagas ir matricines medžiagas. Korpuso skeleto medžiaga aukštesni augalai yra celiuliozė (pluoštas), chemiškai atstovaujanti β-1, 4-D gliukanui. Gliukozės likučių skaičius molekulėje ir, atitinkamai, pačių molekulių ilgis skirtingų augalų ląstelių sienelėse gali labai skirtis, o tai turi įtakos celiuliozės savybėms. Jo gijinės molekulės apvalkale išsidėsčiusios lygiagrečiai viena kitai ir sugrupuotos keliomis dešimtimis taip, kad atsiranda kristalams būdinga pailga trimatė gardelė. Šios kristalinės grupės, kurių formavime dalyvauja kovalentiniai ir vandeniliniai ryšiai, sudaro mikrofibrilių – ploniausių skaidulų, kurios sukuria struktūrinį apvalkalo karkasą, jo „sustiprinimą“ – pagrindą.
Mikrofibrilėse esančių celiuliozės molekulių kristalinė būsena lemia ląstelės sienelės dvigubą lūžimą, kuris stebimas poliarizaciniu mikroskopu. Mikrofibrilėse yra parakristalinės sritys, t.y. sritys su laisvesne lygiagrečia, bet neturinčia trimatės orientacijos, celiuliozės molekulių išsidėstymo. Mikrofibrilių kristališkumo laipsnis, kristalinių ir parakristalinių zonų santykis gana stipriai skiriasi priklausomai nuo augalo rūšies, ląstelės būklės ir apvalkalo tipo, kuris veikia mechanines ir fizines celiuliozes kaip žaliavas. Skirtingose ląstelėse skiriasi ir mikrofibrilių skersmuo (dažniausiai 10-30 nm ribose), ilgis gali siekti kelis mikrometrus. Mikrofibriles galima sujungti į atskiras grupes arba sluoksnius – 0,4-0,5 mikrono storio makrofibriles, matomas šviesos mikroskopu.
25 pav
Ląstelių gyvavimo metu, ypač priklausomai nuo kokios nors specializuotos funkcijos atlikimo, ląstelės sienelėje vyksta antriniai pokyčiai. Dažnai jie yra susiję su radikaliais jo cheminės sudėties, struktūros ir fizikinių bei cheminių savybių pokyčiais.
Miško ruošimas. Ląstelės sienelė inkrustuojama (impregnuojama) specialia medžiaga – ligninu, kuri padidina ląstelės sienelės kietumą, kaloringumą, tankį ir mažina jos plastiškumą bei gebėjimą augti. Lignified ląstelių sienelės nepraranda savo gebėjimo praleisti vandenį ir orą. Jų protoplastas gali išlikti gyvas, nors dažniausiai miršta. Miško ruoša gamtoje yra labai paplitusi. Jis suteikia tvirtumo medžių kamienams ir šakoms. Spygliuočių ir kietmedžio medienoje yra iki 50 % celiuliozės ir 20...30 % lignino. Sudegusios ląstelių sienelės ir daug žolelių, ypač besibaigiant auginimo sezonui.
Zondavimas arba suberinizacija. Dėl gausaus chemiškai atsparios medžiagos - suberino - sluoksniavimosi ląstelės sienelėje atsiranda jo suberinas. Pagal savo cheminę prigimtį suberinas yra artimas riebalams. Tai amorfinis hidrofobinis junginys, susidedantis iš daug polimerų turinčių sočiųjų riebalų rūgščių ir kompleksinių hidroksi rūgščių. Suberinas daugiausia nusėda antrinio vidinio audinio - kamštienos, kuris yra puikus vandens ir šilumos izoliatorius, - ląstelių sienelėse. Mažais kiekiais jis kaupiasi įvairių audinių ląstelių sienelėse. Visiškai užsikimšus, ląstelės protoplastas miršta, nes suberino lamelės neleidžia prasiskverbti vandeniui ir orui, o iš dalies - protoplastas išsaugomas ilgą laiką.
Mineralizacija. Gyvybės procese ūglių paviršinių ląstelių sienelėse gali kauptis mineralinės medžiagos, ypač silicio dioksidas ir kalcio karbonatas.Kai kurios augalų grupės, pavyzdžiui, daugelio rūšių viksvos, žolės, asiūkliai, visada turi vidinio audinio ląstelių. , epidermis, stipriai inkrustuotas druskomis. Be to, gali būti inkrustuota ne tik ląstelės sienelė, bet ir įvairios epidermio ataugos – trichomos. Be to, mineralinės medžiagos nusėda ant vidinio audinio – epidermio – ląstelių paviršiaus. Dėl mineralizacijos ašiniai organai įgauna didesnį mechaninį stiprumą.
Lieknėjimas. Celiuliozės ir pektino pavertimas šalia jų esančiomis gleivėmis ir dantenomis, kurios yra polimeriniai angliavandeniai, pasižymintys gebėjimu stipriai išsipūsti sąlytyje su vandeniu. Gleivės pastebimos sėklų apvalkalo ląstelių ląstelėse, pavyzdžiui, linuose. Gleivių susidarymas turi didelę adaptacinę reikšmę. Sėkloms dygdamos gleivės jas fiksuoja tam tikroje vietoje, lengvai sugeria ir sulaiko drėgmę, saugo sėklas nuo išdžiūvimo, pagerina sodinukų vandens režimą. Šaknies vloskų ląstelių sienelių gleivės užtikrina tvirtą jų ryšį su dirvožemio dalelėmis.
Maceracija yra tarpląstelinės medžiagos ištirpimas, dėl kurio ląstelės atsiskiria. Natūralus maceravimas vyksta brandžiuose vaisiuose. Dirbtinai atliekama, pavyzdžiui, skiliant linus, kad išsiskirtų verpimo žaliavos – karnienos pluoštų ląstelių grupės.
Darbo aprašymas
Būdingas bruožas yra stipraus apvalkalo buvimas augalo ląstelė tai išskiria jį iš gyvūninės ląstelės. Apvalkalas suteikia ląstelei tam tikrą formą ir stiprumą bei apsaugo gyvąjį turinį – protoplastą ir, svarbiausia, plazmalemą, tvirtai prispaustą prie apvalkalo iš vidaus. Specialiose maistinėse terpėse auginamos aukštesniųjų augalų ląstelės, kuriose membrana pašalinama fermentiniu būdu, visada įgauna sferinę formą.