Klausimas 1. Kuo skiriasi eukariotinių ir prokariotinių ląstelių sandara?
Prokariotai neturi tikros formos branduolio (gr. karyon – branduolys). Jų DNR yra viena apskrita molekulė, laisvai išsidėsčiusi citoplazmoje ir neapsupta membranos. At prokariotinės ląstelės nėra plastidžių, mitochondrijų, endoplazminio tinklo, Golgi aparato, lizosomų. Tiek prokariotai, tiek eukariotai turi ribosomas (branduoliniai turi didesnes). Prokariotinės ląstelės žvyneliai yra plonesni ir veikia kitu principu nei eukariotų žiuželis. Eukariotiniai organizmai yra grybai, augalai, gyvūnai – vienaląsčiai ir daugialąsčiai; prokariotai – bakterijos ir melsvadumbliai (cianobakterijos).
2 klausimas. Papasakokite apie pinotą ir fagocitozę. Kuo šie procesai skiriasi?
Ląstelės membrana yra mobilus darinys, galintis užfiksuoti išorinės aplinkos objektus, formuodamas invaginacijas ir ataugas. Šis procesas vadinamas endocitoze. Endocitozės priežastis yra sudėtingos biocheminės reakcijos, vykstančios citoplazmoje ir pirmiausia susijusios su intracelulinių baltymų tretinės struktūros pokyčiais. Jei ląstelė sugauna skysčio lašą - tai yra pinocitozė, jei kieta dalelė - fagocitozė. Dėl to susidaro pinocitinės arba fagocitinės vakuolės (membraninės pūslelės). Procesas, atvirkštinis endocitozė (vakuolių turinio išsiskyrimas iš ląstelės), vadinamas egzocitoze.
3 klausimas. Išplėskite ryšį tarp ląstelės membranos struktūros ir funkcijų.
Yra žinoma, kad bet kurios membranos pagrindas yra dvisluoksnis (dvigubas) fosfolipidų sluoksnis, kuriame hidrofilinės molekulių „galvutės“ (glicerolis) yra pasuktos į išorę, o hidrofobinės liekanos. riebalų rūgštys- viduje. Baltymų molekulės yra susietos su lipidų dvisluoksniu sluoksniu, kuris gali priglusti prie membranos iš abiejų pusių, į ją įsiskverbti ar net prasiskverbti. Ląstelės membranos padėtis prie ląstelės ribos ir aplinką apibrėžia pagrindines jo funkcijas. Tvirtas, elastingas, lengvai atsinaujinantis dvisluoksnis yra barjeras, užtikrinantis tarpląstelinės aplinkos pastovumą ir apsaugantis citoplazmą nuo pašalinių medžiagų įsiskverbimo. Membranos transportavimo funkcija yra selektyvi. Mažos neįkrautos molekulės (02, N2) lengvai prasiskverbia tiesiai per dvisluoksnį sluoksnį. Didesnės ir (arba) įkrautos dalelės (Na +, K +, kai kurie hormonai) praeina per specialias baltymų poras (kanalus) arba pernešamos baltymų nešikliais. Būdama mobili struktūra, ląstelės membrana taip pat gali pernešti medžiagas endo ir egzocitozės būdu.
4 klausimas. Kokios ląstelės organelės yra citoplazmoje?
Organelės, esančios eukariotinių ląstelių citoplazmoje, gali būti suskirstytos į tris grupes: vienmembranes, dvimembranes ir nemembranas. Vienos membranos organelės apima endoplazminį tinklą (lygų ir šiurkštų), Golgi aparatą, lizosomas ir vakuoles. Du membraninės organelės yra plastidai ir mitochondrijos; nemembraniniai – ribosomos, citoskeletas ir ląstelių centras.
5 klausimas. Apibūdinkite citoplazmos organelius ir jų svarbą ląstelės gyvenime.
Endoplazminis tinklas (ER) yra vakuolių, kanalų ir kanalėlių rinkinys. Jis sudaro vieną tinklą citoplazmos viduje, kartu su išorine membrana. branduolinis apvalkalas. Atskirkite lygų ir grubų EPS. Smooth ER dalyvauja lipidų ir angliavandenių sintezėje, taip pat neutralizuoja toksiškos medžiagos. Ribosomos yra ant šiurkštaus ER membranų paviršiaus.
Golgi aparatas yra vienos membranos organelė, kuri yra vienos ląstelės membranos tinklo dalis ir yra plokščių rezervuarų krūva. Čia vyksta galutinis ląstelių atliekų produktų rūšiavimas ir pakavimas į membranines pūsleles (vakuoles). Be kita ko, Golgi aparatas formuoja lizosomas ir užtikrina egzocitozę.
Lizosomos yra mažos membraninės pūslelės, kuriose yra virškinimui reikalingų fermentų maistinių medžiagų. Lizosomos susilieja su endocitine vakuole, sudarydamos virškinimo vakuolę. Jei lizosomų turinys išsiskiria pačios ląstelės viduje, vyksta jos autolizė (savaiminis ląstelės virškinimas).
Mitochondrijos yra klasifikuojamos kaip dviejų membranų organelės. Jų išorinė membrana lygi, o vidinė formuoja raukšles (cristae). Mitochondrijos yra ląstelės energijos stotys, kurių pagrindinė funkcija yra ATP sintezė.
Plastidės yra dviejų membranų augalų ląstelių organelės. Yra trijų tipų plastidai: chloroplastai, chromoplastai ir leukoplastai. Bespalviai leukoplastai kaupia krakmolą; žalieji chloroplastai vykdo fotosintezę; oranžinės, geltonos ir raudonos spalvos chromoplastai suteikia vaisių ir gėlių spalvą (pritraukia apdulkintojus ir sėklų platintojus). Manoma, kad tolimoje praeityje mitochondrijos ir plastidai kilo iš prokariotų, kuriuos „prarijo“ eukariotinė ląstelė ir pateko į simbiozę su ja. Mitochondrijos ir plastidai turi žiedinę DNR, savarankiškai sintetina kai kuriuos baltymus, o jų ribosomos yra mažesnės nei eukariotinių.
Ribosomos yra mažos daugybės nemembraninių organelių, sudarytų iš dviejų subvienetų - didelio ir mažo. Subvienetus sudaro baltymai ir ribosominė RNR. Ribosomų funkcija yra baltymų sintezė. Kai kurios ribosomos yra tiesiai citoplazmoje, o kai kurios yra ant šiurkščiojo ER membranų.
Ląstelių centras – gyvūnų ląstelių, grybų ir nemembraninės struktūros organoidas žemesni augalai. Jį sudaro du centrioliai, savo forma panašūs į cilindrus ir sudaryti iš mažiausių baltymų vamzdelių; dalyvauja formuojant dalijimosi veleną.
Vakuolė yra membraninė pūslelė, užpildyta ląstelių sultimis. Jo turi būti augalo ląstelėje. Vakuolės funkcija yra vandens, druskų ir maistinių medžiagų kaupimasis. Jame taip pat gali būti pigmentų (mėlynos, violetinės) ir kaupiasi medžiagų apykaitos atliekos.
Citoskeletas yra ne membraninė organelė, kuri yra baltyminis laido vamzdelis, esantis šalia membranos ir citoplazmoje. Jų funkcija – palaikyti ląstelės formą, užtikrinti medžiagų pernešimą ląstelėje, taip pat aktyvų ląstelės (amebos, fagocito) judėjimą. Membraninės pūslelės, kuriose yra bet kokių medžiagų, gali judėti išilgai citoskeleto vamzdelių, tarsi ant bėgių.
Kaip atsisiųsti nemokamą esė? . Ir nuoroda į šį rašinį; eukariotinė ląstelė. Citoplazma. Organelės jau yra jūsų žymėse.Papildomi rašiniai šia tema
Klausimas 1. Kur susidaro lizosoma? Lizosomos yra membraninės struktūros, kuriose yra daug aktyvių fermentų, dalyvaujančių skaidant stambiamolekulinius junginius: baltymus, lipidus, angliavandenius. Lizosomos susidaro Golgi komplekse, kur jos patenka ir kur kaupiasi fermentai. 2 klausimas. Kokią funkciją atlieka mitochondrijos? mitochondrijos - ląstelių struktūros padengtas dviguba membrana. Ant vidinės membranos, kurioje yra daug ataugų, yra puiki suma fermentai, dalyvaujantys ATP sintezėje. Vadinasi, pagrindinė funkcija mitochondrijos – aprūpina ląsteles energija per ATP sintezę. Klausimas 3. Kas
1 klausimas. Kokius prokariotų primityvumo požymius, palyginti su eukariotais, galite įvardyti? Prokariotams trūksta branduolio ir chromosomų. Paveldimą prokariotų informaciją vaizduoja viena DNR molekulė, kuri yra uždara žiedo pavidalu ir yra citoplazmoje. Prokariotų ribosomos yra labai mažos. L organelės, apsuptos membranomis (endoplazminis tinklas, Golgi aparatas, mitochondrijos, plastidės), prokariotai iš viso neturi. Eukariotų ląstelėje šios struktūros skirtos įvairių junginių sintezei. Dėl jų nebuvimo prokariotuose reakcijos
1 klausimas. Iš ko susidaro endoplazminio tinklo sienelės ir Golgi kompleksas? Endoplazminio tinklo sieneles ir Golgi kompleksą sudaro vienasluoksnė membrana. 2 klausimas. Įvardykite endoplazminio tinklo funkcijas. Endoplazminis tinklas (ER) sudaro ląstelės transportavimo sistemą. Sklandžiai ER atliekama riebalų ir angliavandenių sintezė. Grubiame (granuliuotame) ER baltymai sintetinami dėl ribosomų, prijungtų prie ER membranų, darbo. 3 klausimas. Kokia yra ribosomų funkcija? Pagrindinė ribosomų funkcija yra baltymų sintezė. 4 klausimas. Kodėl dauguma ribosomų yra endoplazmos kanaluose
Testas tema „Kerpės. Dumbliai 1. Fotosintezė dumbliuose vyksta: a) branduolyje c) citoplazmoje b) chromatoforuose d) chloroplastuose 2. kerpėse kaip vienas organizmas dauginimosi būdas būdingas: a) lytinei c) partenogenezei b) nelytinei d) teisingo atsakymo nėra 3. Kerpės vadinamos oro grynumo rodikliais, nes: a) valo orą b) teršia orą c) sukuria buveinę kiti augalai ir gyvūnai d) auga tik švaraus oro vietose 4. Ar sugeba aktyvus judėjimas jūros dumbliai? a) visi dumbliai gali judėti b) judėti
Klausimas 1. Kokios yra funkcijos išorinė membrana ląstelės? Išorinė ląstelės membrana susideda iš dvigubo lipidų sluoksnio ir baltymų molekulių, kurių dalis yra paviršiuje, o dalis prasiskverbia pro ir per abu lipidų sluoksnius. Išorinė ląstelės membrana atlieka apsauginę funkciją, atskiria ląstelę nuo išorinės aplinkos, neleidžia pažeisti jos turinio. Be to, išorinė ląstelės membrana užtikrina medžiagų transportavimą į ląstelę ir iš jos, leidžia ląstelėms sąveikauti tarpusavyje. 2 klausimas. Kokiais būdais įvairios medžiagos gali patekti į ląstelę? Per išorę
1 klausimas. Kokie yra pagrindiniai gyvybės atsiradimo ir vystymosi Žemėje etapai? Yra daugybė hipotezių, bandančių paaiškinti gyvybės kilmę ir vystymąsi mūsų planetoje. Ir nors jie siūlo skirtingus šios problemos sprendimo būdus, dauguma jų daro prielaidą, kad egzistuoja trys evoliucijos etapai: cheminė, prebiologinė ir biologinė evoliucija. Cheminės evoliucijos stadijoje vyko abiogeninė organinių polimerų sintezė. Antrajame etape susidarė baltymų-nukleorūgščių-lipoidų kompleksai (mokslininkai juos vadino skirtingai: koacervatais, hiperciklais, probionais, progenotais ir.
1 klausimas. Kokias funkcijas atlieka ląstelių centras? Ląstelės centras atlieka ląstelės vidinio skeleto (citoskeleto) formavimo funkciją. Citoskeletas – tai mikrovamzdelių tinklas, prasiskverbiantis į citoplazmą, išlaikantis ląstelės formą, užtikrinantis ląstelės organelių judėjimą, taip pat specializuotų judėjimo organelių – blakstienų ir žvynelių – darbą. Ląstelių centras taip pat užtikrina normalų ląstelių dalijimąsi. Ląstelės centro centrioliai nukrypsta į besidalijančios ląstelės polius ir sudaro dalijimosi veleną, dėl kurio iš vienos motininės ląstelės vėliau susidaro dvi dukterinės ląstelės. Centrioliai yra sudaryti iš cilindrų
Citoplazma- privaloma ląstelės dalis, uždaryta tarp plazminės membranos ir branduolio; Jis skirstomas į hialoplazmą (pagrindinę citoplazmos medžiagą), organelius (nuolatinius citoplazmos komponentus) ir inkliuzus (laikinus citoplazmos komponentus). Cheminė citoplazmos sudėtis: pagrindas yra vanduo (60-90% visos citoplazmos masės), įvairūs organiniai ir neorganiniai junginiai. Citoplazma yra šarminė. Būdingas eukariotinės ląstelės citoplazmos bruožas yra nuolatinis judėjimas ( ciklozė). Jis aptinkamas pirmiausia dėl ląstelių organelių, tokių kaip chloroplastai, judėjimo. Jei citoplazmos judėjimas sustoja, ląstelė miršta, nes tik būdama joje nuolatiniame judėjime, jis gali atlikti savo funkcijas.
Hialoplazma ( citozolis) yra bespalvis, gleivingas, tirštas ir skaidrus koloidinis tirpalas. Būtent jame vyksta visi medžiagų apykaitos procesai, užtikrinamas branduolio ir visų organelių tarpusavio ryšys. Priklausomai nuo skysčio dalies arba didelių molekulių dominavimo hialoplazmoje, išskiriamos dvi hialoplazmos formos: sol- skystesnė hialoplazma ir gelis- tankesnė hialoplazma. Tarp jų galimi abipusiai perėjimai: gelis virsta zoliu ir atvirkščiai.
Citoplazmos funkcijos:
- visų ląstelės komponentų integravimas į vieną sistemą,
- aplinka daugeliui biocheminių ir fiziologinių procesų,
- aplinka organelių egzistavimui ir funkcionavimui.
Ląstelių sienelės
Ląstelių sienelės apriboti eukariotų ląsteles. Kiekvienoje ląstelės membranoje galima išskirti bent du sluoksnius. Vidinis sluoksnis yra greta citoplazmos ir yra pavaizduotas plazmos membrana(sinonimai – plazmolema, ląstelės membrana, citoplazminė membrana), virš kurios susidaro išorinis sluoksnis. IN gyvūnų narvas jis yra plonas ir vadinamas glikokaliksas(susidaro iš glikoproteinų, glikolipidų, lipoproteinų), augalo ląstelėje – storas, vadinamas ląstelių sienelės(susidaro iš celiuliozės).
Visi biologinės membranos turi bendrų struktūrinių savybių ir savybių. Šiuo metu visuotinai priimta membranos struktūros skysčio mozaikinis modelis. Membranos pagrindas yra lipidų dvisluoksnis sluoksnis, kurį daugiausia sudaro fosfolipidai. Fosfolipidai yra trigliceridai, kuriuose viena riebalų rūgšties liekana yra pakeista fosforo rūgšties liekana; molekulės atkarpa, kurioje yra fosforo rūgšties liekana, vadinama hidrofiline galvute, pjūviai, kuriuose yra riebalų rūgščių likučiai, vadinami hidrofobinėmis uodegomis. Membranoje fosfolipidai išsidėstę griežtai tvarkingai: hidrofobinės molekulių uodegos yra viena prieš kitą, o hidrofilinės galvutės – į išorę, vandens link.
Be lipidų, membranoje yra baltymų (vidutiniškai ≈ 60%). Jie lemia daugumą specifinių membranos funkcijų (tam tikrų molekulių pernešimas, reakcijų katalizė, signalų iš aplinkos priėmimas ir konvertavimas ir kt.). Išskirkite: 1) periferiniai baltymai(esantis ant išorinio arba vidinio lipidų dvigubo sluoksnio paviršiaus), 2) pusiau integruoti baltymai(panardintas į lipidų dvigubą sluoksnį į skirtingus gylius), 3) integraliniai arba transmembraniniai baltymai(persiskverbia pro membraną ir per ją, tuo pačiu metu liečiantis tiek su išorine, tiek su vidine ląstelės aplinka). Integraliniai baltymai kai kuriais atvejais vadinami kanalus formuojančiais arba kanalais, nes jie gali būti laikomi hidrofiliniais kanalais, kuriais polinės molekulės patenka į ląstelę (membranos lipidų komponentas jų nepraleistų).
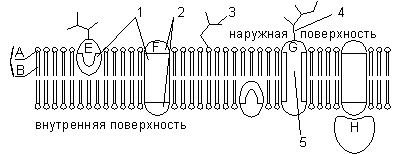
A - hidrofilinė fosfolipido galvutė; C, hidrofobinės fosfolipido uodegos; 1 - hidrofobinės baltymų E ir F sritys; 2, baltymo F hidrofilinės sritys; 3 - šakotoji oligosacharidinė grandinė, prijungta prie lipido glikolipidų molekulėje (glikolipidai yra rečiau nei glikoproteinai); 4 - šakotoji oligosacharidinė grandinė, prijungta prie baltymo glikoproteino molekulėje; 5 - hidrofilinis kanalas (veikia kaip pora, per kurią gali praeiti jonai ir kai kurios polinės molekulės).
Membrana gali turėti angliavandenių (iki 10%). Membranų angliavandenių komponentą sudaro oligosacharidų arba polisacharidų grandinės, susijusios su baltymų molekulėmis (glikoproteinais) arba lipidais (glikolipidais). Iš esmės angliavandeniai yra išoriniame membranos paviršiuje. Angliavandeniai atlieka membranos receptorių funkcijas. Gyvūnų ląstelėse glikoproteinai sudaro kelių dešimčių nanometrų storio epimembraninį kompleksą – glikokaliksą. Jame yra daug ląstelių receptorių, su jo pagalba atsiranda ląstelių adhezija.
Baltymų, angliavandenių ir lipidų molekulės yra judrios, gali judėti membranos plokštumoje. Plazminės membranos storis yra maždaug 7,5 nm.
Membranos funkcijos
Membranos atlieka šias funkcijas:
- ląstelių turinio atskyrimas nuo išorinės aplinkos,
- metabolizmo tarp ląstelės ir aplinkos reguliavimas,
- ląstelės padalijimas į skyrius ("skyrius"),
- „fermentinių konvejerių“ vieta,
- užtikrina ryšį tarp audinių ląstelių daugialąsčiai organizmai(Sukibimas),
- signalo atpažinimas.
Svarbiausias membranos savybė- selektyvus pralaidumas, t.y. membranos yra labai pralaidžios kai kurioms medžiagoms ar molekulėms, o kitoms – prastai (arba visiškai nepralaidžios). Ši savybė yra membranų reguliavimo funkcija, užtikrinanti medžiagų apykaitą tarp ląstelės ir išorinės aplinkos. Procesas, kurio metu medžiagos praeina per ląstelės membraną, vadinamas medžiagų transportavimas. Išskirkite: 1) pasyvus transportas- medžiagų perdavimo procesas, ėjimas be energijos; 2) aktyvus transportas- medžiagų perdavimo procesas, vykstantis su energijos sąnaudomis.
At pasyvus transportas medžiagos juda iš srities, kurioje yra daugiau didelė koncentracijaį plotą su žemesniu, t.y. išilgai koncentracijos gradiento. Bet kuriame tirpale yra tirpiklio ir tirpios medžiagos molekulių. Tirpalų molekulių judėjimo procesas vadinamas difuzija, tirpiklio molekulių judėjimas – osmosu. Jei molekulė yra įkrauta, jos pernešimą veikia elektrinis gradientas. Todėl dažnai kalbama apie elektrocheminį gradientą, sujungiantį abu gradientus. Transporto greitis priklauso nuo gradiento dydžio.
Galima atskirti šių tipų pasyvus transportas: 1) paprasta difuzija- medžiagų pernešimas tiesiai per lipidų dvisluoksnį sluoksnį (deguonis, anglies dioksidas); 2) difuzija membraniniais kanalais- transportavimas per kanalus formuojančius baltymus (Na +, K +, Ca 2+, Cl -); 3) palengvinta difuzija- medžiagų transportavimas naudojant specialius transportavimo baltymus, kurių kiekvienas yra atsakingas už tam tikrų molekulių ar giminingų molekulių grupių (gliukozės, aminorūgščių, nukleotidų) judėjimą; 4) osmosas— vandens molekulių transportavimas (visuose biologines sistemas tirpiklis yra vanduo).
Būtinybė aktyvus transportas atsiranda, kai reikia užtikrinti molekulių pernešimą per membraną prieš elektrocheminį gradientą. Šį transportą atlieka specialūs baltymai-nešėjai, kurių veiklai reikia energijos sąnaudų. Energijos šaltinis yra ATP molekulės. Aktyvus transportas apima: 1) Na + /K + -siurblį (natrio-kalio pompą), 2) endocitozę, 3) egzocitozę.
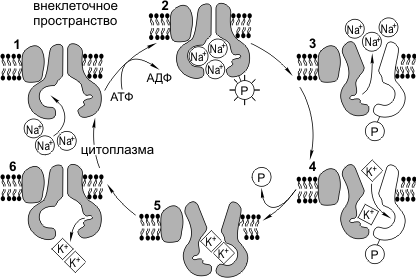
Dirbkite Na + /K + -siurblį. Dėl normalus funkcionavimas ląstelė turi palaikyti tam tikrą K + ir Na + jonų santykį citoplazmoje ir išorinėje aplinkoje. K + koncentracija ląstelės viduje turėtų būti žymiai didesnė nei už jos ribų, o Na + - atvirkščiai. Reikėtų pažymėti, kad Na + ir K + gali laisvai difuzuoti per membranos poras. Na+/K+ siurblys neutralizuoja šių jonų koncentracijų išlyginimą ir aktyviai pumpuoja Na+ iš ląstelės, o K+ – į ląstelę. Na + /K + -siurblys yra transmembraninis baltymas, galintis keisti konformacinius pokyčius, todėl gali prijungti ir K +, ir Na +. Na + /K + -siurblio veikimo ciklą galima suskirstyti į tokias fazes: 1) Na + pridėjimas su viduje membranos, 2) siurblio baltymo fosforilinimas, 3) Na + išsiskyrimas tarpląstelinėje erdvėje, 4) K + prijungimas su lauke membranos, 5) siurblio baltymo defosforilinimas, 6) K + išsiskyrimas viduląstelinėje erdvėje. Natrio-kalio siurblys sunaudoja beveik trečdalį visos ląstelės gyvybei būtinos energijos. Per vieną veikimo ciklą siurblys išpumpuoja 3Na + iš elemento ir pumpuoja 2K +.
Endocitozė- didelių dalelių ir makromolekulių absorbcijos procesas ląstelėje. Yra dviejų tipų endocitozė: 1) fagocitozė- didelių dalelių (ląstelių, ląstelių dalių, makromolekulių) surinkimas ir absorbcija ir 2) pinocitozė- skystos medžiagos (tirpalo, koloidinio tirpalo, suspensijos) surinkimas ir absorbcija. Fagocitozės reiškinį atrado I.I. Mechnikovas 1882. Endocitozės metu plazminė membrana suformuoja invaginaciją, jos kraštai susilieja, o struktūros, atskirtos nuo citoplazmos viena membrana, surišamos į citoplazmą. Daugelis pirmuonių ir kai kurie leukocitai gali fagocitozę. Pinocitozė stebima žarnyno epitelio ląstelėse, kraujo kapiliarų endotelyje.
Egzocitozė- endocitozei atvirkštinis procesas: išskyrimas įvairių medžiagų iš ląstelės. Egzocitozės metu pūslelės membrana susilieja su išorine citoplazmine membrana, pūslelės turinys pašalinamas už ląstelės ribų, o jos membrana įtraukiama į išorinę citoplazminę membraną. Tokiu būdu iš liaukų ląstelių vidinė sekrecija hormonai išsiskiria pirmuoniuose - nesuvirškintų likučių maistas.
Eiti į 5 paskaitos„Ląstelių teorija. Korinio ryšio organizavimo tipai »
Eiti į 7 paskaitos"Eukariotinė ląstelė: organelių struktūra ir funkcijos"
Abstraktus turinys
2.
Eukariotinės ląstelės citoplazma
4. Citoplazminės membranos
4.1. Chemija ir savybės ląstelių membranos
5. Plazminė membrana
5.1. Plazmos membranos augimas
6.1. Transporto funkcijos
6.2. Plazminės membranos receptorių funkcijos
6.3. Tarpląsteliniai kontaktai
6.4. Paprastas kontaktas. Užrakto tipo jungtis
6.6. tarpinis kontaktas
6.7. tarpo kontaktas
Bibliografija
1. Mokslininkų vaidmuo atrandant ląstelių membranas
Iki XVIII amžiaus pabaigos buvo manoma, kad ląstelės buvo atskirtos bendra siena ir negali būti atskirtos viena nuo kitos. Tolimesnėje šio mokslo raidoje svarbų vaidmenį suvaidino prancūzų botaniko Brissot de Mirbel (1776–1854) tyrimai, kuriuose pirmą kartą buvo išreikšta augalų audinių lyginamojo tyrimo idėja. Mirbelis pirmasis pasiūlė, kad gretimos ląstelės yra atskirtos bendra pertvara – membrana.
Vokiečių gamtininkas Rudolphis padarė išvadą, kad ląstelės yra izoliuotos, kad jos turi savo membranas, jas supa iš visų pusių. Bet kokios abejonės, kad ląsteles galima išskirti ir izoliuoti, galutinai išnyko po botaniko I. Moldengauerio (1766-1827) darbų, iš kurių paaiškėjo, kad ląstelė yra nepriklausoma struktūra, padengta membrana. Taigi, susiformavo nauja ląstelės idėja.
Aiškiausiai ją kiek vėliau suformulavo F. Meyenas. Jo apibrėžimas yra toks: „Augalų organizmo ląstelė yra erdvė, visiškai uždengta vegetatyvine membrana“. F. Meyenas aiškiai išdėstė šį požiūrį XIX amžiaus pradžioje. kuris pakeitė pradinį kameros kaip laiko vaizdą. Tačiau specifinis augalų anatomijos turinys vis tiek iš esmės mažai skyrėsi nuo senojo. Tyrėjai toliau sistemingai apibūdino struktūrą įvairūs kūnai augalus, taip pat detalizuoti ir patikslinti ištirtus. Šis laikotarpis pagrįstai laikomas medžiagos rinkimo, daugybės informacijos apie geriausią augalų struktūrą kaupimo laikotarpiu.
Taigi, pats ląstelių buvimo faktas įvairių audinių nekėlė jokių abejonių. Tačiau ar visą augalų organizmų struktūrų įvairovę išsekina tik ląstelės? Klausimas vis dar nebuvo iki galo aiškus. Be to, dauguma mokslininkų į šį klausimą atsakė neigiamai, kartu su ląstelėmis nurodydami augalų kraujagysles ir pluoštus kaip gana savotiškas struktūras. Apibendrindamas visas žinias šioje srityje, Meyenas tiesiogiai parašė, kad augalų audiniai susideda iš ląstelių, skaidulų ir kraujagyslių. Šių pastarųjų atsiradimas iš metamorfizuojančių ląstelių galutinai buvo nustatytas tik kiek vėliau – 30-aisiais. praėjusį šimtmetį.
2. Eukariotinės ląstelės citoplazma
Citoplazma yra ląstelės metabolinis darbo aparatas. Jame susitelkę bendrieji ir specialieji organeliai, pagrindiniai medžiagų apykaitos procesai vyksta citoplazmoje. Pastaraisiais dešimtmečiais ypač didelė pažanga padaryta atliekant atskirų organelių ir citoplazmos metabolinio aparato membraninių sistemų morfofunkcinę analizę. Tai leidžia įsivaizduoti medžiagų apykaitos aparato darbą kaip vientisą sistemą.
Šiuolaikiniuose ląstelės metabolinio aparato tyrimuose ypač ryški tendencija tirti funkcinę morfologinių struktūrų reikšmę – analizuoti funkciją per struktūrinę ir biocheminę šių struktūrų organizaciją. Svarbiausios sėkmės šiuo atžvilgiu buvo pasiektos tiriant pagrindinius baltymų sintezės organelius (ribosomas) ir energijos apykaitą užtikrinančias sistemas (mitochondrijas, chloroplastus ir prokariotinių ląstelių konjuguojančias membranas). Pagrindinis citoplazmos metabolinio aparato tyrimo apibendrinimas yra idėja, kad ji yra plastinė diferencijuota trifazė sistema, susidedanti iš pagrindinės citoplazmos, tarpląstelinių membranų ir membraninės sistemos turinio. Taigi, citoplazmos metabolinis aparatas, viena vertus, yra sudėtingas su specializuotomis struktūromis, atliekančiomis tam tikras funkcijas, ir, kita vertus, tai yra viena vientisa sistema, glaudžiai susijusi su paviršiniu ir branduoliniu aparatu. ląstelė.
Šis požiūris prasidėjo 1960 m. De Duve egzoplazminių membranų sistemų doktrinoje. Vėliau šis metodas buvo sukurtas tiek lizosomų teorijoje, tiek šiuolaikinės idėjos apie Golgi aparatą ir endoplazminį tinklą, taip pat apie dinaminę tarpląstelinių membranų sistemų sąveiką. Labai daug žadanti, mūsų nuomone, yra ir pastaruoju metu susiformavusi mintis apie pagrindinę citoplazmą – hialoplazmą. Priešingai nei ankstesnis požiūris į hialoplazmą kaip vienalytę gyvą koloidinę sistemą, pagrindinė citoplazma aiškinama kaip diferencijuota, nevienalytė citoplazmos fazė, galinti sudaryti sudėtingas struktūras. Su šia hialoplazmos savybe mes jau iš dalies susipažinome su paviršinio aparato submembraninės sistemos pavyzdžiu su dinamiškomis mechaninėmis-cheminėmis ir laikančiomis struktūromis.
Šiuo metu tampa aišku, kad tokios membranomis surištos hialoplazmos sistemos formuojasi ne tik šioje srityje plazminės membranos, bet ir visų citoplazmos membranų, o kartais net ir nemembraninių (ląstelių centro) struktūrų srityje. Šią citoplazmos dalį siūloma išskirti į specialų hialoplazmos posistemį, žymint ją terminu „citozolis“. Natūralu, kad tokio citozolio funkcijos ir morfobiocheminė struktūra grubaus ir lygaus endoplazminio tinklo (ER), Golgi aparato membranų, mitochondrijų, ląstelių centro, branduolio apvalkalo ir kt. srityje bus skirtingos ir specifinės. struktūros, aplink kurias susidaro atitinkamos citozolio dalys.
Atsižvelgiant į organelių, užtikrinančių tarpląstelinį metabolizmą ląstelių citoplazmoje, struktūrinę ir funkcinę įvairovę, citozolio sekcijų, atitinkančių kiekvieną konkretų organoidą ir praktiškai įtrauktų į jo sudėtį, funkcija neapsiribos tik atramine-susitraukiančia funkcija. . Struktūros ir fermentai, sudarantys citozolį, yra specifiniai kiekvienai organolei ir yra tiesiogiai susiję su šiame organoidu vykstančių procesų užtikrinimu. biocheminiai procesai.
Citoplazmos svarstymą patartina pradėti nuo ribosomų - baltymų sintezės molekulinių mašinų - struktūrinės ir biocheminės organizavimo problemos. Šiuo atveju ypač ryškiai išryškėja biocheminių procesų struktūrinės organizavimo reikšmė ląstelių sistemose, kurią tiriant buvo pasiekta didelės sėkmės.
Antroji ląstelių biologijos sritis, kurioje biocheminių procesų struktūrinė organizacija taip pat buvo labai išsamiai ištirta, yra vadinamųjų konjuguojančių membranų - ląstelių energijos apykaitos aparato - analizė. Į juos atsižvelgsime apibūdindami mitochondrijas, plastidus ir kitas citoplazmos struktūras, atsakingas už energijos apykaitą ląstelėje. Toliau aptariame diskrečios pagrindinių membranų analizės rezultatus.
Tikrasis ląstelės kūnas ir jo turinys yra atskirti nuo išorinės aplinkos arba nuo gretimų elementų daugialąsčiuose organizmuose plazmine membrana. Už plazminės membranos ribų, ekstraląstelinis, esantis ląstelių sienelės, arba sienelė, ypač gerai išreikšta augaluose ir prokariotiniuose organizmuose; gyvūnų ląstelėse jo nėra arba jis yra labai silpnai išreikštas. Visas ląstelės vidus, išskyrus branduolį, vadinamas citoplazma. Tai bendras terminas, pabrėžiantis ląstelės padalijimą į du pagrindinius komponentus: citoplazmą ir branduolį.
Eukariotinių ląstelių citoplazma nevienalytė savo struktūra ir sudėtimi ir apima hialoplazmą, membraną ir ne membraninius komponentus. Membraniniai komponentai yra vakuolinė sistema (endoplazminis tinklas, Golgi aparatas, lizosomos, augalų vakuolės), membranos organelės (mitochondrijos ir plastidės). Ne membraniniai komponentai apima centriolius, būdingus gyvūnų ląstelėms, ir ne membraninius makromolekulinius kompleksus ir struktūras, tokias kaip ribosomos, mikrovamzdeliai ir mikrofilamentai. Toks citoplazmos padalijimas į atskirus komponentus nereiškia jų struktūrinio ir funkcinio izoliavimo vienas nuo kito.
3. Hialoplazma – vidinė ląstelės aplinka
Sąvokos „hialoplazma“ (iš hialino – permatoma, skaidri), „bazinė plazma“ arba „citoplazminė matrica“ žymi labai svarbią ląstelės dalį, jos tikrąją, vidinę aplinką. IN elektroninis mikroskopas citoplazminė matrica atrodo kaip vienalytė arba smulkiagrūdė, mažo elektronų tankio medžiaga. Iš senų citoplazmos fizikinių ir cheminių savybių stebėjimų buvo gauta mintis, kad hialoplazma yra sudėtinga koloidinė sistema, apimanti įvairius biopolimerus: baltymus, nukleino rūgštis, polisacharidus ir kt. būklė iki gelio ir atgal. Pavyzdžiui, esant dideliam hidrostatiniam slėgiui citoplazma nekondensuoja, o grįžtamai suskystėja. Šis reiškinys paaiškinamas ryšių tarp molekulių sutrikimu hialoplazmos sudėtyje.
Elektroninių mikroskopinių ir fizikinių-cheminių tyrimų metodų dėka kilo idėjos apie pagrindinę plazmą kaip organizuotą, tvarkingą daugiakomponentę sistemą. Atskiros hialoplazmos zonos gali pakeisti savo agregacijos būseną - priklausomai nuo sąlygų ar funkcinės užduoties. Taigi, žinoma, kad atskiros tubulino baltymų molekulės gali būti išsklaidytos hialoplazmoje, tačiau tam tikrais momentais jos pradeda kauptis ir statyti ilgas vamzdines struktūras – mikrovamzdelius.
Šis mikrovamzdelių savaiminio susibūrimo procesas yra grįžtamasis: pasikeitus ląstelės gyvenimo sąlygoms (padidėjus slėgiui ar pakitus ląstelės membranų pralaidumui), mikrovamzdeliai suyra į monomerines tubulino molekules. Lygiai taip pat, atrodytų, bestruktūriškoje hialoplazmoje gali atsirasti ir suirti įvairūs fibriliniai, siūliniai baltymų molekulių kompleksai. Pagrindinė plazma gali dalyvauti formuojant ląstelių membranas, siūlus ir mikrofilamentus. Amebose išoriniai citoplazmos sluoksniai (ektoplazma) arba citoplazmos zona pseudopodijos srityje praktiškai susideda iš vienos hialoplazmos, ten retai randamos membranos, mitochondrijos ir kitos struktūros. Tačiau paveikus jonus, skatinančius ląstelių pabrinkimą, įvyksta greiti ir staigūs šių citoplazmos atkarpų persitvarkymai – jose atsiranda membraninių mažų vakuolių. Todėl pagrindinėje plazmoje turi būti visi statybiniai blokai (lipidai ir baltymai), kurie būtini membranoms susidaryti.
Fibrilinės citoplazminės struktūros taip pat gali atsirasti dėl specifinių agregacijos procesų hialoplazmoje. Jei iš ląstelių homogenatų nusodinami dideli branduoliai, membranos struktūros ir ribosomos, likusiame supernatante bus pagrindiniai cheminiai hialoplazmos komponentai. Mikromolekulių hialoplazmos sudėtis daugiausia apima įvairius globulinius baltymus ir citoplazminės matricos fermentus. Jie sudaro 20-25 proc. bendras turinys baltymai eukariotų ląstelėje.
IN bakterijų ląstelės, skurdžiai membraninių elementų, hialoplazminiai baltymai gali sudaryti apie 50 % visų baltymų. Svarbiausi matriciniai fermentai yra glikolizės fermentai, cukrų, azoto bazių, aminorūgščių, lipidų ir kitų svarbių junginių metabolizmo fermentai. Matricoje yra fermentų, skirtų aminorūgščių aktyvavimui baltymų sintezės metu, RNR pernešimui. Osmosines ir buferines ląstelės savybes daugiausia lemia hialoplazmos sudėtis ir struktūra.
Svarbiausias hialoplazmos vaidmuo yra tas, kad ši pusiau skysta terpė sujungia visas ląstelių struktūras ir užtikrina jų cheminę sąveiką tarpusavyje. Dauguma tarpląstelinio transportavimo procesų vyksta per hialoplazmą: aminorūgščių, riebalų rūgščių, nukleotidų ir cukrų pernešimas. Hialoplazmoje yra nuolatinis jonų srautas į plazmos membraną ir iš jos, į mitochondrijas, branduolį ir vakuoles. Hialoplazma yra pagrindinė ATP molekulių masės talpykla ir judėjimo zona. Jame nusėda atsarginiai produktai: glikogenas, riebalų lašai. Struktūriškai ir morfologiškai hialoplazma vis dar menkai suprantama.
4. Citoplazminės membranos
Bendras visų ląstelių membranų, išorinės plazmos membranos ir visų tarpląstelinių membranų bei membraninių organelių bruožas yra tas, kad jie yra ploni (6–10 nm) lipoproteinų sluoksniai (lipidai komplekse su baltymais), užsidarę ant savęs. Ląstelėje nėra atvirų membranų su laisvais galais. Ląstelių membranos visada apriboja ertmes ar sritis, uždarydamos jas iš visų pusių ir taip atskirdamos tokių ertmių turinį nuo aplinkos. Taigi plazminė membrana, dengianti visą ląstelės paviršių, turinti sudėtingą formą ir daugybę ataugų, niekur nenutrūksta, ji yra uždaryta. Jis atskiria citoplazmos turinį nuo ląstelę supančios aplinkos.
Intraląstelinės uždaros membranos sudaro pūsleles - sferinės arba suplotos formos vakuoles. Pastaruoju atveju susidaro plokšti membraniniai maišeliai arba talpyklos. Dažnai membranomis apribotos ertmės yra sudėtingos formos, primenančios kempinę ar tinklą, tačiau šiuo atveju tokios ertmės be pertrūkių ribojamos membrana. Tokiuose variantuose membranos taip pat atskiria dvi struktūrines citoplazmos fazes: hialoplazmą nuo vakuolių ir cisternų turinio. Mitochondrijų ir plastidų membranos turi tą pačią savybę: jos atskiria vidinį turinį nuo tarpmembraninių ertmių ir nuo hialoplazmos.
Branduolinė membrana taip pat gali būti pavaizduota kaip perforuotas tuščiaviduris sferinės formos dvigubas membraninis maišas. Branduolinio apvalkalo membranos riboja, atskiria karioplazmą ir chromosomas viena nuo kitos nuo perinuklearinės erdvės ertmės ir nuo hialoplazmos. Šias bendrąsias ląstelių membranų morfologines savybes lemia jų cheminė sudėtis, jų lipoproteinų prigimtis.
4.1. Ląstelių membranų chemija ir savybės
Pagrindinis cheminiai komponentai ląstelių membranos yra lipidai (40%) ir baltymai (60%), be to, daugelyje membranų yra angliavandenių. Lipidai yra didelė grupė organinės medžiagos blogai tirpsta vandenyje (hidrofobiškumas) ir tirpumas organiniuose tirpikliuose ir riebaluose (lipofiliškumas).
Lipidų sudėtis ląstelių membranose yra labai įvairi. Būdingi lipidų, randamų ląstelių membranose, atstovai yra fosfolipidai, sfingomielinai, o iš steroidinių lipidų – cholesterolis. Glicerolipidai yra trihidrolio alkoholio, glicerolio su dviem riebalų rūgštimis ir fosforo rūgšties esteriai, kurie savo ruožtu gali būti susieti su įvairiomis cheminėmis grupėmis (cholinu, serinu, inozitoliu, etanolaminu ir kt.).
Kita membraninių lipidų grupė yra sfingomielinai, kur glicerolis pakeičiamas aminoalkoholiu sfingozinu. Iš lipidų, susijusių su steroidais, daugiausia cholesterolio membranose. IN augalų ląstelės cholesterolio nerandama, jį pakeičia fitosteroliai. Bakterijos neturi sterolių. būdingas bruožas membraniniai lipidai yra jų molekulių padalijimas į dvi funkciškai skirtingas dalis: nepolines uodegas, kurios neturi krūvių ir susideda iš riebalų rūgščių, ir įkrautas polines galvutes. Poliarinės galvutės turi neigiamus krūvius arba gali būti neutralios (jei jos turi ir teigiamą, ir neigiamą krūvį).
Nustatyta, kad ląstelių membranos labai skiriasi viena nuo kitos pagal lipidų sudėtį. Taigi gyvūnų ląstelių plazminėse membranose yra daug cholesterolio (iki 30%), jose mažai lecitino, o mitochondrijų membranose, atvirkščiai, daug fosfolipidų, o cholesterolio – mažai. Iš viso lecitino lipidų kiekis endoplazminio tinklo frakcijose sudaro 60-70% visų fosfolipidų, o plazmos membranoje - 25-35%.
Apskritai plazminė membrana yra apibūdinama didelis kiekis cholesterolio ir sfingolipidų, taip pat sočiųjų ir mononesočiųjų riebalų rūgščių dominavimą fosfolipidų sudėtyje, o mitochondrijose, endoplazminiame tinkle ir daugelyje kitų citoplazminių membranų yra mažai cholesterolio ir sfingolipidų bei santykinai daug polinesočiųjų riebalų rūgščių. Matyt, dėl to citoplazminės membranos yra mažiau standžios nei plazminės membranos, jos yra labiau tirpios.
Skiriasi ir baltymų kiekis ląstelių membranose. Taigi mitochondrijose, palyginti su kitomis membranomis, yra daugiausia baltymų. Membranos labai skiriasi baltymų molekulių rinkiniu. Kai kurie iš jų yra susieti su lipidų galvutėmis joninėmis jungtimis, todėl lengvai išskiriami iš membranų druskos tirpalais. Kiti, sąveikaudami su Mg2+ arba Ca2+ jonais, sudaro druskų ryšius su lipidų polinėmis sritimis. Baltymai taip pat ekstrahuojami naudojant kompleksonus, tokius kaip versene (EDTA). Dauguma baltymų sąveikauja su lipidais membranose, pagrįstose hidrofobiniais ryšiais.
Tapo žinoma, kad daugelis membraninių baltymų, atrodo, susideda iš dviejų dalių: iš regionų, kuriuose gausu polinių (nešančių) aminorūgščių, ir regionų, praturtintų nepolinėmis aminorūgštimis (glicinu, alaninu, valionu, leucinu). Tokie baltymai lipidiniuose membranų sluoksniuose išsidėstę taip, kad jų nepolinės sritys būtų tarsi panardintos į „riebiąją“ membranos dalį, kurioje yra hidrofobinės lipidų sritys. Tokių baltymų poliarinė (hidrofilinė) dalis sąveikauja su lipidų galvutėmis ir pasukama į vandeninę fazę. Todėl tokie baltymai, susiję su lipidais hidrofobinėmis sąveikomis, praktiškai nėra ekstrahuojami vandeninėse fazėse. Juos galima išskirti tik suardant membraną, iš jos ištraukiant lipidus organiniais tirpikliais ar plovikliais.
Autorius biologinis vaidmuo Membraninius baltymus galima suskirstyti į tris grupes: fermentus, receptorių baltymus ir struktūrinius baltymus. Fermentų rinkinys membranose gali būti labai didelis ir įvairus (pavyzdžiui, kepenų ląstelių plazminėje membranoje buvo rasti mažiausiai 24 skirtingi fermentai).
Skirtingos membranos turi būdingą fermentų rinkinį. Kitaip tariant, cheminį membranų heterogeniškumą lemia ne tik lipidai, bet ir baltymai. Visoms ląstelių membranoms būdinga jų lipoproteinų struktūra; skirtumai slypi kiekybiniuose ir kokybiniuose lipidų ir baltymų sudėtis. Pavyzdžiui, lipidų ir baltymų sudėtis ir kiekis bus gana skirtingi endoplazminio tinklo ir vidinės mitochondrijų membranos membranose.
Membranų angliavandenių komponentą daugiausia sudaro glikoproteinai - baltymų molekulės kovalentiškai (skirtingai nuo nukleoproteinų ir lipoproteinų), susietos su angliavandenių grandinėmis.
4.2. Ląstelių membranų struktūra
Idėja, kad ląstelių membranos yra pagrįstos dvigubu lipidų sluoksniu, buvo gauta dar 1920-aisiais. 1925 m. buvo paskelbtas darbas, kuriame nurodyta, kad susidarė ištisinis plazminės membranos bilipidinis sluoksnis. Nepaisant daugybės klaidų, bendra idėja apie bilipidinio sluoksnio egzistavimą pasirodė teisinga. Paaiškėjo, kad jei lipidai ištraukiami iš eritrocitų membranos, o po to dedami ant vandens menisko paviršiaus, tuomet galima apskaičiuoti susidariusio vienarūšio lipidų sluoksnio užimamą plotą. Paaiškėjo, kad šis plotas yra du kartus didesnis už plotą, kurį užima eritrocitų paviršius, iš kurio buvo išskirti lipidai.
Mokslininkai teigia, kad eritrocitų membranose lipidai yra išsidėstę dviem sluoksniais. Be to, paaiškėjo, kad ląstelės paviršiaus įtempis yra žymiai mažesnis nei dirbtinio lipidinio sluoksnio paviršiaus įtempis. Be to, mokslininkai nustatė, kad kai į aliejų pridedama baltymų, paviršiaus įtempis sumažėja iki vertės, būdingos ląstelių paviršiaus įtempimui. Remiantis tuo, buvo padaryta išvada, kad membranoje yra baltymų.
Taigi, kilo hipotezė apie membranos struktūrą, pagal kurią tai yra trijų sluoksnių „sumuštinis“ (baltymas-lipidas-baltymas). Lipidų sluoksnis centre yra bimolekulinis sluoksnis, kuriame hidrofobiniai lipidų molekulių galai yra nukreipti vienas į kitą, o hidrofiliniai galai yra link baltymų sluoksnių, esančių abiejose lipidų sluoksnio pusėse. Ši hipotezė buvo tiesiogiai patvirtinta, kai atsirado galimybė naudoti elektroninę mikroskopiją ląstelių membranoms tirti. Jos dėka ant mielininio nervų apvalkalo membranų buvo galima ypač gerai matyti, kad membrana tikrai trisluoksnė: du tamsūs sluoksniai po 2,5 nm ir tarp jų platesnis šviesus sluoksnis.
Ląstelių membranų trijų sluoksnių sandara gali būti parodyta elektroniniu mikroskopu, naudojant užšaldymo-skilimo metodą, o tai ypač svarbu, nes tokiu atveju galima stebėti specifinius fermentus ar net fermentų sistemas. 60-ųjų pradžioje. Robertsonas suformulavo hipotezę apie vieną, vieningą „elementarią“ biologinę membraną. Klausimas apie kilmę, membranų susidarymą gyvoje ląstelėje dar nėra pakankamai ištirtas. Yra žinoma, kad membraninių elementų skaičius ir dydis ląstelėse gali didėti ir mažėti, t.y. galima kalbėti apie membranogenezę ir membranų destrukciją.
5. Plazminė membrana
Plazminė membrana, arba plazmolema, užima ypatingą vietą tarp įvairių ląstelių membranų. Tai paviršinė periferinė struktūra, ribojanti ląstelę iš išorės, kuri lemia tiesioginį jos ryšį su ekstraląsteline aplinka, taigi ir su visomis ląstelę veikiančiomis medžiagomis ir dirgikliais. Chemiškai plazmos membrana yra lipoproteinų kompleksas.
Pagrindiniai membranos organizavimo principai jau buvo aptarti ankstesniame skyriuje. Jis yra apie 10 nm storio ir yra storiausias iš ląstelių membranų. Pagrindiniai plazmos membranų komponentai yra lipidai (apie 40%), baltymai (daugiau nei 60%) ir angliavandeniai (apie 1%). Kaip jau minėta, plazminėje membranoje, palyginti su kitomis membranomis, yra daugiau cholesterolio, o jos fosfolipiduose vyrauja sočiosios riebalų rūgštys.
Baltymų, sudarančių plazmos membraną, sudėtis yra labai įvairi, tai lemia jos funkcinių apkrovų įvairovė. Tarp fermentų rasta 5 "- nukleotidazė, nuo Mg priklausoma ATPazė, aktyvuota Na ir K jonais, šarminė ir rūgštinė fosfatazė, adenilato ciklazė, RNazė, proteolitinis aktyvumas. Plazmos membranoje visiškai nėra kvėpavimo grandinės fermentų ir glikolitinių fermentų. Tačiau prokariotinėse ląstelėse elektronų transportavimo grandinės ir oksidacinio fosforilinimo elementai yra lokalizuoti būtent plazminės membranos sudėtyje.
Reikia pabrėžti, kad kai kurie baltymai, esantys plazmos membranos frakcijose, gali būti tiesiogiai nesusieti su bilipidiniu sluoksniu. Daugelis fermentų, kurie yra aktyvūs plazmos membranos frakcijose, gali būti lokalizuoti glikokalikse, glikoproteinų komplekse, susijusiame su plazmos membrana. Glikokalikso sudėtyje yra įvairių angliavandenių, kurie gali sudaryti apie 1% sausos membranos masės.
Angliavandeniai sudaro ilgas, išsišakojusias polisacharidų grandines, pritvirtintas prie baltymų sluoksnio, kuris dažnai apibrėžia ląstelės formą. Membraniniai maišeliai čia gali priglusti prie plazminės membranos iš vidaus; šiuo atveju šalia ląstelių paviršiaus yra trys membranos sluoksniai: pati plazminė membrana ir dvi pelikulinių alveolių membranos.
Bato blakstienose sruogelė formuoja sustorėjimus, išsidėsčiusius šešiakampių pavidalu, kurių centre iškyla blakstienos. Pelikulinių formacijų standumas taip pat gali būti siejamas su citoplazmos elementais, esančiais po plazmos membrana, su žievės sluoksniu. Taigi prie membranos esančiose eugleno pelikulo keterose, be membraninių vakuolių, randama lygiagrečių mikrovamzdelių ir mikrofilamentų pluoštų. Šis fibrilinis periferinis sutvirtinimas kartu su sulankstyta daugiasluoksne membranos periferija sukuria standžią granulės struktūrą.
5.1. Plazmos membranos augimas
Po ląstelių dalijimosi didėja augančių dukterinių ląstelių tūris, o dėl to padidėja ląstelės paviršius, padidėja plazminės membranos plotas. Tačiau tai ne vienintelis pavyzdys. staigus augimas tūris ir paviršius. Sparčiai augančių ląstelių paviršius javų kuokelių gijose per 1 valandą gali padidėti 65 kartus, t.y. kiekvieną minutę plazmolema paauga pradine verte. Toks didelis plazmos membranos augimo greitis gali būti paaiškintas tik greitu paruoštų membranos pirmtakų, lipidų ir baltymų įtraukimu į seną lipoproteinų sluoksnį. Panašiu būdu augimas yra visiškai įmanomas, nes yra žinoma, kad ląstelėje vyksta nuolatinis membranų persitvarkymas – kai kurie lipidai pakeičiami kitais, keičiasi jų kiekis, pasikeičia baltymų sudėtis ir tt Todėl membranos yra labai labilios. junginiai.
Tačiau buvo aprašytas ir kitas naujos plazminės membranos susidarymo atvejis. Miksomiceto Physarum polycephalum plazmodis buvo pradurtas stikliniu kapiliaru, o gautas plazmos cilindras, kuriame nebuvo plazminės membranos, buvo užfiksuotas nedelsiant, o po 1, 2, 3 ir 6 s nuo eksperimento pradžios, tada tiriamas. po elektroniniu mikroskopu. Šio grybelio citoplazmoje yra daug mažų pūslelių, kuriose yra gleivių, kurios iš išorės dengia visą plazmodiją. Buvo matyti, kad tokios pūslelės, išsibarsčiusios pagrindinėje plazmoje, susirinko neapsaugotame plazmos krašte, sudarydamos suplotas vakuoles, išsidėsčiusias lygiagrečiai išoriniam eksponuojamo gabalo paviršiui. Tokios plokščios vakuolės susijungė ir suformavo ištisinę membraną – naują plazmos membraną, kuri izoliuoja plazmos komponentus nuo išorinės aplinkos.
Matyt, toks plazminės membranos atsinaujinimo procesas vyksta ne tik jai stipriai pažeidžiant, bet ir nuolat, ląstelės gyvavimo metu. Taigi, dėl egzocitozės, vakuolinės membranos, susiliejančios su plazmine membrana, atrodo įterptos į ją, taip padidindamos bendrą ląstelės paviršiaus plotą. Sekrecijos proceso (vienos iš egzocitozės formų) stebėjimai paskatino daugelį tyrinėtojų manyti, kad ląstelėje vyksta nuolatiniai membraninių elementų srautai: iš plazminės membranos į citoplazmą (endocitozė) ir, atvirkščiai, membraninių struktūrų srautas. iš citoplazmos į ląstelės paviršių (egzocitozė) . Šiame membranų cikle pagrindinis vaidmuo skiriamas Golgi aparato membraninių vakuolių sistemai.
6. Plazminės membranos funkcijos
Plazminė membrana daro visa linija svarbiausios ląstelių funkcijos, kurių vadovaujančios yra citoplazmos medžiagų atribojimo nuo išorinės aplinkos funkcija ir įvairių medžiagų transportavimo tiek į ląstelę, tiek iš jos funkcija. Transporto funkcijos apima pasyvų vandens, jonų, mažos molekulinės masės medžiagų pernešimą ir aktyvų šių medžiagų transportavimą prieš koncentracijos gradientus, taip pat įvairias didelės molekulinės masės junginių ir kompleksų transportavimo formas (endocitozė).
Be to, plazmos membrana dalyvauja pašalinant iš ląstelių susidariusius produktus. Jis taip pat dalyvauja tarpląstelinio biopolimerų skilimo procesuose. Plazminės membranos paviršiuje yra įvairių receptorių struktūrų, kurios specifiškai sąveikauja su tarpląsteliniais faktoriais ir su kaimyninėmis ląstelėmis. Taigi, ląstelės membrana dalyvauja perduodant signalus į ląstelę. Plazminė membrana dalyvauja tarpląstelinėje sąveikoje daugialąsčiuose organizmuose. Atskiros plazminės membranos sekcijos specializuotose gyvūnų ląstelėse dalyvauja specialių ląstelių procesų, tokių kaip mikrovilliukai, blakstienėlės, receptorių ataugos ir kt., konstravimas. Galiausiai plazminė membrana atlieka svarbų vaidmenį ląstelių dalijimuisi.
Šis trumpas plazminės membranos funkcijų išvardijimas toli gražu nėra baigtas, tačiau jis parodo šios periferinės bet kurios ląstelės, nesvarbu, ar tai būtų bakterijų, ar eukariotinių organizmų, struktūros svarbą.
6.1. Transporto funkcijos
Plazminė membrana, kaip ir kitos ląstelių lipoproteininės membranos, yra pusiau laidi. Tai reiškia, kad skirtingos molekulės per ją praeina skirtingu greičiu, ir kuo didesnis molekulių dydis, tuo mažesnis jų prasiskverbimo per membraną greitis. Ši savybė apibrėžia plazmos membraną kaip osmosinį barjerą.
Vanduo ir jame ištirpusios dujos turi didžiausią prasiskverbimo gebą, jonai į membraną prasiskverbia daug lėčiau (apie 104 kartus lėčiau). Todėl, jei ląstelė, pavyzdžiui, eritrocitas, patalpinama į aplinką, kurioje druskų koncentracija mažesnė nei ląstelėje (hipotenzija), vanduo iš išorės pateks į ląstelę, o tai padidins jo tūris ir plazminės membranos plyšimas (hipotoninis „šokas“).
Priešingai, kai eritrocitas dedamas į didesnės koncentracijos druskos tirpalus nei ląstelėje, vanduo iš jo pateks į išorinė aplinka. Tuo pačiu metu ląstelė susiraukšlės, sumažės tūris. Toks pasyvus vandens transportavimas iš ląstelės ir į ląstelę vis dar vyksta nedideliu greičiu. Vadinasi, ląstelės membranoje, jos lipoproteininiame sluoksnyje, yra specialios poros, skirtos vandeniui ir jonams prasiskverbti.
6.2. Plazminės membranos receptorių funkcijos
Šios funkcijos yra susijusios su specialių struktūrų, skirtų specifiniam cheminių ar fizinių veiksnių atpažinimui, lokalizavimu plazmos membranoje. Ląstelės paviršiuje yra daug komponentų – receptorių, kurie lemia specifinių reakcijų su įvairiais agentais galimybę. Kaip tokie receptoriai ląstelės paviršiuje gali veikti membraniniai baltymai arba glikokalikso elementai (polisacharidai, glikoproteinai).
Manoma, kad tokios atskiroms medžiagoms jautrios vietos gali būti išsklaidytos po ląstelės paviršių arba surinktos nedidelėse zonose. Pavyzdžiui, bakterijų ar gyvūnų ląstelių paviršiuje yra ribotas vietų, prie kurių gali prisijungti viruso dalelės, skaičius. Įdomu tai, kad skirtingi virusai paprastai jungiasi prie skirtingų ląstelės periferijos dalių, o ta pati bakterinė ląstelė gali turėti keletą skirtingi tipai receptoriai.
Įvairių imunologinių metodų naudojimas parodė, kad antigeniškai lokalizuota ląstelės paviršiuje veikliosios medžiagos kurie specifiškai reaguoja su antikūnais arba imuninės ląstelės. Daugumoje šių paviršiaus antigenų yra angliavandenių grupių (glikolipidų, glikoproteinų). Antigeniniai bakterijų sienelės komponentai taip pat dažniausiai yra lipopolisacharidai ir glikolipidai. Ląstelės paviršiuje, dažnai tiesiai jos lipoproteininėje membranoje, yra įvairių receptorių, kurie fiziologiškai jungiasi. veikliosios medžiagos pvz., įvairūs hormonai, neurotransmiteriai ir kt.
skirtingos ląstelės gyvūnų organizmai gali turėti skirtingus receptorių rinkinius arba skirtingą to paties receptoriaus jautrumą. Daugelio ląstelių receptorių vaidmuo yra perduoti signalus iš paviršiaus į ląstelę. Šiuo metu yra gerai ištirta stimulų perdavimo į ląsteles sistema, naudojant tam tikrus hormonus, įskaitant peptidines grandines. Nustatyta, kad šie hormonai jungiasi prie specifinių receptorių ląstelės plazminės membranos paviršiuje.
Dėl ląstelių paviršiuje esančių receptorių rinkinių įvairovės ir specifiškumo susidaro labai sudėtinga žymenų sistema, leidžianti atskirti savo ląsteles (to paties individo ar tos pačios rūšies) nuo kitų. Panašios ląstelės sąveikauja viena su kita, todėl paviršiai sukimba (konjugacija pirmuoniuose ir bakterijose, susidaro audinių ląstelių kompleksai). Tokiu atveju ląstelės, kurios skiriasi determinantinių žymenų rinkiniu arba jų nesuvokia, yra pašalinamos iš tokios sąveikos arba sunaikinamos aukštesniuose gyvūnuose dėl imunologinių reakcijų.
Plazminė membrana yra susijusi su specifinių receptorių, kurie reaguoja į fiziniai veiksniai. Taigi plazmos membranoje arba jos dariniuose fotosintetinėse bakterijose ir melsvadumbliuose yra lokalizuoti receptorių baltymai (chlorofilai), sąveikaujantys su šviesos kvantais. Šviesai jautrių gyvūnų ląstelių plazminėje membranoje yra speciali fotoreceptorių baltymų (rodopsino) sistema, kurios pagalba šviesos signalas paverčiamas cheminiu, o tai savo ruožtu sukelia elektros generavimą. impulsas.
6.3. Tarpląsteliniai kontaktai
Plazminė membrana, kaip jau minėta, aktyviai dalyvauja tarpląsteliniuose kontaktuose, susijusiuose su konjugacija. vienaląsčiai organizmai. Daugialąsteliuose organizmuose dėl tarpląstelinės sąveikos susidaro sudėtingi ląstelių ansambliai, kurių priežiūra gali būti vykdoma. Skirtingi keliai. Gimdos, embriono audiniuose, ypač ant ankstyvosios stadijos vystymuisi, ląstelės lieka sujungtos viena su kita, nes jų paviršiai gali sulipti. Šią ląstelių sujungimo savybę galima nustatyti pagal jų paviršiaus savybes, kurios konkrečiai sąveikauja viena su kita.
Šių jungčių mechanizmas vis dar nepakankamai ištirtas, tačiau greičiausiai jį užtikrina lipoproteinų ir plazmos membranų glikokalikso sąveika. Esant tokiai tarpląstelinei embrioninių ląstelių sąveikai tarp plazmos membranų, visada lieka apie 20 nm pločio tarpas, užpildytas glikokaliksu. Audinių apdorojimas fermentais, pažeidžiančiais glikokalikso vientisumą (gleivių, kurios hidroliziškai veikia mucinus, mukopolisacharidus) arba pažeidžiantys plazmos membraną (proteazes), lemia ląstelių izoliaciją viena nuo kitos, jų disociaciją. Tačiau pašalinus disociacijos faktorių ląstelės gali vėl susiburti ir vėl agreguotis.
Taigi, galima atskirti skirtingų spalvų kempinių ląsteles - oranžinę ir geltoną. Paaiškėjo, kad šių ląstelių mišinyje susidaro dviejų tipų agregatai: susidedantys tik iš geltonų ir tik iš oranžinių ląstelių. Tokiu atveju mišrios ląstelių suspensijos savaime susitvarko, atkuria pirminę daugialąstę struktūrą. Panašūs rezultatai gauti su atskirtomis varliagyvių embrionų ląstelių suspensijomis; šiuo atveju vyksta selektyvus erdvinis ektodermos ląstelių atskyrimas nuo endodermos ir nuo mezenchimo.
Be to, jei audiniai naudojami reagregacijai vėlyvieji etapai vystosi embrionai, tada mėgintuvėlyje savarankiškai surenkami įvairūs audinių ir organų specifiškumo ląstelių ansambliai, formuojasi į inkstų kanalėlius panašūs epitelio agregatai ir kt. Daugialąsčių gyvūnų organizmų audinių ir organų jungtis tarp ląstelių gali sudaryti kompleksinis specialus struktūras, kurios vadino tinkamais tarpląsteliniais kontaktais. Šie struktūriniai tarpląsteliniai kontaktai ypač ryškūs viršutiniuose ribiniuose audiniuose, epitelyje.
Gali būti, kad pirminis tarpusavyje sujungtų ląstelių sluoksnio išskyrimas specialių struktūrinių tarpląstelinių kontaktų pagalba užtikrino audinių ir organų formavimąsi ir vystymąsi gyvūnų filogenezėje. Elektroninės mikroskopijos dėka sukaupta daug duomenų apie šių jungiamųjų darinių ultrastruktūrą. Deja, jų biocheminė sudėtis ir molekulinė struktūra dar nėra gerai suprantama.
Tiriant ląstelių ryšius epitelio sluoksniuose, galima rasti tokias struktūras, kurios jungia ląsteles viena su kita: paprastas kontaktas, „užrakto“ tipo jungtis, sandarus kontaktas, tarpinis kontaktas arba adhezijos zona, desmosominis kontaktas. , į plyšį panašus kontaktas.
6.4. Paprastas kontaktas. užrakto tipo jungtis
Paprastas kontaktas randamas tarp daugumos įvairios kilmės greta esančių ląstelių. Dauguma kontaktuojančių epitelio ląstelių paviršiaus taip pat yra sujungti paprastu kontaktu, kai kontaktuojančių ląstelių plazmines membranas skiria 15-20 nm tarpas. Kaip jau minėta, ši erdvė vaizduoja viršmembraninius ląstelių paviršių komponentus. Tarpo tarp ląstelių membranų plotis gali būti didesnis nei 20 nm, suformuojant pratęsimus, ertmes, bet ne mažesnis kaip 10 nm. Iš citoplazmos pusės specialių papildomų struktūrų nesiriboja su šia plazminės membranos zona.
„Užrakto“ tipo jungtis – tai vienos ląstelės plazminės membranos išsikišimas į kitos ląstelės invaginaciją (išsikišimą). Ant pjūvio tokio tipo jungtis primena dailidės siūlę. Tarpmembraninė erdvė ir citoplazma „pilių“ zonoje pasižymi tokiomis pat savybėmis kaip ir paprasto kontakto srityse.
6.5. Tvirtas uždarymo kontaktas
Tai zona, kurioje dviejų plazminių membranų išoriniai sluoksniai yra kuo arčiau. Šiame kontakte dažnai matoma trijų sluoksnių membrana: abiejų membranų du išoriniai osmofiliniai sluoksniai susilieja į bendrą 2–3 nm storio sluoksnį. Membranos susiliejimas nevyksta visoje sandaraus kontakto srityje, o yra taškinių jungčių serija. Iš citoplazmos pusės šioje zonoje dažnai randama daugybė maždaug 8 nm skersmens fibrilių, esančių lygiagrečiai plazmalemos paviršiui.
Šio tipo kontaktai buvo rasti tarp fibroblastų audinių kultūroje, tarp embriono epitelio ir mezenchiminių ląstelių.
6.6. Tarpinis kontaktas (arba klijavimo zona)
Šioje vietoje atstumas tarp membranų yra šiek tiek išplėstas (iki 25-30 nm) ir, skirtingai nei paprastas kontaktas, yra užpildytas tankiu turiniu, greičiausiai baltyminio pobūdžio.
6.7. tarpo kontaktas
Tai 0,5-3 μm ilgio sritis, kurioje plazmines membranas skiria 2-3 nm tarpas, kuris po osmacijos visai struktūrai suteikia septynių sluoksnių išvaizdą. Iš citoplazmos pusės specialių membraninių struktūrų nerasta. Šio tipo ryšys randamas visų tipų audiniuose. Atrodo, kad tarpo jungties funkcinis vaidmuo yra jonų ir molekulių perkėlimas iš ląstelės į ląstelę. Pavyzdžiui, širdies raumenyje veikimo potencialo perdavimas iš ląstelės į ląstelę vyksta per kontakto tipą, kai jonai gali laisvai pereiti per tarpląstelines jungtis. Šio joninio ryšio tarp ląstelių palaikymas priklauso nuo energijos, gaunamos oksidacinio fosforilinimo metu.
Bibliografija
1. Vermelis E. M. Ląstelės doktrinos istorija. M.: Nauka, 1970. 259 p.
2. Guliajevas G. V., Malčenka V. V. Genetikos, citologijos, veisimo, sėklininkystės ir sėklininkystės terminų žodynas. M.: Rosselkhozizdat, 1983. 240 p.
3. De Duve K. Kelionė į gyvos ląstelės pasaulį. M.: Mir, 1987. 253 p.
4. Zavarzin A. A., Kharazova A. D. Bendrosios citologijos pagrindai: vadovėlis. L .: Leidykla Leningradas. un-ta, 1982. 240 p.
5. Nuo molekulių iki žmogaus. / Pagal. red. Naumova N.P.M.: Švietimas, 1973. 480 p.
6. Pausheva Z. P. Seminaras apie augalų citologiją. M.: Agropromizdat, 1988. 271 p.
7. Troshin A. S., Braun A. D., Vakhtin Yu. B., Zhilkin L. N., Sukhanova K. M. Citologija. Maskva: Švietimas, 1970. 304 p.
8. Chencov Yu. S. Bendroji citologija. M.: MGU, 1978. 344 p.
Abstraktus pliusas
plazmolema gyvūnų ląstelių sienelę sudaro membrana, iš išorės padengta 10-20 nm storio glikokalikso sluoksniu. plazmolema atlieka atribojimo, barjero, transportavimo ir receptorių funkcijas. Dėl selektyvaus pralaidumo savybės plazmalemma reguliuoja ląstelės vidinės aplinkos cheminę sudėtį. Plazmalemoje yra receptorių molekulių, kurios selektyviai atpažįsta tam tikras biologiškai aktyvias medžiagas (hormonus). Sluoksniuose ir sluoksniuose kaimyninės ląstelės išsaugomos dėl skirtingos rūšies kontaktai, kuriuos vaizduoja specialią struktūrą turinčios plazmalemos dalys. Iš vidaus žievės (žievės) sluoksnis ribojasi su membrana citoplazma 0,1-0,5 µm storio.
Citoplazma. Citoplazmoje yra keletas formalizuotų struktūrų, kurios turi reguliarius struktūros ir elgesio ypatumus skirtingais ląstelės gyvenimo laikotarpiais. Kiekviena iš šių struktūrų atlieka tam tikrą funkciją. Iš to kilo jų palyginimas su viso organizmo organais, dėl kurių jie gavo pavadinimą organelės, arba organelės. Citoplazmoje nusėda įvairios medžiagos – inkliuzai (glikogenas, riebalų lašai, pigmentai). Citoplazma yra persmelkta membranų endoplazminis Tinklelis.
Endoplazminis tinklas (EMF). Endoplazminis tinklas yra šakotas kanalų ir ertmių tinklas ląstelės citoplazmoje, sudarytas iš membranų. Kanalų membranose yra daug fermentų, užtikrinančių gyvybinę ląstelės veiklą. Yra 2 tipai membranos EMF- lygus ir grubus. Ant membranų lygus endoplazminis tinklas yra fermentų sistemos, susijusios su riebalais ir angliavandenių apykaitą. pagrindinė funkcija šiurkštus endoplazminis tinklas- baltymų sintezė, kuri atliekama ribosomose, pritvirtintose prie membranų. Endoplazminis Tinklelis– tai bendra tarpląstelinė kraujotakos sistema, kurios kanalais ląstelės pernešamos medžiagos ir iš ląstelės į ląstelę.
Ribosomos atlieka baltymų sintezės funkciją. Ribosomos yra sferinės 15-35 nm skersmens dalelės, susidedančios iš 2 nevienodo dydžio subvienetų ir turinčios maždaug vienodą kiekį mRNR baltymų. Ribosomos citoplazmoje yra arba yra pritvirtintos prie išorinio endoplazminio tinklo membranų paviršiaus. Priklausomai nuo sintezuojamo baltymo tipo, ribosomos gali būti sujungtos į kompleksus - poliribosomos. Ribosomos yra visų tipų ląstelėse.
Golgi kompleksas. Pagrindinis konstrukcinis elementas Golgi kompleksas yra lygi membrana, kuri sudaro plokščių cisternų arba didelių vakuolių arba mažų pūslelių paketus. Golgi komplekso cisternos yra sujungtos su endoplazminio tinklo kanalais. Baltymai, polisacharidai, riebalai, susintetinti ant endoplazminio tinklo membranų, yra transportuojami į kompleksą, kondensuojami jo struktūrų viduje ir „supakuojami“ į paslaptį, paruoštą išsiskyrimui, arba panaudojami pačioje ląstelėje per savo gyvavimo laikotarpį.
Mitochondrijos. Bendras mitochondrijų pasiskirstymas gyvūne ir flora atkreipti dėmesį į svarbų vaidmenį mitochondrijosžaisti narve. Mitochondrijos turi sferinio, ovalo ir cilindro formos kūnus, gali būti siūliniai. Mitochondrijų dydis yra 0,2–1 µm skersmens, iki 5–7 µm ilgio. Gijinių formų ilgis siekia 15-20 mikronų. Skirtingų audinių ląstelėse mitochondrijų skaičius nėra vienodas, daugiau jų yra ten, kur intensyvūs sintetiniai procesai (kepenys) arba didelės energijos sąnaudos. Mitochondrijų siena susideda iš 2 membranų – išorinės ir vidinės. Išorinė membrana yra lygi, o pertvaros - keteros, arba cristae, nukrypsta nuo organoido vidaus. Krūtų membranose yra daug fermentų, dalyvaujančių energijos apykaitoje. Pagrindinė mitochondrijų funkcija - ATP sintezė.
Lizosomos- maži ovalūs kūnai, kurių skersmuo apie 0,4 mikrono, apsupti viena trijų sluoksnių membrana. Lizosomose yra apie 30 fermentų, galinčių skaidyti baltymus, nukleino rūgštis, polisacharidus, lipidus ir kitas medžiagas. Medžiagų skaidymas fermentais vadinamas lizė, todėl organoidas pavadintas lizosoma. Manoma, kad lizosomos susidaro iš Golgi komplekso struktūrų arba tiesiogiai iš endoplazminio tinklo. Lizosomų funkcijos : tarpląstelinis maistinių medžiagų virškinimas, pačios ląstelės struktūros sunaikinimas jai žūstant embriono vystymosi metu, kai embriono audiniai pakeičiami nuolatiniais ir daugeliu kitų atvejų.
Centrioliai. Ląstelės centras susideda iš 2 labai mažų cilindrinių kūnų, išdėstytų stačiu kampu vienas kito atžvilgiu. Šie kūnai vadinami centrioliai. Centrio sienelę sudaro 9 poros mikrotubulių. Centrioliai gali savaime susiburti ir yra savaime besidauginančios citoplazmos organelės. Centrioliai atlieka svarbų vaidmenį ląstelių dalijimuisi: jie pradeda augti mikrotubuliams, kurie sudaro dalijimosi veleną.
Šerdis.Šerdis yra svarbiausia komponentas ląstelės. Jame yra DNR molekulių, todėl atlieka dvi pagrindines funkcijas: 1) genetinės informacijos saugojimą ir atkūrimą, 2) ląstelėje vykstančių medžiagų apykaitos procesų reguliavimą. Ląstelė, kuri prarado šerdis, negali egzistuoti. Branduolys taip pat nepajėgus savarankiškai egzistuoti. Dauguma ląstelių turi vieną branduolį, tačiau vienoje ląstelėje galima stebėti 2-3 branduolius, pavyzdžiui, kepenų ląstelėse. Žinomos daugiabranduolės ląstelės, kurių branduolių skaičius per kelias dešimtis. Branduolių forma priklauso nuo ląstelės formos. Branduoliai sferiniai, daugiaskilčiai. Branduolys yra apsuptas membranos, susidedančios iš dviejų membranų, turinčių įprastą trijų sluoksnių struktūrą. Išorinė branduolio membrana padengta ribosomomis, vidinė membrana lygi. Pagrindinis vaidmuo gyvybinėje branduolio veikloje tenka metabolizmui tarp branduolio ir citoplazmos. Branduolio turinys apima branduolio sultis arba karioplazmą, chromatiną ir branduolį. Branduolinių sulčių sudėtis apima įvairius baltymus, įskaitant daugumą branduolinių fermentų, laisvųjų nukleotidų, aminorūgščių, branduolio produktų ir chromatino, judančių iš branduolio į citoplazmą. Chromatinas yra DNR, baltymų ir yra spiralizuotos ir sutankintos chromosomų dalys. branduolys yra tankus suapvalintas kūnas, esantis branduolio sultyse. Branduolių skaičius svyruoja nuo 1 iki 5-7 ar daugiau. Branduoliai yra tik nesidalijančiuose branduoliuose, mitozės metu jie išnyksta, o pasibaigus dalijimui vėl susidaro. Branduolys nėra savarankiška ląstelės organelė; jis neturi membranos ir yra suformuotas aplink chromosomos sritį, kurioje užkoduota rRNR struktūra. Branduolėje susidaro ribosomos, kurios vėliau pereina į citoplazmą. chromatinas vadinami gabalėliais, granulėmis ir į tinklą panašiomis branduolio struktūromis, intensyviai nudažytomis kai kuriais dažais ir savo forma skiriasi nuo branduolio.
2)1. Ląstelių teorija
Ląstelių teorija yra apibendrinta idėja apie ląstelių, kaip gyvų vienetų, struktūrą, jų dauginimąsi ir vaidmenį formuojant daugialąsčius organizmus.
Tam tikrų nuostatų atsiradimas ir formulavimas ląstelių teorija prieš tai buvo gana ilgas stebėjimų apie įvairių vienaląsčių ir daugialąsčių augalų ir gyvūnų organizmų sandaros kaupimo laikotarpis. Šis laikotarpis buvo susijęs su įvairių optinių tyrimų metodų taikymo plėtra ir tobulėjimu.
Robertas Hukas pirmasis, naudodamas didinamuosius lęšius, pastebėjo kamštienos audinio padalijimą į „ląsteles“ arba „ląsteles“. Jo aprašymai paskatino sistemingus augalų anatomijos tyrimus, kurie patvirtino Roberto Hooke'o pastebėjimus ir parodė, kad įvairios augalų dalys yra sudarytos iš glaudžiai išdėstytų „pūslelių“ arba „maišelių“. Vėliau A. Leeuwenhoekas atrado vienaląsčių organizmų pasaulį ir pirmą kartą pamatė gyvūnų ląsteles. Vėliau gyvūnų ląsteles aprašė F. Fontana; tačiau šie ir kiti daugybė tyrimų tuo metu nepadėjo suprasti ląstelių struktūros universalumo, aiškių idėjų apie tai, kas yra ląstelė. Mikroanatomijos ir ląstelių tyrimo pažanga siejama su mikroskopijos raida XIX a. Iki to laiko idėjos apie ląstelių sandarą pasikeitė: ląstelės organizacijoje pagrindiniu dalyku imta laikyti ne ląstelės sienelę, o tikrąjį jos turinį, protoplazmą. Protoplazmoje buvo aptiktas nuolatinis ląstelės komponentas – branduolys. Visi šie daugybė pastebėjimų leido T. Schwann 1838 m. padaryti nemažai apibendrinimų. Jis parodė, kad augalų ir gyvūnų ląstelės iš esmės yra panašios viena į kitą. „T. Schwanno nuopelnas buvo ne tai, kad jis atrado ląsteles kaip tokias, o tai, kad jis išmokė tyrinėtojus suprasti jų prasmę. Šios idėjos buvo toliau plėtojamos R. Virchow darbuose. Ląstelių teorijos sukūrimas tapo svarbiu biologijos įvykiu, vienu iš lemiamų visos gyvos gamtos vienybės įrodymų. Ląstelių teorija turėjo didelę įtaką biologijos raidai, buvo pagrindinis pagrindas plėtoti tokias disciplinas kaip embriologija, histologija ir fiziologija. Ji suteikė pagrindus suprasti gyvybę, paaiškinti organizmų santykius, suprasti individualią raidą.
Pagrindinės ląstelių teorijos nuostatos savo reikšmę išlaikė iki šių dienų, nors jau daugiau nei šimtą penkiasdešimt metų gauta naujos informacijos apie ląstelių sandarą, gyvybinę veiklą ir vystymąsi. Šiuo metu ląstelių teorija teigia:
1) Ląstelė yra elementarus gyvųjų vienetas: - už ląstelės ribų nėra gyvybės.
2) Ląstelė yra viena sistema, susidedanti iš daugelio natūraliai tarpusavyje susijusių elementų, atstovaujančių tam tikrą vientisą darinį, susidedantį iš konjuguotų funkcinių vienetų – organelių arba organelių.
3) Ląstelės yra panašios – homologiškos – struktūra ir pagrindinėmis savybėmis.
4) Ląstelių skaičius didėja dalijant pirminę ląstelę, padvigubėjus jos genetinei medžiagai: ląstelė po ląstelės.
5) Daugialąstis organizmas yra nauja sistema, sudėtingas daugelio ląstelių ansamblis, susijungęs ir integruotas į audinių ir organų sistemas, sujungtas viena su kita cheminių, humoralinių ir nervinių faktorių pagalba.
6) Daugialąsčių organizmų ląstelės yra totipotentinės, t.y. turi visų tam tikro organizmo ląstelių genetines galias, yra lygiavertės genetinės informacijos požiūriu, tačiau skiriasi viena nuo kitos skirtinga įvairių genų raiška, o tai lemia jų morfologinę ir funkcinę įvairovę – diferenciaciją.
Ląstelės, kaip savarankiško gyvybiškai svarbaus vieneto, idėja buvo pateikta T. Schwann darbuose. R. Virchow taip pat tikėjo, kad kiekviena ląstelė neša visas gyvybės savybes: „Ląstelė yra paskutinis morfologinis visų gyvų kūnų elementas, ir mes neturime teisės ieškoti tikros gyvybės veiklos už jos ribų“.
Šiuolaikinis mokslas visiškai įrodė šį teiginį. Populiariojoje literatūroje ląstelė dažnai vadinama „gyvybės atomu“, „gyvybės kvantu“, taip pabrėžiant, kad ląstelė yra mažiausias gyvos materijos vienetas, už kurio ribų gyvybės nėra.
Tokia bendra ląstelės charakteristika savo ruožtu turėtų būti pagrįsta gyvojo apibrėžimu – kas yra gyva, kas yra gyvybė. Labai sunku pateikti galutinį gyvenimo, gyvenimo apibrėžimą.
M.V. Volkenšteinas pateikia tokį gyvybės apibrėžimą: „gyvi organizmai yra atviros, savireguliuojančios ir savaime besidauginančios sistemos, kurių svarbiausios funkcionuojančios medžiagos yra baltymai ir nukleorūgštys“. Gyvoms būtybėms būdinga daugybė kumuliacinių savybių, tokių kaip gebėjimas daugintis, energijos naudojimas ir transformacija, medžiagų apykaita, jautrumas ir kintamumas. Ir tokį šių ženklų derinį galima rasti ant ląstelių lygis. Nėra mažesnio gyvybės vieneto už ląstelę. Mes galime išskirti atskirus komponentus ar net molekules iš ląstelės ir patikrinti, ar daugelis jų turi specifinių funkcinių savybių. Taigi, izoliuotos aktomiozino fibrilės gali susitraukti, reaguodamos į ATP pridėjimą; už ląstelės ribų puikiai „veikia“ daugelis fermentų, dalyvaujančių sudėtingų bioorganinių molekulių sintezėje ar skaidyme; išskirtos ribosomos, esant reikalingiems faktoriams, gali sintetinti baltymus, buvo sukurtos neląstelinės sistemos fermentinei nukleorūgščių sintezei ir kt. Ar visi šie ląstelių komponentai, struktūros, fermentai, molekulės gali būti laikomi gyvais? Ar aktomiozino kompleksą galima laikyti gyvu? Atrodo, kad taip nėra, jau vien todėl, kad jis turi tik dalį gyvųjų savybių rinkinio. Tas pats pasakytina ir apie kitus pavyzdžius. Tik ląstelė kaip tokia yra mažiausias vienetas, turintis visas savybes, atitinkančias „gyvo“ apibrėžimą.
3) Ląstelių paviršiaus aparato (PAC) pagrindas yra lauke ląstelių membrana arba plazmolema. Be plazmalemos, PAC turi epimembraninį kompleksą, o eukariotai taip pat turi submembraninį kompleksą. Pagrindiniai biocheminiai plazmalemos komponentai (iš graikų plazmos – formavimasis ir lemma – apvalkalas, pluta) yra lipidai ir baltymai. Jų kiekybinis santykis daugumoje eukariotų yra 1:1, o prokariotuose plazmos lemoje vyrauja baltymai. Nedidelis angliavandenių kiekis randamas išorinėje ląstelės membranoje ir galima rasti į riebalus panašių junginių (žinduoliams – cholesterolio, riebaluose tirpių vitaminų). 1925 metais E. Gorteris ir F. Grendelis (Olandija) pasiūlė, kad membranos pagrindas yra dvigubas lipidų sluoksnis – bilipidinis sluoksnis. 1935 metais J. Danieli ir G. Dawson pasiūlė pirmąjį erdvinį membranos organizavimo modelį, pavadintą „sumuštinio“ arba „sumuštinio“ modeliu. Jų nuomone, membranos pagrindas yra bilipidinis sluoksnis, o abu sluoksnio paviršiai padengti ištisiniais baltymų sluoksniais. Tolesnis ląstelių membranų, įskaitant plazmalemą, tyrimas parodė, kad beveik visais atvejais jų struktūra yra panaši. 1972 metais S. Singer ir G. Nicholson (JAV) suformulavo koncepciją skysta-mozaikinė struktūra ląstelių membranos (pav.). Pagal šį modelį membranos yra pagrįstos lipidų sluoksnis, tačiau baltymai jame išsidėstę atskirose molekulėse ir kompleksuose, t.y. mozaika (iš prancūzų k. mosaique – mozaika; vaizdas, sudarytas iš atskirų gabalėlių). Visų pirma integralinių (iš lot. integrer – visa) baltymų molekulės gali kirsti bilipidinį sluoksnį, pusiau vientisų – iš dalies panirti į jį, o periferinių (iš graikų kalbos periferija – apskritimas) – išsidėsčiusios jo paviršiuje (pav.). Šiuolaikinė molekulinė biologija patvirtino skysčio mozaikos modelio pagrįstumą, nors buvo atrasti ir kiti ląstelių membranų variantai. Visų pirma, archebakterijose membranos pagrindas yra vienasluoksnis sudėtingų lipidų sluoksnis, o kai kuriose bakterijose citoplazmoje yra membraninių pūslelių, kurių sieneles vaizduoja baltyminis monosluoksnis. Supramembraninis kompleksas ląstelių paviršinis aparatas pasižymi struktūrų įvairove (pav.). Prokariotuose epimembraninį kompleksą daugeliu atvejų sudaro įvairaus storio ląstelės sienelė, kurios pagrindas yra kompleksinis glikoproteinas mureinas (archebakterijose pseudomureinas). Daugelyje eubakterijų išorinę epimembraninio komplekso dalį sudaro kita membrana, kurioje yra daug lipopolisacharidų. Eukariotuose universalus supramembraninio komplekso komponentas yra angliavandeniai – plazmalemos glikolipidų ir glikoproteinų komponentai. Dėl šios priežasties jis iš pradžių buvo vadinamas glikokaliksu (iš graikų glycos - saldus, angliavandenių ir lotynų callum - stora oda, apvalkalas). Be angliavandenių, periferiniai baltymai, esantys virš bilipidinio sluoksnio, yra įtraukti į glikokaliksą. Sudėtingesni epimembraninio komplekso variantai randami augaluose (ląstelių sienelė iš celiuliozės), grybuose ir nariuotakojų (išorinė danga iš chitino). pomembraninė(iš lot. sub - under) kompleksas būdingas tik eukariotų ląstelėms. Jį sudaro įvairios baltyminės gijinės struktūros: plonos fibrilės (iš lotynų kalbos fibrilė – pluoštas, siūlas), mikrofibrilės (iš graikų micros – mažos), skeleto (iš graikiško skeleto – džiovinti) fibrilių ir mikrovamzdelių. Jie yra sujungti vienas su kitu baltymais ir sudaro ląstelės raumenų ir kaulų aparatą. Submembraninis kompleksas sąveikauja su plazmos membranos baltymais, kurie, savo ruožtu, yra susiję su supramembraniniu kompleksu. Dėl to PAH yra struktūriškai vientisa sistema. Tai leidžia atlikti svarbias ląstelei funkcijas: izoliaciją, transportavimą, katalizavimą, receptorių signalizavimą ir kontaktą.
4) Membranose taip pat yra glikolipidų ir cholesterolio. Glikolipidai yra lipidai, prie kurių prisijungę angliavandeniai. Kaip ir fosfolipidai, glikolipidai Yra poliarinės galvos ir nepoliarinės uodegos. Cholesterolis yra artimas lipidams; jo molekulė turi ir polinę dalį.
1. Ląstelių teorijos kūrėjas yra:
R. Virchovas
Į IR. Vernadskis
T.Schwann
2. Ląstelių teorija buvo sukurta:
1839 m
3. Eukariotinių ląstelių citoplazmos membraninių organelių grupė:
Ribosomos, mitochondrijos, plastidai, lizosomos
sferosomos, vakuolės, plastidai
Mikrovamzdeliai, mikrofilamentai, ribosomos, centrosomos
EPS, ribosomos, lizosomos, Golgi aparatas
mitochondrijos, ER, Golgi aparatas
4. Bendrosios paskirties organelės:
miofibrilės
ribosomos, mitochondrijos
EPS, Golgi aparatai
žvyneliai, ribosomos
Vėliavos, miofibrilės
5. Tarpfaziniame branduolyje yra:
Ribosomos
branduoliai
kariolema
branduolinės sultys
chromatinas
6. Chromosomų tipai priklausomai nuo centromeros vietos:
subcentrinis
Telocentrinis
akrocentrinis
metacentrinis
nelygus petys
7. Kiekviena mitozės metafazės chromosoma susideda iš:
DNR, histono baltymai, nehistoniniai baltymai
2x mikrofibrilės
2x chromatidės
4x semichromatids
chromonema
8. Somatinės ląstelės turi chromosomų rinkinį:
poliploidas
diploidinis
Haploidas
tetraploidas
9. Sintezė vyksta sklandžiai EPS:
angliavandenių
10. Chromoplastuose yra pigmento:
Hemoglobinas
likopenas
karotino
ksantofilas
Chlorofilas
11. Ribosomų funkcijos:
hormonai
Angliavandenių sintezė
Nukleino rūgštys
baltymų sintezė
Riebalų sintezė
12. Įtraukimo grupės:
vitaminas
trofinis
mineralinis
sekrecinis, šalinamasis
pigmentas
13. Lytinės ląstelės turi chromosomų rinkinį:
diploidas
poliploidas
haploidinis
14. Eukariotai apima:
cianobakterijos
gyvūnai
kerpės
augalai
15. Eukariotinių ląstelių citoplazmos nemembraninių organelių grupė:
Ribosomos, vakuolės, mikrovamzdeliai
ribosomos, centrosoma
ER, mitochondrijos, ribosomos
mikrovamzdeliai, mikrofibrilės
Mikrofilamentai, plastidai, lizosomos
16. Hialoplazmos funkcijos:
dalyvavimas medžiagų apykaitoje
Sintetinis
ribojantis
homeostatinis
transportas
17. Branduolių funkcijos:
Baltymų sintezė
ribosomų subvienetų susidarymas
transporto RNR sintezė
Mikrofibrilių susidarymas
ribosomų RNR sintezė
18.EPS funkcijos:
Receptorius
transportas
atribojant
Apsauginis
sintetinis
19. Lizosomų funkcijos:
Angliavandenių sintezė ir kaupimas
laikinų embrionų ir lervų organų sunaikinimas
Baltymų, riebalų, angliavandenių sintezė
autolizė, pašalinių dalelių sunaikinimas
tarpląstelinis virškinimas
20. Vakuolės yra:
Gyvūnų ląstelių nemembraninės organelės
Augalų ląstelių membraninės organelės
Bakterijų ląstelių membraninės organelės
Vienaląsčių gyvūnų membraninės organelės
Augalų ląstelių nemembraninės organelės
21. Vakuolių funkcijos:
pašalinti vandens perteklių
palaikyti ląstelės osmosinį slėgį
atlikti tarpląstelinį virškinimą
palaikyti turgorinį slėgį augalų ląstelėse
Vykdyti riebalų sintezę ir kaupimąsi augalų ląstelėse
22. Chloroplastuose yra pigmento:
Ksantofilas
chlorofilas
23. Ribosomos apima:
T-RNR, magnis
Baltymai, DNR, magnis
Tik voverės
Baltymai, i-RNR
baltymai, r-RNR, magnis
24. Mikrofilamentų sudėtis apima baltymus:
25. Mikrotubuliuose yra baltymų:
tubulinas
26. Mikrofibrilėse yra baltymų:
Kolagenas
keratinas
27. Branduolys būdingas ląstelėms:
Visi gyvi organizmai
eukariotas
augalai
prokariotas