AUGALŲ LĄSTELĖS STRUKTŪRA
Augalo ląstelė susideda iš daugiau ar mažiau standžių ląstelių sienelės ir protoplastas. Ląstelės sienelė yra ląstelės sienelė ir citoplazminė membrana. Terminas protoplastas kilęs iš žodžio protoplazma, kuris ilgam laikui naudojamas reikšti visus gyvus dalykus. Protoplastas yra atskiros ląstelės protoplazma.
Protoplastas susideda iš citoplazmos ir branduolio. Citoplazmoje yra organelių (ribosomų, mikrotubulių, plastidžių, mitochondrijų) ir membraninių sistemų (endoplazminis tinklas, diktiosomos). Citoplazma taip pat apima citoplazminę matricą (pagrindinę medžiagą), į kurią panardinami organeliai ir membranų sistemos. Citoplazma yra atskirta nuo ląstelės sienelės plazmos membrana, kuri yra elementari membrana. Skirtingai nuo daugelio gyvūnų ląstelių, augalų ląstelėse yra viena ar daugiau vakuolių. Tai pūslelės, užpildytos skysčiu ir apsuptos elementaria membrana (tonoplastas).
Gyvoje augalo ląstelėje pagrindinė medžiaga yra nuolatiniame judėjime. Organelės dalyvauja judėjime, vadinamame citoplazmos srautu arba cikloze. Ciklozė palengvina medžiagų judėjimą ląstelėje ir jų mainus tarp ląstelės ir aplinkos.
Plazmos membrana. Tai dvisluoksnė fosfolipidinė struktūra. Augalų ląstelėms būdingos plazminės membranos invaginacijos.
Plazminė membrana atlieka šias funkcijas:
Dalyvauja medžiagų mainuose tarp ląstelės ir aplinkos;
Koordinuoja ląstelės sienelės celiuliozės mikrofibrilių sintezę ir surinkimą;
Perduoda hormoninius ir išorinius signalus, kurie kontroliuoja ląstelių augimą ir diferenciaciją.
Šerdis. Tai ryškiausia eukariotinės ląstelės citoplazmos struktūra. Branduolys atlieka dvi svarbias funkcijas:
Kontroliuoja gyvybinę ląstelės veiklą, nustatydama, kurie baltymai ir kokiu laiku turi būti sintetinami;
Saugo genetinę informaciją ir perduoda ją dukterinėms ląstelėms ląstelių dalijimosi metu.
Eukariotinės ląstelės branduolį supa dvi elementarios membranos, kurios sudaro branduolio apvalkalą. Jis persmelktas daugybe porų, kurių skersmuo nuo 30 iki 100 nm, matomos tik elektroniniu mikroskopu. Poros turi sudėtingą struktūrą. išorinė membrana branduolio membrana kai kuriose vietose yra sujungta su endoplazminiu tinklu. Branduolinis apvalkalas gali būti vertinamas kaip specializuota, lokaliai diferencijuota endoplazminio tinklo (ER) dalis.
Specialiais dažais nudažytame branduolyje galima išskirti ploni siūlai o chromatino ir nukleoplazmos (pagrindinės branduolio medžiagos) gabalėliai. Chromatinas susideda iš DNR, susietos su specialiais baltymais, vadinamais histonais. Ląstelių dalijimosi procese chromatinas vis labiau sutankinamas ir surenkamas į chromosomas. DNR koduoja genetinę informaciją.
Organizmai skiriasi chromosomų skaičiumi somatinėse ląstelėse. Pavyzdžiui, kopūstai turi - 20 chromosomų; saulėgrąžos - 34; kviečiai - 42; žmonių – 46, o viena iš paparčio Ophioglossum rūšių – 1250. Lytinės ląstelės (gametos) turi tik pusę chromosomų, būdingų somatinėms kūno ląstelėms. Chromosomų skaičius gametose vadinamas haploidiniu (viengubu), somatinėse ląstelėse – diploidiniu (dvigubu). Ląstelės, turinčios daugiau nei du chromosomų rinkinius, vadinamos poliploidinėmis.
Šviesos mikroskopu matomos sferinės struktūros – branduoliai. Kiekvienas branduolys turi vieną ar daugiau branduolių, kurie matomi nesidalijančiuose branduoliuose. Ribosominė RNR sintetinama branduolyje. Paprastai diploidinių organizmų branduoliuose yra du branduoliai, po vieną kiekvienam haploidinis rinkinys chromosomos. Branduoliai neturi savo membranos. Biochemiškai apibūdinami branduoliai didelė koncentracija RNR, kuri čia yra susijusi su fosfoproteinais. Branduolių dydis priklauso nuo funkcinė būklė ląstelės. pastebėta, kad sparčiai augančioje ląstelėje, kurioje vyksta intensyvūs baltymų sintezės procesai, didėja branduolių dydis. Branduoliuose gaminasi mRNR ir ribosomos, kurios sintetinę funkciją atlieka tik branduolyje.
Nukleoplazmą (karioplazmą) vaizduoja vienalytis skystis, kuriame yra ištirpę įvairūs baltymai, įskaitant fermentus.
Plastidai. Vakuolės, celiuliozės ląstelių sienelės ir plastidai yra būdingi augalų ląstelių komponentai. Kiekviena plastidė turi savo apvalkalą, susidedantį iš dviejų elementarių membranų. Plastido viduje išskiriama membraninė sistema ir įvairaus laipsnio vienalytė medžiaga – stroma. Subrendusios plastidės klasifikuojamos pagal juose esančius pigmentus.
Chloroplastuose, kuriuose vyksta fotosintezė, yra chlorofilų ir karotinoidų. Paprastai turi 4–5 mikronų skersmens disko formą. Vienoje mezofilo ląstelėje (lapo viduryje) gali būti 40 - 50 chloroplastų; mm 2 lapuose – apie 500 000. citoplazmoje chloroplastai dažniausiai išsidėstę lygiagrečiai ląstelės membranai.
Vidinė chloroplasto struktūra yra sudėtinga. Stroma yra persmelkta išsivysčiusios membranų sistemos pūslelių pavidalu – tilakoidų. Kiekvienas tilakoidas susideda iš dviejų membranų. Susidaro tilakoidai viena sistema. Paprastai jie surenkami krūvomis – grūdeliais, primenančiais monetų stulpelius. Atskirų granatų tilakoidai yra tarpusavyje sujungti stromos tilakoidais arba tarpgranaliniais tilakoidais. Chlorofilas ir karotinoidai yra įterpti į tilakoidų membranas. Žaliųjų augalų ir dumblių chloroplastuose dažnai yra krakmolo grūdelių ir mažų lipidų (riebalų) lašelių. Krakmolo grūdai yra laikinos fotosintezės produktų saugyklos. Iš tamsoje išbuvusio augalo chloroplastų jie gali išnykti tik 24 valandas, o vėl atsirasti praėjus 3–4 valandoms po augalo perkėlimo į šviesą.
Chloroplastai yra pusiau autonominiai organeliai ir panašūs į bakterijas. Pavyzdžiui, bakterijų ir chloroplastų ribosomos yra gana panašios. Jie yra mažesni už eukariotų ribosomas. Baltymų sintezę bakterijų ir chloroplastų ribosomose slopina chloramfenikolis, kuris eukariotinėse ląstelėse neveikia. Be to, tiek bakterijos, tiek chloroplastai turi panašų nukleoidų tipą, organizuotą panašiai. Nepaisant to, kad chloroplastų susidarymą ir juose esančių pigmentų sintezę daugiausia kontroliuoja ląstelės chromosomų DNR, vis dėlto, nesant savo DNR, chloroplastai nesusidaro.
Chloroplastai gali būti laikomi pagrindiniais ląstelių organeliais, nes jie yra pirmieji saulės energijos transformacijos grandinėje, dėl kurios žmonija gauna ir maistą, ir kurą. Chloroplastuose vyksta ne tik fotosintezė. Jie taip pat dalyvauja aminorūgščių sintezėje ir riebalų rūgštys, tarnauja kaip laikinųjų krakmolo atsargų saugykla.
Chromoplastai yra pigmentuoti plastidai. Įvairios formos, jos neturi chlorofilo, tačiau sintetina ir kaupia karotinoidus, kurie žiedams, seniems lapams, vaisiams ir šaknims suteikia geltoną, oranžinę, raudoną spalvą. iš chloroplastų gali išsivystyti chromoplastai, kurie tokiu atveju praranda chlorofilą ir vidines membranines struktūras, kaupia karotenoidus. Tai atsitinka daugelio vaisių nokimo metu. Chromoplastai pritraukia vabzdžius ir kitus gyvūnus, su kuriais jie vystėsi kartu.
Leukoplastai yra nepigmentuoti plastidai. Vieni jų sintetina krakmolą (amiloplastus), kiti gali susidaryti įvairių medžiagų, įskaitant lipidus ir baltymus. Šviesoje leukoplastai virsta chloroplastais.
Proplastidai yra mažos, bespalvės arba šviesiai žalios, nediferencijuotos plastidės, randamos meristematinėse (dalijamosiose) šaknų ir ūglių ląstelėse. Jie yra kitų, labiau diferencijuotų plastidų – chloroplastų, chromoplastų ir aminoplastų – pirmtakai. Jei protoplastidų vystymasis vėluoja dėl šviesos trūkumo, juose gali atsirasti vienas ar daugiau prolamelių kūnų, kurie yra pusiau kristalinės vamzdinių membranų sankaupos. Plastidės, kuriose yra prolamelių kūnų, vadinamos etioplastais. Šviesoje etioplastai virsta chloroplastais, o prolamelių kūnų membranos sudaro tilakoidus. Etioplastai susidaro tamsoje esančių augalų lapuose. sėklinių embrionų protoplastai pirmiausia virsta etioplastais, iš kurių vėliau šviesoje vystosi chloroplastai. Plastidams būdingas gana lengvas perėjimas iš vienos rūšies į kitą. Plastidės, kaip ir bakterijos, dauginasi dalijantis į dvi dalis. Meristematinėse ląstelėse protoplastidų dalijimosi laikas maždaug sutampa su ląstelių dalijimosi laiku. Tačiau subrendusiose ląstelėse dauguma Plastidė susidaro dėl subrendusių plastidžių dalijimosi.
Mitochondrijos. Kaip ir chloroplastus, mitochondrijas supa dvi elementarios membranos. Vidinė membrana suformuoja daug raukšlių ir išsikišimų – cristae, kurios žymiai padidina vidinį mitochondrijų paviršių. Jie yra žymiai mažesni už plastidus, apie 0,5 µm skersmens ir įvairaus ilgio bei formos.
Mitochondrijose vyksta kvėpavimo procesas, kurio pasekoje organinės molekulės suskaidomos išsiskiriant energijai ir ją perduodant ATP molekulėms – pagrindiniam visų eukariotinių ląstelių energijos rezervui. Daugumoje augalų ląstelių yra šimtai ar tūkstančiai mitochondrijų. Jų skaičius vienoje ląstelėje nustatomas pagal ląstelės ATP poreikį. Mitochondrijos nuolat juda, juda iš vienos ląstelės dalies į kitą, susilieja viena su kita ir dalijasi. Mitochondrijos dažniausiai susirenka ten, kur reikia energijos. Jei plazminė membrana aktyviai perneša medžiagas iš ląstelės į ląstelę, tai mitochondrijos yra išilgai membranos paviršiaus. Judriuose vienaląsčiuose dumbliuose mitochondrijos kaupiasi žvynelių apačioje, tiekdamos jiems judėjimui reikalingą energiją.
Mitochondrijos, kaip ir plastidai, yra pusiau autonominės organelės, turinčios komponentų, reikalingų jų pačių baltymų sintezei. Vidinė membrana supa skystą matricą, kurioje yra baltymų, RNR, DNR, į bakterijas panašių ribosomų ir įvairių tirpių medžiagų. DNR egzistuoja apskritų molekulių, esančių viename ar keliuose nukleoiduose, pavidalu.
Remiantis bakterijų panašumu su eukariotinių ląstelių mitochondrijomis ir chloroplastais, galima daryti prielaidą, kad mitochondrijos ir chloroplastai atsirado iš bakterijų, kurios „prieglobstį“ rado didesnėse heterotrofinėse ląstelėse – eukariotų pirmtakuose.
Mikrokūnai. Skirtingai nuo plastidžių ir mitochondrijų, kurias riboja dvi membranos, mikrokūnai yra sferinės organelės, apsuptos viena membrana. Mikrokūnai turi granuliuotą (granuliuotą) turinį, kartais juose yra ir kristalinių baltymų intarpų. Mikrokūnai yra susiję su vienu ar dviem endoplazminio tinklo sritimis.
Kai kurie mikrokūnai, vadinami proksisomomis, žaidžia svarbus vaidmuo glikolio rūgšties metabolizme, kuris yra tiesiogiai susijęs su fotokvėpavimu. Žaliuose lapuose jie yra susiję su mitochondrijomis ir chloroplastais. Kituose mikrokūnuose, vadinamuose glioksisomomis, yra fermentų, reikalingų riebalams paversti angliavandeniais. Tai atsitinka daugelyje sėklų dygimo metu.
Vakuolės – tai membrana apribotos ląstelės sritys, užpildytos skysčiu – ląstelės sultimis. Juos supa tonoplastas (vakuolinė membrana).
Jaunoje augalo ląstelėje yra daug mažų vakuolių, kurios ląstelei senstant susilieja į vieną didelę vakuolę. Brandžioje ląstelėje iki 90% jos tūrio gali užimti vakuolė. Šiuo atveju citoplazma plono periferinio sluoksnio pavidalu prispaudžiama prie ląstelės membranos. Ląstelių dydžio padidėjimą daugiausia lemia vakuolės augimas. Dėl to atsiranda turgorinis slėgis ir išlaikomas audinių elastingumas. Tai viena iš pagrindinių vakuolės ir tonoplasto funkcijų.
Pagrindinis sulčių komponentas yra vanduo, likusi dalis skiriasi priklausomai nuo augalo rūšies ir jo fiziologinės būklės. Vakuolėse yra druskų, cukrų, rečiau baltymų. Tonoplastas vaidina aktyvų vaidmenį transportuojant ir kaupiant tam tikrus jonus vakuolėje. Jonų koncentracija ląstelės sultyse gali gerokai viršyti jos koncentraciją aplinkoje. At didelis kiekis Kai kurios medžiagos formuoja kristalus vakuolėse. Labiausiai paplitę kalcio oksalato kristalai yra skirtinga forma.
Vakuolės yra medžiagų apykaitos produktų (metabolizmo) kaupimosi vietos. Tai gali būti baltymai, rūgštys ir net žmonėms toksiškos medžiagos (alkaloidai). Pigmentai dažnai nusėda. Mėlyna, violetinė, violetinė, tamsiai raudona, tamsiai raudona suteikia augalų ląstelėms pigmentų iš antocianinų grupės. Skirtingai nuo kitų pigmentų, jie gerai tirpsta vandenyje ir yra ląstelių sultyse. Jie lemia daugelio daržovių (ridikėlių, ropių, kopūstų), vaisių (vynuogių, slyvų, vyšnių), gėlių (rugiagėlių, pelargonijų, delfinijų, rožių, bijūnų) raudoną ir mėlyną spalvą. Kartais šie pigmentai užmaskuoja chlorofilą lapuose, pavyzdžiui, dekoratyviniame raudonajame kleve. Antocianinai nuspalvina rudens lapus ryškiai raudonos spalvos. Jie susidaro šaltu saulėtu oru, kai lapuose sustoja chlorofilo sintezė. Lapuose, kai nesusidaro antocianinai, sunaikinus chlorofilą, pastebimi geltonai oranžiniai chloroplastų karotenoidai. Lapai ryškiausios spalvos būna šaltą, giedrą rudenį.
Vakuolės dalyvauja makromolekulių sunaikinime, jų komponentų cirkuliacijoje ląstelėje. Ribosomos, mitochondrijos, plastidai, patenkantys į vakuoles, sunaikinami. Pagal šią virškinimo veiklą jas galima palyginti su lizosomomis – gyvūnų ląstelių organelėmis.
Vakuolės susidaro iš endoplazminio tinklo (tinklo)
Ribosomos. Mažos dalelės (17–23 nm), susidedančios iš maždaug vienodo kiekio baltymų ir RNR. Ribosomose aminorūgštys susijungia ir sudaro baltymus. Ląstelėse, kuriose vyksta aktyvi medžiagų apykaita, jų yra daugiau. Ribosomos yra ląstelės citoplazmoje laisvai arba prijungtos prie endoplazminio tinklo (80S). Jų taip pat yra branduolyje (80S), mitochondrijose (70S), plastiduose (70S).
Ribosomos gali sudaryti kompleksą, kuriame vienu metu vyksta identiškų polipeptidų sintezė, informacija apie kurią paimama iš vienos molekulės ir RNR. Toks kompleksas vadinamas poliribosomomis (polisomomis). Ląstelės, kurios sintetina baltymus dideliais kiekiais, turi didelę polisomų sistemą, prie kurios dažnai prisijungia išorinis paviršius branduolio lukštai.
Endoplazminis Tinklelis. Tai sudėtinga neriboto dydžio trimatė membraninė sistema. ER kontekste atrodo kaip dvi elementarios membranos su siaura skaidria erdve tarp jų. ER forma ir ilgis priklauso nuo ląstelės tipo, jos metabolinio aktyvumo ir diferenciacijos stadijos. Ląstelėse, kurios išskiria arba kaupia baltymus, ER yra plokščių maišelių arba cisternų formos, o prie išorinio paviršiaus yra pritvirtinta daug ribosomų. Šis tinklas vadinamas grubiu endoplazminiu tinkleliu. Lygus ER paprastai yra vamzdinės formos. Toje pačioje ląstelėje gali būti šiurkštus ir lygus endoplazminis tinklas. Paprastai tarp jų yra daug ryšių.
Endoplazminis tinklas veikia kaip ląstelės ryšio sistema. Jis yra susijęs su išorinis apvalkalas branduoliai. Tiesą sakant, šios dvi struktūros sudaro vieną membranų sistemą. Kai ląstelės dalijimosi metu plyšta branduolio apvalkalas, jo fragmentai primena ER fragmentus. Endoplazminis tinklas yra sistema, pernešanti medžiagas: baltymus, lipidus, angliavandenius į skirtingas ląstelės dalis. kaimyninių ląstelių endoplazminis tinklas yra sujungtas per citoplazmines sruogas - plazmodesmatas -, kurios praeina per ląstelių membranas.
Endoplazminis tinklas yra pagrindinė sintezės vieta ląstelių membranos. Kai kuriose augalų ląstelėse čia susidaro vakuolių ir mikrokūnų membranos, diktiozomų cisternos.
Goldžio kompleksas. Šis terminas vartojamas apibūdinti visas diktiozomas arba Golgi kūnus ląstelėje. Diktiosomos yra plokščių, disko formos pūslelių arba cisternų grupės, kurios išsišakoja sudėtinga sistema kanalėlių. Aukštesniųjų augalų diktiosomos susideda iš 4–8 cisternų, surinktų kartu.
Paprastai rezervuarų pakuotėje išskiriama besivystanti ir bręstanti pusė. formuojančių cisternų membranos savo struktūra primena ER membranas, o bręstančių cisternų membranos primena plazminę membraną.
Diktiosomos dalyvauja sekrecijoje, o daugumoje aukštesniųjų augalų – ląstelių membranų formavime. Diktiosomų sintetinami ląstelių sienelių polisacharidai kaupiasi pūslelėse, kurios vėliau atsiskiria nuo bręstančių cisternų. Šios sekrecinės pūslelės migruoja ir susilieja su CMP; o juose esantys polisacharidai yra įterpti į ląstelės membraną. Kai kurios medžiagos, besikaupiančios diktiozomose, susidaro kitose struktūrose, pavyzdžiui, ER, o po to pernešamos į diktioomas, kur jos modifikuojamos (modifikuojamos) prieš sekreciją. Pavyzdžiui, glikoproteinai yra svarbūs statybinė medžiaga ląstelės membrana. Baltyminę dalį sintetina šiurkščiojo ER polisomos, angliavandenių dalį – diktiozomose, kur abi dalys susijungdamos sudaro glikoproteinus.
Membranos yra dinamiškos, judrios struktūros, nuolat keičiančios savo formą ir plotą. Endoplazminės sistemos samprata remiasi membranų mobilumu. Pagal šią koncepciją vidinės citoplazmos membranos, išskyrus mitochondrijų ir plastidžių membranas, yra viena visuma ir kyla iš endoplazminio tinklo. Iš endoplazminio tinklo per tarpinę pūslelių stadiją susidaro naujos diktiozomų cisternos, o sekrecinės pūslelės, kurios atsiskiria nuo diktiozomų, ilgainiui prisideda prie plazminės membranos susidarymo. Taigi, endoplazminis tinklas ir diktiosomos sudaro funkcinę visumą, kurioje diktiosomos atlieka svarbų vaidmenį tarpinės struktūrosį endoplazminį tinklą panašias membranas paverčiant membranomis, panašiomis į plazmą. Audiniuose, kurių ląstelės auga ir dalijasi silpnai, membranos komponentai nuolat atnaujinami.
Mikrovamzdeliai randami beveik visose eukariotų ląstelėse. Tai cilindrinės struktūros, kurių skersmuo yra apie 24 nm. Jų ilgis skiriasi. Kiekvienas kanalėlis sudarytas iš baltymo, vadinamo tubulinu, subvienetų. Subvienetai sudaro 13 išilginių gijų, supančių centrinę ertmę. Mikrovamzdeliai yra dinamiškos struktūros, jos reguliariai sunaikinamos ir susidaro tam tikrais ląstelių ciklo etapais. Jie surenkami ypatingos vietos vadinami mikrovamzdelių organizavimo centrais. Augalų ląstelėse jie turi silpnai išreikštą amorfinę struktūrą.
Mikrovamzdelių funkcijos: dalyvauja formuojant ląstelės membraną; nukreipti diktiozomų burbulus į besiformuojančią membraną, pavyzdžiui, verpstės siūlus, kurie susidaro besidalijančioje ląstelėje; vaidina vaidmenį formuojant ląstelės plokštelę (pradinę ribą tarp dukterinių ląstelių). Be to, mikrovamzdeliai svarbus komponentasžvyneliais ir blakstienomis, kurių judėjime jos vaidina svarbų vaidmenį.
Mikrofilamentai, kaip ir mikrovamzdeliai, randami beveik visose eukariotų ląstelėse. Tai ilgi, 5–7 nm storio siūlai, susidedantys iš susitraukiančio baltymo aktino. Mikrofilamentų ryšuliai randami daugelyje aukštesniųjų augalų ląstelių. Matyt, jie vaidina svarbų vaidmenį citoplazmos srovėse. Mikrofilamentai kartu su mikrotubuliais sudaro lankstų tinklą, vadinamą citoskeletu.
Susmulkinta medžiaga nuo seno buvo laikoma vienalyčiu (homogenišku) baltymų turtingu tirpalu, turinčiu nedaug struktūrų arba visai neturinčiu. Tačiau šiuo metu, naudojant aukštos įtampos elektroninį mikroskopą, nustatyta, kad gruntinė medžiaga yra trimatė gardelė, sudaryta iš plonų (3–6 nm skersmens) gijų, užpildančių visą ląstelę. Kiti citoplazmos komponentai, įskaitant mikrovamzdelius ir mikrofilamentus, yra suspenduoti iš šios mikrotrabekulinės gardelės.
Mikrotrabekulinė struktūra yra baltymų gijų gardelė, tarp kurių esanti erdvė užpildyta vandeniu. Kartu su vandeniu grotelės yra gelio konsistencijos, gelis atrodo kaip želatininiai kūnai.
Organelės yra pritvirtintos prie mikrotrabekulinės gardelės. Grotelės suteikia ryšį tarp atskiros dalys ląstelės ir nukreipia intracelulinį transportą.
Lipidų lašeliai yra sferinės struktūros, suteikiančios citoplazmai granuliuotumą augalo ląstelė po šviesos mikroskopu. Elektroninėse mikrografijose jie atrodo amorfiški. Labai panašūs, bet mažesni lašai randami plastiduose.
Lipidų lašai, painiodami juos su organelėmis, vadino juos sferosomomis ir tikėjo, kad juos supa vieno arba dvigubo sluoksnio membrana. Tačiau naujausi duomenys rodo, kad lipidų lašeliai neturi membranų, bet gali būti padengti baltymu.
Ergastinės medžiagos yra protoplasto „pasyvieji produktai“: atsarginės medžiagos arba atliekos. Jie gali atsirasti ir išnykti skirtingi laikotarpiai ląstelių ciklas. Išskyrus krakmolo grūdelius, kristalus, antocianinų pigmentus ir lipidų lašelius. Tai yra dervos, dervos, taninai ir baltymai. Ergastinės medžiagos yra ląstelės membranos dalis, pagrindinė citoplazmos ir organelių, įskaitant vakuoles, medžiaga.
Vėliavos ir blakstienos yra plonos, į plaukus panašios struktūros, besitęsiančios nuo daugelio eukariotinių ląstelių paviršiaus. Jų skersmuo yra pastovus, tačiau ilgis svyruoja nuo 2 iki 150 mikronų. Tradiciškai ilgesnės ir kelios jų vadinamos žiuželiais, o trumpesnės ir gausesnės – blakstienomis. Nėra aiškių skirtumų tarp šių dviejų tipų struktūrų, todėl terminas flagellum vartojamas abiem.
Kai kuriuose dumbliuose ir grybuose žvyneliai yra judėjimo organai, kurių pagalba jie juda vandenyje. Augaluose (pavyzdžiui, samanose, kepenėliuose, paparčiuose, kai kuriuose gimnasėkliuose) žvyneliais yra tik lytinės ląstelės (gametos).
Kiekvienas žvynelis turi tam tikrą organizaciją. Išorinis 9 porų mikrovamzdelių žiedas supa du papildomus mikrovamzdelius, esančius žvynelio centre. Fermentų turinčios „rankenos“ tęsiasi iš kiekvienos išorinės poros vieno mikrotubulo. Tai yra pagrindinis 9+2 organizavimo modelis, randamas visose eukariotinių organizmų žvyneliuose. Manoma, kad žvynelių judėjimas pagrįstas mikrotubulių slydimu, o išorinės mikrovamzdelių poros juda viena iš kitos nesusitraukdamos. Mikrovamzdelių porų slydimas vienas kito atžvilgiu sukelia vietinį žvynelio lenkimą.
Žvyneliai „auga“ iš citoplazminių cilindrinių struktūrų, vadinamų baziniais kūnais, kurie taip pat sudaro bazinę žvynelio dalį. Baziniai kūnai turi vidinė struktūra, panašus į žvynelio struktūrą, išskyrus tai, kad išoriniai kanalėliai surenkami trynukais, o ne poromis, o centrinių kanalėlių nėra.
Ląstelių sienelės. Ląstelės sienelė riboja protoplasto dydį ir neleidžia jam plyšti dėl vandens sugėrimo vakuolės.
Ląstelės sienelė atlieka specifines funkcijas, kurios svarbios ne tik ląstelei ir audiniui, kuriame yra ląstelė, bet ir visam augalui. Ląstelių sienelės atlieka esminį vaidmenį įsisavinant, pernešant ir pašalinant medžiagas, be to, jose gali koncentruotis lizosominė, arba virškinimo veikla.
Ląstelės sienelės komponentai. Tipiškiausias ląstelės sienelės komponentas yra celiuliozė, kuri daugiausia lemia jos architektūrą. Celiuliozės molekulės yra sudarytos iš pasikartojančių gliukozės molekulių, sujungtų galais. Ilgos plonos celiuliozės molekulės sujungiamos į 10–25 nm storio mikrofibriles. Mikrofibrilės susipina ir sudaro plonus siūlus, kurie savo ruožtu gali būti apvynioti vienas aplink kitą, tarsi sruogos virvelėje. Kiekviena tokia „virvė“ arba makrofibrilė yra apie 0,5 mikrono storio, o ilgis siekia 4 mikronus. Makrofibrilės yra tokios pat stiprios kaip vienodo dydžio plieninė viela.
Ląstelės sienelės celiuliozės karkasas užpildytas su juo susipynusiomis celiuliozės matricos molekulėmis. Jį sudaro polisacharidai, vadinami hemiceliuliozės, ir pektinai, arba pektinai, chemiškai labai artimi hemiceliuliozei.
Kitas ląstelės sienelės komponentas – ligninas – yra gausiausias polimeras augalų ląstelėse po celiuliozės. Ligninas padidina sienos standumą ir dažniausiai randamas ląstelėse, kurios atlieka atraminę arba mechaninę funkciją.
Kutinas, suberinas, vaškas – dažniausiai nusėda augalų apsauginių audinių lukštuose. Pavyzdžiui, kutinas randamas epidermio ląstelių membranose, o suberinas – antriniame apsauginiame audinyje – kamštelyje. Abi medžiagos randamos kartu su vašku ir neleidžia augalui pernelyg prarasti vandens.
Ląstelės sienelės sluoksniai. Augalų ląstelių sienelių storis labai skiriasi, priklausomai nuo ląstelių vaidmens augalo struktūroje ir pačios ląstelės amžiaus. Pagal elektroninis mikroskopas augalo ląstelės sienelėje matomi du sluoksniai: mediana (taip pat vadinama tarpląsteline medžiaga) ir pirminė ląstelės sienelė. Daugelis ląstelių kloja kitą sluoksnį – antrinį ląstelių sienelės. Vidutinė plokštė yra tarp gretimų ląstelių pirminių sienelių. Antrinę sienelę, jei yra, ląstelės protoplastas nusodina ant pirminės ląstelės sienelės vidinio paviršiaus.
Vidurinė plokštelė. Vidurinę plokštelę daugiausia sudaro pektinas. Ten, kur tarp dviejų naujai susidariusių ląstelių turėtų atsirasti ląstelės sienelė, pirmiausia pastebimas tankus endoplazminio tinklo kanalėlių rezginys ir Golgi aparato cisterna (diktiosomos). Tada šioje vietoje atsiranda burbuliukai, užpildyti pektinu (iš polisacharidų). Šie burbuliukai yra atskirti nuo Golgi aparato rezervuarų. Ankstyvojoje ląstelės sienelėje yra įvairių polisacharidų, iš kurių pagrindiniai yra pektinai ir hemiceliuliozė. Vėliau jis susideda iš tankesnių medžiagų – celiuliozės ir lignino.
Pirminė ląstelės sienelė. Tai yra celiuliozės apvalkalo sluoksnis, kuris nusėda prieš ląstelių augimą arba jo metu. Be celiuliozės, hemiceliuliozės ir pektino, pirminėse membranose yra glikoproteino. Pirminiai apvalkalai gali sudegti. Pektino komponentas suteikia plastiškumo, todėl pirminis apvalkalas gali išsiplėsti, kai šaknis, stiebas ar lapas pailgėja.
Aktyviai besidalijančios ląstelės (dauguma subrendusių ląstelių, dalyvaujančių fotosintezės, kvėpavimo ir sekrecijos procesuose) turi pirmines membranas. Tokios ląstelės, turinčios pirminę membraną ir gyvą protoplastą, gali prarasti būdinga forma, dalijasi ir diferencijuos į naują ląstelių tipą. Jie dalyvauja žaizdų gijime ir audinių regeneracijoje augaluose.
Pirminės ląstelių membranos nėra vienodo storio per visą ilgį, tačiau turi plonas dalis, kurios vadinamos pirminių porų laukais. Citoplazmos gijos arba plazmodesmata, jungiančios gretimų ląstelių protoplastus, dažniausiai praeina per pirminius porų laukus.
antrinė ląstelės sienelė. Nepaisant to, kad daugelis augalų ląstelių turi tik pirminę sienelę, kai kuriose, protoplastas nusodina antrinę sienelę link ląstelės centro. Paprastai tai įvyksta sustojus ląstelių augimui ir pirminės membranos plotas nebedidėja. Dėl šios priežasties antrinis apvalkalas skiriasi nuo pirminio. Antrinės membranos ypač reikalingos specializuotoms ląstelėms, kurios stiprina augalą ir praleidžia vandenį. Nusodinus antrinę membraną, šių ląstelių protoplastas, kaip taisyklė, miršta. Antriniuose apvalkaluose celiuliozės yra daugiau nei pirminiuose, juose nėra pektino medžiagų ir glikoproteinų. Antrinė membrana sunkiai ištempiama, jos matrica susideda iš hemiceliuliozės.
Antriniame apvalkale galima išskirti tris sluoksnius – išorinį, vidurinį ir vidinį (S 1, S 2, S 3). Sluoksniuota antrinių apvalkalų struktūra žymiai padidina jų stiprumą. Celiuliozės mikrofibrilės antrinėje membranoje nusėda tankiau nei pirminėje. Ligninas - įprastas komponentas antriniai mediniai apvalkalai.
Poros kontaktuojančių ląstelių membranose yra viena priešais kitą. Dvi priešingos poros ir porų membrana sudaro porų porą. Ląstelėse su antrine membrana yra du pagrindiniai porų tipai: paprastos ir apvaduotos. Apribotose porose antrinė sienelė kabo virš porų ertmės. Paprastose porose taip nėra.
Ląstelių sienelių augimas. Kai ląstelė auga, didėja ląstelės membranos storis ir plotas. Lukšto tempimas yra sudėtingas procesas. Jį kontroliuoja protoplastas ir reguliuoja hormonas auksinas.
Ląstelėse, kurios auga tolygiai visomis kryptimis, miofibrilių nusėdimas yra atsitiktinis. Šios miofibrilės sudaro netaisyklingą tinklą. Tokios ląstelės buvo aptiktos stiebo šerdyje, saugojimo audiniuose ir ląstelių auginimo in vitro metu. Pailgėjusiose ląstelėse šoninių apvalkalų miofibrilės nusėda stačiu kampu pailgėjimo ašiai.
Matricinės medžiagos – pektinai, hemiceliuliozės ir glikoproteinai į membraną pernešami diktiozomų pūslelėmis. Tuo pačiu pektinai labiau būdingi augančioms ląstelėms, o neaugančiose ląstelėse vyrauja hemiceliuliozės.
Celiuliozės mikrofibrilės sintezuojamos ląstelės paviršiuje, padedant fermentų kompleksui, susijusiam su plazmine membrana. Mikrofibrilių orientaciją kontroliuoja mikrovamzdeliai, esantys adresu vidinis paviršius plazmos membrana.
Plazmodezma. Tai ploni citoplazmos siūlai, jungiantys kaimyninių ląstelių protoplastus. Plazmodesmatos arba praeina pro ląstelės sienelę bet kur, arba yra susitelkusios pirminiuose porų laukuose arba membranose tarp porų porų. Elektroniniu mikroskopu plazmodesmata atrodo kaip siauri kanalai, iškloti plazmos membrana. Išilgai kanalo ašies nuo vienos ląstelės iki kitos tęsiasi mažesnis cilindrinis vamzdelis – desmotubulas, kuris bendrauja su abiejų gretimų ląstelių endoplazminiu tinklu. Daugelis plazmodesmų susidaro ląstelių dalijimosi metu, kai besivystančios ląstelės plokštelė paima vamzdinį endoplazminį tinklą. Plazmodezmos taip pat gali susidaryti nesidalijančių ląstelių membranose. Šios struktūros užtikrina efektyvų tam tikrų medžiagų perkėlimą iš ląstelės į ląstelę.
Ląstelių dalijimasis. At daugialąsčiai organizmai ląstelių dalijimasis kartu su jų dydžio padidėjimu yra viso organizmo augimo būdas. Dalijimosi metu susidariusios naujos ląstelės savo struktūra ir funkcijomis yra panašios tiek į pirminę, tiek į viena kitą. Eukariotų dalijimosi procesą galima suskirstyti į du iš dalies persidengiančius etapus: mitozę ir citokinezę.
Mitozė – tai dviejų dukterinių branduolių susidarymas iš vieno branduolio, morfologiškai ir genetiškai lygiaverčiai vienas kitam. Citokinezė – tai ląstelės citoplazminės dalies dalijimasis, kai susidaro dukterinės ląstelės.
Ląstelių ciklas. Gyva ląstelė pereina iš eilės įvykių, kurie sudaro ląstelės ciklą. Pačio ciklo trukmė skiriasi priklausomai nuo ląstelės tipo ir išoriniai veiksniai, pavyzdžiui, dėl temperatūros ar saugumo maistinių medžiagų. Paprastai ciklas skirstomas į tarpfazę ir keturias mitozės fazes.
Tarpfazė. Laikotarpis tarp nuoseklių mitozinių dalijimų.
Tarpfazė yra padalinta į tris periodus, žymimus kaip G 1 , S , G 2 .
G 1 periodu, kuris prasideda po mitozės. Per šį laikotarpį padidėja citoplazmos kiekis, įskaitant įvairius organelius. Be to, remiantis šiuolaikine hipoteze, G 1 periodo metu sintetinamos medžiagos, kurios arba stimuliuoja, arba slopina S periodą ir likusį ciklą, taip nulemdamos dalijimosi procesą.
S periodas seka po G1 periodo, tuo metu genetinė medžiaga (DNR) yra dubliuojama.
Per G2 laikotarpį, einantį po S, susidaro struktūros, tiesiogiai susijusios su mitoze, pavyzdžiui, veleno komponentai.
Kai kurios ląstelės praeina neribotą ląstelių ciklų skaičių. Tai vienaląsčiai organizmai ir kai kurios aktyvaus augimo zonų ląstelės (meristemos). Kai kurios specializuotos ląstelės po brendimo praranda gebėjimą daugintis. Trečioji ląstelių grupė, pavyzdžiui, formuojančios žaizdos audinį (kalusą), gebėjimą dalytis išlaiko tik ypatingomis sąlygomis.
Mitozė arba branduolio dalijimasis. Tai nenutrūkstamas procesas, suskirstytas į keturias fazes: profazę, metafazę, anafazę, telofazę. Dėl mitozės tarpfazėje padvigubėjusi genetinė medžiaga po lygiai pasiskirsto tarp dviejų dukterinių branduolių.
Vienas is labiausiai ankstyvieji požymiai ląstelės perėjimas prie dalijimosi yra siauro žiedo formos mikrotubulių juostos atsiradimas tiesiai po plazmine membrana. Šis gana tankus diržas supa branduolį būsimojo mitozinio veleno pusiaujo plokštumoje. Kadangi jis pasirodo prieš profazę, jis vadinamas priešfazės juosta. Jis išnyksta po mitozinio verpstės, dar ilgai prieš atsirandant vėlyvojoje ląstelės plokštelės telofazėje, kuri auga nuo centro iki periferijos ir susilieja su motininės ląstelės membrana toje srityje, kurią anksčiau užėmė priešfazinis diržas.
Profazė. Profazės pradžioje chromosomos primena ilgus siūlus, išsibarsčiusius branduolio viduje. Tada, trumpėjant ir storėjant sruogoms, galima pastebėti, kad kiekviena chromosoma susideda ne iš vienos, o iš dviejų susipynusių sruogų, vadinamų chromatidėmis. Vėlyvojoje fazėje dvi sutrumpintos suporuotos kiekvienos chromosomos chromatidės yra lygiagrečiai, sujungtos siaura atkarpa, vadinama centromera. Jis turi tam tikrą vietą kiekvienoje chromosomoje ir padalija chromosomą į dvi skirtingo ilgio šakas.
Mikrovamzdeliai išsidėstę lygiagrečiai branduolio paviršiui išilgai veleno ašies. Tai yra ankstyviausias mitozinio veleno surinkimo pasireiškimas.
Profazės pabaigoje branduolys palaipsniui praranda aiškų kontūrą ir galiausiai išnyksta. Netrukus po to branduolinis apvalkalas taip pat suyra.
Metafazė. Metafazės pradžioje verpstė, kuri yra trimatė struktūra, plačiausia per vidurį ir siaurėjanti link polių, užima vietą, kurią anksčiau užėmė branduolys. Verpstės pluoštai yra mikrovamzdelių pluoštai. Metafazės metu chromosomos, kurių kiekviena susideda iš dviejų chromatidžių, yra išdėstytos taip, kad jų centromerai būtų suklio pusiaujo plokštumoje. Kiekviena chromosoma savo centromeru yra pritvirtinta prie veleno sriegių. Tačiau kai kurios sruogos pereina iš vieno poliaus į kitą neprisirišusios prie chromosomų.
Kai visos chromosomos yra pusiaujo plokštumoje, metafazė baigiama. Chromosomos yra pasirengusios dalytis.
Anafazė. Kiekvienos chromosomos chromatidės išsiskiria. Dabar tai yra dukterinės chromosomos. Visų pirma, centromera dalijasi, o dvi dukterinės chromosomos traukiamos į priešingus polius. Šiuo atveju centromerai juda į priekį, o chromosomų rankos išsitiesia už nugaros. Prie chromosomų prisirišę verpstės siūlai sutrumpėja, prisidedant prie chromatidų atsiskyrimo ir dukterinių chromosomų judėjimo priešingomis kryptimis.
Telofazė. Telofazėje baigiasi dviejų identiškų chromosomų grupių atskyrimas, o aplink kiekvieną iš jų susidaro branduolinė membrana. Šiurkštus tinklelis aktyviai dalyvauja čia. Dingsta veleno aparatas. Telofazės metu chromosomos praranda kontūrų aiškumą, išsitempia, vėl virsta plonais siūlais. Branduoliai atkuriami. Kai chromosomos tampa nematomos, mitozė baigiasi. Du dukteriniai branduoliai patenka į interfazę. Jie yra genetiškai lygiaverčiai vienas kitam ir pirminiam branduoliui. Tai labai svarbu, nes genetinė programa, o kartu ir visi požymiai, turi būti perduota dukteriniams organizmams.
Mitozės trukmė skiriasi įvairūs organizmai Ir tai priklauso nuo audinio tipo. Tačiau profazė yra ilgiausia, o anafazė – trumpiausia. Šaknies galiuko ląstelėse profazės trukmė yra 1–2 valandos; metafazė - 5 - 15 min; anafazė - 2 - 10 min; telofazė - 10 - 30 min. Tarpfazės trukmė nuo 12 iki 30 valandų.
Daugelyje eukariotinių ląstelių mikrotubulų organizavimo centrai, atsakingi už mitozinio veleno susidarymą, yra susiję su centriolais.
Citokinezė. Tai yra citoplazmos dalijimosi procesas. Daugumoje organizmų ląstelės dalijasi atitraukdamos ląstelės sienelę ir suformuodamos dalijimosi vagą, kuri palaipsniui gilėja ir suspaudžia likusius mitozinio veleno siūlus. Visuose augaluose (briofituose ir kraujagysliniuose augaluose) ir kai kuriuose dumbliuose ląstelės dalijasi dėl ląstelės plokštelės susidarymo.
Ankstyvojoje telofazėje tarp dviejų dukterinių branduolių susidaro statinės formos skaidulų sistema, vadinama fragmoplastu. Fragmoplasto pluoštai, kaip ir mitozinio veleno pluoštai, susideda iš mikrovamzdelių. Fragmoplasto pusiaujo plokštumoje atsiranda nedideli lašeliai. Jie susilieja ir sudaro ląstelės plokštelę, kuri auga tol, kol pasiekia besidalijančios ląstelės sienelę. Tai užbaigia dviejų dukterinių ląstelių atskyrimą. Susiliejantys lašeliai yra burbuliukai, atitrūkę nuo Golgi aparato. Iš esmės juose yra pektino medžiagų, iš kurių susidaro vidurinė plokštelė. Pūslelinės membranos dalyvauja kuriant plazminę membraną abiejose plokštelės pusėse. Tuo pačiu metu iš vamzdinio endoplazminio tinklo fragmentų susidaro plazmodesmata.
Susidarius vidutinei plokštelei, kiekvienas protoplastas ant jos nusodina pirminę membraną. Be to, kiekviena dukterinė ląstelė aplink visą protoplastą nusodina naują membranos sluoksnį, kuris tęsia membraną, kuri atsirado iš ląstelės plokštelės. Augant dukterinėms ląstelėms, pradinis pirminės ląstelės apvalkalas sunaikinamas.
Įvairūs eukariotų mitozės tipai
Aukščiau aprašytas ląstelių dalijimasis augaluose, taip pat ir gyvūnuose, nėra vienintelė netiesioginio ląstelių dalijimosi forma. Paprasčiausias mitozės tipas yra pleuromitozė. Tai primena dvejetainį padalijimą prokariotinės ląstelės, kuriame nukleoidai po replikacijos lieka susieti su plazmos membrana. Membrana pradeda augti tarp DNR surišimo taškų ir taip išplinta chromosomas skirtingos sritys ląstelės. Po to, susiformuojant ląstelių susiaurėjimui, kiekviena iš DNR molekulių bus naujoje atskiroje ląstelėje.
Būdingas eukariotinių ląstelių dalijimasis yra iš mikrovamzdelių sudaryto verpstės susidarymas. Esant uždarai pleuromitozei (ji vadinama uždara, nes chromosomų divergencija vyksta nepažeidžiant branduolio apvalkalo), ne centrioliai, o kitos ant jo esančios struktūros. viduje branduolio membrana. Tai vadinamieji neapibrėžtos morfologijos poliniai kūnai, iš kurių tęsiasi mikrovamzdeliai. Tokių kūnų yra du. Jie skiriasi vienas nuo kito neprarasdami ryšio branduolinis apvalkalas. Dėl to susidaro dvi pusverpstės, susijusios su chromosomomis. Visas mitozinio veleno formavimosi ir chromosomų segregacijos procesas šiuo atveju vyksta po branduolio apvalkalu. Šio tipo mitozė pasitaiko tarp pirmuonių ir yra plačiai paplitusi tarp grybų (chitridijų, zygomicetų, mielių, oomicetų, askomicetų, miksomicetų ir kt.). yra pusiau uždaros pleuromitozės formos, kai susidariusio veleno poliuose suardomas branduolio apvalkalas.
Kita mitozės forma yra ortomitozė. Šiuo atveju COMT yra citoplazmoje, nuo pat pradžių susidaro ne pusverpstės, o dvipolis velenas. Yra trys ortomitozės (įprastos mitozės) formos: pusiau uždara ir uždara. Esant pusiau uždarai ortomitozei, citoplazmoje esančio TsOMT pagalba susidaro bisimetrinis velenas, branduolinis apvalkalas išsaugomas per visą mitozės laikotarpį, išskyrus poliarines zonas. Granuliuotos medžiagos ar net centriolių masės gali būti aptiktos kaip COMT. Ši mitozės forma randama žaliųjų, rudųjų, raudonųjų dumblių zoosporose, kai kuriose apatiniai grybai ir gregarinas. Esant uždarai ortomitozei, visiškai išsaugoma branduolinė membrana, kurioje susidaro tikras velenas. Karioplazmoje susidaro mikrovamzdeliai, rečiau išauga iš vidinės TsOMT, kuri nesusijusi (skirtingai nei pleuromitozė) su branduoline membrana. Šio tipo mitozė būdinga blakstienų mikrobranduolių dalijimuisi, tačiau gali atsirasti ir pirmuoniams. Atviros ortomitozės atveju branduolinis apvalkalas visiškai suyra. Šis ląstelių dalijimosi tipas būdingas gyvūnų organizmams, kai kuriems pirmuoniams ir aukštesnių augalų ląstelėms. Šią mitozės formą savo ruožtu atstovauja astraliniai ir anastraliniai tipai.
Iš trumpos apžvalgos aišku, kad Pagrindinis bruožas Mitozė apskritai yra dalijimosi veleno struktūrų atsiradimas, susidarantis ryšium su TsOMT, kuris yra įvairios struktūros.
Mitozinės figūros morfologija
Mitozinis aparatas ypač gerai išreiškiamas mitozės metafazės stadijoje. Metafazėje, ląstelės pusiaujo plokštumoje, išsidėsčiusios chromosomos, iš kurių į priešingas puses tęsiasi vadinamieji verpstės siūlai, susiliejantys dviejuose skirtinguose mitozinės figūros poliuose. Taigi mitozinis velenas yra chromosomų, polių ir skaidulų rinkinys. Verpstės skaidulos yra pavieniai mikrovamzdeliai arba jų ryšuliai. Mikrovamzdeliai prasideda nuo verpstės polių, o dalis jų eina į centromerus, kur yra chromosomų kinetochorai (kinetochoro mikrovamzdeliai), dalis eina toliau link priešingo poliaus, bet jo nepasiekia. Jie vadinami „tarppoliais mikrotubulais“. Iš polių nukrypsta grupė radialinių mikrovamzdelių, aplink juos suformuodami struktūrą, primenančią „spinduliuojantį švytėjimą“ – tai astraliniai mikrovamzdeliai.
Pagal morfologiją mitozinės figūros skirstomos į astralinius ir anastralinius tipus.
Astralinis verpstės tipas, arba konvergentas, pasižymi tuo, kad jo polius vaizduoja maža zona, į kurią susilieja (susilenkia) mikrovamzdeliai. Paprastai centrosomos, kuriose yra centriolių, yra astralinių verpsčių poliuose. Tačiau yra žinomi centriolinių astralinių mitozių atvejai (kai kurių bestuburių mejozės metu). Be to, pastebimi radialiniai mikrovamzdeliai, sklindantys iš polių, kurie nėra veleno dalis, bet sudaro žvaigždžių zonas - citasters. Šio tipo mitozinis dalijimasis primena hantelį. Mitozinės figūros anastrialinio tipo polių nėra. Verpstės poliarinės sritys čia yra plačios, jos vadinamos poliniais gaubteliais, jose yra centriolių. Šiuo atveju veleno pluoštai nenukrypsta nuo vieno taško, o nukrypsta plačiu priekiu (nukrypsta) nuo visos poliarinių dangtelių zonos. Šio tipo verpstės būdingos besidalijančioms aukštesniųjų augalų ląstelėms, tačiau jos gali būti ir aukštesniuosiuose gyvūnuose. Ankstyvoje žinduolių embriogenezėje centriolinės (divergentinės) mitozės stebimos kiaušialąstės brendimo dalijimosi metu ir I bei II zigotos dalijimosi metu. Tačiau jau trečiojo ląstelių dalijimosi metu ir visose vėlesnėse ląstelėse jie dalijasi dalyvaujant astraliniams verpstams, kurių poliuose visada randama centriolių. Apskritai visoms mitozės formoms chromosomos su jų kinetoforais, poliniais kūnais (centrosomomis) ir verpstės skaidulomis išlieka įprastos struktūros.
Centromerai ir kinetochorai
Centromerai gali turėti skirtinga lokalizacija išilgai chromosomų ilgio. Holocentriniai centromerai susidaro, kai mikrovamzdeliai susijungia per visą chromosomos ilgį (kai kurie vabzdžiai, nematodai, kai kurie augalai). Monocentriniai centromerai – kai mikrovamzdeliai yra susieti su chromosomomis vienoje vietoje. monocentriniai centromerai gali būti taškiniai (pavyzdžiui, kai kuriose pumpurinėse mielėse), kai tik vienas mikrovamzdelis priartėja prie kinetochoro, ir zoninės, kai mikrovamzdelių pluoštas artėja prie kompleksinio kinetochoro. Nepaisant centromerų zonų įvairovės, jos visos yra susijusios su sudėtinga struktūra kinetochore, kurios struktūra ir funkcijos yra labai panašios visuose eukariotuose. Kinetochorai yra specializuotos baltymų struktūros didžiąja dalimi išsidėstę chromosomų centromeruose. Tai sudėtingi kompleksai, susidedantys iš daugelio baltymų. morfologiškai jie yra labai panašūs, turi tą pačią struktūrą – nuo diatomų iki žmonių. Tai yra trijų sluoksnių struktūros: vidinis tankus sluoksnis, esantis greta chromosomos kūno, vidurinis laisvas sluoksnis ir išorinis tankus sluoksnis. Daugelis fibrilių tęsiasi nuo išorinio sluoksnio, sudarydamos vadinamąjį pluoštinį kinetochoro vainiką. IN bendra forma kinetochorai yra plokštelių arba diskų formos, esančios pirminio chromosomos susiaurėjimo zonoje, centromeroje. Paprastai kiekvienoje chromosomoje arba chromatidėje yra vienas kinetochoras. Prieš anafazę kiekvienoje seserinėje chromatidėje yra kinetochorai, kurių kiekviena bendrauja su savo mikrotubulių pluoštu. Augaluose kinetochoras atrodo ne kaip plokštelės, o kaip pusrutuliai. Funkcinis kinetochorų vaidmuo yra surišti seserines chromatides viena su kita, fiksuoti mitozinius mikrotubulus, reguliuoti chromosomų atskyrimą ir faktiškai perkelti chromosomas mitozės metu, dalyvaujant mikrotubulams. Apskritai, baltymų struktūros, kinetochorai S periode dvigubėja, lygiagrečiai chromosomų padvigubėjimui. Tačiau jų baltymai yra chromosomose visais ląstelių ciklo laikotarpiais.
AUGALŲ AUDINIAI
Mokomieji audiniai(meristemos)
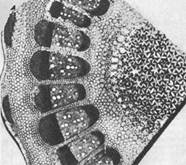
Ugdomieji audiniai augalų kūne yra skirtingos vietos, todėl jie skirstomi į tokias grupes (0 pav.; 1).
1. Viršūninės (viršūninės) meristemos išsidėsčiusios ašinių organų – stiebo, šaknies – viršūnėse, arba viršūnėse. Šių meristemų pagalba augalų vegetatyviniai organai vykdo savo ilgį.
2. Šoninės meristemos būdingos ašiniams organams. Ten jie yra koncentriškai, sankabos pavidalu.
3. Tarpkalarinės, arba tarpkalinės, meristemos kyla iš viršūninių meristemų. Tai ląstelių grupės, kurios dar nesugeba daugintis, bet yra išėjusios į diferenciacijos kelią. Tarp jų nėra pradinių ląstelių, tačiau yra daug specializuotų.
4. Žaizdų meristemos užtikrina pažeistos kūno dalies atstatymą. Regeneracija prasideda dediferenciacija, tai yra atvirkštinis vystymasis nuo specializuotų ląstelių iki meristematinių. Jie virsta felogenu, kuris sudaro kamštį, dengiantį žaizdos paviršių. Dediferencijuotos ląstelės, besidalijančios, gali suformuoti purų parenchiminį audinį – kaliusą. Iš jo tam tikromis sąlygomis susidaro augalų organai.
Integumentiniai audiniai
Jie veikia kaip ribinis barjeras, atskiriantis apatinius audinius nuo aplinką. Pirminius augalo sluoksnius sudaro tik gyvos ląstelės. Antrinis ir tretinis apvalkalas – daugiausia iš mirusiųjų su storomis ląstelių sienelėmis.
Pagrindinės vientisųjų audinių funkcijos:
augalo apsauga nuo išdžiūvimo;
· Apsauga nuo patekimo kenksmingų mikroorganizmų;
· gynyba nuo saulės nudegimas;
· gynyba nuo mechaniniai pažeidimai;
medžiagų apykaitos tarp augalo ir aplinkos reguliavimas;
dirginimo suvokimas.
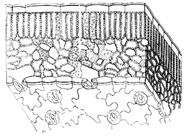
Pirminis vientisas audinys yra epidermis, epidermis. Susideda iš gyvų ląstelių. Susidaro iš viršūninių meristemų. Apima jaunus augančius stiebus ir lapus.
Epidermis susiformavo augaluose, kai jie patenka iš vandens buveinių į sausumą, kad jis neišdžiūtų. Be stomos, visos epidermio ląstelės yra glaudžiai tarpusavyje susijusios. Išorinės pagrindinių ląstelių sienelės yra storesnės nei likusios. Visas paviršius padengtas kutino ir augalinio vaško sluoksniu. Šis sluoksnis vadinamas odele (oda). Jo nėra ant augančių šaknų ir povandeninių augalų dalių. Išdžiūvus gerokai susilpnėja odelės pralaidumas.
Be pagrindinių ląstelių, epidermyje yra ir kitų, ypač plaukelių arba trichomų. Jie yra vienaląsčiai ir daugialąsčiai (2 pav.). Funkciškai jie padidina epidermio paviršių, pavyzdžiui, šaknų augimo zonoje, tarnauja kaip mechaninė apsauga, prilimpa prie atramos, mažina vandens praradimą. Nemažai augalų turi liaukų plaukelius, pavyzdžiui, dilgėlės.
Tik aukštesni augalai epidermyje turi stomatus, kurios reguliuoja vandens ir dujų mainus. Jeigu odelės nėra, vadinasi, ir stomatos nereikia. Stomatai yra ląstelių grupė, sudaranti stomatinį aparatą, susidedantį iš dviejų užpakalinių ląstelių ir greta esančių epidermio ląstelių - šoninių ląstelių. Jie skiriasi nuo pagrindinių epidermio ląstelių (3 pav.). Apsauginės ląstelės skiriasi nuo aplinkinių ląstelių forma ir daugybe chloroplastų bei netolygiai sustorėjusių sienelių. Tie, kurie yra vienas prieš kitą, yra storesni nei kiti (4 pav.). Tarp apsauginių ląstelių susidaro stomatalinis tarpas, kuris veda į substomatalinę erdvę, vadinamą substomataline ertme. Apsauginės ląstelės yra labai fotosintetinės. Juose yra didelis skaičius atsarginis krakmolas ir daug mitochondrijų.
Stomatų skaičius ir pasiskirstymas, stomatalinių aparatų tipai skirtinguose augaluose labai skiriasi. Šiuolaikiniuose briofituose stomatų nėra. Fotosintezę juose vykdo gametofitų karta, o sporofitai negali egzistuoti savarankiškai.
Paprastai stomos yra apatinėje lapo pusėje. Vandens paviršiuje plūduriuojančiuose augaluose – viršutiniame paviršiuje. Javų lapuose stomatai dažnai būna vienodai iš abiejų pusių. Tokie lapai apšviečiami palyginti tolygiai. Ant 1 mm 2 paviršiaus gali būti nuo 100 iki 700 stomatų.
Antrinis vidinis audinys (peridermas). Šis audinys pakeičia epidermį, kai žalia spalva metinius ūglius pakeičia rudi. Jis yra daugiasluoksnis ir susideda iš centrinio kambinių ląstelių sluoksnio – felogeno. Felogeno ląstelės, besidalijančios, kloja į išorę sluoksnį iš fellemo, o į vidų – felodermą (5 pav.).
Phelema arba kamštiena. Iš pradžių jis susideda iš gyvų plonasienių ląstelių. Laikui bėgant jų sienelės impregnuojamos suberinu ir augaliniu vašku ir miršta. Ląstelės turinys užpildytas oru.
Fellema funkcijos:
Apsaugo nuo drėgmės praradimo
apsaugo augalą nuo mechaninių pažeidimų;
Apsaugo nuo patogenų
užtikrina šilumos izoliaciją, nes ląstelės užpildomos oru.
Felogeno ląstelės, esančios pačiame epidermyje, apatiniame poepiderminiame sluoksnyje, rečiau giliuose pirminės žievės sluoksniuose, yra pirminės žievės pagrindas.
Kamštienos sluoksnis nėra nuolatinis. Joje atsiranda lūžių, kurios susisiekia su šalia esančiomis tarpląstelinėmis erdvėmis. Tuo pat metu paviršiuje formuojasi nedideli gumbai – lenticeliai, kurie atmosferos oru komunikuoja tarpląstelinių erdvių erdves (6.7 pav.).
Rudenį po lęšiais esantis felogenas kloja užkamšytų ląstelių sluoksnį, kuris labai sumažina transpiraciją, bet visiškai jos neatmeta. Pavasarį šis sluoksnis suyra iš vidaus. Ant šviesios beržo žievės lęšiai aiškiai matomi tamsių brūkšnelių pavidalu.
Tretinis vientisas audinys (pluta) taip pat būdingas tik sumedėjusioms augalų formoms.
Felogenas pakartotinai nusėda gilesniuose žievės sluoksniuose. Už jos ribų esantys audiniai ilgainiui miršta ir susidaro pluta. Jo ląstelės yra negyvos ir negali ištempti. Tačiau gyvos ląstelės, esančios giliau, dalijasi, todėl padidėja skersinis stiebo dydis. Laikui bėgant, išorinis plutos sluoksnis lūžta. Tokio tarpo atsiradimo laikas yra gana pastovi atskirų augalų reikšmė. Obelėje tai atsitinka septintaisiais gyvenimo metais, skroblėje - penkiasdešimtaisiais. Kai kuriose rūšyse jis visai nevyksta. Pagrindinė pilingo funkcija yra apsauga nuo mechaninių ir terminių pažeidimų.
Parenchima
Tai specializuotų audinių grupė, užpildanti augalo kūno viduje esančias erdves tarp laidžių ir mechaninių audinių (8 pav.). Dažniau parenchimos ląstelės yra suapvalintos, rečiau pailgos formos. Būdingas išsivysčiusių tarpląstelinių erdvių buvimas. Tarpai tarp ląstelių kartu sudaro transporto sistemą – apoplastą. Be to, tarpląstelinės erdvės sudaro augalo „vėdinimo sistemą“. Per stomatas arba lenticelius jie yra sujungti su atmosferos oru ir užtikrina optimalų dujų sudėtis augalo viduje. Išvystytos tarpląstelinės erdvės ypač reikalingos augalams, augantiems užmirkusioje dirvoje, kur sunku normaliai keistis dujomis. Ši parenchima vadinama aerenchima (9 pav.).
Parenchimos elementai, užpildantys tarpus tarp kitų audinių, atlieka ir atramos funkciją. Parenchimos ląstelės yra gyvos, jos neturi storų ląstelių sienelių, kaip sklerenchima. Todėl mechanines savybes suteikia turgoras. Jei vandens kiekis sumažėja, tai sukelia plazmolizę ir augalo vytimą.
Asimiliacinę parenchimą sudaro plonasienės ląstelės, turinčios daug tarpląstelinių erdvių. Šios struktūros ląstelėse yra daug chloroplastų, todėl ji vadinama chlorenchima. Chloroplastai išsidėstę išilgai sienos, vienas kito neužtemdydami. Asimiliacinėje parenchimoje vyksta fotosintezės reakcijos, kurios aprūpina augalą organinėmis medžiagomis ir energija. Fotosintezės procesų rezultatas – galimybė Žemėje egzistuoti visiems gyviems organizmams.
Asimiliaciniai audiniai yra tik apšviestose augalo dalyse, juos nuo aplinkos skiria skaidrus epidermis. Jei epidermį pakeičia nepermatomi antriniai integumentiniai audiniai, asimiliacinė parenchima išnyksta.
Sandėliavimo parenchima tarnauja kaip organinių medžiagų, kurių augalų organizmas laikinai nenaudoja, talpykla. Iš esmės atidėti organinės medžiagos kaip Įvairios rūšys inkliuzai yra pajėgūs bet kuriai ląstelei su gyvu protoplastu, tačiau kai kurios ląstelės tam specializuojasi (10, 11 pav.). Energetiniai junginiai nusėda tik vegetacijos metu, vartojami ramybės laikotarpiu ir ruošiantis kitam vegetacijos sezonui. Todėl rezervinės medžiagos vegetatyviniuose organuose nusėda tik daugiamečiuose augaluose.
Atsargų talpykla gali būti paprasti organai (pabėgimas, šaknis), taip pat specializuoti (šakniastiebiai, gumbai, svogūnėliai). Visi sėkliniai augalai kaupia energetiškai vertingas medžiagas sėklose (skilčialapiuose, endosperme). Daugelis sausringo klimato augalų kaupia ne tik organines medžiagas, bet ir vandenį (12 pav.). Pavyzdžiui, alavijas kaupia vandenį mėsinguose lapuose, kaktusai – ūgliuose.
mechaniniai audiniai
Mechanines augalų ląstelių savybes suteikia:
Kietas ląstelės apvalkalas
turgoriškumas, tai yra ląstelių turgorinė būsena.
Nepaisant to, kad beveik visos audinių ląstelės turi mechaninių savybių, tačiau augale yra audinių, kurių mechaninės savybės yra pagrindinės. Tai yra kolenchima ir sklerenchima. Paprastai jie veikia sąveikaudami su kitais audiniais. Kūno viduje augalai sudaro savotišką rėmą. Todėl jie vadinami sustiprinimu.
Ne visi augalai yra vienodai gerai išreikšti mechaniniai audiniai. Daug mažiau augalų, gyvenančių vandens aplinka nei antžeminės. Priežastis ta vandens augalai reikia mažiau vidinės paramos. Jų kūną daugiausia palaiko aplinkinis vanduo. Oras sausumoje tokios atramos nesukuria, nes jo tankis mažesnis nei vandens. Dėl šios priežasties specializuotų mechaninių audinių buvimas tampa aktualus.
Evoliucijos procese įvyko vidinių atraminių struktūrų tobulėjimas.
Kolenchima. Jį sudaro tik gyvos ląstelės, pailgos išilgai organo ašies. Šio tipo mechaniniai audiniai susidaro labai anksti, pirminio augimo laikotarpiu. Todėl svarbu, kad ląstelės išliktų gyvos ir išlaikytų galimybę ištempti kartu su šalia esančiomis tempimo ląstelėmis.
Kolenchimos ląstelių savybės:
Netolygus lukšto sustorėjimas, dėl kurio vienos jo dalys lieka plonos, o kitos sustorėja;
lukštai nerūgsta.
Kolenchimos ląstelės yra skirtingos viena kitos atžvilgiu. Gretimose ląstelėse ant vienas į kitą atsuktų kampų susidaro sustorėjimai (13 pav.). Tokia kolenchima vadinama kampine kolenchima. Kitu atveju ląstelės yra išdėstytos lygiagrečiais sluoksniais. Ląstelių membranos, esančios priešais šiuos sluoksnius, yra stipriai sustorėjusios. Tai yra lamelinė kolenchima. Ląstelės gali išsidėstyti laisvai, su gausiomis tarpląstelinėmis erdvėmis – tai laisva kolenchima. Tokia kolenchima dažnai aptinkama augaluose užmirkusiose dirvose.
Kolenchima ypač svarbi jauniems augalams, žolinėms formoms, taip pat tose augalų dalyse, kuriose antrinis augimas nevyksta, pavyzdžiui, lapuose. Šiuo atveju jis klojamas labai arti paviršiaus, kartais tiesiog po epidermiu. Jei organas turi veidus, tada išilgai jų keteros randami stori kolenchimo sluoksniai.
Kolenchimos ląstelės veikia tik esant turgorui. Vandens trūkumas sumažina kolenchimo efektyvumą ir augalas laikinai nuvysta, pavyzdžiui, karštą dieną nusvyra agurkų lapai. Užpildžius ląsteles vandeniu, atkuriamos kolenchimos funkcijos.
Sklerenchima. Antrasis mechaninių audinių tipas. Skirtingai nuo kolenchimos, kur visos ląstelės yra gyvos, sklerenchimos ląstelės yra negyvos. Jų sienos labai storos. Jie atlieka mechaninę funkciją. Dėl stipraus membranos sustorėjimo sutrinka medžiagų pernešimas, dėl to protoplastas žūva (14 pav.). Sklerenchimos ląstelių membranų lignifikacija įvyksta, kai augalo organas jau baigė augti. Todėl jie nebetrukdo aplinkinių audinių tempimui.
Priklausomai nuo formos, išskiriami du sklerenchimos ląstelių tipai – skaidulinės ir sklereidinės.
Pluoštai yra stipriai pailgos formos su labai storomis sienelėmis ir maža ertme. Jie yra šiek tiek mažesni nei medienos pluoštai. Dažnai po epidermiu susidaro išilginiai sluoksniai ir sruogos. Floemoje arba ksileme juos galima rasti pavieniui arba grupėmis. Floemoje jie vadinami bastiniais pluoštais (15 pav.), ksileme – libriforminiais pluoštais (16 pav.).
Sklereidai arba akmeninės ląstelės yra suapvalintos arba šakotos ląstelės su galingomis membranomis. Augalo kūne jų galima rasti pavieniui (atraminės ląstelės) (17 pav.) arba grupėmis (18 pav.). Reikia pažymėti, kad mechaninės savybės labai priklauso nuo sklereidų vietos. Dalis sklereidų sudaro ištisinius sluoksnius, pavyzdžiui, riešutų kevaluose arba vaisių (kaulavaisių) kauliukuose.
išskyrimo audiniai
Bet koks gyva ląstelė iš pradžių atlieka medžiagų pašalinimo funkciją, o transportavimas gali vykti tiek palei koncentracijos gradientą, tiek prieš gradientą. Medžiagos, kurios buvo susintetintos ląstelėje, gali būti pašalintos ir vėliau paveiks kitų ląstelių veiklą (fitohormonai, fermentai). Šis procesas vadinamas sekrecija. Tuo atveju, kai išskiriamos medžiagos, kurios yra ląstelės atliekos, procesas vadinamas išskyrimu. Nepaisant to, kad medžiagų išskyrimas į augalų ir gyvūnų ląsteles turi esminį panašumą, augalai turi nemažai savybių, atsirandančių dėl esminių gyvybinės veiklos skirtumų. Metabolinių reakcijų lygis augaluose yra daug mažesnis nei gyvūnų. Todėl išmetama proporcingai mažesni atliekų kiekiai. Kitas bruožas yra tai, kad savarankiškai sintezuoja beveik viską, ko reikia organiniai junginiai, augalai niekada nesudaro perteklinių atsargų. Išsiskyrusios medžiagos gali būti pradinė medžiaga kitoms reakcijoms (pavyzdžiui, CO 2 ir H 2 O).
Jei gyvūnams toksinų išskyrimo procesas yra susijęs su jų pašalinimu iš organizmo, augalai gali to nedaryti, išskirdami nereikalingas medžiagas gyvame protoplaste (įvairių medžiagų pašalinimas į vakuolę), negyvose ląstelėse (daugumoje audinių). daugiametis augalas susideda iš tokių ląstelių), tarpląstelinėse erdvėse.
Išsiskyrusias medžiagas galima suskirstyti į dvi dalis didelės grupės. Pirmoji grupė yra organinės medžiagos, kurias tiesiogiai sintezuoja ląstelė (fermentai, polisacharidai, ligninai, terpenai, pastarieji yra sudedamieji elementai eteriniai aliejai ir derva). Antroji grupė - medžiagos, kurios iš pradžių patenka į ląstelę iš išorės ksilemo ar floemo transportavimo pagalba (vanduo, mineralinės druskos, aminorūgštys, monocukrūs ir kt.). Išskirta paslaptis retai būna vienalytė ir dažniausiai susideda iš mišinio, kuriame vienos medžiagos koncentracija yra didžiausia.
Augalams, skirtingai nei gyvūnams, trūksta pilno išskyrimo sistema. Yra tik specializuotos struktūros, išsibarsčiusios visame augale - idioblastai. Jas formuojančios ląstelės yra mažesnės nei šalia esančios parenchimos ląstelės. Jie turi elektronų tankią citoplazmą su išvystytais endoplazminio tinklo elementais ir Golgi kompleksu. Dažniausiai neišreiškiama centrinė vakuolė. Tarp jų ir kitų gyvų ląstelių šios ląstelės yra sujungtos daugybe plazmodesmų.
Priklausomai nuo vietos, šalinimo audiniai gali būti išoriniai ir vidiniai.
Išoriniai šalinimo audiniai. Liaukiniai plaukai yra epidermio dariniai. Morfologiškai jie yra kintami – gali turėti daugialąstę galvą, būti pailgos, skydo formos ant kojos ir kt. (19 pav.). Tai apima dilgėlių plaukelius.
Hidatodai – tai konstrukcijos, išskiriančios vandens perteklių esant žemai transpiracijai ir didelės drėgmės sąlygoms – gutacija. Šias struktūras sudaro bespalvių gyvų ląstelių grupės su plonomis sienelėmis – epitema. Šis audinys yra greta laidžiojo pluošto (20 pav.). Vanduo išleidžiamas per specialias vandens stomatas, kurios nuo įprastų stomatų skiriasi nejudrumu ir nuolat atsiveriančiu tarpu. Gutacijos skysčio sudėtis labai skiriasi nuo beveik Tyras vanduoį labai sudėtingą medžiagų mišinį.
Nektarai. Daugeliui augalų būdingas skysčio, kuriame yra nuo 7 iki 87% mono- ir oligosacharidų, išsiskyrimas. Šį procesą vykdo specialios struktūros – nektarai. Priklausomai nuo vietos, žiedinės ar žiedinės, skiriami nektarai (21 pav.), taip pat augalai, esantys ant stiebų ir lapų – ekstrafloriniai, arba ekstrafloriniai nektarai. Nektaras gali būti nepakitusios floemo sultys, kurios per tarpląstelinius tarpus patenka į paviršių ir išskiriamos per stomatas. Sudėtingesnius nektarus sudaro liaukų parenchima, padengta epidermiu su liaukiniais plaukais. Nektarą išskiria epidermio ląstelės arba liaukų plaukai. Šiuo atveju išsiskiriantis nektaras skiriasi nuo floemų sulčių. Kadangi jame vyrauja gliukozė, fruktozė, sacharozė, o floemo sultyse – gliukozė. Jonų yra nedidelėmis koncentracijomis. Jonų yra nedidelėmis koncentracijomis. Kad pritrauktų apdulkintojus, nektare gali būti steroidiniai hormonai kurios būtinos vabzdžiams.
Virškinimo liaukos yra vabzdžiaėdžiuose augaluose (saulėraščiuose, nepentiuose ir kt.). Reaguodami į mažų bestuburių prisilietimą, jie išskiria paslaptį, kurioje yra hidrolizinių fermentų, skruzdžių, sviesto ir propiono rūgščių. Dėl to biopolimerai suskaidomi į monomerus, kuriuos augalas naudoja savo reikmėms.
Druskos liaukos vystosi augaluose, augančiuose druskingoje dirvoje. Šios liaukos yra ant lapų, o ištirpusios formos jonų perteklius iškeliamas į paviršių. Druska pirmiausia nusėda ant odelės, vėliau ją nuplauna lietus (22 pav.). Be to, daugelio augalų lapuose yra druskos plaukelių. Kiekvienas plaukas susideda iš dviejų ląstelių: viena sudaro galvą, kita - stiebą. Druskos kaupiasi galvos ląstelės vakuolėje, o kai jų koncentracija tampa didelė, galva nukrenta. Jo vietoje išauga nauja ląstelė, kuri taip pat kaupia druskas. Lapui augant gali kelis kartus nukristi ir formuotis galvutė. Druskos plaukų privalumas yra tas, kad išskirdami druskas jie netenka mažai vandens. Šis reiškinys labai svarbus augalams, augantiems druskingose dirvose, kur gėlo vandens trūksta.
Vidiniai šalinimo audiniai. Vidinės šalinimo struktūros, išsibarsčiusios po visą augalo kūną idioblastų pavidalu, nepašalina medžiagų iš organizmo, kaupdamos jas savyje. Idioblastai yra augalų ląstelės, kurios savo forma, struktūra ar turiniu skiriasi nuo kitų to paties audinio ląstelių. Priklausomai nuo kilmės skiriamos šizogeninės (gr. schiso – suskaidytos) ir lizogeninės (gr. lizė – tirpimas) išskiriamų medžiagų talpyklos (23 pav.).
Šizogeninės talpyklos. Tai daugiau ar mažiau plačios tarpląstelinės erdvės, užpildytos išskiriamomis medžiagomis. Pavyzdžiui, spygliuočių, aralijų, skėčių ir kt. Manoma, kad derva turi baktericidinės savybės, atbaido žolėdžius, todėl augalai jiems nevalgomi.
Vietoje gyvų ląstelių susidaro lizigeninės talpyklos, kurios jose susikaupus medžiagoms žūva ir sunaikinamos. Tokius darinius galima pastebėti citrusinių vaisių žievelėse.
Be pagrindinių tipų, yra tarpinių formų. Melžėjai būna dviejų tipų: šarnyriniai ir nešarnyriniai. Pirmieji susidaro iš daugybės gyvų ląstelių, išdėstytų grandinėmis. Sąlyčio vietoje apvalkalai sunaikinami, protoplastai susilieja ir dėl to susidaro vientisas tinklas. Tokių pieniškų yra Compositae. Nesegmentuotas pieno ląsteles sudaro milžiniška daugiabranduolė ląstelė. Jis atsiranda ankstyvosiose vystymosi stadijose, pailgėja ir šakojasi augalui augant, sudarydamas sudėtingą tinklą, pavyzdžiui, euforbiniuose augaluose (25 pav.).
Pieninių augalų ląstelių sienelės yra stipriai laistomos. Jie nerūgsta, todėl yra plastikiniai. Centrinę lakto dalį užima vakuolė su lateksu, o protoplastas – netoli sienos. Ne visada galima nustatyti ribą tarp protoplasto ir vakuolės. Pienžolės prasiskverbia į meristemą ir auga kartu su augalo organu.
Laidūs audiniai
Šis tipas reiškia sudėtingus audinius, susideda iš skirtingai diferencijuotų ląstelių. Be faktinių laidžių elementų, audinyje yra mechaninių, šalinimo ir saugojimo elementų (26 pav.). Laidūs audiniai sujungia visus augalų organus į vieną sistemą. Yra dviejų tipų laidūs audiniai: ksilemas ir floemas (graikiškai ksilonas – medis; phloios – žievė, bastas). Jie turi tiek struktūrinių, tiek funkcinių skirtumų.
Laidžius ksilemo elementus sudaro negyvos ląstelės. Jie perneša tolimais atstumais vandenį ir jame ištirpusias medžiagas nuo šaknų iki lapų. Laidūs floemo elementai palaiko protoplastą gyvą. Jie perneša didelius atstumus nuo fotosintetinių lapų iki šaknų.
Paprastai ksilemas ir floemas yra augalo kūne tam tikra tvarka, formuojant sluoksnius arba laidžius ryšulius. Priklausomai nuo struktūros, išskiriami keli laidžių ryšulių tipai, kuriems būdingi tam tikros grupės augalai. Šalutiniame atvirame ryšulyje tarp ksilemo ir floemo yra kambis, užtikrinantis antrinį augimą (27-A, 28 pav.). Dvišaliame atvirame ryšulyje floema yra abiejose ksilemo pusėse (27-B, 29 pav.). Uždaruose ryšuliuose kambio nėra, todėl jie negali antrinio sutirštėjimo (27-B, 27-D, 30.31 pav.). Galima rasti dar du koncentrinių ryšulių tipus, kur arba floema supa ksilemą (27-E, 32 pav.), arba ksilemas supa floemą (27-E pav.).
Ksilemas (mediena). Ksilemo vystymasis aukštesniuose augaluose yra susijęs su vandens mainų užtikrinimu. Kadangi vanduo nuolat išsiskiria per epidermį, augalas turi sugerti tą patį drėgmės kiekį ir įpilti į transpiraciją vykdančius organus. Reikėtų atsižvelgti į tai, kad gyvo protoplasto buvimas vandeniui laidžiose ląstelėse labai sulėtintų transportavimą, o negyvos ląstelės čia yra funkcionalesnės. Tačiau negyva ląstelė neturi standumo, todėl membrana turi turėti mechaninių savybių. Pastaba: turgescencija yra augalų ląstelių, audinių ir organų būsena, kai jie tampa elastingi dėl ląstelių turinio spaudimo ant jų elastinių membranų. Iš tiesų, laidžiuosius ksilemo elementus sudaro negyvos ląstelės, ištemptos išilgai organo ašies su storais lignifikuotais apvalkalais.
Iš pradžių ksilemas susidaro iš pirminės meristemos – prokambio, esančio ašinių organų viršūnėse. Pirmiausia skiriasi protoksilema, po to metaksilema. Yra žinomi trys ksilemų susidarymo tipai. Eksarcho tipo protoksilemos elementai pirmiausia atsiranda prokambio pluošto periferijoje, tada metaksilemos elementai atsiranda centre. Jei procesas vyksta priešinga kryptimi (ty nuo centro iki periferijos), tai yra endarchinis tipas. Mesarchinio tipo ksilemas klojamas prokambio pluošto centre, po to jis nusėda tiek link centro, tiek į periferiją.
Šaknims būdingas eksarchinis ksilemo formavimosi tipas, o stiebams – endarchinis. Žemai organizuotuose augaluose ksilemo formavimo būdai yra labai įvairūs ir gali būti sistemingi.
Kai kuriuose augaluose (pavyzdžiui, vienaląsčiuose) visos prokambio ląstelės diferencijuojasi į laidžius audinius, kurie negali antrinio sustorėjimo. Kitose formose (pavyzdžiui, sumedėjusios) tarp ksilemo ir floemo lieka šoninės meristemos (kambis). Šios ląstelės gali dalytis, atnaujindamos ksilemą ir floemą. Šis procesas vadinamas antriniu augimu. Daugelis augalų, augančių palyginti stabiliomis klimato sąlygomis, auga nuolat. Prie sezoninių klimato pokyčių pritaikytomis formomis – periodiškai. Dėl to susidaro aiškiai apibrėžti metiniai augimo žiedai.
Pagrindiniai prokambinių ląstelių diferenciacijos etapai. Jo ląstelės turi plonas membranas, kurios netrukdo joms ištempti organui augant. Tada protoplastas pradeda kloti antrinę membraną. Tačiau šis procesas turi ryškių bruožų. Antrinė membrana nusėda ne ištisiniu sluoksniu, kuris neleistų ląstelei išsitempti, o žiedų pavidalu arba spirale. Ląstelių pailgėjimas nėra sunkus. Jaunose ląstelėse spiralės žiedai arba ritės yra arti vienas kito. Brandžiose ląstelėse jos išsiskiria dėl ląstelių tempimo (33 pav.). Žieduoti ir spiraliniai lukšto sustorėjimai netrukdo augti, tačiau mechaniškai jie yra prastesni už kiautus, kur antrinis sustorėjimas sudaro vientisą sluoksnį. Todėl, nustojus augti ksileme, susidaro elementai su ištisiniu lignifikuotu apvalkalu (metaksilemas). Pažymėtina, kad antrinis sustorėjimas čia ne žiedinis ar spiralinis, o taškuotas, laiptiškas, tinklinis (34 pav.). Jo ląstelės nepajėgia ištempti ir miršta per kelias valandas. Šis procesas netoliese esančiose ląstelėse vyksta koordinuotai. Citoplazmoje atsiranda daug lizosomų. Tada jie suyra, o juose esantys fermentai sunaikina protoplastą. Sunaikinus skersines sienas, viena virš kitos grandinėje išsidėsčiusios ląstelės suformuoja tuščiavidurį indą (35 pav.). Dauguma gaubtasėkliai o kai kurie paparčiai turi indus.
Laidi ląstelė, kuri nesusidaro per perforacijas jos sienelėje, vadinama tracheide. Vandens judėjimas tracheidėmis yra lėtesnis nei per indus. Faktas yra tas, kad tracheidose pirminė membrana niekur nenutrūksta. Tracheidės susisiekia viena su kita per poras. Reikėtų paaiškinti, kad augaluose pora yra tik antrinės membranos įdubimas iki pirminės membranos, o tarp tracheidų nėra perforacijų.
Dažniausios yra ribojamos poros (35-1 pav.). Jų kanalas, nukreiptas į ląstelės ertmę, sudaro pratęsimą - porų kamerą. Daugumos spygliuočių augalų poros ant pirminio apvalkalo turi sustorėjimą – torą, kuris yra savotiškas vožtuvas ir gali reguliuoti vandens transportavimo intensyvumą. Pasislinkdamas toras blokuoja vandens tekėjimą per poras, tačiau po to nebegali grįžti į ankstesnę padėtį, atlikdamas vienkartinį veiksmą.
Poros yra daugiau ar mažiau suapvalintos, pailgos statmenai pailgai ašiai (šių porų grupė primena kopėčias, todėl toks poringumas vadinamas kopėčių poringumu). Transportavimas per poras atliekamas tiek išilgine, tiek skersine kryptimis. Poros yra ne tik tracheidėse, bet ir atskirose kraujagyslių ląstelėse, kurios sudaro indą.
Evoliucijos teorijos požiūriu, tracheidai yra pirmoji ir pagrindinė struktūra, kuri veda vandenį aukštesniųjų augalų kūne. Manoma, kad kraujagyslės atsirado iš tracheidų dėl tarp jų esančių skersinių sienelių lizės (36 pav.). Dauguma paparčių ir gimnasėklių neturi. Jie perkelia vandenį per tracheidą.
Vykstant evoliuciniam vystymuisi, indai atsirado skirtingos grupės augalų pakartotinai, tačiau svarbiausią funkcinę reikšmę jie įgijo gaubtasėkliuose, turinčiuose juos kartu su tracheidėmis. Manoma, kad pažangesnio transporto mechanizmo turėjimas padėjo jiems ne tik išgyventi, bet ir pasiekti didelę formų įvairovę.
Ksilemas yra sudėtingas audinys, be vandeniui laidžių elementų, jame yra ir kitų. Mechanines funkcijas atlieka libriforminiai pluoštai (lot. liber – bastas, forma – forma). Papildomų mechaninių konstrukcijų buvimas yra svarbus, nes, nepaisant sustorėjimo, vandenį nešančių elementų sienelės vis dar yra per plonos. Jie nesugeba savęs išlaikyti didelė masė daugiametis augalas. Pluoštai išsivystė iš tracheidų. Jiems būdingi mažesni dydžiai, lignified (lignified) apvalkalai ir siauros ertmės. Ant sienos galima rasti porų be apvadų. Šios skaidulos nepraleidžia vandens, jų pagrindinė funkcija yra atrama.
Ksileme taip pat yra gyvų ląstelių. Jų masė gali siekti 25% viso medienos tūrio. Kadangi šios ląstelės turi apvalią formą, jos vadinamos medienos parenchima. Augalo kūne parenchima yra dviem būdais. Pirmuoju atveju ląstelės yra išdėstytos vertikalių virvelių pavidalu - tai yra laido parenchima. Kitu atveju parenchima formuoja horizontalius spindulius. Jie vadinami šerdies spinduliais, nes jungia šerdį ir žievę. Šerdis atlieka daugybę funkcijų, įskaitant medžiagų saugojimą.
Floemas (batas). Tai sudėtingas audinys, nes jį sudaro nevienalytės ląstelės. Pagrindinės laidžiosios ląstelės vadinamos sieto elementais (37 pav.). Laidžius ksilemo elementus sudaro negyvos ląstelės, o floeme per funkcionavimo laikotarpį jie išsaugo gyvą, nors ir labai pakitusį protoplastą. Floema atlieka plastikinių medžiagų nutekėjimą iš fotosintezės organų. Visos gyvos augalų ląstelės turi savybę pravesti organines medžiagas. Vadinasi, jei ksilemą galima rasti tik aukštesniuose augaluose, tai organinių medžiagų pernešimas tarp ląstelių taip pat vyksta žemesni augalai.
Ksilemas ir floema išsivysto iš viršūninių meristemų. Pirmajame etape prokambalinėje virvelėje susidaro protoflomas. Augant aplinkiniams audiniams jis išsitempia, o augimui pasibaigus vietoj protofloemos susidaro metafloema.
At įvairios grupės Aukštesniuose augaluose galima rasti dviejų tipų sieto elementus. Paparčiuose ir gimnasėkliuose jį vaizduoja sieto ląstelės. Sietų laukai ląstelėse yra išsibarstę palei šonines sieneles. Protoplastas išlaiko šiek tiek sunaikintą branduolį.
Gaubtasėkliuose sieto elementai vadinami sieto vamzdeliais. Jie bendrauja tarpusavyje per sieto plokštes. Brandžios ląstelės neturi branduolių. Tačiau šalia sieto vamzdelio yra kompanioninė ląstelė, kuri susidaro kartu su sieto vamzdeliu dėl bendros motininės ląstelės mitozinio dalijimosi (38 pav.). Kompanioninė ląstelė turi tankesnę citoplazmą su daugybe aktyvių mitochondrijų, taip pat pilnai funkcionuojantį branduolį, puiki suma plazmodesma (dešimt kartų daugiau nei kitos ląstelės). Kompanioninės ląstelės veikia nebranduolinių sieto vamzdelių ląstelių funkcinį aktyvumą.
Subrendusių sieto ląstelių struktūra turi tam tikrų ypatumų. Vakuolės nėra, todėl citoplazma labai suskystinta. Branduolys gali nebūti (sėkmiuose) arba gali būti susiraukšlėjęs, funkciškai neaktyvus. Ribosomų ir Golgi komplekso taip pat nėra, tačiau gerai išvystytas endoplazminis tinklas, kuris ne tik prasiskverbia į citoplazmą, bet ir per sieto laukų poras patenka į kaimynines ląsteles. Gausiai randama gerai išsivysčiusių mitochondrijų ir plastidžių.
Tarp ląstelių medžiagų transportavimas vyksta per skylutes, esančias ant ląstelių membranų. Tokios angos vadinamos poromis, tačiau skirtingai nei tracheidinės poros, jos yra pro. Daroma prielaida, kad tai labai išsiplėtusios plazmodesmos, ant kurių sienelių nusėda kaliozės polisacharidas. Poros išsidėsčiusios grupėmis, suformuodamos sieto laukus. Primityviose formose sieto laukai atsitiktinai išsibarstę po visą apvalkalo paviršių, labiau išsivysčiusiuose gaubtasėkliuose išsidėstę gretimų ląstelių galuose, besiribojančiose viena su kita, suformuodami sieto plokštę (39 pav.). Jei ant jo yra vienas sieto laukas, jis vadinamas paprastu, jei keli - kompleksiniu.
Tirpalų judėjimo per sieto elementus greitis yra iki 150 cm ∕ val. Tai tūkstantį kartų viršija laisvosios difuzijos greitį. Tikriausiai vyksta aktyvus transportas, o daugybė sieto elementų ir palydovinių ląstelių mitochondrijų tiekia tam reikalingą ATP.
Floemo sieto elementų tarnavimo laikas priklauso nuo šoninių meristemų buvimo. Jei jų yra, tada sieto elementai veikia visą augalo gyvenimą.
Be sieto elementų ir palydovinių ląstelių, floemoje yra karnienos pluoštų, sklereidų ir parenchimos.
Klausimai kontrolei ir kartojimui
Aukštesnių augalų ženklai? Kokios kartos aukštesni augalai vadinami gametofitais? Sporofitas?
Kokios priežastys prisideda prie diferencijuotų sausumos augalų audinių vystymosi.
Išvardykite augalų rūšis.
Kokias meristemų grupes žinote? Kur jie yra augalo kūne?
Pilnamieji audiniai, jų funkcijos, reikšmė.
Organizacija įvairių tipų dengiančius audinius.
Stomatalinio aparato sandara. Išvardykite jums žinomų stomatalinių aparatų tipus. Kaip veikia stomos?
Parenchiminių audinių ypatumai, parenchimo tipai.
Parenchimo ir sklerenchimo požymiai.
Išvardykite augalo išskyrimo struktūrų tipus. Kokia jų reikšmė?
Išskiriamų medžiagų talpyklų formavimo metodai.
Kas yra melžėjai? Į kokias rūšis jie skirstomi?
Kaip organizuojami laidūs audiniai?
Kas yra laidūs pluoštai? Jų rūšys.
Medžiagų judėjimo per ksilemą kryptys? Floemas? Ar transportuojamų medžiagų sudėtis skiriasi ksilemu ir floemu?
Ksilemo, floemo sandara ir funkcijos.
Laidžių floemo elementų organizavimas augaluose iš įvairių sisteminės grupės.
Kaip medžiagos pernešamos floemoje?
Su jais susiję papildomi baltymai, vadinamieji MAP baltymai. Šie baltymai, stabilizuodami mikrovamzdelius, pagreitina tubulino polimerizacijos procesą. Mikrofilamentų struktūra ir funkcijos Mikrofilamentai yra labai plonos ir ilgos siūlinės baltymų struktūros, randamos visoje citoplazmoje. Po plazmine membrana mikrofilamentai sudaro ištisinį rezginį, sudarydami ...

Kaip, pavyzdžiui, krapai, morkos, ropės ir kt., daugeliu atvejų jie turi tą patį audinį, kurį ką tik nurodžiau kamštelyje. Taigi augalo ląstelė buvo atrasta pirmą kartą. Tačiau Hooke'as ilgai negalėjo susitvarkyti su savo mikroskopu: jo galvoje knibždėte knibžda kitų išradimų idėjos (spyruokliniai laikrodžiai, patobulinti kompasai ir kt.), ir jis noriai perdavė tolesnę mikroskopinio ...
Bent šiek tiek supratimo apie tinkamus ląstelių tyrimo metodus. Šviesos mikroskopija Pats seniausias ir kartu labiausiai paplitęs ląstelių tyrimo metodas yra mikroskopija. Galima sakyti, kad ląstelės tyrimo pradžia buvo šviesos optinio mikroskopo išradimas. Neginkluotas žmogaus akis skiriamoji geba yra apie 1/10 mm. Šis...
Su daugybe ląstelių organelių jis suteikia ryšį tarp atskirų tarpląstelinių struktūrų. Lizosomos Lizosomos (iš lis- ir graik. soma – kūnas), gyvūnų ir augalų organizmų ląstelėse esančios struktūros, turinčios fermentų, galinčių skaidyti (ty lizuoti – iš čia ir pavadinimas) baltymus, polisacharidus, peptidus, nukleino rūgštis. Tai labai marga burbulų klasė, kurios dydis yra 0,1–0,4 mikrono, ...
Augalų ląstelės struktūra. Augalų audiniai
AUGALŲ LĄSTELĖS STRUKTŪRA
Augalo ląstelė susideda iš daugiau ar mažiau standžios ląstelės sienelės ir protoplasto. Ląstelės sienelė yra ląstelės sienelė ir citoplazminė membrana. Terminas protoplastas kilęs iš žodžio protoplazma, kuris ilgą laiką buvo vartojamas visiems gyviems dalykams apibūdinti. Protoplastas yra atskiros ląstelės protoplazma.
Protoplastas susideda iš citoplazmos ir branduolio. Citoplazmoje yra organelių (ribosomų, mikrotubulių, plastidžių, mitochondrijų) ir membraninių sistemų (endoplazminis tinklas, diktiosomos). Citoplazma taip pat apima citoplazminę matricą (pagrindinę medžiagą), į kurią panardinami organeliai ir membranų sistemos. Citoplazmą nuo ląstelės sienelės skiria plazminė membrana, kuri yra elementari membrana. Skirtingai nuo daugelio gyvūnų ląstelių, augalų ląstelėse yra viena ar daugiau vakuolių. Tai pūslelės, užpildytos skysčiu ir apsuptos elementaria membrana (tonoplastas).
Gyvoje augalo ląstelėje žemės medžiaga nuolat juda. Organelės dalyvauja judėjime, vadinamame citoplazmos srautu arba cikloze. Ciklozė palengvina medžiagų judėjimą ląstelėje ir jų mainus tarp ląstelės ir aplinkos.
Plazmos membrana. Tai dvisluoksnė fosfolipidinė struktūra. Augalų ląstelėms būdingos plazminės membranos invaginacijos.
Plazminė membrana atlieka šias funkcijas:
Dalyvauja medžiagų mainuose tarp ląstelės ir aplinkos;
Koordinuoja ląstelės sienelės celiuliozės mikrofibrilių sintezę ir surinkimą;
Perduoda hormoninius ir išorinius signalus, kurie kontroliuoja ląstelių augimą ir diferenciaciją.
Šerdis. Tai ryškiausia eukariotinės ląstelės citoplazmos struktūra. Branduolys atlieka dvi svarbias funkcijas:
Kontroliuoja gyvybinę ląstelės veiklą, nustatydama, kurie baltymai ir kokiu laiku turi būti sintetinami;
Saugo genetinę informaciją ir perduoda ją dukterinėms ląstelėms ląstelių dalijimosi metu.
Eukariotinės ląstelės branduolį supa dvi elementarios membranos, kurios sudaro branduolio apvalkalą. Jis persmelktas daugybe porų, kurių skersmuo nuo 30 iki 100 nm, matomos tik elektroniniu mikroskopu. Poros turi sudėtingą struktūrą. Branduolinės membranos išorinė membrana kai kuriose vietose yra sujungta su endoplazminiu tinklu. Branduolinis apvalkalas gali būti vertinamas kaip specializuota, lokaliai diferencijuota endoplazminio tinklo (ER) dalis.
Specialiais dažais nudažytame branduolyje galima išskirti plonus siūlus ir chromatino bei nukleoplazmos (pagrindinės branduolio medžiagos) gumulėlius. Chromatinas susideda iš DNR, susietos su specialiais baltymais, vadinamais histonais. Ląstelių dalijimosi procese chromatinas vis labiau sutankinamas ir surenkamas į chromosomas. DNR koduoja genetinę informaciją.
Organizmai skiriasi chromosomų skaičiumi somatinėse ląstelėse. Pavyzdžiui, kopūstai turi - 20 chromosomų; saulėgrąžos - 34; kviečiai - 42; žmonių – 46, o viena iš paparčio Ophioglossum rūšių – 1250. Lytinės ląstelės (gametos) turi tik pusę chromosomų, būdingų somatinėms kūno ląstelėms. Chromosomų skaičius gametose vadinamas haploidiniu (viengubu), somatinėse ląstelėse – diploidiniu (dvigubu). Ląstelės, turinčios daugiau nei du chromosomų rinkinius, vadinamos poliploidinėmis.
Šviesos mikroskopu matomos sferinės struktūros – branduoliai. Kiekvienas branduolys turi vieną ar daugiau branduolių, kurie matomi nesidalijančiuose branduoliuose. Ribosominė RNR sintetinama branduolyje. Paprastai diploidinių organizmų branduoliai turi du branduolius, po vieną kiekvienam haploidiniam chromosomų rinkiniui. Branduoliai neturi savo membranos. Biochemiškai branduoliams būdinga didelė RNR koncentracija, kuri čia susijusi su fosfoproteinais. Branduolių dydis priklauso nuo ląstelės funkcinės būklės. pastebėta, kad sparčiai augančioje ląstelėje, kurioje vyksta intensyvūs baltymų sintezės procesai, didėja branduolių dydis. Branduoliuose gaminasi mRNR ir ribosomos, kurios sintetinę funkciją atlieka tik branduolyje.
Nukleoplazmą (karioplazmą) vaizduoja vienalytis skystis, kuriame yra ištirpę įvairūs baltymai, įskaitant fermentus.
Plastidai. Vakuolės, celiuliozės ląstelių sienelės ir plastidai yra būdingi augalų ląstelių komponentai. Kiekviena plastidė turi savo apvalkalą, susidedantį iš dviejų elementarių membranų. Plastido viduje išskiriama membraninė sistema ir įvairaus laipsnio vienalytė medžiaga – stroma. Subrendusios plastidės klasifikuojamos pagal juose esančius pigmentus.
Chloroplastuose, kuriuose vyksta fotosintezė, yra chlorofilų ir karotinoidų. Paprastai turi 4–5 mikronų skersmens disko formą. Vienoje mezofilo ląstelėje (lapo viduryje) gali būti 40 - 50 chloroplastų; mm2 lapo – apie 500 000. citoplazmoje chloroplastai dažniausiai išsidėstę lygiagrečiai ląstelės membranai.
Vidinė chloroplasto struktūra yra sudėtinga. Stroma yra persmelkta išsivysčiusios membranų sistemos pūslelių pavidalu – tilakoidų. Kiekvienas tilakoidas susideda iš dviejų membranų. Tilakoidai sudaro vieną sistemą. Paprastai jie surenkami krūvomis – grūdeliais, primenančiais monetų stulpelius. Atskirų granatų tilakoidai yra tarpusavyje sujungti stromos tilakoidais arba tarpgranaliniais tilakoidais. Chlorofilas ir karotinoidai yra įterpti į tilakoidų membranas. Žaliųjų augalų ir dumblių chloroplastuose dažnai yra krakmolo grūdelių ir mažų lipidų (riebalų) lašelių. Krakmolo grūdai yra laikinos fotosintezės produktų saugyklos. Iš tamsoje išbuvusio augalo chloroplastų jie gali išnykti tik 24 valandas, o vėl atsirasti praėjus 3–4 valandoms po augalo perkėlimo į šviesą.
Chloroplastai yra pusiau autonominiai organeliai ir panašūs į bakterijas. Pavyzdžiui, bakterijų ir chloroplastų ribosomos yra gana panašios. Jie yra mažesni už eukariotų ribosomas. Baltymų sintezę bakterijų ir chloroplastų ribosomose slopina chloramfenikolis, kuris eukariotinėse ląstelėse neveikia. Be to, tiek bakterijos, tiek chloroplastai turi panašų nukleoidų tipą, organizuotą panašiai. Nepaisant to, kad chloroplastų susidarymą ir juose esančių pigmentų sintezę daugiausia kontroliuoja ląstelės chromosomų DNR, vis dėlto, nesant savo DNR, chloroplastai nesusidaro.
Chloroplastai gali būti laikomi pagrindiniais ląstelių organeliais, nes jie yra pirmieji saulės energijos transformacijos grandinėje, dėl kurios žmonija gauna ir maistą, ir kurą. Chloroplastuose vyksta ne tik fotosintezė. Jie taip pat dalyvauja aminorūgščių ir riebalų rūgščių sintezėje, tarnauja kaip laikinų krakmolo atsargų saugykla.
Chromoplastai yra pigmentuoti plastidai. Įvairios formos, jos neturi chlorofilo, tačiau sintetina ir kaupia karotinoidus, kurie žiedams, seniems lapams, vaisiams ir šaknims suteikia geltoną, oranžinę, raudoną spalvą. iš chloroplastų gali išsivystyti chromoplastai, kurie tokiu atveju praranda chlorofilą ir vidines membranines struktūras, kaupia karotenoidus. Tai atsitinka daugelio vaisių nokimo metu. Chromoplastai pritraukia vabzdžius ir kitus gyvūnus, su kuriais jie vystėsi kartu.
Leukoplastai yra nepigmentuoti plastidai. Kai kurie iš jų sintetina krakmolą (amiloplastus), kiti gali sudaryti įvairias medžiagas, įskaitant lipidus ir baltymus. Šviesoje leukoplastai virsta chloroplastais.
Proplastidai yra mažos, bespalvės arba šviesiai žalios, nediferencijuotos plastidės, randamos meristematinėse (dalijamosiose) šaknų ir ūglių ląstelėse. Jie yra kitų, labiau diferencijuotų plastidų – chloroplastų, chromoplastų ir aminoplastų – pirmtakai. Jei protoplastidų vystymasis vėluoja dėl šviesos trūkumo, juose gali atsirasti vienas ar daugiau prolamelių kūnų, kurie yra pusiau kristalinės vamzdinių membranų sankaupos. Plastidės, kuriose yra prolamelių kūnų, vadinamos etioplastais. Šviesoje etioplastai virsta chloroplastais, o prolamelių kūnų membranos sudaro tilakoidus. Etioplastai susidaro tamsoje esančių augalų lapuose. sėklinių embrionų protoplastai pirmiausia virsta etioplastais, iš kurių vėliau šviesoje vystosi chloroplastai. Plastidams būdingas gana lengvas perėjimas iš vienos rūšies į kitą. Plastidės, kaip ir bakterijos, dauginasi dalijantis į dvi dalis. Meristematinėse ląstelėse protoplastidų dalijimosi laikas maždaug sutampa su ląstelių dalijimosi laiku. Tačiau subrendusiose ląstelėse dauguma plastidų susidaro dėl subrendusių plastidų dalijimosi.
Mitochondrijos. Kaip ir chloroplastus, mitochondrijas supa dvi elementarios membranos. Vidinė membrana suformuoja daug raukšlių ir išsikišimų – cristae, kurios žymiai padidina vidinį mitochondrijų paviršių. Jie yra žymiai mažesni už plastidus, apie 0,5 µm skersmens ir įvairaus ilgio bei formos.
Mitochondrijose vyksta kvėpavimo procesas, kurio pasekoje organinės molekulės suskaidomos išsiskiriant energijai ir ją perduodant ATP molekulėms – pagrindiniam visų eukariotinių ląstelių energijos rezervui. Daugumoje augalų ląstelių yra šimtai ar tūkstančiai mitochondrijų. Jų skaičius vienoje ląstelėje nustatomas pagal ląstelės ATP poreikį. Mitochondrijos nuolat juda, juda iš vienos ląstelės dalies į kitą, susilieja viena su kita ir dalijasi. Mitochondrijos dažniausiai susirenka ten, kur reikia energijos. Jei plazminė membrana aktyviai perneša medžiagas iš ląstelės į ląstelę, tai mitochondrijos yra išilgai membranos paviršiaus. Judriuose vienaląsčiuose dumbliuose mitochondrijos kaupiasi žvynelių apačioje, tiekdamos jiems judėjimui reikalingą energiją.
Mitochondrijos, kaip ir plastidai, yra pusiau autonominės organelės, turinčios komponentų, reikalingų jų pačių baltymų sintezei. Vidinė membrana supa skystą matricą, kurioje yra baltymų, RNR, DNR, į bakterijas panašių ribosomų ir įvairių tirpių medžiagų. DNR egzistuoja apskritų molekulių, esančių viename ar keliuose nukleoiduose, pavidalu.
Remiantis bakterijų panašumu su eukariotinių ląstelių mitochondrijomis ir chloroplastais, galima daryti prielaidą, kad mitochondrijos ir chloroplastai atsirado iš bakterijų, kurios „prieglobstį“ rado didesnėse heterotrofinėse ląstelėse – eukariotų pirmtakuose.
Mikrokūnai. Skirtingai nuo plastidžių ir mitochondrijų, kurias riboja dvi membranos, mikrokūnai yra sferinės organelės, apsuptos viena membrana. Mikrokūnai turi granuliuotą (granuliuotą) turinį, kartais juose yra ir kristalinių baltymų intarpų. Mikrokūnai yra susiję su vienu ar dviem endoplazminio tinklo sritimis.
Kai kurie mikrokūnai, vadinami proksisomomis, atlieka svarbų vaidmenį glikolio rūgšties metabolizme, kuris yra tiesiogiai susijęs su fotokvėpavimu. Žaliuose lapuose jie yra susiję su mitochondrijomis ir chloroplastais. Kituose mikrokūnuose, vadinamuose glioksisomomis, yra fermentų, reikalingų riebalams paversti angliavandeniais. Tai atsitinka daugelyje sėklų dygimo metu.
Vakuolės - Tai membrana atribotos ląstelės sritys, užpildytos skysčiu – ląstelių sultimis. Juos supa tonoplastas (vakuolinė membrana).
Jaunoje augalo ląstelėje yra daug mažų vakuolių, kurios ląstelei senstant susilieja į vieną didelę vakuolę. Brandžioje ląstelėje iki 90% jos tūrio gali užimti vakuolė. Šiuo atveju citoplazma plono periferinio sluoksnio pavidalu prispaudžiama prie ląstelės membranos. Ląstelių dydžio padidėjimą daugiausia lemia vakuolės augimas. Dėl to atsiranda turgorinis slėgis ir išlaikomas audinių elastingumas. Tai viena iš pagrindinių vakuolės ir tonoplasto funkcijų.
Pagrindinis sulčių komponentas yra vanduo, likusi dalis skiriasi priklausomai nuo augalo rūšies ir jo fiziologinės būklės. Vakuolėse yra druskų, cukrų, rečiau baltymų. Tonoplastas vaidina aktyvų vaidmenį transportuojant ir kaupiant tam tikrus jonus vakuolėje. Jonų koncentracija ląstelės sultyse gali gerokai viršyti jos koncentraciją aplinkoje. Esant dideliam tam tikrų medžiagų kiekiui, vakuolėse susidaro kristalai. Labiausiai paplitę yra kalcio oksalato kristalai, kurie turi skirtingą formą.
Vakuolės yra medžiagų apykaitos produktų (metabolizmo) kaupimosi vietos. Tai gali būti baltymai, rūgštys ir net žmonėms toksiškos medžiagos (alkaloidai). Pigmentai dažnai nusėda. Mėlyna, violetinė, violetinė, tamsiai raudona, tamsiai raudona suteikia augalų ląstelėms pigmentų iš antocianinų grupės. Skirtingai nuo kitų pigmentų, jie gerai tirpsta vandenyje ir yra ląstelių sultyse. Jie lemia daugelio daržovių (ridikėlių, ropių, kopūstų), vaisių (vynuogių, slyvų, vyšnių), gėlių (rugiagėlių, pelargonijų, delfinijų, rožių, bijūnų) raudoną ir mėlyną spalvą. Kartais šie pigmentai užmaskuoja chlorofilą lapuose, pavyzdžiui, dekoratyviniame raudonajame kleve. Antocianinai nuspalvina rudens lapus ryškiai raudonai. Jie susidaro šaltu saulėtu oru, kai lapuose sustoja chlorofilo sintezė. Lapuose, kai nesusidaro antocianinai, sunaikinus chlorofilą, pastebimi geltonai oranžiniai chloroplastų karotenoidai. Lapai ryškiausios spalvos būna šaltą, giedrą rudenį.
Vakuolės dalyvauja makromolekulių sunaikinime, jų komponentų cirkuliacijoje ląstelėje. Ribosomos, mitochondrijos, plastidai, patenkantys į vakuoles, sunaikinami. Pagal šią virškinimo veiklą jas galima palyginti su lizosomomis – gyvūnų ląstelių organelėmis.
Vakuolės susidaro iš endoplazminio tinklo (tinklo)
Ribosomos. Mažos dalelės (17–23 nm), susidedančios iš maždaug vienodo kiekio baltymų ir RNR. Ribosomose aminorūgštys susijungia ir sudaro baltymus. Ląstelėse, kuriose vyksta aktyvi medžiagų apykaita, jų yra daugiau. Ribosomos yra ląstelės citoplazmoje laisvai arba prijungtos prie endoplazminio tinklo (80S). Jų taip pat yra branduolyje (80S), mitochondrijose (70S), plastiduose (70S).
Ribosomos gali sudaryti kompleksą, kuriame vienu metu vyksta identiškų polipeptidų sintezė, informacija apie kurią paimama iš vienos molekulės ir RNR. Toks kompleksas vadinamas poliribosomomis (polisomomis). Ląstelės, kurios sintetina baltymus dideliais kiekiais, turi didelę polisomų sistemą, kuri dažnai yra pritvirtinta prie išorinio branduolio apvalkalo paviršiaus.
Endoplazminis Tinklelis. Tai sudėtinga neriboto dydžio trimatė membraninė sistema. ER kontekste atrodo kaip dvi elementarios membranos su siaura skaidria erdve tarp jų. ER forma ir ilgis priklauso nuo ląstelės tipo, jos metabolinio aktyvumo ir diferenciacijos stadijos. Ląstelėse, kurios išskiria arba kaupia baltymus, ER yra plokščių maišelių arba cisternų formos, o prie išorinio paviršiaus yra pritvirtinta daug ribosomų. Šis tinklas vadinamas grubiu endoplazminiu tinkleliu. Lygus ER paprastai yra vamzdinės formos. Toje pačioje ląstelėje gali būti šiurkštus ir lygus endoplazminis tinklas. Paprastai tarp jų yra daug ryšių.
Endoplazminis tinklas veikia kaip ląstelės ryšio sistema. Jis yra susijęs su išoriniu branduolio apvalkalu. Tiesą sakant, šios dvi struktūros sudaro vieną membranų sistemą. Kai ląstelės dalijimosi metu plyšta branduolio apvalkalas, jo fragmentai primena ER fragmentus. Endoplazminis tinklas yra sistema, pernešanti medžiagas: baltymus, lipidus, angliavandenius į skirtingas ląstelės dalis. kaimyninių ląstelių endoplazminis tinklas yra sujungtas per citoplazmines sruogas - plazmodesmatas -, kurios praeina per ląstelių membranas.
Endoplazminis tinklas yra pagrindinė ląstelių membranų sintezės vieta. Kai kuriose augalų ląstelėse čia susidaro vakuolių ir mikrokūnų membranos, diktiozomų cisternos.
Goldžio kompleksas.Šis terminas vartojamas apibūdinti visas diktiozomas arba Golgi kūnus ląstelėje. Diktiosomos yra plokščių, disko formos pūslelių arba cisternų grupės, kurios išsišakoja išilgai kraštų į sudėtingą kanalėlių sistemą. Aukštesniųjų augalų diktiosomos susideda iš 4–8 cisternų, surinktų kartu.
Paprastai rezervuarų pakuotėje išskiriama besivystanti ir bręstanti pusė. formuojančių cisternų membranos savo struktūra primena ER membranas, o bręstančių cisternų membranos primena plazminę membraną.
Diktiosomos dalyvauja sekrecijoje, o daugumoje aukštesniųjų augalų – ląstelių membranų formavime. Diktiosomų sintetinami ląstelių sienelių polisacharidai kaupiasi pūslelėse, kurios vėliau atsiskiria nuo bręstančių cisternų. Šios sekrecinės pūslelės migruoja ir susilieja su CMP; o juose esantys polisacharidai yra įterpti į ląstelės membraną. Kai kurios medžiagos, besikaupiančios diktiozomose, susidaro kitose struktūrose, pavyzdžiui, ER, o po to pernešamos į diktioomas, kur jos modifikuojamos (modifikuojamos) prieš sekreciją. Pavyzdžiui, glikoproteinai yra svarbi ląstelės membranos statybinė medžiaga. Baltyminę dalį sintetina šiurkščiojo ER polisomos, angliavandenių dalį – diktiozomose, kur abi dalys susijungdamos sudaro glikoproteinus.
Membranos yra dinamiškos, judrios struktūros, nuolat keičiančios savo formą ir plotą. Endoplazminės sistemos samprata remiasi membranų mobilumu. Pagal šią koncepciją vidinės citoplazmos membranos, išskyrus mitochondrijų ir plastidžių membranas, yra viena visuma ir kyla iš endoplazminio tinklo. Iš endoplazminio tinklo per tarpinę pūslelių stadiją susidaro naujos diktiozomų cisternos, o sekrecinės pūslelės, kurios atsiskiria nuo diktiozomų, ilgainiui prisideda prie plazminės membranos susidarymo. Taigi, endoplazminis tinklas ir diktiosomos sudaro funkcinį vienetą, kuriame diktiosomos atlieka tarpinių struktūrų vaidmenį membranas, tokias kaip endoplazminis tinklas, paverčiant tokiomis membranomis kaip plazma. Audiniuose, kurių ląstelės auga ir dalijasi silpnai, membranos komponentai nuolat atnaujinami.
mikrovamzdeliai randama beveik visose eukariotinėse ląstelėse. Tai cilindrinės struktūros, kurių skersmuo yra apie 24 nm. Jų ilgis skiriasi. Kiekvienas kanalėlis sudarytas iš baltymo, vadinamo tubulinu, subvienetų. Subvienetai sudaro 13 išilginių gijų, supančių centrinę ertmę. Mikrovamzdeliai yra dinamiškos struktūros, jos reguliariai sunaikinamos ir susidaro tam tikrais ląstelių ciklo etapais. Jų surinkimas vyksta specialiose vietose, kurios vadinamos mikrovamzdelių organizavimo centrais. Augalų ląstelėse jie turi silpnai išreikštą amorfinę struktūrą.
Mikrovamzdelių funkcijos: dalyvauja formuojant ląstelės membraną; nukreipti diktiozomų burbulus į besiformuojančią membraną, pavyzdžiui, verpstės siūlus, kurie susidaro besidalijančioje ląstelėje; vaidina vaidmenį formuojant ląstelės plokštelę (pradinę ribą tarp dukterinių ląstelių). Be to, mikrovamzdeliai yra svarbi žvynelių ir blakstienų sudedamoji dalis, kurių judėjime jie atlieka svarbų vaidmenį.
mikrofilamentai, kaip ir mikrovamzdeliai, jų yra beveik visose eukariotinėse ląstelėse. Tai ilgi, 5–7 nm storio siūlai, susidedantys iš susitraukiančio baltymo aktino. Mikrofilamentų ryšuliai randami daugelyje aukštesniųjų augalų ląstelių. Matyt, jie vaidina svarbų vaidmenį citoplazmos srovėse. Mikrofilamentai kartu su mikrotubuliais sudaro lankstų tinklą, vadinamą citoskeletu.
Bazinė medžiaga ilgą laiką buvo laikomas vienalyčiu (homogenišku) daug baltymų turinčiu tirpalu, turinčiu nedidelį skaičių struktūrų ar net bestruktūrių. Tačiau šiuo metu, naudojant aukštos įtampos elektroninį mikroskopą, nustatyta, kad gruntinė medžiaga yra trimatė gardelė, sudaryta iš plonų (3–6 nm skersmens) gijų, užpildančių visą ląstelę. Kiti citoplazmos komponentai, įskaitant mikrovamzdelius ir mikrofilamentus, yra suspenduoti iš šios mikrotrabekulinės gardelės.
Mikrotrabekulinė struktūra yra baltymų gijų gardelė, tarp kurių esanti erdvė užpildyta vandeniu. Kartu su vandeniu grotelės yra gelio konsistencijos, gelis atrodo kaip želatininiai kūnai.
Organelės yra pritvirtintos prie mikrotrabekulinės gardelės. Grotelės užtikrina ryšį tarp atskirų ląstelės dalių ir nukreipia tarpląstelinį transportą.
Lipidų lašai- sferinės formos struktūros, suteikiančios augalo ląstelės citoplazmos granuliuotumą šviesos mikroskopu. Elektroninėse mikrografijose jie atrodo amorfiški. Labai panašūs, bet mažesni lašai randami plastiduose.
Lipidų lašai, painiodami juos su organelėmis, vadino juos sferosomomis ir tikėjo, kad juos supa vieno arba dvigubo sluoksnio membrana. Tačiau naujausi duomenys rodo, kad lipidų lašeliai neturi membranų, bet gali būti padengti baltymu.
Ergastinės medžiagos - tai yra protoplasto „pasyvieji produktai“: atsarginės medžiagos arba atliekos. Jie gali atsirasti ir išnykti skirtingais ląstelių ciklo laikotarpiais. Išskyrus krakmolo grūdelius, kristalus, antocianinų pigmentus ir lipidų lašelius. Tai yra dervos, dervos, taninai ir baltymai. Ergastinės medžiagos yra ląstelės membranos dalis, pagrindinė citoplazmos ir organelių, įskaitant vakuoles, medžiaga.
Vėliava ir blakstienos - tai plonos, į plaukus panašios struktūros, besitęsiančios nuo daugelio eukariotinių ląstelių paviršiaus. Jų skersmuo yra pastovus, tačiau ilgis svyruoja nuo 2 iki 150 mikronų. Tradiciškai ilgesnės ir kelios jų vadinamos žiuželiais, o trumpesnės ir gausesnės – blakstienomis. Nėra aiškių skirtumų tarp šių dviejų tipų struktūrų, todėl terminas flagellum vartojamas abiem.
Kai kuriuose dumbliuose ir grybuose žvyneliai yra judėjimo organai, kurių pagalba jie juda vandenyje. Augaluose (pavyzdžiui, samanose, kepenėliuose, paparčiuose, kai kuriuose gimnasėkliuose) žvyneliais yra tik lytinės ląstelės (gametos).
Kiekvienas žvynelis turi tam tikrą organizaciją. Išorinis 9 porų mikrovamzdelių žiedas supa du papildomus mikrovamzdelius, esančius žvynelio centre. Fermentų turinčios „rankenos“ tęsiasi iš kiekvienos išorinės poros vieno mikrotubulo. Tai yra pagrindinis 9+2 organizavimo modelis, randamas visose eukariotinių organizmų žvyneliuose. Manoma, kad žvynelių judėjimas pagrįstas mikrotubulių slydimu, o išorinės mikrovamzdelių poros juda viena iš kitos nesusitraukdamos. Mikrovamzdelių porų slydimas vienas kito atžvilgiu sukelia vietinį žvynelio lenkimą.
Žvyneliai „auga“ iš citoplazminių cilindrinių struktūrų, vadinamų baziniais kūnais, kurie taip pat sudaro bazinę žvynelio dalį. Baziniai kūnai turi vidinę struktūrą, panašią į žvynelinę, išskyrus tai, kad išoriniai kanalėliai yra išsidėstę trynukais, o ne poromis, o centrinių kanalėlių nėra.
Ląstelių sienelės. Ląstelės sienelė riboja protoplasto dydį ir neleidžia jam plyšti dėl vandens sugėrimo vakuolės.
Ląstelės sienelė atlieka specifines funkcijas, kurios svarbios ne tik ląstelei ir audiniui, kuriame yra ląstelė, bet ir visam augalui. Ląstelių sienelės atlieka esminį vaidmenį įsisavinant, pernešant ir pašalinant medžiagas, be to, jose gali koncentruotis lizosominė, arba virškinimo veikla.
Ląstelės sienelės komponentai. Tipiškiausias ląstelės sienelės komponentas yra celiuliozė, kuri daugiausia lemia jos architektūrą. Celiuliozės molekulės yra sudarytos iš pasikartojančių gliukozės molekulių, sujungtų galais. Ilgos plonos celiuliozės molekulės sujungiamos į 10–25 nm storio mikrofibriles. Mikrofibrilės susipina ir sudaro plonus siūlus, kurie savo ruožtu gali būti apvynioti vienas aplink kitą, tarsi sruogos virvelėje. Kiekviena tokia „virvė“ arba makrofibrilė yra apie 0,5 mikrono storio, o ilgis siekia 4 mikronus. Makrofibrilės yra tokios pat stiprios kaip vienodo dydžio plieninė viela.
Ląstelės sienelės celiuliozės karkasas užpildytas su juo susipynusiomis celiuliozės matricos molekulėmis. Jį sudaro polisacharidai, vadinami hemiceliuliozės, ir pektinai, arba pektinai, chemiškai labai artimi hemiceliuliozei.
Kitas ląstelės sienelės komponentas – ligninas – yra gausiausias polimeras augalų ląstelėse po celiuliozės. Ligninas padidina sienos standumą ir dažniausiai randamas ląstelėse, kurios atlieka atraminę arba mechaninę funkciją.
Kutinas, suberinas, vaškas – dažniausiai nusėda augalų apsauginių audinių lukštuose. Pavyzdžiui, kutinas randamas epidermio ląstelių membranose, o suberinas – antriniame apsauginiame audinyje – kamštelyje. Abi medžiagos randamos kartu su vašku ir neleidžia augalui pernelyg prarasti vandens.
Ląstelės sienelės sluoksniai. Augalų ląstelių sienelių storis labai skiriasi, priklausomai nuo ląstelių vaidmens augalo struktūroje ir pačios ląstelės amžiaus. Elektroniniu mikroskopu augalo ląstelės sienelėje matomi du sluoksniai: mediana (taip pat vadinama tarpląsteline medžiaga) ir pirminė ląstelės sienelė. Daugelis ląstelių kloja kitą sluoksnį – antrinę ląstelės sienelę. Vidutinė plokštė yra tarp gretimų ląstelių pirminių sienelių. Antrinę sienelę, jei yra, ląstelės protoplastas nusodina ant pirminės ląstelės sienelės vidinio paviršiaus.
Vidurinė plokštelė. Vidurinę plokštelę daugiausia sudaro pektinas. Ten, kur tarp dviejų naujai susidariusių ląstelių turėtų atsirasti ląstelės sienelė, pirmiausia pastebimas tankus endoplazminio tinklo kanalėlių rezginys ir Golgi aparato cisterna (diktiosomos). Tada šioje vietoje atsiranda burbuliukai, užpildyti pektinu (iš polisacharidų). Šie burbuliukai yra atskirti nuo Golgi aparato rezervuarų. Ankstyvojoje ląstelės sienelėje yra įvairių polisacharidų, iš kurių pagrindiniai yra pektinai ir hemiceliuliozė. Vėliau jis susideda iš tankesnių medžiagų – celiuliozės ir lignino.
Pirminė ląstelės sienelė. Tai yra celiuliozės apvalkalo sluoksnis, kuris nusėda prieš ląstelių augimą arba jo metu. Be celiuliozės, hemiceliuliozės ir pektino, pirminėse membranose yra glikoproteino. Pirminiai apvalkalai gali sudegti. Pektino komponentas suteikia plastiškumo, todėl pirminis apvalkalas gali išsiplėsti, kai šaknis, stiebas ar lapas pailgėja.
Aktyviai besidalijančios ląstelės (dauguma subrendusių ląstelių, dalyvaujančių fotosintezės, kvėpavimo ir sekrecijos procesuose) turi pirmines membranas. Tokios ląstelės, turinčios pirminę membraną ir gyvą protoplastą, gali prarasti jiems būdingą formą, dalytis ir diferencijuotis į naują ląstelių tipą. Jie dalyvauja žaizdų gijime ir audinių regeneracijoje augaluose.
Pirminės ląstelių membranos nėra vienodo storio per visą ilgį, tačiau turi plonas dalis, kurios vadinamos pirminių porų laukais. Citoplazmos gijos arba plazmodesmata, jungiančios gretimų ląstelių protoplastus, dažniausiai praeina per pirminius porų laukus.
antrinė ląstelės sienelė. Nepaisant to, kad daugelis augalų ląstelių turi tik pirminę sienelę, kai kuriose, protoplastas nusodina antrinę sienelę link ląstelės centro. Paprastai tai įvyksta sustojus ląstelių augimui ir pirminės membranos plotas nebedidėja. Dėl šios priežasties antrinis apvalkalas skiriasi nuo pirminio. Antrinės membranos ypač reikalingos specializuotoms ląstelėms, kurios stiprina augalą ir praleidžia vandenį. Nusodinus antrinę membraną, šių ląstelių protoplastas, kaip taisyklė, miršta. Antriniuose apvalkaluose celiuliozės yra daugiau nei pirminiuose, juose nėra pektino medžiagų ir glikoproteinų. Antrinė membrana sunkiai ištempiama, jos matrica susideda iš hemiceliuliozės.
Antriniame apvalkale galima išskirti tris sluoksnius – išorinį, vidurinį ir vidinį (S1, S2, S3). Sluoksniuota antrinių apvalkalų struktūra žymiai padidina jų stiprumą. Celiuliozės mikrofibrilės antrinėje membranoje nusėda tankiau nei pirminėje. Ligninas yra įprastas antrinių medienos odelių komponentas.
Poros kontaktuojančių ląstelių membranose yra viena priešais kitą. Dvi priešingos poros ir porų membrana sudaro porų porą. Ląstelėse su antrine membrana yra du pagrindiniai porų tipai: paprastos ir apvaduotos. Apribotose porose antrinė sienelė kabo virš porų ertmės. Paprastose porose taip nėra.
Ląstelių sienelių augimas. Kai ląstelė auga, didėja ląstelės membranos storis ir plotas. Lukšto tempimas yra sudėtingas procesas. Jį kontroliuoja protoplastas ir reguliuoja hormonas auksinas.
Ląstelėse, kurios auga tolygiai visomis kryptimis, miofibrilių nusėdimas yra atsitiktinis. Šios miofibrilės sudaro netaisyklingą tinklą. Tokios ląstelės buvo aptiktos stiebo šerdyje, saugojimo audiniuose ir ląstelių auginimo in vitro metu. Pailgėjusiose ląstelėse šoninių apvalkalų miofibrilės nusėda stačiu kampu pailgėjimo ašiai.
Matricinės medžiagos – pektinai, hemiceliuliozės ir glikoproteinai į membraną pernešami diktiozomų pūslelėmis. Tuo pačiu pektinai labiau būdingi augančioms ląstelėms, o neaugančiose ląstelėse vyrauja hemiceliuliozės.
Celiuliozės mikrofibrilės sintezuojamos ląstelės paviršiuje, padedant fermentų kompleksui, susijusiam su plazmine membrana. Mikrofibrilių orientaciją kontroliuoja mikrotubulės, esančios šalia vidinio plazmos membranos paviršiaus.
Plazmodezma. Tai ploni citoplazmos siūlai, jungiantys kaimyninių ląstelių protoplastus. Plazmodesmatos arba praeina pro ląstelės sienelę bet kur, arba yra susitelkusios pirminiuose porų laukuose arba membranose tarp porų porų. Elektroniniu mikroskopu plazmodesmata atrodo kaip siauri kanalai, iškloti plazmos membrana. Išilgai kanalo ašies nuo vienos ląstelės iki kitos tęsiasi mažesnis cilindrinis vamzdelis – desmotubulas, kuris bendrauja su abiejų gretimų ląstelių endoplazminiu tinklu. Daugelis plazmodesmų susidaro ląstelių dalijimosi metu, kai besivystančios ląstelės plokštelė paima vamzdinį endoplazminį tinklą. Plazmodezmos taip pat gali susidaryti nesidalijančių ląstelių membranose. Šios struktūros užtikrina efektyvų tam tikrų medžiagų perkėlimą iš ląstelės į ląstelę.
Ląstelių dalijimasis. Daugialąsčiuose organizmuose ląstelių dalijimasis kartu su jų dydžio padidėjimu yra viso organizmo augimo būdas. Dalijimosi metu susidariusios naujos ląstelės savo struktūra ir funkcijomis yra panašios tiek į pirminę, tiek į viena kitą. Eukariotų dalijimosi procesą galima suskirstyti į du iš dalies persidengiančius etapus: mitozę ir citokinezę.
Mitozė – tai dviejų dukterinių branduolių susidarymas iš vieno branduolio, morfologiškai ir genetiškai lygiaverčiai vienas kitam. Citokinezė – tai ląstelės citoplazminės dalies dalijimasis, kai susidaro dukterinės ląstelės.
Ląstelių ciklas. Gyva ląstelė pereina iš eilės įvykių, kurie sudaro ląstelės ciklą. Pačio ciklo trukmė skiriasi priklausomai nuo ląstelės tipo ir išorinių veiksnių, tokių kaip temperatūra ar maistinių medžiagų prieinamumas. Paprastai ciklas skirstomas į tarpfazę ir keturias mitozės fazes.
Tarpfazė. Laikotarpis tarp nuoseklių mitozinių dalijimų.
Tarpfazė yra padalinta į tris periodus, žymimus kaip G1, S, G2.
G1 periodu, kuris prasideda po mitozės. Per šį laikotarpį padidėja citoplazmos kiekis, įskaitant įvairius organelius. Be to, pagal dabartinę hipotezę G1 periodo metu sintetinamos medžiagos, kurios arba stimuliuoja, arba slopina S periodą ir likusį ciklą, taip nulemdamos dalijimosi procesą.
S laikotarpis seka G1 periodą, tuo metu genetinė medžiaga (DNR) yra dubliuojama.
Per G2 laikotarpį, einantį po S, susidaro struktūros, kurios tiesiogiai dalyvauja mitozėje, pavyzdžiui, veleno komponentai.
Kai kurios ląstelės praeina neribotą ląstelių ciklų skaičių. Tai vienaląsčiai organizmai ir kai kurios aktyvaus augimo zonų ląstelės (meristemos). Kai kurios specializuotos ląstelės po brendimo praranda gebėjimą daugintis. Trečioji ląstelių grupė, pavyzdžiui, formuojančios žaizdos audinį (kalusą), gebėjimą dalytis išlaiko tik ypatingomis sąlygomis.
mitozė, arba branduolio dalijimasis. Tai nenutrūkstamas procesas, suskirstytas į keturias fazes: profazę, metafazę, anafazę, telofazę. Dėl mitozės tarpfazėje padvigubėjusi genetinė medžiaga po lygiai pasiskirsto tarp dviejų dukterinių branduolių.
Vienas iš pirmųjų ląstelių perėjimo prie dalijimosi požymių yra siauros žiedinės mikrovamzdelių juostos atsiradimas tiesiai po plazmos membrana. Šis gana tankus diržas supa branduolį būsimojo mitozinio veleno pusiaujo plokštumoje. Kadangi jis pasirodo prieš profazę, jis vadinamas priešfazės juosta. Jis išnyksta po mitozinio verpstės, dar ilgai prieš atsirandant vėlyvojoje ląstelės plokštelės telofazėje, kuri auga nuo centro iki periferijos ir susilieja su motininės ląstelės membrana toje srityje, kurią anksčiau užėmė priešfazinis diržas.
Profazė. Profazės pradžioje chromosomos primena ilgus siūlus, išsibarsčiusius branduolio viduje. Tada, trumpėjant ir storėjant sruogoms, galima pastebėti, kad kiekviena chromosoma susideda ne iš vienos, o iš dviejų susipynusių sruogų, vadinamų chromatidėmis. Vėlyvojoje fazėje dvi sutrumpintos suporuotos kiekvienos chromosomos chromatidės yra lygiagrečiai, sujungtos siaura atkarpa, vadinama centromera. Jis turi tam tikrą vietą kiekvienoje chromosomoje ir padalija chromosomą į dvi skirtingo ilgio šakas.
Mikrovamzdeliai išsidėstę lygiagrečiai branduolio paviršiui išilgai veleno ašies. Tai yra ankstyviausias mitozinio veleno surinkimo pasireiškimas.
Profazės pabaigoje branduolys palaipsniui praranda aiškų kontūrą ir galiausiai išnyksta. Netrukus po to branduolinis apvalkalas taip pat suyra.
Metafazė. Metafazės pradžioje verpstė, kuri yra trimatė struktūra, plačiausia per vidurį ir siaurėjanti link polių, užima vietą, kurią anksčiau užėmė branduolys. Verpstės pluoštai yra mikrovamzdelių pluoštai. Metafazės metu chromosomos, kurių kiekviena susideda iš dviejų chromatidžių, yra išdėstytos taip, kad jų centromerai būtų suklio pusiaujo plokštumoje. Kiekviena chromosoma savo centromeru yra pritvirtinta prie veleno sriegių. Tačiau kai kurios sruogos pereina iš vieno poliaus į kitą neprisirišusios prie chromosomų.
Kai visos chromosomos yra pusiaujo plokštumoje, metafazė baigiama. Chromosomos yra pasirengusios dalytis.
Anafazė. Kiekvienos chromosomos chromatidės išsiskiria. Dabar tai yra dukterinės chromosomos. Visų pirma, centromera dalijasi, o dvi dukterinės chromosomos traukiamos į priešingus polius. Šiuo atveju centromerai juda į priekį, o chromosomų rankos išsitiesia už nugaros. Prie chromosomų prisirišę verpstės siūlai sutrumpėja, prisidedant prie chromatidų atsiskyrimo ir dukterinių chromosomų judėjimo priešingomis kryptimis.
AUGALŲ LĄSTELĖS STRUKTŪRA
Augalo ląstelė susideda iš daugiau ar mažiau standžios ląstelės sienelės ir protoplasto. Ląstelės sienelė yra ląstelės sienelė ir citoplazminė membrana. Terminas protoplastas kilęs iš žodžio protoplazma, kuris ilgą laiką buvo vartojamas visiems gyviems dalykams apibūdinti. Protoplastas yra atskiros ląstelės protoplazma.
Protoplastas susideda iš citoplazmos ir branduolio. Citoplazmoje yra organelių (ribosomų, mikrotubulių, plastidžių, mitochondrijų) ir membraninių sistemų (endoplazminis tinklas, diktiosomos). Citoplazma taip pat apima citoplazminę matricą (pagrindinę medžiagą), į kurią panardinami organeliai ir membranų sistemos. Citoplazmą nuo ląstelės sienelės skiria plazminė membrana, kuri yra elementari membrana. Skirtingai nuo daugelio gyvūnų ląstelių, augalų ląstelėse yra viena ar daugiau vakuolių. Tai pūslelės, užpildytos skysčiu ir apsuptos elementaria membrana (tonoplastas).
Gyvoje augalo ląstelėje žemės medžiaga nuolat juda. Organelės dalyvauja judėjime, vadinamame citoplazmos srautu arba cikloze. Ciklozė palengvina medžiagų judėjimą ląstelėje ir jų mainus tarp ląstelės ir aplinkos.
Plazmos membrana. Tai dvisluoksnė fosfolipidinė struktūra. Augalų ląstelėms būdingos plazminės membranos invaginacijos.
Plazminė membrana atlieka šias funkcijas:
Dalyvauja medžiagų mainuose tarp ląstelės ir aplinkos;
Koordinuoja ląstelės sienelės celiuliozės mikrofibrilių sintezę ir surinkimą;
Perduoda hormoninius ir išorinius signalus, kurie kontroliuoja ląstelių augimą ir diferenciaciją.
Šerdis. Tai ryškiausia eukariotinės ląstelės citoplazmos struktūra. Branduolys atlieka dvi svarbias funkcijas:
Kontroliuoja gyvybinę ląstelės veiklą, nustatydama, kurie baltymai ir kokiu laiku turi būti sintetinami;
Saugo genetinę informaciją ir perduoda ją dukterinėms ląstelėms ląstelių dalijimosi metu.
Eukariotinės ląstelės branduolį supa dvi elementarios membranos, kurios sudaro branduolio apvalkalą. Jis persmelktas daugybe porų, kurių skersmuo nuo 30 iki 100 nm, matomos tik elektroniniu mikroskopu. Poros turi sudėtingą struktūrą. Branduolinės membranos išorinė membrana kai kuriose vietose yra sujungta su endoplazminiu tinklu. Branduolinis apvalkalas gali būti vertinamas kaip specializuota, lokaliai diferencijuota endoplazminio tinklo (ER) dalis.
Specialiais dažais nudažytame branduolyje galima išskirti plonus siūlus ir chromatino bei nukleoplazmos (pagrindinės branduolio medžiagos) gumulėlius. Chromatinas susideda iš DNR, susietos su specialiais baltymais, vadinamais histonais. Ląstelių dalijimosi procese chromatinas vis labiau sutankinamas ir surenkamas į chromosomas. DNR koduoja genetinę informaciją.
Organizmai skiriasi chromosomų skaičiumi somatinėse ląstelėse. Pavyzdžiui, kopūstai turi - 20 chromosomų; saulėgrąžos - 34; kviečiai - 42; žmonių – 46, o viena iš paparčio Ophioglossum rūšių – 1250. Lytinės ląstelės (gametos) turi tik pusę chromosomų, būdingų somatinėms kūno ląstelėms. Chromosomų skaičius gametose vadinamas haploidiniu (viengubu), somatinėse ląstelėse – diploidiniu (dvigubu). Ląstelės, turinčios daugiau nei du chromosomų rinkinius, vadinamos poliploidinėmis.
Šviesos mikroskopu matomos sferinės struktūros – branduoliai. Kiekvienas branduolys turi vieną ar daugiau branduolių, kurie matomi nesidalijančiuose branduoliuose. Ribosominė RNR sintetinama branduolyje. Paprastai diploidinių organizmų branduoliai turi du branduolius, po vieną kiekvienam haploidiniam chromosomų rinkiniui. Branduoliai neturi savo membranos. Biochemiškai branduoliams būdinga didelė RNR koncentracija, kuri čia susijusi su fosfoproteinais. Branduolių dydis priklauso nuo ląstelės funkcinės būklės. pastebėta, kad sparčiai augančioje ląstelėje, kurioje vyksta intensyvūs baltymų sintezės procesai, didėja branduolių dydis. Branduoliuose gaminasi mRNR ir ribosomos, kurios sintetinę funkciją atlieka tik branduolyje.
Nukleoplazmą (karioplazmą) vaizduoja vienalytis skystis, kuriame yra ištirpę įvairūs baltymai, įskaitant fermentus.
Plastidai. Vakuolės, celiuliozės ląstelių sienelės ir plastidai yra būdingi augalų ląstelių komponentai. Kiekviena plastidė turi savo apvalkalą, susidedantį iš dviejų elementarių membranų. Plastido viduje išskiriama membraninė sistema ir įvairaus laipsnio vienalytė medžiaga – stroma. Subrendusios plastidės klasifikuojamos pagal juose esančius pigmentus.
Chloroplastuose, kuriuose vyksta fotosintezė, yra chlorofilų ir karotinoidų. Paprastai turi 4–5 mikronų skersmens disko formą. Vienoje mezofilo ląstelėje (lapo viduryje) gali būti 40 - 50 chloroplastų; mm2 lapo – apie 500 000. citoplazmoje chloroplastai dažniausiai išsidėstę lygiagrečiai ląstelės membranai.
Vidinė chloroplasto struktūra yra sudėtinga. Stroma yra persmelkta išsivysčiusios membranų sistemos pūslelių pavidalu – tilakoidų. Kiekvienas tilakoidas susideda iš dviejų membranų. Tilakoidai sudaro vieną sistemą. Paprastai jie surenkami krūvomis – grūdeliais, primenančiais monetų stulpelius. Atskirų granatų tilakoidai yra tarpusavyje sujungti stromos tilakoidais arba tarpgranaliniais tilakoidais. Chlorofilas ir karotinoidai yra įterpti į tilakoidų membranas. Žaliųjų augalų ir dumblių chloroplastuose dažnai yra krakmolo grūdelių ir mažų lipidų (riebalų) lašelių. Krakmolo grūdai yra laikinos fotosintezės produktų saugyklos. Iš tamsoje išbuvusio augalo chloroplastų jie gali išnykti tik 24 valandas, o vėl atsirasti praėjus 3–4 valandoms po augalo perkėlimo į šviesą.
Chloroplastai yra pusiau autonominiai organeliai ir panašūs į bakterijas. Pavyzdžiui, bakterijų ir chloroplastų ribosomos yra gana panašios. Jie yra mažesni už eukariotų ribosomas. Baltymų sintezę bakterijų ir chloroplastų ribosomose slopina chloramfenikolis, kuris eukariotinėse ląstelėse neveikia. Be to, tiek bakterijos, tiek chloroplastai turi panašų nukleoidų tipą, organizuotą panašiai. Nepaisant to, kad chloroplastų susidarymą ir juose esančių pigmentų sintezę daugiausia kontroliuoja ląstelės chromosomų DNR, vis dėlto, nesant savo DNR, chloroplastai nesusidaro.
Chloroplastai gali būti laikomi pagrindiniais ląstelių organeliais, nes jie yra pirmieji saulės energijos transformacijos grandinėje, dėl kurios žmonija gauna ir maistą, ir kurą. Chloroplastuose vyksta ne tik fotosintezė. Jie taip pat dalyvauja aminorūgščių ir riebalų rūgščių sintezėje, tarnauja kaip laikinų krakmolo atsargų saugykla.
Chromoplastai yra pigmentuoti plastidai. Įvairios formos, jos neturi chlorofilo, tačiau sintetina ir kaupia karotinoidus, kurie žiedams, seniems lapams, vaisiams ir šaknims suteikia geltoną, oranžinę, raudoną spalvą. iš chloroplastų gali išsivystyti chromoplastai, kurie tokiu atveju praranda chlorofilą ir vidines membranines struktūras, kaupia karotenoidus. Tai atsitinka daugelio vaisių nokimo metu. Chromoplastai pritraukia vabzdžius ir kitus gyvūnus, su kuriais jie vystėsi kartu.
Leukoplastai yra nepigmentuoti plastidai. Kai kurie iš jų sintetina krakmolą (amiloplastus), kiti gali sudaryti įvairias medžiagas, įskaitant lipidus ir baltymus. Šviesoje leukoplastai virsta chloroplastais.
Proplastidai yra mažos, bespalvės arba šviesiai žalios, nediferencijuotos plastidės, randamos meristematinėse (dalijamosiose) šaknų ir ūglių ląstelėse. Jie yra kitų, labiau diferencijuotų plastidų – chloroplastų, chromoplastų ir aminoplastų – pirmtakai. Jei protoplastidų vystymasis vėluoja dėl šviesos trūkumo, juose gali atsirasti vienas ar daugiau prolamelių kūnų, kurie yra pusiau kristalinės vamzdinių membranų sankaupos. Plastidės, kuriose yra prolamelių kūnų, vadinamos etioplastais. Šviesoje etioplastai virsta chloroplastais, o prolamelių kūnų membranos sudaro tilakoidus. Etioplastai susidaro tamsoje esančių augalų lapuose. sėklinių embrionų protoplastai pirmiausia virsta etioplastais, iš kurių vėliau šviesoje vystosi chloroplastai. Plastidams būdingas gana lengvas perėjimas iš vienos rūšies į kitą. Plastidės, kaip ir bakterijos, dauginasi dalijantis į dvi dalis. Meristematinėse ląstelėse protoplastidų dalijimosi laikas maždaug sutampa su ląstelių dalijimosi laiku. Tačiau subrendusiose ląstelėse dauguma plastidų susidaro dėl subrendusių plastidų dalijimosi.
Mitochondrijos. Kaip ir chloroplastus, mitochondrijas supa dvi elementarios membranos. Vidinė membrana suformuoja daug raukšlių ir išsikišimų – cristae, kurios žymiai padidina vidinį mitochondrijų paviršių. Jie yra žymiai mažesni už plastidus, apie 0,5 µm skersmens ir įvairaus ilgio bei formos.
Mitochondrijose vyksta kvėpavimo procesas, kurio pasekoje organinės molekulės suskaidomos išsiskiriant energijai ir ją perduodant ATP molekulėms – pagrindiniam visų eukariotinių ląstelių energijos rezervui. Daugumoje augalų ląstelių yra šimtai ar tūkstančiai mitochondrijų. Jų skaičius vienoje ląstelėje nustatomas pagal ląstelės ATP poreikį. Mitochondrijos nuolat juda, juda iš vienos ląstelės dalies į kitą, susilieja viena su kita ir dalijasi. Mitochondrijos dažniausiai susirenka ten, kur reikia energijos. Jei plazminė membrana aktyviai perneša medžiagas iš ląstelės į ląstelę, tai mitochondrijos yra išilgai membranos paviršiaus. Judriuose vienaląsčiuose dumbliuose mitochondrijos kaupiasi žvynelių apačioje, tiekdamos jiems judėjimui reikalingą energiją.
Mitochondrijos, kaip ir plastidai, yra pusiau autonominės organelės, turinčios komponentų, reikalingų jų pačių baltymų sintezei. Vidinė membrana supa skystą matricą, kurioje yra baltymų, RNR, DNR, į bakterijas panašių ribosomų ir įvairių tirpių medžiagų. DNR egzistuoja apskritų molekulių, esančių viename ar keliuose nukleoiduose, pavidalu.
Remiantis bakterijų panašumu su eukariotinių ląstelių mitochondrijomis ir chloroplastais, galima daryti prielaidą, kad mitochondrijos ir chloroplastai atsirado iš bakterijų, kurios „prieglobstį“ rado didesnėse heterotrofinėse ląstelėse – eukariotų pirmtakuose.
Mikrokūnai. Skirtingai nuo plastidžių ir mitochondrijų, kurias riboja dvi membranos, mikrokūnai yra sferinės organelės, apsuptos viena membrana. Mikrokūnai turi granuliuotą (granuliuotą) turinį, kartais juose yra ir kristalinių baltymų intarpų. Mikrokūnai yra susiję su vienu ar dviem endoplazminio tinklo sritimis.
Kai kurie mikrokūnai, vadinami proksisomomis, atlieka svarbų vaidmenį glikolio rūgšties metabolizme, kuris yra tiesiogiai susijęs su fotokvėpavimu. Žaliuose lapuose jie yra susiję su mitochondrijomis ir chloroplastais. Kituose mikrokūnuose, vadinamuose glioksisomomis, yra fermentų, reikalingų riebalams paversti angliavandeniais. Tai atsitinka daugelyje sėklų dygimo metu.
Vakuolės - Tai membrana atribotos ląstelės sritys, užpildytos skysčiu – ląstelių sultimis. Juos supa tonoplastas (vakuolinė membrana).
Jaunoje augalo ląstelėje yra daug mažų vakuolių, kurios ląstelei senstant susilieja į vieną didelę vakuolę. Brandžioje ląstelėje iki 90% jos tūrio gali užimti vakuolė. Šiuo atveju citoplazma plono periferinio sluoksnio pavidalu prispaudžiama prie ląstelės membranos. Ląstelių dydžio padidėjimą daugiausia lemia vakuolės augimas. Dėl to atsiranda turgorinis slėgis ir išlaikomas audinių elastingumas. Tai viena iš pagrindinių vakuolės ir tonoplasto funkcijų.
Pagrindinis sulčių komponentas yra vanduo, likusi dalis skiriasi priklausomai nuo augalo rūšies ir jo fiziologinės būklės. Vakuolėse yra druskų, cukrų, rečiau baltymų. Tonoplastas vaidina aktyvų vaidmenį transportuojant ir kaupiant tam tikrus jonus vakuolėje. Jonų koncentracija ląstelės sultyse gali gerokai viršyti jos koncentraciją aplinkoje. Esant dideliam tam tikrų medžiagų kiekiui, vakuolėse susidaro kristalai. Labiausiai paplitę yra kalcio oksalato kristalai, kurie turi skirtingą formą.
Vakuolės yra medžiagų apykaitos produktų (metabolizmo) kaupimosi vietos. Tai gali būti baltymai, rūgštys ir net žmonėms toksiškos medžiagos (alkaloidai). Pigmentai dažnai nusėda. Mėlyna, violetinė, violetinė, tamsiai raudona, tamsiai raudona suteikia augalų ląstelėms pigmentų iš antocianinų grupės. Skirtingai nuo kitų pigmentų, jie gerai tirpsta vandenyje ir yra ląstelių sultyse. Jie lemia daugelio daržovių (ridikėlių, ropių, kopūstų), vaisių (vynuogių, slyvų, vyšnių), gėlių (rugiagėlių, pelargonijų, delfinijų, rožių, bijūnų) raudoną ir mėlyną spalvą. Kartais šie pigmentai užmaskuoja chlorofilą lapuose, pavyzdžiui, dekoratyviniame raudonajame kleve. Antocianinai nuspalvina rudens lapus ryškiai raudonai. Jie susidaro šaltu saulėtu oru, kai lapuose sustoja chlorofilo sintezė. Lapuose, kai nesusidaro antocianinai, sunaikinus chlorofilą, pastebimi geltonai oranžiniai chloroplastų karotenoidai. Lapai ryškiausios spalvos būna šaltą, giedrą rudenį.
Vakuolės dalyvauja makromolekulių sunaikinime, jų komponentų cirkuliacijoje ląstelėje. Ribosomos, mitochondrijos, plastidai, patenkantys į vakuoles, sunaikinami. Pagal šią virškinimo veiklą jas galima palyginti su lizosomomis – gyvūnų ląstelių organelėmis.
Vakuolės susidaro iš endoplazminio tinklo (tinklo)
Ribosomos. Mažos dalelės (17–23 nm), susidedančios iš maždaug vienodo kiekio baltymų ir RNR. Ribosomose aminorūgštys susijungia ir sudaro baltymus. Ląstelėse, kuriose vyksta aktyvi medžiagų apykaita, jų yra daugiau. Ribosomos yra ląstelės citoplazmoje laisvai arba prijungtos prie endoplazminio tinklo (80S). Jų taip pat yra branduolyje (80S), mitochondrijose (70S), plastiduose (70S).
Ribosomos gali sudaryti kompleksą, kuriame vienu metu vyksta identiškų polipeptidų sintezė, informacija apie kurią paimama iš vienos molekulės ir RNR. Toks kompleksas vadinamas poliribosomomis (polisomomis). Ląstelės, kurios sintetina baltymus dideliais kiekiais, turi didelę polisomų sistemą, kuri dažnai yra pritvirtinta prie išorinio branduolio apvalkalo paviršiaus.
Endoplazminis Tinklelis. Tai sudėtinga neriboto dydžio trimatė membraninė sistema. ER kontekste atrodo kaip dvi elementarios membranos su siaura skaidria erdve tarp jų. ER forma ir ilgis priklauso nuo ląstelės tipo, jos metabolinio aktyvumo ir diferenciacijos stadijos. Ląstelėse, kurios išskiria arba kaupia baltymus, ER yra plokščių maišelių arba cisternų formos, o prie išorinio paviršiaus yra pritvirtinta daug ribosomų. Šis tinklas vadinamas grubiu endoplazminiu tinkleliu. Lygus ER paprastai yra vamzdinės formos. Toje pačioje ląstelėje gali būti šiurkštus ir lygus endoplazminis tinklas. Paprastai tarp jų yra daug ryšių.
Endoplazminis tinklas veikia kaip ląstelės ryšio sistema. Jis yra susijęs su išoriniu branduolio apvalkalu. Tiesą sakant, šios dvi struktūros sudaro vieną membranų sistemą. Kai ląstelės dalijimosi metu plyšta branduolio apvalkalas, jo fragmentai primena ER fragmentus. Endoplazminis tinklas yra sistema, pernešanti medžiagas: baltymus, lipidus, angliavandenius į skirtingas ląstelės dalis. kaimyninių ląstelių endoplazminis tinklas yra sujungtas per citoplazmines sruogas - plazmodesmatas -, kurios praeina per ląstelių membranas.
Endoplazminis tinklas yra pagrindinė ląstelių membranų sintezės vieta. Kai kuriose augalų ląstelėse čia susidaro vakuolių ir mikrokūnų membranos, diktiozomų cisternos.
Goldžio kompleksas.Šis terminas vartojamas apibūdinti visas diktiozomas arba Golgi kūnus ląstelėje. Diktiosomos yra plokščių, disko formos pūslelių arba cisternų grupės, kurios išsišakoja išilgai kraštų į sudėtingą kanalėlių sistemą. Aukštesniųjų augalų diktiosomos susideda iš 4–8 cisternų, surinktų kartu.
Paprastai rezervuarų pakuotėje išskiriama besivystanti ir bręstanti pusė. formuojančių cisternų membranos savo struktūra primena ER membranas, o bręstančių cisternų membranos primena plazminę membraną.
Diktiosomos dalyvauja sekrecijoje, o daugumoje aukštesniųjų augalų – ląstelių membranų formavime. Diktiosomų sintetinami ląstelių sienelių polisacharidai kaupiasi pūslelėse, kurios vėliau atsiskiria nuo bręstančių cisternų. Šios sekrecinės pūslelės migruoja ir susilieja su CMP; o juose esantys polisacharidai yra įterpti į ląstelės membraną. Kai kurios medžiagos, besikaupiančios diktiozomose, susidaro kitose struktūrose, pavyzdžiui, ER, o po to pernešamos į diktioomas, kur jos modifikuojamos (modifikuojamos) prieš sekreciją. Pavyzdžiui, glikoproteinai yra svarbi ląstelės membranos statybinė medžiaga. Baltyminę dalį sintetina šiurkščiojo ER polisomos, angliavandenių dalį – diktiozomose, kur abi dalys susijungdamos sudaro glikoproteinus.
Membranos yra dinamiškos, judrios struktūros, nuolat keičiančios savo formą ir plotą. Endoplazminės sistemos samprata remiasi membranų mobilumu. Pagal šią koncepciją vidinės citoplazmos membranos, išskyrus mitochondrijų ir plastidžių membranas, yra viena visuma ir kyla iš endoplazminio tinklo. Iš endoplazminio tinklo per tarpinę pūslelių stadiją susidaro naujos diktiozomų cisternos, o sekrecinės pūslelės, kurios atsiskiria nuo diktiozomų, ilgainiui prisideda prie plazminės membranos susidarymo. Taigi, endoplazminis tinklas ir diktiosomos sudaro funkcinį vienetą, kuriame diktiosomos atlieka tarpinių struktūrų vaidmenį membranas, tokias kaip endoplazminis tinklas, paverčiant tokiomis membranomis kaip plazma. Audiniuose, kurių ląstelės auga ir dalijasi silpnai, membranos komponentai nuolat atnaujinami.
mikrovamzdeliai randama beveik visose eukariotinėse ląstelėse. Tai cilindrinės struktūros, kurių skersmuo yra apie 24 nm. Jų ilgis skiriasi. Kiekvienas kanalėlis sudarytas iš baltymo, vadinamo tubulinu, subvienetų. Subvienetai sudaro 13 išilginių gijų, supančių centrinę ertmę. Mikrovamzdeliai yra dinamiškos struktūros, jos reguliariai sunaikinamos ir susidaro tam tikrais ląstelių ciklo etapais. Jų surinkimas vyksta specialiose vietose, kurios vadinamos mikrovamzdelių organizavimo centrais. Augalų ląstelėse jie turi silpnai išreikštą amorfinę struktūrą.
Mikrovamzdelių funkcijos: dalyvauja formuojant ląstelės membraną; nukreipti diktiozomų burbulus į besiformuojančią membraną, pavyzdžiui, verpstės siūlus, kurie susidaro besidalijančioje ląstelėje; vaidina vaidmenį formuojant ląstelės plokštelę (pradinę ribą tarp dukterinių ląstelių). Be to, mikrovamzdeliai yra svarbi žvynelių ir blakstienų sudedamoji dalis, kurių judėjime jie atlieka svarbų vaidmenį.
mikrofilamentai, kaip ir mikrovamzdeliai, jų yra beveik visose eukariotinėse ląstelėse. Tai ilgi, 5–7 nm storio siūlai, susidedantys iš susitraukiančio baltymo aktino. Mikrofilamentų ryšuliai randami daugelyje aukštesniųjų augalų ląstelių. Matyt, jie vaidina svarbų vaidmenį citoplazmos srovėse. Mikrofilamentai kartu su mikrotubuliais sudaro lankstų tinklą, vadinamą citoskeletu.
Bazinė medžiaga ilgą laiką buvo laikomas vienalyčiu (homogenišku) daug baltymų turinčiu tirpalu, turinčiu nedidelį skaičių struktūrų ar net bestruktūrių. Tačiau šiuo metu, naudojant aukštos įtampos elektroninį mikroskopą, nustatyta, kad gruntinė medžiaga yra trimatė gardelė, sudaryta iš plonų (3–6 nm skersmens) gijų, užpildančių visą ląstelę. Kiti citoplazmos komponentai, įskaitant mikrovamzdelius ir mikrofilamentus, yra suspenduoti iš šios mikrotrabekulinės gardelės.
Mikrotrabekulinė struktūra yra baltymų gijų gardelė, tarp kurių esanti erdvė užpildyta vandeniu. Kartu su vandeniu grotelės yra gelio konsistencijos, gelis atrodo kaip želatininiai kūnai.
Organelės yra pritvirtintos prie mikrotrabekulinės gardelės. Grotelės užtikrina ryšį tarp atskirų ląstelės dalių ir nukreipia tarpląstelinį transportą.
Lipidų lašai- sferinės formos struktūros, suteikiančios augalo ląstelės citoplazmos granuliuotumą šviesos mikroskopu. Elektroninėse mikrografijose jie atrodo amorfiški. Labai panašūs, bet mažesni lašai randami plastiduose.
Lipidų lašai, painiodami juos su organelėmis, vadino juos sferosomomis ir tikėjo, kad juos supa vieno arba dvigubo sluoksnio membrana. Tačiau naujausi duomenys rodo, kad lipidų lašeliai neturi membranų, bet gali būti padengti baltymu.
Ergastinės medžiagos - tai yra protoplasto „pasyvieji produktai“: atsarginės medžiagos arba atliekos. Jie gali atsirasti ir išnykti skirtingais ląstelių ciklo laikotarpiais. Išskyrus krakmolo grūdelius, kristalus, antocianinų pigmentus ir lipidų lašelius. Tai yra dervos, dervos, taninai ir baltymai. Ergastinės medžiagos yra ląstelės membranos dalis, pagrindinė citoplazmos ir organelių, įskaitant vakuoles, medžiaga.
Vėliava ir blakstienos - tai plonos, į plaukus panašios struktūros, besitęsiančios nuo daugelio eukariotinių ląstelių paviršiaus. Jų skersmuo yra pastovus, tačiau ilgis svyruoja nuo 2 iki 150 mikronų. Tradiciškai ilgesnės ir kelios jų vadinamos žiuželiais, o trumpesnės ir gausesnės – blakstienomis. Nėra aiškių skirtumų tarp šių dviejų tipų struktūrų, todėl terminas flagellum vartojamas abiem.
Kai kuriuose dumbliuose ir grybuose žvyneliai yra judėjimo organai, kurių pagalba jie juda vandenyje. Augaluose (pavyzdžiui, samanose, kepenėliuose, paparčiuose, kai kuriuose gimnasėkliuose) žvyneliais yra tik lytinės ląstelės (gametos).
Kiekvienas žvynelis turi tam tikrą organizaciją. Išorinis 9 porų mikrovamzdelių žiedas supa du papildomus mikrovamzdelius, esančius žvynelio centre. Fermentų turinčios „rankenos“ tęsiasi iš kiekvienos išorinės poros vieno mikrotubulo. Tai yra pagrindinis 9+2 organizavimo modelis, randamas visose eukariotinių organizmų žvyneliuose. Manoma, kad žvynelių judėjimas pagrįstas mikrotubulių slydimu, o išorinės mikrovamzdelių poros juda viena iš kitos nesusitraukdamos. Mikrovamzdelių porų slydimas vienas kito atžvilgiu sukelia vietinį žvynelio lenkimą.
Žvyneliai „auga“ iš citoplazminių cilindrinių struktūrų, vadinamų baziniais kūnais, kurie taip pat sudaro bazinę žvynelio dalį. Baziniai kūnai turi vidinę struktūrą, panašią į žvynelinę, išskyrus tai, kad išoriniai kanalėliai yra išsidėstę trynukais, o ne poromis, o centrinių kanalėlių nėra.
Ląstelių sienelės. Ląstelės sienelė riboja protoplasto dydį ir neleidžia jam plyšti dėl vandens sugėrimo vakuolės.
Ląstelės sienelė atlieka specifines funkcijas, kurios svarbios ne tik ląstelei ir audiniui, kuriame yra ląstelė, bet ir visam augalui. Ląstelių sienelės atlieka esminį vaidmenį įsisavinant, pernešant ir pašalinant medžiagas, be to, jose gali koncentruotis lizosominė, arba virškinimo veikla.
Ląstelės sienelės komponentai. Tipiškiausias ląstelės sienelės komponentas yra celiuliozė, kuri daugiausia lemia jos architektūrą. Celiuliozės molekulės yra sudarytos iš pasikartojančių gliukozės molekulių, sujungtų galais. Ilgos plonos celiuliozės molekulės sujungiamos į 10–25 nm storio mikrofibriles. Mikrofibrilės susipina ir sudaro plonus siūlus, kurie savo ruožtu gali būti apvynioti vienas aplink kitą, tarsi sruogos virvelėje. Kiekviena tokia „virvė“ arba makrofibrilė yra apie 0,5 mikrono storio, o ilgis siekia 4 mikronus. Makrofibrilės yra tokios pat stiprios kaip vienodo dydžio plieninė viela.
Ląstelės sienelės celiuliozės karkasas užpildytas su juo susipynusiomis celiuliozės matricos molekulėmis. Jį sudaro polisacharidai, vadinami hemiceliuliozės, ir pektinai, arba pektinai, chemiškai labai artimi hemiceliuliozei.
Kitas ląstelės sienelės komponentas – ligninas – yra gausiausias polimeras augalų ląstelėse po celiuliozės. Ligninas padidina sienos standumą ir dažniausiai randamas ląstelėse, kurios atlieka atraminę arba mechaninę funkciją.
Kutinas, suberinas, vaškas – dažniausiai nusėda augalų apsauginių audinių lukštuose. Pavyzdžiui, kutinas randamas epidermio ląstelių membranose, o suberinas – antriniame apsauginiame audinyje – kamštelyje. Abi medžiagos randamos kartu su vašku ir neleidžia augalui pernelyg prarasti vandens.
Ląstelės sienelės sluoksniai. Augalų ląstelių sienelių storis labai skiriasi, priklausomai nuo ląstelių vaidmens augalo struktūroje ir pačios ląstelės amžiaus. Elektroniniu mikroskopu augalo ląstelės sienelėje matomi du sluoksniai: mediana (taip pat vadinama tarpląsteline medžiaga) ir pirminė ląstelės sienelė. Daugelis ląstelių kloja kitą sluoksnį – antrinę ląstelės sienelę. Vidutinė plokštė yra tarp gretimų ląstelių pirminių sienelių. Antrinę sienelę, jei yra, ląstelės protoplastas nusodina ant pirminės ląstelės sienelės vidinio paviršiaus.
Vidurinė plokštelė. Vidurinę plokštelę daugiausia sudaro pektinas. Ten, kur tarp dviejų naujai susidariusių ląstelių turėtų atsirasti ląstelės sienelė, pirmiausia pastebimas tankus endoplazminio tinklo kanalėlių rezginys ir Golgi aparato cisterna (diktiosomos). Tada šioje vietoje atsiranda burbuliukai, užpildyti pektinu (iš polisacharidų). Šie burbuliukai yra atskirti nuo Golgi aparato rezervuarų. Ankstyvojoje ląstelės sienelėje yra įvairių polisacharidų, iš kurių pagrindiniai yra pektinai ir hemiceliuliozė. Vėliau jis susideda iš tankesnių medžiagų – celiuliozės ir lignino.
Pirminė ląstelės sienelė. Tai yra celiuliozės apvalkalo sluoksnis, kuris nusėda prieš ląstelių augimą arba jo metu. Be celiuliozės, hemiceliuliozės ir pektino, pirminėse membranose yra glikoproteino. Pirminiai apvalkalai gali sudegti. Pektino komponentas suteikia plastiškumo, todėl pirminis apvalkalas gali išsiplėsti, kai šaknis, stiebas ar lapas pailgėja.
Aktyviai besidalijančios ląstelės (dauguma subrendusių ląstelių, dalyvaujančių fotosintezės, kvėpavimo ir sekrecijos procesuose) turi pirmines membranas. Tokios ląstelės, turinčios pirminę membraną ir gyvą protoplastą, gali prarasti jiems būdingą formą, dalytis ir diferencijuotis į naują ląstelių tipą. Jie dalyvauja žaizdų gijime ir audinių regeneracijoje augaluose.
Pirminės ląstelių membranos nėra vienodo storio per visą ilgį, tačiau turi plonas dalis, kurios vadinamos pirminių porų laukais. Citoplazmos gijos arba plazmodesmata, jungiančios gretimų ląstelių protoplastus, dažniausiai praeina per pirminius porų laukus.
antrinė ląstelės sienelė. Nepaisant to, kad daugelis augalų ląstelių turi tik pirminę sienelę, kai kuriose, protoplastas nusodina antrinę sienelę link ląstelės centro. Paprastai tai įvyksta sustojus ląstelių augimui ir pirminės membranos plotas nebedidėja. Dėl šios priežasties antrinis apvalkalas skiriasi nuo pirminio. Antrinės membranos ypač reikalingos specializuotoms ląstelėms, kurios stiprina augalą ir praleidžia vandenį. Nusodinus antrinę membraną, šių ląstelių protoplastas, kaip taisyklė, miršta. Antriniuose apvalkaluose celiuliozės yra daugiau nei pirminiuose, juose nėra pektino medžiagų ir glikoproteinų. Antrinė membrana sunkiai ištempiama, jos matrica susideda iš hemiceliuliozės.
Antriniame apvalkale galima išskirti tris sluoksnius – išorinį, vidurinį ir vidinį (S1, S2, S3). Sluoksniuota antrinių apvalkalų struktūra žymiai padidina jų stiprumą. Celiuliozės mikrofibrilės antrinėje membranoje nusėda tankiau nei pirminėje. Ligninas yra įprastas antrinių medienos odelių komponentas.
Poros kontaktuojančių ląstelių membranose yra viena priešais kitą. Dvi priešingos poros ir porų membrana sudaro porų porą. Ląstelėse su antrine membrana yra du pagrindiniai porų tipai: paprastos ir apvaduotos. Apribotose porose antrinė sienelė kabo virš porų ertmės. Paprastose porose taip nėra.
Ląstelių sienelių augimas. Kai ląstelė auga, didėja ląstelės membranos storis ir plotas. Lukšto tempimas yra sudėtingas procesas. Jį kontroliuoja protoplastas ir reguliuoja hormonas auksinas.
Ląstelėse, kurios auga tolygiai visomis kryptimis, miofibrilių nusėdimas yra atsitiktinis. Šios miofibrilės sudaro netaisyklingą tinklą. Tokios ląstelės buvo aptiktos stiebo šerdyje, saugojimo audiniuose ir ląstelių auginimo in vitro metu. Pailgėjusiose ląstelėse šoninių apvalkalų miofibrilės nusėda stačiu kampu pailgėjimo ašiai.
Matricinės medžiagos – pektinai, hemiceliuliozės ir glikoproteinai į membraną pernešami diktiozomų pūslelėmis. Tuo pačiu pektinai labiau būdingi augančioms ląstelėms, o neaugančiose ląstelėse vyrauja hemiceliuliozės.
Celiuliozės mikrofibrilės sintezuojamos ląstelės paviršiuje, padedant fermentų kompleksui, susijusiam su plazmine membrana. Mikrofibrilių orientaciją kontroliuoja mikrotubulės, esančios šalia vidinio plazmos membranos paviršiaus.
Plazmodezma. Tai ploni citoplazmos siūlai, jungiantys kaimyninių ląstelių protoplastus. Plazmodesmatos arba praeina pro ląstelės sienelę bet kur, arba yra susitelkusios pirminiuose porų laukuose arba membranose tarp porų porų. Elektroniniu mikroskopu plazmodesmata atrodo kaip siauri kanalai, iškloti plazmos membrana. Išilgai kanalo ašies nuo vienos ląstelės iki kitos tęsiasi mažesnis cilindrinis vamzdelis – desmotubulas, kuris bendrauja su abiejų gretimų ląstelių endoplazminiu tinklu. Daugelis plazmodesmų susidaro ląstelių dalijimosi metu, kai besivystančios ląstelės plokštelė paima vamzdinį endoplazminį tinklą. Plazmodezmos taip pat gali susidaryti nesidalijančių ląstelių membranose. Šios struktūros užtikrina efektyvų tam tikrų medžiagų perkėlimą iš ląstelės į ląstelę.
Ląstelių dalijimasis. Daugialąsčiuose organizmuose ląstelių dalijimasis kartu su jų dydžio padidėjimu yra viso organizmo augimo būdas. Dalijimosi metu susidariusios naujos ląstelės savo struktūra ir funkcijomis yra panašios tiek į pirminę, tiek į viena kitą. Eukariotų dalijimosi procesą galima suskirstyti į du iš dalies persidengiančius etapus: mitozę ir citokinezę.
Mitozė – tai dviejų dukterinių branduolių susidarymas iš vieno branduolio, morfologiškai ir genetiškai lygiaverčiai vienas kitam. Citokinezė – tai ląstelės citoplazminės dalies dalijimasis, kai susidaro dukterinės ląstelės.
Ląstelių ciklas. Gyva ląstelė pereina iš eilės įvykių, kurie sudaro ląstelės ciklą. Pačio ciklo trukmė skiriasi priklausomai nuo ląstelės tipo ir išorinių veiksnių, tokių kaip temperatūra ar maistinių medžiagų prieinamumas. Paprastai ciklas skirstomas į tarpfazę ir keturias mitozės fazes.
Tarpfazė. Laikotarpis tarp nuoseklių mitozinių dalijimų.
Tarpfazė yra padalinta į tris periodus, žymimus kaip G1, S, G2.
G1 periodu, kuris prasideda po mitozės. Per šį laikotarpį padidėja citoplazmos kiekis, įskaitant įvairius organelius. Be to, pagal dabartinę hipotezę G1 periodo metu sintetinamos medžiagos, kurios arba stimuliuoja, arba slopina S periodą ir likusį ciklą, taip nulemdamos dalijimosi procesą.
S laikotarpis seka G1 periodą, tuo metu genetinė medžiaga (DNR) yra dubliuojama.
Per G2 laikotarpį, einantį po S, susidaro struktūros, kurios tiesiogiai dalyvauja mitozėje, pavyzdžiui, veleno komponentai.
Kai kurios ląstelės praeina neribotą ląstelių ciklų skaičių. Tai vienaląsčiai organizmai ir kai kurios aktyvaus augimo zonų ląstelės (meristemos). Kai kurios specializuotos ląstelės po brendimo praranda gebėjimą daugintis. Trečioji ląstelių grupė, pavyzdžiui, formuojančios žaizdos audinį (kalusą), gebėjimą dalytis išlaiko tik ypatingomis sąlygomis.
mitozė, arba branduolio dalijimasis. Tai nenutrūkstamas procesas, suskirstytas į keturias fazes: profazę, metafazę, anafazę, telofazę. Dėl mitozės tarpfazėje padvigubėjusi genetinė medžiaga po lygiai pasiskirsto tarp dviejų dukterinių branduolių.
Vienas iš pirmųjų ląstelių perėjimo prie dalijimosi požymių yra siauros žiedinės mikrovamzdelių juostos atsiradimas tiesiai po plazmos membrana. Šis gana tankus diržas supa branduolį būsimojo mitozinio veleno pusiaujo plokštumoje. Kadangi jis pasirodo prieš profazę, jis vadinamas priešfazės juosta. Jis išnyksta po mitozinio verpstės, dar ilgai prieš atsirandant vėlyvojoje ląstelės plokštelės telofazėje, kuri auga nuo centro iki periferijos ir susilieja su motininės ląstelės membrana toje srityje, kurią anksčiau užėmė priešfazinis diržas.
Profazė. Profazės pradžioje chromosomos primena ilgus siūlus, išsibarsčiusius branduolio viduje. Tada, trumpėjant ir storėjant sruogoms, galima pastebėti, kad kiekviena chromosoma susideda ne iš vienos, o iš dviejų susipynusių sruogų, vadinamų chromatidėmis. Vėlyvojoje fazėje dvi sutrumpintos suporuotos kiekvienos chromosomos chromatidės yra lygiagrečiai, sujungtos siaura atkarpa, vadinama centromera. Jis turi tam tikrą vietą kiekvienoje chromosomoje ir padalija chromosomą į dvi skirtingo ilgio šakas.
Mikrovamzdeliai išsidėstę lygiagrečiai branduolio paviršiui išilgai veleno ašies. Tai yra ankstyviausias mitozinio veleno surinkimo pasireiškimas.
Profazės pabaigoje branduolys palaipsniui praranda aiškų kontūrą ir galiausiai išnyksta. Netrukus po to branduolinis apvalkalas taip pat suyra.
Metafazė. Metafazės pradžioje verpstė, kuri yra trimatė struktūra, plačiausia per vidurį ir siaurėjanti link polių, užima vietą, kurią anksčiau užėmė branduolys. Verpstės pluoštai yra mikrovamzdelių pluoštai. Metafazės metu chromosomos, kurių kiekviena susideda iš dviejų chromatidžių, yra išdėstytos taip, kad jų centromerai būtų suklio pusiaujo plokštumoje. Kiekviena chromosoma savo centromeru yra pritvirtinta prie veleno sriegių. Tačiau kai kurios sruogos pereina iš vieno poliaus į kitą neprisirišusios prie chromosomų.
Kai visos chromosomos yra pusiaujo plokštumoje, metafazė baigiama. Chromosomos yra pasirengusios dalytis.
Anafazė. Kiekvienos chromosomos chromatidės išsiskiria. Dabar tai yra dukterinės chromosomos. Visų pirma, centromera dalijasi, o dvi dukterinės chromosomos traukiamos į priešingus polius. Šiuo atveju centromerai juda į priekį, o chromosomų rankos išsitiesia už nugaros. Prie chromosomų prisirišę verpstės siūlai sutrumpėja, prisidedant prie chromatidų atsiskyrimo ir dukterinių chromosomų judėjimo priešingomis kryptimis.
Telofazė. Telofazėje baigiasi dviejų identiškų chromosomų grupių atskyrimas, o aplink kiekvieną iš jų susidaro branduolinė membrana. Šiurkštus tinklelis aktyviai dalyvauja čia. Dingsta veleno aparatas. Telofazės metu chromosomos praranda kontūrų aiškumą, išsitempia, vėl virsta plonais siūlais. Branduoliai atkuriami. Kai chromosomos tampa nematomos, mitozė baigiasi. Du dukteriniai branduoliai patenka į interfazę. Jie yra genetiškai lygiaverčiai vienas kitam ir pirminiam branduoliui. Tai labai svarbu, nes genetinė programa, o kartu ir visi požymiai, turi būti perduota dukteriniams organizmams.
Mitozės trukmė skirtinguose organizmuose skiriasi ir priklauso nuo audinio tipo. Tačiau profazė yra ilgiausia, o anafazė – trumpiausia. Šaknies galiuko ląstelėse profazės trukmė yra 1–2 valandos; metafazė - 5 - 15 min; anafazė - 2 - 10 min; telofazė - 10 - 30 min. Tarpfazės trukmė nuo 12 iki 30 valandų.
Daugelyje eukariotinių ląstelių mikrotubulų organizavimo centrai, atsakingi už mitozinio veleno susidarymą, yra susiję su centriolais.
Citokinezė. Tai yra citoplazmos dalijimosi procesas. Daugumoje organizmų ląstelės dalijasi atitraukdamos ląstelės membraną ir suformuodamos dalijimosi vagą, kuri palaipsniui gilėja, suspaudžiant likusius mitozinio verpstės siūlus.Visuose augaluose (briofituose ir kraujagysliniuose) ir kai kuriuose dumbliuose ląstelės dalijasi dėl to, kad susidaro ląstelių plokštelė.
Ankstyvojoje telofazėje tarp dviejų dukterinių branduolių susidaro statinės formos skaidulų sistema, vadinama fragmoplastu. Fragmoplasto pluoštai, kaip ir mitozinio veleno pluoštai, susideda iš mikrovamzdelių. Fragmoplasto pusiaujo plokštumoje atsiranda nedideli lašeliai. Jie susilieja ir sudaro ląstelės plokštelę, kuri auga tol, kol pasiekia besidalijančios ląstelės sienelę. Tai užbaigia dviejų dukterinių ląstelių atskyrimą. Susiliejantys lašeliai yra burbuliukai, atitrūkę nuo Golgi aparato. Iš esmės juose yra pektino medžiagų, iš kurių susidaro vidurinė plokštelė. Pūslelinės membranos dalyvauja kuriant plazminę membraną abiejose plokštelės pusėse. Tuo pačiu metu iš vamzdinio endoplazminio tinklo fragmentų susidaro plazmodesmata.
Susidarius vidutinei plokštelei, kiekvienas protoplastas ant jos nusodina pirminę membraną. Be to, kiekviena dukterinė ląstelė aplink visą protoplastą nusodina naują membranos sluoksnį, kuris tęsia membraną, kuri atsirado iš ląstelės plokštelės. Augant dukterinėms ląstelėms, pradinis pirminės ląstelės apvalkalas sunaikinamas.
Įvairūs eukariotų mitozės tipai
Aukščiau aprašytas ląstelių dalijimasis augaluose, taip pat ir gyvūnuose, nėra vienintelė netiesioginio ląstelių dalijimosi forma. Paprasčiausias mitozės tipas yra pleuromitozė. Jis primena dvejetainį prokariotinių ląstelių dalijimąsi, kai nukleoidai po replikacijos lieka susieti su plazmos membrana. Membrana pradeda augti tarp DNR surišimo taškų ir taip pasklinda chromosomas į skirtingas ląstelės dalis. Po to, susiformuojant ląstelių susiaurėjimui, kiekviena iš DNR molekulių bus naujoje atskiroje ląstelėje.
Būdingas eukariotinių ląstelių dalijimasis yra iš mikrovamzdelių sudaryto verpstės susidarymas. At uždara pleuromitozė(vadinama uždara, nes chromosomų divergencija vyksta nepažeidžiant branduolio apvalkalo) kaip mikrovamzdelių organizavimo centrai (MCMT) dalyvauja ne centrioliai, o kitos struktūros, esančios vidinėje branduolio membranos pusėje. Tai vadinamieji neapibrėžtos morfologijos poliniai kūnai, iš kurių tęsiasi mikrovamzdeliai. Tokių kūnų yra du. Jie skiriasi vienas nuo kito neprarasdami ryšio su branduolio apvalkalu. Dėl to susidaro dvi pusverpstės, susijusios su chromosomomis. Visas mitozinio veleno formavimosi ir chromosomų segregacijos procesas šiuo atveju vyksta po branduolio apvalkalu. Šio tipo mitozė pasitaiko tarp pirmuonių ir yra plačiai paplitusi tarp grybų (chitridijų, zygomicetų, mielių, oomicetų, askomicetų, miksomicetų ir kt.). atitinka formas pusiau uždara pleuromitozė kai susidariusios verpstės poliuose sunaikinamas branduolio apvalkalas.
Kita mitozės forma yra ortomitozė.Šiuo atveju COMT yra citoplazmoje, nuo pat pradžių susidaro ne pusverpstės, o dvipolis velenas. Yra trys formos ortomitozė ( normali mitozė) pusiau uždara Ir uždaryta. Esant pusiau uždarai ortomitozei, citoplazmoje esančio TsOMT pagalba susidaro bisimetrinis velenas, branduolinis apvalkalas išsaugomas per visą mitozės laikotarpį, išskyrus poliarines zonas. Granuliuotos medžiagos ar net centriolių masės gali būti aptiktos kaip COMT. Ši mitozės forma randama žaliųjų, rudųjų ir raudonųjų dumblių zoosporose, kai kuriuose žemesniuosiuose grybuose ir gregarinuose. Esant uždarai ortomitozei, visiškai išsaugoma branduolinė membrana, kurioje susidaro tikras velenas. Karioplazmoje susidaro mikrovamzdeliai, rečiau išauga iš vidinės TsOMT, kuri nesusijusi (skirtingai nei pleuromitozė) su branduoline membrana. Šio tipo mitozė būdinga blakstienų mikrobranduolių dalijimuisi, tačiau gali atsirasti ir pirmuoniams. Atviros ortomitozės atveju branduolinis apvalkalas visiškai suyra. Šis ląstelių dalijimosi tipas būdingas gyvūnų organizmams, kai kuriems pirmuoniams ir aukštesnių augalų ląstelėms. Šią mitozės formą savo ruožtu atstovauja astraliniai ir anastraliniai tipai.
Iš trumpai apžvelgtos medžiagos matyti, kad pagrindinis mitozės bruožas apskritai yra dalijimosi verpstės struktūrų atsiradimas, kurios susidaro ryšium su CMT, kurios skiriasi savo struktūra.
Mitozinės figūros morfologija
Mitozinis aparatas ypač gerai išreiškiamas mitozės metafazės stadijoje. Metafazėje, ląstelės pusiaujo plokštumoje, išsidėsčiusios chromosomos, iš kurių į priešingas puses tęsiasi vadinamieji verpstės siūlai, susiliejantys dviejuose skirtinguose mitozinės figūros poliuose. Taigi mitozinis velenas yra chromosomų, polių ir skaidulų rinkinys. Verpstės skaidulos yra pavieniai mikrovamzdeliai arba jų ryšuliai. Mikrovamzdeliai prasideda nuo veleno polių, o kai kurie jų eina į centromerus kur yra įsikūrę kinetochores chromosomų (kinetochorų mikrotubulių), kai kurios pereina toliau link priešingo poliaus, bet jo nepasiekia. Jie vadinami „tarppoliais mikrotubulais“. Iš polių nukrypsta grupė radialinių mikrovamzdelių, aplink juos suformuodami struktūrą, primenančią „spinduliuojantį švytėjimą“ – tai astraliniai mikrovamzdeliai.
Pagal morfologiją mitozinės figūros skirstomos į astralinius ir anastralinius tipus.
Astralinis verpstės tipas, arba konvergentas, pasižymi tuo, kad jo polius vaizduoja maža zona, į kurią susilieja (susilenkia) mikrovamzdeliai. Paprastai centrosomos, kuriose yra centriolių, yra astralinių verpsčių poliuose. Tačiau yra žinomi centriolinių astralinių mitozių atvejai (kai kurių bestuburių mejozės metu). Be to, pastebimi radialiniai mikrovamzdeliai, sklindantys iš polių, kurie nėra veleno dalis, bet sudaro žvaigždžių zonas - citasters. Šio tipo mitozinis dalijimasis primena hantelį. Mitozinės figūros anastrialinio tipo polių nėra. Verpstės poliarinės sritys čia yra plačios, jos vadinamos poliniais gaubteliais, jose yra centriolių. Šiuo atveju veleno pluoštai nenukrypsta nuo vieno taško, o nukrypsta plačiu priekiu (nukrypsta) nuo visos poliarinių dangtelių zonos. Šio tipo verpstės būdingos besidalijančioms aukštesniųjų augalų ląstelėms, tačiau jos gali būti ir aukštesniuosiuose gyvūnuose. Ankstyvoje žinduolių embriogenezėje centriolinės (divergentinės) mitozės stebimos kiaušialąstės brendimo dalijimosi metu ir I bei II zigotos dalijimosi metu. Tačiau jau trečiojo ląstelių dalijimosi metu ir visose vėlesnėse ląstelėse jie dalijasi dalyvaujant astraliniams verpstams, kurių poliuose visada randama centriolių. Apskritai visoms mitozės formoms chromosomos su jų kinetoforais, poliniais kūnais (centrosomomis) ir verpstės skaidulomis išlieka įprastos struktūros.
Centromerai ir kinetochorai
Centromerai gali būti skirtingos lokalizacijos išilgai chromosomų ilgio. Holocentriniai centromerai susidaro, kai mikrovamzdeliai susijungia per visą chromosomos ilgį (kai kurie vabzdžiai, nematodai, kai kurie augalai). Monocentriniai centromerai – kai mikrovamzdeliai yra susieti su chromosomomis vienoje vietoje. monocentriniai centromerai gali būti taškiniai (pavyzdžiui, kai kuriose pumpurinėse mielėse), kai tik vienas mikrovamzdelis priartėja prie kinetochoro, ir zoninės, kai mikrovamzdelių pluoštas artėja prie kompleksinio kinetochoro. Nepaisant centromerų zonų įvairovės, visos jos yra susijusios su sudėtinga kinetochoro struktūra, kurios struktūra ir funkcijos yra labai panašios visuose eukariotuose. Kinetochorai yra specialios baltymų struktūros, dažniausiai išsidėsčiusios chromosomų centromerinėse zonose. Tai sudėtingi kompleksai, susidedantys iš daugelio baltymų. morfologiškai jie yra labai panašūs, turi tą pačią struktūrą – nuo diatomų iki žmonių. Tai yra trijų sluoksnių struktūros: vidinis tankus sluoksnis, esantis greta chromosomos kūno, vidurinis laisvas sluoksnis ir išorinis tankus sluoksnis. Daugelis fibrilių tęsiasi nuo išorinio sluoksnio, sudarydamos vadinamąjį pluoštinį kinetochoro vainiką. Bendra forma kinetochorai atrodo kaip plokštelės ar diskai, esantys pirminio chromosomos susiaurėjimo zonoje, centromeroje. Paprastai kiekvienoje chromosomoje arba chromatidėje yra vienas kinetochoras. Prieš anafazę kiekvienoje seserinėje chromatidėje yra kinetochorai, kurių kiekviena bendrauja su savo mikrotubulių pluoštu. Augaluose kinetochoras atrodo ne kaip plokštelės, o kaip pusrutuliai. Funkcinis kinetochorų vaidmuo yra surišti seserines chromatides viena su kita, fiksuoti mitozinius mikrotubulus, reguliuoti chromosomų atskyrimą ir faktiškai perkelti chromosomas mitozės metu, dalyvaujant mikrotubulams. Apskritai, baltymų struktūros, kinetochorai S periode dvigubėja, lygiagrečiai chromosomų padvigubėjimui. Tačiau jų baltymai yra chromosomose visais ląstelių ciklo laikotarpiais.
AUGALŲ AUDINIAI
Ugdomieji audiniai (meristemos)
Ugdomieji audiniai augalų organizme išsidėstę skirtingose vietose, todėl skirstomi į tokias grupes (0;1 pav.).
Viršūninės (apikos) meristemos išsidėsčiusios ašinių organų – stiebo, šaknies – viršūnėse arba viršūnėse. Šių meristemų pagalba augalų vegetatyviniai organai vykdo savo ilgį.
Šoninės meristemos būdingos ašiniams organams. Ten jie yra koncentriškai, sankabos pavidalu.
Tarpkalarinės arba tarpkalinės meristemos kyla iš viršūninių meristemų. Tai ląstelių grupės, kurios dar nesugeba daugintis, bet yra išėjusios į diferenciacijos kelią. Tarp jų nėra pradinių ląstelių, tačiau yra daug specializuotų.
Žaizdų meristemos atkuria pažeistą kūno dalį. Regeneracija prasideda dediferenciacija, tai yra atvirkštinis vystymasis nuo specializuotų ląstelių iki meristematinių. Jie virsta felogenu, kuris sudaro kamštį, dengiantį žaizdos paviršių. Dediferencijuotos ląstelės, besidalijančios, gali suformuoti purų parenchiminį audinį – kaliusą. Iš jo tam tikromis sąlygomis susidaro augalų organai.
Integumentiniai audiniai
Jie veikia kaip ribinis barjeras, atskiriantys apatinius audinius nuo aplinkos. Pirminius augalo sluoksnius sudaro tik gyvos ląstelės. Antrinis ir tretinis apvalkalas – daugiausia iš mirusiųjų su storomis ląstelių sienelėmis.
Pagrindinės vientisųjų audinių funkcijos:
augalo apsauga nuo išdžiūvimo;
apsauga nuo kenksmingų mikroorganizmų patekimo;
apsauga nuo saulės nudegimo;
apsauga nuo mechaninių pažeidimų;
medžiagų apykaitos tarp augalo ir aplinkos reguliavimas;
dirginimo suvokimas.
Pirminis vientisas audinys yra epidermis, epidermis. Susideda iš gyvų ląstelių. Susidaro iš viršūninių meristemų. Apima jaunus augančius stiebus ir lapus.
Epidermis susiformavo augaluose, kai jie patenka iš vandens buveinių į sausumą, kad jis neišdžiūtų. Be stomos, visos epidermio ląstelės yra glaudžiai tarpusavyje susijusios. Išorinės pagrindinių ląstelių sienelės yra storesnės nei likusios. Visas paviršius padengtas kutino ir augalinio vaško sluoksniu. Šis sluoksnis vadinamas odele (oda). Jo nėra ant augančių šaknų ir povandeninių augalų dalių. Išdžiūvus gerokai susilpnėja odelės pralaidumas.
Be pagrindinių ląstelių, epidermyje yra ir kitų, ypač plaukelių arba trichomų. Jie yra vienaląsčiai ir daugialąsčiai (2 pav.). Funkciškai jie padidina epidermio paviršių, pavyzdžiui, šaknų augimo zonoje, tarnauja kaip mechaninė apsauga, prilimpa prie atramos, mažina vandens praradimą. Nemažai augalų turi liaukų plaukelius, pavyzdžiui, dilgėlės.
Tik aukštesni augalai epidermyje turi stomatus, kurios reguliuoja vandens ir dujų mainus. Jeigu odelės nėra, vadinasi, ir stomatos nereikia. Stomatai yra ląstelių grupė, sudaranti stomatinį aparatą, susidedantį iš dviejų užpakalinių ląstelių ir greta esančių epidermio ląstelių - šoninių ląstelių. Jie skiriasi nuo pagrindinių epidermio ląstelių (3 pav). Apsauginės ląstelės skiriasi nuo aplinkinių ląstelių forma ir daugybe chloroplastų bei netolygiai sustorėjusių sienelių. Tie, kurie susiduria vienas su kitu, yra storesni nei kiti (4 pav.). Tarp apsauginių ląstelių susidaro stomatalinis tarpas, kuris veda į substomatalinę erdvę, vadinamą substomataline ertme. Apsauginės ląstelės yra labai fotosintetinės. Juose yra daug atsarginio krakmolo ir daug mitochondrijų.
Stomatų skaičius ir pasiskirstymas, stomatalinių aparatų tipai skirtinguose augaluose labai skiriasi. Šiuolaikiniuose briofituose stomatų nėra. Fotosintezę juose vykdo gametofitų karta, o sporofitai negali egzistuoti savarankiškai.
Paprastai stomos yra apatinėje lapo pusėje. Vandens paviršiuje plūduriuojančiuose augaluose – viršutiniame paviršiuje. Javų lapuose stomatai dažnai būna vienodai iš abiejų pusių. Tokie lapai apšviečiami palyginti tolygiai. Ant 1 mm2 paviršiaus gali būti nuo 100 iki 700 stomatų.
Antrinis vidinis audinys (peridermas). Šis audinys pakeičia epidermį, kai vienmečių ūglių žalia spalva pasikeičia į rudą. Jis yra daugiasluoksnis ir susideda iš centrinio kambinių ląstelių sluoksnio – felogeno. Felogeno ląstelės, besidalijančios, išorėje kloja sluoksnį fellemo, o viduje – felodermą. (5 pav.).
Phelema arba kamštiena. Iš pradžių jis susideda iš gyvų plonasienių ląstelių. Laikui bėgant jų sienelės impregnuojamos suberinu ir augaliniu vašku ir miršta. Ląstelės turinys užpildytas oru.
Fellema funkcijos:
apsaugo nuo drėgmės praradimo;
apsaugo augalą nuo mechaninių pažeidimų;
apsaugo nuo patogenų;
užtikrina šilumos izoliaciją, nes ląstelės užpildomos oru.
Felogeno ląstelės, esančios pačiame epidermyje, apatiniame poepiderminiame sluoksnyje, rečiau giliuose pirminės žievės sluoksniuose, yra pirminės žievės pagrindas.
Kamštienos sluoksnis nėra nuolatinis. Joje atsiranda lūžių, kurios susisiekia su šalia esančiomis tarpląstelinėmis erdvėmis. Tuo pačiu metu paviršiuje susidaro maži gumbai – lęšiai, kurie tarpląstelines erdves susisiekia su atmosferos oru. (6,7 pav.).
Rudenį po lęšiais esantis felogenas kloja užkamšytų ląstelių sluoksnį, kuris labai sumažina transpiraciją, bet visiškai jos neatmeta. Pavasarį šis sluoksnis suyra iš vidaus. Ant šviesios beržo žievės lęšiai aiškiai matomi tamsių brūkšnelių pavidalu.
Tretinis vientisas audinys (pluta) taip pat būdingas tik sumedėjusioms augalų formoms.
Felogenas pakartotinai nusėda gilesniuose žievės sluoksniuose. Už jos ribų esantys audiniai ilgainiui miršta ir susidaro pluta. Jo ląstelės yra negyvos ir negali ištempti. Tačiau gyvos ląstelės, esančios giliau, dalijasi, todėl padidėja skersinis stiebo dydis. Laikui bėgant, išorinis plutos sluoksnis lūžta. Tokio tarpo atsiradimo laikas yra gana pastovi atskirų augalų reikšmė. Obelėje tai atsitinka septintaisiais gyvenimo metais, skroblėje - penkiasdešimtaisiais. Kai kuriose rūšyse jis visai nevyksta. Pagrindinė pilingo funkcija yra apsauga nuo mechaninių ir terminių pažeidimų.
Parenchima
Tai specializuotų audinių grupė, užpildanti augalo kūno viduje esančias erdves tarp laidžių ir mechaninių audinių. (8 pav.). Dažniau parenchimos ląstelės yra suapvalintos, rečiau pailgos formos. Būdingas išsivysčiusių tarpląstelinių erdvių buvimas. Tarpai tarp ląstelių kartu sudaro transporto sistemą – apoplastą. Be to, tarpląstelinės erdvės sudaro augalo „vėdinimo sistemą“. Per stomatas arba lenticelius jie susijungia su atmosferos oru ir užtikrina optimalią dujų sudėtį augalo viduje. Išvystytos tarpląstelinės erdvės ypač reikalingos augalams, augantiems užmirkusioje dirvoje, kur sunku normaliai keistis dujomis. Ši parenchima vadinama aerenchima. (9 pav.).
Parenchimos elementai, užpildantys tarpus tarp kitų audinių, atlieka ir atramos funkciją. Parenchimos ląstelės yra gyvos, jos neturi storų ląstelių sienelių, kaip sklerenchima. Todėl mechanines savybes suteikia turgoras. Jei vandens kiekis sumažėja, tai sukelia plazmolizę ir augalo vytimą.
Asimiliacinę parenchimą sudaro plonasienės ląstelės, turinčios daug tarpląstelinių erdvių. Šios struktūros ląstelėse yra daug chloroplastų, todėl ji vadinama chlorenchima. Chloroplastai išsidėstę išilgai sienos, vienas kito neužtemdydami. Asimiliacinėje parenchimoje vyksta fotosintezės reakcijos, kurios aprūpina augalą organinėmis medžiagomis ir energija. Fotosintezės procesų rezultatas – galimybė Žemėje egzistuoti visiems gyviems organizmams.
Asimiliaciniai audiniai yra tik apšviestose augalo dalyse, juos nuo aplinkos skiria skaidrus epidermis. Jei epidermį pakeičia nepermatomi antriniai integumentiniai audiniai, asimiliacinė parenchima išnyksta.
Sandėliavimo parenchima tarnauja kaip organinių medžiagų, kurių augalų organizmas laikinai nenaudoja, talpykla. Iš esmės bet kuri ląstelė, turinti gyvą protoplastą, gali nusodinti organines medžiagas įvairių rūšių inkliuzų pavidalu, tačiau kai kurios ląstelės tam specializuojasi. (10, 11 pav.). Energetiniai junginiai nusėda tik vegetacijos metu, vartojami ramybės laikotarpiu ir ruošiantis kitam vegetacijos sezonui. Todėl rezervinės medžiagos vegetatyviniuose organuose nusėda tik daugiamečiuose augaluose.
Atsargų talpykla gali būti paprasti organai (pabėgimas, šaknis), taip pat specializuoti (šakniastiebiai, gumbai, svogūnėliai). Visi sėkliniai augalai kaupia energetiškai vertingas medžiagas sėklose (skilčialapiuose, endosperme). Daugelis augalų sausringuose klimatuose kaupia ne tik organines medžiagas, bet ir vandenį. (12 pav.). Pavyzdžiui, alavijas kaupia vandenį mėsinguose lapuose, kaktusai – ūgliuose.
mechaniniai audiniai
Mechanines augalų ląstelių savybes suteikia:
kietas ląstelės apvalkalas;
turgoriškumas, tai yra ląstelių turgorinė būsena.
Nepaisant to, kad beveik visos audinių ląstelės turi mechaninių savybių, tačiau augale yra audinių, kurių mechaninės savybės yra pagrindinės. Tai yra kolenchima ir sklerenchima. Paprastai jie veikia sąveikaudami su kitais audiniais. Kūno viduje augalai sudaro savotišką rėmą. Todėl jie vadinami sustiprinimu.
Ne visi augalai turi vienodai aiškiai apibrėžtus mechaninius audinius. Vandens aplinkoje gyvenantiems augalams vidinės paramos reikia daug mažiau nei sausumos augalams. Priežastis ta, kad vandens augalams reikia mažiau vidinės paramos. Jų kūną daugiausia palaiko aplinkinis vanduo. Oras sausumoje tokios atramos nesukuria, nes jo tankis mažesnis nei vandens. Dėl šios priežasties specializuotų mechaninių audinių buvimas tampa aktualus.
Evoliucijos procese įvyko vidinių atraminių struktūrų tobulėjimas.
Kolenchima. Jį sudaro tik gyvos ląstelės, pailgos išilgai organo ašies. Šio tipo mechaniniai audiniai susidaro labai anksti, pirminio augimo laikotarpiu. Todėl svarbu, kad ląstelės išliktų gyvos ir išlaikytų galimybę ištempti kartu su šalia esančiomis tempimo ląstelėmis.
Kolenchimos ląstelių savybės:
netolygus lukšto sustorėjimas, dėl to kai kurios jo dalys lieka plonos, o kitos sustorėja;
lukštai nerūgsta.
Kolenchimos ląstelės yra skirtingos viena kitos atžvilgiu. Gretimose ląstelėse kampuose, nukreiptuose vienas į kitą, susidaro sustorėjimai ( ryžių. 13). Tokia kolenchima vadinama kampine kolenchima. Kitu atveju ląstelės yra išdėstytos lygiagrečiais sluoksniais. Ląstelių membranos, esančios priešais šiuos sluoksnius, yra stipriai sustorėjusios. Tai yra lamelinė kolenchima. Ląstelės gali išsidėstyti laisvai, su gausiomis tarpląstelinėmis erdvėmis – tai laisva kolenchima. Tokia kolenchima dažnai aptinkama augaluose užmirkusiose dirvose.
Kolenchima ypač svarbi jauniems augalams, žolinėms formoms, taip pat tose augalų dalyse, kuriose antrinis augimas nevyksta, pavyzdžiui, lapuose. Šiuo atveju jis klojamas labai arti paviršiaus, kartais tiesiog po epidermiu. Jei organas turi veidus, tada išilgai jų keteros randami stori kolenchimo sluoksniai.
Kolenchimos ląstelės veikia tik esant turgorui. Vandens trūkumas sumažina kolenchimo efektyvumą ir augalas laikinai nuvysta, pavyzdžiui, karštą dieną nusvyra agurkų lapai. Užpildžius ląsteles vandeniu, atkuriamos kolenchimos funkcijos.
Sklerenchima. Antrasis mechaninių audinių tipas. Skirtingai nuo kolenchimos, kur visos ląstelės yra gyvos, sklerenchimos ląstelės yra negyvos. Jų sienos labai storos. Jie atlieka mechaninę funkciją. Stiprus membranos sustorėjimas sukelia medžiagų transportavimo pažeidimą, dėl kurio protoplastas miršta ( ryžių. 14). Sklerenchimos ląstelių membranų lignifikacija įvyksta, kai augalo organas jau baigė augti. Todėl jie nebetrukdo aplinkinių audinių tempimui.
Priklausomai nuo formos, išskiriami du sklerenchimos ląstelių tipai – skaidulinės ir sklereidinės.
Pluoštai yra stipriai pailgos formos su labai storomis sienelėmis ir maža ertme. Jie yra šiek tiek mažesni nei medienos pluoštai. Dažnai po epidermiu susidaro išilginiai sluoksniai ir sruogos. Floemoje arba ksileme juos galima rasti pavieniui arba grupėmis. Floemoje jie vadinami karnienos pluoštais ( ryžių. 15), o ksileme - libriforminiai pluoštai ( ryžių. 16).
Sklereidai arba akmeninės ląstelės yra suapvalintos arba šakotos ląstelės su galingomis membranomis. Augalo kūne jų galima rasti pavieniui (palaikančios ląstelės) ( pav.17) arba grupės (18 pav.). Reikia pažymėti, kad mechaninės savybės labai priklauso nuo sklereidų vietos. Dalis sklereidų sudaro ištisinius sluoksnius, pavyzdžiui, riešutų kevaluose arba vaisių (kaulavaisių) kauliukuose.
išskyrimo audiniai
Bet kuri gyva ląstelė iš pradžių atlieka medžiagų pašalinimo funkciją, o transportavimas gali vykti tiek palei koncentracijos gradientą, tiek prieš gradientą. Medžiagos, kurios buvo susintetintos ląstelėje, gali būti pašalintos ir vėliau paveiks kitų ląstelių veiklą (fitohormonai, fermentai). Šis procesas vadinamas sekrecija. Tuo atveju, kai išskiriamos medžiagos, kurios yra ląstelės atliekos, procesas vadinamas išskyrimu. Nepaisant to, kad medžiagų išskyrimas į augalų ir gyvūnų ląsteles turi esminį panašumą, augalai turi nemažai savybių, atsirandančių dėl esminių gyvybinės veiklos skirtumų. Metabolinių reakcijų lygis augaluose yra daug mažesnis nei gyvūnų. Todėl išmetama proporcingai mažesni atliekų kiekiai. Kitas bruožas yra tai, kad augalai, savarankiškai sintetindami beveik visus būtinus organinius junginius, niekada nesudaro perteklinių jų atsargų. Išsiskyrusios medžiagos gali būti pradinė medžiaga kitoms reakcijoms (pavyzdžiui, CO2 ir H2O).
Jei gyvūnams toksinų išskyrimo procesas yra susijęs su jų pašalinimu iš organizmo, augalai gali to nedaryti, išskirdami nereikalingas medžiagas gyvame protoplaste (įvairių medžiagų pašalinimas į vakuolę), negyvose ląstelėse (daugumoje daugiamečio augalo audinių). augalas susideda iš tokių ląstelių), tarpląstelinėse erdvėse.
Išskiriamos medžiagos gali būti suskirstytos į dvi dideles grupes. Pirmoji grupė – organinės medžiagos, kurias tiesiogiai sintetina ląstelė (fermentai, polisacharidai, ligninai, terpenai, pastarieji yra eterinių aliejų ir dervų sudedamosios dalys). Antroji grupė – medžiagos, kurios iš pradžių į ląstelę patenka iš išorės ksilemo ar floemo transportavimo pagalba (vanduo, mineralinės druskos, aminorūgštys, monosacharidai ir kt.). Išskirta paslaptis retai būna vienalytė ir dažniausiai susideda iš mišinio, kuriame vienos medžiagos koncentracija yra didžiausia.
Augalai, skirtingai nei gyvūnai, neturi pilnos šalinimo sistemos. Yra tik specializuotos struktūros, išsibarsčiusios visame augale - idioblastai. Jas formuojančios ląstelės yra mažesnės nei šalia esančios parenchimos ląstelės. Jie turi elektronų tankią citoplazmą su išvystytais endoplazminio tinklo elementais ir Golgi kompleksu. Dažniausiai nėra išreikšta, yra centrinė vakuolė. Tarp jų ir kitų gyvų ląstelių šios ląstelės yra sujungtos daugybe plazmodesmų.
Priklausomai nuo vietos, šalinimo audiniai gali būti išoriniai ir vidiniai.
Išoriniai šalinimo audiniai. Liaukiniai plaukai yra epidermio dariniai. Morfologiškai jie yra kintami – gali turėti daugialąstę galvą, būti pailgos, skydo formos ant kojos ir kt. (19 pav.). Tai apima dilgėlių plaukelius.
Hidatodai – tai konstrukcijos, išskiriančios vandens perteklių esant žemai transpiracijai ir didelės drėgmės sąlygoms – gutacija. Šias struktūras sudaro bespalvių gyvų ląstelių grupės su plonomis sienelėmis – epitema. Šis audinys yra greta laidžiojo pluošto (20 pav.). Vanduo išleidžiamas per specialias vandens stomatas, kurios nuo įprastų stomatų skiriasi nejudrumu ir nuolat atsiveriančiu tarpu. Gutavimo skysčio sudėtis labai skiriasi nuo beveik gryno vandens iki labai sudėtingo medžiagų mišinio.
Nektarai. Daugeliui augalų būdingas skysčio, kuriame yra nuo 7 iki 87% mono- ir oligosacharidų, išsiskyrimas. Šį procesą vykdo specialios struktūros – nektarai. Priklausomai nuo vietos, žiedinės ar žiedinės, skiriami nektarai (21 pav.), taip pat augalai, esantys ant stiebų ir lapų – ekstrafloriniai, arba ekstrafloriniai nektarai. Nektaras gali būti nepakitusios floemo sultys, kurios per tarpląstelinius tarpus patenka į paviršių ir išskiriamos per stomatas. Sudėtingesnius nektarus sudaro liaukų parenchima, padengta epidermiu su liaukiniais plaukais. Nektarą išskiria epidermio ląstelės arba liaukų plaukai. Šiuo atveju išsiskiriantis nektaras skiriasi nuo floemų sulčių. Kadangi jame vyrauja gliukozė, fruktozė, sacharozė, o floemo sultyse – gliukozė. Jonų yra nedidelėmis koncentracijomis. Jonų yra nedidelėmis koncentracijomis. Norint pritraukti apdulkintojus, nektare gali būti steroidinių hormonų, būtinų vabzdžiams.
Virškinimo liaukos yra vabzdžiaėdžiuose augaluose (saulėraščiuose, nepentiuose ir kt.). Reaguodami į mažų bestuburių prisilietimą, jie išskiria paslaptį, kurioje yra hidrolizinių fermentų, skruzdžių, sviesto ir propiono rūgščių. Dėl to biopolimerai suskaidomi į monomerus, kuriuos augalas naudoja savo reikmėms.
Druskos liaukos vystosi augaluose, augančiuose druskingoje dirvoje. Šios liaukos yra ant lapų, o ištirpusios formos jonų perteklius iškeliamas į paviršių. Druska pirmiausia nusėda ant odelės, vėliau ją nuplauna lietus (22 pav.). Be to, daugelio augalų lapuose yra druskos plaukelių. Kiekvienas plaukas susideda iš dviejų ląstelių: viena sudaro galvą, kita - stiebą. Druskos kaupiasi galvos ląstelės vakuolėje, o kai jų koncentracija tampa didelė, galva nukrenta. Jo vietoje išauga nauja ląstelė, kuri taip pat kaupia druskas. Lapui augant gali kelis kartus nukristi ir formuotis galvutė. Druskos plaukų privalumas yra tas, kad išskirdami druskas jie netenka mažai vandens. Šis reiškinys labai svarbus augalams, augantiems druskingose dirvose, kur gėlo vandens trūksta.
Vidiniai šalinimo audiniai. Vidinės šalinimo struktūros, išsibarsčiusios po visą augalo kūną idioblastų pavidalu, nepašalina medžiagų iš organizmo, kaupdamos jas savyje. Idioblastai yra augalų ląstelės, kurios savo forma, struktūra ar turiniu skiriasi nuo kitų to paties audinio ląstelių. Priklausomai nuo kilmės skiriamos šizogeninės (gr. schiso – suskaidytos) ir lizogeninės (gr. lizė – tirpimas) išskiriamų medžiagų talpyklos (23 pav.).
Šizogeninės talpyklos. Tai daugiau ar mažiau plačios tarpląstelinės erdvės, užpildytos išskiriamomis medžiagomis. Pavyzdžiui, spygliuočių, aralijų, skėčių ir kt. Manoma, kad derva pasižymi baktericidinėmis savybėmis, atbaido žolėdžius, todėl augalai jiems nevalgomi.
Vietoje gyvų ląstelių susidaro lizigeninės talpyklos, kurios jose susikaupus medžiagoms žūva ir sunaikinamos. Tokius darinius galima pastebėti žievėje
AUGALŲ LĄSTELĖS STRUKTŪRA
Augalo ląstelė susideda iš daugiau ar mažiau standžių ląstelių sienelės Ir protoplastas. Ląstelės sienelė yra ląstelės sienelė ir citoplazminė membrana. Terminas protoplastas kilęs iš žodžio protoplazma, kuris nuo seno buvo vartojamas visiems gyviems dalykams apibūdinti. Protoplastas yra atskiros ląstelės protoplazma.
Protoplastas sudarytas iš citoplazma Ir branduoliai. Citoplazmoje yra organelių (ribosomų, mikrotubulių, plastidžių, mitochondrijų) ir membraninių sistemų (endoplazminis tinklas, diktiosomos). Citoplazma taip pat apima citoplazminę matricą ( malta medžiaga), kurioje panardintos organelės ir membranų sistemos. Citoplazma yra atskirta nuo ląstelės sienelės plazmos membrana, kuri yra elementari membrana. Skirtingai nuo daugelio gyvūnų ląstelių, augalų ląstelėse yra viena ar daugiau vakuolės. Tai skysčiu užpildytos pūslelės, apsuptos elementaria membrana ( tonoplastas).
Gyvoje augalo ląstelėje žemės medžiaga nuolat juda. Judėjimu, vadinamu citoplazmine srove arba ciklozė, dalyvauja organelės. Ciklozė palengvina medžiagų judėjimą ląstelėje ir jų mainus tarp ląstelės ir aplinkos.
Plazmos membrana. Tai dvisluoksnė fosfolipidinė struktūra. Augalų ląstelėms būdingos plazminės membranos invaginacijos.
Plazminė membrana atlieka šias funkcijas:
Dalyvauja medžiagų mainuose tarp ląstelės ir aplinkos;
Koordinuoja ląstelės sienelės celiuliozės mikrofibrilių sintezę ir surinkimą;
Perduoda hormoninius ir išorinius signalus, kurie kontroliuoja ląstelių augimą ir diferenciaciją.
Šerdis. Tai ryškiausia eukariotinės ląstelės citoplazmos struktūra. Branduolys atlieka dvi svarbias funkcijas:
Kontroliuoja gyvybinę ląstelės veiklą, nustatydama, kurie baltymai ir kokiu laiku turi būti sintetinami;
Saugo genetinę informaciją ir perduoda ją dukterinėms ląstelėms ląstelių dalijimosi metu.
Eukariotinės ląstelės branduolys yra apsuptas dviejų elementarių membranų, kurios susidaro branduolinis apvalkalas. Jis persmelktas daugybe porų, kurių skersmuo nuo 30 iki 100 nm, matomos tik elektroniniu mikroskopu. Poros turi sudėtingą struktūrą. Branduolinės membranos išorinė membrana kai kuriose vietose yra sujungta su endoplazminiu tinklu. Branduolinis apvalkalas gali būti vertinamas kaip specializuota, lokaliai diferencijuota endoplazminio tinklo (ER) dalis.
Specialiais dažais nudažytoje šerdyje galima išskirti plonus siūlus ir gumuliukus chromatinas Ir nukleoplazma(pagrindinė branduolio medžiaga). Chromatinas susideda iš DNR, susietos su specialiais baltymais, vadinamais histonais. Ląstelių dalijimosi procese chromatinas tampa vis tankesnis ir kaupiasi chromosomos. DNR koduoja genetinę informaciją.
Organizmai skiriasi chromosomų skaičiumi somatinėse ląstelėse. Pavyzdžiui, kopūstai turi - 20 chromosomų; saulėgrąžos - 34; kviečiai - 42; žmonių – 46, ir viena iš paparčio rūšių Ophioglossum – 1250. Lytinės ląstelės (gametos) turi tik pusę chromosomų, būdingų somatinėms kūno ląstelėms. Chromosomų skaičius gametose vadinamas haploidas(pavieniai), somatinėse ląstelėse - diploidas(dvigubas). Ląstelės, turinčios daugiau nei du chromosomų rinkinius, vadinamos poliploidas.
Šviesos mikroskopu galima pamatyti sferines struktūras - branduoliai. Kiekvienas branduolys turi vieną ar daugiau branduolių, kurie matomi nesidalijančiuose branduoliuose. Ribosominė RNR sintetinama branduolyje. Paprastai diploidinių organizmų branduoliai turi du branduolius, po vieną kiekvienam haploidiniam chromosomų rinkiniui. Branduoliai neturi savo membranos. Biochemiškai branduoliams būdinga didelė RNR koncentracija, kuri čia susijusi su fosfoproteinais. Branduolių dydis priklauso nuo ląstelės funkcinės būklės. pastebėta, kad sparčiai augančioje ląstelėje, kurioje vyksta intensyvūs baltymų sintezės procesai, didėja branduolių dydis. Branduoliuose gaminasi mRNR ir ribosomos, kurios sintetinę funkciją atlieka tik branduolyje.
Nukleoplazmą (karioplazmą) vaizduoja vienalytis skystis, kuriame yra ištirpę įvairūs baltymai, įskaitant fermentus.
Plastidai. Vakuolės, celiuliozės ląstelės sienelės ir plastidai yra būdingi augalų ląstelių komponentai. Kiekviena plastidė turi savo apvalkalą, susidedantį iš dviejų elementarių membranų. Plastido viduje išskiriama membraninė sistema ir įvairaus laipsnio vienalytė medžiaga - stroma. Subrendusios plastidės klasifikuojamos pagal juose esančius pigmentus.
chloroplastai, kuriose vyksta fotosintezė, turi chlorofilų ir karotinoidų. Paprastai turi 4–5 mikronų skersmens disko formą. Vienoje mezofilo ląstelėje (lapo viduryje) gali būti 40 - 50 chloroplastų; mm 2 lapuose – apie 500 000. citoplazmoje chloroplastai dažniausiai išsidėstę lygiagrečiai ląstelės membranai.
Vidinė chloroplasto struktūra yra sudėtinga. Stroma yra persmelkta išvystyta membranų sistema burbuliukų pavidalu - tilakoidai. Kiekvienas tilakoidas susideda iš dviejų membranų. Tilakoidai sudaro vieną sistemą. Paprastai jie surenkami į krūvas - grūdai primenančios monetų stulpelius. Atskirų granatų tilakoidai yra tarpusavyje sujungti stromos tilakoidais arba tarpgranaliniais tilakoidais. Chlorofilas ir karotinoidai yra įterpti į tilakoidų membranas. Žaliųjų augalų ir dumblių chloroplastuose dažnai yra krakmolo grūdelių ir mažų lipidų (riebalų) lašelių. Krakmolo grūdai yra laikinos fotosintezės produktų saugyklos. Iš tamsoje išbuvusio augalo chloroplastų jie gali išnykti tik 24 valandas, o vėl atsirasti praėjus 3–4 valandoms po augalo perkėlimo į šviesą.
Chloroplastai yra pusiau autonominiai organeliai ir panašūs į bakterijas. Pavyzdžiui, bakterijų ir chloroplastų ribosomos yra gana panašios. Jie yra mažesni už eukariotų ribosomas. Baltymų sintezę bakterijų ir chloroplastų ribosomose slopina chloramfenikolis, kuris eukariotinėse ląstelėse neveikia. Be to, tiek bakterijos, tiek chloroplastai turi panašų nukleoidų tipą, organizuotą panašiai. Nepaisant to, kad chloroplastų susidarymą ir juose esančių pigmentų sintezę daugiausia kontroliuoja ląstelės chromosomų DNR, vis dėlto, nesant savo DNR, chloroplastai nesusidaro.
Chloroplastai gali būti laikomi pagrindiniais ląstelių organeliais, nes jie yra pirmieji saulės energijos transformacijos grandinėje, dėl kurios žmonija gauna ir maistą, ir kurą. Chloroplastuose vyksta ne tik fotosintezė. Jie taip pat dalyvauja aminorūgščių ir riebalų rūgščių sintezėje, tarnauja kaip laikinų krakmolo atsargų saugykla.
Chromoplastai - pigmentuoti plastidai. Įvairios formos, jos neturi chlorofilo, tačiau sintetina ir kaupia karotinoidus, kurie žiedams, seniems lapams, vaisiams ir šaknims suteikia geltoną, oranžinę, raudoną spalvą. iš chloroplastų gali išsivystyti chromoplastai, kurie tokiu atveju praranda chlorofilą ir vidines membranines struktūras, kaupia karotenoidus. Tai atsitinka daugelio vaisių nokimo metu. Chromoplastai pritraukia vabzdžius ir kitus gyvūnus, su kuriais jie vystėsi kartu.
Leukoplastai - nepigmentuotos plastidės. Kai kurie iš jų sintetina krakmolą ( amiloplastai), kiti gali sudaryti įvairias medžiagas, įskaitant lipidus ir baltymus. Šviesoje leukoplastai virsta chloroplastais.
proplastidai - mažos bespalvės arba šviesiai žalios nediferencijuotos plastidės, kurios randamos meristematinėse (dalijamosiose) šaknų ir ūglių ląstelėse. Jie yra kitų, labiau diferencijuotų plastidų – chloroplastų, chromoplastų ir aminoplastų – pirmtakai. Jei protoplastidų vystymasis vėluoja dėl šviesos trūkumo, vienas ar daugiau prolameliniai kūnai, kurios yra pusiau kristalinės vamzdinių membranų sankaupos. Plastidės, turinčios prolamelių kūnus, vadinamos etioplastai.Šviesoje etioplastai virsta chloroplastais, o prolamelių kūnų membranos sudaro tilakoidus. Etioplastai susidaro tamsoje esančių augalų lapuose. sėklinių embrionų protoplastai pirmiausia virsta etioplastais, iš kurių vėliau šviesoje vystosi chloroplastai. Plastidams būdingas gana lengvas perėjimas iš vienos rūšies į kitą. Plastidės, kaip ir bakterijos, dauginasi dalijantis į dvi dalis. Meristematinėse ląstelėse protoplastidų dalijimosi laikas maždaug sutampa su ląstelių dalijimosi laiku. Tačiau subrendusiose ląstelėse dauguma plastidų susidaro dėl subrendusių plastidų dalijimosi.
Mitochondrijos. Kaip ir chloroplastus, mitochondrijas supa dvi elementarios membranos. Vidinė membrana sudaro daugybę raukšlių ir išsikišimų - krista, kurios žymiai padidina vidinį mitochondrijų paviršių. Jie yra žymiai mažesni už plastidus, apie 0,5 µm skersmens ir įvairaus ilgio bei formos.
Mitochondrijose vyksta kvėpavimo procesas, kurio pasekoje organinės molekulės suskaidomos išsiskiriant energijai ir ją perduodant ATP molekulėms – pagrindiniam visų eukariotinių ląstelių energijos rezervui. Daugumoje augalų ląstelių yra šimtai ar tūkstančiai mitochondrijų. Jų skaičius vienoje ląstelėje nustatomas pagal ląstelės ATP poreikį. Mitochondrijos nuolat juda, juda iš vienos ląstelės dalies į kitą, susilieja viena su kita ir dalijasi. Mitochondrijos dažniausiai susirenka ten, kur reikia energijos. Jei plazminė membrana aktyviai perneša medžiagas iš ląstelės į ląstelę, tai mitochondrijos yra išilgai membranos paviršiaus. Judriuose vienaląsčiuose dumbliuose mitochondrijos kaupiasi žvynelių apačioje, tiekdamos jiems judėjimui reikalingą energiją.
Mitochondrijos, kaip ir plastidai, yra pusiau autonominės organelės, turinčios komponentų, reikalingų jų pačių baltymų sintezei. Vidinė membrana supa skystą matricą, kurioje yra baltymų, RNR, DNR, į bakterijas panašių ribosomų ir įvairių tirpių medžiagų. DNR egzistuoja apskritų molekulių, esančių viename ar keliuose nukleoiduose, pavidalu.
Remiantis bakterijų panašumu su eukariotinių ląstelių mitochondrijomis ir chloroplastais, galima daryti prielaidą, kad mitochondrijos ir chloroplastai atsirado iš bakterijų, kurios „prieglobstį“ rado didesnėse heterotrofinėse ląstelėse – eukariotų pirmtakuose.
Mikrokūnai. Skirtingai nuo plastidų ir mitochondrijų, kurias riboja dvi membranos, mikrokūnas yra sferinės organelės, apsuptos viena membrana. Mikrokūnai turi granuliuotą (granuliuotą) turinį, kartais juose yra ir kristalinių baltymų intarpų. Mikrokūnai yra susiję su vienu ar dviem endoplazminio tinklo sritimis.
Kai kurie mikroorganizmai vadinami proksisomos, vaidina svarbų vaidmenį glikolio rūgšties metabolizme, kuris yra tiesiogiai susijęs su fotokvėpavimu. Žaliuose lapuose jie yra susiję su mitochondrijomis ir chloroplastais. Kiti mikroorganizmai, vadinami glioksisomos, yra fermentų, reikalingų riebalams paversti angliavandeniais. Tai atsitinka daugelyje sėklų dygimo metu.
Vakuolės - Tai ląstelės sritys, kurias riboja membrana, užpildyta skysčiu. ląstelių sultys. Jie yra apsupti tonoplastas(vakuolinė membrana).
Jaunoje augalo ląstelėje yra daug mažų vakuolių, kurios ląstelei senstant susilieja į vieną didelę vakuolę. Brandžioje ląstelėje iki 90% jos tūrio gali užimti vakuolė. Šiuo atveju citoplazma plono periferinio sluoksnio pavidalu prispaudžiama prie ląstelės membranos. Ląstelių dydžio padidėjimą daugiausia lemia vakuolės augimas. Dėl to atsiranda turgorinis slėgis ir išlaikomas audinių elastingumas. Tai viena iš pagrindinių vakuolės ir tonoplasto funkcijų.
Pagrindinis sulčių komponentas yra vanduo, likusi dalis skiriasi priklausomai nuo augalo rūšies ir jo fiziologinės būklės. Vakuolėse yra druskų, cukrų, rečiau baltymų. Tonoplastas vaidina aktyvų vaidmenį transportuojant ir kaupiant tam tikrus jonus vakuolėje. Jonų koncentracija ląstelės sultyse gali gerokai viršyti jos koncentraciją aplinkoje. Esant dideliam tam tikrų medžiagų kiekiui, vakuolėse susidaro kristalai. Labiausiai paplitę yra kalcio oksalato kristalai, kurie turi skirtingą formą.
Vakuolės yra medžiagų apykaitos produktų (metabolizmo) kaupimosi vietos. Tai gali būti baltymai, rūgštys ir net žmonėms toksiškos medžiagos (alkaloidai). Pigmentai dažnai nusėda. Mėlyna, violetinė, violetinė, tamsiai raudona, tamsiai raudona suteikia augalų ląstelėms pigmentų iš antocianinų grupės. Skirtingai nuo kitų pigmentų, jie gerai tirpsta vandenyje ir yra ląstelių sultyse. Jie lemia daugelio daržovių (ridikėlių, ropių, kopūstų), vaisių (vynuogių, slyvų, vyšnių), gėlių (rugiagėlių, pelargonijų, delfinijų, rožių, bijūnų) raudoną ir mėlyną spalvą. Kartais šie pigmentai užmaskuoja chlorofilą lapuose, pavyzdžiui, dekoratyviniame raudonajame kleve. Antocianinai nuspalvina rudens lapus ryškiai raudonai. Jie susidaro šaltu saulėtu oru, kai lapuose sustoja chlorofilo sintezė. Lapuose, kai nesusidaro antocianinai, sunaikinus chlorofilą, pastebimi geltonai oranžiniai chloroplastų karotenoidai. Lapai ryškiausios spalvos būna šaltą, giedrą rudenį.
Vakuolės dalyvauja makromolekulių sunaikinime, jų komponentų cirkuliacijoje ląstelėje. Ribosomos, mitochondrijos, plastidai, patenkantys į vakuoles, sunaikinami. Pagal šią virškinimo veiklą juos galima palyginti su lizosomos- gyvūnų ląstelių organelės.
Vakuolės susidaro iš endoplazminio tinklo (tinklo)
Ribosomos. Mažos dalelės (17–23 nm), susidedančios iš maždaug vienodo kiekio baltymų ir RNR. Ribosomose aminorūgštys susijungia ir sudaro baltymus. Ląstelėse, kuriose vyksta aktyvi medžiagų apykaita, jų yra daugiau. Ribosomos yra ląstelės citoplazmoje laisvai arba prijungtos prie endoplazminio tinklo (80S). Jų taip pat yra branduolyje (80S), mitochondrijose (70S), plastiduose (70S).
Ribosomos gali sudaryti kompleksą, kuriame vienu metu vyksta identiškų polipeptidų sintezė, informacija apie kurią paimama iš vienos molekulės ir RNR. Toks kompleksas vadinamas poliribosomos (polisomos). Ląstelės, kurios sintetina baltymus dideliais kiekiais, turi didelę polisomų sistemą, kuri dažnai yra pritvirtinta prie išorinio branduolio apvalkalo paviršiaus.
Endoplazminis Tinklelis. Tai sudėtinga neriboto dydžio trimatė membraninė sistema. ER kontekste atrodo kaip dvi elementarios membranos su siaura skaidria erdve tarp jų. ER forma ir ilgis priklauso nuo ląstelės tipo, jos metabolinio aktyvumo ir diferenciacijos stadijos. Ląstelėse, kurios išskiria arba kaupia baltymus, ER yra plokščių maišelių arba cisternos, su daugybe ribosomų, pritvirtintų prie išorinio paviršiaus. Šis tinklelis vadinamas šiurkštus endoplazminis tinklas. Lygus ER paprastai yra vamzdinės formos. Toje pačioje ląstelėje gali būti šiurkštus ir lygus endoplazminis tinklas. Paprastai tarp jų yra daug ryšių.
Endoplazminis tinklas veikia kaip ląstelės ryšio sistema. Jis yra susijęs su išoriniu branduolio apvalkalu. Tiesą sakant, šios dvi struktūros sudaro vieną membranų sistemą. Kai ląstelės dalijimosi metu plyšta branduolio apvalkalas, jo fragmentai primena ER fragmentus. Endoplazminis tinklas yra sistema, pernešanti medžiagas: baltymus, lipidus, angliavandenius į skirtingas ląstelės dalis. kaimyninių ląstelių endoplazminis tinklas yra sujungtas per citoplazmines grandines - plazmodesmata - kurie praeina pro ląstelių sieneles.
Endoplazminis tinklas yra pagrindinė ląstelių membranų sintezės vieta. Kai kuriose augalų ląstelėse vakuolių ir mikrokūnų membranos, cisternos diktiosomos.
Goldžio kompleksas.Šis terminas vartojamas visiems diktiozomas, arba golgi kūnas, narve. Diktiosomos yra plokščių, disko formos pūslelių arba cisternų grupės, kurios išsišakoja išilgai kraštų į sudėtingą kanalėlių sistemą. Aukštesniųjų augalų diktiosomos susideda iš 4–8 cisternų, surinktų kartu.
Paprastai rezervuarų pakuotėje išskiriama besivystanti ir bręstanti pusė. formuojančių cisternų membranos savo struktūra primena ER membranas, o bręstančių cisternų membranos primena plazminę membraną.
Diktiosomos dalyvauja sekrecijoje, o daugumoje aukštesniųjų augalų – ląstelių membranų formavime. Diktiosomų sintetinami ląstelių sienelių polisacharidai kaupiasi pūslelėse, kurios vėliau atsiskiria nuo bręstančių cisternų. Šios sekrecinės pūslelės migruoja ir susilieja su CMP; o juose esantys polisacharidai yra įterpti į ląstelės membraną. Kai kurios medžiagos, besikaupiančios diktiozomose, susidaro kitose struktūrose, pavyzdžiui, ER, o po to pernešamos į diktioomas, kur jos modifikuojamos (modifikuojamos) prieš sekreciją. Pavyzdžiui, glikoproteinai yra svarbi ląstelės membranos statybinė medžiaga. Baltyminę dalį sintetina šiurkščiojo ER polisomos, angliavandenių dalį – diktiozomose, kur abi dalys susijungdamos sudaro glikoproteinus.
Membranos yra dinamiškos, judrios struktūros, nuolat keičiančios savo formą ir plotą. Koncepcija pagrįsta membranos mobilumu endoplazminė sistema. Pagal šią koncepciją vidinės citoplazmos membranos, išskyrus mitochondrijų ir plastidžių membranas, yra viena visuma ir kyla iš endoplazminio tinklo. Iš endoplazminio tinklo per tarpinę pūslelių stadiją susidaro naujos diktiozomų cisternos, o sekrecinės pūslelės, kurios atsiskiria nuo diktiozomų, ilgainiui prisideda prie plazminės membranos susidarymo. Taigi, endoplazminis tinklas ir diktiosomos sudaro funkcinį vienetą, kuriame diktiosomos atlieka tarpinių struktūrų vaidmenį membranas, tokias kaip endoplazminis tinklas, paverčiant tokiomis membranomis kaip plazma. Audiniuose, kurių ląstelės auga ir dalijasi silpnai, membranos komponentai nuolat atnaujinami.
mikrovamzdeliai randama beveik visose eukariotinėse ląstelėse. Tai cilindrinės struktūros, kurių skersmuo yra apie 24 nm. Jų ilgis skiriasi. Kiekvienas kanalėlis sudarytas iš baltymo, vadinamo, subvienetų tubulinas. Subvienetai sudaro 13 išilginių gijų, supančių centrinę ertmę. Mikrovamzdeliai yra dinamiškos struktūros, jos reguliariai sunaikinamos ir susidaro tam tikrais ląstelių ciklo etapais. Jų surinkimas vyksta specialiose vietose, kurios vadinamos mikrovamzdelių organizavimo centrais. Augalų ląstelėse jie turi silpnai išreikštą amorfinę struktūrą.
Mikrovamzdelių funkcijos: dalyvauja formuojant ląstelės membraną; nukreipti diktiozomų burbulus į besiformuojančią membraną, pavyzdžiui, verpstės siūlus, kurie susidaro besidalijančioje ląstelėje; vaidina vaidmenį formuojant ląstelės plokštelę (pradinę ribą tarp dukterinių ląstelių). Be to, mikrovamzdeliai yra svarbi žvynelių ir blakstienų sudedamoji dalis, kurių judėjime jie atlieka svarbų vaidmenį.
mikrofilamentai, kaip ir mikrovamzdeliai, jų yra beveik visose eukariotinėse ląstelėse. Tai ilgi, 5–7 nm storio siūlai, susidedantys iš susitraukiančio baltymo aktino. Mikrofilamentų ryšuliai randami daugelyje aukštesniųjų augalų ląstelių. Matyt, jie vaidina svarbų vaidmenį citoplazmos srovėse. Mikrofilamentai kartu su mikrotubuliais sudaro lankstų tinklą, vadinamą citoskeletas.
Bazinė medžiaga ilgą laiką buvo laikomas vienalyčiu (homogenišku) daug baltymų turinčiu tirpalu, turinčiu nedidelį skaičių struktūrų ar net bestruktūrių. Tačiau šiuo metu, naudojant aukštos įtampos elektroninį mikroskopą, nustatyta, kad gruntinė medžiaga yra trimatė gardelė, sudaryta iš plonų (3–6 nm skersmens) gijų, užpildančių visą ląstelę. Kiti citoplazmos komponentai, įskaitant mikrotubulus ir mikrofilamentus, yra suspenduoti mikrotrabekulinis tinklelis.
Mikrotrabekulinė struktūra yra baltymų gijų gardelė, tarp kurių esanti erdvė užpildyta vandeniu. Kartu su vandeniu grotelės yra gelio konsistencijos, gelis atrodo kaip želatininiai kūnai.
Organelės yra pritvirtintos prie mikrotrabekulinės gardelės. Grotelės užtikrina ryšį tarp atskirų ląstelės dalių ir nukreipia tarpląstelinį transportą.
Lipidų lašai- sferinės formos struktūros, suteikiančios augalo ląstelės citoplazmos granuliuotumą šviesos mikroskopu. Elektroninėse mikrografijose jie atrodo amorfiški. Labai panašūs, bet mažesni lašai randami plastiduose.
Lipidų lašai, painiodami juos su organelėmis, vadino juos sferosomomis ir tikėjo, kad juos supa vieno arba dvigubo sluoksnio membrana. Tačiau naujausi duomenys rodo, kad lipidų lašeliai neturi membranų, bet gali būti padengti baltymu.
Ergastinės medžiagos - tai yra protoplasto „pasyvieji produktai“: atsarginės medžiagos arba atliekos. Jie gali atsirasti ir išnykti skirtingais ląstelių ciklo laikotarpiais. Išskyrus krakmolo grūdelius, kristalus, antocianinų pigmentus ir lipidų lašelius. Tai yra dervos, dervos, taninai ir baltymai. Ergastinės medžiagos yra ląstelės membranos dalis, pagrindinė citoplazmos ir organelių, įskaitant vakuoles, medžiaga.
Vėliava ir blakstienos - tai plonos, į plaukus panašios struktūros, besitęsiančios nuo daugelio eukariotinių ląstelių paviršiaus. Jų skersmuo yra pastovus, tačiau ilgis svyruoja nuo 2 iki 150 mikronų. Tradiciškai ilgesnės ir kelios jų vadinamos žiuželiais, o trumpesnės ir gausesnės – blakstienomis. Nėra aiškių skirtumų tarp šių dviejų tipų struktūrų, todėl terminas vartojamas abiem. žvyneliai.
Kai kuriuose dumbliuose ir grybuose žvyneliai yra judėjimo organai, kurių pagalba jie juda vandenyje. Augaluose (pavyzdžiui, samanose, kepenėliuose, paparčiuose, kai kuriuose gimnasėkliuose) žvyneliais yra tik lytinės ląstelės (gametos).
Kiekvienas žvynelis turi tam tikrą organizaciją. Išorinis 9 porų mikrovamzdelių žiedas supa du papildomus mikrovamzdelius, esančius žvynelio centre. Fermentų turinčios „rankenos“ tęsiasi iš kiekvienos išorinės poros vieno mikrotubulo. Tai yra pagrindinis 9+2 organizavimo modelis, randamas visose eukariotinių organizmų žvyneliuose. Manoma, kad žvynelių judėjimas pagrįstas mikrotubulių slydimu, o išorinės mikrovamzdelių poros juda viena iš kitos nesusitraukdamos. Mikrovamzdelių porų slydimas vienas kito atžvilgiu sukelia vietinį žvynelio lenkimą.
Vėliavos „auga“ iš citoplazminių cilindrinių struktūrų, vadinamų baziniai kūnai, suformuota ir bazinė žvynelio dalis. Baziniai kūnai turi vidinę struktūrą, panašią į žvynelinę, išskyrus tai, kad išoriniai kanalėliai yra išsidėstę trynukais, o ne poromis, o centrinių kanalėlių nėra.
Ląstelių sienelės. Ląstelės sienelė riboja protoplasto dydį ir neleidžia jam plyšti dėl vandens sugėrimo vakuolės.
Ląstelės sienelė atlieka specifines funkcijas, kurios svarbios ne tik ląstelei ir audiniui, kuriame yra ląstelė, bet ir visam augalui. Ląstelių sienelės atlieka esminį vaidmenį įsisavinant, pernešant ir pašalinant medžiagas, be to, jose gali koncentruotis lizosominė, arba virškinimo veikla.
Ląstelės sienelės komponentai. Tipiškiausias ląstelės sienelės komponentas yra celiuliozė, kas daugiausia lemia jo architektūrą. Celiuliozės molekulės yra sudarytos iš pasikartojančių gliukozės molekulių, sujungtų galais. Ilgos plonos celiuliozės molekulės sujungiamos į mikrofibrilės storis 10 - 25 nm. Mikrofibrilės susipina ir sudaro plonus siūlus, kurie savo ruožtu gali būti apvynioti vienas aplink kitą, tarsi sruogos virvelėje. Kiekviena tokia „virvė“ arba makrofibrilė yra apie 0,5 mikrono storio, o ilgis siekia 4 mikronus. Makrofibrilės yra tokios pat stiprios kaip vienodo dydžio plieninė viela.
Ląstelės sienelės celiuliozės karkasas užpildytas su juo susipynusiomis celiuliozės matricos molekulėmis. Jame yra polisacharidų, vadinamų hemiceliuliozės, Ir pektinai, arba pektinai, chemiškai labai artimas hemiceliuliozei.
Kitas ląstelės sienelės komponentas yra ligninas - yra labiausiai augalų ląstelėse esantis polimeras po celiuliozės. Ligninas padidina sienos standumą ir dažniausiai randamas ląstelėse, kurios atlieka atraminę arba mechaninę funkciją.
Cutinas, suberinas, vaškas- dažniausiai nusėda augalų apsauginių audinių lukštuose. Pavyzdžiui, kutinas randamas epidermio ląstelių membranose, o suberinas – antriniame apsauginiame audinyje – kamštelyje. Abi medžiagos randamos kartu su vašku ir neleidžia augalui pernelyg prarasti vandens.
Ląstelės sienelės sluoksniai. Augalų ląstelių sienelių storis labai skiriasi, priklausomai nuo ląstelių vaidmens augalo struktūroje ir pačios ląstelės amžiaus. Elektroniniu mikroskopu augalo ląstelės sienelėje matomi du sluoksniai: vidurinė plokštelė(taip pat vadinama tarpląsteline medžiaga), ir pirminė ląstelės sienelė. Daugelis ląstelių kloja kitą sluoksnį - antrinė ląstelės sienelė. Vidutinė plokštė yra tarp gretimų ląstelių pirminių sienelių. Antrinę sienelę, jei yra, ląstelės protoplastas nusodina ant pirminės ląstelės sienelės vidinio paviršiaus.
Vidurinė plokštelė. Vidurinę plokštelę daugiausia sudaro pektinas. Ten, kur tarp dviejų naujai susidariusių ląstelių turėtų atsirasti ląstelės sienelė, pirmiausia pastebimas tankus endoplazminio tinklo kanalėlių rezginys ir Golgi aparato cisterna (diktiosomos). Tada šioje vietoje atsiranda burbuliukai, užpildyti pektinu (iš polisacharidų). Šie burbuliukai yra atskirti nuo Golgi aparato rezervuarų. Ankstyvojoje ląstelės sienelėje yra įvairių polisacharidų, iš kurių pagrindiniai yra pektinai ir hemiceliuliozė. Vėliau jis susideda iš tankesnių medžiagų – celiuliozės ir lignino.
Pirminė ląstelės sienelė. Tai yra celiuliozės apvalkalo sluoksnis, kuris nusėda prieš ląstelių augimą arba jo metu. Be celiuliozės, hemiceliuliozės ir pektino, pirminėse membranose yra glikoproteino. Pirminiai apvalkalai gali sudegti. Pektino komponentas suteikia plastiškumo, todėl pirminis apvalkalas gali išsiplėsti, kai šaknis, stiebas ar lapas pailgėja.
Aktyviai besidalijančios ląstelės (dauguma subrendusių ląstelių, dalyvaujančių fotosintezės, kvėpavimo ir sekrecijos procesuose) turi pirmines membranas. Tokios ląstelės, turinčios pirminę membraną ir gyvą protoplastą, gali prarasti jiems būdingą formą, dalytis ir diferencijuotis į naują ląstelių tipą. Jie dalyvauja žaizdų gijime ir audinių regeneracijoje augaluose.
Pirminės ląstelių membranos nėra vienodo storio per visą ilgį, tačiau turi plonas dalis, vadinamus pirminiai porų laukai. Citoplazmos gijos arba plazmodesmata, jungiančios gretimų ląstelių protoplastus, dažniausiai praeina per pirminius porų laukus.
antrinė ląstelės sienelė. Nepaisant to, kad daugelis augalų ląstelių turi tik pirminę sienelę, kai kuriose, protoplastas nusodina antrinę sienelę link ląstelės centro. Paprastai tai įvyksta sustojus ląstelių augimui ir pirminės membranos plotas nebedidėja. Dėl šios priežasties antrinis apvalkalas skiriasi nuo pirminio. Antrinės membranos ypač reikalingos specializuotoms ląstelėms, kurios stiprina augalą ir praleidžia vandenį. Nusodinus antrinę membraną, šių ląstelių protoplastas, kaip taisyklė, miršta. Antriniuose apvalkaluose celiuliozės yra daugiau nei pirminiuose, juose nėra pektino medžiagų ir glikoproteinų. Antrinė membrana sunkiai ištempiama, jos matrica susideda iš hemiceliuliozės.
Antriniame apvalkale galima išskirti tris sluoksnius – išorinį, vidurinį ir vidinį (S 1, S 2, S 3). Sluoksniuota antrinių apvalkalų struktūra žymiai padidina jų stiprumą. Celiuliozės mikrofibrilės antrinėje membranoje nusėda tankiau nei pirminėje. Ligninas yra įprastas antrinių medienos odelių komponentas.
Poros kontaktuojančių ląstelių membranose yra viena priešais kitą. Susidaro dvi priešingos poros ir porų membrana porą kartų. Ląstelėse su antrine membrana yra du pagrindiniai porų tipai: paprastas ir su apvadu. Apribotose porose antrinė membrana kabo porų ertmė. Paprastose porose taip nėra.
Ląstelių sienelių augimas. Kai ląstelė auga, didėja ląstelės membranos storis ir plotas. Lukšto tempimas yra sudėtingas procesas. Jį kontroliuoja protoplastas ir reguliuoja hormonas auksinas.
Ląstelėse, kurios auga tolygiai visomis kryptimis, miofibrilių nusėdimas yra atsitiktinis. Šios miofibrilės sudaro netaisyklingą tinklą. Tokios ląstelės randamos kamieno šerdyje, saugojimo audiniuose ir ląstelių auginimo in vitro metu. Pailgėjusiose ląstelėse šoninių apvalkalų miofibrilės nusėda stačiu kampu pailgėjimo ašiai.
Matricinės medžiagos – pektinai, hemiceliuliozės ir glikoproteinai į membraną pernešami diktiozomų pūslelėmis. Tuo pačiu pektinai labiau būdingi augančioms ląstelėms, o neaugančiose ląstelėse vyrauja hemiceliuliozės.
Celiuliozės mikrofibrilės sintezuojamos ląstelės paviršiuje, padedant fermentų kompleksui, susijusiam su plazmine membrana. Mikrofibrilių orientaciją kontroliuoja mikrotubulės, esančios šalia vidinio plazmos membranos paviršiaus.
Plazmodezma. Tai ploni citoplazmos siūlai, jungiantys kaimyninių ląstelių protoplastus. Plazmodesmatos arba praeina pro ląstelės sienelę bet kur, arba yra susitelkusios pirminiuose porų laukuose arba membranose tarp porų porų. Elektroniniu mikroskopu plazmodesmata atrodo kaip siauri kanalai, iškloti plazmos membrana. Mažesnis cilindrinis vamzdis driekiasi išilgai kanalo ašies iš vienos ląstelės į kitą - desmotube, kuri bendrauja su abiejų gretimų ląstelių endoplazminiu tinklu. Daugelis plazmodesmų susidaro ląstelių dalijimosi metu, kai besivystančios ląstelės plokštelė paima vamzdinį endoplazminį tinklą. Plazmodezmos taip pat gali susidaryti nesidalijančių ląstelių membranose. Šios struktūros užtikrina efektyvų tam tikrų medžiagų perkėlimą iš ląstelės į ląstelę.
Ląstelių dalijimasis. Daugialąsčiuose organizmuose ląstelių dalijimasis kartu su jų dydžio padidėjimu yra viso organizmo augimo būdas. Dalijimosi metu susidariusios naujos ląstelės savo struktūra ir funkcijomis yra panašios tiek į pirminę, tiek į viena kitą. Eukariotų dalijimosi procesą galima suskirstyti į du iš dalies sutampančius etapus: mitozė Ir citokinezė.
Mitozė- tai dviejų dukterinių branduolių susidarymas iš vieno branduolio, morfologiškai ir genetiškai lygiaverčiai vienas kitam. Citokinezė - Tai ląstelės citoplazminės dalies dalijimasis, kai susidaro dukterinės ląstelės.
Ląstelių ciklas. Gyva ląstelė pereina iš eilės įvykių, kurie sudaro ląstelės ciklą. Pačio ciklo trukmė skiriasi priklausomai nuo ląstelės tipo ir išorinių veiksnių, tokių kaip temperatūra ar maistinių medžiagų prieinamumas. Paprastai ciklas skirstomas į tarpfazė ir keturios fazės mitozė.
Tarpfazė. Laikotarpis tarp nuoseklių mitozinių dalijimų.
Tarpfazė yra padalinta į tris periodus, žymimus kaip G 1 , S , G 2 .
G 1 periodu, kuris prasideda po mitozės. Per šį laikotarpį padidėja citoplazmos kiekis, įskaitant įvairius organelius. Be to, remiantis šiuolaikine hipoteze, G 1 periodo metu sintetinamos medžiagos, kurios arba stimuliuoja, arba slopina S periodą ir likusį ciklą, taip nulemdamos dalijimosi procesą.
S periodas seka po G1 periodo, tuo metu genetinė medžiaga (DNR) yra dubliuojama.
Per G2 laikotarpį, einantį po S, susidaro struktūros, kurios tiesiogiai dalyvauja mitozėje, pavyzdžiui, veleno komponentai.
Kai kurios ląstelės praeina neribotą ląstelių ciklų skaičių. Tai vienaląsčiai organizmai ir kai kurios aktyvaus augimo zonų ląstelės (meristemos). Kai kurios specializuotos ląstelės po brendimo praranda gebėjimą daugintis. Trečioji ląstelių grupė, pavyzdžiui, formuojančios žaizdos audinį (kalusą), gebėjimą dalytis išlaiko tik ypatingomis sąlygomis.
mitozė, arba branduolio dalijimasis. Tai nuolatinis procesas, suskirstytas į keturis etapus: profazė, metafazė, anafazė, telofazė. Dėl mitozės tarpfazėje padvigubėjusi genetinė medžiaga po lygiai pasiskirsto tarp dviejų dukterinių branduolių.
Vienas iš pirmųjų ląstelių perėjimo prie dalijimosi požymių yra siauros žiedinės mikrovamzdelių juostos atsiradimas tiesiai po plazmos membrana. Šis gana tankus diržas supa branduolį būsimojo mitozinio veleno pusiaujo plokštumoje. Kadangi jis pasirodo prieš profazę, jis vadinamas priešfazinis diržas. Jis išnyksta po mitozinio verpstės, dar ilgai prieš atsirandant vėlyvojoje ląstelės plokštelės telofazėje, kuri auga nuo centro iki periferijos ir susilieja su motininės ląstelės membrana toje srityje, kurią anksčiau užėmė priešfazinis diržas.
Profazė. Profazės pradžioje chromosomos primena ilgus siūlus, išsibarsčiusius branduolio viduje. Tada, sruogoms trumpėjant ir storėjant, matyti, kad kiekviena chromosoma susideda ne iš vienos, o iš dviejų susipynusių sruogų, vadinamų chromatidės. Vėlyvojoje fazėje dvi sutrumpintos suporuotos kiekvienos chromosomos chromatidės yra lygiagrečiai viena šalia kitos, sujungtos siaura atkarpa, vadinama centromeras. Jis turi tam tikrą vietą kiekvienoje chromosomoje ir padalija chromosomą į dvi skirtingo ilgio šakas.
Mikrovamzdeliai išsidėstę lygiagrečiai branduolio paviršiui išilgai veleno ašies. Tai yra ankstyviausias mitozinio veleno surinkimo pasireiškimas.
Profazės pabaigoje branduolys palaipsniui praranda aiškų kontūrą ir galiausiai išnyksta. Netrukus po to branduolinis apvalkalas taip pat suyra.
Metafazė. Iš pradžių metafazinis velenas, kuri vaizduoja trimatę struktūrą, plačiausią per vidurį ir siaurėjančią link polių, užima vietą, kurią anksčiau užėmė branduolys. Verpstės pluoštai yra mikrovamzdelių pluoštai. Metafazės metu chromosomos, kurių kiekviena susideda iš dviejų chromatidžių, yra išdėstytos taip, kad jų centromerai būtų suklio pusiaujo plokštumoje. Kiekviena chromosoma savo centromeru yra pritvirtinta prie veleno sriegių. Tačiau kai kurios sruogos pereina iš vieno poliaus į kitą neprisirišusios prie chromosomų.
Kai visos chromosomos yra pusiaujo plokštumoje, metafazė baigiama. Chromosomos yra pasirengusios dalytis.
Anafazė. Kiekvienos chromosomos chromatidės išsiskiria. dabar tai vaikas chromosomos. Visų pirma, centromera dalijasi, o dvi dukterinės chromosomos traukiamos į priešingus polius. Šiuo atveju centromerai juda į priekį, o chromosomų rankos išsitiesia už nugaros. Prie chromosomų prisirišę verpstės siūlai sutrumpėja, prisidedant prie chromatidų atsiskyrimo ir dukterinių chromosomų judėjimo priešingomis kryptimis.
Telofazė. Telofazėje baigiasi dviejų identiškų chromosomų grupių atskyrimas, o aplink kiekvieną iš jų susidaro branduolinė membrana. Šiurkštus tinklelis aktyviai dalyvauja čia. Dingsta veleno aparatas. Telofazės metu chromosomos praranda kontūrų aiškumą, išsitempia, vėl virsta plonais siūlais. Branduoliai atkuriami. Kai chromosomos tampa nematomos, mitozė baigiasi. Du dukteriniai branduoliai patenka į interfazę. Jie yra genetiškai lygiaverčiai vienas kitam ir pirminiam branduoliui. Tai labai svarbu, nes genetinė programa, o kartu ir visi požymiai, turi būti perduota dukteriniams organizmams.
Mitozės trukmė skirtinguose organizmuose skiriasi ir priklauso nuo audinio tipo. Tačiau profazė yra ilgiausia, o anafazė – trumpiausia. Šaknies galiuko ląstelėse profazės trukmė yra 1–2 valandos; metafazė - 5 - 15 min; anafazė - 2 - 10 min; telofazė - 10 - 30 min. Tarpfazės trukmė nuo 12 iki 30 valandų.
Daugelyje eukariotinių ląstelių mikrotubulų organizavimo centrai, atsakingi už mitozinio veleno susidarymą, yra susiję su centrioliai.
Citokinezė. Tai yra citoplazmos dalijimosi procesas. Daugumoje organizmų ląstelės dalijasi atitraukdamos ląstelės membraną ir suformuodamos dalijimosi vagą, kuri palaipsniui gilėja, suspaudžiant likusius mitozinio verpstės siūlus.Visuose augaluose (briofituose ir kraujagysliniuose) ir kai kuriuose dumbliuose ląstelės dalijasi dėl susidarymo. ląstelių plokštelė.
Ankstyvojoje telofazėje tarp dviejų dukterinių branduolių susidaro statinės formos skaidulų sistema, vadinama fragmoplastas. Fragmoplasto pluoštai, kaip ir mitozinio veleno pluoštai, susideda iš mikrovamzdelių. Fragmoplasto pusiaujo plokštumoje atsiranda nedideli lašeliai. Jie susilieja ir sudaro ląstelės plokštelę, kuri auga tol, kol pasiekia besidalijančios ląstelės sienelę. Tai užbaigia dviejų dukterinių ląstelių atskyrimą. Susiliejantys lašeliai yra burbuliukai, atitrūkę nuo Golgi aparato. Iš esmės juose yra pektino medžiagų, iš kurių susidaro vidurinė plokštelė. Pūslelinės membranos dalyvauja kuriant plazminę membraną abiejose plokštelės pusėse. Tuo pačiu metu iš vamzdinio endoplazminio tinklo fragmentų susidaro plazmodesmata.
Susidarius vidutinei plokštelei, kiekvienas protoplastas ant jos nusodina pirminę membraną. Be to, kiekviena dukterinė ląstelė aplink visą protoplastą nusodina naują membranos sluoksnį, kuris tęsia membraną, kuri atsirado iš ląstelės plokštelės. Augant dukterinėms ląstelėms, pradinis pirminės ląstelės apvalkalas sunaikinamas.
Įvairūs eukariotų mitozės tipai
Aukščiau aprašytas ląstelių dalijimasis augaluose, taip pat ir gyvūnuose, nėra vienintelė netiesioginio ląstelių dalijimosi forma. Paprasčiausias mitozės tipas yra pleuromitozė. Tai primena dvejetainį padalijimą prokariotinės ląstelės, kuriame nukleoidai po replikacijos lieka susieti su plazmos membrana. Membrana pradeda augti tarp DNR surišimo taškų ir taip pasklinda chromosomas į skirtingas ląstelės dalis. Po to, susiformuojant ląstelių susiaurėjimui, kiekviena iš DNR molekulių bus naujoje atskiroje ląstelėje.
Būdinga padalijimui eukariotinės ląstelės yra iš mikrovamzdelių pastatytos verpstės susidarymas. At uždara pleuromitozė(vadinama uždara, nes chromosomų divergencija vyksta nepažeidžiant branduolio apvalkalo) kaip mikrovamzdelių organizavimo centrai (MCMT) dalyvauja ne centrioliai, o kitos struktūros, esančios vidinėje branduolio membranos pusėje. Tai vadinamieji neapibrėžtos morfologijos poliniai kūnai, iš kurių tęsiasi mikrovamzdeliai. Tokių kūnų yra du. Jie skiriasi vienas nuo kito neprarasdami ryšio su branduolio apvalkalu. Dėl to susidaro dvi pusverpstės, susijusios su chromosomomis. Visas mitozinio veleno formavimosi ir chromosomų segregacijos procesas šiuo atveju vyksta po branduolio apvalkalu. Šio tipo mitozė pasitaiko tarp pirmuonių ir yra plačiai paplitusi tarp grybų (chitridijų, zygomicetų, mielių, oomicetų, askomicetų, miksomicetų ir kt.). atitinka formas pusiau uždara pleuromitozė kai susidariusios verpstės poliuose sunaikinamas branduolio apvalkalas.
Kita mitozės forma yra ortomitozė.Šiuo atveju COMT yra citoplazmoje, nuo pat pradžių susidaro ne pusverpstės, o dvipolis velenas. Yra trys formos ortomitozė ( normali mitozė) pusiau uždara Ir uždaryta. Esant pusiau uždarai ortomitozei, citoplazmoje esančio TsOMT pagalba susidaro bisimetrinis velenas, branduolinis apvalkalas išsaugomas per visą mitozės laikotarpį, išskyrus poliarines zonas. Granuliuotos medžiagos ar net centriolių masės gali būti aptiktos kaip COMT. Ši mitozės forma randama žaliųjų, rudųjų ir raudonųjų dumblių zoosporose, kai kuriuose žemesniuosiuose grybuose ir gregarinuose. Esant uždarai ortomitozei, visiškai išsaugoma branduolinė membrana, kurioje susidaro tikras velenas. Karioplazmoje susidaro mikrovamzdeliai, rečiau išauga iš vidinės TsOMT, kuri nesusijusi (skirtingai nei pleuromitozė) su branduoline membrana. Šio tipo mitozė būdinga blakstienų mikrobranduolių dalijimuisi, tačiau gali atsirasti ir pirmuoniams. Atviros ortomitozės atveju branduolinis apvalkalas visiškai suyra. Šis ląstelių dalijimosi tipas būdingas gyvūnų organizmams, kai kuriems pirmuoniams ir aukštesnių augalų ląstelėms. Šią mitozės formą savo ruožtu atstovauja astraliniai ir anastraliniai tipai.
Iš trumpai apžvelgtos medžiagos matyti, kad pagrindinis mitozės bruožas apskritai yra dalijimosi verpstės struktūrų atsiradimas, kurios susidaro ryšium su CMT, kurios skiriasi savo struktūra.
Mitozinės figūros morfologija
Mitozinis aparatas ypač gerai išreiškiamas mitozės metafazės stadijoje. Metafazėje, ląstelės pusiaujo plokštumoje, išsidėsto chromosomos, iš kurių atsiranda vadinamoji. veleno sriegiai, susiliejančios dviejuose skirtinguose mitozinės figūros poliuose. Taigi mitozinis velenas yra chromosomų, polių ir skaidulų rinkinys. Verpstės skaidulos yra pavieniai mikrovamzdeliai arba jų ryšuliai. Mikrovamzdeliai prasideda nuo veleno polių, o kai kurie jų eina į centromerus kur yra įsikūrę kinetochores chromosomų (kinetochorų mikrotubulių), kai kurios pereina toliau link priešingo poliaus, bet jo nepasiekia. Jie vadinami „tarppoliais mikrotubulais“. Iš polių nukrypsta grupė radialinių mikrovamzdelių, aplink juos suformuodami struktūrą, primenančią „spinduliuojantį švytėjimą“ – tai astraliniai mikrovamzdeliai.
Pagal morfologiją mitozinės figūros skirstomos į astralinius ir anastralinius tipus.
Astralinis verpstės tipas, arba konvergentas, pasižymi tuo, kad jo polius vaizduoja maža zona, į kurią susilieja (susilenkia) mikrovamzdeliai. Paprastai centrosomos, kuriose yra centriolių, yra astralinių verpsčių poliuose. Tačiau yra žinomi centriolinių astralinių mitozių atvejai (kai kurių bestuburių mejozės metu). Be to, pastebimi radialiniai mikrovamzdeliai, sklindantys iš polių, kurie nėra veleno dalis, bet sudaro žvaigždžių zonas - citasters. Šio tipo mitozinis dalijimasis primena hantelį. Mitozinės figūros anastrialinio tipo polių nėra. Verpstės poliarinės sritys čia yra plačios, jos vadinamos poliniais gaubteliais, jose yra centriolių. Šiuo atveju veleno pluoštai nenukrypsta nuo vieno taško, o nukrypsta plačiu priekiu (nukrypsta) nuo visos poliarinių dangtelių zonos. Šio tipo verpstės būdingos besidalijančioms aukštesniųjų augalų ląstelėms, tačiau jos gali būti ir aukštesniuosiuose gyvūnuose. Ankstyvoje žinduolių embriogenezėje centriolinės (divergentinės) mitozės stebimos kiaušialąstės brendimo dalijimosi metu ir I bei II zigotos dalijimosi metu. Tačiau jau trečiojo ląstelių dalijimosi metu ir visose vėlesnėse ląstelėse jie dalijasi dalyvaujant astraliniams verpstams, kurių poliuose visada randama centriolių. Apskritai visoms mitozės formoms chromosomos su jų kinetoforais, poliniais kūnais (centrosomomis) ir verpstės skaidulomis išlieka įprastos struktūros.
Centromerai ir kinetochorai
Centromerai gali būti skirtingos lokalizacijos išilgai chromosomų ilgio. holocentrinis centromerai atsiranda, kai mikrovamzdeliai susijungia per visą chromosomos ilgį (kai kurie vabzdžiai, nematodai, kai kurie augalai). Monocentrinis centromerai – kai mikrovamzdeliai yra susieti su chromosomomis vienoje srityje. monocentriniai centromerai gali būti tiksliai nustatyti(pavyzdžiui, kai kuriose pumpurinėse mielėse), kai tik vienas mikrovamzdelis priartėja prie kinetochoro ir zoninis, kur mikrotubulių pluoštas artėja prie sudėtingo kinetochoro. Nepaisant centromerų zonų įvairovės, jos visos yra susijusios su sudėtinga struktūra. kinetochore, kurios struktūra ir funkcijos yra labai panašios į visus eukariotus. Kinetochorai yra specialios baltymų struktūros, dažniausiai išsidėsčiusios chromosomų centromerinėse zonose. Tai sudėtingi kompleksai, susidedantys iš daugelio baltymų. morfologiškai jie yra labai panašūs, turi tą pačią struktūrą – nuo diatomų iki žmonių. Tai yra trijų sluoksnių struktūros: vidinis tankus sluoksnis, esantis greta chromosomos kūno, vidurinis laisvas sluoksnis ir išorinis tankus sluoksnis. Daugelis fibrilių tęsiasi nuo išorinio sluoksnio, sudarydamos vadinamąjį pluoštinį kinetochoro vainiką. Bendra forma kinetochorai atrodo kaip plokštelės ar diskai, esantys pirminio chromosomos susiaurėjimo zonoje, centromeroje. Paprastai kiekvienoje chromosomoje arba chromatidėje yra vienas kinetochoras. Prieš anafazę kiekvienoje seserinėje chromatidėje yra kinetochorai, kurių kiekviena bendrauja su savo mikrotubulių pluoštu. Augaluose kinetochoras atrodo ne kaip plokštelės, o kaip pusrutuliai. Funkcinis kinetochorų vaidmuo yra surišti seserines chromatides viena su kita, fiksuoti mitozinius mikrotubulus, reguliuoti chromosomų atskyrimą ir faktiškai perkelti chromosomas mitozės metu, dalyvaujant mikrotubulams. Apskritai, baltymų struktūros, kinetochorai S periode dvigubėja, lygiagrečiai chromosomų padvigubėjimui. Tačiau jų baltymai yra chromosomose visais ląstelių ciklo laikotarpiais.
AUGALŲ AUDINIAI
Ugdomieji audiniai (meristemos)
Ugdomieji audiniai augalų organizme išsidėstę skirtingose vietose, todėl skirstomi į tokias grupes (0;1 pav.).
Viršūninės (viršūninės) meristemos išsidėsčiusios ašinių organų – stiebo, šaknies – viršūnėse arba viršūnėse. Šių meristemų pagalba augalų vegetatyviniai organai vykdo savo ilgį.
Šoninės meristemos būdingas ašiniams organams. Ten jie yra koncentriškai, sankabos pavidalu.
tarpkalnis, arba tarpkalariniai, meristemos kilę iš viršūninių meristemų. Tai ląstelių grupės, kurios dar nesugeba daugintis, bet yra išėjusios į diferenciacijos kelią. Tarp jų nėra pradinių ląstelių, tačiau yra daug specializuotų.
žaizdos meristemos atkurti pažeistą kūno dalį. Regeneracija prasideda dediferenciacija, tai yra atvirkštinis vystymasis nuo specializuotų ląstelių iki meristematinių. Jie virsta fellogen, kurios formuojasi kamštiena dengiantis žaizdos paviršių. Dediferencijuotos ląstelės, besidalijančios, gali sudaryti laisvą parenchiminį audinį - nuospauda. Iš jo tam tikromis sąlygomis susidaro augalų organai.
Integumentiniai audiniai
Jie veikia kaip ribinis barjeras, atskiriantys apatinius audinius nuo aplinkos. Pirminius augalo sluoksnius sudaro tik gyvos ląstelės. Antrinis ir tretinis apvalkalas – daugiausia iš mirusiųjų su storomis ląstelių sienelėmis.
Pagrindinės vientisųjų audinių funkcijos:
augalo apsauga nuo išdžiūvimo;
apsauga nuo kenksmingų mikroorganizmų patekimo;
apsauga nuo saulės nudegimo;
apsauga nuo mechaninių pažeidimų;
medžiagų apykaitos tarp augalo ir aplinkos reguliavimas;
dirginimo suvokimas.
Pirminis vientisas audinys – epidermis, epidermis . Susideda iš gyvų ląstelių. Susidaro iš viršūninių meristemų. Apima jaunus augančius stiebus ir lapus.
Epidermis susiformavo augaluose, kai jie patenka iš vandens buveinių į sausumą, kad jis neišdžiūtų. Be stomos, visos epidermio ląstelės yra glaudžiai tarpusavyje susijusios. Išorinės pagrindinių ląstelių sienelės yra storesnės nei likusios. Visas paviršius padengtas kutino ir augalinio vaško sluoksniu. Šis sluoksnis vadinamas odelė(oda). Jo nėra ant augančių šaknų ir povandeninių augalų dalių. Išdžiūvus gerokai susilpnėja odelės pralaidumas.
Išskyrus pagrindinės ląstelės, ypač epidermyje yra kitų plaukeliai, arba trichomes. Jie yra vienaląsčiai ir daugialąsčiai (2 pav.). Funkciškai jie padidina epidermio paviršių, pavyzdžiui, šaknų augimo zonoje, tarnauja kaip mechaninė apsauga, prilimpa prie atramos, mažina vandens praradimą. Nemažai augalų turi liaukiniai plaukai pvz dilgėlių.
Tik aukštesniuose augaluose epidermyje yra stomata kurie reguliuoja vandens ir dujų mainus. Jeigu odelės nėra, vadinasi, ir stomatos nereikia. Stomatai yra susidarančių ląstelių grupė stomatalinis aparatas, kuri susideda iš dviejų sargybos ląstelės ir greta esančios epidermio ląstelės šoninės ląstelės. Jie skiriasi nuo pagrindinių epidermio ląstelių (3 pav ). Apsauginės ląstelės skiriasi nuo aplinkinių ląstelių forma ir daugybe chloroplastų bei netolygiai sustorėjusių sienelių. Tie, kurie susiduria vienas su kitu, yra storesni nei kiti (4 pav.) . Tarp apsauginių ląstelių susidaro stomatos anga, kuris veda į substomatalinė erdvė paskambino substomatalinė ertmė. Apsauginės ląstelės yra labai fotosintetinės. Juose yra daug atsarginio krakmolo ir daug mitochondrijų.
Stomatų skaičius ir pasiskirstymas, stomatalinių aparatų tipai skirtinguose augaluose labai skiriasi. Šiuolaikiniuose briofituose stomatų nėra. Fotosintezę juose vykdo gametofitų karta, o sporofitai negali egzistuoti savarankiškai.
Paprastai stomos yra apatinėje lapo pusėje. Vandens paviršiuje plūduriuojančiuose augaluose – viršutiniame paviršiuje. Javų lapuose stomatai dažnai būna vienodai iš abiejų pusių. Tokie lapai apšviečiami palyginti tolygiai. Ant 1 mm 2 paviršiaus gali būti nuo 100 iki 700 stomatų.
Antrinis vidinis audinys (peridermas). Šis audinys pakeičia epidermį, kai vienmečių ūglių žalia spalva pasikeičia į rudą. Jis yra daugiasluoksnis ir susideda iš centrinio kambinių ląstelių sluoksnio - felogenas. Felogeno ląstelės, besidalijančios, kloja sluoksnį išorėje Fellems, ir viduje - Felodermija(5 pav.).
Fellema, arba kamštiena. Iš pradžių jis susideda iš gyvų plonasienių ląstelių. Laikui bėgant jų sienelės impregnuojamos suberinu ir augaliniu vašku ir miršta. Ląstelės turinys užpildytas oru.
Fellema funkcijos:
apsaugo nuo drėgmės praradimo;
apsaugo augalą nuo mechaninių pažeidimų;
apsaugo nuo patogenų;
užtikrina šilumos izoliaciją, nes ląstelės užpildomos oru.
Felogeno ląstelės, esančios pačiame epidermyje, apatiniame poepiderminiame sluoksnyje, rečiau giliuose pirminės žievės sluoksniuose, yra pirminės žievės pagrindas.
Kamštienos sluoksnis nėra nuolatinis. Joje atsiranda lūžių, kurios susisiekia su šalia esančiomis tarpląstelinėmis erdvėmis. Tuo pačiu metu ant paviršiaus susidaro maži gumbai - lęšiai, kurios jungia tarpląstelines erdves su atmosferos oru (6,7 pav.).
Rudenį po lęšiais esantis felogenas kloja užkamšytų ląstelių sluoksnį, kuris labai sumažina transpiraciją, bet visiškai jos neatmeta. Pavasarį šis sluoksnis suyra iš vidaus. Ant šviesios beržo žievės lęšiai aiškiai matomi tamsių brūkšnelių pavidalu.
Tretinis vientisas audinys (pluta), taip pat būdinga tik sumedėjusioms augalų formoms.
Felogenas pakartotinai nusėda gilesniuose žievės sluoksniuose. Už jos ribų esantys audiniai ilgainiui miršta ir susidaro pluta. Jo ląstelės yra negyvos ir negali ištempti. Tačiau gyvos ląstelės, esančios giliau, dalijasi, todėl padidėja skersinis stiebo dydis. Laikui bėgant, išorinis plutos sluoksnis lūžta. Tokio tarpo atsiradimo laikas yra gana pastovi atskirų augalų reikšmė. Obelėje tai atsitinka septintaisiais gyvenimo metais, skroblėje - penkiasdešimtaisiais. Kai kuriose rūšyse jis visai nevyksta. Pagrindinė pilingo funkcija yra apsauga nuo mechaninių ir terminių pažeidimų.
Parenchima
Tai specializuotų audinių grupė, užpildanti augalo kūno viduje esančias erdves tarp laidžių ir mechaninių audinių. (8 pav.). Dažniau parenchimos ląstelės yra suapvalintos, rečiau pailgos formos. Būdingas išsivysčiusių tarpląstelinių erdvių buvimas. Tarpai tarp ląstelių kartu sudaro transporto sistemą - apoplastas Be to, tarpląstelinės erdvės sudaro augalo „vėdinimo sistemą“. Per stomatas arba lenticelius jie susijungia su atmosferos oru ir užtikrina optimalią dujų sudėtį augalo viduje. Išvystytos tarpląstelinės erdvės ypač reikalingos augalams, augantiems užmirkusioje dirvoje, kur sunku normaliai keistis dujomis. Ši parenchima vadinama aerenchima(9 pav.).
Parenchimos elementai, užpildantys tarpus tarp kitų audinių, atlieka ir atramos funkciją. Parenchimos ląstelės yra gyvos, jos neturi storų ląstelių sienelių, kaip sklerenchima. Todėl mechanines savybes suteikia turgoras. Jei vandens kiekis sumažėja, tai sukelia plazmolizę ir augalo vytimą.
Asimiliacinė parenchima susidaro plonasienių ląstelių su daugybe tarpląstelinių erdvių. Šios struktūros ląstelėse yra daug chloroplastų, todėl ji vadinama chlorenchima. Chloroplastai išsidėstę išilgai sienos, vienas kito neužtemdydami. Asimiliacinėje parenchimoje vyksta fotosintezės reakcijos, kurios aprūpina augalą organinėmis medžiagomis ir energija. Fotosintezės procesų rezultatas – galimybė Žemėje egzistuoti visiems gyviems organizmams.
Asimiliaciniai audiniai yra tik apšviestose augalo dalyse, juos nuo aplinkos skiria skaidrus epidermis. Jei epidermį pakeičia nepermatomi antriniai integumentiniai audiniai, asimiliacinė parenchima išnyksta.
Sandėliavimo parenchima tarnauja kaip organinių medžiagų, kurių augalo organizmas laikinai nenaudoja, rezervuaras. Iš esmės bet kuri ląstelė, turinti gyvą protoplastą, gali nusodinti organines medžiagas įvairių rūšių inkliuzų pavidalu, tačiau kai kurios ląstelės tam specializuojasi. (10, 11 pav.). Energetiniai junginiai nusėda tik vegetacijos metu, vartojami ramybės laikotarpiu ir ruošiantis kitam vegetacijos sezonui. Todėl rezervinės medžiagos vegetatyviniuose organuose nusėda tik daugiamečiuose augaluose.
Atsargų talpykla gali būti paprasti organai (pabėgimas, šaknis), taip pat specializuoti (šakniastiebiai, gumbai, svogūnėliai). Visi sėkliniai augalai kaupia energetiškai vertingas medžiagas sėklose (skilčialapiuose, endosperme). Daugelis augalų sausringuose klimatuose kaupia ne tik organines medžiagas, bet ir vandenį. (12 pav.). Pavyzdžiui, alavijas kaupia vandenį mėsinguose lapuose, kaktusai – ūgliuose.
mechaniniai audiniai
Mechanines augalų ląstelių savybes suteikia:
kietas ląstelės apvalkalas;
turgoriškumas, tai yra ląstelių turgorinė būsena.
Nepaisant to, kad beveik visos audinių ląstelės turi mechaninių savybių, tačiau augale yra audinių, kurių mechaninės savybės yra pagrindinės. Tai kolenchima Ir sklerenchima. Paprastai jie veikia sąveikaudami su kitais audiniais. Kūno viduje augalai sudaro savotišką rėmą. Todėl jie vadinami sustiprinimas.
Ne visi augalai turi vienodai aiškiai apibrėžtus mechaninius audinius. Vandens aplinkoje gyvenantiems augalams vidinės paramos reikia daug mažiau nei sausumos augalams. Priežastis ta, kad vandens augalams reikia mažiau vidinės paramos. Jų kūną daugiausia palaiko aplinkinis vanduo. Oras sausumoje tokios atramos nesukuria, nes jo tankis mažesnis nei vandens. Dėl šios priežasties specializuotų mechaninių audinių buvimas tampa aktualus.
Evoliucijos procese įvyko vidinių atraminių struktūrų tobulėjimas.
Kolenchima. Jį sudaro tik gyvos ląstelės, pailgos išilgai organo ašies. Šio tipo mechaniniai audiniai susidaro labai anksti, pirminio augimo laikotarpiu. Todėl svarbu, kad ląstelės išliktų gyvos ir išlaikytų galimybę ištempti kartu su šalia esančiomis tempimo ląstelėmis.
Kolenchimos ląstelių savybės:
netolygus lukšto sustorėjimas, dėl to kai kurios jo dalys lieka plonos, o kitos sustorėja;
lukštai nerūgsta.
Kolenchimos ląstelės yra skirtingos viena kitos atžvilgiu. Gretimose ląstelėse kampuose, nukreiptuose vienas į kitą, susidaro sustorėjimai ( ryžių. 13 ). Ši kolenchima vadinama kampas. Kitu atveju ląstelės yra išdėstytos lygiagrečiais sluoksniais. Ląstelių membranos, esančios priešais šiuos sluoksnius, yra stipriai sustorėjusios. Tai lamelinė kolenchima. Ląstelės gali būti išdėstytos laisvai, su daugybe tarpląstelinių erdvių yra laisva kolenchima. Tokia kolenchima dažnai aptinkama augaluose užmirkusiose dirvose.
Kolenchima ypač svarbi jauniems augalams, žolinėms formoms, taip pat tose augalų dalyse, kuriose antrinis augimas nevyksta, pavyzdžiui, lapuose. Šiuo atveju jis klojamas labai arti paviršiaus, kartais tiesiog po epidermiu. Jei organas turi veidus, tada išilgai jų keteros randami stori kolenchimo sluoksniai.
Kolenchimos ląstelės veikia tik esant turgorui. Vandens trūkumas sumažina kolenchimo efektyvumą ir augalas laikinai nuvysta, pavyzdžiui, karštą dieną nusvyra agurkų lapai. Užpildžius ląsteles vandeniu, atkuriamos kolenchimos funkcijos.
Sklerenchima. Antrasis mechaninių audinių tipas. Skirtingai nuo kolenchimos, kur visos ląstelės yra gyvos, sklerenchimos ląstelės yra negyvos. Jų sienos labai storos. Jie atlieka mechaninę funkciją. Stiprus membranos sustorėjimas sukelia medžiagų transportavimo pažeidimą, dėl kurio protoplastas miršta ( ryžių. 14 ). Sklerenchimos ląstelių membranų lignifikacija įvyksta, kai augalo organas jau baigė augti. Todėl jie nebetrukdo aplinkinių audinių tempimui.
Priklausomai nuo formos, išskiriami du sklerenchimos ląstelių tipai – skaidulinės ir sklereidinės.
skaidulų turi stipriai pailgą formą su labai storomis sienelėmis ir maža ertme. Jie yra šiek tiek mažesni nei medienos pluoštai. Dažnai po epidermiu susidaro išilginiai sluoksniai ir sruogos. Floemoje arba ksileme juos galima rasti pavieniui arba grupėmis. Floemoje jie vadinami šerno pluoštai(ryžių. 15 ), ir ksileme libriforminiai pluoštai(ryžių. 16 ).
sklereidai, arba akmeninės ląstelės, yra vaizduojamos suapvalintomis arba šakotomis ląstelėmis su galingais apvalkalais. Augalo kūne jų galima rasti pavieniui (palaikančios ląstelės) ( pav.17 ) arba grupės (18 pav.). Reikia pažymėti, kad mechaninės savybės labai priklauso nuo sklereidų vietos. Dalis sklereidų sudaro ištisinius sluoksnius, pavyzdžiui, riešutų kevaluose arba vaisių (kaulavaisių) kauliukuose.
išskyrimo audiniai
Bet kuri gyva ląstelė iš pradžių atlieka medžiagų pašalinimo funkciją, o transportavimas gali vykti tiek palei koncentracijos gradientą, tiek prieš gradientą. Medžiagos, kurios buvo susintetintos ląstelėje, gali būti pašalintos ir vėliau paveiks kitų ląstelių veiklą (fitohormonai, fermentai). Toks procesas vadinamas sekrecija. Tuo atveju, kai išsiskiria medžiagos, kurios yra ląstelės atliekos, procesas vadinamas išskyrimas. Nepaisant to, kad medžiagų išskyrimas į augalų ir gyvūnų ląsteles turi esminį panašumą, augalai turi nemažai savybių, atsirandančių dėl esminių gyvybinės veiklos skirtumų. Metabolinių reakcijų lygis augaluose yra daug mažesnis nei gyvūnų. Todėl išmetama proporcingai mažesni atliekų kiekiai. Kitas bruožas yra tai, kad augalai, savarankiškai sintetindami beveik visus būtinus organinius junginius, niekada nesudaro perteklinių jų atsargų. Išsiskyrusios medžiagos gali būti pradinė medžiaga kitoms reakcijoms (pavyzdžiui, CO 2 ir H 2 O).
Jei gyvūnams toksinų išskyrimo procesas yra susijęs su jų pašalinimu iš organizmo, augalai gali to nedaryti, išskirdami nereikalingas medžiagas gyvame protoplaste (įvairių medžiagų pašalinimas į vakuolę), negyvose ląstelėse (daugumoje daugiamečio augalo audinių). augalas susideda iš tokių ląstelių), tarpląstelinėse erdvėse.
Išskiriamos medžiagos gali būti suskirstytos į dvi dideles grupes. Pirmoji grupė – organinės medžiagos, kurias tiesiogiai sintetina ląstelė (fermentai, polisacharidai, ligninai, terpenai, pastarieji yra eterinių aliejų ir dervų sudedamosios dalys). Antroji grupė – medžiagos, kurios iš pradžių į ląstelę patenka iš išorės ksilemo ar floemo transportavimo pagalba (vanduo, mineralinės druskos, aminorūgštys, monosacharidai ir kt.). Išskirta paslaptis retai būna vienalytė ir dažniausiai susideda iš mišinio, kuriame vienos medžiagos koncentracija yra didžiausia.
Augalai, skirtingai nei gyvūnai, neturi pilnos šalinimo sistemos. Visoje gamykloje yra tik specializuotos struktūros - idioblastai. Jas formuojančios ląstelės yra mažesnės nei šalia esančios parenchimos ląstelės. Jie turi elektronų tankią citoplazmą su išvystytais endoplazminio tinklo elementais ir Golgi kompleksu. Dažniausiai nėra išreikšta, yra centrinė vakuolė. Tarp jų ir kitų gyvų ląstelių šios ląstelės yra sujungtos daugybe plazmodesmų.
Priklausomai nuo vietos, išskyrimo audiniai gali būti išorinis ir vidinis.
Išoriniai šalinimo audiniai.liaukiniai plaukai yra dariniai epidermis. Morfologiškai jie yra kintami – gali turėti daugialąstę galvą, būti pailgos, skydo formos ant kojos ir kt. (19 pav.). Tai apima dilgėlių plaukelius.
Hidatodai- tai struktūros, kurios išskiria vandens perteklių esant žemai transpiracijai ir didelei drėgmei - gutacija. Šias struktūras sudaro bespalvių gyvų ląstelių grupės su plonomis sienelėmis - epitema.Šis audinys yra greta laidžiojo pluošto (20 pav.). Vanduo išleidžiamas per specialias vandens stomatas, kurios nuo įprastų stomatų skiriasi nejudrumu ir nuolat atsiveriančiu tarpu. Gutavimo skysčio sudėtis labai skiriasi nuo beveik gryno vandens iki labai sudėtingo medžiagų mišinio.
Nektarai. Daugeliui augalų būdingas skysčio, kuriame yra nuo 7 iki 87% mono- ir oligosacharidų, išsiskyrimas. Šį procesą vykdo specialios struktūros – nektarai. Priklausomai nuo vietos, yra gėlė, arba gėlių, nektarai (21 pav.), taip pat augalai, esantys ant stiebų, lapų - ekstraflora, arba ekstrafloriniai nektarai. Nektaras gali atstovauti Tai nepakitusios floemo sultys, kurios per tarpląstelinius tarpus patenka į paviršių ir išskiriamos per stomatas. Sudėtingesnius nektarus sudaro liaukų parenchima, padengta epidermiu su liaukiniais plaukais. Nektarą išskiria epidermio ląstelės arba liaukų plaukai. Šiuo atveju išsiskiriantis nektaras skiriasi nuo floemų sulčių. Kadangi jame vyrauja gliukozė, fruktozė, sacharozė, o floemo sultyse – gliukozė. Jonų yra nedidelėmis koncentracijomis. Jonų yra nedidelėmis koncentracijomis. Norint pritraukti apdulkintojus, nektare gali būti steroidinių hormonų, būtinų vabzdžiams.
Virškinimo liaukos yra vabzdžiaėdžiuose augaluose (rasa, nepentis ir kt.). Reaguodami į mažų bestuburių prisilietimą, jie išskiria paslaptį, kurioje yra hidrolizinių fermentų, skruzdžių, sviesto ir propiono rūgščių. Dėl to biopolimerai suskaidomi į monomerus, kuriuos augalas naudoja savo reikmėms.
Druskos liaukos vystosi augaluose, augančiuose druskingoje dirvoje. Šios liaukos yra ant lapų, o ištirpusios formos jonų perteklius iškeliamas į paviršių. Druska pirmiausia nusėda ant odelės, vėliau ją nuplauna lietus (22 pav.). Be to, daugelis augalų turi ant lapų druskos plaukeliai. Kiekvienas plaukas susideda iš dviejų ląstelių: viena sudaro galvą, kita - stiebą. Druskos kaupiasi galvos ląstelės vakuolėje, o kai jų koncentracija tampa didelė, galva nukrenta. Jo vietoje išauga nauja ląstelė, kuri taip pat kaupia druskas. Lapui augant gali kelis kartus nukristi ir formuotis galvutė. Druskos plaukų privalumas yra tas, kad išskirdami druskas jie netenka mažai vandens. Šis reiškinys labai svarbus augalams, augantiems druskingose dirvose, kur gėlo vandens trūksta.
Vidiniai šalinimo audiniai. Vidinės šalinimo struktūros, išsibarsčiusios po visą augalo kūną idioblastų pavidalu, nepašalina medžiagų iš organizmo, kaupdamos jas savyje. Idioblastai yra augalų ląstelės, kurios savo forma, struktūra ar turiniu skiriasi nuo kitų to paties audinio ląstelių. Priklausomai nuo kilmės skiriamos šizogeninės (gr. schiso – suskaidytos) ir lizogeninės (gr. lizė – tirpimas) išskiriamų medžiagų talpyklos (23 pav.).
Šizogeninės talpyklos. Tai daugiau ar mažiau plačios tarpląstelinės erdvės, užpildytos išskiriamomis medžiagomis. Pavyzdžiui, spygliuočių, aralijų, skėčių ir kt. Manoma, kad derva pasižymi baktericidinėmis savybėmis, atbaido žolėdžius, todėl augalai jiems nevalgomi.
Lizigeninės talpyklos susidaro vietoje gyvų ląstelių, kurios žūva ir sunaikinamos jose susikaupus medžiagoms. Tokius darinius galima pastebėti citrusinių vaisių žievelėse.
Be pagrindinių tipų, yra tarpinių formų. pieniškas- yra dviejų tipų: sujungti Ir nesegmentuoti. Pirmieji susidaro iš daugybės gyvų ląstelių, išdėstytų grandinėmis. Sąlyčio vietoje apvalkalai sunaikinami, protoplastai susilieja ir dėl to susidaro vientisas tinklas. Tokių pieniškų yra Compositae. nesegmentuoti laktiferiai susidaro iš milžiniškos daugiabranduolės ląstelės. Jis atsiranda ankstyvosiose vystymosi stadijose, pailgėja ir šakojasi augalui augant, sudarydamas sudėtingą tinklą, pavyzdžiui, euforbiniuose augaluose (25 pav.).
Pieninių augalų ląstelių sienelės yra stipriai laistomos. Jie nerūgsta, todėl yra plastikiniai. Centrinę lakto dalį užima vakuolė su lateksu, o protoplastas – netoli sienos. Ne visada galima nustatyti ribą tarp protoplasto ir vakuolės. Pienžolės prasiskverbia į meristemą ir auga kartu su augalo organu.
Laidūs audiniai
Šis tipas priklauso sunku audinių, susideda iš skirtingai diferencijuotų ląstelių. Be faktinių laidžių elementų, audinyje yra mechaninių, šalinimo ir saugojimo elementų (26 pav.). Laidūs audiniai sujungia visus augalų organus į vieną sistemą. Yra dviejų tipų laidūs audiniai: ksilemas Ir floem(gr. ksilonas – medis; phloios – žievė, bastas). Jie turi tiek struktūrinių, tiek funkcinių skirtumų.
Laidžius ksilemo elementus sudaro negyvos ląstelės. Jie perneša tolimais atstumais vandenį ir jame ištirpusias medžiagas nuo šaknų iki lapų. Laidūs floemo elementai palaiko protoplastą gyvą. Jie perneša didelius atstumus nuo fotosintetinių lapų iki šaknų.
Paprastai ksilemas ir floemas yra augalo kūne tam tikra tvarka, formuodami sluoksnius arba laidūs ryšuliai. Priklausomai nuo sandaros, išskiriami keli laidžių ryšulių tipai, būdingi tam tikroms augalų grupėms. Užstatu atvirame ryšulyje tarp ksilemo ir floemo yra kambis, užtikrinantis antrinį augimą (27-A, 28 pav.). Dvišaliame atvirame ryšulyje floema yra ksilemo atžvilgiu iš abiejų pusių (27-B, 29 pav.). uždari ryšuliai neturi kambio, todėl negali antrinio sutirštėjimo (27-B, 27-D, 30.31 pav.). Yra dar du tipai koncentriniai ryšuliai, kur arba floema supa ksilemą (27-E pav., 32), arba ksilemas supa floemą (27-E pav.).
Ksilemas (mediena). Ksilemo vystymasis aukštesniuose augaluose yra susijęs su vandens mainų užtikrinimu. Kadangi vanduo nuolat išsiskiria per epidermį, augalas turi sugerti tą patį drėgmės kiekį ir įpilti į transpiraciją vykdančius organus. Reikėtų atsižvelgti į tai, kad gyvo protoplasto buvimas vandeniui laidžiose ląstelėse labai sulėtintų transportavimą, o negyvos ląstelės čia yra funkcionalesnės. Tačiau negyvos ląstelės neturi drebulys , todėl apvalkalas turi turėti mechaninių savybių. Pastaba: turgescencija - augalų ląstelių, audinių ir organų būklė, kai jie tampa elastingi dėl ląstelių turinio spaudimo ant jų elastinių membranų. Iš tiesų, laidžiuosius ksilemo elementus sudaro negyvos ląstelės, ištemptos išilgai organo ašies su storais lignifikuotais apvalkalais.
Iš pradžių ksilemas susidaro iš pirminės meristemos – prokambio, esančio ašinių organų viršūnėse. Pirmiausia išskiria protoksilemas, tada metaksilema. Yra žinomi trys ksilemų susidarymo tipai. At eksarchas tipo, protoksilemo elementai pirmiausia atsiranda prokambio pluošto periferijoje, vėliau metaksilemo elementai atsiranda centre. Jei procesas vyksta priešinga kryptimi (t. y. iš centro į periferiją), tada tai endarchinis tipo. At mesarch tipas ksilemas dedamas prokambio pluošto centre, po to jis nusėda tiek link centro, tiek į periferiją.
Šaknims būdingas eksarchinis ksilemo formavimosi tipas, o stiebams – endarchinis. Žemai organizuotuose augaluose ksilemo formavimo būdai yra labai įvairūs ir gali būti sistemingi.
Kai kuriuose augaluose (pavyzdžiui, vienaląsčiuose) visos prokambio ląstelės diferencijuojasi į laidžius audinius, kurie negali antrinio sustorėjimo. Kitose formose (pavyzdžiui, sumedėjusios) tarp ksilemo ir floemo lieka šoninės meristemos (kambis). Šios ląstelės gali dalytis, atnaujindamos ksilemą ir floemą. Toks procesas vadinamas antrinis augimas. Daugelis augalų, augančių palyginti stabiliomis klimato sąlygomis, auga nuolat. Prie sezoninių klimato pokyčių pritaikytomis formomis – periodiškai. Dėl to susidaro aiškiai apibrėžti metiniai augimo žiedai.
Pagrindiniai prokambinių ląstelių diferenciacijos etapai. Jo ląstelės turi plonas membranas, kurios netrukdo joms ištempti organui augant. Tada protoplastas pradeda kloti antrinę membraną. Tačiau šis procesas turi ryškių bruožų. Antrinė membrana nusėda ne ištisiniu sluoksniu, kuris neleistų ląstelei išsitempti, o žiedų pavidalu arba spirale. Ląstelių pailgėjimas nėra sunkus. Jaunose ląstelėse spiralės žiedai arba ritės yra arti vienas kito. Brandžiose ląstelėse jos išsiskiria dėl ląstelių tempimo (33 pav.). Žieduoti ir spiraliniai lukšto sustorėjimai netrukdo augti, tačiau mechaniškai jie yra prastesni už kiautus, kur antrinis sustorėjimas sudaro vientisą sluoksnį. Todėl, nustojus augti ksileme, susidaro elementai su ištisiniu lignifikuotu apvalkalu ( metaksilema). Pažymėtina, kad antrinis sustorėjimas čia ne žiedinis ar spiralinis, o taškuotas, laiptiškas, tinklinis (34 pav.). Jo ląstelės nepajėgia ištempti ir miršta per kelias valandas. Šis procesas netoliese esančiose ląstelėse vyksta koordinuotai. Citoplazmoje atsiranda daug lizosomų. Tada jie suyra, o juose esantys fermentai sunaikina protoplastą. Sunaikinus skersines sienas, viena virš kitos grandinėje išsidėsčiusios ląstelės suformuoja tuščiavidurį indą (35 pav.). Dauguma gaubtasėklių ir kai kurie paparčiai turi indus.
Laidi ląstelė, kuri nesusiformuoja per perforacijas jos sienelėje, vadinama tracheidinė. Vandens judėjimas tracheidėmis yra lėtesnis nei per indus. Faktas yra tas, kad tracheidose pirminė membrana niekur nenutrūksta. Tracheidės bendrauja tarpusavyje nuo. Reikėtų paaiškinti, kad augaluose pora yra tik antrinės membranos įdubimas iki pirminės membranos, o tarp tracheidų nėra perforacijų.
Dažniausios yra ribojamos poros (35-1 pav.). Jų kanalas, nukreiptas į ląstelės ertmę, sudaro pratęsimą - porų kamera. Daugumos spygliuočių augalų poros ant pirminio apvalkalo yra sustorėjusios - Torus, kuris yra savotiškas vožtuvas ir gali reguliuoti vandens transportavimo intensyvumą. Pasislinkdamas toras blokuoja vandens tekėjimą per poras, tačiau po to nebegali grįžti į ankstesnę padėtį, atlikdamas vienkartinį veiksmą.
Poros yra daugiau ar mažiau suapvalintos, pailgos statmenai pailgai ašiai (šių porų grupė primena kopėčias, todėl toks poringumas vadinamas kopėčių poringumu). Transportavimas per poras atliekamas tiek išilgine, tiek skersine kryptimis. Poros yra ne tik tracheidėse, bet ir atskirose kraujagyslių ląstelėse, kurios sudaro indą.
Evoliucijos teorijos požiūriu, tracheidai yra pirmoji ir pagrindinė struktūra, kuri veda vandenį aukštesniųjų augalų kūne. Manoma, kad kraujagyslės atsirado iš tracheidų dėl tarp jų esančių skersinių sienelių lizės (36 pav.). Dauguma paparčių ir gimnasėklių neturi. Jie perkelia vandenį per tracheidą.
Evoliucinio vystymosi procese kraujagyslės atsirado ne kartą įvairiose augalų grupėse, tačiau svarbiausią funkcinę reikšmę jie įgijo gaubtasėkliuose, turinčiuose juos kartu su tracheidėmis. Manoma, kad pažangesnio transporto mechanizmo turėjimas padėjo jiems ne tik išgyventi, bet ir pasiekti didelę formų įvairovę.
Ksilemas yra sudėtingas audinys, be vandeniui laidžių elementų, jame yra ir kitų. Mechanines funkcijas atlieka libriforminiai pluoštai ( lat. liber – lub, forma – forma). Papildomų mechaninių konstrukcijų buvimas yra svarbus, nes, nepaisant sustorėjimo, vandenį nešančių elementų sienelės vis dar yra per plonos. Jie nesugeba savarankiškai išlaikyti didelės daugiamečio augalo masės. Pluoštai išsivystė iš tracheidų. Jiems būdingi mažesni dydžiai, lignified (lignified) apvalkalai ir siauros ertmės. Ant sienos galima rasti porų be apvadų. Šios skaidulos nepraleidžia vandens, jų pagrindinė funkcija yra atrama.
Ksileme taip pat yra gyvų ląstelių. Jų masė gali siekti 25% viso medienos tūrio. Kadangi šios ląstelės turi apvalią formą, jos vadinamos medienos parenchima. Augalo kūne parenchima yra dviem būdais. Pirmuoju atveju ląstelės yra išdėstytos vertikalių sruogų pavidalu - tai yra sruogų parenchima. Kitu atveju parenchima formuoja horizontalius spindulius. Jie vadinami šerdies spinduliai, nes jie sujungia šerdį ir žievę. Šerdis atlieka daugybę funkcijų, įskaitant medžiagų saugojimą.
Floemas (batas). Tai sudėtingas audinys, nes jį sudaro nevienalytės ląstelės. Pagrindinės laidžiosios ląstelės vadinamos sieto elementai(37 pav.). Laidžius ksilemo elementus sudaro negyvos ląstelės, o floeme per funkcionavimo laikotarpį jie išsaugo gyvą, nors ir labai pakitusį protoplastą. Floema atlieka plastikinių medžiagų nutekėjimą iš fotosintezės organų. Visos gyvos augalų ląstelės turi savybę pravesti organines medžiagas. Vadinasi, jei ksilemą galima rasti tik aukštesniuose augaluose, tai organinių medžiagų pernešimas tarp ląstelių taip pat vyksta žemesniuose augaluose.
Ksilemas ir floema išsivysto iš viršūninių meristemų. Pirmajame etape, prokamybinėje virvelėje, a protofloemas. Kai aplinkiniai audiniai auga, jis išsitempia, o kai augimas baigiasi, vietoj protoflemos susidaro metafloema.
Įvairiose aukštesniųjų augalų grupėse galima rasti dviejų tipų sieto elementus. Paparčiuose ir gimnasėkliuose jis vaizduojamas sieto ląstelės. Sietų laukai ląstelėse yra išsibarstę palei šonines sieneles. Protoplastas išlaiko šiek tiek sunaikintą branduolį.
Gaubtasėkliuose vadinami sieto elementais sieto vamzdeliai. Jie bendrauja tarpusavyje per sieto plokštes. Brandžios ląstelės neturi branduolių. Tačiau šalia sieto vamzdis yra kompaniono ląstelė, kuris susidaro kartu su sieto vamzdeliu dėl bendros motininės ląstelės mitozinio dalijimosi (38 pav.). Kompanioninė ląstelė turi tankesnę citoplazmą su daugybe aktyvių mitochondrijų, taip pat pilnai funkcionuojantį branduolį, daugybę plazmodesmų (dešimt kartų daugiau nei kitose ląstelėse). Kompanioninės ląstelės veikia nebranduolinių sieto vamzdelių ląstelių funkcinį aktyvumą.
Subrendusių sieto ląstelių struktūra turi tam tikrų ypatumų. Vakuolės nėra, todėl citoplazma labai suskystinta. Branduolys gali nebūti (sėkmiuose) arba gali būti susiraukšlėjęs, funkciškai neaktyvus. Ribosomų ir Golgi komplekso taip pat nėra, tačiau gerai išvystytas endoplazminis tinklas, kuris ne tik prasiskverbia į citoplazmą, bet ir per sieto laukų poras patenka į kaimynines ląsteles. Gausiai randama gerai išsivysčiusių mitochondrijų ir plastidžių.
Tarp ląstelių medžiagų transportavimas vyksta per skylutes, esančias ant ląstelių membranų. Tokios angos vadinamos poromis, tačiau skirtingai nei tracheidinės poros, jos yra pro. Daroma prielaida, kad tai labai išsiplėtusios plazmodesmos, ant kurių sienelių nusėda kaliozės polisacharidas. Poros išsidėsčiusios grupėmis, formuojasi sieto laukai. Primityviose formose sietų laukai atsitiktinai išsibarstę po visą apvalkalo paviršių, labiau išsivysčiusiuose gaubtasėkliuose jie yra gretimų ląstelių galuose, besiribojančiose viena su kita, sudarydami sieto plokštelė(39 pav.). Jei ant jo yra vienas sieto laukas, jis vadinamas paprastu, jei keli - kompleksiniu.
Tirpalų judėjimo per sieto elementus greitis yra iki 150 cm ∕ val. Tai tūkstantį kartų viršija laisvosios difuzijos greitį. Tikriausiai vyksta aktyvus transportas, o daugybė sieto elementų ir palydovinių ląstelių mitochondrijų tiekia tam reikalingą ATP.
Floemo sieto elementų tarnavimo laikas priklauso nuo šoninių meristemų buvimo. Jei jų yra, tada sieto elementai veikia visą augalo gyvenimą.
Be sieto elementų ir palydovinių ląstelių, floemoje yra karnienos pluoštų, sklereidų ir parenchimos.
Klausimai kontrolei ir kartojimui
Aukštesnių augalų ženklai? Kokios kartos aukštesni augalai vadinami gametofitais? Sporofitas?
Kokios priežastys prisideda prie diferencijuotų sausumos augalų audinių vystymosi.
Išvardykite augalų rūšis.
Kokias meristemų grupes žinote? Kur jie yra augalo kūne?
Pilnamieji audiniai, jų funkcijos, reikšmė.
Įvairių tipų vientisų audinių organizavimas.
Stomatalinio aparato sandara. Išvardykite jums žinomų stomatalinių aparatų tipus. Kaip veikia stomos?
Parenchiminių audinių ypatumai, parenchimo tipai.
Parenchimo ir sklerenchimo požymiai.
Išvardykite augalo išskyrimo struktūrų tipus. Kokia jų reikšmė?
Išskiriamų medžiagų talpyklų formavimo metodai.
Kas yra melžėjai? Į kokias rūšis jie skirstomi?
Kaip organizuojami laidūs audiniai?
Kas yra laidūs pluoštai? Jų rūšys.
Medžiagų judėjimo per ksilemą kryptys? Floemas? Ar transportuojamų medžiagų sudėtis skiriasi ksilemu ir floemu?
Ksilemo, floemo sandara ir funkcijos.
Floemą laidžių elementų organizavimas augaluose iš įvairių sisteminių grupių.
Specializuotas ir bakterinis, turi išorinę ląstelės sienelę. Aukštesnėse ląstelėse Organelių struktūra ir funkcijos ląstelės. G. Mendelio dėsniaiKontrolinio darbo >>
Tipas audiniai. Mūsų artimiausi giminaičiai, kiti žinduoliai, struktūra ir... , susidedantis iš vieno ląstelės. Augalai taip pat susideda iš įvairių ... mikrovamzdelių, centriolių ląstelių centras. IN daržovių ląstelės Taip pat išskiriami chloroplastai, kuriuose ...
Raudona spalva. IN audinių augalai antraceno darinių galima rasti ... antraceno darinių kaip ir vaistinėje daržoviųžaliavos, ir ... yra apsuptos 2-3, rečiau 4 ląstelės epidermis. Vienaląsčiai plaukai, ... junginiai, pagrindu pastatai iš kurių antraceno struktūra...