lauke citoplazminė membrana yra plona plėvelė. Jo storis apie 7-10 nm. Filmas žiūrimas tik elektroninis mikroskopas. Toliau apsvarstykite, kas yra citoplazminė membrana. Filmo funkcijos taip pat bus aprašytos straipsnyje.
Struktūra
Kokia yra citoplazminės membranos sudėtis? Filmo struktūra gana įvairi. Pagal cheminė organizacija, tai baltymų ir lipidų kompleksas. Ląstelės citoplazminė membrana apima dvisluoksnį sluoksnį. Jis veikia kaip pagrindas. Be to, citoplazmos membranoje yra cholesterolio ir glikolipidų. Šios medžiagos yra amfipatrinės. Kitaip tariant, jie turi hidrofobinius („bijo drėgmės“) ir hidrofilinius („mėgsta vandenį“) galus. Pastarieji (fosfatų grupė) yra nukreipti į išorę nuo membranos, pastarieji (likučiai iš riebalų rūgštys) yra orientuoti vienas į kitą. Dėl to susidaro lipidinis bipolinis sluoksnis. Lipidų molekulės yra mobilios. Jie gali judėti savo viename sluoksnyje arba (kas yra retai) iš vieno į kitą. 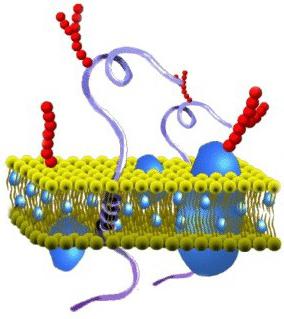 Lipidų sluoksnis gali būti kieto arba skysto kristalo būsenos. Monosluoksniai yra asimetriški. Tai reiškia, kad jie turi skirtingą lipidų sudėtį. Dėl šios savybės citoplazminės membranos turi specifiškumą net vienoje ląstelėje. Baltymai yra antrasis privalomas plėvelės komponentas. Daugelis šių junginių gali judėti membranos plokštumoje arba suktis aplink savo ašį. Tačiau jie negali pereiti iš vienos dvisluoksnės dalies į kitą. Apsauga vidinė aplinka- pagrindinė užduotis, kurią atlieka citoplazminė membrana. Be to, filmo struktūra suteikia srautą įvairūs procesai. Baltymai yra atsakingi už tam tikrų užduočių atlikimą. Lipidų dėka suteikiamos plėvelės struktūrinės savybės.
Lipidų sluoksnis gali būti kieto arba skysto kristalo būsenos. Monosluoksniai yra asimetriški. Tai reiškia, kad jie turi skirtingą lipidų sudėtį. Dėl šios savybės citoplazminės membranos turi specifiškumą net vienoje ląstelėje. Baltymai yra antrasis privalomas plėvelės komponentas. Daugelis šių junginių gali judėti membranos plokštumoje arba suktis aplink savo ašį. Tačiau jie negali pereiti iš vienos dvisluoksnės dalies į kitą. Apsauga vidinė aplinka- pagrindinė užduotis, kurią atlieka citoplazminė membrana. Be to, filmo struktūra suteikia srautą įvairūs procesai. Baltymai yra atsakingi už tam tikrų užduočių atlikimą. Lipidų dėka suteikiamos plėvelės struktūrinės savybės.
Citoplazminė membrana: funkcijos
Pagrindinės užduotys yra šios:
- Barjeras. Apsauginė plėvelė užtikrina aktyvų, pasyvų, selektyvų, reguliuojamą jungčių keitimąsi su išorinė aplinka. Dėl selektyvaus pralaidumo ląstelė ir jos skyriai yra atskiriami ir aprūpinami reikalingomis medžiagomis.
- Transportas. Per plėvelę vyksta junginių perėjimas iš ląstelės į ląstelę. Dėl to tiekiami maistiniai junginiai, pašalinami galutiniai metabolizmo produktai ir atsiranda sekrecija. skirtingos medžiagos. Be to, susidaro joniniai gradientai, palaikoma optimali jonų koncentracija ir pH. Jie būtini aktyviai ląstelių fermentų veiklai.
Pagalbinės užduotys
Ypatingos savybės
Konkrečios membranos funkcijos apima:
Papildoma informacija
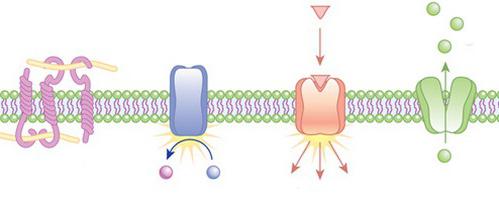
Jei dėl vienokių ar kitokių priežasčių kai kurios dalelės negali prasiskverbti pro fosfolipidų dvisluoksnį sluoksnį (pavyzdžiui, dėl hidrofilinių savybių, nes citoplazminė membrana viduje yra hidrofobinė ir nepraleidžia tokių junginių, arba dideli dydžiai pačios dalelės), tačiau jos būtinos, gali prasiskverbti pasitelkus specialius nešiklius (transporterius) ir kanalinius baltymus. Arba jų prasiskverbimas atliekamas endocitozės būdu. 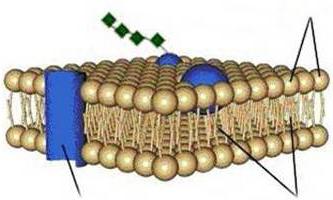 Pasyviojo transportavimo procese medžiagos difuzijos būdu kerta lipidų sluoksnį. Tokiu atveju energija nėra švaistoma. Supaprastinta difuzija gali būti vienas iš tokio mechanizmo variantų. Jo metu tam tikra molekulė palengvina medžiagos praėjimą. Jis gali turėti kanalą, galintį praleisti tik to paties tipo daleles. Aktyvus transportas sunaudoja energiją. Taip yra dėl to, kad šis procesas atliekamas atsižvelgiant į koncentracijos gradientą. Citoplazminėje membranoje yra specialūs siurblio baltymai, įskaitant ATPazę, kuri skatina aktyvų kalio patekimą ir natrio jonų išsiskyrimą.
Pasyviojo transportavimo procese medžiagos difuzijos būdu kerta lipidų sluoksnį. Tokiu atveju energija nėra švaistoma. Supaprastinta difuzija gali būti vienas iš tokio mechanizmo variantų. Jo metu tam tikra molekulė palengvina medžiagos praėjimą. Jis gali turėti kanalą, galintį praleisti tik to paties tipo daleles. Aktyvus transportas sunaudoja energiją. Taip yra dėl to, kad šis procesas atliekamas atsižvelgiant į koncentracijos gradientą. Citoplazminėje membranoje yra specialūs siurblio baltymai, įskaitant ATPazę, kuri skatina aktyvų kalio patekimą ir natrio jonų išsiskyrimą.
Modeliai
Jų yra keletas:
- "Sumuštinio modelis". Idėją apie trijų sluoksnių visų membranų struktūrą mokslininkai Dawsonas ir Danieli išreiškė 1935 m. Jų nuomone, plėvelės struktūra buvo tokia: baltymai-lipidai-baltymai. Ši idėja gyvavo jau seniai.
- „Skystai-mozaikinė struktūra“.Šį modelį Nicholsonas ir Singer aprašė 1972 m. Pagal jį baltymų molekulės nesudaro ištisinio sluoksnio, o yra panardintos į bipolinį lipidų sluoksnį mozaikos pavidalu į skirtingus gylius. Šis modelis laikomas universaliausiu.
- „Baltymų-kristalų struktūra“. Pagal šį modelį membranos susidaro dėl baltymų ir lipidų molekulių susipynimo, kurios yra sujungtos hidrofilinių-hidrofobinių ryšių pagrindu.
Jis išsivystė taip, kad kiekvienos jos sistemos funkcija buvo ląstelių, sudarančių šios sistemos organus ir audinius, sumos funkcijos rezultatas. Kiekviena kūno ląstelė turi aibę struktūrų ir mechanizmų, leidžiančių jai pačiai vykdyti medžiagų apykaitą ir atlikti savo funkcijas.
Ląstelėje yra citoplazminė arba paviršinė membrana; citoplazma, turinti daugybę organelių, inkliuzų, citoskeleto elementų; branduolys, kuriame yra branduolio genomas. Ląstelių organelės ir branduolys citoplazmoje yra apriboti vidinėmis membranomis. Kiekviena ląstelės struktūra joje atlieka savo funkciją, ir visos jos kartu užtikrina ląstelės gyvybingumą ir specifinių jos funkcijų atlikimą.
Pagrindinis vaidmuo įgyvendinant ląstelių funkcijos ir jų reguliavimas priklauso ląstelės citoplazminei membranai.
Bendrieji citoplazminės membranos sandaros principai
Visos ląstelių membranos turi tą patį struktūrinį principą.(1 pav.), kuris yra pagrįstas fizikines ir chemines savybes sudėtingi lipidai ir baltymai, sudarantys jų sudėtį. Ląstelių membranos yra vandeninėje terpėje, o norint suprasti fizikinius ir cheminius reiškinius, turinčius įtakos jų struktūrinei organizacijai, naudinga aprašyti lipidų ir baltymų molekulių sąveiką su vandens molekulėmis ir tarpusavyje. Atsižvelgiant į šią sąveiką, taip pat atsiranda keletas ląstelių membranų savybių.
Yra žinoma, kad ląstelės plazminę membraną vaizduoja dvigubas sudėtingų lipidų sluoksnis, dengiantis ląstelės paviršių per visą jos ilgį. Norint sukurti lipidų dvigubą sluoksnį, pagal prigimtį gali būti atrinktos ir į jo struktūrą įtrauktos tik tos lipidų molekulės, kurios turi amfifilinių (amfipatinių) savybių. Fosfolipidų ir cholesterolio molekulės atitinka šias sąlygas. Jų savybės tokios, kad viena molekulės dalis (glicerolis – fosfolipidams, o ciklopentanas – cholesteroliui) turi polinių (hidrofilinių), o kita (riebalų rūgščių radikalai) – nepolinių (hidrofobinių) savybių.
Ryžiai. 1. Ląstelės citoplazminės membranos sandara.
Jei tam tikras skaičius fosfolipidų ir cholesterolio molekulių dedamas į vandeninę terpę, jie spontaniškai pradės kauptis į tvarkingas struktūras ir sudarys uždarus burbulus ( liposomos), kuriame yra dalis vandens aplinka, o paviršius pasidengia ištisiniu dvigubu sluoksniu ( dvisluoksnis) fosfolipidų molekulės ir cholesterolis. Įvertinus fosfolipidų ir cholesterolio molekulių erdvinio išsidėstymo šiame dvisluoksnyje pobūdį, aišku, kad šių medžiagų molekulės yra hidrofilinėmis dalimis link išorinės ir vidinės vandens erdvės, o hidrofobinės – priešingomis kryptimis – dvisluoksnio viduje. .
Dėl ko šių lipidų molekulės spontaniškai sudaro dvisluoksnes struktūras, panašias į dvisluoksnę struktūrą vandeninėje terpėje ląstelės membrana? Erdvinį amfifilinių lipidų molekulių išsidėstymą vandeninėje terpėje lemia vienas iš termodinamikos reikalavimų. Labiausiai tikėtina erdvinė struktūra, kurią lipidų molekulės susidarys vandeninėje terpėje struktūra su minimalia laisvąja energija.
Toks laisvos energijos minimumas vandens lipidų erdvinėje struktūroje bus pasiektas, kai tiek hidrofilinės, tiek hidrofobinės molekulių savybės bus realizuotos atitinkamų tarpmolekulinių ryšių pavidalu.
Nagrinėjant sudėtingų amfifilinių lipidų molekulių elgesį vandenyje, kai kurie ląstelių membranų savybės. Yra žinoma, kad jeigu mechaniškai pažeista plazminė membrana(pavyzdžiui, pradurkite jį elektrodu arba pašalinkite branduolį per punkciją ir įdėkite kitą branduolį į ląstelę), tada akimirksniu dėl lipidų ir vandens tarpmolekulinės sąveikos jėgų membrana spontaniškai atkurs vientisumą. Tų pačių jėgų įtakoje galima stebėti dviejų membranų dviejų sluoksnių susiliejimas, kai jie liečiasi(pvz., pūslelės ir presinapsinės membranos sinapsėse). Membranų gebėjimas susijungti tiesioginio kontakto metu yra membranos struktūros atsinaujinimo, membranos komponentų pernešimo iš vienos tarpląstelinės erdvės į kitą mechanizmų, taip pat endo- ir egzocitozės mechanizmų dalis.
Tarpmolekulinių ryšių energija lipidų dvisluoksnyje labai žemai, todėl susidaro sąlygos greitam lipidų ir baltymų molekulių judėjimui membranoje ir membranos struktūros pasikeitimui jai veikiant. mechaninės jėgos, slėgis, temperatūra ir kiti veiksniai. Dvigubo lipidinio sluoksnio buvimas membranoje sudaro uždarą erdvę, izoliuoja citoplazmą nuo supančios vandens aplinkos ir sukuria kliūtį laisvam vandens ir jame tirpių medžiagų judėjimui per ląstelės membraną. Lipidų dvigubo sluoksnio storis yra apie 5 nm.
Ląstelių membranose taip pat yra baltymų. Jų molekulės yra 40-50 kartų didesnės tūrio ir masės nei membraninių lipidų molekulės. Dėl baltymų membranos storis siekia 7-10 nm. Nepaisant to, kad bendroji baltymų ir lipidų masė daugumoje membranų yra beveik vienoda, baltymų molekulių skaičius membranoje yra dešimt kartų mažesnis nei lipidų molekulių.
Kas atsitiks, jei baltymo molekulė bus patalpinta į liposomų dvisluoksnį fosfolipidinį sluoksnį, išorinį ir vidiniai paviršiai kurie yra poliniai, o intralipidas yra nepolinis? Veikiant lipidų, baltymų ir vandens tarpmolekulinės sąveikos jėgoms, susiformuos tokia erdvinė struktūra, kurioje nepolinės peptidinės grandinės sritys bus linkusios nusėsti lipidinio dvisluoksnio sluoksnio gylyje, o poliniai užims poziciją ant vieno iš dvisluoksnio sluoksnio paviršių ir taip pat gali būti panardinti į išorinę arba vidinę liposomos vandeninę aplinką. Labai panašus baltymų molekulių išsidėstymo pobūdis vyksta ir ląstelių membranų lipidiniame dvisluoksnyje (1 pav.).
Paprastai baltymų molekulės yra lokalizuotos membranoje atskirai viena nuo kitos. Labai silpnos hidrofobinės sąveikos tarp lipidų molekulių angliavandenilių radikalų ir baltymo molekulės nepolinių sričių (lipidų-lipidų, lipidų ir baltymų sąveikos), atsirandančios nepolinėje lipidų dvisluoksnio dalyje, jėgos netrukdo šiluminės difuzijos procesams. šios molekulės dvisluoksnėje struktūroje.
Subtiliais tyrimo metodais ištyrus ląstelių membranų struktūrą, paaiškėjo, kad ji labai panaši į tą, kurią vandens aplinkoje spontaniškai susidaro fosfolipidai, cholesterolis ir baltymai. 1972 m. Singer ir Nichols pasiūlė skysčio mozaikinį ląstelės membranos struktūros modelį ir suformulavo pagrindinius jo principus.
Pagal šį modelį visų ląstelių membranų struktūrinis pagrindas yra į skystį panašus ištisinis dvigubas amfipatinių fosfolipidų, cholesterolio, glikolipidų molekulių sluoksnis, spontaniškai jį susidarantis vandens aplinkoje. Lipidų dvisluoksnyje asimetriškai išsidėsčiusios baltymų molekulės, kurios atlieka specifines receptorių, fermentines ir transportavimo funkcijas. Baltymų ir lipidų molekulės yra judrios ir gali atlikti sukamuosius judesius, difunduoti dvisluoksnio plokštumoje. Baltymų molekulės gali keisti savo erdvinę struktūrą (konformaciją), pasislinkti ir keisti savo padėtį lipidiniame dvisluoksniame membranos sluoksnyje, pasinerdamos į skirtingą gylį arba plūduriuodamos iki jo paviršiaus. Membranos lipidinio dvigubo sluoksnio struktūra yra nevienalytė. Jame yra sričių (domenų), vadinamų „plaustais“, kurios yra praturtintos sfingolipidais ir cholesteroliu. „Praustai“ faze skiriasi nuo likusios membranos, kurioje jie yra, būsenos. Membranų struktūrinės savybės priklauso nuo jų atliekamos funkcijos ir funkcinės būklės.
Ląstelių membranų sudėties tyrimas patvirtino, kad pagrindiniai jų komponentai yra lipidai, kurie sudaro apie 50% plazminės membranos masės. Apie 40–48 % membranos masės sudaro baltymai, o 2–10 % – angliavandeniai. Angliavandenių likučiai įeina į baltymus, sudarydami glikoproteinus, arba į lipidus, sudarydami glikolipidus. Fosfolipidai yra pagrindiniai struktūriniai lipidai plazminės membranos ir sudaro 30–50 % jų masės.
Glikolipidų molekulių angliavandenių likučiai dažniausiai yra išoriniame membranos paviršiuje ir panardinami į vandeninę terpę. Jie žaidžia svarbus vaidmuo tarpląstelinėje, ląstelių-matricos sąveikoje ir imuninės sistemos ląstelių atpažįstant antigenus. Cholesterolio molekulės, įterptos į fosfolipidų dvigubą sluoksnį, padeda išlaikyti tvarkingą fosfolipidų riebalų rūgščių grandinių išsidėstymą ir jų skystųjų kristalų būseną. Dėl didelio fosfolipiduose esančių riebalų rūgščių acilo radikalų konformacinio mobilumo jie sudaro gana laisvą lipidinio dvisluoksnio sluoksnį ir jame gali susidaryti struktūriniai defektai.
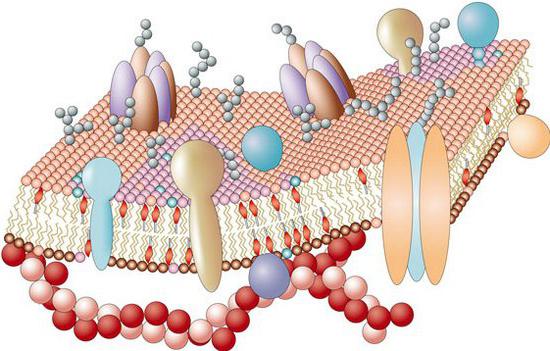
Baltymų molekulės gali prasiskverbti per visą membraną taip, kad jų galinės dalys išsikištų už skersinių ribų. Tokie baltymai vadinami transmembraninis, arba integralas. Membranose taip pat yra baltymų, kurie tik iš dalies panardinami į membraną arba yra ant jos paviršiaus.
Daug specifinės membranų funkcijos yra nulemtos baltymų molekulių, kurių lipidų matrica yra tiesioginė mikroaplinka, o baltymų molekulių funkcijų įgyvendinimas priklauso nuo jo savybių. Iš svarbiausių membraninių baltymų funkcijų galima išskirti: receptorius – prisijungimą prie tokių signalinių molekulių kaip neurotransmiteriai, hormonai, ingerleukinai, augimo faktoriai ir signalo perdavimas į ląstelės postreceptorines struktūras; fermentinis - tarpląstelinių reakcijų katalizė; struktūrinis - dalyvavimas formuojant pačios membranos struktūrą; transportavimas – medžiagų pernešimas per membranas; kanalų formavimas – joninių ir vandens kanalų susidarymas. Baltymai kartu su angliavandeniais dalyvauja vykdant sukibimą, lipnumą, ląstelių klijavimą imuninės reakcijos, sujungdami ląsteles į sluoksnius ir audinius, užtikrina ląstelių sąveiką su tarpląsteline matrica.
Membraninių baltymų (receptorių, fermentų, nešėjų) funkcinį aktyvumą lemia jų gebėjimas lengvai keisti savo erdvinę struktūrą (konformaciją), kai jie sąveikauja su signalinėmis molekulėmis, fiziniai veiksniai arba keičiant mikroaplinkos aplinkos savybes. Energija, reikalinga šiems konformaciniams baltymų struktūros pokyčiams atlikti, priklauso ir nuo intramolekulinių sąveikos jėgų. atskiri skyriai peptidų grandinę ir membranos lipidų, tiesiogiai supančių baltymą, takumo (mikroklampumo) laipsnį.
Angliavandeniai glikolipidų ir glikoproteinų pavidalu sudaro tik 2-10% membranos masės; jų skaičius skirtingos ląstelės keičiamas. Jų dėka atliekamos tam tikros tarpląstelinės sąveikos rūšys, jos dalyvauja ląstelėje atpažįstant svetimus antigenus ir kartu su baltymais sukuria savotišką savo ląstelės paviršiaus membranos antigeninę struktūrą. Pagal tokius antigenus ląstelės atpažįsta viena kitą, susijungia į audinius ir trumpam laikui sulipti, kad perduotų signalines molekules viena kitai.
Dėl mažos į membraną įtrauktų medžiagų sąveikos energijos ir santykinio jų išsidėstymo tvarkingumo ląstelės membrana įgyja nemažai savybių ir funkcijų, kurių negalima redukuoti į paprastą ją sudarančių medžiagų savybių sumą. Nežymus poveikis membranai, prilyginamas baltymų ir lipidų tarpmolekulinių ryšių energijai, gali lemti baltymų molekulių konformacijos pokyčius, jonų kanalų pralaidumą, membranos receptorių savybių pokyčius ir daugybę kitų ląstelių funkcijų. membrana ir pati ląstelė. Didelis jautrumas konstrukciniai komponentai plazminė membrana turi lemiamas ląstelėje suvokiant informacinius signalus ir paverčiant juos ląstelės atsakais.
Ląstelės citoplazminės membranos funkcijos
Citoplazminė membrana atlieka daugybę funkcijų, kurios užtikrina gyvybiškai svarbius ląstelės poreikius. ir ypač daug funkcijų, reikalingų ląstelei suvokti ir perduoti informacinius signalus.
Tarp svarbiausių plazminės membranos funkcijų yra:
- ląstelės atribojimas nuo aplinkos, išlaikant formą, tūrį ir reikšmingus ląstelių turinio ir tarpląstelinės erdvės skirtumus;
- medžiagų perkėlimas į ląstelę ir iš jos, remiantis selektyvaus pralaidumo, aktyvaus ir kitų transportavimo būdų savybėmis;
- transmembraninio elektrinio potencialo skirtumo (membranos poliarizacijos) palaikymas ramybės būsenoje, jo kitimas veikiant įvairiems poveikiams ląstelei, sužadinimo generavimas ir laidumas;
- dalyvavimas aptinkant (priimant) signalus fizinė prigimtis, signalines molekules dėl sensorinių ar molekulinių receptorių susidarymo ir signalų perdavimo į ląstelę;
- tarpląstelinių kontaktų (glaudaus, tarpo ir desmosominio kontakto) susidarymas susidariusių audinių sudėtyje arba įvairių audinių ląstelių sukibimo metu;
- hidrofobinės mikroaplinkos sukūrimas su membrana susijusių fermentų aktyvumui pasireikšti;
- užtikrinant ląstelės imuninį specifiškumą dėl baltyminio arba glikoproteininio pobūdžio antigenų buvimo membranos struktūroje. Imuninis specifiškumas yra svarbus, kai ląstelės susijungia į audinį ir sąveikauja su imuninės sistemos stebėjimo ląstelėmis organizme.
Aukščiau pateiktas ląstelių membranų funkcijų sąrašas rodo, kad jos dalyvauja įgyvendinant ne tik ląstelių funkcijas, bet ir pagrindinius organų, audinių ir viso organizmo gyvybinės veiklos procesus. Nežinant daugelio reiškinių ir procesų, kuriuos suteikia membranos struktūros, neįmanoma suprasti ir sąmoningai atlikti kai kurių diagnostinės procedūros Ir medicinines priemones. Pavyzdžiui, už teisingas pritaikymas daug vaistinių medžiagų būtina žinoti, kiek kiekvienas iš jų prasiskverbia pro ląstelių membranas iš kraujo į audinių skystis ir į ląsteles.
Jo storis yra 8-12 nm, todėl jo neįmanoma apžiūrėti šviesos mikroskopu. Membranos struktūra tiriama elektroniniu mikroskopu.
Plazminę membraną sudaro du lipidų sluoksniai – lipidinis sluoksnis arba dvisluoksnis. Kiekviena lipidų molekulė susideda iš hidrofilinės galvos ir hidrofobinės uodegos. biologinės membranos lipidai išsidėstę galva į išorę, uodegos į vidų.
Daugybė baltymų molekulių yra panardintos į bilipidinį sluoksnį. Kai kurie iš jų yra membranos paviršiuje (išorėje arba vidinėje), kiti prasiskverbia pro membraną.
Plazminės membranos funkcijos
Membrana apsaugo ląstelės turinį nuo pažeidimų, palaiko ląstelės formą, selektyviai praeina reikalingų medžiagų ląstelės viduje ir pašalina medžiagų apykaitos produktus, taip pat užtikrina ryšį tarp ląstelių.
Užtvarinė, ribojanti membranos funkcija suteikia dvigubą lipidų sluoksnį. Jis neleidžia ląstelės turiniui plisti, susimaišyti su aplinką arba intersticinis skystis, ir neleidžia pavojingoms medžiagoms prasiskverbti į ląstelę.
Nemažai svarbiausių citoplazminės membranos funkcijų atliekama dėl į ją panardintų baltymų. Receptorių baltymų pagalba ląstelė gali suvokti įvairius dirginimus savo paviršiuje. Transporto baltymai sudaro ploniausius kanalus, kuriais kalio, kalcio, natrio ir kiti mažo skersmens jonai patenka į ląstelę ir iš jos. Fermentiniai baltymai užtikrina gyvybinius procesus pačioje ląstelėje.
Didelės maisto dalelės, kurios negali praeiti plonais membraniniais kanalais, patenka į ląstelę fagocitozės arba pinocitozės būdu. Dažnas vardasšie procesai vadinami endocitoze.
Kaip atsiranda endocitozė – didelių maisto dalelių įsiskverbimas į ląstelę
Maisto dalelė liečiasi su išorinė membrana ląstelėse, o šioje vietoje susidaro invaginacija. Tada dalelė, apsupta membranos, patenka į ląstelę, susidaro virškinimo vakuolė, o į susidariusią pūslelę prasiskverbia virškinimo fermentai.
Baltieji kraujo kūneliai, galintys sugauti ir virškinti svetimas bakterijas, vadinami fagocitais.
Pinocitozės atveju membranos invaginacija užfiksuoja ne kietąsias daleles, o skysčio lašelius su joje ištirpusiomis medžiagomis. Šis mechanizmas yra vienas iš pagrindinių medžiagų prasiskverbimo į ląstelę būdų.
Augalų ląstelės, padengtos membrana kietu ląstelės sienelės sluoksniu, nėra pajėgios fagocitozei.
Atvirkštinis endocitozės procesas yra egzocitozė. Ląstelėje susintetintos medžiagos (pavyzdžiui, hormonai) supakuojamos į membranines pūsleles, priartėja prie membranos, įterpiamos į ją, o pūslelės turinys išstumiamas iš ląstelės. Taigi ląstelė gali atsikratyti ir nereikalingų medžiagų apykaitos produktų.
pagrindu Plazlema, kaip ir kitos ląstelių membranos (pavyzdžiui, mitochondrijos, plastidės ir kt.), yra lipidų sluoksnis, turintis dvi molekulių eiles (1 pav.). Kadangi lipidų molekulės yra polinės (vienas iš jų polių yra hidrofilinis, tai yra, jį traukia vanduo, o kitas yra hidrofobinis, tai yra, jį atstumia vanduo), jie yra tam tikra tvarka. Vieno sluoksnio molekulių hidrofiliniai galai yra nukreipti į vandeninę terpę - į ląstelės citoplazmą, o kito sluoksnio - į išorę iš ląstelės - į tarpląstelinę medžiagą (daugialąsčiuose organizmuose) arba į vandeninę terpę (vienaląsčiuose organizmuose). ).
Ryžiai. 1. Ląstelės membranos sandara pagal skystįmozaikinis modelis. Baltymai ir glikoproteinai panardinami į dvigubąlipidų molekulių sluoksnis, nukreiptas į jų hidrofilinesgalai (apskritimai) į išorę ir hidrofobiniai (banguotos linijos) -giliai į membraną
Jie išskiria periferinius baltymus (jie yra tik vidiniu arba išorinis paviršius membranos), vientisas nye (jie yra tvirtai įterpti į membraną, panardinti į ją, gali pakeisti savo padėtį priklausomai nuo būsenos ląstelės). Membraninių baltymų funkcijos: receptorinis, struktūrinis(palaiko ląstelės formą), fermentinis, lipnus, antigeninis, transportinis.
Baltymų molekulės yra mozaikiškai įterptos į bimolekulinį lipidų sluoksnį. SU lauke gyvūnų ląstelė polisacharidų molekulės prisijungia prie lipidų ir plazmos membranos baltymų molekulių, sudarydamos glikolipidus ir glikoproteinus.
Šis agregatas sudaro glikokalikso sluoksnį. Plazlemos receptorių funkcija yra susijusi su ja (žr. toliau); taip pat gali kaupti įvairias ląstelės naudojamas medžiagas. Be to, glikokaliksas padidina mechaninį plazmalemos stabilumą.
Augalų ir grybų ląstelėse taip pat yra ląstelių sienelės, kuris atlieka pagalbinį ir apsauginį vaidmenį. Augaluose jį sudaro celiuliozė, o grybuose - chitinas.
Elementariosios membranos struktūrinė schema yra skysta-mozaikinė: riebalai sudaro skystų kristalų rėmą, o baltymai yra mozaikiškai įterpti į jį ir gali keisti savo padėtį.
Svarbiausia membranos funkcija: skatina skyrimąsi – poląstelės turinio padalijimas į atskiras ląsteles, besiskiriančias cheminės ar fermentinės sudėties detalėmis. Taip pasiekiamas aukštas bet kurios eukariotinės ląstelės vidinio turinio tvarkingumas. Suskirstymas prisideda ląstelėje vykstančių procesų erdvinis atskyrimas ke. Atskirą skyrių (ląstelę) vaizduoja kokia nors membranos organelė (pavyzdžiui, lizosoma) arba jos dalis (cristae, apribotas vidine mitochondrijų membrana).
Kitos funkcijos:
1) barjeras (vidinio ląstelės turinio ribojimas);
2) struktūrinis (suteikia ląstelėms tam tikrą formąatsakomybė už atliktas funkcijas);
3) apsauginis (dėl selektyvaus pralaidumo, priėmimoir membranos antigeniškumas);
4) reguliavimo (selektyvaus pralaidumo reguliavimas už įvairių medžiagų(pasyvus transportavimas be energijos sąnaudų pagal difuzijos arba osmoso dėsnius ir aktyvus transportavimas su energijos sąnaudomis pinocitozės, endo- ir egzocitozės būdu, natrio-kalio siurblio veikimas, fagocitozė)). Ištisas ląsteles arba dideles daleles pasiglemžia fagocitozė (pavyzdžiui, atminkite, kad amebomis maitinasi ar fagocitozė, kurią sukelia apsauginės bakterijų kraujo ląstelės). Pinocitozės metu absorbuojamos mažos dalelės arba lašeliai skysta medžiaga. Abiem procesams būdinga tai, kad absorbuojamos medžiagos yra apsuptos invaginuojančia išorine membrana, kurioje susidaro vakuolė, kuri vėliau juda į ląstelės citoplazmos gelmes. Egzocitozė – fagocitozei ir pinocitozei priešinga kryptimi vykstantis procesas (taip pat ir aktyvus transportas) (13 pav.). Jis gali būti naudojamas išvesti nesuvirškintų likučių maistas pirmuoniuose arba sekrecinėje ląstelėje susidariusios biologiškai aktyvios medžiagos.
5) lipni funkcija (visos ląstelės yra tarpusavyje sujungtos per tam tikrus kontaktus (sandarus ir laisvus));
6) receptorius (dėl periferinių membranų baltymų darbo). Yra nespecifinių receptorių, kurie suvokia kelis dirgiklius (pavyzdžiui, šalčio ir karščio termoreceptoriai), ir specifinių, kurie suvokia tik vieną dirgiklį (akies šviesą suvokiančios sistemos receptoriai);
7) elektrogeninis (ląstelės paviršiaus elektrinio potencialo pokytis dėl kalio ir natrio jonų persiskirstymo (membranos potencialas). nervų ląstelės yra 90 mV));
8) antigeninis: susijęs su glikoproteinais ir membraniniais polisacharidais. Kiekvienos ląstelės paviršiuje yra baltymų molekulių, kurios būdingos tik šio tipo ląstelėms. Su jų pagalba Imuninė sistema gebantis atskirti savas ir nesavas ląsteles. Vyksta medžiagų apykaita tarp ląstelės ir aplinkos Skirtingi keliai- pasyvus ir aktyvus.