Mitochondrijos. Gyvūnų ir augalų ląstelių citoplazmoje yra vadinamosios energetinės organelės – mitochondrijos (iš graikų „mitos“ – siūlas, „chondrion“ – grūdas). Mitochondrijų forma yra skirtinga, jos gali būti ovalios, lazdelės formos, siūlinės, kurių vidutinis skersmuo yra 1 µm, o ilgis – 7 µm. Mitochondrijų skaičius priklauso nuo ląstelės funkcinio aktyvumo ir gali siekti dešimtis tūkstančių skraidančių vabzdžių raumenyse.
Vidinė struktūra mitochondrijos (11, 12; 15 pav.) buvo tiriamos elektroniniu mikroskopu. Elektroninės mikrografijos rodo, kad mitochondrijas išoriškai riboja išorinė membrana, kurios struktūra iš esmės tokia pati kaip ir plazminės membranos. Po išorine membrana yra vidinė membrana, kuri sudaro daugybę raukšlių - cristae. Mitochondrijų viduje yra RNR, DNR ir ribosomos, kurios skiriasi nuo citoplazminių. Jo membranose yra įmontuoti specifiniai fermentai, kurių pagalba energija paverčiama mitochondrijomis. maistinių medžiagųį ATP energiją, būtiną ląstelės ir viso organizmo gyvybei.
Ryžiai. 15. Mitochondrijų sandaros schema
Plastidai. Tai organelės, būdingos tik augalų ląstelėms. Yra trijų tipų plastidai: žali chloroplastai, spalvoti (bet ne žali) chromoplastai ir bespalviai leukoplastai.
Chloroplastas(16 pav.) savo forma primena 4-6 mikronų skersmens diską ar rutulį su dviguba membrana – išorine ir vidine. Chloroplasto viduje yra DNR, ribosomos ir specialios membranos struktūros – grūdeliai, sujungti tarpusavyje ir su vidiniu memoranu bei chloroplastu, kiekvienas chloroplastas turi apie 50 grūdelių, išsidėsčiusių šaškių lentos raštu, kad geriau užfiksuotų šviesą. Gran membranose yra žalio pigmento chlorofilo. Chloroplastuose esančio chlorofilo dėka paverčiama energija saulės šviesaį cheminę ATP energiją. ATP energija naudojama chloroplastuose sintezei organiniai junginiai pirmiausia angliavandeniai.
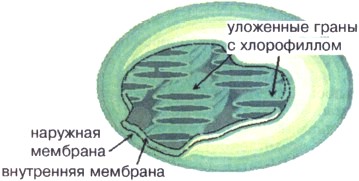
Ryžiai. 16. Chloroplasto sandaros schema
Chromoplastai. Pigmentai raudonos ir geltona spalva, esantys chromoplastuose, įvairioms augalų dalims suteikia raudoną ir geltoną spalvą. Morkų šaknys, pomidorų vaisiai yra spalvoti dėl chromoplastuose esančių pigmentų. Chromoplastų, turinčių skirtingų pigmentų, derinys sukuria daugybę gėlių ir augalų vaisių spalvų.
Leukoplastai yra rezervinės maistinės medžiagos – krakmolo – kaupimosi vieta. Ypač daug leukoplastų yra bulvių gumbų ląstelėse. Šviesoje leukoplastai gali virsti chloroplastais (dėl to bulvių gumbai pažaliuoja). Rudenį chloroplastai virsta chromoplastais, o žalieji lapai ir vaisiai tampa geltoni ir raudoni.
Nuolatinis ląstelės judėjimas. Judėjimo organelės. Daugelis vienaląsčių ir daugialąsčiai organizmai turi galimybę judėti. Tai reiškia tiek ląstelės judėjimą erdvėje, tiek tarpląstelinį jos organelių judėjimą. Skystoje terpėje ląstelių judėjimas vykdomas judant žvyneliams ir blakstienoms (10, 2, 8, 11 pav.). Taip juda daugelis vienaląsčių organizmų, pavyzdžiui, žalioji euglena, žiogelis, infuzorija ir kt. Kai kurios bakterijų rūšys taip pat juda žiuželių pagalba, ilgos ir lanksčios, kurios greitai sukasi, užtikrindamos ląstelių progresavimą. Amebos ir kai kurie kiti pirmuonys, taip pat specializuotos daugialąsčių organizmų ląstelės (pavyzdžiui, limfocitai) juda ląstelių paviršiuje susidariusių ataugų pagalba.
Ląstelė yra viduje nuolatiniame judėjime. Fagocitozės ir pinocitozės metu į ląstelę išsikiša plazminė membrana, juda lizosomos, Golgi komplekso pūslelės, mitochondrijos, galiausiai – pati citoplazma.
Ląstelių judėjimą užtikrina citoskeletas, susidedantis iš mikrotubulių, mikrofilamentų ir ląstelių centras. Mikrovamzdeliai yra ilgi tuščiaviduriai 25 nm skersmens cilindrai, kurių sienelės sudarytos iš baltymų. Gyvūnų ir augalų ląstelių žvyneliai ir blakstienos susideda iš lygiagrečių mikrotubulių. Flagella skiriasi nuo blakstienų tik ilgiu. Taigi žinduolių spermatozoidai turi vieną iki 100 mikronų ilgio žvynelį. Blakstienos yra daugiau nei 10 kartų trumpesnės už žvynelius; vienoje ląstelėje yra keli tūkstančiai blakstienų. Mikrofilamentai yra labai plonos struktūros, sudarytos iš tūkstančių aktino baltymų molekulių, sujungtų viena su kita. Raumenų ląstelėse jie kartu su kitais baltymų gijomis – miozinu patenka į kompleksus, kurie užtikrina šių ląstelių susitraukimo funkciją.
Visų organizmų ląstelių citoplazmoje, šalia branduolio, yra ląstelių centras, dalyvaujantis ląstelių dalijimuisi. Gyvūnų ląstelių ir kai kurių augalų ląstelių centro sudėtis apima centriolę (11, 3 pav.). Centriolė – porų susidarymas. Jame yra du cilindrai, pagaminti iš mikrovamzdelių ir išdėstyti statmenai vienas kitam.
Ląstelių inkliuzai. Galiausiai reikėtų pasakyti apie daugybę intarpų citoplazmoje. Inkliuzai vadinami nenuolatinėmis citoplazmos struktūromis, kurios, skirtingai nei organelės, ląstelės gyvavimo metu arba atsiranda, arba išnyksta. Tankiuose granulių pavidalo inkliuzuose yra rezervinių maistinių medžiagų (krakmolo, baltymų, cukraus, riebalų) arba ląstelių atliekų, kurių dėl vienokių ar kitokių priežasčių negalima iš karto pašalinti. Visi augalų ląstelių plastidai turi savybę sintetinti ir kaupti atsargines maistines medžiagas.
IN augalų ląstelės rezervinių maisto medžiagų kaupimasis vyksta ir vakuolėse – membraniniuose maišeliuose su vandeninis tirpalas druskos ir organiniai junginiai, kurie dažnai užima beveik visą ląstelės tūrį, nustumdami branduolį ir citoplazmą į plazmos membraną.
- Kodėl mitochondrijos vadinamos ląstelės „galia“?
- Kokios ląstelės struktūros prisideda prie jos judėjimo?
- Kas yra ląstelių inkliuzai? Koks jų vaidmuo ląstelėje?
- Apibūdinkite mitochondrijų ir chloroplastų struktūrines ypatybes, susijusias su jų funkcijomis ląstelėje.
Ląstelių centras- optiniu mikroskopu matoma organelė gyvūnų ląstelėse ir žemesni augalai. Paprastai jis yra šalia branduolio arba geometriniame ląstelės centre ir susideda iš dviejų lazdelės formos centriolių kūnų, kurių dydis yra apie 0,3–1 mikrono. Elektroniniu mikroskopu buvo nustatyta, kad centriolė yra cilindras, kurio sieneles sudaro devynios poros labai plonų vamzdelių. Cilindro viduryje yra ertmė, užpildyta vienalyte mase. Centrolių porą supa šviesesnė zona – centrosfera.
Žaidžia ląstelių centras svarbus vaidmuo chromosomų judėjimo metu mitozės metu. Su tuo siejamas kai kurių ląstelių gebėjimas aktyviai judėti. Tai įrodo faktas, kad judriųjų ląstelių (pirmuonių, spermatozoidų) žvynelių arba blakstienų apačioje yra tokios pat struktūros, kaip ir ląstelės centras, dariniai.
Judėjimo organelės. Ląstelės gali judėti specializuotų organelių, tarp kurių yra blakstienos ir žvyneliai, pagalba. Ląstelių blakstienos visada yra daug (pirmuoiuose jų skaičius yra šimtai ir tūkstančiai), o ilgis yra 10–15 mikronų. Vėliavos dažniausiai būna 1-8, jų ilgis 20-50 mikronų. Blakstienos ir žvynelių struktūra tiek augalų, tiek gyvūnų ląstelėse yra panaši. Po elektroniniu mikroskopu buvo nustatyta, kad mikrovamzdeliai driekiasi per visą savo ilgį. Du iš jų yra centre, o aplink juos išilgai periferijos yra dar 9 poros mikrovamzdelių. Visa ši struktūra yra padengta citoplazminė membrana kuri yra ląstelės membranos pratęsimas. Žvynelių ir blakstienų judėjimas užtikrina ne tik ląstelių judėjimą erdvėje, bet ir judėjimą įvairių medžiagų ląstelių paviršiuje, taip pat maisto dalelių patekimas į ląstelę. Blakstienų ir žvynelių apačioje yra baziniai kūnai, kurie taip pat susideda iš mikrotubulių. Manoma, kad baziniai kūnai yra žvynelių ir blakstienų mikrotubulių formavimosi centras. Savo ruožtu baziniai kūnai dažnai kilę iš ląstelės centro.
Nemažai vienaląsčių organizmų ir kai kurių daugialąsčių ląstelių neturi specialių judėjimo organelių ir juda pseudopodijų (pseudopodijų) pagalba. Judėjimas pseudopodijos pagalba vadinamas ameboidiniu judėjimu. Jis pagrįstas specialių baltymų, vadinamų susitraukiančiais baltymais, molekulių judėjimu.
Inkliuzai dažniausiai randami augalų ląstelėse ir skiriasi nuo organelių tuo, kad yra laikini, nenuolatiniai dariniai. Jų skaičius priklauso nuo medžiagų apykaitos intensyvumo ir organizmo būklės. Paprastai jie matomi optiniame mikroskope įvairių dydžių ir formų grūdelių ar lašelių pavidalu. Autorius cheminė sudėtis atskirti: angliavandenių, riebalų ir baltymų intarpus. Angliavandenių ir baltymų intarpai atrodo kaip grūdai, o riebalų – kaip lašeliai. Augalų ląstelėse angliavandeniai dažniausiai nusėda krakmolo grūdelių pavidalu, o gyvūnuose - glikogeno. Baltymų grūdai dideliais kiekiais randami gyvūnų kiaušinių citoplazmoje trynio pavidalu. Daugelis jų yra augalų sėklose. Ląstelėse yra riebalų lašelių jungiamasis audinys gyvūnų ir augalų sėklos. Augalų ląstelėse yra kristalinių intarpų (druskų organinės rūgštys). Tam tikromis sąlygomis ląstelė gali panaudoti visų tipų inkliuzus savo gyvybinės veiklos metu ir vėl kauptis.
Daugelio augalų ląstelių citoplazmoje yra specialių darinių – vakuolių. Tai talpyklos, užpildytos įvairių cheminių medžiagų tirpalų mišiniu. Ypač didelės vakuolės susidaro senose ląstelėse.
|
Kriterijai Palyginimui |
Ribosomų tipai |
|
|
prokariotinis tipas |
eukariotinis tipas |
|
|
Suradimas ląstelėse |
Eubakterijų ir cianobakterijų ribosomos plastidinės ribosomos Mitochondrijų Ribosomos Archebakterijų ribosomos (panašios į eukariotų) |
Citoplazminis eukariotinės ribosomos: Gyvūnai, Ir augalai |
|
Santykis RNR: baltymai | ||
|
rRNR kiekis subvienetas |
1 1500-1600 nukleotidų ilgio molekulė (mažiausiai - 950 mitochondrijų ribosomose) |
1 molekulė, apie 1800 nukleotidų ilgio |
|
rRNR kiekis dideliame subvienetas |
2 molekulės: 1 molekulė apie 3000 nukleotidų ilgio, 1 molekulė apie 120 nukleotidų ilgio |
3 molekulės: 1 molekulė, apie 4-5 tūkst. nukleotidų ilgio, 1 molekulė apie 120 nukleotidų ilgio, 1 molekulė, kurios ilgis yra apie 160 nukleotidų (ši molekulė panaši į vieną iš prokariotinės rRNR sričių) |
|
baltymai (beveik visos molekulės yra unikalios) |
Maždaug 20 molekulių mažame subvienete ir 30 molekulių dideliame |
Maždaug 30 molekulių viename mažas subvienetas ir 40 molekulių didelėje |
Ląstelių centras
Ląstelės centras yra organelė, kuri kontroliuoja mikrotubulių susidarymą ir augimą eukariotinės ląstelės citoskelete.(38 pav.)
Ryžiai. 38. Ląstelių centras 1 - mikrotubulių trynukai; 2 - radialiniai stipinai; 3 - centrinė "krepšelio rato" konstrukcija; 4 - palydovas; 5 - lizosoma; 6 - Golgi komplekso diktiosomos; 7 - besiribojanti pūslelė; 8 - granuliuoto endoplazminio tinklo cisterna; 10 - mitochondrija; 11 - liekamasis korpusas; 12 - mikrovamzdeliai; 13 - karoteca (pagal R. Krstic, su pakeitimais).
Pagrindinės ląstelės centro funkcijos yra citoskeleto mikrotubulių sistemų, judėjimo organelių ir dalijimosi veleno formavimas.
Ląstelės centro pagrindas yra centriolių pora (kartais jų skaičius siekia 4-6-8 poliploidinėse ląstelėse), esančios ląstelės centre.
Centrioles pirmasis atrado ir aprašė V. Flemmingas (1875), tačiau detaliau jie buvo ištirti tik elektroninio mikroskopo pagalba. Centrioliai yra glaudžiai susiję su branduolinis apvalkalas, o daugelyje žemesniųjų eukariotų (pirmuonių, dumblių, grybų, mezokariotų) centrioliai yra įmontuoti į branduolio apvalkalą.
Ląstelių centras visada randamas daugialąsčių gyvūnų ląstelėse. Prokariotams trūksta ląstelių centro ir tubulino. Grybų, dumblių ir vienaląsčių gyvūnų ląstelėse ląstelių centras randamas ne visada, o ląstelėse aukštesni augalai nerasta (su retomis išimtimis). Nesant ląstelinio centro, jo funkcijas eukariotuose atlieka mikrotubulių formavimo centras – CMTC.
Centriolė yra tuščiaviduris cilindras, kurio skersmuo apie 0,15 mikronų ir 0,3-0,5 mikronų ilgio (rečiau - keli mikronai). Centrolių sienelės sudarytos iš 9 trynukai tubulino mikrovamzdeliai, sujungti dyneino tilteliais. Paprastai centrioliai yra išdėstyti poromis: viena centriolė yra pagrindinė centriolė, o kita - dukterinė centriolė; dukterinė centriolė yra statmena pirminei centriolei. Ši centriolių pora diplominis- turi T arba L formos formą. Diplosomą supa tankus citoplazmos sluoksnis centrosfera, arba centrosoma. Ląstelių dalijimosi metu tankios mikrovamzdelių gijos nukrypsta nuo centrosferos ir susidaro astrosfera.
Centriolių padvigubėjimas įvyksta maždaug tarpfazės viduryje: motinos ir dukterinės centriolės išsiskiria, o iš kiekvienos centriolės atsiranda naujas centriolių pumpurai. At skirtingos grupės organizmams, šis procesas turi savo ypatybes.
Be organelių, ląstelėse yra ląstelių inkliuzų. Jų gali būti ne tik citoplazmoje, bet ir kai kuriose organelėse, pavyzdžiui, mitochondrijose ir plastidėse.
Kas yra ląstelių inkliuzai?
Tai dariniai, kurie nėra nuolatiniai. Skirtingai nuo organoidų, jie nėra tokie stabilūs. Be to, jie turi daug paprastesnę struktūrą ir atlieka pasyvias funkcijas, tokias kaip, pavyzdžiui, atsarginė kopija.
Kaip jie statomi?
Dauguma jų yra lašo formos, tačiau kai kurios gali būti ir kitos, pavyzdžiui, panašios į dėmę. Kalbant apie dydį, jis gali skirtis. Ląstelių inkliuzai gali būti arba mažesni už organelius, arba tokio pat dydžio arba net didesni.
Jas daugiausia sudaro viena specifinė medžiaga, dažniausiai organinė. Tai gali būti riebalai arba angliavandeniai arba baltymai. 
klasifikacija
Priklausomai nuo to, iš kur kilusi medžiaga, iš kurios jie sudaryti, yra šie ląstelių intarpų tipai:
- egzogeninis;
- endogeninis;
- virusinis.
Egzogeniniai ląstelių inkliuzai yra sukurti iš cheminiai junginiai kurie pateko į kamerą iš išorės. Tie, kurie susidaro iš pačios ląstelės gaminamų medžiagų, vadinamos endogeninėmis. Virusiniai intarpai, nors juos sintetina pati ląstelė, tačiau tai atsiranda dėl viruso DNR patekimo į ją. Ląstelė tiesiog paima ją savo DNR ir iš jos sintetina viruso baltymą.
Priklausomai nuo ląstelių inkliuzų atliekamų funkcijų, jie skirstomi į pigmentinius, sekrecinius ir trofinius.
Be to, inkliuzai skirstomi į tipus, atsižvelgiant į konkrečius cheminius junginius, iš kurių jie susideda. 
Ląstelių intarpai: funkcijos
Jie gali atlikti tris funkcijas. Apsvarstykite juos lentelėje
Visa tai yra nenuolatinių darinių ląstelėje funkcijos.
Gyvūnų ląstelių inkliuzai
Gyvūno citoplazmoje yra ir trofinių, ir pigmentinių intarpų. Kai kurios ląstelės taip pat turi sekrecinių ląstelių.
Trofinės gyvūnų ląstelėse yra glikogeno įtraukimas. Jie yra maždaug 70 nm dydžio granulės formos. 
Glikogenas yra pagrindinė gyvūno atsarginė medžiaga. Kaip duota medžiaga organizmas kaupia gliukozę. Gliukozės ir gliukogeno apykaitą reguliuoja du hormonai: insulinas ir gliukagonas. Juos abu gamina kasa. Insulinas yra atsakingas už glikogeno susidarymą iš gliukozės, o gliukagonas, priešingai, dalyvauja gliukozės sintezėje.
Dauguma glikogeno inkliuzų randami kepenų ląstelėse. Jų taip pat yra dideliais kiekiais raumenyse, įskaitant širdį. Kepenų ląstelių glikogeno intarpai yra maždaug 70 nm dydžio granulių pavidalo. Jie susirenka į mažas grupes. Glikogeno intarpai miocituose ( raumenų ląstelės) yra apvalios formos. Jie yra pavieniai, šiek tiek didesni už ribosomas.
Taip pat apibūdinamos gyvūnų ląstelės lipidų intarpai. Tai taip pat yra trofiniai inkliuzai, kurių dėka kūnas gali gauti energijos Skubus atvėjis. Jie susideda iš riebalų ir yra ašaros formos. Iš esmės tokie intarpai yra riebalinio jungiamojo audinio ląstelėse - lipocituose. Yra dviejų tipų riebalinis audinys: baltas ir rudas. Baltuosiuose lipocituose yra vienas didelis riebalų lašas, rudose ląstelėse yra daug mažų.
Kalbant apie pigmento intarpus, gyvūnų ląstelėms būdingos tos, kurios susideda iš melanino. Dėl šios medžiagos akies rainelė, oda ir kitos kūno dalys turi tam tikrą spalvą. Kuo daugiau melanino inkliuzų ląstelėse, tuo tamsesnė kas susideda iš šių ląstelių.
Kitas pigmentas, kurio galima rasti gyvūnų ląstelėse, yra lipofuscinas. Ši medžiaga yra geltonai rudos spalvos. Senstant organams, jis kaupiasi širdies raumenyje ir kepenyse.
Augalų ląstelių inkliuzai
Jame taip pat yra ląstelių inkliuzų, kurių struktūrą ir funkcijas mes svarstome
Pagrindiniai trofiniai intarpai šiuose organizmuose yra krakmolo grūdeliai. Savo forma augalai kaupia gliukozę. Paprastai krakmolo inkliuzai yra lęšinės, sferinės arba kiaušinio formos. Jų dydis gali skirtis priklausomai nuo augalo tipo ir organo, kurio ląstelėse jie yra. Jis gali būti nuo 2 iki 100 mikronų.
Lipidų inkliuzai būdingas ir augalų ląstelėms. Jie yra antroji pagal dažnumą trofiniai inkliuzai. Jie turi sferinę formą ir ploną membraną. Jie kartais vadinami sferosomomis.
Baltymų intarpai yra tik augalų ląstelėse, gyvūnams jos nebūdingos. Jie sudaryti iš paprastų baltymų, vadinamų baltymais. Baltymų inkliuzai yra dviejų tipų: aleurono grūdeliai ir baltymų kūnai. Aleurono grūduose gali būti kristalų arba tiesiog amorfinių baltymų. Taigi, pirmasis vadinamas sudėtingu, o antrasis - paprastas. Paprasti aleurono grūdeliai, susidedantys iš amorfinių baltymų, yra mažiau paplitę.
Kalbant apie pigmentų inkliuzus, augalams būdinga plastoglobulės. Juose yra karotinoidų. Tokie inkliuzai būdingi plastidams.
Ląstelių inkliuzai, kurių struktūrą ir funkcijas svarstome, daugiausia susideda iš organinių cheminių junginių, tačiau augalų ląstelėse yra ir tokių, kurie susidaro iš Ne organinės medžiagos. Tai kalcio oksalato kristalai. 
Jų yra tik ląstelės vakuolėse. Šie kristalai gali būti pačios įvairiausios formos ir dažnai būna individualūs tam tikrų tipų augalai.
Gyvi organizmai, susidedantys iš vienos ląstelės, dažnai turi galimybę aktyviai judėti. Evoliucijos procese atsiradę judėjimo mechanizmai yra labai įvairūs.
Judėjimo organelės yra pseudopodijos (sarkoduose ir kai kuriuose žvyneliuose), žvyneliai (žievelėse) arba blakstienėlės (blakstienose).
Be to, judėjimas gali būti atliekamas dėl specialių mikropluoštų - mionemų, esančių po dangteliais citoplazmoje (apikomple), susitraukimo rezultatas.
Ameboidų judėjimas gavo savo pavadinimą nuo paprasčiausio organizmo – amebos. (žr. priedą Nr. 6). Judėjimo organai ameboje yra netikros kojos - pseudopodijos, kurios yra citoplazmos išsikišimai. Jie susidaro skirtingos vietos citoplazmos paviršius. Jie gali išnykti ir vėl atsirasti kitur.
Pseudopodia(iš pseudo... ir graikų pūliai, genties p. podos - pėda) (pseudopods), laikinos citoplazminės ataugos vienaląsčiuose organizmuose (šakniastiebiuose, sporozoanuose ir kt.) ir kai kuriose daugialąsčių organizmų ląstelėse (leukocituose, makrofaguose ir kt.) . Jie skirti judėti ir užfiksuoti maistą ir kitas daleles.
Judėjimas žvynelių pagalba būdingas daugeliui vienaląsčių dumblių (pavyzdžiui, chlamidomonų), pirmuonių (pavyzdžiui, žaliųjų euglenų) ir bakterijų. Šių organizmų judėjimo organai yra žvyneliai – citoplazminės ataugos citoplazmos paviršiuje.
Flagella (Žr. priedą Nr. 7). Dydžiai nuo 2-5 mikronų iki 1 mm. Jie juda vienos ar kelių žvynelių pagalba, kurių ilgis vienam individui gali būti skirtingas. Tik labai kelios rūšys gali sudaryti pseudopodijas. Paprastai jie veisiasi aseksualiai dėl išilginio ląstelių dalijimosi; kai kurie matė lytinis dauginimasis. Didžioji dauguma vėliavėlių yra laisvai gyvenančios rūšys, randamos jūrų ir gėluosiuose vandenyse, taip pat dirvožemyje. Planktoninės formos vaidina svarbų vaidmenį medžiagų cikle; yra daug rūšių biologiniai rodikliai vandens tarša.
Ląstelių struktūra.
Ląstelė, elementari gyvoji sistema, visų gyvūnų ir augalų sandaros ir gyvybės pagrindas. Ląstelės egzistuoja kaip savarankiški organizmai (pavyzdžiui, pirmuonys, bakterijos) ir kaip dalis daugialąsčių organizmų, kuriuose yra lytinės ląstelės, kurios tarnauja dauginimuisi, ir kūno ląstelės (somatinės), skirtingos sandaros ir funkcijų (pavyzdžiui, nervų, kaulų). , raumenys , sekrecijos).
Ląstelių dydis svyruoja nuo 0,1–0,25 mikronų (kai kurios bakterijos) iki 155 mm (stručio kiaušinis su lukštu).
Bet kurio organizmo ląstelė yra pilnavertė gyvoji sistema. Jį sudaro trys neatsiejamai susijusios dalys: membrana, citoplazma ir branduolys. (Žr. priedą Nr. 9).
1. Ląstelių membrana.
Ląstelės membrana tiesiogiai sąveikauja su išorine aplinka ir sąveikauja su kaimyninėmis ląstelėmis (daugialąsčiuose organizmuose).
Ląstelės sienelė turi sudėtinga struktūra. Jį sudaro išorinis sluoksnis ir plazminė membrana, esanti apačioje.
Gyvūnų ir augalų ląstelės skiriasi savo išorinio sluoksnio struktūra (žr. priedą Nr. 10). Augaluose, taip pat bakterijose, melsvadumbliuose ir grybuose, yra ląstelių paviršiuje tankus apvalkalas, arba ląstelės sienelę. Daugumoje augalų jis susideda iš skaidulų. Ląstelės sienelė atlieka nepaprastai svarbų vaidmenį: ji yra išorinis karkasas, apsauginis apvalkalas, suteikia augalų ląstelių turgorą: pro ląstelės sienelę praeina vanduo, druskos, daugelio organinių medžiagų molekulės.
Išorinis gyvūnų ląstelių paviršiaus sluoksnis, skirtingai nei augalų ląstelių sienelės, yra labai plonas ir elastingas. Jis nėra matomas šviesos mikroskopu ir susideda iš įvairių polisacharidų ir baltymų. Paviršinis gyvūnų ląstelių sluoksnis vadinamas glikokaliksas.
Glikokaliksas visų pirma atlieka tiesioginio gyvūnų ląstelių ryšio su išorine aplinka, su visomis ją supančiomis medžiagomis, funkciją. Turėdamas nereikšmingą storį (mažiau nei 1 µm), išorinis gyvūnų ląstelių sluoksnis neatlieka pagalbinio vaidmens, kuris būdingas augalų ląstelių sienelėms. Glikokalikso, taip pat augalų ląstelių sienelių susidarymas vyksta dėl pačių ląstelių gyvybinės veiklos.
2. Citoplazma.
Citoplazma (iš cito... ir plazma), terminą „citoplazma“ pasiūlė E. Strasburgeris (1882).
Atribota nuo išorinė aplinka plazmos membrana, citoplazma yra vidinė pusiau skysta ląstelių aplinka (žr. priedą Nr. 8). citoplazmoje eukariotinės ląstelės išsidėsto branduolys ir įvairios organelės.
Branduolys yra centrinėje citoplazmos dalyje. Jame taip pat yra įvairių inkliuzų – ląstelių veiklos produktų, vakuolių, taip pat mažiausių vamzdelių ir gijų, sudarančių ląstelės skeletą.
Vakuolės (Žr. priedą Nr. 8)(pranc. vakuolė, iš lot. vacuus – tuščia), ertmės gyvūnų ir augalų ląstelėse arba vienaląsčiai organizmai. Yra virškinimo ir susitraukiančios (pulsuojančios) vakuolės, kurios reguliuoja osmosinį slėgį ir padeda pašalinti iš organizmo skilimo produktus. (žr. priedą Nr. 11).
Pagrindinės citoplazmos medžiagos sudėtyje vyrauja baltymai.
Pagrindiniai medžiagų apykaitos procesai vyksta citoplazmoje, ji sujungia branduolį ir visas organeles į vieną visumą, užtikrina jų sąveiką, ląstelės, kaip vientisos vientisos gyvos sistemos, veiklą.
Eksperimentiniu būdu galima gauti gyvų nebranduolinių citoplastų ląstelių, kurios per 1-3 dienas gali susintetinti baltymus, lipidus, ATP. Tada, žinoma, jie miršta dėl to, kad nesant branduolio neįmanoma susintetinti naujų RNR.
Citoplazmos tūris skirtingos ląstelės skiriasi: limfocituose jis yra maždaug lygus branduolio tūriui, o kepenų ląstelėse citoplazma sudaro 94% viso ląstelių tūrio.
Formaliai citoplazmoje išskiriamos trys dalys: organelės, inkliuzai ir hialoplazma.
Organelės- komponentai, būtini bet kuriai ląstelei, be kurių ląstelė negali palaikyti savo egzistavimo.
Vakuolinės sistemos vienos membranos organelės apima endoplazminį tinklą, lizosomas, Golgi aparatą, peroksisomas ir plazminę membraną.
Dviejų membranų organelės yra mitochondrijos ir plastidai. Ši grupė taip pat apima ląstelės branduolys.
bendra nuosavybė membranos organelės yra tai, kad jos yra sudarytos iš lipoproteininių membranų, kurios užsidaro pačios, todėl sudaro uždaras ertmes ir taip padalija citoplazmą į funkciškai skirtingų skyrių grupes.
Ribosomos, gyvūnų ląstelių centrosomos priklauso nemembraninėms organelėms.
Inclusions_ _ ne visada randami ir yra atsarginių medžiagų nuosėdos (glikogenas, trynys) arba medžiagų apykaitos produktų sankaupos (pigmentai, druskos kristalai augaluose).
Hialoplazma(iš "hualine" - skaidrus) - tai yra pagrindinė plazma, citozolis, tiesa vidinė aplinka ląstelės. Hialoplazmos sudėtis yra labai sudėtinga, o konsistencija artėja prie gelio.
Geliai- struktūrinės koloidinės sistemos su skysta dispersine terpe, kuri, veikiama išorinių ar vidinių veiksnių gali pakeisti savo agregacijos būseną ir pereiti į skystesnę fazę – solą. Panašūs gelio-zolio perėjimai gali vykti citoplazmoje veikiant aktino baltymui, o jo būsena pakinta skirtingose ląstelės dalyse, o tai užtikrina visos ląstelės judėjimą. Kai fibrilinis aktinas sąveikauja su fibrino tipo baltymais, gelis stabilizuojamas, o prisijungus prie baltymų, kurių aktyvumas priklauso nuo Ca jonų koncentracijos (pavyzdžiui, gelsolino), visa sistema pereina į skystą būseną.
Ląstelės skeleto elementų, įskaitant jo komponentą aktiną, sunkumas gali labai pasikeisti ląstelės ciklo metu.
Citoplazmos funkcinė reikšmė yra labai didelė.
Hialoplazmoje, be įvairių neorganinių junginių jonų, yra fermentų, dalyvaujančių aminorūgščių, nukleotidų sintezėje, riebalų rūgštys, cukrus. Ribosomose ir poliribosomose, esančiose ant membranų, sintetinami įvairūs baltymai, užtikrinantys ląstelių metabolizmą.
3. Endoplazminis tinklas.
Visa vidinė citoplazmos zona užpildyta daugybe mažų kanalų ir ertmių, kurių sienelės yra membranos, panašios į plazmos membraną. Šie kanalai šakojasi, jungiasi vienas su kitu ir sudaro tinklą, vadinamą endoplazminiu arba endoplazminiu tinklu.
Endoplazminis Tinklelis (žr. priedą Nr. 12) nevienalytės struktūros. Žinomi du jo tipai – granuliuotas (šiurkštus) ir lygus.
Granuliuoto tinklo kanalų ir ertmių membranose yra daug mažų apvalių kūnelių - ribosomų, kurios suteikia membranoms grubią išvaizdą.
Lygiojo endoplazminio tinklo membranos savo paviršiuje neneša ribosomų.
Endoplazminis tinklas atlieka daugybę skirtingų funkcijų. Pagrindinė granuliuoto endoplazminio tinklo funkcija yra dalyvavimas baltymų sintezėje, kuri atliekama ribosomose.
Ant lygaus endoplazminio tinklo membranų sintetinami lipidai ir angliavandeniai. Visi šie sintezės produktai kaupiasi kanaluose ir ertmėse, o vėliau pernešami į įvairias ląstelių organeles, kur sunaudojami arba kaupiasi citoplazmoje kaip ląstelių inkliuzai.
Endoplazminis tinklas jungia pagrindinius ląstelės organelius (žr. priedą Nr. 8).
4. Biologinės membranos.
Biologinės membranos (lot. „membrana“ – oda, plėvelė), plonos (ne daugiau kaip 10 nm storio) lipoproteininės plėvelės, susidedančios iš dvigubo lipidų molekulių sluoksnio, į kurį įeina įvairių baltymų molekulės. Jie yra ląstelių paviršiuje (plazmos membranoje) ir tarpląstelinėse dalelėse (branduoluose, mitochondrijose ir kt.). Pagal svorį, priklausomai nuo membranos tipo, lipidai sudaro 25-60%. o baltymų dalis – 40-75 proc. Daugelyje membranų yra angliavandenių, kurių kiekis gali siekti 2-10%.
Ląstelės sveikata, jos gyvenimo trukmė labai priklauso nuo membranos būklės.
a) Membranų savybės.
Visiškas vandens pralaidumas. Membrana visada leidžia vandenį į ląstelę arba išeina, priklausomai nuo to, kur vandens koncentracija yra didesnė. Toks medžiagos judėjimas iš didelės koncentracijos srities į mažesnės koncentracijos sritį vadinamas difuzija. Medžiagos difuzijai nereikia energijos.
Atrankinis tirpių medžiagų laidumas:
Neigiamo krūvio dalelės greičiau ir lengviau prasiskverbia pro membraną. Riebaluose tirpios medžiagos lengviau prasiskverbia pro membraną nei vandenyje tirpios medžiagos. Mažos molekulės lengviau prasiskverbia pro membraną nei didelės.
b) Aktyvus medžiagų pernešimas.
Kai kurios medžiagos gali prasiskverbti pro membraną priešinga jų difuzijai kryptimi, ty iš žemos vietos į vietą, kurioje yra daugiau didelė koncentracija. Aktyviuoju transportavimu iš ląstelės nuolat pašalinamas natrio, vandenilio ir chloro jonų perteklius. O fosfatai, gliukozė, aminorūgštys, atvirkščiai, aktyviai prasiskverbia į citoplazmą. Aktyvus transportas visada yra susijęs su energijos sąnaudomis.
Ląstelės gyvavimo metu membrana gali būti iš dalies prarasta dėl virškinimo vakuolių (pūslelių) susidarymo. Dėl specialių organelių darbo membrana reguliariai atkuriama. sintetinant membranines vakuoles. Šios vakuolės prilimpa bet kurioje ląstelės membranos vietoje, atkurdamos jos buvusius matmenis ir savybes.
c) Membraniniai lipidai.
Lipidų sudėtis biologinės membranos labai įvairus. būdingi atstovai Ląstelių membranos lipidai yra fosfolipidai, sfingomielinai ir cholesterolis (steroidinis lipidas).
būdingas bruožas membraniniai lipidai yra jų molekulių padalijimas į dvi funkciškai skirtingas dalis: nepolines, be krūvio uodegas, susidedančias iš riebalų rūgščių, ir įkrautas polines galvutes.
Poliarinės galvos neša neigiami krūviai arba gali būti neutralus. Nepolinių uodegų buvimas paaiškina gerą lipidų tirpumą riebaluose ir organiniuose tirpikliuose. Eksperimente, maišant iš membranų išskirtus lipidus su vandeniu, galima gauti apie 7,5 nm storio bimolekulinius sluoksnius arba membranas, kurių periferinės sluoksnio zonos yra hidrofilinės poliarinės galvutės, o centrinė zona – neįkrautos lipidų molekulių uodegos. . Visos natūralios ląstelių membranos turi tą pačią struktūrą.
Ląstelių membranų lipidų sudėtis labai skiriasi. Pavyzdžiui, plazminės membranos gyvūnų ląstelėse gausu cholesterolio (iki 30 proc.) ir mažai lecitino, o mitochondrijų membranose daug fosfolipidų ir mažai cholesterolio.
Lipidų molekulės gali judėti išilgai lipidų sluoksnio, gali suktis aplink savo ašį, taip pat judėti iš sluoksnio į sluoksnį. „Lipidų ežere“ plūduriuojantys baltymai taip pat turi tam tikrą šoninį mobilumą. Lipidų sudėtis abiejose membranos pusėse skiriasi, o tai lemia bilipidinio sluoksnio struktūros asimetriją.
d) Membraniniai baltymai.
Baltymų molekulės, kurios yra daug sunkesnės ir didesnės už lipidų molekules, yra tarsi įsiterpusios į membranos bilipidinį sluoksnį. Dauguma baltymų molekulių yra iš dalies panardintos į membraną ir yra susijusios su lipidais per hidrofobinę sąveiką. Tai yra vadinamieji integraliniai baltymai.
Kiti baltymai yra susiję tik su lipidų molekulių poliniais „galais“ ir yra bilipidinio sluoksnio paviršiuje, o kiti pro membraną prasiskverbia pro ir kiaurai. Asimetrija membranų struktūroje ryškiausiai pasireiškia integralinių baltymų atžvilgiu. Jie yra griežtai orientuoti: jų N-galai žiūri į vakuolių ertmę arba į ekstraląstelinę aplinką.
Membranos labai skiriasi baltymų sudėtimi, kuri iš tikrųjų lemia jų funkcines savybes. Visus membraninius baltymus galima suskirstyti į tris grupes: fermentus, receptorių baltymus ir struktūrinius baltymus.
Skirtingos membranos turi būdingą fermentų rinkinį. Pavyzdžiui, nuo K-Na priklausoma ATPazė, kuri dalyvauja jonų pernešime, yra lokalizuota plazmos membranoje. Receptoriniai baltymai specifiškai jungiasi prie tam tikrų medžiagų ir tarsi jas „atpažįsta“: tai yra hormonų receptorių baltymai, skirti atpažinti gretimų ląstelių paviršių, virusus ir kt.
Struktūriniai baltymai suteikia membranos stiprumą ir yra susiję su įvairiomis baltymų struktūromis citoplazmoje. Pavyzdžiui, epitelio ląstelėse specialūs plazmos membranos baltymai jungiasi prie citoskeleto elementų ir dalyvauja formuojant daugybę tarpląstelinių jungčių, tokių kaip desmosomos, adheziniai kontaktai ir kt.
e) Membraniniai angliavandeniai.
Jie daugiausia susiję su membranomis. glikoproteinai - baltymų molekulės, kovalentiškai susietos su angliavandenių grandinėmis. Paprastai angliavandenių molekulės yra išoriniuose membranų sluoksniuose. Dažniausiai tai yra trumpos linijinės arba šakotos grandinės, kurios apima galaktozę, maniozę, fruktozę, sacharozę, N-acetilgliukozaminą. arabinozės. ksilozė ir kt.
f) Ląstelių membranų sintezė.
Visos ląstelių membranos (išskyrus mitochondrijų ir plastidines membranas) yra pastatytos granuliuotame endoplazminiame tinkle. Nuo jo atsiskiria nedidelės membraninės vakuolės, kurios susilieja su Golgi aparato membranomis. Membraninės pūslelės, kurias savo ruožtu gamina Golgi aparatas, yra naudojamos plazminei membranai, sekrecinėms vakuolėms ir visiems kitiems ląstelės membranos komponentams sukurti.
5. Plazminė membrana.
plazmos membrana (Žr. priedą Nr. 8)(plazmalema, ląstelės membrana), po glikokaliksu ir ląstelių sienelės yra plazminė membrana, kuri supa augalų ir gyvūnų ląstelių protoplazmą. Plazminės membranos storis apie 10 nm, jos sandaros ir funkcijų tyrimas įmanomas tik elektroninio mikroskopo pagalba.
Tarnauja ne tik kaip mechaninis barjeras, bet, svarbiausia, riboja laisvą dvipusį mažos ir didelės molekulinės masės medžiagų patekimą į ląstelę ir iš jos. Be to, plazmolema veikia kaip struktūra, kuri „atpažįsta“ įvairias cheminių medžiagų ir reguliuojant selektyvų šių medžiagų transportavimą į ląstelę. Kaip ir kitos ląstelių membranos, jis atsiranda ir atnaujinamas dėl sintetinė veikla endoplazminis tinklas ir turi panašią į juos struktūrą.
a) Plazlemos barjerinis transportavimo vaidmuo.
Plazminės membranos mechaninį stabilumą lemia ne tik pačios membranos, bet ir šalia esančios glikokalikso bei citoplazmos žievės sluoksnio savybės.
Išorinis plazminės membranos paviršius (žr. priedą Nr. 13). padengtas biriu pluoštiniu 3-4 nm storio medžiagos sluoksniu – glikokaliksu. Jį sudaro išsišakojusios membranos vientisų baltymų polisacharidinės grandinės, tarp kurių gali išsidėstyti ląstelės išskiriami glikolipidai ir proteoglikanai. Nedelsiant aptinkami kai kurie ląsteliniai hidroliziniai fermentai, dalyvaujantys tarpląsteliniame medžiagų skaidyme (tarpląsteliniame virškinime, pavyzdžiui, žarnyno epitelyje).
0,1-0,5 µm storio citoplazmos žievės sluoksnyje nėra ribosomų ir membraninių struktūrų, tačiau jame gausu aktino mikrofilamentų.
Plazminė membrana, kaip ir kitos lipoproteininės ląstelių membranos, yra pusiau laidi. Didžiausia prasiskverbimo galia pasižymi vanduo ir jame ištirpusios dujos. Jonų pernešimas gali vykti pasyviai, nenaudojant energijos. Šiuo atveju kai kurie membranos transportavimo baltymai sudaro molekulinius kompleksus, kanalus, kuriais jonai praeina pro membraną paprastos difuzijos būdu.
Kitais atvejais specialūs membraniniai nešikliai selektyviai jungiasi prie vieno ar kito jono ir perneša jį per membraną. Šis pernešimo būdas vadinamas aktyviuoju transportavimu ir atliekamas naudojant baltymų jonų siurblius. Pavyzdžiui, išleisdama 1 ATP molekulę, K-Na siurblio sistema per vieną ciklą iš ląstelės išpumpuoja 3 Na jonus ir pumpuoja 2 K jonus prieš koncentracijos gradientą.
Kartu su aktyviu jonų transportavimu per plazmalemą prasiskverbia įvairūs cukrūs, nukleotidai ir aminorūgštys. Makromolekulės, tokios kaip baltymai, neprasiskverbia pro membraną. Jos, kaip ir didesnės medžiagos dalelės, pernešamos į ląstelę endocitozė . Endocitozės metu tam tikra plazmalemos dalis užfiksuoja, apgaubia ekstraląstelinę medžiagą ir uždaro ją į membranos vakuolę. Ši vakuolė yra endosoma - susilieja citoplazmoje su pirmine lizosoma ir įvyksta sugautos medžiagos virškinimas.
Endocitozė formaliai skirstoma į fagocitozę (stambių dalelių absorbciją ląstelėje) ir pinocitozę (tirpų absorbciją).
Plazminė membrana taip pat dalyvauja pašalinant medžiagas iš ląstelės egzocitozė - atvirkštinis endocitozės procesas.
b) Plazlemos receptorių vaidmuo.
Ląstelės išorinės membranos baltymai nešikliai taip pat yra receptoriai, atpažįstantys tam tikrus jonus ir su jais sąveikaujantys. Membraniniai baltymai arba glikokalikso elementai gali veikti kaip receptoriai ląstelės paviršiuje. Tokios vietos, jautrios atskiroms medžiagoms, yra išsklaidytos po ląstelės paviršių arba surenkamos mažose zonose.
Daugelio ląstelių receptorių vaidmuo yra ne tik surišti specifines medžiagas, bet ir perduoti signalus iš paviršiaus į ląstelę. Pavyzdžiui, hormonui veikiant ląstelę, įvykių grandinė klostosi taip: hormono molekulė specifiškai sąveikauja su plazmos membranos receptoriaus baltymu ir, neįsiskverbdama į ląstelę, aktyvuoja fermentą, sintetinantį ciklinį AMP. Pastarasis aktyvina arba slopina tarpląstelinį fermentą arba fermentų grupę.
Ląstelės paviršiuje esančių receptorių rinkinių įvairovė ir specifiškumas lemia labai sudėtinga sistemažymenys, leidžiantys ląstelėms atskirti „savuosius“ (to paties individo ar tos pačios rūšies) nuo „svetimų“.
c) tarpląsteliniai ryšiai.
Daugialąsčiuose organizmuose dėl tarpląstelinės sąveikos susidaro sudėtingi ląstelių ansambliai. Glikokaliksas, esantis arti ląstelių viena kitos, užtikrina ląstelių sukibimą, nes jame yra transmembraninių glikoproteinų kadherinų. Tai paprastas tarpląstelinis kontaktas, kuriame tarpas tarp ląstelių yra 10-20 nm.
Epitelyje dažnai yra tanki arba blokuojanti jungtis, kurioje abiejų plazmos membranų išoriniai sluoksniai yra kuo arčiau, o jų sąlyčio taškuose yra vientisų membranų baltymų rutuliukai. Toks kontaktas yra nepralaidus molekulėms ir jonams, uždaro tarpląstelines ertmes.
Inkaro jungtys, arba kontaktai, ne tik jungia gretimų ląstelių plazmines membranas, bet ir jungiasi su fibriliniais citoskeleto elementais. Pavyzdžiui, desmosomoms, kurios atrodo kaip plokštelės ar mygtukai, tarpląstelinei erdvei būdingas tankus desmogleino glikoproteinų sluoksnis.
Citoplazminėje pusėje desmoplakino baltymo sluoksnis, susietas su citoskeleto tarpinėmis gijomis, yra greta plazmalemos.
Tarpų sandūros laikomos ląstelių komunikacijos jungtimis. Tarpo kontakto zonoje gali būti nuo 20-30 iki kelių tūkstančių jungtys - cilindrinės baltymų struktūros su vidinis kanalas 2 nm skersmens. Kiekvienas konneksonas sudarytas iš 6 jungiamųjų baltymų subvienetų. Konneksonai atlieka tiesioginių tarpląstelinių kanalų, per kuriuos jonai ir mažos molekulinės masės medžiagos gali sklisti iš ląstelės į ląstelę, vaidmenį.
6. Lizosomos.
Lizosomos (iš liz ir graikų soma – kūnas). Jie yra maži apvalūs kūnai. Kiekviena lizosoma nuo citoplazmos yra atskirta membrana. Lizosomos viduje yra fermentų, kurie gali skaidyti (t.y. lizuoti – iš čia ir pavadinimas) baltymus, polisacharidus, peptidus, nukleino rūgštis. (žr. priedą Nr. 8).
Tai labai marga 0,1–0,4 µm dydžio pūslelių klasė, kurią riboja viena membrana (apie 7 nm storio), kurių viduje yra nevienalytis turinys. Jie susidaro veikiant endoplazminiam tinklui ir Golgi aparatui ir šiuo požiūriu primena sekrecines vakuoles.
Pagrindinis jų vaidmuo yra dalyvavimas tiek egzogeninių, tiek endogeninių biologinių makromolekulių intraląstelinio skilimo procesuose. būdingas bruožas lizosomos yra tai, kad jose yra apie 40 hidrolizinių fermentų: proteinazių, nukleazių, fosfatazių, glikozidazių ir kt., kurių optimalus yra 5 pH.
Lizosomose rūgštinė aplinkos vertė susidaro dėl to, kad jų membranose yra protonų „siurblys“, kuris sunaudoja ATP energiją.
Be to, į lizosomų membranas yra įmontuoti baltymai-nešikliai, kad jie galėtų transportuoti iš lizosomos į hidrolizės produktų citoplazmą: suskaidytų molekulių monomerai - aminorūgštys, cukrūs, nukleotidai, lipidai. Kad nesusivirškintų, lizosomų membraniniai elementai yra apsaugoti oligosacharidais, kurie neleidžia hidrolazėms su jais sąveikauti.
Tarp skirtingos morfologijos lizosomų dalelių išskiriami keturi tipai: pirminės ir antrinės lizosomos, autofagosomos ir liekamieji kūnai.
pirminės lizosomos.
Paprastai tai yra mažos membraninės pūslelės, kurių skersmuo yra apie 100 nm, su bestruktūriu turiniu, kuriame yra aktyvios rūgštinės fosfatazės, lizosomų žymeklio. Įrodyta, kad lizosomų fermentai, kaip įprasta, sintetinami granuliuotame tinkle, o Golgi aparate yra supakuoti į membranines pūsleles. Visas pirminių lizosomų susidarymo būdas labai panašus į sekrecinių granulių susidarymą, pavyzdžiui, kasos ląstelėse.
antrinės lizosomos.
Fagocitinės arba pinocitinės vakuolės susilieja su pirminėmis lizosomomis ir sudaro antrines lizosomas. Prasideda ląstelės absorbuotų substratų skaidymo procesas, veikiant hidrolazėms, esančioms pirminėje lizosomoje. Biogeninės medžiagos suskaidomos iki monomerų, kurie per lizosomų membraną pernešami į citoplazmą, kur vėl panaudojamos, įtraukiamos į įvairias sintetines ir medžiagų apykaitos procesai. Ląstelių lizosomų dydžio ir struktūros įvairovė pirmiausia siejama su antrinių lizosomų – endocitinių vakuolių susiliejimo su pirminėmis lizosomomis produktais – įvairove.
Liekamieji kūnai.
Biogeninių makromolekulių skilimas lizosomų viduje gali nesibaigti. Tokiu atveju lizosomos ertmėje kaupiasi nesuvirškinti produktai, o antrinė lizosoma tampa likutiniu kūnu (telolisosoma). Jų turinys sutankinamas ir pertvarkomas. Dažnai liekamuosiuose kūnuose yra antrinė nesuvirškintų lipidų struktūra, sudaranti sudėtingas sluoksnines struktūras. Ten taip pat nusėda pigmentai.
Autolizosomos.
Pagal morfologiją autolizosomos (autofagosomos) priskiriamos antrinėms lizosomoms, tačiau šių dalelių viduje yra fragmentų ar net ištisų citoplazminių struktūrų, tokių kaip mitochondrijos, plastidės, ribosomos, tinklinio audinio elementai ir kt. Daroma prielaida, kad autofagocitozės procesas yra susijęs su pakitusių, „sulaužytų“ ląstelių komponentų atranka ir sunaikinimu.
7. Golgi aparatas.
K. Golgi vardu pavadintas Golgi aparatas (Golgi kompleksas) – ląstelės organoidas.
Daugelyje gyvūnų ląstelių, pavyzdžiui, nervų ląstelėse, jis yra sudėtingo tinklo, esančio aplink branduolį, forma. Augalų ir pirmuonių ląstelėse Golgi aparatą vaizduoja atskiri pjautuvo arba lazdelės formos kūnai. Šio organoido struktūra yra panaši augalų ir gyvūnų organizmų ląstelėse, nepaisant jo formos įvairovės. (žr. priedą Nr. 8).
Golgi aparato sudėtis apima: ertmes, apribotas membranomis ir išdėstytas grupėmis (po 5-10); dideli ir maži burbuliukai, esantys ertmių galuose. Visi šie elementai sudaro vieną kompleksą.
Golgi aparatas atlieka daug svarbių funkcijų.
Atneša baltymus į galutinę darbinę formą, pavyzdžiui, kai kuriuos baltymus „susiuva“ į didelius baltymų kompleksai, prie kai kurių baltymų prijungia reikiamus metalo jonus.
Sudaro membranines pūsleles, kurios, palikdamos Golgi kompleksą, arba atkuria ląstelės membraną, arba virsta lizosomomis. Lizosomos yra ląstelės membraninės organelės, atstovaujančios mikroskopines pūsleles, užpildytas virškinimo fermentais.
Jie atlieka virškinimo ir apsaugines funkcijas. Pavyzdžiui, žinduolių kasos ląstelėse, virškinimo fermentai kurios kaupiasi organoido ertmėse.
Jie gali sulipti su virškinimo vakuole, supildami į ją virškinimo fermentus. Ląstelei kontaktuojant su svetima medžiaga arba su svetima ląstele, lizosomos sulimpa su ląstelės membrana išleisdami savo fermentus už ląstelės ribų. Lizosomų fermentai taip pat gali dalyvauti „užprogramuotoje savo ląstelių mirtyje“.
Dėl Golgi aparato veiklos atsinaujina ir auga plazmos membrana.
8. Ribosomos.
Ribosomos randamos visų organizmų ląstelėse. Tai mikroskopiniai suapvalintos formos kūnai, kurių skersmuo 15-20 nm. Kiekviena ribosoma susideda iš dviejų nevienodo dydžio dalelių – mažų ir didelių. (žr. priedą Nr. 14).
Vienoje ląstelėje yra daug tūkstančių ribosomų, jos yra arba ant granuliuoto endoplazminio tinklo membranų, arba laisvai guli citoplazmoje.
Ribosomos sudarytos iš baltymų ir RNR. Ribosomų funkcija yra baltymų sintezė. Baltymų sintezė - sunkus procesas, kurią vykdo ne viena ribosoma, o visa grupė, įskaitant iki kelių dešimčių kombinuotų ribosomų. Ši ribosomų grupė vadinama polisomas .
Susintetinti baltymai pirmiausia kaupiami endoplazminio tinklo kanaluose ir ertmėse, o vėliau transportuojami į organelius ir ląstelių vietas, kur jie suvartojami.
Endoplazminis tinklas ir ribosomos (Žr. priedą Nr. 8), esantis ant jo membranų, yra vienas baltymų biosintezės ir transportavimo aparatas.
9. Mitochondrijos.
Membranos savaime besidalijančios organelės. Daugumos gyvūnų ir augalų ląstelių citoplazmoje yra nedideli kūnai (0,2-7 mikronai) – mitochondrijos (gr. „mitos“ – siūlas, „chondrion“ – grūdas, granulė).
Mitochondrijos yra aiškiai matomos šviesos mikroskopu, su kuriuo galite pamatyti jų formą, vietą, suskaičiuoti skaičių (žr. priedą Nr. 8). Vidinė mitochondrijų struktūra buvo tiriama naudojant elektroninį mikroskopą.
Mitochondrijų apvalkalas susideda iš dviejų membranų – išorinės ir vidinės. Išorinė plėvelė lygi, nesudaro jokių raukšlių ir ataugų. Priešingai, vidinė membrana sudaro daugybę raukšlių, nukreiptų į mitochondrijų ertmę. Vidinės membranos raukšlės vadinamos cristae (lot. „crista“ - šukos, atauga) Juose vyksta pieno rūgšties oksidacijos procesas, dėl kurio išsiskiria energija, kaupiama ATP pavidalu.
Kritų skaičius skirtingų ląstelių mitochondrijose nėra vienodas. Jų gali būti nuo kelių dešimčių iki kelių šimtų, o ypač daug kristalų yra aktyviai veikiančių ląstelių mitochondrijose, pavyzdžiui, raumenų ląstelėse.
Mitochondrijos vadinamos ląstelių „elektrinėmis“, nes jų pagrindinė funkcija yra adenozino trifosfato (ATP) sintezė. Ši rūgštis sintetinama visų organizmų ląstelių mitochondrijose ir yra universalus energijos šaltinis, būtinas ląstelės ir viso organizmo gyvybiniams procesams įgyvendinti.
Mitochondrijos, kaip ir bakterijos, dauginasi tiesioginio dalijimosi būdu.
10. Ląstelės centras.
Ląstelėse šalia branduolio yra organelė, vadinama ląstelės centru. Pagrindinę ląstelės centro dalį sudaro du maži kūnai (kartais daugiau) - centrioliai esantis nedideliame suspaustos citoplazmos plote. Kiekviena centriolė turi iki 1 µm ilgio cilindro formą, kurios sieneles sudaro mikrovamzdeliai, išsidėstę išilgai cilindro ašies. Centriolėse yra baltymų ir didelis skaičius RNR. Ląstelėje yra dvi poros centriolių. Kiekvienoje centriolių poroje yra statmenos viena kitai.
Centrioliai vaidina svarbų vaidmenį ląstelių dalijimuisi. Jie sudaro ilgus baltymų siūlus, vadinamąjį dalijimosi verpstę. Ląstelių dalijimosi metu centrioliai nukrypsta link jo polių, o tai lemia dalijimosi veleno orientaciją.
11. Plastidai.
Plastidės yra membraninės savaime besidalančios ląstelių organelės. Skirtingai nuo visų aukščiau aptartų organelių, plastidai randami tik augalų ląstelėse.
Savo struktūra jie primena mitochondrijas: juos sudaro dvi membranos, išorinė lygi ir vidinė, sudarančios daugybę plokščių ataugų - tilakoidai . Visi tilakoidai yra sukrauti kaip monetų krūvos. Kiekvienas kaminas vadinamas briaunotas . Tarp granos yra vidinis plastido skystis, vadinamas stroma . Jame yra savo DNR, kurios struktūra panaši į bakterijų.
Plastidės dauginasi kaip bakterijos tiesioginio dalijimosi būdu.
Pagal konstrukcines ypatybes išskiriami trys pagrindiniai plastidžių tipai (Žr. priedą Nr. 15): žalia - chloroplastai; raudona, oranžinė ir geltona - chromoplastai; bespalviai – leukoplastai.
a) chloroplastas.
Šių organelių yra lapų ir kitų žalių augalų organų ląstelėse (ūgliuose, nesubrendusiuose vaisiuose), taip pat įvairiuose dumbliuose.
Chloroplastų dydis yra 4-6 mikronai, dažniausiai jie turi ovalo formos. Aukštesniuose augaluose viena ląstelė paprastai turi kelias dešimtis chloroplastų.
Žalia spalva chloroplastai priklauso nuo juose esančio chlorofilo pigmento kiekio.
Chloroplastas yra pagrindinė augalų ląstelių organelė, kurioje vyksta fotosintezė, t.y. organinių medžiagų (angliavandenių) susidarymas iš neorganinių medžiagų (CO2 ir H20), naudojant saulės šviesos energiją.
Chloroplastai yra struktūriškai panašūs į mitochondrijas. (žr. priedą Nr. 16). Chloroplastas nuo citoplazmos yra atskirtas dviem membranomis – išorine ir vidine. Išorinė membrana yra lygi, be raukšlių ir ataugų, o vidinė sudaro daug sulenktų ataugų, nukreiptų į chloroplasto vidų. Todėl chloroplasto viduje yra sutelkta daug membranų, kurios sudaro specialias struktūras - graną. Chlorofilo molekulės yra granulių membranose, nes čia vyksta fotosintezė.
ATP taip pat sintetinamas chloroplastuose. Tarp vidinių chloroplasto membranų yra DNR, RNR ir ribosomos. Vadinasi, chloroplastuose, taip pat ir mitochondrijose, vyksta šių organelių veiklai reikalingo baltymo sintezė. Chloroplastai dauginasi dalijantis (žr. priedą Nr. 17).
b) Chromoplastai.
Chromoplastai yra plastidai, kurių sudėtyje yra į riebalus panašių pigmentų, kurie nuspalvina ląstelę geltona, oranžine ir raudona. Jų yra prinokusiuose augalų vaisiuose, suteikiant jiems atitinkamą spalvą, taip pat rudenį lapuočių medžių lapuose.
c) Leukoplastai.
Leukoplastai yra bespalviai plastidai. Ant vidinės membranos juose nėra jokių pigmentų. Ląstelėje jie yra atsakingi už polisacharidų (krakmolo) sintezę ir kaupimąsi. Daug jų randama požeminių ūglių (bulvių gumbų, topinambų) ląstelėse, taip pat vaisiuose ir sėklose.
d) Plastidinės transformacijos.
Chloroplastai gali lengvai išsigimti į kitų tipų plastidus. Tai stebime, kai rudenį pagelsta ir parausta bręstantys vaisiai ar lapai. Tamsoje chloroplastai gali pakeisti spalvą ir virsti leukoplastais. Tačiau šie procesai yra negrįžtami: leukoplastai ir chromoplastai niekada nevirsta chloroplastais.
Branduolys (ląstelės branduolys) privalomoji dalis daugelio vienaląsčių ir visų daugialąsčių organizmų ląstelės. Dydžiai nuo 1 mikrono (kai kuriuose pirmuoniuose) iki 1 mm (kai kurių žuvų ir varliagyvių kiaušiniuose). Terminą „branduolys“ (lot. Nucleus) pirmą kartą pavartojo R. Brownas 1833 m., kai aprašė augalų ląstelėse pastebėtas sferines struktūras. (žr. priedą Nr. 8).
a) branduolinis apvalkalas.
Ląstelės branduolio vidinę erdvę nuo citoplazmos skiria branduolinė membrana, susidedanti iš dviejų membranų. Branduolinės membranos membranos savo struktūra panašios į kitus ląstelės membraninius komponentus ir yra sukonstruotos pagal tą patį principą: tai plonos lipoproteininės plėvelės, susidedančios iš dvigubo lipidų molekulių sluoksnio, į kurį įterptos baltymų molekulės.
Tarpas tarp vidinės ir išorinės branduolinės membranos vadinamas perinuklearinis.
Daugybė ribosomų dažniausiai būna išorinės branduolinės membranos paviršiuje, kartais galima stebėti tiesioginį šios membranos perėjimą į ląstelės granuliuoto endoplazminio tinklo kanalų sistemą.
Vidinė branduolio membrana yra surišta plonu pluoštiniu baltymų sluoksniu - branduolinė lamina sudarytas iš laminų baltymų. Tankus branduolio sluoksnio fibrilių tinklas gali užtikrinti branduolio vientisumą, net ir eksperimente ištirpus branduolio apvalkalo lipidinėms membranoms. SU viduje chromatino kilpos, užpildančios branduolį, yra pritvirtintos prie plokštelės.
Branduolinė membrana turi apie 90 nm skersmens skylutes, susidariusias susiliejus išorinei ir vidinei branduolių membranoms. Tokios skylės branduolio apvalkale yra apsuptos sudėtingų baltymų struktūrų, vadinamų branduolinių porų kompleksu.
Aštuoni baltymų subvienetai, sudarantys branduolio poras, yra aplink branduolio apvalkalo perforaciją žiedų pavidalu, kurių skersmuo apie 120 nm, stebimas elektroniniu mikroskopu abiejose branduolio apvalkalo pusėse. Porų komplekso baltymų subvienetai turi ataugas, nukreiptas į poros centrą, kur kartais matoma 10–40 nm skersmens „centrinė granulė“.
Branduolinių porų dydis ir jų struktūra yra standartiniai visoms eukariotų ląstelėms. Branduolinių porų skaičius priklauso nuo ląstelių metabolinio aktyvumo: kuo aukštesnis sintetinių procesų lygis ląstelėje, tuo daugiau porų tenka ląstelės branduolio paviršiaus vienetui.
Branduolinio citoplazminio transportavimo procese branduolio poros veikia kaip tam tikras molekulinis sietas, pasyviai praleidžiančios jonus ir mažas molekules (cukrus, nukleotidus, ATP ir kt.) išilgai koncentracijos gradiento ir vykdančios aktyvų selektyvų didelių molekulių transportavimą. baltymų ir ribonukleoproteinų, tai yra, ribonukleino rūgšties kompleksai (RNR) su baltymais. Taigi, pavyzdžiui, baltymai, pernešami į branduolį iš citoplazmos, kur yra sintetinami, turi turėti tam tikras apie 50 aminorūgščių sekas (vadinamosios NLS sekos), „atpažįstamos“ branduolinių porų komplekso. Šiuo atveju branduolinių porų kompleksas, naudodamas energiją ATP pavidalu, aktyviai perkelia baltymą iš citoplazmos į branduolį.
b) Chromatinas.
Ląstelės branduolys yra beveik visos ląstelės genetinės informacijos talpykla, todėl pagrindinis ląstelės branduolio turinys yra chromatinas: dezoksiribonukleino rūgšties (DNR) ir įvairių baltymų kompleksas.
Branduolyje ir ypač mitozinėse chromosomose chromatino DNR pakartotinai sulankstoma, supakuota specialiu būdu, kad aukštas laipsnis tankinimas. Juk visos ilgos DNR gijos, kurių bendras ilgis, pavyzdžiui, pas žmogų yra apie 164 cm, turi būti patalpintos į ląstelės branduolį, kurio skersmuo siekia vos kelis mikrometrus. Ši užduotis išsprendžiama nuosekliai pakuojant DNR į chromatiną specialių baltymų pagalba.
Didžioji chromatino baltymų dalis yra histono baltymai, kurie yra rutulinių chromatino subvienetų, vadinamų, dalis. nukleosomos . Iš viso yra 5 histono baltymų tipai.
Nukleosoma yra cilindro formos dalelė, susidedanti iš 8 histonų molekulių, kurių skersmuo yra apie 10 nm, ant kurių „suvyniota“ šiek tiek mažiau nei du DNR molekulės apsisukimai. IN elektroninis mikroskopas toks dirbtinai dekondensuotas chromatinas atrodo kaip „karoliukai ant virvelės“.
Gyvame branduolyje nukleozomų ląstelės yra glaudžiai sujungtos viena su kita, padedant kitam jungikliui histono baltymui, sudarydamos vadinamąją elementariąją chromatino fibrilę, kurios skersmuo 30 nm. Kiti nehistoniniai baltymai, sudarantys chromatiną, užtikrina tolesnį sutankinimą. y., stacking, fibril chromatin, kuris pasiekia savo didžiausios vertės ląstelių dalijimosi metu mitozinėse arba mejotinėse chromosomose.
Ląstelės branduolyje chromatinas yra tiek tankaus kondensuoto chromatino pavidalu, kuriame tankiai supakuotos 30 nm elementarios fibrilės, tiek homogeninio difuzinio chromatino pavidalu. Kiekybinis šių dviejų tipų chromatino santykis priklauso nuo ląstelės metabolinio aktyvumo pobūdžio, jos diferenciacijos laipsnio. Pavyzdžiui, paukščių eritrocitų branduoliuose, kuriuose nevyksta aktyvūs replikacijos ir transkripcijos procesai, yra praktiškai tik tankus kondensuotas chromatinas.
Dalis chromatino išlaiko kompaktišką, kondensuotą būseną viso ląstelės ciklo metu – šis chromatinas vadinamas heterochromatinas ir nuo euchromatino skiriasi daugybe savybių.
c) Replikacija ir transkripcija.
Eukariotinėse ląstelėse paprastai yra keletas chromosomų (nuo dviejų iki kelių šimtų), kurios praranda savo kompaktišką formą branduolyje (tarpfazėje, tai yra tarp mitozinių padalijimų), atsipalaiduoja ir užpildo branduolio tūrį chromatino pavidalu. Nepaisant dekondensuotos būsenos, kiekviena chromosoma branduolyje užima griežtai apibrėžtą vietą ir yra sujungta su branduolio apvalkalu sluoksniu. griežtai pritvirtintas vidinis paviršius branduolio apvalkalai, tokios chromosomų struktūros kaip centromerai ir telomerai.
Tam tikrame ląstelės gyvavimo ciklo etape, sintetiniame periode, vyksta replikacija, t.y., visa branduolio DNR padvigubėja, o chromatinas tampa dvigubai didesnis. Šiam procesui reikalingi baltymai, žinoma, patenka iš citoplazmos per branduolio poras. Taigi ląstelė ruošiasi būsimam ląstelių dalijimuisi – mitozei, kai viso DNR branduolyje grįš į pradinį lygį.
Genetinės informacijos, esančios DNR genų pavidalu, įgyvendinimas prasideda nuo transkripcijos, t.y., nuo pasiuntinio RNR (i-RNR) sintezės - tikslių genų kopijų, pagal kurias baltymai vėliau bus pastatyti citoplazmoje ant ribosomų.
Šis procesas vyksta įvairiuose branduolio tūrio taškuose, kurie morfologiškai nesiskiria nuo aplinkinio chromatino. Dažniausiai galima stebėti difuzinę transkripciją, t.y. dekondensuotas chromatinas.
Be chromatino, sudarančio chromosomas, eukariotų branduoliuose paprastai yra vienas ar daugiau branduolių.
d) Branduolinio baltymo matrica.
Norint atlikti replikacijos, transkripcijos procesus, taip pat išlaikyti tam tikrą chromosomų padėtį branduolio tūryje, yra rėmo baltymų struktūros, vadinamos branduolinio baltymo matrica. Tokią matricą sudaro mažiausiai trys morfologiniai komponentai:
periferinis pluoštinis sluoksnis – sluoksniai;
vidinė arba tarpchromatinė branduolio matrica
o branduolio matrica.
Stebėjimai rodo, kad branduolinės matricos komponentai nėra standžios sustingusios struktūros, jos yra dinamiškos ir gali labai keistis priklausomai nuo funkcinės savybės branduoliai. Buvo įrodyta, kad baltymų matrica turi daug taškų, stipriai jungiasi su pagrindine DNR, kuri savo ruožtu turi tam reikalingas specialias nukleotidų sekas.
13. Branduoliai.
a) Branduolių skaičius ląstelėje.
Pradedant žaliais dumbliais, grybais ir žemesniaisiais pirmuoniais ir baigiant aukštesni organizmai, visos ląstelės turi privalomas intranuklearines struktūras – branduolius (žr. priedą Nr. 18).Ši taisyklė turi daugybę išimčių, kurios tik pabrėžia branduolio svarbą ir būtinumą gyvenimo ciklas ląstelės. Tokios išimtys apima skylančių kiaušinėlių ląsteles, kuriose ankstyvosiose embriogenezės stadijose branduolių nėra, arba ląstelės, kurios baigė vystytis ir negrįžtamai specializuotos, pavyzdžiui, kai kurios kraujo ląstelės.
Branduolių skaičius ląstelėje gali būti įvairus, tačiau jų skaičius viename branduolyje priklauso nuo ląstelės genų balanso. Nustatyta, kad dalyvauja branduolių susidarymas tam tikros vietos kai kurios chromosomos, kurių ryšį su branduoliu galima gerai atsekti telofazėje ir profazėje. Tokios chromosomos, kaip taisyklė, turi antrinius susiaurėjimus, kurių zonos yra vietos, kuriose vyksta branduolių vystymasis. McClintock (1934) pavadino šias chromosomų dalis „branduoliniai organizatoriai“.
Antrinių susiaurėjimų vietos ypač būdingos branduolio organizatorių išsidėstymui, tačiau pastarieji kartais gali būti chromosomų galuose arba keliose vietose išilgai chromosomos ilgio.
Iš viso branduolio branduolyje yra nustatomas pagal branduolio organizatorių skaičių ir didėja priklausomai nuo branduolio ploidiškumo. Tačiau dažnai branduolių skaičius viename branduolyje yra mažesnis už skaičių branduolių organizatoriai. Įrodyta, kad branduoliai gali susilieti; be to, formuojant vieną branduolį kartais dalyvauja keli organizatoriai.
Net M. S. Navashino (1934) darbuose buvo parodyta, kad chromosomų lokusas, kuris m. normaliomis sąlygomis suformuoja didelį branduolį, tampa neaktyvus, kai po hibridizacijos kitos chromosomos branduolyje atsiranda „stipresnis“ lokusas. Tai, kad tam tikromis sąlygomis kai kurių branduolių organizatorių aktyvumas gali būti slopinamas arba kitų, anksčiau buvusių latentinėje, paslėptoje būsenoje, aktyvumas gali padidėti, rodo, kad ląstelėse palaikomas tam tikras branduolinės medžiagos kiekio balansas. arba, kitaip tariant, branduolių gaminami „bendrieji“ produktai.
Remiantis aukščiau pateiktais faktais, galima padaryti tokias išvadas:
Þ nukleolių susidarymas ir jų skaičius yra susijęs su tam tikrų chromosomų sekcijų – branduolio organizatorių, kurie yra išsidėstę, veikla. didžiąja dalimi antrinių susiaurėjimų zonose;
Þ branduolių skaičiaus pokyčiai ląstelėse šio tipo gali atsirasti dėl branduolių susiliejimo arba dėl ląstelės chromosomų pusiausvyros poslinkių.
b) Branduolio fiziologija ir chemija.
Branduolys, palyginti su kitais ląstelės komponentais, apibūdinamas kaip tankiausia struktūra, turinti didžiausią RNR koncentraciją. didelis aktyvumas RNR sintezei.
RNR koncentracija branduoliuose visada yra didesnė už RNR koncentraciją kituose ląstelės komponentuose, todėl RNR koncentracija branduolyje gali būti 2-8 kartus didesnė nei branduolyje ir 1-3 kartus didesnė nei citoplazma. RNR koncentracijos santykis pelių kepenų ląstelių branduolyje, branduolyje ir citoplazmoje yra 1:7, 3:4, 1, kasos ląstelėse - 1:9, 6:6, 6.
Branduolyje DNR nerandama, bet nepaisant to, tiriant fiksuotas ląsteles, aplink branduolį visada išskiriama chromatino zona. Šis perinukleolinis chromatinas, remiantis elektronine mikroskopija, yra neatskiriama sudėtinga struktūra branduolys.
Branduolys yra viena iš aktyviausių ląstelių vietų, skirtų pirmtakams įtraukti į RNR. Nukleolinė RNR yra citoplazminės RNR pirmtakas.
Citoplazminė RNR sintetinama branduolyje.
c) nukleolinė RNR.
Vertinant bendras turinys baltymų, RNR ir DNR nukleolinėse frakcijose galima pastebėti, kad RNR sudaro apie 10% visos branduolio masės.
Kadangi didžioji dalis citoplazminės RNR yra ribosominė RNR, galime teigti, kad nukleolinė RNR priklauso šiai klasei.
Idėją, kad branduolys yra rRNR sintezės ir ribosomų susidarymo vieta, patvirtino tai, kad RNP dalelės buvo išskirtos iš nukleolinių preparatų, kurie tiek RNR sudėtimi (pagal sedimentacijos savybes), tiek dydžiu gali būti apibūdinami kaip. ribosomos arba jų pirmtakai.su skirtingais sedimentacijos koeficientais.
d) branduolio DNR.
Biocheminiais tyrimais išskirtuose branduoliuose buvo rastas tam tikras DNR kiekis, kurį galima identifikuoti su perinukleoliniu chromatinu arba su nukleolinių chromosomų organizatoriais. Išskirtuose branduoliuose DNR yra 5-12% sausos masės ir 6-17% visos branduolio DNR.
Branduolinio organizatoriaus DNR yra ta pati DNR, ant kurios vyksta branduolio sintezė, t.y. ribosomų, RNR.
Taigi iš biocheminių tyrimų kilo mintis, kad branduolio DNR yra lokalizuota daugybė identiškų rRNR sintezės genų. RRNR sintezė vyksta formuojant didžiulį pirmtaką ir jo tolesnį transformavimą (brendimą) į trumpesnes RNR molekules dideliems ir mažiems ribosomų subvienetams.
Tyrinėdami tritonų oocitų branduolius, mokslininkai susidūrė su įdomiu reiškiniu – branduolių pertekliumi. Kiaušialąstės augimo metu atsiranda iki 1000 mažų branduolių, nesusijusių su chromosomomis. Būtent šiuos branduolius nustatė O. Milleris, kartu didėja ir rDNR kiekis viename oocito branduolyje. Šis reiškinys buvo pavadintas stiprinimas . Tai slypi tame, kad vyksta per didelis branduolio organizatoriaus zonos replikacija, daugybė kopijų išsiskiria iš chromosomų ir tampa papildomai veikiančiais branduoliais. Toks procesas yra būtinas tam, kad vienoje kiaušialąstelėje susikauptų didžiulis (10 12) ribosomų skaičius, kuris užtikrins tolesnį embriono vystymąsi. ankstyvosios stadijos net nesant naujos ribosomų sintezės. Pertekliniai branduoliai išnyksta po kiaušinėlio brendimo.
e) Branduolių ultrastruktūra.
Studijuojant didelis skaičius įvairios ląstelės Gyvūnuose ir augaluose pastebima pluoštinė arba tinklinė branduolių struktūra, uždaryta daugiau ar mažiau tankioje difuzinėje masėje. Šioms dalims buvo pasiūlyti pavadinimai: pluoštinė dalis - nukleonema o difuzinė, vienalytė dalis – amorfinė medžiaga, arba amorfinė dalis . Beveik tuo pačiu metu atlikti elektroniniai mikroskopiniai tyrimai taip pat atskleidė pluoštinę-gijinę branduolių struktūrą.
Tačiau tokia siūlinė branduolio struktūra ne visada aiškiai išreikšta. Kai kuriose ląstelėse susilieja atskiros nukleonemų gijos, o branduoliai gali būti visiškai vienalyčiai.
Atidžiau patyrus branduolį, matyti, kad pagrindinis konstrukciniai komponentai branduolys – tankios apie 15 nm skersmens granulės ir plonos 4-8 nm storio fibrilės. Daugeliu atvejų (žuvų ir varliagyvių oocitai, meristematinės augalų ląstelės) fibrilinis komponentas surenkamas į tankią centrinę zoną (šerdį), kurioje nėra granulių, o granulės užima periferinę branduolio zoną. Daugeliu atvejų (pavyzdžiui, augalų šaknų ląstelėse) papildomos struktūros šioje granuliuotoje zonoje nepastebima.
Nustatyta, kad amorfinės branduolių sritys yra nevienalytės. Jų struktūroje atsiskleidžia žemos spalvos zonos – fibriliniai centrai – ir juos supančios tamsesnės sritys, kurios taip pat turi fibrilinę struktūrą.
Be šių dviejų branduolio komponentų Pastaruoju metu daug dėmesio buvo skirta perinukleolinio chromatino struktūrai. Šis chromatinas ir intranukleolinis DNR tinklas yra vieninga sistema ir yra neatskiriama branduolio sudedamoji dalis.
Granulės ir fibrilinė dalis susideda iš ribonukleoproteinų.
Buvo parodyta, kad rDNR yra šviesos fibriliniuose centruose.