Ne kartą minėjome, kad vienas ar kitas iš išvardytų tyrinėtojų ląstelėse pastebėjo branduolius.
Kadangi Schleideno darbe, į kurį kreipiamės toliau, branduoliui teikiama ypatinga reikšmė, tada, nukrypdami nuo chronologinio pateikimo, čia nagrinėsime šios svarbiausios ląstelės dalies atradimo istoriją. Būtent branduolys padėjo Schwannui palyginti gyvūnų ir augalų ląsteles, todėl branduolio atradimas žymi svarbiausią ląstelės teorijos raidos etapą.
Branduolius žuvų eritrocituose pirmą kartą pamatė Löwenhoekas 1700 m. ir pavaizdavo juos paveiksle. Vėliau tame pačiame objekte – daugelio stuburinių ir bestuburių eritrocituose – Hewsonas (1777) nubraižė branduolius. Šio išsilavinimo vertė tame ankstyvas laikotarpis Mikroskopijos kilmės, žinoma, negalėjo įvertinti nei patys autoriai, nei jų amžininkai. Fontana savo angių nuodų tyrime, vaizduodama epidermio epitelio ląsteles ir eritrocitus, traukia branduolio ląsteles ir trumpai mini „tekste jas; bet ir tuo metu (Fontano veikalas buvo išleistas 1781 m.), kai mikroskopinis tyrimas gyvūnų audinių, Fontano atradimo nepavyko suprasti.
Tuo pačiu metu kai kurie mokslininkai stebėjo kiaušinių branduolius. Cavolini (Filippo Cavolini, 1756-1810) pamatė branduolius žuvų kiaušiniuose (1787); ir Poly (Poli, 1791) pažymėjo moliuskų kiaušinėlių branduolius. Jų stebėjimai praėjo be pėdsakų, neatkreipdami į save dėmesio.
Tyrime apie paukščių kiaušinį (1825 m.) Purkinė aprašė „embrioninę pūslelę“ (vesicula germinativa). Tai buvo paukščio kiaušinio branduolys. Pagal Purkine aprašymą, tai „suspaustas sferinis burbulas, padengtas ploniausiu apvalkalu. Jame yra sava limfa, ji yra įtraukta į baltąjį krūties gumburą ir yra pilna generacinės galios, todėl aš ją pavadinau „embrionine pūslele“. Purkyne prisirišo prie savo atrasto išsilavinimo didelę reikšmę; po jo vėlesni tyrinėtojai šio paslaptingo „burbulo“ nebeignoravo. Taigi Purkinet atradimas nepraėjo be pėdsakų, kaip Cavolini ir Poli pastebėjimai, tačiau „gemalinės pūslelės“ reikšmė ilgą laiką liko neaiški, nes suprantant kiaušinėlio dalis, iš taško. „ląstelės“ koncepcijos požiūriu, teisingu keliu buvo apibrėžta tik po Schwann tyrimų.
Augaluose pirmąjį ląstelės branduolio vaizdą Baueris padarė 1802 m., tačiau šis piešinys buvo paskelbtas tik 1830 m. (J. Baker, 1949). Meyen (1830) vienoje figūroje rodo branduolį. Tyrime apie marchantia Mirbelis (1831-1832) taip pat vaizduoja branduolį, suteikdamas jam rutulio pavadinimą; jį matė ir prancūzų botanikas Brognardas (Adolphe Brogniart, 1801-1876). Tačiau šie pirmieji augalų ląstelių branduolių stebėjimai nebuvo įvertinti pačių stebėtojų ir taip pat nepatraukė dėmesio.
Branduolio kaip nepakeičiamos augalo ląstelės dalies pripažinimas yra anglų botaniko Roberto Browno (Robert Brown, 1773-1858) nuopelnas.
Pradėdamas savo botanikos darbą kelionės į Australiją metu sukurtų kolekcijų aprašymu, Brownas pereina prie tyrimo. anatominė struktūra augalai. Savo darbe jis nekėlė grynai morfologinių problemų; anatominės studijos jam yra augalų taksonomijos tyrimo įrankis, tačiau šiuose darbuose Brownas daro išskirtinius botaninius atradimus, susijusius su augalų reprodukcija. 1833 metais buvo išleistas Browno darbas „Apie orchidėjų apvaisinimo organus ir metodą“ (pranešta Linnean Society Londone jau 1831 m. lapkričio mėn.). Brownas šiame straipsnyje rašo, kad kiekvienoje epidermio ląstelėje jis pastebėjo „vieną suapvalintą areolę, paprastai tamsesnę už ląstelės membraną. Ši areola yra daugiau ar mažiau granuliuota, šiek tiek išgaubta ir, nors atrodo, kad ji yra ant paviršiaus, iš tikrųjų ją dengia išorinė ląstelės plokštelė. Jo padėtis ląstelėje nėra pastovi; tačiau dažnai centre arba šalia jo“ (p. 710). Šią areolę arba ląstelės branduolį (brandulį), kaip Brownas kitaip vadina šiuo dariniu, jis buvo pastebėtas ne tik epidermio ląstelėse; jis pamatė branduolį parenchimoje, vidinėse augalų dalių ląstelėse, „ypač kai jose nėra granuliuotos medžiagos“. Brownas, nors ir atsargiai, rodo, kad branduolys yra paprastas neatskiriama dalis ląstelės. Jis neturi kategoriško teiginio, kad branduolys yra privaloma ląstelės organelė; taip pat Brownas savo darbe nepateikia ląstelių branduolių vaizdų. Nepaisant to, Browno tyrimuose pirmą kartą branduolys minimas ne kaip atsitiktinis darinys ląstelėje, o pasirodo kaip kažkokia esminė dalis, svarbi ląstelės gyvybei.
Meyen, „Fitotomijos“ autorius - darbas, kuris buvo aptartas anksčiau - vėlesniame vadove " Nauja sistema Augalų fiziologija“ (1837-1839) branduolį mini kaip nuolatinę ląstelės dalį, kurios reikšmė lieka paslaptinga. Iš tikrųjų tik Negeli darbai (S. Nageli, 1844) įrodė visuotinį ląstelių branduolių pasiskirstymą ne tik žydinčių augalų, bet ir dumblių, grybų, samanų ir kt. ląstelėse. žemesni augalai.
Gyvūnų histologijoje terminą „branduolys“ įvedė Valentinas. Pranešime „O smulki struktūra jutimo organai“ (1836) Valentinas apie junginės epitelį rašė: „Jį sudaro rombinės arba kvadratinės apvalios, glaudžiai viena šalia kitos esančios ląstelės, kurių ribas sudaro paprastos siūlinės linijos; kiekvienoje ląstelėje be išimties yra kiek tamsesnis ir kompaktiškesnis apvalios arba pailgos apvalios formos branduolys (branduolys). Didžiąja dalimi jis užima ląstelės vidurį, susideda iš smulkiagrūdžios medžiagos, tačiau viduje yra tobulai apvalus kūnas, kuris, vadovaudamasis tuo pačiu modeliu, sudaro savotišką antrąjį branduolį jo viduje“ (p. 143). Šis aprašymas rodo, kad Valentinas aiškiai pastebėjo epitelio ląstelių branduolius. Branduolio viduje Valentinas pamatė branduolį; matyt, tai buvo pirmasis šios intrabranduolinės struktūros aprašymas.
Aiškų epitelio ląstelių branduolių aprašymą ir vaizdą pateikė Henle (1837). Bakeris (1949) teisingai pažymi, kad Valentino ir Henle darbai pradeda branduolinių ląstelių erą gyvūnų histologijoje.
1838 m. Muller's Archiv pasirodė jauno botaniko Schleideno straipsnis pavadinimu „Fitogenezės medžiagos“ (Beitrage zur Phytogenesis). Šis darbas tradiciškai laikomas gairės ląstelinio mokymo raidoje, o jo autorius kartu su Schwannu pripažįstamas kūrėju ląstelių teorija. Schleideno reikšmė ląstelių teorijos istorijoje yra neginčijama, tačiau mokomojoje, populiariojoje, o kartais ir istorinėje literatūroje ši reikšmė nušviečiama paviršutiniškai ir neteisingai. Schleidenas kartais priskiriamas beveik atradimui augalų ląstelės, Štai kodėl. reikia suprasti, kokia iš tikrųjų yra šio mokslininko reikšmė ląstelių mokymo istorijoje, kur yra tiesa legendoje, kuri susiformavo aplink jo kūrybą ir tradiciškai pereina iš vadovėlio į vadovėlį.
Matthias Schleidenas(Matthias Jacob Schleiden, 1804-1881) yra didžiausias atstovas Praėjusio amžiaus vidurio vokiečių botanika. Iš pradžių baigė Teisės fakultetą ir vertėsi advokato praktika. Neturėdamas sėkmės šioje veikloje, Schleidenas 1831 m. metė teisę ir pradėjo studijuoti mediciną ir gamtos mokslai. Nuo 1840 m. buvo botanikos profesorius Jenoje, kur išbuvo iki 1862 m. Tai pagrindinis laikotarpis. kūrybinė veikla Schleiden. 1842 m. buvo paskelbtas pagrindinis jo veikalas „Mokslinės botanikos pagrindai“, suvaidinęs svarbų vaidmenį tolesnių botaninių tyrimų kryptimi. Vietoj gamtos-filosofinių samprotavimų Schleidenas reikalauja įvado į botaniką tikslūs metodai augalų sandaros ir funkcijos tyrimai; jis ypač pabrėžė būtinybę atkreipti dėmesį į raidos istoriją, kurioje įžvelgė daugelio prieštaringų problemų sprendimo raktą. Filosofinės Schleideno pozicijos, išdėstytos jo raštuose, nėra originalios ir atskleidžia Kanto filosofijos pėdsaką. 1862–1864 metais Schleidenas buvo antropologijos profesorius Dorpate (dabar Tartu, Estijos TSR), 1864 metais paliko Dorpatą dėl susidūrimo su bažnyčios ratais ir tuo pačiu sustojo. pedagoginė veikla. Būdamas ne tik serialo autoriumi mokslo darbai, bet ir daug populiarių kūrinių, Schleidenas buvo plačiai žinomas.
„Fitogenezės medžiagos“ yra antrasis Schleideno, kuris tada dar buvo pradedančiojo botaniko, darbas. Tai apie 40 puslapių apimties straipsnis, prie kurio pridedamos dvi lentelės. Bendras pagrindinis žmogaus proto dėsnis, – taip Schleidenas pradeda savo darbą, – dėsnis, lemiantis jo nenugalimą pažinimo vienybės troškimą ir sukurti analogiją tiek apskritai moksle, tiek organizmų srityje. dideli departamentai - gyvūnų ir augalų karalystė, - paskatino ne kartą nagrinėti šią temą. Tiek daug minčių tuo buvo užimta, bet – to negalima paneigti – visi ligšioliniai bandymai šiuo atžvilgiu žlugo ir buvo kliedesiai. Priežastis slypi tame, kad individo samprata, ta prasme, kad ji taikoma gyvūnų gamtoje, nėra pritaikyta augalų pasaulyje. Didžiausias dalykas, ką galima pasakyti apie individą šia prasme, yra žemiausi augalai, kai kurie dumbliai ir grybai, susidedantys tik iš vienos ląstelės. Bet kiekvienas labiau išsivystęs augalas yra visiškai individualizuotų uždarų individualybių visuma... kurios yra ląstelės“ (p. 137). Mes sąmoningai tai atsinešėme ilga citata, kuris yra Schleideno straipsnio pradžia, siekiant parodyti, kaip jam svetima idėja apie gyvūnų ir augalų mikroskopinės struktūros vienybę, išreikštą ląstelių struktūroje. Tuo tarpu būtent ši idėja yra kertinis akmuo ląstelių teorijai, kurios vienu iš bendraautorių dažniausiai laikomas Schleidenas.
Norint teisingai įvertinti Schleideno darbą, reikia prisiminti ląstelių teorijos padėtį botanikoje iki 1837 m., kai Schleidenas baigė savo darbą. Visiškai klaidinga mintis, kad Schleidenas įrodė visuotinį ląstelių pasiskirstymą augaluose ar net atrado ląsteles augaluose. Tai yra tikrovės iškraipymas. istorinė raida Mokslai. Praėjusio amžiaus trečiojo dešimtmečio pradžioje botanikoje buvo sukurta visapusiška ląstelės, kaip elementarios struktūros, idėja. flora; Schleidenas savo darbe laikosi šios pozicijos kaip nepajudinamai įtvirtintos išvados. Netgi tokios neląstelinės augalų dalys, kaip anksčiau atrodė, kaip vandenį laikantys medienos indai, šiuo metu laikomos modifikuotomis, savotiškai diferencijuotomis ir susiliejusiomis ląstelėmis. Schleidenui nereikėjo nustatyti visuotinio ląstelių pasiskirstymo augaluose: šios pozicijos nustatymas buvo, kaip matėme, kolektyvinė daugelio botanikų galaktikos darbų sėkmė praėjusio amžiaus pirmąjį ketvirtį.
K. A. Timirjazevas (1920) teisingai rašė apie posakį „ląstelės atradimas“: „Bet faktas yra tas, kad ląstelės niekas neatrado“ (p. 79), taip pabrėždamas, kad ląstelės „atradimas“ nėra nuopelnas. koks nors mokslininkas. Taip pat netiesa, kad Schleidenas, kaip rašo Aschoffas (1938), sukūrė „visa apimančios konstrukcijos iš visų augalų ląstelių“ doktriną (p. 177). Ir šiuo atžvilgiu teisus K. A. Timirjazevas, kuris rašė: „Šleidenas paprastai laikomas šios ląstelės doktrinos, kuri pasirodė tokia turtinga vaisingiausių apibendrinimų, kūrėju. Tačiau vargu ar tai teisinga... Iškalbingas, aistringas rutinos ir sąstingio priešininkas Schleidenas, kaip kadaise Baconas, galėtų pagrįstai pasakyti apie save, kad yra trimitininkas, šauklys, buccinatorius, skelbdamas apie šios doktrinos atsiradimą, tačiau tai patvirtinantys faktiniai duomenys jau egzistavo anksčiau...“ (p. 75). Būdinga, kad Ungeris (Unger, 1846) savo botanikos pagrinduose, nubrėždamas ląstelės, kaip universalios elementarios organizmų struktūros, padėtį, literatūrinėje nuorodoje nurodo Schwanną ir Köllikerį, šiuo aspektu net nepaminėdamas Schleideno.
Pati ląstelės koncepcija Schleiden nesiskiria nuo idėjų, kurios susiformavo anksčiau ir buvo atspindėtos Meyeno vadovėlyje (1830 m.) dar prieš tai, kai Schleidenas net pradėjo studijuoti botaniką. Šių idėjų lygmenyje Schleidenas ląstelę laikė burbulu arba kamera, kurią riboja apvalkalas, kurio viduje gali būti turinys. Šis ląstelės „turinys“ (ateities protoplazma!) patraukė ir Meyeno dėmesį, kuris jam skyrė daug tyrimų, tačiau nesuprato šio pagrindinio ląstelių komponento reikšmės. Schleidenas taip pat matė augalų ląstelių protoplazmą, tačiau nesuprato ląstelės „turinio“ prasmės. Jam tai guma (Gummi) arba želė (Gallerte). Dalis protoplazmos Schleiden priskiriama ląstelės sienelei. Pastarasis, jo nuomone, susideda iš dviejų sluoksnių, tarp jų yra ląstelės branduolys – „ditoblastas“, kuris niekada nebūna ląstelės viduje, o visada yra uždarytas ląstelės sienelėje „taip, kad ląstelės sienelė skyla į dvi dalis. plokštelės, iš kurių viena patenka į citoblasto išorę, o kita į vidų. Tas, kuris eina su viduje, dažniausiai minkštesnis ir želatinesnis“ (p. 142). Iš Schleideno piešinių akivaizdu, koks „vidinis sluoksnis ląstelių sienelės» jis paėmė augalų ląstelių protoplazmos parietalinį sluoksnį.
Kokią užduotį savo darbe iškėlė Schleidenas? „Kiekviena ląstelė“, rašo jis, „gyvena dvigubą gyvenimą: visiškai nepriklausomą, susietą tik su savo vystymusi, ir kitą priklausomą, nes ji yra neatsiejama augalo dalis. Tačiau nesunku pastebėti, kad tiek augalų fiziologijai, tiek lyginamajai fiziologijai atskirų ląstelių gyvybiniai procesai paprastai turėtų būti pirmoje vietoje, turėtų būti neišvengiamas pagrindas, ir šiuo atveju pirmiausia keliamas klausimas. : kaip tai savita mažas organizmas, ląstelė?" (p. 138). Ši užduotis, ląstelės genezė, yra Schleideno darbo pagrindas. Genetinis momentas šia prasme buvo iškeltas anksčiau, tačiau negalima paneigti, kad Schleidenas pagal savo laiką šią problemą iškėlė aiškiau nei jo pirmtakai.
Atsakydamas į pateiktą klausimą, Schleidenas plėtoja savo ląstelių formavimosi teoriją. Šioje teorijoje pagrindinis vaidmuo kuriant naujas ląsteles priskiriamas branduoliui. Kaip matėme, jis buvo atrastas gerokai anksčiau nei Schleideno darbas, tačiau nesulaukė jokios aiškios interpretacijos. Schleideno teigimu, branduolys yra „citoblastas“ – ląstelės buvęs. Schleideno sukurtą ląstelių formavimosi teoriją galima trumpai apibūdinti taip.
Gumoje, greta iš vidaus su jau esamų ląstelių sienelėmis, susidaro grūdeliai; Schleidenas jas vadina gleivėmis ir mano, kad šie grūdeliai kondensacijos būdu sudaro branduolius, o tada susidaro branduolys, kuris aplink branduolį atrodo kaip granuliuotos nuosėdos. Branduolio paviršiuje, viena vertus, iš „gleivių“ vėl susidaro apvalkalas; jis riboja citoblastą, ir taip atsiranda sienelė, kurioje branduolys yra uždarytas sienelės storiu. Ši erdvė yra nauja ląstelė. Todėl, pasak Schleideno, dukterinės ląstelės atsiranda motininių ląstelių viduje. Naujų ląstelių, kurios gali išsivystyti vienoje motinos ląstelėje, skaičiaus, taip pat šios motinos auklėjamosios ląstelės likimo Schleidenas neaptaria.
Tai yra ląstelių formavimosi teorijos esmė, „puikaus Schleideno tyrimo, kuris atskleidė tiek daug šios srities šviesos“, esmė – tokį Schleideno darbo apibūdinimą pateikė Theodoras Schwannas. Kaip netrukus buvo parodyta, Schleideno teorija remiasi klaidingai interpretuotais pastebėjimais. Būtent šią neteisingą ląstelių formavimosi teoriją iš Schleiden perėmė jo draugas Švanas, ir tai buvo silpniausia Schwanno mokymo vieta. Sachsas savo botanikos istorijoje apibūdina Schleideno teoriją tokiais šiurkščiais žodžiais: „Schleideno ląstelių formavimosi teorija atsirado dėl nesuprantamos neaiškių stebėjimų ir išankstinių nuomonių sankaupos, be to, ji labai primena senąsias Sprengelio ir Trevirano teorijas. 76 p.). Pats Schleidenas atkakliai gynė savo citogenezės teoriją ir citavo ją net 4-ajame leidime. „Mokslinės botanikos pagrindai“ (1861).
Savo straipsnyje Schleidenas, be nagrinėjamos ląstelių formavimosi teorijos, nagrinėja spiralinių kraujagyslių sienelių sustorėjimų atsiradimą ir plėtoja teorinius argumentus apie augalų darbą. Schleideno kūryboje šioje dalyje nėra nieko iš esmės naujo, o kadangi ši straipsnio dalis nėra tiesiogiai susijusi su mūsų tema, tai nėra reikalo prie jos gilintis.
Koks turėtų būti įvertinimas istorinis aspektas Schleideno vaidmuo plėtojant ląstelių mokslą? Martinas Heidenhainas (M. Heidenhainas, 1899) praėjusio amžiaus pabaigoje pastebėjo ekvivalentiškos vertės idėjos neteisingumą Schleideno ir Schwanno ląstelių teorijos istorijoje. Vėliau šį klausimą vėl ryžtingai iškėlė čekų histologas Studnichka (1933), didelis ląstelių teorijos istorijos žinovas, kritine literatūros analize. Iš tiesų, tradicinis Schleiden ir Schwann vardų palyginimas, paprastai pateikiami kaip ląstelių teorijos „bendraautoriai“, nėra pateisinamas kruopščiu šaltinių tyrimu. Schleidenas nebuvo ląstelių teorijos bendraautoris; jam, kaip matėme, buvo visiškai svetima pagrindinė šios teorijos idėja - mikroskopinės elementariosios gyvūnų ir augalų struktūros vienybė; jis nėra ląstelių doktrinos botanikos srityje kūrėjas, nes pagrindinės šios doktrinos nuostatos buvo parengtos anksčiau nei jis. Tai būtina pabrėžti, nes tiek užsienio, tiek mūsų literatūroje apie Šleideno vardą buvo sukurta „legenda“, kurios mokslo istorijoje tiek daug dėl nepakankamos pažinties su originalais. Studnichka minėtame straipsnyje apie Schleideną citavo ištraukas iš kelių dešimčių užsienio histologijos ir biologijos žinynų ir net iš specialių straipsnių apie ląstelių mokslo istoriją, kur Schleideno vaidmuo yra visiškai klaidingas ir kartojama legenda, kad mokslas. skolingas Schleidenui už atradimą ląstelių struktūra augaluose, kad Schleidenas ir Schwannas sukūrė ląstelių teoriją ir tt. Studnicho paminėtą nepagrįstų, o kartais tiesiog juokingų teiginių apie Schleideno vaidmenį sąrašą, deja, galima pridėti nemažą sąrašą citatų iš naujesnių vadovėlių ir net ypatingi kūriniai, įskaitant raštus, kurie pretenduoja į istorinius kūrinius tiek mūsų, tiek mūsų šalyje užsienio literatūra. Istorinis Schleideno kūrybos vaidmuo neabejotinas, tačiau šis vaidmuo skiriasi nuo to, kas paprastai nušviečiama. Schleidenui priskiriamas genetinio požiūrio įvedimas į audinių ir ląstelių teoriją. Panašaus požiūrio bandymų buvo dar prieš Šleideną (Vilkas, Mirbelas, Sprengelis, Treviranas; gyvūnų histologijoje – Valentinas), tačiau tuo metu jie negalėjo būti tokie veiksmingi kaip Schleideno darbai, atsiradę, kai ląstelės idėja kaip pagrindinė augalų struktūra jau buvo įprasta. Be genetinio požiūrio Schwann nebūtų galėjęs sukurti nuoseklios ląstelių teorijos, pagrįstos įtikinamais to meto duomenimis. Tik remdamasis audinių ir ląstelių vystymosi istorija, Schwannas sugebėjo parodyti įvairių elementarių struktūrų „atitikimą“, galėjo įrodyti jų homologiją. Žinoma, Schleideno darbai suvaidino reikšmingą vaidmenį nukreipiant Schwanno mintį į panašų tyrimų kelią.
Bet tai dar ne viskas. Kad būtų galima įtikinamai parodyti jų homologiją, remiantis elementariųjų struktūrų raidos istorija, reikėjo rasti pagrindinį bruožą ir, atsižvelgiant į jį kaip pagrindinę grandį, išnarplioti sudėtingų santykių tarp elementariųjų struktūrų raizginį gyvūne. audinių. Schwann išmoko šios pagrindinės savybės iš Schleiden. Tai yra esmė. Ląstelės viduje įvairių audinių išoriškai gali labai skirtis vienas nuo kito, tačiau ryškus branduolių panašumas, padedantis homologuoti išoriškai nepanašius darinius. Branduolys buvo žinomas ir augalų ląstelėse, ir gyvūnų struktūrose iki Šleideno. Tačiau tik jo darbe branduolys įgavo pagrindinės besivystančios ląstelės ypatybės reikšmę. Šis ženklas tarnavo kaip svirtis Schwannui, kurią suvokęs jis sugebėjo sukurti ląstelių teoriją.
Tai yra Schleideno reikšmė ląstelių mokslo istorijoje. Jo negalima statyti šalia Schwann, jis nebuvo ląstelių teorijos bendraautoris, tačiau jo darbas buvo būtina grandis tyrimų grandinėje, parengusioje medžiagą, be kurios Schwanno genijus, ko gero, būtų bejėgis. kad jo suformuluotus apibendrinimus padarytų ląstelių teorijos pavidalu. Virchow (1859) teisingai tai išreiškė, nurodydamas, kad Schwann stovėjo „ant Schleideno pečių“.
Jei radote klaidą, pažymėkite teksto dalį ir spustelėkite Ctrl + Enter.
Ląstelės branduolio B diagrama ląstelių biologija, šerdis(lot. branduolys) – ląstelių organelės, esančios daugumoje eukariotų ląstelių ir turinčios branduolinių genų, sudarančių dauguma genetinė medžiaga. Branduolys atlieka dvi pagrindines funkcijas: nukreipia chemines reakcijas citoplazmoje ir saugo informaciją, reikalingą ląstelių dalijimuisi.
Branduolį apie 1833 m. atrado botanikas Robertas Brownas.
Be ląstelės genomo, branduolyje yra tam tikrų baltymų, kurie reguliuoja genetinės informacijos skaitymą. Genų nuskaitymas branduoliniame lygmenyje traukia sudėtingus procesus transkripcija, pirminis mRNR apdorojimas ir subrendusios mRNR eksportas į citoplazmą.
Šerdies skersmuo paprastai yra 8-25 mikrometrai. Jį supa dviguba membrana, vadinama branduoliniu apvalkalu. Branduolinės poros tam tikrais intervalais praeina pro vidinę ir išorinę membranas. Branduolinis apvalkalas reguliuoja ir palengvina transportavimą tarp branduolio ir citoplazmos, atskirdamas cheminės reakcijos vykstančios citoplazmoje dėl reakcijų, vykstančių branduolyje. Išorinė membrana yra ištisinė su grubiu endoplazminiu tinklu (RER) ir gali turėti susijusių ribosomų. Erdvė tarp dviejų membranų (vadinama „perinuklearine erdve“) yra ištisinė su RER liumenais. Branduolinė pusė branduolinis apvalkalas apsuptas gijų tinklo, vadinamo branduoliniu sluoksniu.
Vidinė dalis Branduolys turi vieną ar daugiau branduolių, apsuptų matricos, vadinamos nukleoplazma. Nukleoplazma yra gelio pavidalo skystis (šiuo požiūriu panašus į citoplazmą), kuriame yra ištirpę daug medžiagų. Šios medžiagos apima nukleotidų trifosfatą, signalines molekules, DNR, RNR ir baltymus (fermentus ir filamentus).
Branduolys ir endomembraninė sistema Genetinė medžiaga branduolyje yra chromatino arba baltymo ir DNR komplekso pavidalu. DNR yra kaip visa linija atskiros molekulės, žinomos kaip chromosomos. Yra dviejų tipų chromatinas: euchromatinas ir heterochromatinas. Euchromatinas yra ne tokia kompaktiška DNR forma, euchromatino formos DNR regionuose yra genų, kuriuos dažnai skaito ląstelė.
Heterochromatine DNR yra kompaktiškiau. Heterochromatino formos DNR regionuose yra genų, kurių ląstelė neskaito šiame vystymosi etape (šis heterochromatino tipas žinomas kaip fakultatyvinis heterochromatinas) arba yra regionai, sudarantys chromosomų telomerus ir centromerus (šio tipo heterochromatinas). yra žinomas kaip konstruktyvus heterochromatinas). At daugialąsčiai organizmai, ląstelės yra labai specializuotos atlikti specifines funkcijas, todėl reikalingi ir skaitomi skirtingi genų rinkiniai. Todėl DNR sritys, kurios yra heterochromatino pavidalu, priklauso nuo ląstelės tipo.
Branduolys yra tanki branduolio struktūra, kurioje yra surinkti ribosomos elementai. Branduolys gali turėti vieną ar daugiau branduolių.
III. ląstelės branduolys.
Branduolį 1831 metais atrado ir aprašė anglų botanikas R. Brownas.
Jis būdingas visoms eukariotų ląstelėms, išskyrus brandžius žinduolių eritrocitus ir augalų sieto vamzdelius. Ląstelės gali būti vienabranduolinės (dažniausiai); daugiabranduolės (žmonių ir žinduolių kepenų ir raumenų ląstelės, grybai ir kt.). Ląstelės branduolio forma ir dydis yra labai įvairus ir priklauso nuo organizmo tipo, taip pat nuo tipo, amžiaus ir funkcinė būklė ląstelės. Apvaliose arba daugiakampėse ląstelėse dažniausiai būna rutuliškos, pailgose – lazdelės arba ovalios, leukocituose – letenėlės ar net daugiaskilties (kai kurių vabzdžių ir vorų voratinklinių liaukų ląstelėse). Branduolio skersmuo nuo 3 iki 10 mikronų (5 - 20 mikronų)
Autorius cheminė sudėtis branduolys skiriasi nuo likusios ląstelės dalies didelis kiekis DNR (15 - 30%) ir RNR (12%). 99% ląstelės DNR yra sutelkta branduolyje, kur ji kartu su baltymais (histonais ir nehistonais) sudaro kompleksus – dezoksiribonukleoproteinus (DNP).
Branduolio funkcijos:
1) genetinės informacijos saugojimas ir dauginimas;
2) ląstelėje vykstančių medžiagų apykaitos procesų reguliavimas.
Ląstelės gyvavimo ciklas (ląstelių ciklas) apima du laikotarpius:
1) dalijimasis, dėl kurio susidaro dvi dukterinės ląstelės;
2) laikotarpis tarp dviejų padalijimų, kuris vadinamas - tarpfazė.
Interfazės metu ląstelė ruošiasi dalytis: ląstelė auga, vyksta DNR replikacija ir įvairių medžiagų reikalinga dalijimuisi, kaupiama energija. Dalijimosi procese ląstelių struktūrosšerdys patiria reikšmingų pokyčių.
Tarpfaziniame branduolyje yra: branduolio membrana; branduolinės sultys; branduoliai ir chromatinas.
branduolinis apvalkalas(kardiolema) vaizduoja dvi elementarios membranos, tarp kurių yra perinuklearinė erdvė . Išorinė branduolinės membranos membrana, besiribojanti su hialoplazma, turi sudėtingą sulankstytą struktūrą, kai kuriose vietose sujungta su EPS kanalais. Susiformuoja perinuklearinė erdvė ir ER viena sistema komunikacijos kanalai. Ribosomos yra ant išorinės branduolio membranos, vidinė membrana yra lygi. Branduolinė membrana persmelkta daug porų, skersmuo apie 30 - 100 nm. Jų skaičius skiriasi priklausomai nuo branduolio funkcinio aktyvumo. Jaunose ląstelėse visada yra daugiau porų nei senose.
Branduolinio apvalkalo funkcija:
1. metabolizmo tarp branduolio ir citoplazmos reguliavimas;
2. apsauginė funkcija.
Medžiagų mainai tarp branduolio ir citoplazmos vyksta keliais būdais:
1. Pro poras, užtikrinančias selektyvų pralaidumą;
Pavyzdys: mRNR ir ribosomų subdalelių išskyrimas į citoplazmą arba ribosomų baltymų, nukleotidų ir molekulių, reguliuojančių DNR aktyvumą, patekimas į branduolį.
2. Išvarstydami branduolinės membranos ataugas ir iškilimus.
3. Difuzijos būdu per branduolio apvalkalą.
Nepaisant aktyvaus medžiagų apykaitos tarp branduolio ir citoplazmos, branduolio membrana atskiria branduolio turinį nuo citoplazmos, todėl atsiranda speciali intrabranduolinė aplinka, kuri skiriasi nuo aplinkinės citoplazmos.
branduolinės sultys(nukleoplazma; karioplazma) – vienalytė masė, užpildanti tarpą tarp branduolio struktūrų (chromatino ir nukleolių), susidedanti iš baltymų (fermentų), nukleotidų, aminorūgščių ir Skirtingos rūšys RNR (mRNR, tRNR, rRNR) ir DNR, taip pat tarpinius medžiagų apykaitos produktus. Branduolinės sultys atlieka branduolinių struktūrų ryšį ir mainus su ląstelės citoplazma.
Chromatinas atstovauja dezoksiribonukleoproteinas(DNP), aptinkamas šviesiu mikroskopu mažų granulių arba gumulėlių plonų sruogelių (fibrilių) pavidalu. Chromatino pagrindas yra nukleoproteinai – ilgos siūlinės DNR molekulės (apie 40%), susijungusios su specifinėmis baltymai – histonai(40%) kurių sudėtyje yra didelis skaičius lizinas ir argininas.
Branduolio dalijimosi procese nukleoproteinai spiralizuojasi, trumpėja ir taip sutankina (100-500 kartų) į kompaktiškas lazdelės formos chromosomas.
Chromosomos(graikų chromas – dažai ir soma – korpusas) buvo taip pavadinti dėl gebėjimo intensyviai dažytis. Chromosomose taip pat yra RNR, rūgščių baltymų, lipidų ir mineralai(Ca ir Mg jonai), taip pat fermentas DNR polimerazė, būtinas DNR replikacijai. Interfazės metu chromosomos matomos tik apačioje elektroninis mikroskopas ir yra DNP, vadinamas chromatinu (despiralizuota chromosomų būsena).
Metafazinė chromosoma susideda iš dviejų išilginių DNP gijų - chromatidų, sujungtų viena su kita pirminio susiaurėjimo srityje - centromerai . Centrinėje centromero dalyje yra kinetochores prie kurių mitozės metu prisitvirtina dalijimosi verpstės gijų mikrovamzdeliai. Kai kurios chromosomos turi antrinis susiaurėjimas, nesusijęs su veleno sriegio tvirtinimu. Ši chromosomų sritis kontroliuoja branduolio (branduolių organizatoriaus) sintezę ir vadinama palydovu. Centromeras padalija chromosomą į dvi rankas.
Centromero vieta lemia 3 pagrindinius chromosomų tipus:
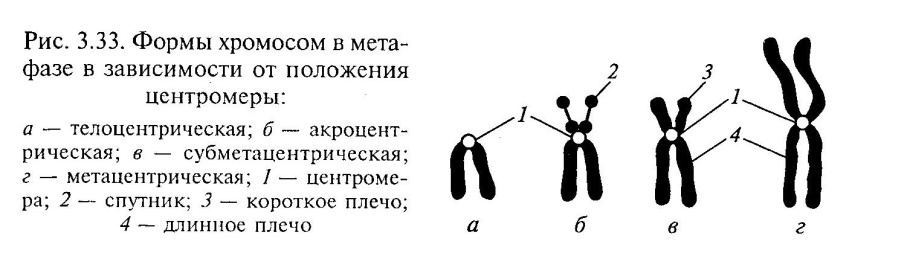 vienodo peties
(metacentrinis) - vienodo ilgio pečiais;
vienodo peties
(metacentrinis) - vienodo ilgio pečiais;
Submetacentrinis arba nelygūs pečiai – nevienodo ilgio pečiai;
strypo formos- su vienu ilgu, o antru labai trumpu pečiu (akrocentriniu).
Chromosomų struktūra aiškiai matoma mitozės metafazės stadijoje. Chromosomų tyrimas leido nustatyti šiuos faktus:
ü Visose bet kurio augalo ar gyvūno organizmo somatinėse ląstelėse chromosomų skaičius yra vienodas;
ü Lytinėse ląstelėse visada yra perpus mažiau chromosomų nei tam tikro tipo organizmo somatinėse ląstelėse;
ü Visi tai pačiai rūšiai priklausantys organizmai turi vienodą chromosomų skaičių ląstelėse.
Pavyzdys: arklio apvaliosios kirmėlės - 2; Drosophila musė - 8; Šimpanzės - 48; Pelenai - 46; Vyras - 46; balandis - 80; Karpiai - 104; Radioliarija (protistas) - 1000 - 1600.
Chromosomų skaičius nepriklauso nuo organizacijos aukščio ir ne visada rodo filogenetinius ryšius: tokį patį skaičių galima rasti rūšyse, kurios sistemingai yra labai nutolusios viena nuo kitos ir labai skiriasi artimos kilmės organizmuose. Taigi chromosomų skaičius nėra rūšiai būdingas bruožas. Tačiau chromosomų rinkinio, kaip visumos, charakteristika yra būdinga rūšiai, t.y. būdingas tik vienos rūšies augalams ar gyvūnams.
Kariotipas- somatinės ląstelės chromosomų rinkinio kiekybinių (skaičius ir dydis) ir kokybinių (formos) požymių rinkinys. Chromosomų skaičius kariotipe visada yra lygus. Taip yra dėl to, kad somatinėse ląstelėse yra 2 tos pačios formos ir dydžio chromosomos. Vienas kyla iš tėvo organizmo, antrasis iš motinos.
Chromosomos, kurios yra vienodo dydžio ir formos bei turi tuos pačius genus, vadinamos homologiškas.
Chromosomų skaičius brandžiose lytinėse ląstelėse vadinamas haploidas (viengubas) ir žymimas Lotyniška raidė n. Somatinėse ląstelėse yra dvigubas chromosomų skaičius - diploidinis rinkinys – pažymima 2n. Ląstelės, turinčios daugiau nei du chromosomų rinkinius, vadinamos poliploidas (4n, 8n, 16n ir kt.)
Branduoliai -šios yra suapvalintos, stipriai sutankintos, neribotos 1–2 μm ir didesnio skersmens ląstelės branduolio membraninėmis sritimis. Branduolių forma, dydis ir skaičius priklauso nuo branduolio funkcinės būklės: kuo didesnis branduolys, tuo didesnis jo aktyvumas. Branduolys gali būti nuo 1 iki 10 branduolių, o kartais, pavyzdžiui, mielių branduoliuose, jų visiškai nėra. Branduolys turi apie 80 % baltymų, 10-15 % RNR, kai kurių DNR ir kitų cheminių komponentų.
Branduoliai yra tik nesidalijančiuose branduoliuose, dalijimosi metu jie išnyksta, o pasibaigus dalijimui vėl susidaro aplink tam tikras chromosomų dalis – genus, vadinamus branduolio organizatoriais.
Branduolys RNR jungiasi su baltymais, todėl susidaro ribonukleoproteinai – ribosomų pirmtakai. Pastarieji pro branduolinės membranos poras patenka į citoplazmą, kur baigiasi jų formavimasis. Taigi, branduolys yra rRNR ir ribosomų sankaupa skirtingi etapai dariniai.
Branduolį 1833 metais atrado ir aprašė anglas R. Brownas. Branduolys yra visuose eukariotinės ląstelės, išskyrus subrendusius eritrocitus ir augalų sietus. Ląstelės dažniausiai turi vieną branduolį, tačiau kartais randama ir daugiabranduolių.
Branduolys yra sferinis arba ovalo formos. Kai kurios ląstelės turi segmentuotus branduolius. Branduolių skersmuo yra nuo 3 iki 10 mikronų.
Branduolys yra būtinas ląstelės gyvybei. Jis reguliuoja ląstelių veiklą. Branduolys saugo paveldimą informaciją, esančią DNR. Ši informacija branduolio dėka perduodama dukterinėms ląstelėms ląstelių dalijimosi metu. Branduolys lemia ląstelėje sintetinamų baltymų specifiškumą. Branduolys turi daug baltymų, reikalingų jo funkcijoms. RNR sintetinama branduolyje.
Branduolys turi branduolinę membraną, kuri atskiria jį nuo citoplazmos, karioplazmos (branduolių sulčių), vieno ar kelių branduolių, chromatino
Branduolinis apvalkalas sudarytas iš dviejų membranų. Jame yra poros, kurios žaidžia svarbus vaidmuo transportuojant medžiagas į citoplazmą ir iš jos. Poros nėra nuolatiniai dariniai. Jų skaičius skiriasi priklausomai nuo branduolio funkcinio aktyvumo. Didžiausio branduolinio aktyvumo laikotarpiu porų skaičius didėja. Branduolinis apvalkalas yra tiesiogiai susijęs su endoplazminiu tinklu.
Įjungta išorinė membrana branduolinis apvalkalas, su lauke yra ribosomų, kurios sintetina specifinius baltymus, kurie susidaro tik branduolio apvalkalo ribosomose.
Branduolinės sultys (karioplazma) – vidinis branduolio turinys, yra baltymų, nukleotidų, jonų tirpalas, klampesnis už hialoplazmą. Jame
taip pat yra fibrilinių baltymų. Karioplazmoje yra nukleolių ir chromatino. Branduolinės sultys suteikia normalus funkcionavimas genetinė medžiaga.
Branduoliai yra privalomas branduolio komponentas, jie randami tarpfaziniuose branduoliuose ir yra maži kūnai, sferinės formos. Branduoliai yra tankesni už branduolį. Branduoliuose vyksta rRNR, kitų tipų RNR sintezė ir ribosomų subvienetų susidarymas.
Branduolių atsiradimas yra susijęs su tam tikromis chromosomų zonomis, vadinamomis branduolio organizatoriais. Branduolių skaičių lemia branduolio organizatorių skaičius. Juose yra rRNR genų.
Chromatinas (dažyta medžiaga) yra tanki branduolio medžiaga, gerai nudažyta pagrindiniais dažais. Chromatinas susideda iš DNR molekulių komplekse su baltymais (histonais ir nehistonais), RNR.
Nesiskiriančiuose (tarpfaziniuose) branduoliuose chromatinas gali tolygiai užpildyti branduolio tūrį, būdamas dekondensuotoje būsenoje. Šis difuzinis chromatinas (euchromatinas) yra genetiškai aktyvus. DNR molekulės, turinčios paveldimos informacijos, gali padvigubėti replikacijos metu, galimas genetinės informacijos perkėlimas (transkripcija) iš DNR į mRNR.
Kartais tarpfaziniame branduolyje matomi chromatino gumulėliai, kurie yra kondensuoto chromatino (heterochromatino) sritys. Tai neaktyvios sritys. Pavyzdžiui, ląstelėse moteriškas kūnas, kur yra dvi X chromosomos, viena yra aktyvios difuzinės būsenos, o antroji yra neaktyvios, kondensuotos būsenos.
Branduolio dalijimosi metu chromatinas dažosi intensyviau, jis kondensuojasi – susidaro labiau spiralizuoti (susisukę) siūlai, vadinami chromosomomis.
Chromosomos yra sintetiškai neaktyvios. Chromosomų struktūrą geriausia ištirti didžiausio jų kondensacijos momentu, ty mitozės metafazėje ir anafazės pradžioje.
Kiekviena chromosoma mitozės metafazėje susideda iš dviejų chromatidžių, susidariusių dėl reduplikacijos ir sujungtų centromeru (pirminis susiaurėjimas). Centrinėje centromero dalyje yra kinetochorai, prie kurių mitozės metu prisitvirtina verpstės gijų mikrovamzdeliai (47 pav.). Anafazėje chromatidės yra atskirtos viena nuo kitos. Iš jų susidaro dukterinės chromosomos, turinčios tą pačią genetinę informaciją. Centromeras padalija chromosomą į dvi rankas. Chromosomos su vienodomis rankomis vadinamos lygiarankiais arba metacentrinėmis, kurių rankos nevienodo ilgio - nelygios rankos - submetacentrinėmis, su viena trumpa, o antroji beveik nepastebima - lazdelės formos arba akrocentrinėmis.
Kai kurios chromosomos turi antrinį susiaurėjimą, kuris atskiria palydovą. Antriniai susiaurėjimai vadinami branduolio organizatoriais. Juose, tarpfazėje, susidaro branduolys. Branduolio organizatoriuose yra DNR
atsakingas už rRNR sintezę. Chromosomų rankos baigiasi regionais, vadinamais telomerais, kurie negali prisijungti prie kitų chromosomų.
Kinetochorai yra chromosomų centromerinėje srityje. / - kinetochoras, 2 - kinetochoro mikrovamzdelių pluoštas; 3 - chromatidas.
Chromosomų skaičius, dydis ir forma rinkinyje skirtingi tipai gali skirtis. Chromosomų rinkinio požymių rinkinys vadinamas kariotipu.
Chromosomų rinkinys yra specifinis ir pastovus kiekvienos rūšies individams. Žmonės turi 46 chromosomas, pelės – 40 chromosomų ir t.t.
Somatinėse ląstelėse, turinčiose diploidinį chromosomų rinkinį, chromosomos yra suporuotos. Jie vadinami homologiniais. Viena chromosoma poroje kilusi iš motinos organizmas, kitas – iš tėvo.
Chromosomų struktūros ar jų skaičiaus pokyčiai atsiranda dėl mutacijų.
Kiekviena rinkinio chromosomų pora yra individuali. Chromosomos iš skirtingų porų vadinamos nehomologinėmis.
Kariotipe išskiriamos lytinės chromosomos (žmonėms tai yra X chromosoma ir Y chromosoma) ir autosomos (visos kitos).
Lytinės ląstelės turi haploidinis rinkinys chromosomos.
Chromosomos pagrindas yra DNR molekulė, susieta su baltymais (histonais ir kt.) į nukleoproteiną.
Pagrindinė molekulinės biologijos pozicija, kurią suformulavo F. Crickas, teigia, kad genetinės informacijos perdavimas vykdomas:
1) iš DNR į DNR replikacijos būdu;
2) iš DNR per mRNR (mRNR) į baltymus.
Nukleino rūgščių makromolekulių savaiminio dauginimosi (replikacijos) procesas užtikrina tikslų genetinės informacijos kopijavimą ir jos perdavimą iš kartos į kartą.
Komplementarumo principas, kuriuo grindžiama DNR molekulės struktūra, leidžia suprasti, kaip sintetiniame tarpfazių periode sintezuojamos naujos molekulės. gyvenimo ciklas ląstelės prieš dalijimąsi.