Yra trys kilmės hipotezės eukariotinės ląstelės:
- simbiotinė hipotezė, arba simbiogenezė,
- invaginatyvus,
- chimerinis.
Iki šiol mokslo pasaulyje pripažįstama pagrindinė eukariotų kilmės hipotezė simbiogenezė.
Remiantis simbiogeneze, eukariotų ląstelių organelės, tokios kaip chloroplastai ir žvyneliai, atsirado įvedant vieną prokariotą į kitą, didesnę prokariotinę ląstelę, kuri atliko šeimininko vaidmenį.
Simbiotinėje hipotezėje kyla sunkumų paaiškinant eukariotinių ląstelių branduolio kilmę ir klausimą, kuris prokariotas vis dėlto buvo šeimininkas. Eukariotų genomo ir baltymų molekulinės analizės duomenys rodo, kad, viena vertus, tai buvo archejoms artimas organizmas (anksčiau jos priklausė bakterijoms, vėliau buvo atskirtos į atskirą šaką). Kita vertus, eukariotuose yra baltymų (ir už jų sintezę atsakingų genų), kurie būdingi tobuloms kitoms prokariotų grupėms.
Pagal invaginacijos hipotezę apie eukariotinių ląstelių kilmę, jų organelės susiformavo įsiskverbus cito plazmos membrana vėliau atskiriant šias struktūras. Susidarė kažkas panašaus į rutulius, apsuptus membranos ir turinčius citoplazmos viduje bei čia užfiksuotus junginius ir struktūras. Priklausomai nuo to, kas pateko į vidų, susidarė skirtingos organelės.
Prokariotai tikrų organelių neturi, jų funkcijas atlieka membranos invaginacijos. Todėl tokį suvarstymą lengva įsivaizduoti. Taip pat invaginacijos hipotezės naudai yra citoplazminės membranos ir dvigubų organelių membranų panašumas.
Invaginacijos požiūriu branduolio kilmė lengvai paaiškinama, tačiau nepaaiškinama, kodėl branduolio-citoplazminio komplekso genomas ir ribosomos skiriasi nuo esančių chloroplastuose ir mitochondrijose (prisiminkime, kad juose taip pat yra DNR ir ribosomų). Be to, šiose organelėse baltymų biosintezės sistema (DNR, RNR, ribosomos) panaši į prokariotų.
Šis skirtumas gerai paaiškinamas tik pirmosios, simbiotinės, hipotezės požiūriu. Anot jos, vienaip ar kitaip aerobinis prokariotas pateko į anaerobinį prokariotą. Jis nebuvo virškinamas, o, priešingai, pradėjo valgyti ląstelės šeimininkės sąskaita. Savo ruožtu jis naudojo deguonį energijai gauti, o šis oksidacijos būdas yra daug efektyvesnis, energijos perteklių jis atidavė prokariotiniam šeimininkui, kuris šiuo atveju taip pat buvo naudingas. Atsirado simbiozė. Vėliau įvestas prokariotas tapo paprastesnis, dalis jo genomo migravo į šeimininko ląstelę, ji nebegalėjo egzistuoti savarankiškai.
Panašiai simbiogenezė paaiškina chloroplastų kilmę. Jau buvo introdukuoti tik fotosintezę galintys prokariotai (panašūs į melsvadumblius).
Pirmieji eukariotai be fotosintetinių simbiontų davė pradžią gyvūnams, kuriuose jie atsirado – augalus.
Šiuo metu yra pirmuonių (vienaląsčių eukariotų organizmų), kurie neturi mitochondrijų ar chloroplastų. Tačiau vietoj jų citoplazmoje įsikuria simbiontai prokariotai, atlikdami atitinkamas funkcijas. Šis faktas, taip pat mitochondrijų ir plastidų baltymų biosintezės sistemos panašumas su prokariotais, yra laikomi simbiogenezės įrodymu. Įrodymas taip pat yra tai, kad mitochondrijos ir chloroplastai dauginasi patys, ląstelė jų niekada nesukuria nuo nulio.
Trečiosios, chimerinės, eukariotinių ląstelių kilmės hipotezės naudai sakoma didelis dydis jų genomas, tūkstančius ar daugiau kartų viršijantis bakterijų, taip pat sintezuojamų baltymų įvairovė. skirtingos grupės prokariotai. Aišku, kad eukariotų evoliucijos metu jų genomas komplikavosi, padvigubėjo, jame atsirado daug reguliuojančių genų. Tačiau vis tiek pradinis genomo dydžio padidėjimas galėjo įvykti dėl kelių prokariotų genomų derinio.
Galbūt senovėje tam tikras prokariotas įgijo fagocitozės gebėjimą ir taip maitindamasis, be kita ko, pasisavino kitus prokariotus, kurie ne visada buvo virškinami. Jų genome buvo šeimininkui naudingi genai, ir jis įtraukė juos į savo genomą. Galbūt kai kurie viduje buvę prokariotai tapo organelėmis, o tai sujungia chimerinę hipotezę su simbiogeneze.
Kadangi visos aukščiau išvardintos hipotezės turi stiprią ir silpnosios pusės, ir taip pat iš esmės neatmeta vienas kito pozicijų, tada, mūsų nuomone, daugelio veiksnių, aprašytų skirtingomis hipotezėmis, derinys gali turėti įtakos eukariotinių ląstelių struktūrų kilmei.
Taip pat reikėtų pažymėti, kad pagal simbiotinę hipotezę kitų membraninių organelių, tokių kaip vakuolės, Golgi kompleksas ir kt., kilmė gali būti laikoma tolesniu supaprastinimu, pavyzdžiui, mitochondrijų.
Taip pat pažymime, kad skirtingų prokariotų baltymų derinys eukariotuose gali visai nereikšti pirmųjų chimerizmo. Gali būti, kad tam tikras fermentines funkcijas gali atlikti tik tam tikros struktūros baltymai. Ir eukariotų evoliucija ne kartą pasiekė tai, nepaisant prokariotų grupių, kurios nėra jų protėviai, evoliucijos. Tarkime, įvyko konvergencija molekuliniame lygmenyje.
Eukariotų atsiradimas yra svarbus įvykis. Ji pakeitė biosferos struktūrą ir atvėrė iš esmės naujas progresyvios evoliucijos galimybes. Eukariotinė ląstelė yra ilgos prokariotų pasaulio, pasaulio, kuriame įvairūs mikrobai prisitaikė vienas prie kito ir ieškojo būdų, kaip veiksmingai bendradarbiauti, evoliucijos rezultatas.
laiko juostos kontūras (pakartojimas)

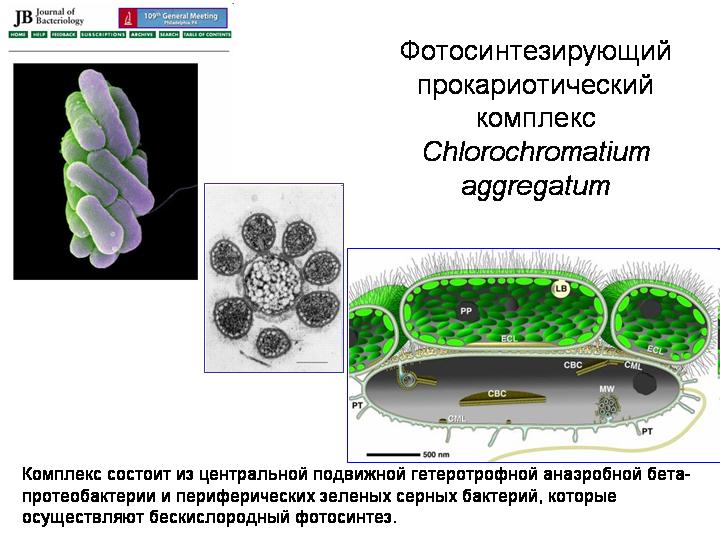
Fotosintetinis prokariotų kompleksas Chlorochromatium aggregatum.
Eukariotai atsirado dėl kelių rūšių prokariotų simbiozės. Prokariotai apskritai yra gana linkę į simbiozę (žr. 3 skyrių „Sudėtingumo gimimas“). Čia yra įdomi simbiotinė sistema, žinoma kaip Chlorochromatium aggregatum. Gyvena giliuose ežeruose, kur gylyje yra anoksinės sąlygos. Centrinis komponentas yra mobili heterotrofinė beta-proteobakterija. Aplink jį yra nuo 10 iki 60 fotosintetinių žaliosios sieros bakterijų. Visi komponentai yra sujungti ataugomis išorinė membrana centrinė bakterija. Bendruomenės prasmė ta, kad mobiliosios beta-proteobakterijos visą kompaniją tempia į išrankioms sieros bakterijoms gyvybei palankias vietas, o sieros bakterijos užsiima fotosinteze ir aprūpina save bei beta-proteobakterijas maistu. Galbūt kai kurios senovinės maždaug tokio tipo mikrobų asociacijos buvo eukariotų protėviai.
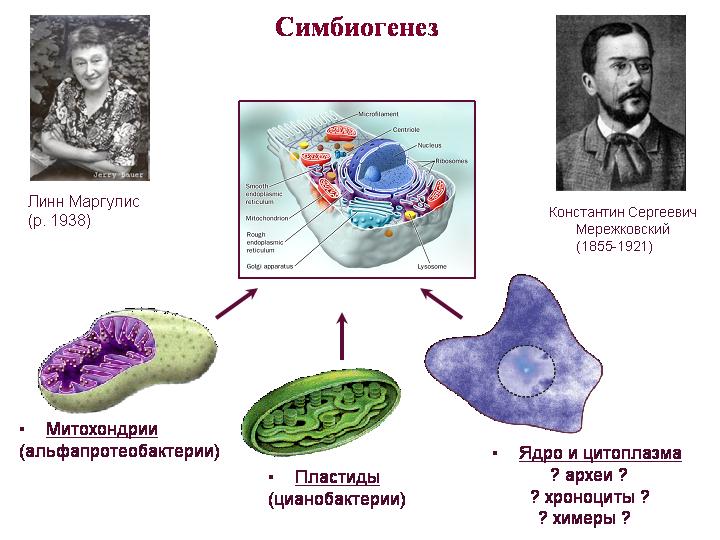
Simbiogenezės teorija. Merežkovskis, Margulis. Mitochondrijos yra alfa-proteobakterijų palikuonys, plastidės yra cianobakterijų palikuonys. Sunkiau suprasti, kas buvo viso kito, tai yra, citoplazmos ir branduolio, protėvis. Eukariotų branduolys ir citoplazma sujungia archėjos ir bakterijų savybes, taip pat turi daug unikalių savybių.
Apie mitochondrijas. Galbūt tai buvo mitochondrijų (o ne branduolio) įsigijimas, kuris buvo pagrindinis eukariotų vystymosi momentas. Dauguma protėvių mitochondrijų genų buvo perkelti į branduolį, kur juos kontroliavo branduolinės reguliavimo sistemos. Šie mitochondrinės kilmės branduoliniai genai koduoja ne tik mitochondrijų baltymus, bet ir daugybę citoplazmoje veikiančių baltymų. Tai rodo, kad mitochondrijų simbiontas vaidino svarbesnį vaidmenį formuojant eukariotinę ląstelę, nei tikėtasi.
Dviejų skirtingų genomų sambūvis vienoje ląstelėje reikalavo sukurti veiksmingą jų reguliavimo sistemą. O norint efektyviai valdyti didelio genomo darbą, reikia išskirti genomą nuo citoplazmos, kurioje vyksta medžiagų apykaita ir tūkst. cheminės reakcijos. Branduolinis apvalkalas tiesiog atskiria genomą nuo turbulentinių citoplazmos cheminių procesų. Simbiontų (mitochondrijų) įgijimas galėtų tapti svarbiu stimulu vystytis branduolio ir genų reguliavimo sistemoms.
Tas pats pasakytina apie seksualinį dauginimąsi. Jūs galite gyventi be lytinio dauginimosi tol, kol jūsų genomas yra pakankamai mažas. Organizmai, turintys didelį genomą, bet neturintys lytinio dauginimosi, yra pasmerkti greitam išnykimui, išskyrus retas išimtis.
![]()
Alfaproteobakterijos – šiai grupei priklausė mitochondrijų protėviai.
Rhodospirillum yra nuostabus mikroorganizmas, galintis gyventi dėl fotosintezės, taip pat ir anaerobinėmis sąlygomis, ir kaip aerobinis heterotrofas, ir netgi kaip aerobinis chemoautotrofas. Pavyzdžiui, jis gali augti dėl oksidacijos smalkės CO nenaudojant jokių kitų energijos šaltinių. Be viso to, jis taip pat žino, kaip fiksuoti atmosferos azotą. Tai yra, jis yra aukščiausias laipsnis universalus organizmas.
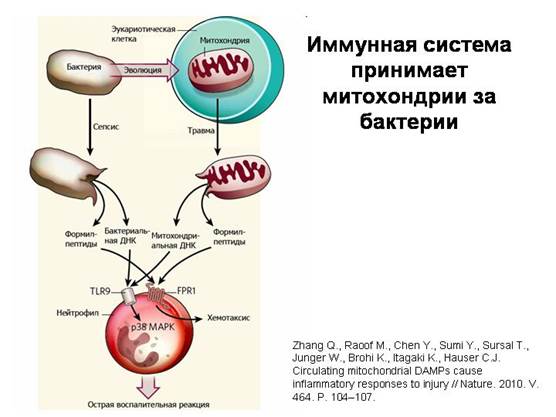
Imuninė sistema painioja mitochondrijas su bakterijomis. Pažeistos mitochondrijos, patekusios į kraują traumos metu, išskiria būdingas molekules, kurios randamos tik bakterijose ir mitochondrijose (bakterijų tipo žiedinė DNR ir baltymai, kurių viename galų yra speciali modifikuota aminorūgštis formilmetioninas). Taip yra dėl to, kad baltymų sintezės aparatas mitochondrijose išliko toks pat kaip ir bakterijose. Ląstelės Imuninė sistema- neutrofilai - į šias mitochondrines medžiagas reaguoja taip pat, kaip ir į bakterines, ir tų pačių receptorių pagalba. Tai aiškiausias mitochondrijų bakterinės prigimties patvirtinimas.
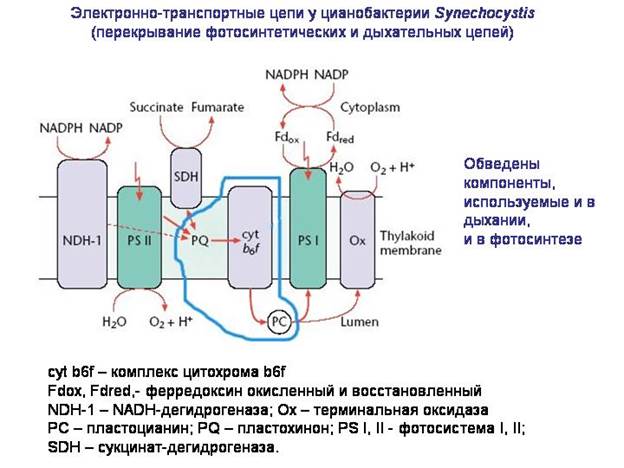
Pagrindinė funkcija mitochondrijos – deguonies kvėpavimas. Labiausiai tikėtina, kad anaerobinio branduolio ir citoplazmos protėvio susiejimo su „protomitochondrijomis“ stimulas buvo būtinybė apsisaugoti nuo toksiškas veiksmas deguonies.
Iš kur bakterijos, įskaitant alfaproteobakterijas, gavo reikalingas molekulines sistemas deguonies kvėpavimas? Atrodo, kad jie buvo pagrįsti molekulinėmis fotosintezės sistemomis. Elektronų transportavimo grandinė, kuri susidarė bakterijose kaip fotosintezės aparato dalis, buvo pritaikyta kvėpuoti deguonimi. Kai kuriose bakterijose elektronų transportavimo grandinių dalys vis dar naudojamos vienu metu tiek fotosintezei, tiek kvėpavimui. Labiausiai tikėtina, kad mitochondrijų protėviai buvo aerobinės heterotrofinės alfa-proteobakterijos, kurios, savo ruožtu, kilo iš fotosintetinių alfa-proteobakterijų, tokių kaip Rhodospirillum.
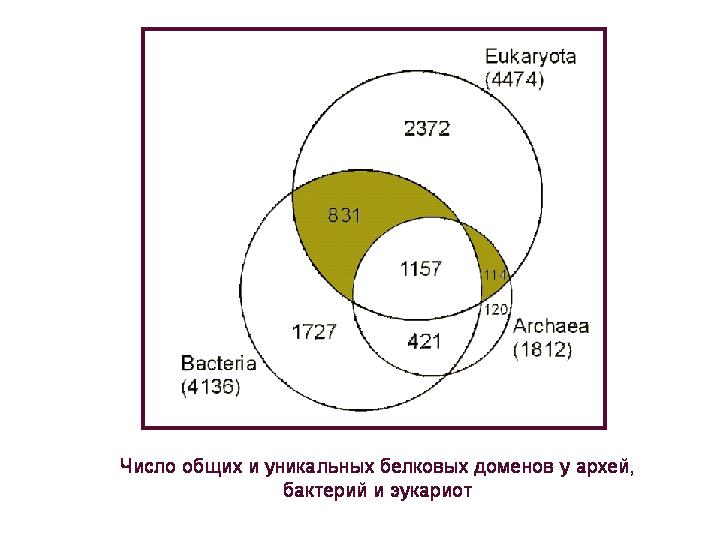
Bendrų ir unikalių baltymų domenų skaičius archėjose, bakterijose ir eukariotuose. Baltymų domenas yra baltymo molekulės dalis, turinti specifinę funkciją ir būdingą struktūrą, tai yra aminorūgščių seka. Kiekvienas baltymas, kaip taisyklė, turi vieną ar daugiau šių struktūrinių ir funkcinių vienetų arba domenų.
4,5 tūkstančio baltymų domenų, kuriuos turi eukariotai, galima suskirstyti į 4 grupes: 1) yra tik eukariotuose, 2) bendri visoms trims superkaralystėms, 3) bendri eukariotams ir bakterijoms, bet jų nėra archea; 4) būdingas eukariotams ir archėjoms, bet nėra bakterijose. Mes apsvarstysime du naujausios grupės(paveiksle jie paryškinti spalva), nes apie šiuos baltymus galima gana užtikrintai kalbėti apie jų kilmę: atitinkamai bakterinę arba archeologinę.
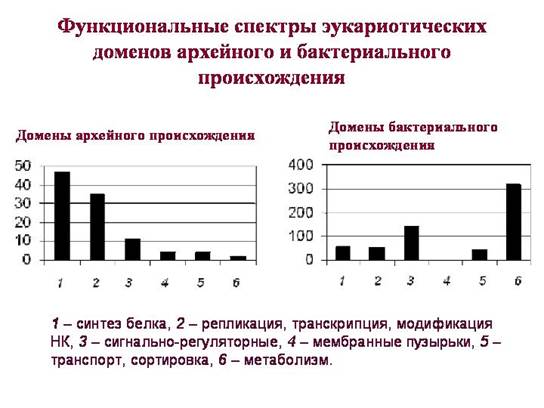
Pagrindinis momentas tuo, kad eukariotų domenai, tariamai paveldėti iš bakterijų ir iš archėjų, atlieka labai skirtingas funkcijas. Iš archajų paveldėti domenai (jų funkcinis spektras parodytas kairiajame grafike) vaidina pagrindinį vaidmenį eukariotinės ląstelės gyvenime. Tarp jų vyrauja sritys, susijusios su genetinės informacijos saugojimu, dauginimu, organizavimu ir skaitymu. Dauguma "archealinių" domenų priklauso toms funkcinėms grupėms, kuriose horizontalūs genų mainai prokariotuose vyksta rečiausiai. Matyt, eukariotai šį kompleksą gavo tiesioginio (vertikalaus) paveldėjimo būdu iš archėjų.
Tarp bakterinės kilmės sričių yra ir su informaciniais procesais susijusių baltymų, tačiau jų nedaug. Dauguma jų veikia tik mitochondrijose arba plastiduose. Citoplazmos eukariotinės ribosomos yra archealinės kilmės, mitochondrijų ir plastidų ribosomos – bakterinės.
Tarp eukariotų bakterijų sričių signalą reguliuojančių baltymų dalis yra daug didesnė. Iš bakterijų eukariotai paveldėjo daug baltymų, atsakingų už ląstelių atsako į veiksnius mechanizmus išorinė aplinka. Taip pat – daug baltymų, susijusių su medžiagų apykaita (plačiau žr. 3 skyrių „Sudėtingumo gimimas“).
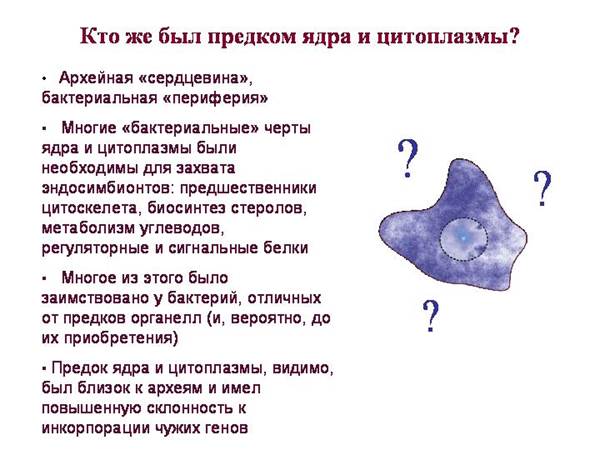
Eukariotai turi:
Archealinė „šerdis“ (darbo su genetine informacija ir baltymų sintezės mechanizmai)
Bakterijų „periferija“ (metabolizmo ir signalų reguliavimo sistemos)
Paprasčiausias scenarijus: ARCHEIA prarijo BAKTERIJAS (mitochondrijų ir plastidžių protėvius) ir visas jo bakteriniai požymiai pirktas iš jų.
· Šis scenarijus yra pernelyg paprastas, nes eukariotai turi daug bakterijų baltymų, kurie negalėjo būti pasiskolinti iš mitochondrijų ar plastidų protėvių.
Eukariotai turi daug „bakterijų“ domenų, kurie nėra būdingi nei cianobakterijoms (plastidų protėviams), nei alfaproteobakterijoms (mitochondrijų protėviams). Jie buvo gauti iš kai kurių kitų bakterijų.
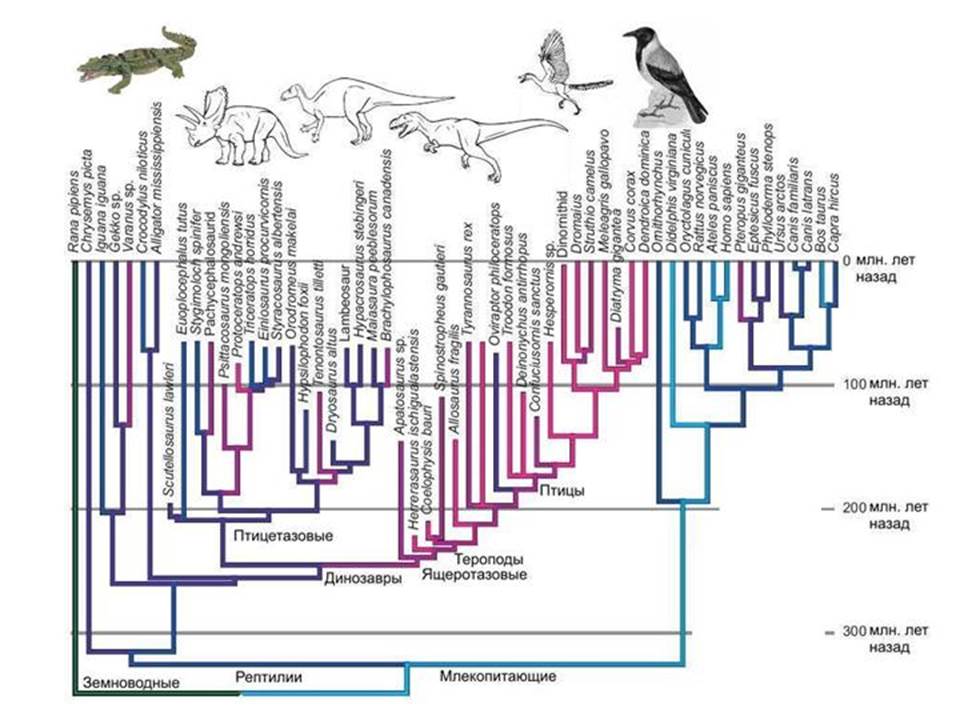
Paukščiai ir dinozaurai. Protoeukariotus atkurti sunku. Akivaizdu, kad senovės prokariotų grupė, iš kurios atsirado branduolys ir citoplazma, turėjo nemažai unikalių savybių, kurių iki šių dienų išlikę prokariotai neturi. O kai bandome rekonstruoti šio protėvio išvaizdą, susiduriame su tuo, kad hipotezių erdvės pasirodo per didelės.
Analogija. Yra žinoma, kad paukščiai kilo iš dinozaurų, o ne iš kai kurių nežinomų dinozaurų, o iš visiškai tam tikra grupė- Maniraptor dinozaurai, priklausantys teropodams, o teropodai, savo ruožtu, yra viena iš driežų dinozaurų grupių. Buvo rasta daug pereinamųjų formų tarp neskraidančių dinozaurų ir paukščių.
Bet ką galėtume pasakyti apie paukščių protėvius, jei nebūtų iškasenų? IN geriausiu atveju sužinotume, kad artimiausi paukščių giminaičiai – krokodilai. Bet ar galėtume atkurti tiesioginių paukščių protėvių, tai yra, dinozaurų, išvaizdą? Vargu ar. Bet kaip tik tokioje padėtyje atsiduriame, kai bandome atkurti branduolio ir citoplazmos protėvio išvaizdą. Akivaizdu, kad tai buvo kai kurių prokariotinių dinozaurų grupė, išnykusi grupė, kuri, skirtingai nei tikri dinozaurai, nepaliko ryškių pėdsakų geologiniuose įrašuose. Šiuolaikinė archėja eukariotų atžvilgiu yra panaši šiuolaikiniai krokodilai paukščių link. Pabandykite atkurti dinozaurų struktūrą pažindami tik paukščius ir krokodilus.
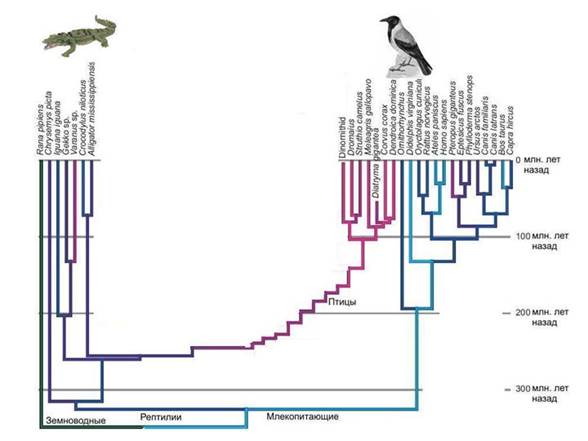
Argumentas už tai, kad Prekambrijoje gyveno daug mikrobų, nepanašių į dabartinius. Proterozojaus stromatolitai buvo daug sudėtingesni ir įvairesni nei šiuolaikiniai. Stromatolitai yra mikrobų bendruomenių gyvybinės veiklos produktas. Ar tai nereiškia, kad proterozojaus mikrobai taip pat buvo įvairesni nei šiuolaikiniai, o daugelis proterozojaus mikrobų grupių tiesiog neišliko iki šių dienų?

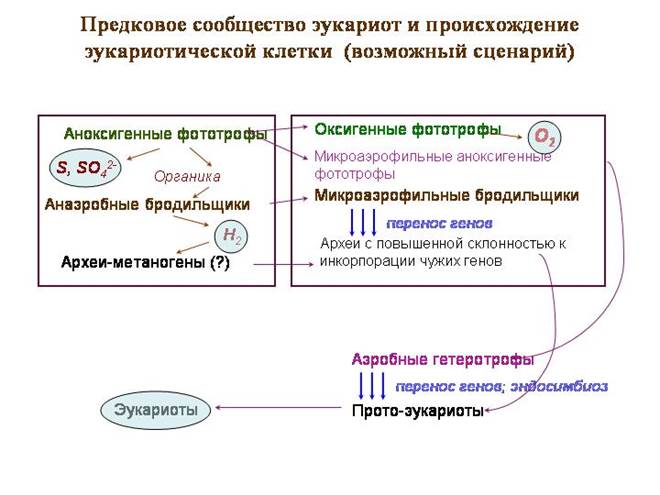
Eukariotų protėvių bendruomenė ir eukariotinės ląstelės kilmė (galimas scenarijus)
Hipotetinė „protėvių bendruomenė“ yra tipiškas bakterijų kilimėlis, tik jo viršutinėje dalyje gyveno melsvadumblių protėviai, kurie dar nebuvo perėję į deguonies fotosintezę. Jie užsiėmė anoksigenine fotosinteze.Elektronų donoras buvo ne vanduo, o sieros vandenilis. Kaip šalutinis produktas išsiskyrė siera ir sulfatai.
Antrajame sluoksnyje gyveno purpurinės fotosintetinės bakterijos, įskaitant alfaproteobakterijas, mitochondrijų protėvius. Violetinės bakterijos naudoja ilgos bangos šviesą (raudoną ir infraraudonąją). Šios bangos turi geriausią prasiskverbimo galią. Violetinės bakterijos vis dar dažnai gyvena po cianobakterijų sluoksniu. Violetinės alfaproteobakterijos taip pat naudoja vandenilio sulfidą kaip elektronų donorą.
Trečiame sluoksnyje buvo fermentuojančios bakterijos, kurios apdorojo organines medžiagas; kai kurie iš jų kaip atliekas išmetė vandenilį. Taip buvo sukurta bazė sulfatus redukuojančioms bakterijoms. Gali būti ir metanogeninių archėjų. Tarp čia gyvenusių archejų buvo branduolio ir citoplazmos protėviai.
Kriziniai įvykiai prasidėjo cianobakterijoms perėjus į deguonies fotosintezę. Kaip elektronų donoras, vietoj vandenilio sulfido buvo pradėtos naudoti cianobakterijos grynas vanduo. Tai atvėrė puikias galimybes, bet taip pat Neigiamos pasekmės. Vietoj sieros ir sulfatų fotosintezės metu pradėjo išsiskirti deguonis – medžiaga, kuri yra itin toksiška visiems senovės žemės gyventojams.
Pirmieji su šiais nuodais susidūrė jo gamintojai – melsvadumbliai. Jie bene pirmieji sukūrė apsaugos nuo jos priemones. Elektronų transportavimo grandinės, kurios tarnavo fotosintezei, buvo modifikuotos ir pradėjo tarnauti aerobiniam kvėpavimui. Pirminis tikslas, matyt, buvo ne gauti energijos, o tik neutralizuoti deguonį.
Netrukus ir antrojo bendruomenės sluoksnio – purpurinių bakterijų – gyventojai turėjo sukurti panašias gynybines sistemas. Kaip ir cianobakterijos, jos sukūrė aerobinio kvėpavimo sistemas, pagrįstas fotosintezės sistemomis. Būtent purpurinės alfaproteobakterijos sukūrė tobuliausią kvėpavimo grandinę, kuri dabar funkcionuoja eukariotų mitochondrijose.
Trečiame bendruomenės sluoksnyje laisvo deguonies atsiradimas turėjo sukelti krizę. Naudojami metanogenai ir daugelis sulfatų reduktorių molekulinis vandenilis hidrogenazės fermentų pagalba. Tokie mikrobai negali gyventi aerobinėmis sąlygomis, nes deguonis slopina hidrogenazes. Daugelis bakterijų, gaminančių vandenilį, savo ruožtu neauga aplinkoje, kurioje nėra jį naudojančių mikroorganizmų. Iš fermentatorių bendruomenė, matyt, išlaikė formas, kurios kaip galutinius produktus išskiria mažai organinių junginių (piruvato, laktato, acetato ir kt.). Šie fermentatoriai sukūrė savo apsaugos nuo deguonies priemones, ne tokias veiksmingas. Tarp išgyvenusiųjų buvo archajų – branduolio ir citoplazmos protėvių.
Galbūt šiuo krizės momentu įvyko esminis įvykis - susilpnėjo eukariotų protėvių genetinė izoliacija ir prasidėjo aktyvus svetimų genų skolinimasis. Protoeukariotai įtraukė įvairių fermentatorių genus, kol patys tapo mikroaerofiliniais fermentatoriais, fermentuojančiais angliavandenius iki piruvato ir pieno rūgšties.
Trečiojo sluoksnio gyventojai – eukariotų protėviai – dabar tiesiogiai bendravo su naujaisiais antrojo sluoksnio gyventojais – aerobinėmis alfaproteobakterijomis, išmokusiomis energijai panaudoti deguonį. Protoeukariotų ir alfaproteobakterijų metabolizmas tapo vienas kitą papildantis, o tai sudarė prielaidas simbiozei. Ir pati alfaproteobakterijų vieta bendruomenėje (tarp viršutinio sluoksnio, išskiriančio deguonį, ir apatinio sluoksnio) lėmė jų, kaip eukariotų protėvių „gynėjų“ nuo deguonies pertekliaus, vaidmenį.
Tikėtina, kad protoeukariotai prarijo ir įgijo daug įvairių bakterijų kaip endosimbiontų. Tokio pobūdžio eksperimentai vis dar vyksta. vienaląsčiai eukariotai, kurie turi didžiulę tarpląstelinių simbiontų įvairovę. Iš šių eksperimentų aljansas su aerobinėmis alfaproteobakterijomis pasirodė esąs sėkmingiausias.
Biologija ir genetika
Pagal simbiotinę hipotezę, kad šiuo metu populiarios eukariotinės ląstelės korpuskulinės organelės, turinčios savo genomą, pasižymi nepriklausoma kilme ir yra kilę iš prokariotinės ląstelės simbiontai. Iš pradžių, viena vertus, informacijos kiekis ląstelės šeimininkės genomuose ir, kita vertus, numanomų centriolių ir chloroplastų mitochondrijų simbiontuose, buvo panašus. Ateityje simbiontų genomai gali prarasti dalį savo genetinių funkcijų, kai genų blokai patenka į genomą ...
Eukariotinių ląstelių kilmės hipotezės.
1. Simbiotinis.
Pagal šiuo metu populiarią simbiotinę hipotezę, eukariotinės ląstelės korpuskulinės organelės, turinčios savo genomą, pasižymi savarankiška kilme ir yra kilusios iš prokariotinių simbiontinių ląstelių. Manoma, kad šeimininko ląstelė buvo anaerobinis prokariotas, galintis judėti tik ameboidais. Mitochondrijos ir žvyneliai atsirado pasikeitus šeimininko ląstelės simbiontams aerobiniams prokariotams ir bakterijoms, panašioms į šiuolaikines spirochetas. Žaliųjų augalų ląstelių chloroplastai atsirado iš simbiontų - melsvadumblių prokariotinių ląstelių. Ypatingą reikšmę turėjo tai, kad ląstelė evoliucijos metu įgijo žvynelius, kurių baziniai kūnai buvo arti centriolių. Tai leido sustiprėti motorinė veikla ir sukurti mitozės mechanizmą.
Branduolio kilmės klausimas yra sunkus. Iš pradžių informacijos kiekis ląstelės-šeimininkės genomuose, viena vertus, ir numanomų mitochondrijų, centriolių ir chloroplastų simbiontuose, kita vertus, buvo panašus. Ateityje simbiontų genomai gali prarasti dalį savo genetinių funkcijų, genų blokams perkeliant į ląstelės šeimininkės genomą. Tai lydėjo branduolinio genomo tūrio padidėjimas, ląstelių funkcijų genetinio reguliavimo centralizavimas ir mitochondrijų bei kitų korpuskulinių organelių autonomijos praradimas. Tačiau negalima atmesti, kad branduolys taip pat yra tarpląstelinio simbionto darinys. Šiurkštaus ir lygaus citoplazminio tinklo intraplazminių membranų, sluoksninio komplekso ir pūslelių-vakuolinių struktūrų kilmė paaiškinama evoliucinėmis išorinės membranos transformacijomis. branduolinis apvalkalas. Simbiotinė hipotezė nepaaiškina daugelio faktų. Taigi baltymas bacilinas, sudarantis šiuolaikinių prokariotų blakstienas ir žvynelius, skiriasi nuo eukariotų tubulino baltymo. Bakterijos neturėjo „9 + 2“ arba „9 + 0“ mikrotubulių sudėties struktūrų, būdingų eukariotinių ląstelių žvyneliams, blakstienoms, baziniams kūnams ar centriolėms.
2. Invaginacija.
Invaginacijos hipotezė apie eukariotinės ląstelės kilmę kyla iš to, kad protėvių forma buvo aerobinis prokariotas. Jame buvo keli genomai, prie kurių kiekvienas buvo prijungtas ląstelės membrana. Korpuskulinės organelės su DNR, taip pat branduolys, atsirado dėl apvalkalo fragmentų invaginacijos ir surišimo kartu su genomu, o po to įvyko funkcinė specializacija į branduolį, mitochondrijas, chloroplastus, branduolinio genomo komplikacija ir citoplazminių membranų vystymasis. Ši hipotezė patenkinamai paaiškina 2 membranų buvimą branduolio, mitochondrijų ir chporoplastų apvalkale. Ji susiduria su sunkumais paaiškindama baltymų biosintezės proceso detalių skirtumus korpuskulinėse organelėse ir eukariotinės ląstelės citoplazmoje. Mitochondrijose ir chloroplastuose šis procesas tiksliai atitinka šiuolaikinėse prokariotinėse ląstelėse vykstantį procesą. Simbiotinės ir invaginacijos hipotezės neišsemia visų požiūrių į eukariotinio tipo ląstelių organizacijos kilmę.
3. Daugialąstės atsiradimas.
Heterotrofinių pirmuonių kolonijinės formos buvo daugialąsčių organizmų protėviai.
1 etapas: šalia kiekvieno žvynelio buvo citoplazmos atauga apykaklė arba kišenė, į kurią buvo įstumtos maisto dalelės (fagocitozė).
2 etapas: ląstelių specializacija fagocitai (fagocitinė-virškinimo funkcija) ir kinocitai (funkcijos judėjimas; liko periferijoje). Tarpt. sluoksnis fagocitoblastas + išorinis kinocitoblastas = fagocitella arba parenchimelė (Mechnikovas).
3 etapas: viename poliuje susidaro burna. skylė => atsirado kempinės, koelenteratai ir kt.
Anot Haeckel: rutulys.jūros kolonija, užpildant skystį, susidaro burbulas panašus organizmas (blastea) invaginacija – dvisluoksnė būtybė (gastrea); (Haeckel manė, kad pirmasis buvo pilvo virškinimas).
4. Daugialąsčio ypatumai.
Daugialąsčiai gyvūnai sudaro didžiausią gyvų organizmų grupę planetoje, kurioje yra daugiau nei 1,5 milijono rūšių. Pradedant savo kilmę iš paprasčiausių, jie patyrė reikšmingų pokyčių evoliucijos procese, susijusiame su organizacijos komplikacija.
Vienas iš svarbiausių daugialąsčių organizmų organizavimo bruožų yra jų kūno ląstelių morfologinis ir funkcinis skirtumas. Evoliucijos eigoje panašios daugialąsčių gyvūnų kūno ląstelės specializavosi atlikdamos tam tikras funkcijas, dėl kurių susiformavo audiniai.
Derinami įvairūs audiniai prie organų ir organų į organų sistemas.Įgyvendinti tarpusavio santykius ir koordinuoti jų darbą,reguliavimo sistemosnervų ir endokrininės sistemos. Nervų dėka humoralinis reguliavimas visų sistemų veikla, daugialąstelis organizmas veikia kaip vientisa biologinė sistema.
Daugialąsčių gyvūnų grupės klestėjimas yra susijęs su komplikacija anatominė struktūra Ir fiziologines funkcijas. Taigi, kūno dydžio padidėjimas paskatino vystymąsi virškinimo kanalas, kuri leido jiems valgyti didelę maisto medžiagą, tiekiančią didelį kiekį energijos visiems gyvybės procesams įgyvendinti. Išsivysčiusios raumenų ir skeleto sistemos užtikrino organizmų judėjimą, tam tikros kūno formos palaikymą, apsaugą ir palaikymą organams. Gebėjimas aktyvus judėjimas leido gyvūnams ieškoti maisto, rasti pastogę ir įsikurti.
Didėjant gyvūnų kūno dydžiui, atsirado poreikis išvaizdaiintratransportinės kraujotakos sistemos,gyvybei palaikyti nutolusiems nuo kūno paviršiaus audiniams ir organams maistinių medžiagų, deguonies, taip pat pašalinant galutinius medžiagų apykaitos produktus.
Tokia kraujotakos transporto sistema tapo skystu audinių krauju.
Kvėpavimo aktyvumo intensyvėjimas vyko lygiagrečiai su laipsnišku vystymusinervų sistema ir jutimo organai. Buvo judėjimas centriniai skyriai nervų sistemą iki priekinio gyvūno kūno galo, dėl ko galvos dalis tapo izoliuota. Tokia gyvūno priekinės kūno dalies struktūra leido jam gauti informaciją apie pokyčius aplinką ir tinkamai į juos reaguoti.
Daugialąsčiai gyvūnai yra nepaprastai įvairios sandaros, gyvenimo ypatybių, skiriasi dydžiu, kūno svoriu ir kt.
Pamoka atitinka programą ir vadovėlį I.N. Ponomareva, kurios numato citologijos studijas 11 klasėje. Pamoka vyksta mokslinės diskusijos forma ir yra skirta 90 minučių.
Pamokos tikslai: susipažinimas su svarbiausios hipotezės apie eukariotinės ląstelės ir daugialąsčių organizmų kilmę; gebėjimo ginčytis ir apginti savo požiūrį ugdymas; viešosios diskusijos kultūros puoselėjimas.
Įranga: lentelės, kuriose pavaizduota ląstelės sandara ir atskiros jos organelės, eukariotinės ląstelės atsiradimo diagrama, piešiniai, vaizduojantys trichoplaksą ir įvairias dumblių formas.
UŽSIĖMIMŲ LAIKOTARPIU
Mokytojas. Tobulėjant mokslui apie ląstelę – citologiją ir atsirandant naujiems šio elementoriaus tyrimų rezultatams biologinė sistema tarp mokslininkų kilo karštos diskusijos, peraugusios į rimtas mokslines diskusijas. Per visą citologijos kaip mokslo istoriją beveik bet kuri nauja hipotezė susilaukė ir šalininkų, ir priešininkų.
Mokslinė diskusija – tai viešas klausimo ar problemos aptarimas. Šiandien pamokoje turėsime diskusiją, kurios metu aptarsime du kritiniais klausimais citologija: pirmųjų eukariotų kilmės problema ir pirmųjų daugialąsčių organizmų kilmės problema.
Jūs žinote, kad klasė anksčiau buvo suskirstyta į grupes, kurios pristatys ir gins skirtingus požiūrius šiais dviem klausimais, kuriuos aptariame. Leiskite jums priminti šių grupių pavadinimus.
1. Eukariotinės ląstelės ir jos organelių kilmės autogeninės hipotezės šalininkai.
2. Simbiogeninės eukariotinės ląstelės ir jos organelių kilmės hipotezės šalininkai.
3. Pirmųjų vienaląsčių organizmų kolonijinės kilmės hipotezės šalininkai.
4. Daugialąsčių organizmų atsiradimo iš blakstienų, jų ląstelių ląstelėjimo, šalininkai.
Kiekvienos grupės atstovai trumpame pristatyme išdėstys vienokio ar kitokio požiūrio į kiekvieną iš aptartų problemų esmę, o vėliau atsakys tiek į savo mokslinių „oponentų“, tiek į visų klasėje dalyvaujančių klausimus, pastabas, prieštaravimus.
Kilmės problema membranos organelės o eukariotinių ląstelių branduoliai yra viena iš aktualiausių šiuolaikinės citologijos diskusijų temų. Norėdami išspręsti šią problemą, moksle buvo pasiūlytos kelios skirtingos hipotezės. Pažvelkime į du svarbiausius.
Autogeninė eukariotinės ląstelės kilmės hipotezė
Pirmosios grupės mokinių atstovų kalba
Pirmieji eukariotai Žemėje pasirodė prieš 1,1–1,4 milijardo metų (proterozojaus eroje). Tai buvo kitas svarbus Prekambrijos evoliucijos etapas po fotosintezės atsiradimo.
Autogeninė, arba nuoseklioji, hipotezė teigia, kad sudėtinga eukariotinė ląstelė išsivystė tiesiai iš prokariotinės ląstelės: dėl plazminės membranos invaginacijos, po kurios ji persitvarkė atlikti tam tikras funkcijas, atsirado svarbiausios organelės. Tokios sudėtingos organelės kaip mitochondrijos ir plastidai yra kilę iš prokariotuose esančios vamzdinės struktūros intraplazminių membranų struktūrų.
Klausimai
1. Kas palaiko šį požiūrį? Ar yra pereinamųjų formų, t.y. prokariotinės ląstelės su eukariotinių organelių „rudimentais“?
Atsakymas. Tarp gyvų organizmų tokių reliktų formų nėra. Tačiau prisiminkime mezosomų buvimą bakterijų ląstelėse, t.y. vamzdinės ir vezikulinės (sferinės) formos membraninės struktūros, susidarančios plazminei membranai įsiskverbus į citoplazmą. Daroma prielaida, kad mezosomos dalyvauja formuojantis ląstelių sienelėms, DNR replikacijai ir kituose procesuose. Visai įmanoma, kad būtent mezosomų pagrindu pamažu atsirado ir diferencijuodavosi membranų kompleksas, sukeldamas įvairius eukariotinės ląstelės organelius.
2. Yra žinoma, kad daugelio organelių, ypač mitochondrijų ir chloroplastų, biocheminė sudėtis labai skiriasi nuo citoplazmos plazminės membranos sudėties. Ar tai nereiškia, kad jų kilmė nėra iš originalo ląstelės membrana- plazminės membranos?
Atsakymas. Iš tiesų mitochondrijų ir chloroplastų baltymų sudėtis yra unikali. Tačiau šį originalumą galima įgyti antrą kartą, tai gali būti šių organelių prisitaikymo prie tam tikrų funkcijų atlikimo rezultatas.
Mokytojas. Išklausykime simbiogeninės eukariotų kilmės šalininkų argumentus ir išsiaiškinkime, kuri hipotezė klasėje populiaresnė.
Simbiogeninė eukariotinės ląstelės kilmės hipotezė
Antros grupės mokinių atstovų kalba
Simbiogeninė hipotezė (dabar dažnai vadinama teorija) kyla iš to, kad eukariotai yra skirtingų prokariotų simbiozės rezultatas.
Jau XX amžiaus pradžioje. Rusijos botanikai A.S. Famintsyn, B.M. Kozo-Polyansky ir K.S. Merežkovskis iškėlė hipotezę, kad žaliųjų augalų (eukariotų) ląstelė gavo plastidų dėl chlorofilo neturinčios ląstelės simbiozės su mėlynai žalių dumblių ląstelėmis. Ši hipotezė apie simbiogeninę eukariotinės ląstelės kilmę pralenkė savo laiką, buvo pamiršta ir vėl patraukė dėmesį XX amžiaus viduryje.
Pavadintą hipotezę sukūrė Lynn Sagan-Margulis (1983). Remiantis šia hipoteze, didelės prokariotinės bakterijos pirminė ląstelė, patekusi į simbiozę su melsvadumblių ląstelėmis, įgijo plastidų. Simbiozė su heterotrofinėmis prokariotinėmis ląstelėmis paskatino jų transformaciją į mitochondrijas. Kai kurios ląstelės, būdamos heterotrofos, užfiksavo kitas, mažesnes bakterijas, kurias jos nežinomų priežasčių nesuvirškino. Pagautos mažos prokariotinės ląstelės sugebėjo absorbuoti deguonį. Ši savybė buvo naudinga ląstelei, kuri apėmė bakteriją, nes suteikė jai daug daugiau energijos, sukauptos ATP molekulėse.
Klausimai
1. Kokie įrodymai pateikiami plastidžių ir mitochondrijų simbiogeninės kilmės naudai?
Atsakymas.Šios nuomonės įrodymai yra tokie.
1. Mitochondrijas ir chloroplastus gaubia dviguba membrana.
2. Šios organelės dauginasi dalijantis ir neatsiskiria nuo kitų membraninių organelių.
3. Mitochondrijos ir chloroplastai turi savo genetinę medžiagą, kuri koduoja tik jų pačių baltymus.
4. Šių organelių DNR yra apskritos struktūros, kaip ir prokariotų, o ne linijinė, kaip eukariotų.
5. Mitochondrijos ir chloroplastai turi savo RNR ir baltymų sintezės mašinas, o jų ribosomos yra labiau prokariotinės nei eukariotinės.
6. Kai kurie šių organelių baltymai savo pirmine struktūra yra panašūs į panašius bakterijų baltymus ir nėra panašūs į atitinkamus citoplazmos baltymus.
2. Yra žinoma, kad ne visi mitochondrijų ir chloroplastų baltymai sintetinami pagal jų pačių DNR, dalis šių organelių baltymų yra užkoduoti branduolio DNR. Kaip galite paaiškinti šį faktą?
Atsakymas. Iš tiesų, tik dalis jų baltymų yra užkoduota mitochondrijų ir chloroplastų DNR, o likusieji yra užkoduoti ląstelės branduolio DNR. Matyt, evoliucijos eigoje dalis genetinės medžiagos „ištekėjo“ iš mitochondrijų ir chloroplastų genomų į branduolio genomą. Chloroplasto genomas yra didesnis nei mitochondrijų genomas, o žemesniųjų eukariotų (pavyzdžiui, mielių) mitochondrijų genomas yra didesnis nei žinduolių, o tai netiesiogiai patvirtina pirmiau minėtą prielaidą.
3. Žinome, kad, be mitochondrijų ir chloroplastų, nedidelis DNR kiekis eukariotų ląstelėje taip pat yra centriolėse ir žvynelių bazėje. Ar jie taip pat atsirado simbiogeniškai?
Atsakymas. Taip. Simbiozė su į spirochetas panašiomis bakterijomis gali lemti žvynelių atsiradimą ir, kita vertus, kai kurių į spirochetas panašių ląstelių transformaciją į centrioles. Šiuolaikinė simbiogenezės koncepcija teigia, kad eukariotinė ląstelė susiformavo dėl kelių vienas po kito einančių simbiogenezės veiksmų.
4. Kaip atsirado likusios eukariotinės ląstelės organelės (Golgi kompleksas, EPS, lizosomos)?
Atsakymas. Daroma prielaida, kad endoplazminis tinklas, lizosomos ir Golgi aparatas galėjo atsirasti dėl plazmos membranos invaginacijos.
5. Kaip, remiantis simbiogenine hipoteze, galėjo atsirasti branduolys, apsuptas dvigubos membranos?
Atsakymas. Daroma prielaida, kad branduolys taip pat išsivystė dėl DNR išskyrimo iš citoplazmos, invaginuojant plazmos membraną ir apgaubiant branduolinę medžiagą.
6. Iš jūsų atsakymų į paskutinius du klausimus matyti, kad eukariotinės ląstelės atsiradimas nėra tik kelių prokariotų paveldimos simbiozės rezultatas. Ką tu į tai pasakysi?
Atsakymas. Taip tai yra. Simbiogeninės hipotezės šalininkai pripažįsta, kad kai kurios eukariotinės ląstelės struktūros atsirado dėl jos savaiminio vystymosi ir komplikacijų.
7. Atsinešėte biocheminius, genetinius, elektronų mikroskopinius duomenis Pastaraisiais metais eukariotinės ląstelės simbiogenetinės kilmės naudai. Tačiau ar yra evoliucinių reliktų, vadinamųjų gyvųjų fosilijų, rodančių ryšius tarp prokariotų ir eukariotų?
Atsakymas. Tokie ryšiai pastebimi tarp augalų ir gyvūnų organizmų. Pavyzdžiui, laisvai gyvenanti ameba Pelomyxa palustris(Pelomyx pelkė) neturi mitochondrijų, tačiau turi simbiotinių bakterijų, kurios suteikia jai kvėpavimą.
Mokytojas. Man atrodo, kad antros grupės atstovams buvo užduota pakankamai klausimų. Kokios hipotezės šalininkai, Jūsų nuomone, išsakė įtikinamesnius argumentus ir kokia hipotezė yra labiau pripažinta moksle?
Iš tiesų, simbiogeninė hipotezė suranda daugybę šalininkų. Tačiau reikia atsiminti, kad simbiogeninės hipotezės naudai išreikšti samprotavimai neturi eksperimentinio patvirtinimo, todėl jos negalima laikyti teorija. Mano nuomone, taip pat svarbu prisiminti, kad simbiozė suvaidino svarbų, bet ne vienintelį vaidmenį eukariotinės ląstelės atsiradime. Svarba formuojantis eukariotams, vyko ir prokariotinių ląstelių savaiminio vystymosi procesai.
Pereikime prie kita problema- daugialąsčių organizmų kilmės problema.
Pirmųjų metazoanų kolonijinės kilmės hipotezės
Mokytojas. Citologijos ir molekulinės biologijos raidos pažanga įrodė vienaląsčių ir daugialąsčių organizmų struktūrinį ir biocheminį ryšį. Visa tai patvirtino daugialąsčių gyvūnų kilmės hipotezę iš vienaląsčių formų. Pagrindiniai moksliniai ginčai yra susiję su klausimais, iš kurių pirmuonių atsirado daugialąsčiai gyvūnai ir kaip atrodė pirmieji daugialąsčiai gyvūnai? Yra daug hipotezių apie daugialąsčių gyvūnų kilmę iš pirmuonių. Panagrinėkime kai kuriuos iš jų.
Trečios grupės mokinių atstovų pasisakymas
Teisingai pastebėta, kad yra daug hipotezių apie daugialąsčių gyvūnų kilmę iš pirmuonių. Labiausiai argumentuota iš jų, mūsų nuomone, turėtų būti laikoma kolonijine hipoteze.
Pirmąją kolonijinę daugialąsčių organizmų kilmės hipotezę pasiūlė Ernstas Haeckelis. Jis sukūrė savo „gastrėjos teoriją“, pagal kurią bendras visų daugialąsčių gyvūnų protėvis panašus į laisvai plaukiančią koelenteratų lervą – koralus su dviem ląstelių sluoksniais.
Rusų biologas Ilja Iljičius Mečnikovas iškėlė kitą hipotezę: daugialąsčiai gyvūnai atsirado iš kolonijinių žvynelių pirmuonių, galinčių fagocituoti, t.y. maisto paėmimas ląstelėje su tarpląsteliniu virškinimu. Šioje teorijoje daroma prielaida, kad paviršinės ląstelės, paėmusios maistą, giliai nugrimzta į kolonijos kūną, kad būtų virškinamos, todėl atsiranda vietos alkanoms ląstelėms. Rezultatas yra organizmas, kurio vidinio sluoksnio ląstelės virškina maistą, o išorinis sluoksnis, susidedantis iš alkanų ląstelių, jį sugauna. Taip pat atlieka priėmimo, judėjimo ir apsaugos funkcijas. Mechnikovas siūlomą organizmą pavadino fagocetella (organizmas, susidedantis iš ląstelių, dalyvaujančių fagocitoze).
Galiausiai trečiąją hipotezę iškėlė vokiečių zoologas O. Byuchli. Anot jos, pirminis daugialąstelis organizmas susidėjo iš dviejų ląstelių sluoksnių, be to, apatine puse jis šliaužė žeme ir viršutinė pusė turėjo apsaugines ir jautrias funkcijas. Sutikęs didelių dydžių maistą, toks organizmas apsivyniojo aplink jį ir suvirškino jį apatiniu sluoksniu. Bütschli šį tariamą organizmą pavadino placula (gyvūnas gaubiančio pyrago pavidalu).
Klausimai
1. Kuriai iš šių trijų hipotezių teikiate pirmenybę? Kuris iš jų yra labiausiai pripažintas moksle?
Atsakymas. Sunku teikti pirmenybę kokiai nors hipotezei, nes nėra paleontologinių duomenų apie pirmuosius daugialąsčių gyvūnų evoliucijos žingsnius. Mokslininkai bando rasti patvirtinimą hipotezėms apie daugialąsčių gyvūnų kolonijinę kilmę, lygindami hipotetinius organizmus su gyvomis primityviomis daugialąsčių gyvūnų formomis.
Primityviausias šiandien žinomas daugialąstis gyvūnas yra Trichoplax (1 pav.). Tai aprašė Artemijus Vasiljevičius Ivanovas (1973). Trichoplax buvo rastas Europos Atlanto vandenyno pakrantėje. Šis gyvūnas atrodo kaip plona plokštelė iš dviejų sluoksnių ląstelių su žvyneliais. Jo kūno ertmėje yra atskiros virškinimo ląstelės. Tarp dviejų ląstelių sluoksnių yra ląstelės, kurios atrodo kaip amebos.
Išstudijavęs Trichoplax, A.V. Ivanovas padarė išvadą, kad ji labai panaši į hipotetinę fagocetelą I.I. Mechnikovas ir iš tikrųjų yra jo gyvas modelis. Kita vertus, Trichoplax taip pat atrodo kaip plokštelė. Taigi, atradus Trichoplax, panašų į koelenteratų lervą, sujungiamos Haeckel (1866), Mechnikovo (1877) ir Buchli (1884) hipotezės.
2. Ilgą laiką primityvių daugialąsčių gyvūnų kilmė buvo siejama su žaliosiomis kolonijinėmis vėliavėlėmis, tokiomis kaip eudorina, volvox, pandorina. Ar dabar jie nėra laikomi formomis, pereinančiomis į daugialąstį?
Atsakymas. Iš tiesų, daugialąsčių gyvūnų kilmė nebėra siejama su žaliomis kolonijinėmis žvyneliais. Šis požiūris pagrįstas tuo, kad jų individualus vystymasis ir dauginimasis yra artimi dumblių, o ne primityvių daugialąsčių gyvūnų.
3. Kokie primityvūs daugialąsčiai gyvūnai sukėlė hipotetinį kolonijinį organizmą? Ar tai buvo kempinės, koelenteratai ar kokio nors kito gyvūno atstovai?
Atsakymas. Daroma prielaida, kad iš į fagocetellas panašių protėvių vienu metu kilę kelių rūšių daugialąsčiai gyvūnai, turintys skirtingą organizuotumo lygį: kempinės, koelenteratai ir primityvūs trisluoksniai gyvūnai, artimi plokščiosioms kirmėlėms priklausančioms nežarnyno planarijoms. Kempinės ir koelenteratai yra aklavietės evoliucijos šakos. Siaura šių tipų specializacija, prisitaikymas prie nejudančio ar pasyvaus gyvenimo būdo uždarė galimybes toliau progresuoti.
4. Vis kalbate apie pirmųjų daugialąsčių gyvūnų kilmę, tačiau daugialąstis būdingas ne tik gyvūnų karalystės atstovams. Kaip gali būti daugialąsčiai grybai, augalai?
Atsakymas.Šiuolaikinis mokslas mano, kad augalai neturi ryškios ribos tarp vienaląsčių ir daugialąsčių. Daugialąstis ne kartą atsirado nepriklausomai vienas nuo kito skirtinguose augalų skyriuose. Taigi auksiniuose, daugiasluoksniuose, pirofitiniuose, žaliuosiuose dumbliuose stebimos rūšys, turinčios siūlinę, kolonijinę daugialąstę ir sifoninę struktūrą (kūną sudaro viena daugiabranduolė milžiniška ląstelė). Tokių formų buvimas, mūsų nuomone, rodo bandymus prasibrauti į kitą organizacijos lygmenį – daugialąstį, kur veikia skirtingos ląstelės. įvairios funkcijos(2 pav.).
Ryžiai. 2. Įvairios formos dumbliai (1 - vienaląsčiai;
2 - kolonijinis; 3 - kelių branduolių; 4 - siūlinis)
Šiuolaikiniam mokslui prieinamos informacijos apie daugialąsčių formų vystymąsi Prekambrijoje yra nedaug. Tai ypač pasakytina apie grybus. Priežastis – prastas šių organizmų liekanų išsaugojimas. Kaip ir daugelyje augalų, grybai pereina iš neląstelinės struktūros (sifoninės, kaip, pavyzdžiui, pelėsinio grybelio gleivinėje) prie daugialąsčių. Taigi, mūsų nuomone, tam tikrame grybų vystymosi etape (maždaug prieš 1,3 milijardo metų) kai kuriuose iš jų sifoninis grybiena pavirto į daugialąstį darinį.
Mokytojas. Apibendrinkime tai, kas buvo pasakyta. Daugialąsčiai organizmai, pasak šiuolaikinis mokslas, anksčiau atsirado tarp augalų ir grybų. Gyvūnuose pirmasis daugialąsčių formų atsirado maždaug prieš 700 milijonų metų. Daugialąsčių gyvūnų pradžią davė senovės heterotrofiniai kolonijiniai žvyneliai.
Pirmųjų metazoanų nekolonijinės kilmės hipotezė
Ketvirtos grupės mokinių atstovų kalba
Ankstesnės grupės atstovai tvirtina, kad tolimi daugialąsčių gyvūnų protėviai buvo pirmuonių kolonijos. Nors tai įprastas, bet ne vienintelis požiūris.
Pateikiame hipotezę, leidžiančią manyti, kad evoliucijos procese pavieniai pirmuonys buvo visiškai transformuoti į daugialąsčius tvarinius. Šią idėją iškėlė garsus serbų zoologas I. Hadžis. Jo nuomone, daugialąsčiai gyvūnai išsivystė iš daugiabranduolių blakstienų.
Blakstienos turi gana sudėtingą struktūrą. Jų citoplazmą vaizduoja du sluoksniai - periferinis ir centrinis, kuriame vyksta tarpląstelinis virškinimas. Blakstienos turi ląstelinę burną, ryklę, susitraukiančios vakuolės su pritraukiamaisiais kanalais. Visos šios įvairiai diferencijuotos dalys vienaląstis organizmas- organelės - Hadji laiko daugialąsčio gyvūno organų prototipu. Taip, jis taip galvoja oda daugialąsčiai būtybės atsirado iš išorinio citoplazmos sluoksnio (ektoplazmos), o jų žarnos – iš vidinio citoplazmos sluoksnio (entoplazmos).
Perėjimas iš vienaląstės į daugialąstę gali įvykti blakstienų kūne iš karto, susidarant ląstelių riboms aplink atskirus branduolius ir gretimas citoplazmos dalis. Šis tariamas procesas vadinamas „ląstelėjimu“ (iš lotyniško žodžio ląstelė- ląstelė), o pati hipotezė vadinama ląstelėjimo teorija.
Klausimai
1. Ar I. Hadji hipotezė turi kokį nors pagrindą žemesnių daugialąsčių organizmų embriologijoje, kaip, pavyzdžiui, I. I. hipotezė? Mechnikovas?
Atsakymas. Deja, tokių lyginamosios embriologijos duomenų nėra. Niekas niekada nepastebėjo blakstienų ląstelėjimo proceso gamtoje.
2. Ar nemanote, kad Hadži hipotezė neatlaiko kritikos, visų pirma dėl to, kad pats principas, kuriuo ji grindžiama, yra žiaurus. Ląstelėjimo hipotezė, mūsų nuomone, prieštarauja pagrindinėms ląstelių teorijos nuostatoms, būtent: ji prilygina vienos ląstelės dalis daugialąsčių organizmų audiniams ir organams. Ką galite pasakyti apie tai?
Atsakymas. Nebūtume tokie kategoriški vertindami. Hadži hipotezė blakstienų organelių netapatina su daugialąsčių organizmų audiniais ir organais. Ji tik skaičiuoja galimas išsilavinimas panašūs organai, kurių pagrindą sudaro blakstienos organelės.
3. Kuo dar remiasi Hadži hipotezė, be to, kad turi blakstienas sudėtinga struktūra ir jų struktūros galėtų tapti daugialąsčių gyvūnų organais?
Atsakymas. Netiesioginis hipotezės patvirtinimas yra tam tikras panašumas tarp blakstienų ir turbelių - primityvus plokščiųjų kirmėlių: artimi dydžiai, ta pati kūno forma, daugybės konstrukcijų vieta ir kt. (3 ir 4 pav.).
Mokytojas. Išklausėme argumentų už kolonijinę ir nekolonijinę pirmųjų daugialąsčių gyvūnų kilmę, susipažinome su pažiūromis apie daugialąsčių augalų ir grybų kilmę. Kokia, jūsų nuomone, hipotezė – daugialąsčių gyvūnų kolonijinė ar nekolonijinė kilmė – yra labiau pagrįsta ir įrodyta?
Žinoma, pirmųjų daugialąsčių gyvūnų kolonijinės kilmės hipotezė. Būtent ši hipotezė yra pripažinta plačiausiai mokslo pasaulyje. Tačiau reikia atsiminti, kad antroji hipotezė nuolat vystosi ir kinta.
Apibendrinant pamoką
1. Įvertinimas pamokai.
2. Namų darbai:
- studijuoti pastraipą „Diskusuotinos citologijos problemos“;
f – namuose susipažinti su viena iš XVIII amžiuje aptartų citologijos problemų, kuri buvo vykdoma tarp skirtingų preformizmo šalininkų; parengti argumentų sistemą skirtingų požiūrių naudai.