Kiekvienos ląstelės citoplazma yra apsupta membrana, kuri ją riboja aplinką. Citoplazminė membrana (CPM) yra išskirtinai daugiafunkcinė struktūra:
1. MUT suvokia visą cheminę informaciją patekęs į ląstelę iš išorinės aplinkos.
2. Ji yra pagrindinė osmosinis barjeras dėl kurių ląstelės viduje palaikomas tam tikras osmosinis slėgis.
Ląstelės sienelė daugiausia sudaryta iš peptidoglikano, sumaišyto su teiko rūgštimi, kuri kartu sulaiko skirtingus sluoksnius. Kryžminio susiejimo kiekis yra didesnis, o sienelė storesnė nei gramneigiamų ląstelių sienelių. Gram-teigiamos ląstelės yra sudarytos beveik vien iš peptidoglikano, bet svarbi struktūra Gramteigiamų ląstelių sienelėse randama techoinė rūgštis. Tai glicerolio arba ribitolio fosfodiesterio polimeras, susietas fosfato grupėmis. Teichoinė rūgštis yra kovalentiškai susieta su muramo rūgštimi ir teka kartu su įvairiais peptidoglikano tinklo sluoksniais.
3. MUT kartu su ląstelės sienele dalyvauja augimo ir ląstelių dalijimosi reguliavimas bakterijos.
4. CPM dalyvauja reguliuojant chromosomų ir plazmidžių replikacijos ir segregacijos procesus (jos yra susijusios su jo receptoriais).
5. MUT yra reikšminga suma fermentaiįskaitant elektronų perdavimo sistemas (TsPM - energijos gamybos vieta bakterijose).
Teichoinė rūgštis stabilizuoja ląstelės sienelę ir stiprina ją. Cheminė formulė Teicho rūgštis parodyta 25 paveiksle. Teicho rūgštis yra ilga, plona molekulė, susipynusi per peptidoglikaną. Gramneigiamų ląstelių sienelės turi daugiau sudėtinga struktūra nei gramteigiamų organizmų. Už citoplazminės membranos yra periplazma, kurioje yra plonas peptidoglikano sluoksnis. Peptidoglikane gramneigiamose ląstelėse yra mažiau kryžminio ryšio nei gramteigiamose ląstelėse be peptidų jungiklio.
6. Su MUT susijusios žvyneliai ir jų judėjimo reguliavimo aparatai.
7. CPM dalyvauja procesuose transporto(įskaitant aktyvų) maistinių medžiagųį ląstelę, o atliekos, įskaitant fermentus ir egzotoksinus, iš ląstelės patenka į aplinką. Jame yra baltymų, dalyvaujančių palengvintoje difuzijoje ir aktyviame transporte.
Su peptidoglikanu kovalentiškai susietas Browno lipoproteinas, turintis hidrofobinį inkarą, kuris padeda stipriai surišti peptidoglikaną su išorine membrana. 26 paveiksle parodyta gramneigiamos ląstelės sienelės vieta. Gramneigiamų bakterijų ląstelės sienelėje yra daug mažiau peptidoglikano ir ją supa išorinė membrana. Tarp peptidoglikano yra daug mažiau kryžminių ryšių.
Gramneigiamų bakterijų išorinė membrana yra dar vienas lipidų dvisluoksnis sluoksnis, panašus į citoplazminę membraną, ir joje taip pat yra baltymų. Tai barjeras baltymams ir neleidžia nusodinti fermentams, išsiskiriantiems į periplazmą. Šis atstūmimas leidžia šioms bakterijoms išgyventi šioje aplinkoje.
8.
MUT groja svarbus vaidmuo skirstant ir stabilizuojant ribosomos.
9.
CPM dalyvauja ląstelės sienelės komponentų sintezėje.
10.
CPM dalyvauja formuojant mezosomas (mezosomos susidaro dėl CPM vietos invaginacijos į citoplazmą, jos yra atviros periplazminėje erdvėje).
Kaip veikia membrana molekulinis lygis daugybė jo funkcijų – viena iš aktualiausius klausimus šiuolaikinė biologija. CPM dalis sudaro apie 10 % sausos bakterijų masės. Jame yra 25-40 proc. fosfolipidai, sudarantys du sluoksnius, 20-75% baltymų ir iki 6% angliavandenių. molekules fosfolipidai yra asimetriški: galvos, turinčios elektros krūvį, yra hidrofilinės; uodegos yra neutralios ir hidrofobinės. Fosfolipidai yra supakuoti į membraną taip: jų poliarinės hidrofilinės galvutės yra pasuktos į išorę ir sudaro du CPM sluoksnius - vidinį ir išorinį, o nepolinės hidrofobinės uodegos yra paslėptos membranos storyje. Elektronų difrakcijos modeliuose CPM yra trijų sluoksnių struktūra, susidedanti iš dviejų 2 lygiagrečiai tamsūs sluoksniai ir juos skiriantis šviesus sluoksnis. Šis sluoksnis yra pralaidesnis elektronams nei sluoksniai, susidedantys iš su baltymais susijusių fosfolipidų poliarinių galų. CPM funkcijų specifiškumas labai priklauso nuo juose esančių baltymų rinkinio. Jų vieta CPM yra savotiška: kai kurie baltymai prasiskverbia per visą dvigubą lipidų sluoksnį, tam tikra baltymų dalis yra susijusi arba tik su vidiniu, arba tik su išorinis paviršius membranos. Tai išplaukia iš to, kad membranos ir citoplazmos sąveika, viena vertus, membrana ir išorinė aplinka o kita vertus, lemia įvairius, nors ir tarpusavyje susijusius, jo gyvybės palaikymo procesus: palengvintą difuziją, aktyvųjį transportavimą, elementarios biologinės membranos.
Jis kuria Platus pasirinkimas fiziologinės reakcijosįskaitant karščiavimo sukėlimą, baltųjų kraujo kūnelių kiekio pokyčius, nutekėjimą nuo kraujagyslės, naviko nekrozė ir sumažinta kraujo spaudimas sukeliantis kraujagyslių kolapsą ir galiausiai šoką. Išorinėje membranoje yra mažiau baltymų ir mažiau unikalių baltymų tipų nei citoplazminėje membranoje. Porins yra ypač svarbius komponentus dėl jų vaidmens išorinės membranos pralaidumui mažoms molekulėms. Porinai yra baltymai, kurie sudaro pakankamai dideles poras išorinėje membranoje, kad galėtų praeiti mažiausioms hidrofilinėms molekulėms.
Esant pernelyg dideliam augimui (palyginti su ląstelės sienelės augimu), citoplazminėje membranoje susidaro invaginacijos – invaginacijos sudėtingai susisukusių membraninių struktūrų pavidalu, vadinamos. mezosomos. Mažiau sudėtingos susuktos struktūros vadinamos intracitoplazminėmis membranomis. Mezosomų vaidmuo ir intracitoplazminės membranos nebuvo iki galo išaiškintos. Netgi teigiama, kad jie yra artefaktas, kuris atsiranda paruošus (fiksavus) preparatą elektroninei mikroskopijai. Nepaisant to, manoma, kad citoplazminės membranos dariniai dalyvauja ląstelių dalijimuisi, teikdami energiją ląstelės sienelės sintezei, dalyvauja medžiagų sekrecijoje, sporų formavime, t.y. procesuose, kuriuose sunaudojama daug energijos.
Ant pav. 28 parodyta porino struktūra molekuliniu mastu. Visi žinomi porinai turi panašią struktūrą, o baltyme yra centrinis kanalas, leidžiantis molekulėms praeiti. Tai leidžia šias molekules transportuoti į periplazminę erdvę, kad jos galėtų būti perneštos per citoplazmą. plazmos membrana. Kai kurios išorinės membranos porinai yra bendro pobūdžio, pasižymi paprasto dydžio ir krūvio atskyrimu, tačiau turi mažai specifiškumo. Kiti porinai yra konkretesni.
Labai didelės arba hidrofobinės molekulės negali prasiskverbti pro išorinę membraną, todėl išorinė membrana tarnauja kaip pralaidumo barjeras bent kai kurioms molekulėms. Porino molekulinė struktūra. Vaizdas iš vidaus yra už ląstelės ribų, žiūrint į membranos paviršių. Žvilgsnis yra perspektyva iš išorės. Porinas turi tris baltymų subvienetus, o tikroji pora yra centrinė trikampė sritis viršutinėje plokštėje, kurią sudaro trys subvienetai.
Citoplazma užima didžiąją dalį bakterijos ląstelės ir susideda iš tirpių baltymų, ribonukleino rūgščių, inkliuzų ir daugybės mažų granulių – ribosomų, atsakingų už baltymų sintezę (vertimą).
Ribosomos bakterijų dydis yra apie 20 nm ir koeficientas sedimentacija 70S, priešingai nei 8OS ribosomos, būdingos eukariotinės ląstelės. Todėl kai kurie antibiotikai jungiasi prie bakterijų ribosomų ir slopina bakterijų baltymų sintezę nepaveikdami baltymų sintezės eukariotinėse ląstelėse. Bakterijų ribosomos gali išsiskirti į du subvienetus – 50S ir 30S. Ribosominė RNR (rRNR.) – konservatyvūs bakterijų elementai (evoliucijos „molekulinis laikrodis“). 16S rRNR. yra mažojo ribosomų subvieneto dalis, o 23S rRNR yra didelio ribosomų subvieneto dalis. 16S rRNR tyrimas. yra genosistematikos pagrindas, leidžiantis įvertinti organizmų giminingumo laipsnį.
Taip pat yra ir kitų rūšių baltymų išorinė membrana kurie dalyvauja įvairios funkcijos. Kai kuriuose patogenuose yra išorinės membranos baltymų, kurie padeda jiems neutralizuoti šeimininko apsaugą. Gramneigiamų ir gramneigiamų bakterijų ląstelių sienelių skirtumai labai įtakoja mikrobų sėkmę jų aplinkoje. Stora gramneigiamų ląstelių sienelė leidžia joms geriau susidoroti sausomis sąlygomis, nes sumažina vandens praradimą. 3 lentelėje apibendrinamas skirtumas tarp gramneigiamų ir gramneigiamų ląstelių sienelių.
Citoplazmoje yra įvairių įtraukimas granulių pavidalu glikogenas, polisacharidai, beta-hidroksisviesto rūgštis ir polifosfatai (volutinas). Visos rezervinės medžiagos ląstelėse yra chemiškai inertiškos formos. Ši sąlyga neleidžia pažeisti ląstelių turinio osmostazės. Kai kurie inkliuzai tiesiog guli citoplazmoje, kiti yra apsupti plona membrana. Membrana dažniausiai yra baltyminio pobūdžio, tačiau kartais joje gali būti lipidų. Jie kaupiasi su maistinių medžiagų pertekliumi aplinkoje ir veikia kaip rezervinės medžiagos mitybos ir energijos poreikiams tenkinti.
3 lentelė Ląstelės sienelės savybės
Gramneigiamų ir gramneigiamų ląstelių sienelių skirtumų santrauka. Daugumos bakterijų ląstelių sienelė turi lemiamas ląstelių išlikimui, tačiau yra keletas bakterijų, kurios neturi ląstelių sienelių. Mikoplazmų rūšys yra dažni pavyzdžiai, o kai kurios gali būti tarpląsteliniai patogenai, augantys jų šeimininkų viduje. Ląstelių sienelės čia nereikalingos, nes ląstelės gyvena tik kontroliuojamoje kitų ląstelių osmosinėje aplinkoje.
Voliutinas turi afinitetą pagrindiniams dažams ir yra lengvai aptinkamas naudojant specialius dažymo metodus (pavyzdžiui, pagal Neisser) metachromatinių granulių pavidalu. Toluidino mėlyna arba metileno mėlyna nudažo volutiną raudonai violetiškai, o bakterijų citoplazmą – mėlynai. Būdingas volutino granulių išsidėstymas difterijos baciloje atsiskleidžia intensyviai dažytų ląstelės polių pavidalu. Metachromatinis volutino dažymas yra susijęs su didelis kiekis polimerizuotas neorganinis polifosfatas. Pagal elektronų mikroskopiją jie atrodo kaip elektronų tankios 0,1–1,0 µm dydžio granulės.
Tikriausiai jie kažkada turėjo galimybę suformuoti ląstelės sienelę, tačiau, kai jų gyvenimo būdas tapo buvimu kitose ląstelėse, jie prarado gebėjimą kurti sienas. Atsižvelgiant į šį labai ribotą kitų ląstelių gyvenimo būdą, šie mikrobai taip pat yra labai maži. Jiems nereikia visų rūšių biosintetinių fermentų genų, nes jie gali pavogti galutinius šių kelių komponentus iš šeimininko. Taip pat jiems nereikia daug koduojančių genų Skirtingi keliaiįvairiems anglies, azoto ir energijos šaltiniams, nes jų tarpląstelinė aplinka yra visiškai nuspėjama.
Nukleoidas yra bakterijos branduolio atitikmuo. 1956 m. bakterijų ląstelėse, kur yra bakterijų chromosoma, buvo aptikta „branduolinė zona“ arba nukleoidas. Bakterijų DNR randama apskrito ir linijinio pavidalo. E. coli ląstelėms buvo įrodyta, kad DNR egzistuoja apskritos molekulės pavidalu, o Borrelia burgdorferi 1989 m. buvo įrodyta, kad ląstelėse yra linijinės formos DNR. Nukleoidas, skirtingai nei eukariotinis branduolys, neturi branduolinis apvalkalas, branduolys ir pagrindiniai baltymai (histonai). Paprastai į bakterinė ląstelė yra viena chromosoma, bet jei dalijimasis sutrikęs, gali būti 4 ar daugiau vienos chromosomos kopijų. Kai kurių rūšių ląstelėse buvo aptiktos dvi ar trys neidentiškos chromosomos. Vibrio, Leptospira interrogans rūšys turi dvi apskritas chromosomas. Todėl mintis, kad bakterijos turi tik vieną chromosomą, laikoma pasenusia. Nukleoidas aptinkamas šviesos mikroskopu po dažymo specifiniais DNR metodais: pagal Feulgeną arba Romanovsky-Giemsa. Itin plonų bakterijų sekcijų elektronų difrakcijos modeliuose nukleoidas turi šviesių zonų pavidalą su fibrilinėmis, į siūlus panašiomis DNR struktūromis, susietomis su tam tikromis vietomis su citoplazmine membrana arba mezosoma, dalyvaujančia chromosomų replikacijoje.
Sterolių buvimas membranoje prisideda prie jų ilgaamžiškumo, nes padeda padidinti jėgas, laikančias membraną kartu. Nes nebūtų žmonių, augalų, gyvūnų, grybų ar pirmuonių. Visi šie organizmai yra Gyva būtybė, be bakterijų, susideda iš eukariotinių ląstelių.
Ir šio tipo ląstelių genezė nebūtų įmanoma be simbiozės. Eukariotinių ląstelių susidarymas moksle aiškinamas vadinamąja endosimbiontų teorija. Pagal šią teoriją sudėtingos ląstelės su ląstelės branduolys, atskirti nuo plazmos ir ląstelių organelių, susidarė iš paprastesnės struktūros prokariotų simbiozės. Tik bakterijos, kurios gyveno kaip endosimbiontas didesnėje ląstelėje, sudarė šių dienų eukariotinių ląstelių organelius.
Be nukleoido, bakterijų ląstelė turi ekstrachromosominių paveldimumo faktorių. - plazmidės, kurie yra kovalentiškai užsidarę DNR žiedai.Jie neužkoduoja funkcijų, kurios yra būtinos bakterijos ląstelės gyvybei, bet suteikia bakterijai pranašumų, kai yra veikiamos nepalankios egzistavimo sąlygos.
Mitochondrijų susidarymas
Šis organinių medžiagų kaupimasis yra sveikintinas heterotrofinių ląstelių maistinių medžiagų šaltinis. Ląstelės įgyja dydį ir, be to, gali įsisavinti organinės medžiagos fagocitozė, gebanti įsisavinti organinės ląstelės taip pat smulkesnės ląstelės fagocitizuojamos. Paprastai šios ląstelės yra maisto pavidalu, jos yra virškinamos.
Tačiau kartais nutinka taip, kad paimtos gyvos bakterijos viduje išgyvena didesnė ląstelė ir bendrauti su savo šeimininku. Vystosi simbiozė, kurioje ląstelė-šeimininkė gali efektyviau naudotis maisto produktai naudojant fermentus iš aerobinių bakterijų kvėpavimo grandinės. Kita vertus, endosimbiontas yra labiau apsaugotas didesnės ląstelės viduje nuo aplinkos pokyčių. Kita galimas šaltinis mitochondrijas įsiveržs bakteriniai parazitai šeimininko ląstelėje ir dėl to kils simbiozė.
Kapsulė, mikrokapsulė, gleivės. Kapsulė- gleivinės struktūra, kurios storis didesnis nei 0,2 mikrono, tvirtai susieta su bakterijų ląstelės sienele ir turinti aiškiai apibrėžtas išorines ribas. Kapsulė išsiskiria tepinėliais-atspaudais nuo patologinės medžiagos. Ląstelių kolonijų paviršius su kapsulėmis atrodo lygus, drėgnas, blizgus.
Organų ląstelės su daugybe membranų
Nėra visiškai tiksliai žinoma, ar eukariotai iš tikrųjų atsirado tokiu būdu, tačiau yra daug faktų, patvirtinančių endosimbiontų teoriją. Taip pat organelės negali susidaryti iš genetinės branduolio informacijos, o dauginasi dalijantis. Dalijimosi procesas panašus į bakterijų dalijimosi procesą. Ląstelių organelių ribosomos yra struktūriškai panašios į bakterines, o ne eukariotų ribosomas.
Endosimbiozės randamos ir šiandien. Taigi, kai kurios jūrų diatomijos gali gyventi simbiozėje su cianobakterijų trūkumu. Vėlgi, simbiontas yra integruotas į šeimininko ląstelės plazmą. Todėl ir šiandien tokia simbiozė kartojasi kaip procesas, buvęs viso aukštesniojo gyvenimo pradžioje.
Susidaro daug bakterijų mikrokapsulė- gleivinės susidarymas, kurio storis mažesnis nei 0,2 mikrono, aptiktas tik elektroniniu mikroskopu. Gleives reikėtų skirti nuo kapsulės – gleivinių egzopolisacharidų, kurie neturi aiškių išorinių ribų. Glebės tirpsta vandenyje.
Gleiviniai egzopolisacharidai būdingi Pseudomonas aeruginosa gleivinės padermėms, kurios dažnai randamos cistine fibroze sergančių pacientų skrepliuose. Bakteriniai egzopolisacharidai dalyvauja sukibime (prilimpa prie substratų); jie taip pat vadinami glikokaliksu. Be bakterijų atliekamos egzopolisacharidų sintezės, yra ir kitas jų susidarymo mechanizmas: tarpląstelinių bakterijų fermentų veikimas disacharidus. Dėl to susidaro dekstranai ir levanai.
Jam būdingas branduolio nebuvimas. Pagrindinis jų padalijimas vyksta į dvi ir Archaea. IN Kasdienybė Prokariotai mums žinomi kaip bakterijos. Dažnai žodis „bakterijos“ arba bakterijos tiesiogiai prilyginamas patogenams. Mažiausiai iš visų prokariotų yra patologiniai – tai yra patogeniški.
Ląstelės sienelė supa citoplazminę membraną ir vidinė dalis ląstelės. Vidaus apdailos nėra. Visi prokariotai turi tą pačią pagrindinę struktūrą. Daugumos prokariotų ląstelės sienelė yra už plazmos membranos ribų ir susideda iš angliavandenių darinio. Atsižvelgiant į mureino sluoksnių skaičių ir išsidėstymą, skiriamos gramteigiamos ir gramneigiamos bakterijos.
- Ląstelę supa plazminė membrana.
- Po šio tipo archeja neturi ląstelės sienelės.
Kapsulė ir gleivės apsaugo bakterijas nuo pažeidimų ir išsausėjimo, nes, būdamos hidrofilinės, gerai suriša vandenį ir neleidžia veikti apsauginiams makroorganizmo ir bakteriofagų faktoriams.
Flagella bakterijos lemia bakterijos ląstelės mobilumą. Flagella yra ploni siūlai, kilę iš citoplazmos membranos turi ilgesnis nei pats narvas. Žvyneliai yra 12–20 nm storio ir 3–15 µm ilgio. Jie sudaryti iš 3 dalių.:
Iš esmės ląstelė, apsupta membranos, toliau neskirstoma. Apskritai, yra mechanizmas, leidžiantis judėti. Jis yra ant citoplazminės membranos, esančios ant kito ląstelės sluoksnio. Jis pagamintas iš muraine. Ant jo guli išorinis apvalkalas. Apgultas vienaląstis įvedė žaliųjų dumblių, kad sutvirtintų. Simbiozės priežastys drastiškų pokyčių abiejose ląstelėse. vienaląstis organizmas endosimbiontu persijungia tarp plėšrūno fazės ir šeimininko fazės, perjungdamas savo energijos balansą iš heterotrofinio į autotrofinį, t.y. nuo tiekimo iki saulės energijos.
1. - spiralinis siūlas,
2. - kabliukas
3. - bazinis korpusas, kuriame yra lazdelė su specialiais diskais (1 pora diskų gramteigiamų bakterijų ir 2 poros gramneigiamų bakterijų) ir motoriniais baltymais (1 pav.).
diskaižvyneliai prisitvirtina prie citoplazminės membranos ir ląstelių sienelės. Taip sukuriamas elektros variklio su strypu efektas – rotorius, kuris suka žvynelį. Protonų potencialų skirtumas citoplazminėje membranoje naudojamas kaip energijos šaltinis. Sukimosi mechanizmą užtikrina protonų ATP sintetazė. Žvynelinės sukimosi greitis gali siekti 100 aps./min. Bakterijos turi dešinę besisukančius žvynelius. Jei bakterija turi keletą žvynelių, jos pradeda sinchroniškai suktis, susipindamos į vieną ryšulį, sudarydamos savotišką sraigtą.
Dviejų ląstelių adaptacijos procese augalas iš pirmuonių iškyla su dalelėmis; Tyrėjai pavadino juos Hatena, pernelyg paslaptinga vokiečių kalba. Šis stebėjimas yra dar vienas endosimbiozės teorijos patvirtinimas, pagal kurią visų augalų ir gyvūnų ląstelių pirmtakai susidarė endosimbiozės būdu.
Kokias organeles gavo pirmtakai augalų ląstelės pagal Endosombiontentoria Endosymbiosis? Įvardykite keturias šių organelių savybes, kurios palaiko endosimbiotinę teoriją! Kiekvienu atveju pagrindžiame, kodėl charakteristiką laikote endosimbiontų teorijos įrodymu. Kiek naujų Khaten dumblių atradimas gali būti laikomas endosimbiontų teorijos pagrindu? Endosimbiozės su aerobinėmis bakterijomis metu pirminiai eukariotai įgijo oksidacinio skaidymosi gebėjimą per simbiozę su cianobakterijomis, gebėjimą fotosintezei. Jie negali vėl pasirodyti. Tai rodo, kad jie iš pradžių nebuvo kilę kaip šeimininkų ląstelių struktūros. Mitochondrijos ir chloroplastai turi savo baltymų sintezės mechanizmus, o jų ribosomos yra sukurtos kaip prokariotai. Abu juos supa dvi membranos, kurių vidinė yra panaši į prokariotinė membrana, išorinis iš ląstelės membrana. Svarbus endosimbiotinės teorijos bruožas yra tai, kad tarp prokariotinių ir eukariotinių ląstelių nėra tarpinių formų, tačiau yra tarpinių formų tarp endosimbiotinių organizmų ir organelių. Ši pereinamoji forma yra stebimi vienaląsčiai dumbliai: po dietos transformacijos vienaląsčiai priklauso nuo simbiozės su žaliais dumbliais, tuo tarpu tik žali dumbliai yra gyvybinga.
- Kokių ypatingų įgūdžių įgijote?
- Mitochondrijos ir chloroplastai yra mikroorganizmų palikuonys.
- Mitochondrijos atsiranda iš bakterijų, chloroplastai – iš cianobakterijų.
- Abu atsiranda tik atskiriant bendraamžius.
Flagella yra sudaryta iš baltymų flagellinas(nuo. žvyneliai- flagellum), kuris yra antigenas - vadinamasis H-antigenas. Flagellino subvienetai yra suvynioti.
Žvynelių skaičius bakterijose Įvairios rūšys skiriasi nuo vieno (monotrichas) Vibrio cholerae iki dešimties ir šimtų žvynelių, besitęsiančių palei bakterijos perimetrą (peritrichas), adresu coli, protea ir kt. lopotrichas viename ląstelės gale turi ryšulį žvyneliais. amfitrichai priešinguose ląstelės galuose turi vieną žvynelį arba žvynelių ryšulį.
Flagelos aptinkamos naudojant sunkiaisiais metalais apipurkštų preparatų elektroninę mikroskopiją arba po apdorojimo šviesos mikroskopu. specialius metodus remiantis marinavimu ir adsorbcija įvairių medžiagų, dėl ko padidėja žvynelių storis (pavyzdžiui, pasidabravus).
villi, arba gėrė(fimbriae) – gijiniai dariniai, kt plonas ir trumpas, nei žvyneliai. Pili tęsiasi nuo ląstelės paviršiaus ir susideda iš Pilina voverė, kuri organizuota spiraline struktūra. Pilių architektūra varijuoja nuo plonų siūlų iki storų tvirtų strypo formos darinių su ašinėmis skylutėmis. Jie turi antigeninį aktyvumą. Išskirti gerti, atsakingas už sukibimą y., kad bakterijos prisitvirtintų prie paveiktos ląstelės, taip pat gertų, atsakingos už mitybą, vandens-druskos mainai, Ir seksualinis (F-drink) arba gėrė 4 rūšį, arba konjuguotą, gėrė.
Paprastai gėrė gausiai- keli šimtai ląstelėje. Tačiau paprastai vienoje ląstelėje yra 1-3 lytinės pilies: jas formuoja vadinamosios „vyriškos“ donorinės ląstelės, turinčios pernešamų plazmidžių (F-, R-, Co/-plazmidės). Išskirtinis lytinių pilių bruožas yra jų sąveika su specialiais „vyriškais“ sferiniais bakteriofagais, kurie intensyviai adsorbuojasi ant lytinių pilių (pav.).
Pagrindinė pilių paskirtis – palaikyti konkrečias tvirtinimo struktūras (adhezines) jų galuose. Adhezinai yra tarpininkai bakterijų kontaktuose, kontaktuojant su negyvais objektais, audiniais ir jautrių organizmų ląstelėmis. Šeimininkų audinių kolonizavimas bakterijų patogenais paprastai priklauso nuo stereocheminio panašumo tarp adhezino ir atitinkamo šeimininko ląstelės receptoriaus architektūros (pav.). Taigi pilių funkcionavimo tyrimas leis ne tik giliau suprasti kolonizacijos ir signalizacijos mechanizmą, bet ir sukurti naujų kartų antimikrobinius vaistus.
ginčų - savotiška ramybės bakterijų forma, turinti gramteigiamą ląstelių sienelės sandaros tipą (pav.).
Sporos susidaro, kai Ne palankiomis sąlygomis bakterijų buvimas (džiūvimas, maistinių medžiagų trūkumas ir kt.). Bakterijos ląstelės viduje susidaro viena spora (endospora). Sporų susidarymas prisideda prie rūšies išsaugojimo, o ne Xia dauginimosi būdas, kaip grybai.
sporas formuojančios genties bakterijos bacila, kurios sporos dydis neviršija ląstelės skersmens, vadinamas bacilos. sporas formuojančios bakterijos, kurios sporos dydis viršija ląstelės skersmenį, todėl jos įgauna verpstės formą, vadinamos klostridijos, tokios kaip genties bakterijos Clostridium(lot. klostridiumas- suklys). Sporos yra atsparios rūgštims, todėl jie dažomi Aujeskio metodu arba Ziehl-Nelsen metodu raudonai, o vegetatyvinė ląstelė – mėlyna spalva.
Sporuliacija, sporų forma ir vieta ląstelėje (vegetatyvinė) yra bakterijų rūšies savybė, leidžianti jas atskirti viena nuo kitos.
Ginčo forma gali būti ovalios, sferinės; išdėstymas ląstelėje :
1. terminalas, t.y., lazdos gale (stabligės sukėlėjoje),
2. subterminalas- arčiau lazdelės galo (dėl botulizmo sukėlėjų, dujų gangrena)
3. centralizuotai e (juodligės baciloje).
Procesas sporų susidarymas (sporuliacija) pereina eilę etapų, kurių metu atsiskiria dalis bakterijos vegetatyvinės ląstelės citoplazmos ir chromosomos, apsuptos įaugusios citoplazminės membranos, - prospore.
Prosporus apsuptas dviejų citoplazminių membranų, tarp kurių susidaro storas pakitęs žievės (žievės) peptidoglikano sluoksnis. Iš vidaus jis liečiasi su sporos ląstelės sienele, o iš išorės – su vidiniu sporos apvalkalu. Išorinį sporos apvalkalą sudaro vegetatyvinė ląstelė.
Kai kurių bakterijų sporos turi papildomą dangą - egzosporiumas.
Taip susidaro daugiasluoksnis prastai pralaidus apvalkalas. Sporuliaciją lydi intensyvus prosporos vartojimas, o vėliau atsirandantis dipikolino rūgšties ir kalcio jonų sporų apvalkalas. Sporos įsigyja karščiui atsparus, kuris yra susijęs su kalcio dipikolinato buvimu jame.
Sporos gali išlikti ilgą laiką dėl daugiasluoksnio apvalkalo, kalcio dipikolinato, mažas turinys vandens ir vangių medžiagų apykaitos procesų. Dirvožemyje, pavyzdžiui, patogenai juodligė stabligė gali išlikti dešimtmečius.
Esant palankioms sąlygoms, sporos sudygsta trimis nuosekliais etapais (pav.):
1. aktyvinimas
2. inicijavimas
3 . augimas
Tokiu atveju iš vienos sporos susidaro viena bakterija.
Aktyvinimas - jis paruoštas dygti. 60-80°C temperatūroje sporos suaktyvėja dygimui.
Iniciacija daigumas trunka keletą minučių. Būdingas augimo etapas staigus augimas, lydimas lukšto sunaikinimo ir sodinuko išlaisvinimo.
Klausimai savikontrolei
| | | | | | | | | 10 | | | | | |
Tipiškos bakterinės ląstelės struktūra parodyta fig. 2.3. Ant pav. 2.4 parodyta lazdelės formos bakterijos pjūvio elektroninė mikrografija. Matote, kokia paprasta yra bakterinė ląstelė, ypač lyginant su eukariotų ląstelėmis (7.5 ir 7.6 pav.).
Ryžiai. 2.3. Apibendrinta lazdelės formos bakterijų ląstelės sandaros schema. Dešinėje yra struktūros, esančios kiekvienoje ląstelėje, kairėje - ne visose ląstelėse. Flagellum kartais vienas, kaip Rhizobium, arba keli, kaip Azotobacter; paprastai jis yra ilgesnis už ląstelę. Kapsulė gali būti gleivėtas, kaip ir Azotobacter; jei kapsulė yra laisva, tada ji vadinama gleivinis sluoksnis. vamzdinės arba maišo formos fotosintetinės membranos turintys pigmentų yra plazminės membranos invaginacijos; fotosintetinėse bakterijose, tokiose kaip Chromatium, tokios membranos yra išsibarsčiusios po visą citoplazmą. Skaičius pili, arba pili, gali siekti nuo vieno iki kelių šimtų, kaip, pavyzdžiui, in Escherichia coli, Salmonella. Mezosoma yra daugialypis plazmos membranos įsiskverbimas, kaip, pavyzdžiui, Bacillus subtilis. ląstelių sienelės kietas ir jame yra mureino. Ribosomos, esantis visoje citoplazmoje, yra mažesnio dydžio nei eukariotų. Iš rezervinės maistinės medžiagos bakterijų ląstelėse galima aptikti lipidų, glikogeno, polifosfatų (volutino granulių). Citoplazma sudėtyje nėra organelių; yra fermentų ir kt.
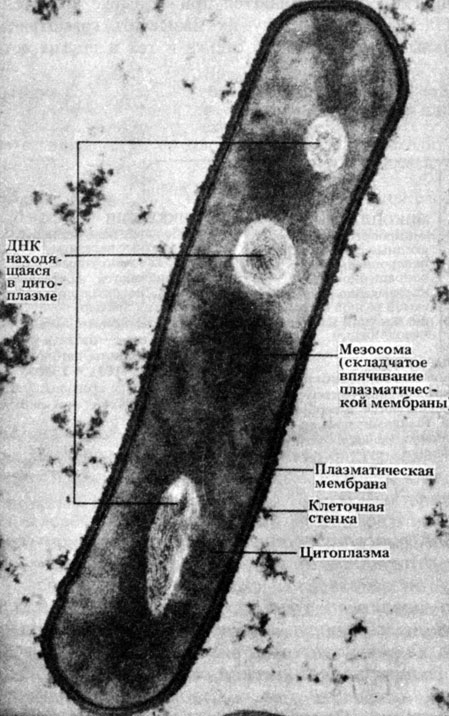
Ryžiai. 2.4. Tipiškos lazdelės formos bakterijos Bacillus subtilis pjūvio elektroninė mikrografija. Šviesiose srityse yra DNR. × 50 000
Kapsulės ir gleivinės sluoksniai
Kapsulės ir gleivinės sluoksniai yra tam tikrų bakterijų gleivinės ar lipnios išskyros; tokios išskyros aiškiai matomos po neigiamo kontrastavimo (kai dažymas yra ne preparatas, o fonas). Kapsulė yra gana storas ir kompaktiškas, ir gleivių sluoksnis daug laisvesnis. Kai kuriais atvejais gleivės padeda formuoti kolonijas iš atskirų ląstelių. Tiek kapsulė, tiek gleivinės sluoksniai tarnauja kaip papildoma ląstelių apsauga. Pavyzdžiui, įkapsuliuotos pneumokokų padermės laisvai dauginasi žmogaus organizme ir sukelia plaučių uždegimą, o nekapsuliuotas padermes lengvai užpuola ir sunaikina fagocitai, todėl yra visiškai nekenksmingos.
ląstelių sienelės
Ląstelės sienelė suteikia ląstelei tam tikrą formą ir standumą. Jis aiškiai matomas ant pjūvio (2.4 pav.). Kaip ir augalų, bakterijų ląstelės sienelė apsaugo nuo osmosinio patinimo ir ląstelių plyšimo, kai, kaip dažnai nutinka, jos patenka į hipotoninę aplinką (Priedo A.1.5 skirsnis). Vanduo, kitos mažos molekulės ir įvairūs jonai lengvai prasiskverbia pro mažytes ląstelės sienelės poras, tačiau didelės baltymų ir nukleorūgščių molekulės pro jas nepraeina. Be to, ląstelės sienelė turi antigeninių savybių, kurias jai suteikia joje esantys baltymai ir polisacharidai.
Pagal ląstelės sienelės sandarą bakterijas galima suskirstyti į dvi grupes. Kai kurie yra nudažyti gramais, todėl jie vadinami gramteigiamas, o kiti nuplaunant dažus tampa bespalviai (2.7 sk.), todėl jie vadinami gramneigiamas. Abiejų jų ląstelės sienelėje yra speciali standi gardelė, susidedanti iš mureina. Mureino molekulė yra reguliarus lygiagrečių polisacharidinių grandinių tinklas, susietas viena su kita trumpomis peptidinėmis grandinėmis. Taigi kiekvieną ląstelę supa tinklinis maišelis, sudarytas tik iš vienos molekulės. (Mureino polisacharidinė dalis aprašyta 5.7 lentelėje).
Gramteigiamose bakterijose, tokiose kaip Lactobacillus, į mureino tinklą yra įterptos kitos medžiagos, daugiausia polisacharidai ir baltymai. Taip aplink ląstelę susidaro gana storas ir standus paketas. Gramneigiamos bakterijos, tokios kaip Escherichia coli arba Azotobacter, turi daug plonesnę ląstelių sienelę, tačiau ji yra sudėtingesnė. Šių bakterijų mureino sluoksnis iš išorės padengtas minkštu ir lygiu lipidų sluoksniu. Tai apsaugo juos nuo lizocimas. Lizocimo yra seilėse, ašarose ir kituose kūno skysčiuose, taip pat baltymuose vištienos kiaušinis. Jis katalizuoja tam tikrų ryšių tarp angliavandenių liekanų hidrolizę ir taip suskaido mureino polisacharidinį stuburą. Ląstelės sienelė plyšta, o jei ląstelė yra hipotoniniame tirpale, įvyksta jos lizė (ląstelė osmosiškai išsipučia ir sprogsta). Lipidų sluoksnis suteikia ląstelių atsparumą penicilinui. Šis antibiotikas neleidžia susidaryti kryžminiams ryšiams gramteigiamų bakterijų ląstelės sienelėje, todėl augančios ląstelės tampa jautresnės osmosiniam šokui.
Flagella
Daugelis bakterijų yra judrios, o toks judrumas atsiranda dėl to, kad jose yra viena ar daugiau žvynelių. Bakterinės žvyneliai yra daug paprastesni nei eukariotų (17.6.2 skyrius, 2.1 lentelė), savo struktūra primena vieną iš eukariotų žiuželių mikrovamzdelių. Flagella yra sudaryta iš identiškų sferinių baltymų subvienetų flagellinas(panašus į raumenų aktiną), kurie išsidėstę spirale ir sudaro tuščiavidurį cilindrą, kurio skersmuo apie 10-20 nm. Nepaisant banguotos žvynelių formos, jos yra gana standžios.
Žvyneliai yra varomi unikalaus mechanizmo. Žuvyklos pagrindas, matyt, sukasi taip, kad žvynelis tarsi įsukamas į terpę nesudarant atsitiktinių dūžių ir taip judina ląstelę į priekį. Tai, matyt, vienintelė gamtoje žinoma konstrukcija, kurioje naudojamas rato principas. Kita įdomi savybė flagella yra atskirų flagelino subvienetų gebėjimas tirpale spontaniškai susiburti į spiralinius siūlus. Spontaniškas savęs surinkimas- Labai svarbus turtas daug sudėtingų biologinių struktūrų. Šiuo atveju savaiminis susijungimas yra visiškai dėl flagelino aminorūgščių sekos (pirminės struktūros).
Judrios bakterijos gali judėti reaguodamos į tam tikrus dirgiklius, t. y. jos gali taksi. Pavyzdžiui, aerobinės bakterijos turi teigiamą aerotaksį (t. y. jos nuplaukia ten, kur aplinkoje yra daugiau deguonies), o mobiliosios fotosintetinės bakterijos turi teigiamą fototaksį (t. y. jos plaukia link šviesos).
Flagelles lengviausia pamatyti elektroniniu mikroskopu (2.5 pav.), naudojant metalo nusodinimo techniką (A.2.5 skyrius).
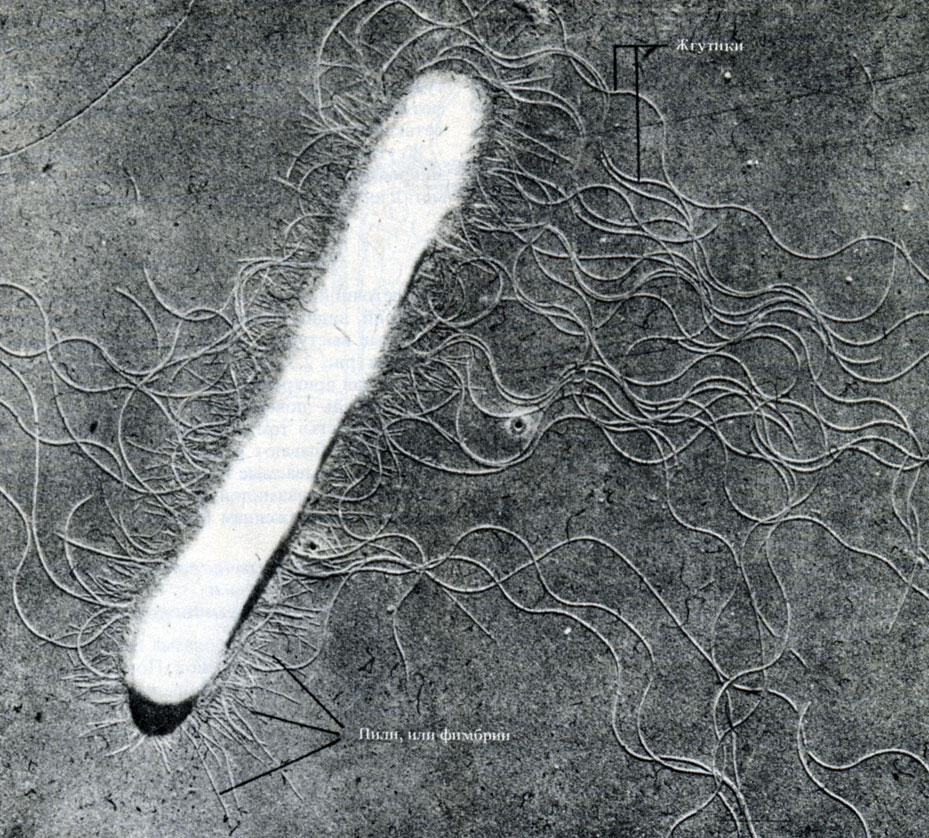
Ryžiai. 2.5. Lazdelės formos bakterijos mikrografija, gauta naudojant permatomą šviesą elektroninis mikroskopas. Aiškiai matoma ląstelės sienelė, fimbrijos ir ilgos banguotos žvyneliai, × 28000
Pili, arba fimbrija
Ant kai kurių gramneigiamų bakterijų ląstelės sienelės matomos plonos ataugos (lazdelės formos baltymų išsikišimai), kurie vadinami. išgėrė arba fimbriae(2.5 pav.). Jie yra trumpesni ir plonesni už žvynelius ir padeda ląstelėms pritvirtinti viena prie kitos arba prie kokio nors paviršiaus, suteikdamos specifinį „lipnumą“ jas turinčioms padermėms. Yra gėrimų skirtingo tipo. Įdomiausios yra vadinamosios F-pili, kurios yra užkoduotos specialios plazmidės (2.2.4 skyrius) ir yra susijusios su lytiniu bakterijų dauginimu.
Plazminė membrana, mezosomos ir fotosintezės membranos
Kaip ir visos ląstelės, bakterijų protoplazma yra apsupta pusiau pralaidžia membrana. Bakterijų plazminės membranos savo struktūra ir funkcijomis nesiskiria nuo eukariotinių ląstelių membranų (7.2.1 skyrius). Kai kuriose bakterijose plazminė membrana išsiveržia į ląstelę ir sudaro mezosomas ir (arba) fotosintezės membranas.
mezosomos- sulankstytos membranos struktūros (2.3 ir 2.4 pav.), kurių paviršiuje yra kvėpavimo procese dalyvaujančių fermentų. Todėl mezosomas galima vadinti primityviomis organelėmis. Ląstelių dalijimosi metu mezosomos prisijungia prie DNR, kuri, atrodo, palengvina dviejų dukterinių DNR molekulių atskyrimą po replikacijos ir skatina pertvaros susidarymą tarp dukterinių ląstelių.
Fotosintetinėse bakterijose plazmos membranos maišeliuose, vamzdeliuose ar sluoksniuotose invaginacijose yra fotosintetinių pigmentų (įskaitant bakteriochlorofilą). Panašios membranos dariniai taip pat dalyvauja azoto fiksavime.
genetinė medžiaga
Bakterijų DNR vaizduoja pavienės apskritos maždaug 1 mm ilgio molekulės. Kiekviena tokia molekulė susideda iš maždaug 5 × 10 6 bazių porų. Bendras DNR (genomo) kiekis bakterinėje ląstelėje yra daug mažesnis nei eukariotų ląstelėje, taigi ir joje užkoduotos informacijos kiekis yra mažesnis. Vidutiniškai tokioje DNR yra keli tūkstančiai genų, tai yra apie 500 kartų mažiau nei žmogaus ląstelėje (taip pat žr. 2.1 lentelę ir 2.3 pav.).
Ribosomos
Žiūrėkite lentelę. 2.1 (baltymų biosintezė) ir pav. 2.3.
ginčų
Kai kurios bakterijos (dažniausiai priklausančios Clostridium arba Bacillus genčiai) sudaro endosporas, t. y. sporas, esančias ląstelės viduje. Endosporos yra storasienės, ilgaamžės dariniai, itin atsparūs karščiui ir trumpųjų bangų spinduliuotei. Ląstelės viduje jie išsidėstę skirtingai, o tai labai svarbi tokių bakterijų identifikavimo ir sisteminimo savybė (2.6 pav.). Jei iš visos ląstelės susidaro rami, stabili struktūra, tai vadinama cista. Cistos sudaro kai kurias Azotobacter rūšis.
Ryžiai. 2.6. Įvairios formos bakterijų, pavyzdžiui, kelių labiausiai paplitusių naudingų ir patogeninių mikrobų tipų.
A. Cocci (sferinis)
Pavyzdys yra auksinis stafilokokas, gyvenantis nosiaryklėje; skirtingos stafilokokų padermės sukelia furunkulozę, plaučių uždegimą, apsinuodijimą maistu ir kitas ligas.
Pavyzdžiui, daugelis Streptococcus rūšių; kažkokia priežastis užkrečiamos ligos viršuje kvėpavimo takai; pavyzdžiui, S. pyogenes sukelia gerklės skausmą ir skarlatina; S. thermophilus suteikia jogurtui pikantišką skonį; S. lactis – žr. sekciją. 2.3.4
Šiai genčiai priklauso vienintelė rūšis Diplococcus pneumoniae (pneumococcus), pneumonijos sukėlėjas*.
* (D. pneumoniae sukelia lobarinę pneumoniją ir daugybę sunkių ūminių piogeninių infekcijų: meningitą, septicemiją, empiemą ir peritonitą. - Maždaug vert.)
B. Bacili (lazdelės formos)
![]()
Pavyzdžiai yra Escherichia coli (paprastas žarnyno simbiontas); Lactobacillus žr. 2.3.4; Salmonella typhi yra vidurių šiltinės sukėlėjas.
Pavyzdžiai yra Azotobacter, azotą fiksuojanti bakterija; Bacillus anthracis yra juodligės sukėlėjas.
Bacilos su endosporomis (sporos yra skirtingose padėtyse, turi skirtingų dydžių ir forma)
Jis yra centre ir nesukelia ląstelės patinimo, pavyzdžiui, Bacillus anthracis, juodligės sukėlėjo.
Įsikūręs motininės ląstelės gale, suteikia jį būdinga forma būgnų lazdelė, pavyzdžiui, Clostridium tetani, stabligės sukėlėjas.
Sporos yra subterminalinėje padėtyje, sukeldamos ląstelės patinimą, pavyzdžiui, Clostridium botulinum (sporos taip pat gali užimti centrinę vietą) - mirtinos ligos sukėlėjas. apsinuodijimas maistu- botulizmas.
B. Spirilli (spiralė)
Pavyzdys yra Spirillum; Spirochetų ląstelių forma yra labai panaši, tačiau skiriasi jų judėjimo būdas, pvz. Blyški treponema- sifilio sukėlėjas.
D. Vibrios (trumpos lazdos, visada išlenktos kablelio pavidalu)
Pavyzdys – Vibrio cholerae – choleros sukėlėjas; turi vieną žvynelį.