Siųsti savo gerą darbą žinių bazėje yra paprasta. Naudokite žemiau esančią formą
Studentai, magistrantai, jaunieji mokslininkai, kurie naudojasi žinių baze savo studijose ir darbe, bus jums labai dėkingi.
Įvadas
1.Ląstelių komponentų sandara ir funkcijos
2. Mitozinis ciklas. Veiksniai, turintys įtakos mitoziniam aktyvumui
3. Idėjų apie mirtingumą kritika paveldimos ligos
Literatūra
Įvadas
Narvas buvo aptiktas XVII amžiaus antroje pusėje. Ląstelės tyrimas ypač stipriai vystėsi XIX amžiaus antroje pusėje, siejant su kūryba ląstelių teorija. Ląstelių lygis tyrimai tapo pagrindiniu principu svarbiausių biologines disciplinas. Biologijoje susiformavo naujas skyrius – citologija. Citologijos tyrimo objektas yra daugialąsčių organizmų ląstelės, taip pat organizmai, kurių kūną atstovauja viena ląstelė. Citologija tiria struktūrą, cheminę sudėtį, jų dauginimosi būdus, prisitaikymo savybes. Šiame darbe bus nagrinėjama ląstelės komponentų struktūra ir funkcijos.
Mitozė yra netiesioginis ląstelių dalijimasis, dėl kurio iš pirminės ląstelės atsiranda dvi naujos, turinčios lygiai tą patį genų rinkinį. Mitozinis ciklas – procesų visuma, kurios metu iš vienos ląstelės susidaro dvi naujos ląstelės, apima mitozės periodą ir dalį tarpfazės. Mūsų darbo šiuo klausimu tikslas taip pat bus išanalizuoti veiksnius, turinčius įtakos mitoziniam aktyvumui.
Paveldimumas yra pagrindinė visų gyvų būtybių savybė, kuri slypi tame, kad visi gyvi organizmai geba kaupti informaciją apie savo sandarą ir perduoti šią informaciją kitoms kartoms. Žmonija nuėjo ilgą kelią sunkus kelias teisingai suprasti paveldimumo priežastis ir dėsnius. Mūsų darbo tikslas paveldimumo aspektu bus išnagrinėti idėjų apie paveldimų ligų mirtingumą kritiką, kuri ypač aktuali šių dienų medicinos tyrimams.
1.Ląstelių komponentų sandara ir funkcijos
Teorinis citologijos pagrindas yra ląstelių teorija. Ląstelių teoriją 1838 metais suformulavo T. Schwann, nors pirmosios dvi ląstelės teorijos nuostatos priklauso M. Schleidenui, tyrinėjusiam augalų ląsteles. Žinomas gyvūnų ląstelių sandaros specialistas T. Schwannas 1838 m., remdamasis M. Schleideno darbų duomenimis ir savo paties tyrimų rezultatais, padarė tokias išvadas:
Ląstelė yra mažiausia struktūrinis vienetas gyvieji organizmai.
Ląstelės susidaro dėl gyvų organizmų veiklos.
Gyvūnų ir augalų ląstelės turi daugiau panašumų nei skirtumų.
Daugialąsčių organizmų ląstelės yra tarpusavyje susijusios struktūriškai ir funkciškai.
Tolesnis struktūros ir gyvenimo veiklos tyrimas leido apie tai daug sužinoti. Tai padėjo tobulėti mikroskopiniai metodai, tyrimo metodai ir daugybė talentingų tyrinėtojų atėjimas į citologiją. Išsamiai ištirta branduolio sandara, atlikta tokių svarbių biologinių procesų kaip mitozė, mejozė, apvaisinimas citologinė analizė. Tapo žinoma pati ląstelės mikrostruktūra. Buvo atrastos ir aprašytos ląstelių organelės. Programa citologiniai tyrimai XX amžius iškėlė uždavinį išsiaiškinti ir tiksliau atskirti ląstelės savybes. Iš čia Ypatingas dėmesys pradėtas skirti ląstelės cheminės sudėties ir medžiagų įsisavinimo ląstelės aplinkoje mechanizmui tirti.
Visi šie tyrimai leido padauginti ir išplėsti ląstelių teorijos nuostatas, kurių pagrindiniai postulatai šiuo metu atrodo taip:
Ląstelė yra pagrindinis ir struktūrinis visų gyvų organizmų vienetas.
Ląstelės susidaro tik iš ląstelių dalijimosi rezultatu.
Visų organizmų ląstelės yra panašios sandaros, cheminė sudėtis, pagrindinės fiziologinės funkcijos.
Daugialąsčių organizmų ląstelės sudaro vieną funkcinį kompleksą.
Visų gyvų būtybių ląsteles žemėje galima iš esmės suskirstyti į dvi skirtingi tipai: branduolinis (eukariotinis) ir nebranduolinis (prokariotinis). Prokariotinės ląstelės yra seniausios mūsų planetoje, tai bakterijų ir melsvadumblių ląstelės. Jie pasižymi šiomis savybėmis:
Šerdies nebuvimas.
Apvalios DNR buvimas.
Daugkartinis tų pačių genų pasikartojimas DNR.
Trūksta savaime besidalijančių ląstelių organelių: centriolių, mitochondrijų, plastidžių.
Ląstelių dalijimasis amitozės būdu (tiesioginis dalijimasis).
Iš eukariotinių ląstelių susidaro augalai, grybai ir gyvūnai. Jie pasirodė vėliau nei prokariotai. Jiems būdingi tokie požymiai kaip:
Branduolio, kuriame visada yra DNR molekulės, buvimas. Kai kurios ląstelės netenka branduolio antrą kartą (žinduolių eritrocitai ir trombocitai).
DNR visada yra vienos ar kelių gijų, atvirų galuose.
Kiekvienos DNR molekulės genai, kaip taisyklė, nesikartoja.
Ląstelės visada turi savaime besidalijančių organelių, kurios turi savo DNR molekules: centriolių, mitochondrijų, plastidžių. Pastarieji randami tik augalų ląstelėse.
Ląstelių dalijimasis mitozės būdu (netiesioginis dalijimasis), ko pasekoje visi genai tolygiai pasiskirsto tarp naujų ląstelių.
Eukariotinės ląstelės yra dešimtis ir šimtus kartų didesnės už prokariotines.
Leiskite mums išsamiau apsvarstyti eukariotinės ląstelės struktūrą.
Ląstelė turi membraną, citoplazmą ir branduolį.
Membrana yra keturių sluoksnių struktūros ląstelės organelės. Išorinis ir vidinis sluoksniai yra baltymingi. Tarp jų yra du sluoksniai į riebalus panašių medžiagų – lipoidų. Vienas iš lipoidų molekulės galų turi aiškiai apibrėžtas hidrofobines savybes. Membranoje visi lipoidai išsidėstę taip, kad kiekvienas sluoksnis hidrofobiniais galais būtų nukreiptas priešinga kryptimi nei kitas. IN skirtingos vietos Ląstelės membranoje yra specialių didelių baltymų molekulių, kurios užima visą jos storį. Daugelio ląstelių membranos iš išorės yra padengtos papildomais apsauginiais apvalkalais, susidedančiais iš angliavandenių (pavyzdžiui, iš celiuliozės augalų ląstelėse) arba iš sudėtingų medžiagų - gliukoproteinų (blakstienų ir žievelių gabalėlių). Ląstelės sveikata, jos gyvenimo trukmė labai priklauso nuo membranos būklės.
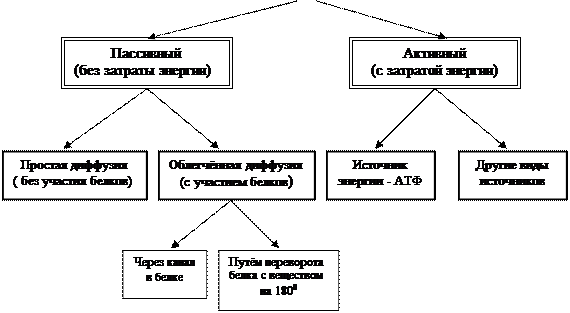
Visiškas vandens pralaidumas. Membrana visada leidžia vandenį į ląstelę arba išeina, priklausomai nuo to, kur vandens koncentracija yra didesnė. Toks medžiagos judėjimas iš didelės koncentracijos srities į mažesnės koncentracijos sritį vadinamas difuzija. Medžiagos difuzijai nereikia energijos.
Atrankinis tirpių medžiagų laidumas:
Neigiamo krūvio dalelės greičiau ir lengviau prasiskverbia pro membraną.
Riebaluose tirpios medžiagos lengviau prasiskverbia pro membraną nei vandenyje tirpios medžiagos.
Mažos molekulės lengviau prasiskverbia pro membraną nei didelės.
Aktyvus medžiagų pernešimas. Kai kurios medžiagos gali prasiskverbti pro membraną priešinga jų difuzijai kryptimi, ty iš žemos vietos į vietą, kurioje yra daugiau didelė koncentracija. Aktyviuoju transportavimu iš ląstelės nuolat pašalinamas natrio, vandenilio ir chloro jonų perteklius. O fosfatai, gliukozė, aminorūgštys, atvirkščiai, aktyviai prasiskverbia į citoplazmą. Aktyvus transportas visada yra susijęs su energijos sąnaudomis.
Membrana reguliariai atkuriama dėl specialių organelių, sintezuojančių membranos vakuoles, darbo. Daugelis membranų nepadengtos tankūs kriauklės, gali suformuoti laikinas ataugas, vadinamas pseudopodijomis (pseudopodijomis).
Membranos funkcijos:
Fagocitozė yra kietų maisto dalelių gaudymas pseudopodais. Dėl to susidaro virškinimo vakuolė, plūduriuojanti citoplazmoje.
Pinocitozė yra tirpių medžiagų absorbcija.
Apsauginis. Membrana apsaugo ląstelę nuo pašalinių, pavojingų medžiagų patekimo į ją.
Kvėpavimo. Per membraną į ląstelę patenka deguonis ir išsiskiria anglies dioksidas.
Homeostatinis. Homeostazė – tai gebėjimas išlaikyti santykinai pastovią sudėtį. Dėl savo savybių (selektyvaus medžiagų įsisavinimo ir aktyvaus pernešimo) membrana užtikrina ląstelei savo sudėties pastovumą.
Integruojantis. Ląstelės tarpusavyje bendrauja naudodamos membranas. Per membraną viena ląstelė gali perduoti įvairią informaciją kitai ląstelei. Ši informacija gali būti perduodama tiek naudojant elektrinius impulsus, tiek naudojant cheminių medžiagų(hormonai, mediatoriai).
Citoplazma – ląstelių sultys, ląstelės skystis. Jame yra vandens, jame ištirpusių neorganinių ir organinių medžiagų, taip pat įvairių atskirų struktūrų, vadinamų organelėmis:
Ribosomos yra ląstelių organelės, susidedančios iš dviejų dalelių, didelių ir mažų. Kiekvieną dalelę sudaro baltymai ir ribosominė RNR. Ribosomos vykdo baltymų sintezę. sintezuojamas branduolyje.
Endoplazminis tinklas (ER) yra ląstelės membraninė organelė, atstovaujanti daugybei kanalų ir ertmių iš membranų, savo struktūra panašių į ląstelės membraną. Pagal struktūrą ir funkcijas jis skirstomas į du tipus: grubus ER – paviršiuje yra ribosomų ir yra baltymų sintezės vieta; sklandžiai ER – neturi ribosomų, yra angliavandenių, lipoidų ir riebalų sintezės vieta. Išorėje ER kontaktuoja su ląstelės membrana, viduje – su branduoline membrana.
Golgi aparatas yra endoplazminiame tinkle. Turi membraninę struktūrą. Tai atrodo kaip daugybės maišelių, ertmių, vakuolių sankaupa. Atlieka daug funkcijų:
Atneša baltymus į galutinę darbinę formą, kai kuriuos baltymus į didelius baltymų kompleksai, prie kai kurių baltymų prijungia reikiamus metalo jonus.
Sudaro membranines pūsleles, kurios, palikdamos Golgi kompleksą, arba atkuria ląstelės membraną, arba virsta lizosomomis.
Lizosomos yra ląstelės membraninės organelės, atstovaujančios mikroskopines pūsleles, užpildytas virškinimo fermentais. Jie atlieka virškinimo ir apsaugines funkcijas. Jie gali sulipti su virškinimo vakuole, supilti į ją virškinimo fermentai. Ląstelei kontaktuojant su svetima medžiaga arba su svetima ląstele, lizosomos prilimpa prie ląstelės membranos, išskirdamos savo fermentus už ląstelės ribų. Lizosomų fermentai taip pat gali būti susiję su užprogramuota ląstelių mirtimi.
Mitochondrijos yra membraninės savaime besidalijančios organelės. Jas sudaro du membranų sluoksniai: išorinis lygus ir vidinis, turintis daugybę ataugų mitochondrijų viduje. Tokios vidinės membranos ataugos vadinamos cristae. Juose vyksta pieno rūgšties oksidacijos procesas, dėl kurio išsiskiria energija, kaupiama ATP pavidalu (oksidacinis fosforilinimas). Todėl svarbiausia mitochondrijų funkcija yra energija. Mitochondrijos turi savo DNR molekules, kurios savo struktūra nesiskiria nuo bakterijų DNR. Mitochondrijos, kaip ir bakterijos, dauginasi tiesioginio dalijimosi būdu.
Plastidės yra membraninės savaime besidalančios ląstelių organelės. Skirtingai nuo visų aukščiau aptartų organelių, plastidai randami tik augalų ląstelėse. Savo struktūra jie primena mitochondrijas: juos sudaro dvi membranos, išorinė lygi ir vidinė, sudarančios daugybę plokščių ataugų - tilakoidų. Visi tilakoidai yra sukrauti kaip monetų krūvos. Kiekvienas krūvas vadinamas aspektu. Tarp granos yra vidinis plastido skystis, vadinamas stroma. Jame yra savo DNR, kurios struktūra panaši į bakterijų. Plastidės dauginasi kaip bakterijos tiesioginio dalijimosi būdu.
Centrioliai yra savaime besidalijančios organelės gyvūnų ląstelėse ir kai kuriose žemesni augalai. Kiekvieną centriolę sudaro trumpas tuščiaviduris cilindras, kurio sieneles sudaro mikrovamzdeliai, išsidėstę išilgai cilindro ašies. Centriolėse yra baltymų ir nedidelis kiekis RNR. Ląstelėje yra dvi poros centriolių.
2. Mitozinis ciklas. Veiksniai, turintys įtakos mitoziniam aktyvumui
Mitozinis ciklas – procesų visuma, kurios metu iš vienos ląstelės susidaro dvi naujos ląstelės. Mitozinis ciklas apima mitozės laikotarpį ir dalį tarpfazės. - laikotarpis tarp dalijimų, kai vyksta pasiruošimas kitai mitozei. Mitozinis ciklas yra ląstelės gyvavimo ciklo dalis; greitai besidalijančiose ląstelių populiacijose (pavyzdžiui, traiško kiaušinėlio blastomeruose) mitozinis ciklas beveik sutampa su gyvenimo ciklas ląstelės.
Mitozė yra netiesioginis ląstelių dalijimasis, dėl kurio iš pirminės ląstelės atsiranda dvi naujos, turinčios lygiai tą patį genų rinkinį.
Mitozė trunka 1-2 valandas ir vyksta keturiomis fazėmis, iš kurių pirmoji ir paskutinė yra ilgiausios.
mitozės fazės.
Profazė. Yra chromatino siūlų kondensacija, tai yra jų pakuotė. Susidaro sustorėjusios chromosomos, aiškiai matomos šviesos mikroskopu (su specialiu atspalviu). RNR ir baltymų sintezė baigiasi. Branduolio apvalkalas sunaikinamas. Suformuojamas velenas.
Metafazė. Visos chromosomos juda į ląstelės centrą, esantį išilgai jos pusiaujo. Kiekvieną chromosomą sudaro dvi aiškiai apibrėžtos dukterinės chromatidės, kurias sudaro dukterinė DNR, susidaranti dėl motinos dauginimosi. Kiekviena dukterinių chromatidžių pora yra sujungta viena su kita plona pertrauka, vadinama centromera. Tai motinos DNR dalis, kurioje reduplikacija dar neįvyko. Kiekviena centromera turi prie jo pritvirtintą savo veleno sriegį.
Anafazė. Dukterinės chromatidės atsiskiria viena nuo kitos dėl centromerų reduplikacijos ir greitai nukrypsta į priešingus ląstelės polius. Dabar kiekvienas polius turi savo chromatidžių rinkinį. Abiejuose šiuose rinkiniuose yra tie patys genai, nes visos dukterinės chromatidės, susidarančios motinos DNR dauginimosi metu, yra viena kitos kopijos.
Telofazė. Poliuose chromatidinės ląstelės išsivynioja į chromatino gijas. RNR ir baltymų sintezė atsinaujina. Aplink kiekvieną dukterinių chromatidžių rinkinį susidaro jų pačių branduoliniai apvalkalai. Narvas suvarstomas išilgai pusiaujo. Susidaro dvi naujos ląstelės.
Dėl mitozinio dalijimosi dvi ląstelės yra genetiškai visiškai vienodos. Tai įmanoma tik dviem procesais:
DNR replikacija paremta komplementarumo principu.
kiekvienos dukterinių chromatidžių poros divergencija į naujas ląsteles.
Mitozinių ląstelių dalijimasis vyksta:
adresu nelytinis dauginimasis augalų grybai ir gyvūnai,
visų daugialąsčių organizmų embrioniniame ir poembrioniniame vystymesi iš apvaisinto kiaušinėlio,
žaizdų gijimo, kraujo ląstelių susidarymo, odos ir žarnyno epitelio ląstelių augimo ir kitų procesų metu.
Apšvitinus labai daug to paties tipo ląstelių, buvo nustatyta, kad veikiant skirtingi tipai spinduliuotė, grįžtamojo ląstelių dalijimosi slopinimo trukmė ir ląstelių, kuriose dalijimasis visiškai sustojo, procentas didėja didėjant radiacijos dozei. Didėjant radiacijos dozei, vis daugiau ląstelių praranda gebėjimą daugintis arba bent jau laikinai nustoja dalytis. Vienas iš šio ląstelių gebėjimo daugintis tiek vienaląstėse, tiek audinių ląstelėse pažeidimo rodiklių aukštesni organizmai yra milžiniškų ląstelių formų atsiradimas.
Kai kurie radiaciniai-biocheminiai pakitimai atsiranda jau po santykinai mažų dozių poveikio, kiti – tik veikiant terpei ar didelėmis dozėmis radiacija. Tarp medžiagų apykaitos sutrikimų, atsirandančių veikiant jonizuojanti radiacija, į pirmąją vietą reikėtų kelti jautriausio radiacijai substrato – nukleino rūgščių – pažeidimą. radiacijos sužalojimas nukleorūgščių sintezės slopinimo forma negali būti laikoma tiesiogine ląstelių dalijimosi slopinimo ar chromosomų plyšimo priežastimi, dėl ko gali atsirasti didelių jų morfologinių sutrikimų, nustatytų mitozių metu po švitinimo. Kitų medžiagų apykaitos tipų, pavyzdžiui, angliavandenių, pažeidimai suteikia teisę kalbėti apie labai mažą radijo jautrumą. Pakeitimai angliavandenių apykaitą po švitinimo, ypač anaerobinės glikolizės slopinimo, paprastai pastebimas tik po 5000–20000 r. dozių poveikio; ląstelių kvėpavimo pažeidimas paprastai stebimas dėl dar didesnių dozių poveikio - nuo 20 000 iki 100 000 r.
Veikiant mažoms radiacijos dozėms, stebimas ląstelių dalijimosi slopinimas. At didelėmis dozėmis ląstelės galiausiai praranda gebėjimą daugintis. Laikinas mitozės slopinimas ir visiškas sterilumas negali būti dėl vieno mechanizmo, nepaisant to, kad abu šie reiškiniai iš pirmo žvilgsnio gali atrodyti gana susiję.
Iš radiacijos kokybės, išskyrus funkciniai pokyčiai, taip pat priklauso tam tikrų tipų spindulių chromosomų aberacijos. Ląstelių populiacijose, turinčiose mitozinį ląstelių dalijimąsi, po švitinimo pirmiausia pastebimas trumpalaikis mitozių dažnio padidėjimas, o vėliau - sumažėjimas iki tam tikros minimalios vertės.
Tam tikri chromosomų pokyčių tipai būdingi pirminiam ir antriniam radiacijos poveikiui.
Pirminio ir antrinio poveikio chromosomų pokyčių mechanizmas skiriasi. Pirminiam poveikiui būdingi chromosomų pokyčiai daugiausia atsiranda tose ląstelėse, kurios švitinimo metu turėjo mitozinį aktyvumą ir buvo metafazės stadijoje. Tam tikra dalis šių ląstelių turi mitozių, kurių dažnis mažėja dėl švitinimo. Kitose mitotiškai besidalijančiose ląstelėse, pasiekusiose arba perėjusiose metafazės stadiją, mitozės tęsiasi, bet lėčiau.
Tam tikra spinduliuotė yra susijusi su ląstelių dalijimusi (mitoze), kurią atrado ir išmatavo A.G. Gurvichas. Jis pavadino tai „mitogenetiniu“. Nustatyta, kad jei kitos ląstelės patenka į šią spinduliuotę, padidėja jų mitozė, tai yra, skatinamas jų augimas.
3. Idėjų apie paveldimų ligų mirtingumą kritika
Dar visai neseniai net tarp gydytojų vyravo nuomonė apie paveldimų ligų mirtingumą, jų profilaktikos ir gydymo neįmanomumą. Šiandien kai kurioms ligoms jau rasti gydymo metodai. Fenilketonurija nustatoma vidutiniškai kiekvienam iš 10 tūkstančių naujagimių. Kadangi organizme nėra fermento, kuris kontroliuoja aminorūgšties fenilalanino pavertimą tirozinu, fenilalanino koncentracija padidėja dešimt kartų. Dalis jo išsiskiria su šlapimu, o likusi dalis paverčiama fenilpiruvo, fenilacto, fenilpieno rūgštimis ir kitomis medžiagomis. Tai sukelia daugybę antrinių biocheminiai pokyčiai dėl to sutrinka smegenų brendimas. Nukrypimai į psichinis vystymasis vaikas tampa pastebimas tik sulaukus 6 mėn. Dauguma vaikų, sergančių fenilketonurija, užaugo protiškai neįgalūs. Dabar tokį medžiagų apykaitos sutrikimą šalina dieta be baltymų, kurios laikosi vaikas iki 6-8 metų. Baltyminės medžiagos pateikiamos tik specialių preparatų pavidalu, iš kurių pašalintas fenilalaninas. Ligos diagnozė gana paprasta: ji pagrįsta teigiama kokybine šlapimo reakcija su fenilpiruvo rūgštimi.
Kita paveldima liga – galaktozemija – dažnai pasireiškia jau pirmosiomis vaiko gyvenimo dienomis vėmimu, stipriu vangumu, hipotenzija, gelta, traukuliais. Jei liga vystosi palaipsniui, pagrindiniai simptomai nustatomi šiek tiek vėliau. Tai apima kataraktą, protinis atsilikimas ir lėtinis kepenų pažeidimas – hepatitas. Pacientams yra padidėjęs galaktozės kiekis ( pieno cukraus), sumažėja gliukozės kiekis. Jei diagnozė nustatoma laiku ir pieno produktai neįtraukiami į vaiko racioną, vaiko vystymasis yra normalus.
Pateikėme tik du pavyzdžius. Yra daug daugiau paveldimų ligų. Pasaulio sveikatos organizacijos duomenimis, apie 4% naujagimių kenčia nuo tam tikrų genetinių defektų. Bet prie jų būtina pridėti ir tuos vaikus, kuriems liga pasireiškia ne iš karto po gimimo, o vėlesniame amžiuje. Todėl tolesnė medicininės genetikos plėtra, genetinių žinių sklaida ne tik tarp gydytojų, bet ir tarp gyventojų yra svarbus uždavinys. Ne dieviškas išankstinis nusistatymas, bet tikros priežastys yra kiekvienos rūšies paveldimos patologijos pagrindas. Kova su šiomis ligomis vykdoma dviem būdais. Pirmasis būdas – kryptingas aplinkos sąlygų keitimas, dėl kurio ligos vystymasis tampa neįmanomas. Antroji – prevencija per medicininį genetinį gyventojų konsultavimą.
Koreguojami paveldimi medžiagų apykaitos sutrikimai speciali dieta: organizmo nevirškinamų medžiagų pašalinimas iš maisto arba, priešingai, trūkstamų. Koreguojami daugelis kalbos, klausos, regos organų defektų chirurginiu būdu. Nežinodamas tikrosios priežastys gimus vaikams su įgimtomis deformacijomis, žmonės tai dažnai laikė „Dievo bausme už nuodėmes“ arba sunkių nelaimių pranašu. J. W. Ballantyne knygoje The Teratological Records of the Chaldeans (1894) pateikia pavyzdžių įvairios interpretacijos ir prognozės, susijusios su deformacijų gimimu: "Jei kuri nors moteris pagimdys vaiką, kuris neturi šnervių, šaliai grės nelaimė, o vyro namai bus sugriauti. Jei kuri nors moteris pagimdys vaiką, kuris neturi nosis, bėda ištiks šalį ir mirs namo šeimininkas.Jei kuri nors moteris pagimdys vaiką be varpos, tai namo šeimininkas pasiims gausų derlių iš laukų.Jei moteris pagimdys vaiką, kurio lytis nėra aiškiai nurodyta, šalį ištiks nelaimės ir negandos, o vyrą lydės nelaimė“.
Neurozių kilmėje ir reaktyviosios psichozės vaidinti pagrindinį vaidmenį psichinė trauma kurios kartais tik provokuoja paveldimas polinkisį ligą. Pagal kilmę psichinė liga derinys vaidina svarbų vaidmenį priežastiniai veiksniai Su individualios savybės asmuo. Pavyzdžiui, ne visi sifiliu sergantys asmenys suserga sifiline psichoze, o tik nedaugeliui pacientų, sergančių smegenų ateroskleroze, išsivysto demencija arba haliucinacinė-kliedesinė psichozė.
Šiandien daugelis paveldimų negimusio vaiko ligų diagnozuojamos nėštumo pradžioje, tiriant vaisiaus vandenis. Tai leidžia pradėti laiku gydyti jei įmanoma ir tikslinga, arba nutraukti nėštumą, kad negimtų neįgalus vaikas. Tėvai turi būti informuoti apie jiems gresiančios nelaimės laipsnį, kad galėtų priimti teisingą sprendimą.
išvadas
Taigi visi organizmai susideda iš tų pačių dalių – ląstelių; jie formuojasi ir auga pagal tuos pačius dėsnius. Bendras elementariųjų organizmo dalių vystymosi principas yra ląstelių formavimasis. Kiekviena ląstelė tam tikrose ribose yra individas, savotiška nepriklausoma visuma. Tačiau šie asmenys veikia kartu, kad susidarytų harmoningas audinys. Visi audiniai sudaryti iš ląstelių. Augalų ląstelėse vykstantys procesai sumažėja iki šių: naujų ląstelių atsiradimas, ląstelių dydžio padidėjimas, ląstelių turinio pasikeitimas, ląstelės sienelės sustorėjimas.
Ląstelių teorijos sukūrimo dėka tapo aišku, kad ląstelė yra svarbiausia visų gyvų organizmų sudedamoji dalis. Ląstelės sudaro audinius ir organus. Vystymasis visada prasideda nuo vienos ląstelės, todėl galima sakyti, kad ji yra daugialąsčio organizmo pirmtakas. Struktūra ir funkcijos ląstelių komponentai buvo svarstomi šiame darbe.
Kaip matote, gebėjimas dalytis - svarbiausias turtas ląstelės. Be dalijimosi neįmanoma įsivaizduoti vienaląsčių būtybių skaičiaus padidėjimo, sudėtingo daugialąsčio organizmo išsivystymo iš vieno apvaisinto kiaušinėlio, ląstelių, audinių ir net per organizmo gyvavimo laiką prarastų organų atsinaujinimo.
Ląstelių dalijimasis vyksta etapais. Kiekviename padalijimo etape vyksta tam tikri procesai. Dėl jų padvigubėja genetinė medžiaga (DNR sintezė) ir jos pasiskirstymas tarp dukterinių ląstelių. Ląstelės gyvenimo laikotarpis nuo vieno dalijimosi iki kito vadinamas ląstelės ciklu. Mitozės metu ląstelė pereina eilę nuoseklių fazių, dėl kurių kiekviena dukterinė ląstelė gauna tokį patį chromosomų rinkinį, koks buvo motininėje ląstelėje.
Yra 4 mitozės fazės: profazė, metafazė, anafazė ir telofazė. Tarp veiksnių, turinčių įtakos mitoziniam aktyvumui, ypatingą dėmesį skyrėme radiaciniam-biocheminiam poveikiui.
Dar visai neseniai net tarp medikų vyravusi nuomonė apie paveldimų ligų mirtingumą, jų profilaktikos ir gydymo neįmanomumą sulaukia vis daugiau kritikos. Ir šiandien kai kurioms ligoms gydymo metodai jau rasti.
Be to, šiuo metu daugelis paveldimų negimusio vaiko ligų gali būti diagnozuojamos nėštumo pradžioje, ištyrus vaisiaus vandenis, o tai leidžia laiku pradėti gydymą, jei reikia, arba nutraukti nėštumą, kad būtų išvengta kūdikio gimimo. neįgalus vaikas.
Literatūra
1. Adamčikas M.V. – Pasaulio enciklopedija. Biologija / Red. Anikejevas V.I. - M.: Šiuolaikinis rašytojas, 2006 m.
2. Kalyuzhny V.G. Biologijos vadovas. (Serija „Vadovėliai ir mokymo priemonės“) – Rostovas prie Dono, Finiksas, 2004 m.
3. Giliarov M.S. Biologija. Didelis enciklopedinis žodynas. – 3 leidimas. - (Aukso fondas). - M.: Didžioji rusų enciklopedija, 2005 m.
4.B. Smelovas. Ląstelių struktūra. Laikraštis „Biologija“. - Nr.36/2001.
5.P. Piniginė. Augalų ląstelės doktrina. - http://bio.1september.ru/article.php?ID=200204304
6.I. Bologova. Ląstelė yra struktūrinis ir funkcinis gyvojo vienetas. - http://bio.1september.ru/article.php?ID=200200603
7. Grekova T.I. Ateizmas ir medicina. - http://lib.metromir.ru/author5836
8. N. Greenas, W. Stoutas, D. Tayloras. Biologija. M.: Mir, 1996 m.
Panašūs dokumentai
Ląstelės, jos sandaros ir komponentų tyrimo istorija ir pagrindiniai etapai. Ląstelių teorijos turinys ir reikšmė, žymūs mokslininkai, prisidėję prie jos kūrimo. Simbiotinė teorija (chloroplastai ir mitochondrijos). Eukariotinės ląstelės kilmė.
pristatymas, pridėtas 2016-04-20
Ląstelių tyrimo metodai, jų priklausomybė nuo mikroskopo objektyvo tipo. Ląstelių teorijos pozicijos. gyvūnų ląstelės ir augalinės kilmės. Fagocitozė – ląstelės absorbcija iš aplinką tankios dalelės. Požiūriai į paveldimų ligų gydymą.
pristatymas, pridėtas 2014-12-09
Eukariotinės ląstelės branduolys. Ląstelės, turinčios daugiau nei du chromosomų rinkinius. Dalijimosi procesas eukariotuose. Jungtinės homologinių chromosomų poros. Augalų ląstelių ontogeniškumas. Ląstelių atskyrimo procesas dėl medianinės sluoksnio sunaikinimo.
santrauka, pridėta 2011-01-28
Ląstelių ciklo periodai ir fazės. Nuoseklus ciklo laikotarpių perėjimas per ląstelę nepraleidžiant arba grįžtant į ankstesnius etapus. Pradinės ląstelės padalijimas į dvi dukterines ląsteles. Ciklinai ir nuo ciklino priklausomos kinazės; eukariotų ląstelių dalijimasis; mitozė.
kontrolinis darbas, pridėtas 2009-11-21
Ląstelių teorijos kūrėjai. Archėjų ir melsvadumblių ypatybės. Gyvų organizmų filogenija. Eukariotinės ląstelės sandara. Membranų mobilumas ir sklandumas. Golgi aparato funkcijos. Simbiotinė pusiau autonominių organelių kilmės teorija.
pristatymas, pridėtas 2014-04-14
Ląstelės sandaros elementai ir jų charakteristikos. Membranos, branduolio, citoplazmos, ląstelės centro, ribosomų, endoplazminio tinklo, Golgi komplekso, lizosomų, mitochondrijų ir plastidžių funkcijos. Skirtingų organizmų karalysčių atstovų ląstelių sandaros skirtumai.
pristatymas, pridėtas 2013-11-26
Ląstelių pažeidimo tipai. Lėtinio ląstelių pažeidimo stadijos. Ląstelių mirties tipai. Nekrozė ir apoptozė. Pažeidimų patogenezė ląstelių membranos. Labai specializuotos ląstelės aukštas lygis intraląstelinė regeneracija. Jungiamojo audinio būklės.
pristatymas, pridėtas 2013-11-03
Elementarus genetinis ir struktūrinis-funkcinis biologinė sistema. Ląstelių teorija. Tipai ląstelių organizacija. Prokariotinės ląstelės struktūros ypatumai. Eukariotinės ląstelės organizavimo principai. Paveldimas ląstelių aparatas.
kontrolinis darbas, pridėtas 2014-12-22
Struktūra gyvūnų ląstelė. Pagrindinės ląstelių teorijos nuostatos, prokariotų ir eukariotų samprata. Citoplazmos ir endoplazminio tinklo struktūra. Žmogaus chromosomų rinkinys. Ląstelių dalijimosi metodai (amitozė, mitozė ir mejozė) ir jų cheminė sudėtis.
pristatymas, pridėtas 2013-10-09
Suskirstymas eukariotinės ląstelės organizacijoje. Linijiniai eukariotinės ląstelės matmenys. Branduolinis-citoplazminis santykis. Įvairios formos chondrioma. Kardiomiocitų mitochondrijų sistema. Žmonių mitochondrijų ligų požymiai.
Ląstelė. Ląstelinių elementų sandara ir sąveika. Ląstelių patologijos elementai.
(Rengiant paskaitą dalyvavo prof. Semjonovas V.V., ast. Košpajeva E.S., as. Koločkova E.V.)
SVARSTYTI KLAUSIMAI.
1. ĮVADAS
2. Struktūriniai komponentai ląstelės.
2.1. Plazminė membrana ir jos vaidmuo ląstelės gyvenime.
2.1.1. Plazminės membranos struktūra.
2.1.2. Plazminės membranos funkcijos.
A. transportavimo funkcija.
b. Informacijos perdavimas per plazmos membrana.
2.1.3. Membraninių komponentų (lipidų, baltymų ir glikoproteinų) dalyvavimas patologiniuose procesuose.
A. lipidų peroksidacija.
b. Fosfolipazės aktyvumas.
V. Glikoproteinai ir naviko procesas
2.2. Endoplazminis Tinklelis.
2.3. Golgi kompleksas.
2.4. Lizosomos.
2.5. Ribosomos.
2.6. Mitochondrijos.
2.7. Ląstelių centras.
2.8. Branduolys yra ląstelės valdymo sistema.
2.8.1. Branduolinis apvalkalas.
2.8.2. Nukleoplazma (kariolimfa, branduolio sultys).
2.8.3. Morfofunkcinės charakteristikos ir chromosomų klasifikacija.
A. Chromatinas yra euchromatinas ir heterochromatinas.
b. Nukleosoma.
2.8.4. Branduolys.
2.8.5. branduolinė matrica.
2.9. citoplazminiai intarpai.
A. trofiniai intarpai.
b. Sekretorė.
V. Pigmentuotas.
d) išskyrimo.
Šioje paskaitoje nutolsime nuo tradicinio morfofunkcinių (struktūros ir funkcijos) ląstelės ypatybių pateikimo, būdingo mokyklinei mokymo metodikai. Mes sutelksime dėmesį į funkcinę ląstelės ir jos elementų elgesio pusę. Tai darydami pradedame nuo trys svarbūs medicinos biologijos principais. Pirma, ląstelė, kaip vientisa sistema, yra ne išorinėje natūralioje aplinkoje, o vientiso organizmo viduje. , savotiškoje skystoje tarpląstelinėje terpėje. Ir tai nėra atsitiktinumas. Gyvų būtybių evoliucija prasidėjo vandens aplinkoje. Beveik visi pagrindiniai ląstelės morfologiniai dariniai susiformavo vandens aplinkoje. Jie yra tam pritaikyti. Perėjus į sausumą, į dujinę aplinką, reikėjo gerokai pakeisti visą vandens aplinkai pritaikytų gyvių organizaciją. Dabar sunku įsivaizduoti, dėl kokių priežasčių į sausumą patekusių organizmų evoliucija dujinėje aplinkoje negalėjo visiškai pašalinti visų adaptacijų, kurias organizmai gavo būdami skystoje aplinkoje. Gali būti, kad evoliucijai dujinėje aplinkoje prireikė tokių radikalių prie vandens aplinkos pritaikytų gyvųjų sistemų pertvarkymų, kad jie tiesiog nebuvo suderinami su gyvybe. Pagal kitą versiją, prie vandens aplinkos prisitaikiusių organizmų evoliucija dujų aplinkoje buvo neįmanoma. Arba buvo kokių nors kitų priežasčių, bet bet kuriuo atveju Gamta padarė kompromisą – į sausumą išlindę organizmai turėjo pirminio vandenyno aplinką, prie kurios jie yra prisitaikę. Tiesą sakant, kraujas, limfa, tarpląstelinis skystis savo sudėtimi primena mūsų vystymosi lopšį, pirminį vandenyną, kuriame įvyko pirmasis mūsų evoliucijos etapas. Ląstelė, būdama kūno viduje, praktiškai nesiliečia su išorine aplinka; Visa jos veikla vyksta intersticinis skystis, kuri užtikrina ne tik ląstelės egzistavimą, bet ir yra jos medžiagų apykaitos restruktūrizavimo iniciatorė. Šis pertvarkymas perkelia ląstelę į naują gyvenimo būdą, į kitokį funkcinė būklė. Tačiau toks perėjimas galimas dėl antrojo svarbaus principo – ląstelės subląstelinių organelių struktūra ir funkcijos nėra griežtai nulemtos, jos plastiškos, galinčios keistis tam tikrose ribose. O kadangi tarpląsteliniai elementai dalyvauja įvairiuose biocheminiuose procesuose, bet kokie ląstelės struktūros pokyčiai tikrai lems šios struktūros atliekamų funkcijų pasikeitimą. Gyvui būdinga, kad vienas ir tas pats ląstelinis elementas gali atlikti kelias funkcijas. Pavyzdžiui, tai gerai iliustruojama toliau, kai išvardijamos, pavyzdžiui, organelių funkcijos. Ir galiausiai būtina prisiminti trečiąjį principą: visi tarpląsteliniai elementai ir procesai yra viena tarpusavyje susijusi sistema,šis elementų ir procesų rinkinys gali būti pavaizduotas kaip tam tikras tinklas, kuriame vienos ląstelės ar mazgo pasikeitimas lemia visos tarpląstelinės organizacijos ir jos funkcijos pasikeitimą. Šis principas turi didelę reikšmę medicinoje, nes kartais atsirandantys pakitimai būna tokie stiprūs, kad sutrinka sklandi vidinė ląstelės organizacija. Šiuo atveju procesai, atsakingi už savireguliaciją, prisitaikymą ir kt ląstelių funkcijos gali tapti patologijos vystymosi arena iš pradžių tarpląstelinių elementarių struktūrų ir procesų lygmeniu, vėliau visos ląstelės patologijos lygmeniu, kaip elementari savireguliuojanti gyvoji sistema, o vėliau susivienijusių ląstelinių darinių lygmeniu. pagal baigtinę funkciją.
2. Eukariotinės ląstelės struktūriniai komponentai
Įjungta lauke ląstelių (1 pav.) yra išorinė plazminė membrana, kuri atskiria ląstelę nuo išorinė aplinka. Po juo yra citoplazma ir branduolys. Branduolys ne visada yra ląstelės centre. Tais atvejais, kai intensyviai dirbama vienoje kameros dalyje, pvz. vyksta aktyvus absorbcijos procesas maistinių medžiagų išeikvojus energiją, branduolys pasislenka į priešingą „neveikiančią“ ląstelės sekciją, o mitochondrijos susitelkia „darbinėje“ sekcijoje. Savo ruožtu citoplazma ir branduolys susideda iš kelių komponentų, kurie parodyti 1 paveiksle.
Ląstelės struktūrinė organizacija yra pagrįsta membranos principas. Tai reiškia, kad membranos yra esminė ląstelės struktūros dalis. Jie atskiria ląstelę nuo supančios tarpląstelinės aplinkos, padalija ląstelę į atskirus skyrius – skyrius. Šiose izoliuotose srityse yra specifinių medžiagų apykaitos procesai. Ši evoliucijos diferenciacija padidino ląstelės organizaciją, bet tuo nesibaigė. Laikui bėgant kai kurie biocheminiai procesai skyriuose tapo tokie specifiniai, kad iškilo būtinybė sukurti labiau izoliuotas, beveik autonomiškas struktūras – organelius. Membraną turinčios organelės apima mitochondrijas, plastidus, lizosomas, endoplazminį tinklą ir Golgi kompleksą. Šiose tilto galvutėse buvo gyvybiškai svarbūs organizmui procesai. Tuo pačiu metu buvo kuriamos struktūros, kurios neturėjo membraninio apvalkalo, bet atliko ir tam tikras ląstelių funkcijas – ribosomos, ląstelės centras ir kt.. Tuo pačiu metu ląstelės citoplazma išlieka izoliuota endoplazminio tinklo membrana, t. nors ir ne visiškai, į atskirus skyrius, kuriuose vyksta tam tikros ląstelių metabolizmo grandys, pavyzdžiui, glikolizė. Visos biomembranos pastatytos maždaug vienodai, todėl nagrinėsime tik plazminės membranos struktūrą.

http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image004.gif" border="0" width="104" height="55 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image006.gif" border="0" alt="Parašas:" width="126" height="64 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image008.gif" border="0" width="204" height="48 src=">!}
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image010.gif" border="0" width="12" height="87 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image012.gif" border="0" width="12" height="99 src=">http://xn--d1aacnkch5m.xn-- p1ai/14-bez-rubriki/images/image014.gif" border="0" width="12" height="13 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez- rubriki/images/image016.gif" border="0" width="12" height="81 src=">http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image011. gif" border="0" width="12" height="27 src=">
Lipidai Branduolinės sultys
Baltymų hialoplazma
Organelės Chromosomos
Polisacharidų intarpai
branduolio membrana
Ryžiai. 1. Bendras planas eukariotinės ląstelės struktūra.
2.1. Plazma membrana, jos vaidmuo ląstelės gyvenime
2.1.1. Plazminės membranos struktūra.
Membrana susideda iš dviejų lipidų molekulių sluoksnių (dvisluoksnio), į kuriuos įterpti baltymai. Angliavandeniai yra prijungti prie kai kurių lipidų ir baltymų molekulių. Jų nedaug. Membranos storis yra apie 10 nm (0,00001 mm). Pagrindinė membranos dalis yra ištisinis fosfolipidų molekulių sluoksnis. Tuo skystisį sluoksnį panardintos skirtingos struktūros ir baltymų molekulių funkcijų molekulės. Baltymai ne visiškai padengia lipidų dvisluoksnį sluoksnį, bet yra jame atskirai arba grupėmis. Apskritai tai primena mozaika(2. B. C pav.). Šiuo atžvilgiu vadinamas šiuo metu priimtas membranos modelis skysta mozaika. Baltymai yra pajėgūs judėti palei lipidų sluoksnį. Lipidų sluoksnio molekulės taip pat juda. Akivaizdu, kad membranos molekulių judėjimas keičia pastarųjų fizikines ir chemines charakteristikas, o tai savo ruožtu keičia membranos funkcines charakteristikas. Reikėtų pažymėti, kad daugumos ląstelių plazminė membrana neturi tobulo rutulio formos. Atvirkščiai, joje daug iškilimų, įdubimų, kurie nuolat keičia savo formą ir dydį. Gauta į Pastaruoju metu Rezultatai šiek tiek pakoregavo membranų struktūros teoriją. Buvo įrodyta, kad ne visi membranos baltymai gali judėti, o kai kurios membranų dalys savo struktūra skiriasi nuo klasikinio lipidinio dvisluoksnio.
Fosfolipidų molekulė atrodo kaip galva su dviem uodegomis (2 pav. A). Galvutė (glicerolis) tirpsta vandenyje, hidrofilinė, uodegos (riebalų rūgštys) netirpsta vandenyje, hidrofobinės. Todėl būdami vandenyje, molekulės spontaniškai vandens fazės atžvilgiu užima tam tikrą padėtį. Pradedant nuo vandens molekulių, uodegos išsidėsčiusios giliai lipidiniame sluoksnyje, o vandenyje tirpios galvutės atsuktos į išorinę ir vidinę vandens aplinką (2. B pav.). Lipidų dvisluoksnis vadinamas matrica. Ypatingas dėmesys skiriamas lipidų buvimui ląstelių membranose, kurių uodegoje yra riebalų rūgščių, kurių struktūroje yra dvigubų jungčių, esančių per CH2 grupę (- CH = CH - CH = CH - CH -). Tokios riebalų rūgštys vadinamos nesočiosiomis. Šias rūgštis labiausiai veikia reaktyviosios deguonies rūšys (ROS), kurių nuolat yra visų gyvų būtybių organizme. Jų skaičius ypač didėja su įvairių ligų. Tai gali sukelti neigiamas poveikis kuriuos aptarsime toliau.
Matricoje įsiterpę baltymai (2c pav.) išsidėsto įvairiai. Vieni vidiniame ir išoriniame lipidinio sluoksnio paviršiuje yra membraniniai arba paviršiaus baltymai, kiti yra pusiau panardinti į membraną – pusiau integraliniai baltymai, o kiti prasiskverbia per visą membraną – integraliniai baltymai. Paprastai pusiau integraliniai ir integraliniai baltymai yra sujungti vienu terminu - vidiniais baltymais, nes juos sunku atskirti vienas nuo kito. Integraliniai baltymai dažniausiai randami membranose. Jie gali būti pavaizduoti kaip viena molekulė ir atlikti bet kurią vieną funkciją, arba kaip baltymų grupė (ansamblis). Kiekvienas ansamblio narys atlieka griežtai apibrėžtą vaidmenį. Šie kompleksai taip pat atlieka vieną ar kelias galutines funkcijas.
A B BMsoNormalTable" style="border-collapse: colapse; kraštinė: nėra paraštė-kairė: 6,75 tšk paraštė dešinėje: 6,75 pt;" align="left">
Fosfolipidas
1 galva,
2- arklio uodega
lipidų bisluoksnis
(matrica)
Baltymų molekulių vieta lipidų sluoksnio atžvilgiu: 1 - membraniniai baltymai, 2 - pusiau panardintas baltymas, 3 - integruotas baltymas
Ryžiai. 2. Plazminės membranos schema.
Pažymėtina, kad membranoje funkcionuojantys vientisieji ir paviršiniai baltymai-fermentai gana dažnai keičia savo padėtį. Kai kuriais atvejais sunku nustatyti, kokiam tipui (paviršiniam ar integralui) priklauso tam tikras membranos baltymas. Pavyzdžiui, fermentas fosfolipazė A, nusėdęs ant membranos, yra paviršiaus baltymas, bet tada jis aktyvuojamas, tampa vientisu baltymu ir, sąveikaudamas su dvisluoksniais lipidais, susidaro iš jų. arachidono rūgštis(3 pav.). Pastarieji palieka membraną ir virsta kitais aktyviais junginiais, kurie dalyvauja įvairių patologinių procesų vystyme.
Na+, K+- ATP-azė Adrenoreceptorius Adenilato ciklazė
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image020.gif" border="0" width="12" height="79 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image022.jpg" border="0" width="576" height="193 src="> Ca2+-ATP-ase
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image030.gif" border="0" width="12" height="39 src="> cAMP
Na+, K+ Ca2+ arachidono
rūgštis G-baltymas Glikogenas
Ryžiai. 3. Hipotetinė kai kurių membraninių baltymų lokalizacijos schema
Atvirkščiai, baltymai, dalyvaujantys medžiagų judėjime per membraną, pavyzdžiui, baltymai, dalyvaujantys palengvintoje Na+, K+-ATPazės ar Ca2+-ATPazės difuzijoje, paprastai nekeičia savo padėties, veikdami kaip integralūs baltymai (1 pav.). . 3). Ir galiausiai, kaip jau minėjome, membranoje gali būti sudėtingų kelių baltymų kompleksų, sujungtų į vieną ansamblį viena užduotimi. Tokie kompleksai apima baltymus, dalyvaujančius informacijos signalo perdavimu per membraną (3 pav.). Pastarieji apima kompleksą, kuriame yra trys baltymai - adrenoreceptorius, G-baltymas ir adenilato ciklazė. Visi šie baltymai turi reikšmingas esant normaliai ląstelių veiklai ir esant patologijai. Apie tai kalbėsime žemiau.
Be lipidų ir baltymų, membranoje yra ir angliavandenių, tačiau jie yra ne kaip savarankiški komponentai, o kaip lipidų (glikolipidų) arba baltymų (glikoproteinų) komponentai. Angliavandeniai yra išoriniame plazmalemos paviršiuje.
2.1.2. Plazminės membranos funkcijos.
Plazminė membrana atlieka daugybę funkcijų. Mes išvardijame svarbiausius.
Medžiagų pernešimas per membraną. Medžiagos pernešamos per membraną į abi membranos puses.
Informacijos perdavimas per membraną. Ant membranos informacija iš išorės yra suvokiama, konvertuojama ir perduodama į ląstelę arba iš ląstelės. Membraniniai receptoriai čia vaidina svarbų vaidmenį.
· Apsauginis vaidmuo. a) apsaugo ląstelės turinį nuo mechaniniai pažeidimai, cheminiai reagentai ir biologinė agresija, pavyzdžiui, dėl virusų įsiskverbimo ir kt .;
b) c daugialąstelis organizmas plazmos membranos receptoriai formuoja organizmo imuninę būklę;
c) daugialąsčiame organizme membrana užtikrina fagocitozės reakciją.
- Fermentinis – membranose yra įvairių fermentų (pavyzdžiui, fosfolipazės A ir kt.), kurie atlieka visa linija fermentinės reakcijos. Citoplazmos membranoje esantys glikoproteinai ir glikolipidai kontaktuoja su kitų ląstelių membranomis.
Pažvelkime į kai kurias iš šių funkcijų atidžiau.
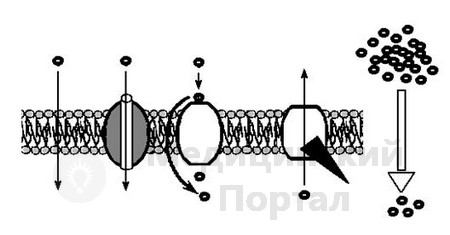
A. transportavimo funkcija. Per membraną juda į ląstelę ir iš jos įvairių medžiagų, įskaitant narkotikus. Priklausomai nuo per membraną pernešamų molekulių dydžio, išskiriami du transportavimo tipai: nepažeidžiant membranos vientisumo ir pažeidžiant membranos vientisumą. Pirmojo tipo transportas gali būti vykdomas dviem būdais – be energijos sąnaudų (pasyvus transportas) ir su energijos sąnaudomis (aktyvus transportas) (žr. 4 pav.). Pasyvus transportas atsiranda dėl difuzijos išilgai elektrocheminio gradiento dėl Brauno atomų ir molekulių judėjimo. Šio tipo transportavimas gali būti atliekamas tiesiogiai per lipidų sluoksnį, nedalyvaujant baltymams ir angliavandeniams, arba naudojant specialius baltymus - translokazes. Riebaluose tirpių medžiagų molekulės daugiausia pernešamos per lipidų sluoksnį, o mažos neįkrautos arba silpnai įkrautos molekulės, tokios kaip vanduo, deguonis, anglies dioksidas, azotas, karbamidas, riebalų rūgštys, taip pat daugelis organinių junginių (pvz. vaistai) gerai tirpsta riebaluose. Translokazės gali transportuoti medžiagą per membranas mažesnės koncentracijos link, neeikvodamos energijos, naudodamos du skirtingus mechanizmus – kanalu, kuris praeina baltymo viduje, arba sujungdamos iš membranos išsikišusios baltymo dalį su medžiaga, kompleksą paversdamos 1800 m. ir medžiagos atskyrimas nuo voverės. Medžiagų difuzija per membraną dalyvaujant baltymams yra svarbi tuo, kad tai vyksta daug greičiau paprasta difuzija per lipidų sluoksnį nedalyvaujant baltymams. Todėl difuzija, kurioje dalyvauja translokazės, vadinama palengvinta difuzija. Pagal šį principą kai kurie jonai (pavyzdžiui, chloro jonai) ir polinės molekulės, taip pat gliukozė yra pernešami į ląstelę.
Aktyvus medžiagų pernešimas per membraną pasižymi trimis savybėmis:
1. Aktyvus transportavimas vykdomas atsižvelgiant į koncentracijos gradientą.
2. Atlieka baltymas nešiklis.
3. Ateina su energijos kaina.
Energija aktyviai perduodant medžiagas yra būtina, kad medžiaga būtų perkelta prieš jos koncentracijos gradientą. Aktyvios perdavimo sistemos dažnai vadinamos membraniniais siurbliais. Energiją šiose sistemose galima gauti iš įvairių šaltinių, dažniausiai toks šaltinis yra ATP. Fosfatinių jungčių skilimą ATP atlieka integruota baltymo-fermento ATP-azė. Todėl šis fermentas randamas daugelio ląstelių membranoje vientiso baltymo pavidalu. Svarbu, kad šis fermentas ne tik atpalaiduotų energiją iš ATP, bet ir vykdytų materijos judėjimą. Todėl aktyvioji perdavimo sistema dažniausiai susideda iš vieno baltymo – ATPazės, kuri gauna energiją ir judina medžiagą. Kitaip tariant, judėjimo procesas ir energijos tiekimas ATPazėje yra konjuguoti. Priklausomai nuo to, kokias medžiagas pumpuoja ATP-azė, siurbliai vadinami arba Na+,K+-ATPazė arbaCa2+-ATPazė. Pirmieji reguliuoja natrio ir kalio kiekį ląstelėje, antrieji – kalcį (šio tipo siurblys dažniausiai yra EPS kanaluose). Iš karto atkreipkime dėmesį į tai, kas svarbu medicinos darbuotojai faktas: sėkmingam kalio-natrio siurblio veikimui ląstelė išleidžia apie 30 proc. bazinė medžiagų apykaitos energija. Tai labai didelis tūris. Ši energija išleidžiama tam tikroms natrio ir kalio koncentracijoms palaikyti ląstelėje ir tarpląstelinėje erdvėje; - ląstelėje yra daugiau kalio nei tarpląstelinėje erdvėje, natrio, atvirkščiai, tarpląstelinėje erdvėje yra daugiau nei ląstelėje. Šis pasiskirstymas, toli gražu ne osmosinė pusiausvyra, suteikia daugiausiai optimalus režimas ląstelių darbas.
 |
Ryžiai. 4. Medžiagų pernešimo per membraną tipų klasifikacija.
Aktyvaus pernešimo metu per membraną juda neorganiniai jonai, aminorūgštys ir cukrus, beveik visos vaistinės medžiagos, turinčios polines molekules. para-aminobenzenkarboksirūgštis, sulfonamidai, jodas, širdies glikozidai, B grupės vitaminai, kortikosteroidiniai hormonai ir kt.
Medžiagų pernešimo per membraną proceso vizualiai iliustracijai pateikiame (su nedideliais pakeitimais) 5 paveikslą, paimtą iš B. Albertso ir kitų lyderiais laikomų mokslininkų knygos „Molekulinė ląstelės biologija“ (1983). teorijos raidą
 http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image036.gif" border="0" width="38" height="19 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image038.gif" border="0" width="33" height="16 src="> vežėjo voverė
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image036.gif" border="0" width="38" height="19 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image038.gif" border="0" width="33" height="16 src="> vežėjo voverė
Pasyvus transportas Aktyvus transportas
5 pav. Daug mažų neįkrautų molekulių laisvai praeina per lipidų dvisluoksnį sluoksnį. Įkrautos molekulės, didelės neįkrautos molekulės ir kai kurios mažos neįkrautos molekulės praeina per membranas per kanalus ar poras arba naudojant specifinius nešiklius. Pasyvus transportas visada nukreiptas prieš elektrocheminį gradientą link pusiausvyros. Aktyvus transportavimas vyksta prieš elektrocheminį gradientą ir reikalauja energijos sąnaudų.
transmembraninis pernešimas, atspindi pagrindinius medžiagų pernešimo per membraną tipus. Reikėtų pažymėti, kad baltymai, dalyvaujantys transmembraniniame transporte, yra integralūs baltymai ir dažniausiai juos atstovauja vienas sudėtingas baltymas.
Didelės molekulinės masės baltymų molekulės ir kitos didelės molekulės per membraną patenka į ląstelę endocitozės būdu (pinocitozė, fagocitozė ir endocitozė), o iš ląstelės – egzocitozės būdu. Visais atvejais šie procesai skiriasi nuo minėtųjų tuo, kad pernešta medžiaga (dalelė, vanduo, mikroorganizmai ir kt.) pirmiausia supakuojama į membraną ir tokia forma pernešama į ląstelę arba iš jos išsiskiria. Pakavimo procesas gali vykti tiek plazminės membranos paviršiuje, tiek ląstelės viduje.
b. Informacijos perdavimas per plazmos membraną.
Be baltymų, dalyvaujančių medžiagų pernešime per membraną, joje buvo nustatyti sudėtingi kelių baltymų kompleksai. Erdviškai atskirtus juos vienija viena baigtinė funkcija. Kompleksiniai baltymų ansambliai apima baltymų kompleksą, atsakingą už labai galingos biologiškai aktyvios medžiagos - cAMP (ciklinio adenozino monofosfato) - gamybą ląstelėje. Šiame baltymų ansamblyje yra ir paviršiaus, ir vientisų baltymų. Pavyzdžiui, vidiniame membranos paviršiuje yra paviršiaus baltymas, kuris vadinamas G - baltymu. Šis baltymas palaiko ryšį tarp dviejų gretimų integruotų baltymų – baltymo, vadinamo adrenalino receptoriumi, ir baltymo – fermento – adenilato ciklazės. Adrenoreceptorius gali susijungti su adrenalinu, kuris iš kraujo patenka į tarpląstelinę erdvę ir susijaudina. Šis sužadinimo G-baltymas pereina į adenilato ciklazę – fermentą, galintį gaminti veikliąją medžiagą cAMP. Pastaroji patenka į ląstelės citoplazmą ir aktyvuoja joje įvairius fermentus. Pavyzdžiui, suaktyvinamas fermentas, kuris suskaido glikogeną į gliukozę. Gliukozės susidarymas padidina mitochondrijų aktyvumą ir ATP, kuris patenka į visus ląstelių skyrius kaip energijos nešiklis, sintezę, sustiprindamas lizosomų, natrio-kalio ir kalcio membraninių siurblių, ribosomų ir kt. . galiausiai padidina beveik visų organų, ypač raumenų, gyvybinę veiklą. Šis pavyzdys, nors ir labai supaprastintas, parodo, kaip membranos veikla yra susijusi su kitų ląstelės elementų darbu. Namų ūkio lygmeniu ši sudėtinga schema atrodo gana paprasta. Įsivaizduokite, kad šuo staiga užpuolė žmogų. Atsiradęs baimės jausmas veda į adrenalino išsiskyrimą į kraują. Pastarasis jungiasi prie plazmos membranos adrenoreceptorių, o keičiasi cheminė struktūra receptorius. Tai savo ruožtu lemia G baltymo struktūros pokyčius. Pakeistas G baltymas gali aktyvuoti adenilato ciklazę, kuri padidina cAMP gamybą. Pastarasis skatina gliukozės susidarymą iš glikogeno. Dėl to sustiprėja daug energijos sunaudojančios ATP molekulės sintezė. Patobulintas švietimas energijos žmonėms raumenyse sukelia greitą ir stipri reakcijaį šuns puolimą (lėkimas, gynyba, kova ir kt.).
2.1.3. Membraninių komponentų (lipidų, baltymų ir glikoproteinų) dalyvavimas patologiniuose procesuose.
Visų pirma, reikia pažymėti, kad beveik bet koks poveikis neigiami veiksniai membranoje vyksta du procesai
1. Suaktyvinama lipidų peroksidacija.
2. Suaktyvėja fosfolipazės veikla.
Pirmasis procesas susijęs su lipidų sluoksniu, antrasis - baltymų sluoksniu.
A. lipidų peroksidacija.
Jau sakėme, kad fosfolipidinės membranos molekulės pagrindas yra glicerolis ir 2 riebalų rūgštys. Šios rūgštys gali būti sočiosios arba nesočiosios. Nesočiųjų rūgščių molekulėje yra dvigubų jungčių tarp anglies atomų (c - C \u003d C - C \u003d C - C \u003d C -). Tokių ryšių nebuvimas apibūdina sočiąsias riebalų rūgštis (-C-C-C-C-C-). Nesočiosios riebalų rūgštys lengvai sąveikauja su reaktyviosiomis deguonies rūšimis (ROS), kurios gali patekti į membraną iš ląstelės ar tarpląstelinės erdvės. ROS yra stiprūs reagentai ir reaguoja su beveik visais žinomais organiniais junginiais. (baltymai, esteriai, aminorūgštys, DNR, RNR ir kt.). ROS yra pavojingos, nes jų reakcijos nekontroliuojamos, reaguoja su visomis organinėmis molekulėmis be išimties, su kuriomis susitinka. Šios reakcijos veda į sunaikinimą organinės medžiagos, specifinės veiklos praradimas. Ląstelės viduje ROS susidaro įvairiose vietose – organelėse, citoplazmoje, tokiais kiekiais, kurie nekelia pavojaus ląstelei. Taip yra dėl to, kad ląstelėje yra galinga apsaugos nuo ROS sistema – vitaminai, kai kurie ląstelių apykaitos produktai, baltymai ir kiti junginiai. Jei ląstelė patenka nepalankios sąlygos( traumos, virusinės ar užkrečiamos ligos, autoimuniniai konfliktai ir kt.), tada ROS generacija ląstelės viduje didėja ir jų lygis pradeda viršyti fiziologines galimybes. apsauginis barjeras. Šiuo atveju ROS pasiskirsto visame ląstelės tūryje. Patekę į membraną, jie pirmiausia sąveikauja su nesočiosiomis riebalų rūgštimis. Dėl to susidaro įvairūs membranai nebūdingi junginiai. Kai kurie iš jų yra labai reaktyvūs ir vadinami laisvaisiais radikalais. Jie itin pavojingi, nes geba sąveikauti su beveik bet kokiais organiniais junginiais – baltymais, riebalais, angliavandeniais, DNR, RNR ir kt. su kuriais jie bendrauja. Būdami membranoje, laisvieji radikalai sąveikauja su visomis organinėmis membranos molekulėmis, pažeisdami jos vientisumą. Laisvieji radikalai gali palikti membraną ląstelės viduje – tada sąveikauja su įvairiais citoplazmos komponentais, organelėmis, branduoliu. Jei jie patenka į tarpląstelinę erdvę, tada jie gali patekti į kraują nešiotojų pagalba ir išplisti visame kūne. Atkreipkite dėmesį, kad laisvieji radikalai, kaip taisyklė, yra stiprūs mutagenai, keliantys grėsmę lytinių ląstelių genetinei medžiagai (galima paveldimos patologijos atsiradimas) ir somatinėms ląstelėms (somatinę ląstelę galima paversti piktybine). dėl mutacijos). Yra ir kitas pavojus. Dėl sąveikos laisvieji radikalai su nesočiosiomis riebalų rūgštimis gali susidaryti vandenyje tirpūs junginiai, dėl kurių per membraną gali nekontroliuojamai prasiskverbti įvairiausių junginių, tokių kaip jonai. Kitaip tariant, ištisiniame dvigubame lipidų sluoksnyje susidaro kanalas, per kurį į ląstelę gali patekti ir iš jos išeiti įvairūs jonai, ypač natrio ir kalio. Paprastai fiziologinėmis sąlygomis ląstelėje kalio koncentracija yra didesnė nei tarpląstelinėje erdvėje, o natrio koncentracija, priešingai, ląstelėje yra mažesnė nei tarpląstelinėje erdvėje. Ši būsena yra būtina, kad ląstelė normaliai funkcionuotų, ir ją palaiko Nuolatinis darbas specialus kalio-natrio siurblys, esantis membranos storyje. Natrio ir kalio jonai praeina per jį griežtai dozuotu būdu. Jei atsiras nenatūralus kanalas, juo prasidės nekontroliuojamas natrio ir kalio judėjimas. Į ląstelę patekęs natris susijungs su chloru ir sudarys druską. Osmosinis slėgis ląstelės viduje padidės ir vanduo tekės į ląstelę. Ląstelė išsipūs ir užblokuos siauras kapiliarų lovas. Dėl to sumažės kitų ląstelių aprūpinimas deguonimi ir maistinėmis medžiagomis. Ląstelėse trūkstant deguonies ir mitybos, ROS generacija automatiškai padidės ir jos vėl pateks į membraną iš citoplazmos, kur vėl sąveikaus su nesočiosiomis riebalų rūgštimis. Kaip minėjome anksčiau, tai sukels laisvųjų radikalų susidarymą ir nenatūralių kanalų atsiradimą membranoje. Tai sustiprins natrio patekimą į ląstelę ir joje susidarys NaCl, iškart prasidės vandens judėjimas į ląstelę, ji išsipūs, o po to susiaurės artimiausi kapiliarai. ir Kaimyninėse ląstelėse vėl pradės trūkti deguonies ir maistinių medžiagų. Scenarijus kartosis. Mes palietėme tik nedidelį fragmentą įvykių, kurie vyksta membranoje patologijos metu. Bet tai taip pat aiškiai matoma svarbus principas būdingas bet kuriai patologijai – procesas, atsiradęs vienoje ląstelės vietoje, dėl tam tikrų priežasčių, vėliau plinta, gaudydamas kitus objektus, sustiprėja ir galiausiai gali sukelti nepataisomų pasekmių. Aišku, kaip svarbu šioje besiskleidžiančių procesų grandinėje rasti silpnąją grandį, kurią įtakojant būtų galima blokuoti patologijos vystymąsi.
b. Fosfolipazės aktyvumas.
Šis procesas yra susijęs su fermento, esančio ant membranos – fosfolipazės A, aktyvavimu. Apie šį fermentą jau rašėme aukščiau – jis kai kuriuos matricos lipidus paverčia arachidono rūgštis. Pastarieji lipidų dvisluoksnį sluoksnį palieka ląstelės citoplazmoje ir virsta daugybe aktyvių junginių, kurių dalis lieka ląstelėje, keičia jos metabolizmą, dalis palieka ląstelę į tarpląstelinę erdvę, paveikdama kaimynines ląsteles, pernešdama jas į naujas veikimo režimas. Kita dalis veikliosios medžiagos patenka į kraują, plinta visame kūne ir veikia tolimas ląsteles, taip pat modifikuodamas jų funkciją. Pateiksime pavyzdį (6 pav.). bronchuose, aplink kraujagyslės, yra daug putliųjų ląstelių. Per
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image045.gif" border="0" width="464" height="137 src="> mRNR ribosoma
Ryžiai. 7. Endoplazminis tinklas:
A – sklandžiai ER fragmentai; B – grubaus EPS fragmentai. C – veikianti ribosoma esant grubiam ER.
Lygioje ER membranoje yra fermentų rinkinys, kuris sintetina riebalus ir paprasti angliavandeniai, taip pat organizmui būtinų steroidinių hormonų. Ypač atkreiptinas dėmesys į tai, kad kepenų ląstelių lygaus EPS membranoje yra fermentų sistema, skaidanti į ląstelę patekusias svetimas medžiagas (ksenobiotikus), įskaitant vaistinius junginius. Sistema susideda iš įvairių baltymų fermentų (oksidatorių, reduktorių, acetilatorių ir kt.).
Ksenobiotinis arba vaistinė medžiaga(LP), nuosekliai sąveikaudamas su tam tikrais fermentais, keičia savo cheminę struktūrą. Dėl to galutinis produktas gali išlaikyti specifinį aktyvumą, gali tapti neaktyvus, arba atvirkščiai, įgyti naują savybę – tapti toksiškas organizmui. ER esanti fermentų sistema, vykdanti cheminę ksenobiotikų (arba LS) transformaciją, vadinama sistema biotransformacija.Šiuo metu šiai sistemai teikiama didelė reikšmė, nes. specifinis vaistų aktyvumas (baktericidinis aktyvumas ir kt.) organizme ir jų toksiškumas priklauso nuo jo darbo intensyvumo ir kiekybinio tam tikrų fermentų kiekio jame. http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image047.gif" border="0" width="216" height="105 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image049.jpg" border="0" width="287" height="252 src="> Citoskeleto elementai
 |
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image052.gif" border="0" width="107" height="50 src="> Ribosoma
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image054.gif" border="0" width="31" height="53 src=">
Branduolinė ląstelė
Ryžiai. 8. Scheminis ląstelės vidaus vaizdas (ne pagal mastelį).
Reikėtų pažymėti svarbų EPS vaidmenį kuriant visas tarpląstelines membranas. Čia prasideda pats pirmasis tokios statybos etapas.
EPS taip pat vaidina svarbų vaidmenį keičiantis kalcio jonais. Šis jonas turi didelę reikšmę reguliuojant ląstelių metabolizmą, keičiant membranų kanalų pralaidumą, aktyvinant įvairius junginius citoplazmoje ir kt. Smooth ER yra kalcio jonų saugykla. Jei reikia, kalcis išsiskiria ir dalyvauja ląstelės gyvenime. Ši funkcija labiausiai būdinga raumenų ER. Kalcio jonų išsiskyrimas iš EPS yra ryšys sudėtingas procesas raumenų susitraukimai.
Pažymėtina glaudus EPS ryšys su mitochondrijomis – ląstelės energijos stotimis. Sergant ligomis, susijusiomis su energijos trūkumu, ribosomos atsiskiria nuo šiurkščios ER membranos. Pasekmes nuspėti nesunku – sutrinka eksportui skirtų baltymų sintezė. O kadangi virškinimo fermentai yra tarp tokių baltymų, tai sergant ligomis, susijusiomis su energijos trūkumu, sutriks virškinimo liaukų darbas ir dėl to nukentės viena pagrindinių organizmo funkcijų – virškinimo. Atsižvelgiant į tai, turėtų būti sukurta farmakologinė gydytojo taktika.
2.3. Golgi kompleksas
liaukose vidinė sekrecija, pavyzdžiui, kasoje kai kurios pūslelės, atsiskirdamos nuo EPS, išsilygina, susilieja su kitomis pūslelėmis, persidengia viena su kita, kaip blynai krūvoje, sudarydami Golgi kompleksą (CG). Jį sudaro keli konstrukciniai elementai – rezervuarai, burbuliukai ir vamzdeliai (9 pav.). Visus šiuos elementus sudaro vienasluoksnė skystos mozaikos membrana. Talpyklose burbuliukų turinys „brandina“. Pastarieji yra surišti iš komplekso ir juda citozolyje išilgai mikrovamzdelių, fibrilių ir gijų. Tačiau pagrindinis burbuliukų kelias yra judėti link plazminės membranos. Susiliedamos su juo pūslelės ištuština savo turinį su virškinimo fermentais į tarpląstelinę erdvę (10 pav.). Iš jo fermentai patenka į lataką ir supilami į žarnyną. Išskyrimo procesas su CG sekrecijos pūslelėmis vadinamas egzocitoze.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image056.gif" border="0" width="150" height="18 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image058.gif" border="0" width="34" height="12 src="> EPS
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image060.gif" border="0" width="12" height="39 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image062.jpg" border="0" width="336" height="226 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image064.gif" border="0" width="47" height="12 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image066.gif" border="0" width="27" height="12 src="> Membrana
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image072.gif" border="0" width="27" height="12 src=">ląstelių membranos iš
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image074.gif" border="0" width="51" height="18 src=">burbulinė membrana.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image076.gif" border="0" width="26" height="67 src=">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image078.gif" border="0" width="155" height="14 src=">iš KG
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image080.jpg" border="0" width="456" height="259 src=">
Mikroorganizmai
Ištirpęs
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image084.gif" border="0" width="17" height="25 src=">medžiagos
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image086.gif" border="0" width="19" height="29 src="> 4
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image088.gif" border="0" width="34" height="25 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image090.gif" border="0" width="39" height="11 src=">http://xn--d1aacnkch5m.xn-- p1ai/14-bez-rubriki/images/image092.gif" border="0" width="31" height="48 src="> 1a 4a
Baltymai, riebalai Lizosomų fragmentai
mitochondrijų angliavandenių
Ryžiai. 12. Lizosomų funkcijos:
1, 1a - hialoplazmos organinių medžiagų panaudojimas; 2, 2a - pinocitinių pūslelių turinio panaudojimas; 3, 3a - fagocitinių pūslelių turinio panaudojimas; 4, 4a - fermentinis pažeistų mitochondrijų skilimas. 3a - fagosomos.
nye organinių junginių, kurie, patekę į citoplazmą, tampa ląstelių metabolizmo dalyviais. Kai kuriose ląstelėse lizosomose esančios biogeninės makromolekulės gali nesibaigti. Tokiu atveju nesuvirškinti produktai kaupiasi lizosomos ertmėje. Tokia lizosoma vadinama likutiniu kūnu. Ten taip pat nusėda pigmentai. Žmonėms, organizmui senstant, senėjimo pigmentas lipofuscinas kaupiasi smegenų ląstelių, kepenų ir raumenų skaidulų likutiniuose kūnuose.
Jeigu tai, kas išdėstyta, sąlyginai galima apibūdinti kaip lizosomų veikimą ląstelės lygmenyje, tai kita šių organelių veiklos pusė pasireiškia viso organizmo, jo sistemų ir organų lygmeniu. Visų pirma, tai susiję su organų, kurie miršta embriogenezės metu (pavyzdžiui, buožgalvio uodega), diferencijuojant tam tikrų audinių ląsteles (kremzlės pakeitimas kaulu) ir kt.
Atsižvelgiant į didelę lizosomų fermentų svarbą ląstelės gyvenime, galima daryti prielaidą, kad bet koks jų darbo sutrikimas gali sukelti sunkios pasekmės. Jei genas, kontroliuojantis bet kurio lizosomų fermento sintezę, bus pažeistas, pastarųjų struktūra bus sutrikdyta. Tai lems tai, kad „nesuvirškinti“ produktai kaupsis lizosomose. Jei ląstelėje tokių lizosomų yra per daug, ląstelė pažeidžiama ir dėl to sutrinka atitinkamų organų darbas. paveldimos ligos vystosi pagal šį scenarijų, vadinamos „lizosomų kaupimosi ligomis“.
Taip pat reikėtų atkreipti dėmesį į lizosomų dalyvavimą formuojant imuninė būklė organizmas (13 pav.). Patekęs į organizmą antigenas (pavyzdžiui, mikroorganizmo toksinas) daugiausia (apie 90%) sunaikinamas, o tai apsaugo ląsteles nuo žalingo jo poveikio. Kraujyje likusias antigeno molekules absorbuoja (pinocitozės arba fagocitozės būdu) makrofagai arba specialios ląstelės su išvystyta lizosomų sistema.
Bakterija
 |
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image099.gif" border="0" width="444" height="244 src="> Antigenas
Makrofagas
pinocitozė
pinocitinis
Lizosoma
Antigeno peptidiniai fragmentai
Ryžiai. 13. Antigeno peptidų fragmentų susidarymas makrofage
(svarstyklių nepastebėta).
tema. Pinocitinė pūslelė arba fagosoma su antigenu susijungia su lizosoma, o pastarosios fermentai suskaido antigeną į fragmentus, kurie turi didesnį antigeninį aktyvumą ir mažesnį toksiškumą nei pirminis mikrobinis antigenas. Šie fragmentai yra dideliais kiekiais iškeliamas į ląstelių paviršių, ir įvyksta galingas aktyvavimas imuninės sistemos organizmas. Akivaizdu, kad antigeninių savybių sustiprėjimas (nesant toksinio poveikio) dėl lizosominio gydymo žymiai pagreitins apsauginių imuninių atsakų į šį mikroorganizmą vystymąsi. Procesas, kurio metu lizosomos suskaido antigeną į peptidų fragmentus, vadinamas antigenų apdorojimas. Reikėtų pažymėti, kad EPS ir Golgi kompleksas yra tiesiogiai susiję su šiuo reiškiniu.
Ir, galiausiai, pastaruoju metu buvo plačiai svarstomas lizosomų ir ląstelės fagocituojamų mikroorganizmų ryšio klausimas. Kaip minėjome anksčiau, fagosomos ir lizosomos susiliejimas sukelia mikroorganizmų virškinimą fagolizosomoje. Tai yra labiausiai palankus rezultatas. Tačiau galimi ir kiti santykiai. Taigi kai kurie patogeniniai (patogeniniai) mikroorganizmai, prasiskverbdami į fagosomos viduje esančią ląstelę, išskiria medžiagas, kurios blokuoja lizosomų susiliejimą su fagosoma. Tai leidžia jiems išgyventi fagosomose. Tačiau ląstelių (fagocitų) gyvenimo trukmė su absorbuotais mikroorganizmais yra trumpa, jos suyra, į kraują išskirdamos fagosomas su mikrobais. Į kraują patekę mikroorganizmai vėl gali išprovokuoti ligos atkrytį (grįžimą). Galimas ir kitas variantas, kai sunaikintų fagocitų dalis, įskaitant fagosomas su mikrobais, reabsorbuoja kiti fagocitai, vėl lieka gyvi ir naujoje ląstelėje. Ciklas gali kartotis pakankamai ilgas laikas. Buvo aprašytas šiltinės atvejis senyvo amžiaus ligoniui, kuris, būdamas jaunas Raudonosios armijos karys, susirgo šiltine, kovodamas Pirmojoje kavalerijos armijoje. Po daugiau nei penkiasdešimties metų pasikartojo ne tik ligos simptomai – net kliedesiniai regėjimai sugrąžino senuką į pilietinio karo erą. Esmė ta, kad patogenai šiltinės turi galimybę blokuoti fagosomų ir lizosomų sujungimo procesą.
Lizosomų funkcijos:
virškinimo (virškina citoplazmos dalis ir mikroorganizmus, aprūpina elementariais organiniais junginiais ląstelės poreikiams),
panaudojimas (išvalo citoplazmą nuo suirusių dalių),
dalyvauti pašalinant mirštančias ląsteles ir organus,
Apsauginis (mikroorganizmų virškinimas, dalyvavimas imuninės reakcijos organizmas).
2.5. Ribosomos.
Tai ląstelės baltymų sintezės aparatas. Ribosomoje yra du subvienetai, didelis ir mažas. Subvienetai turi sudėtingą konfigūraciją (žr. 14 pav.) ir susideda iš baltymų ir ribosominės RNR (rRNR). Ribosominė RNR tarnauja kaip tam tikras karkasas, ant kurio yra pritvirtintos baltymų molekulės.
Ribosomų susidarymas vyksta ląstelės branduolio branduolyje (šis procesas bus aptartas toliau). Susidarę dideli ir maži subvienetai per branduolio poras išeina į citoplazmą.
Citoplazmoje ribosomos yra disocijuotos arba išsklaidytos būsenos, tai disocijuotos ribosomos. Šioje būsenoje jie negali prisitvirtinti prie membranos. Tai nėra ribosomos darbinė būsena. Darbinėje būsenoje ribosoma yra organoidas, susidedantis iš dviejų tarpusavyje sujungtų subvienetų, tarp kurių praeina mRNR grandinė. Tokios ribosomos gali laisvai „plaukti“ citozolyje, jos vadinamos laisvos ribosomos arba pritvirtinti prie įvairių membranų,
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image106.gif" border="0" width="444" height="240 src=">yra vadinamas polisomas(15 pav.).
Baltymų sintezės pabaiga Baltymų sintezės pradžia
Ryžiai. 15. Polisomų baltymų sintezės schema.
Paveiksle polisomas sudarytas iš penkių skirtingų ribosomų.
Paprastai baltymai sintetinami ant grubios ER membranų eksportui, o hialoplazmoje – ląstelės poreikiams. Jei ligos metu nustatomas ribosomų atsiskyrimas nuo membranų ir jų perėjimas į hialoplazmą, tai gali būti laikoma gynybinė reakcija- viena vertus, ląstelės mažina baltymų eksportą ir padidina baltymų sintezę vidiniams poreikiams tenkinti. Kita vertus, toks ribosomų atsiskyrimas rodo artėjantį ląstelės energijos trūkumą, nes ribosomų pritvirtinimui ir sulaikymui ant membranų reikalinga energija, kurios pagrindinis tiekėjas ląstelėje yra ATP. ATP trūkumas natūraliai lemia ne tik ribosomų atsiskyrimą nuo membranos, bet ir laisvų ribosomų nesugebėjimą prisitvirtinti prie membranos. Dėl to efektyvus baltymų generatorius – grubus ER – pašalinamas iš ląstelės molekulinės ekonomikos. Energijos trūkumas laikomas rimtas pažeidimas ląstelių metabolizmas, dažniausiai susijęs su nuo energijos priklausomų procesų veiklos pažeidimu (pavyzdžiui, mitochondrijose).
Ribosomoje yra trys skirtingos vietos, prie kurių jungiasi RNR – viena yra pasiuntinio RNR (mRNR arba mRNR) ir dvi pernešimo RNR. Pirmasis yra didelio ir mažo subvieneto sąlyčio taške. Iš pastarųjų dviejų vienoje vietoje yra tRNR molekulė ir susidaro ryšiai tarp aminorūgščių (peptidiniai ryšiai), todėl ji vadinama P centru. Jis yra mažame subvienete. O antroji skirta naujai atvežtai tRNR molekulei, pakrautai aminorūgštimi, laikyti. Jis vadinamas A centru.
Reikia pabrėžti, kad baltymų sintezės metu kai kurie antibiotikai gali blokuoti šį procesą (apie tai plačiau aptarsime aprašydami vertimą).
2.6. Mitochondrijos.
Jos vadinamos „ląstelės energijos stotimis“. Eukariotuose glikolizės, Krebso ciklo ir kitų biocheminių reakcijų procese susidaro daug elektronų ir protonų. Dalis jų dalyvauja įvairiose biocheminėse reakcijose, kita dalis kaupiasi specialiuose junginiuose. Yra keli. Svarbiausi iš jų yra NADH ir NADPH (nikotinamido adenino dinukleotidas ir nikotinamido adenino dinukleotido fosfatas). Šie NAD ir NADP pavidalo junginiai yra akceptoriai – savotiški elektronų ir protonų „spąstai“. Prie jų prijungę elektronus ir protonus, jie virsta NADH ir NADPH ir jau yra elementariųjų dalelių donorai. „Pagaudami“ juos įvairiose ląstelės vietose, jie daleles perneša į įvairūs skyriai citoplazmą ir, suteikdami jas biocheminių reakcijų poreikiams, užtikrina nenutrūkstamą medžiagų apykaitos eigą. Tie patys junginiai tiekia elektronus ir protonus į mitochondrijas iš citoplazmos ir iš mitochondrijų matricos, kur yra galingas elementariųjų dalelių generatorius – Krebso ciklas. NADH ir NADPH, integruodamiesi į elektronų transportavimo grandinę (žr. toliau), daleles perduoda ATP sintezei. Energija iš ATP paimama visiems procesams, vykstantiems ląstelėje su energijos sąnaudomis.
Mitochondrijose yra dvi skystos mozaikos membranos. Tarp jų yra tarpmembraninė erdvė. Vidinė membrana turi klostes – cristae (16 pav.). Vidinis kristos paviršius išmargintas grybo formos kūnais, turinčiais stiebą ir galvą.
Grybų kūnuose ATP sintetinamas. Pačiame vidinės mitochondrijų membranos storyje yra fermentų kompleksų, kurie perneša elektronus iš NADH2 į deguonį. Šie kompleksai vadinami kvėpavimo grandinė arba atsinaujinimo grandinė
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image108.gif" border="0" width="51" height="12 src="> Ribosoma
tekstas-dekoravimas: pabraukti;"> Nesidalančiose ląstelėse šį kompleksą sudaro du identiškai išsidėstę centrioliai - motininė ir dukterinė Viena centriolė yra cilindras, kurio sienelė sudaro devynias grupes, grupėje yra trys mikrovamzdeliai. Iš viso 9 trynukai.Centrioliai išsidėstę vienas kitam statmenai (17 pav.).Organoidas dalyvauja formuojant citoskeleto mikrovamzdelius.Judančiose ląstelėse mikrovamzdeliai nuolat skaidomi ir vėl formuojasi.Specializuotose ląstelėse (neuronai), mikrotubuliai yra gana stabilūs.
besidalijančiose ląstelėse abu centrioliai pirmiausia išsiskiria išilgai polių, o tarp jų susidaro mikrovamzdelių sistema. Mikrovamzdeliai savaime susirenka iš baltymų blokų. Vienas iš baltymų, įtrauktų į bloką, vadinamas tubulinas. Centrioliai ir
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image117.jpg" border="0" width="312" height="215 src=">
Ryžiai. 17. Centrioliai: ląstelės centras (A), susidedantis iš dviejų statmenų centriolių (B, C).
mikrovamzdelių sistema tarp centriolių primena verpstę. Iš čia ir kilo visos figūros pavadinimas – padalijimo verpstė. Mikrovamzdeliai mikroskopu atrodo kaip gijos, todėl kartais jie vadinami verpstės gijomis. Mikrovamzdeliai yra prijungti prie chromosomų centromerų ir dalyvauja chromatidžių „traukime“ išilgai besidalijančios ląstelės polių.
Pasibaigus ląstelių dalijimuisi abiejose dukterinėse ląstelėse, prie kiekvieno centriolio susidaro nauja dukterinė kopija, kuri iš karto orientuojasi statmenai motinai. Naujų centriolių dauginimasis atliekamas savaime susirenkant iš baltymų subvienetų, esančių citoplazmoje.
Ląstelių centro funkcijos yra įvairios. Pagrindiniai iš jų yra dalyvavimas ląstelių dalijimuisi ir ląstelių organelių, pino- ir fagosomų transportavimas per citoplazmą. Neseniai pasirodė duomenų, rodančių, kad centrioliai yra pagrindiniai ląstelės erdvės ir laiko koordinatoriai.
Reikėtų pažymėti, kad procesas, vedantis į dalijimosi veleno susidarymą, yra labai jautrus įvairių narkotikų. Vienas iš rudens kolchicum alkaloidų – kolchicinas, derinamas su tubulinas, neleidžia susidaryti mitoziniam velenui ir blokuoja ląstelių dalijimąsi. Tokios medžiagos vadinamos antimitozinėmis medžiagomis. Jie įtraukia vaistai vinblastinas ir vinkristinas, kurie plačiai naudojami vėžio terapijoje.
Sergant šia liga, ląstelės piktybinis navikas intensyviai ir nekontroliuojamai daugintis; dalijimosi veleno pažeidimas sustabdo jų dalijimąsi.
2.8. Branduolys yra ląstelės valdymo sistema
Jį sudaro šios dalys: branduolio membrana, karioplazma (kariolimfa), chromosomos (ląstelių dalijimosi metu) arba chromatinas (tarpfazėje), branduolys ir branduolio skeletas (matrica).
Beveik visa eukariotinės ląstelės DNR tarpfazės metu yra sutelkta branduolyje, kurio tūris sudaro apie 10% viso ląstelės tūrio. Paprastai ląstelėse yra tik vienas branduolys, tačiau kai kurios subrendusios ląstelės gali ir neturėti branduolio. Tai apima žmogaus (žinduolių) periferinio kraujo eritrocitus. Yra ląstelių, turinčių du (kepenų ląstelės, blakstienas) arba daug branduolių (skersinis raumuo).
Branduolys yra apsuptas branduolio membrana, jame yra chromatino, taip pat vienas ar daugiau branduolių (18 pav.).
2.8.1. branduolinis apvalkalas
Susideda iš dviejų membranų – išorinės ir vidinės. Tarpas tarp jų vadinamas perinukleariniu. Išorinė membrana gali patekti į ER membranas. Tai leidžia nuolat keistis perinuklearinės erdvės ir EPS kanalų turiniu. Be to, išorinė membrana gali suformuoti pūsleles, kurios yra įterptos į CG. Galimas ir atvirkštinis procesas – CG suformuotos pūslelės įtraukiamos į branduolio membraną, išliedamos turinį į perinuklearinę erdvę.
Branduolinė membrana yra persmelkta porų, per kurias iš branduolio į citoplazmą pernešama iRNR molekulė, kompleksiniai dariniai iš rRNR ir baltymų (ribosomų pirmtakai) bei kai kurios medžiagos. Poros yra sudėtingas kompleksas, susidedantis iš kelių baltymų rutuliukų.
Iš vidaus vidinės branduolinės membranos paviršių dengia baltymų tinklas - branduolio sluoksnis, dalyvaujantis chromatino ir porų komplekso organizavime. Ląstelių dalijimosi metu lamina dalyvauja sunaikinime branduolinis apvalkalas.
2.8.2. Nukleoplazma (kariolimfa, branduolio sultys) - vidinė aplinka ląstelės
Nestruktūrinės masės pavidalu jis supa chromosomas ir branduolį. Karioplazmos klampumas yra maždaug toks pat kaip hialoplazmos, tačiau rūgštingumas yra didesnis. Jį sudaro vanduo (pagrindinė dalis) ir jame ištirpę organiniai ir neorganiniai junginiai. Daroma prielaida, kad karioplazmoje vyksta specifiniai biocheminiai procesai.
Grubus XPS
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image122.gif" border="0" width="98" height="45 src="> 1
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image124.gif" border="0" width="38" height="41 src=">http://xn- -d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image126.gif" border="0" width="125" height="12 src=">left">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image115.gif" border="0" width="63" height="12 src=">left">
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image134.gif" border="0" height="12 src=">membrana
Sudėtingas
tekstas-dekoravimas: pabraukti;"> Interfazės metu genų raiška (funkcija, darbas) yra maksimali ir chromosomos atrodo kaip plonos gijos. Dekondensuojamos tos gijos atkarpos, kuriose vyksta RNR sintezė, o atkarpos, kuriose vyksta sintezė neatsiranda, priešingai, sutirštėja (19 pav.).
Padalijimo metu kai DNR chromosomose praktiškai nefunkcionuoja, chromosomos yra tankūs kūnai, panašūs į "X" arba "Y". Taip yra dėl stiprios DNR kondensacijos chromosomose.
Ypač svarbu suprasti, kad paveldima medžiaga skirtingai pateikiama ląstelėse, kurios yra tarpfazėje ir dalijimosi metu. Tarpfazėje ląstelėje aiškiai matomas branduolys, paveldima medžiaga, kurioje jį vaizduoja chromatinas. Chromatinas savo ruožtu susideda iš iš dalies kondensuotų chromosomų gijų. Jeigu laikytume ląstelę dalijimosi metu, kai branduolio nebėra, tai visa paveldima medžiaga susitelkia chromosomose, kurios maksimaliai kondensuojasi (20 pav.).
Visų chromosomų gijų, susidedančių iš DNR ir įvairių baltymų, visuma eukariotinių ląstelių branduoliuose vadinama chromatinu (žr. 19 pav. B). Chromatinas dar skirstomas į euchromatinas ir heterochromatinas. Pirmasis yra silpnai nudažytas dažais, nes. yra plonų nekondensuotų chromosomų gijų. Priešingai, heterochromatine yra kondensuotas, taigi ir gerai nudažytas chromosomų siūlas. Nekondensuotose chromatino dalyse yra DNR, kurioje veikia genai (t. y. vyksta RNR sintezė).
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image137.gif" border="0" width="43" height="54 src="> 1 branduolys
A B C
Ryžiai. 19. Chromosomos tarpfazėje.
A – tarpfazėje esančios ląstelės branduolio izoliuota chromosomos grandinė. 1- kondensuotas plotas; 2 - nekondensuotas plotas.
B - tarpfazėje iš ląstelės branduolio išskirtos kelios chromosomų grandinės. 1 - kondensuotas plotas; 2 - nekondensuotas plotas. B - ląstelės branduolys su chromosomų gijomis tarpfazėje. 1 - kondensuotas plotas; 2 - nekondensuotas plotas; 1 ir 2, branduolinis chromatinas.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image150.gif" border="0" width="39" height="12 src=">Chromosomos branduolys
Ryžiai. 20. Dvi paveldimos medžiagos būsenos ląstelėse ląstelių cikle: A - tarpfazėje paveldima medžiaga yra chromosomose, kurios dalinai dekondensuotos ir išsidėsčiusios branduolyje; B – ląstelių dalijimosi metu paveldima medžiaga palieka branduolį, chromosomos išsidėsčiusios citoplazmoje.
Reikia atsiminti, kad jei genas veikia, tada šio regiono DNR yra dekondensuota. Ir atvirkščiai, genų DNR kondensacija rodo genų aktyvumo blokadą. DNR sekcijų kondensacijos ir dekondensacijos reiškinį dažnai galima aptikti, kai ląstelėje reguliuojamas genų aktyvumas (įsijungimas arba išjungimas).
Chromatino (toliau jas vadinsime tarpfazinėmis chromosomomis) ir besidalijančios ląstelės chromosomų (toliau jas vadinsime metafazinėmis chromosomomis) submolekulinė struktūra dar nėra iki galo išaiškinta. Tačiau akivaizdu, kad val skirtingos valstybės ląstelės (tarpfazė ir dalijimasis), paveldimos medžiagos organizacija skiriasi. Tarpfazės (IC) ir metafazės chromosomos (MX) yra pagrįstos nukleosomos. Nukleosoma susideda iš centrinės baltymo dalies, aplink kurią yra apvyniota DNR grandinė. Centrinę dalį sudaro aštuonios histono baltymų molekulės – H2A, H2B, H3, H4 (kiekvieną histoną vaizduoja dvi molekulės). Šiuo atžvilgiu nukleosomos šerdis vadinama tetrameras, oktameras arba šerdis. DNR molekulė spiralės pavidalu apsivynioja aplink šerdį 1,75 karto ir eina į gretimą šerdį, apsivynioja aplink ją ir eina į kitą. Taip sukuriama savotiška figūra, primenanti siūlą (DNR), ant kurio suverti karoliukai (nukleosomos).
Tarp nukleozomų yra DNR, vadinama saitas. Kitas histonas H1 gali prisijungti prie jo. Jei jungiasi prie linkerio vietos, tai DNR susilinksta ir susisuka į spiralę (21 pav. B). Histonas H1 dalyvauja sudėtingame DNR kondensacijos procese, kurio metu karoliukų virtinė susisuka į 30 nm storio spiralę. Ši spiralė vadinama solenoidas. Chromosomų sruogos tarpfazinėse ląstelėse susideda iš karoliukų ir solenoidų. Metafazėse chromosomose solenoidas susisuka į superspiralę, kuri susijungia su tinklelio struktūra (pagaminta iš baltymų), sudarydama kilpas, kurios jau telpa chromosomos pavidalu. Toks pakavimas sukelia beveik 5000 kartų DNR sutankinimą metafazės chromosomoje. 23 paveiksle parodyta nuoseklaus chromatino lankstymo schema. Akivaizdu, kad DNR spiralizacijos procesas IC ir MX yra daug sudėtingesnis, tačiau tai, kas buvo pasakyta, leidžia suprasti labiausiai Bendri principai chromosomų pakavimas.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image156.gif" border="0" width="64" height="19 src="> Н1
Ryžiai. 21. Nukleosomų sandara:
A – nekondensuotoje chromosomoje. Histonas H1 nėra susijęs su linkerio DNR. B – kondensuotoje chromosomoje. Histonas H1 yra susijęs su linkerio DNR.
Reikėtų pažymėti, kad kiekviena chromosoma metafazėje susideda iš dviejų chromatidžių, laikomų kartu centromerai(pirminis susiaurėjimas). Kiekviena iš šių chromatidžių yra pagrįsta dukterinėmis DNR molekulėmis, supakuotomis atskirai. Po sutankinimo proceso šviesos mikroskopu jie tampa aiškiai atskiriami kaip vienos chromosomos chromatidės. Pasibaigus mitozei, jie išsisklaido į dukterines ląsteles. Kadangi vienos chromosomos chromatidės atsiskiria viena nuo kitos, jos jau vadinamos chromosomomis, tai yra, chromosomoje yra arba dvi chromatidės prieš dalijimąsi, arba viena (bet ji jau vadinama chromosoma) po padalijimo.
Kai kurios chromosomos, be pirminio susiaurėjimo, turi ir antrinį susiaurėjimą. Ji taip pat vadinama branduolio organizatorius. Tai plonas chromosomos siūlas, kurio gale dedamas palydovas. Antrinis susiaurėjimas, kaip ir pagrindinė chromosoma, susideda iš DNR, ant kurios yra genai, atsakingi už ribosomų RNR sintezę. Chromosomos gale yra sritis, vadinama telomeras. Atrodo, kad tai „užsandarina“ chromosomą. Jei telomeras netyčia nutrūksta, susidaro „lipnus“ galas, kuris gali jungtis su tuo pačiu kitos chromosomos galu.
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image158.gif" border="0" width="105" height="33 src="> Chromosomų siūlas
Chromosoma
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image161.gif" border="0" width="87" height="12 src="> Karoliukų siūlas
Vadinama visų ląstelės metafazių chromosomų visuma, jų forma ir morfologija kariotipas. Pagal formą išskiriami trys chromosomų tipai – metacentrinės, submetacentrinės ir akrocentrinės (23 pav.). Metacentrinėse chromosomose centromeras
2.8.4. branduolys
Tai tankus, gerai nudažytas kūnas, esantis branduolio viduje. Jame yra DNR, RNR ir baltymų. Branduolio pagrindas yra branduolio organizatoriai – DNR sekcijos, kuriose yra daug rRNR genų kopijų. Ribosomų RNR sintezė vyksta branduolio organizatorių DNR. Prie jų prisijungia baltymai ir susidaro kompleksinis darinys – ribonukleoproteino (RNP) dalelės. Tai yra mažų ir didelių ribosomų subvienetų pirmtakai (arba pusgaminiai). RNP susidarymo procesas daugiausia vyksta periferinėje branduolio dalyje. ri pirmtakai
http://xn--d1aacnkch5m.xn--p1ai/14-bez-rubriki/images/image170.jpg" border="0" width="288" height="99 src="> Palydovas
Ribosomos
Ribosomų pirmtakai
Ryžiai. 24. Ribosomų susidarymas branduolio branduolyje.
Branduolio dydis atspindi jo funkcinio aktyvumo laipsnį, kuris labai skiriasi įvairios ląstelės ir gali skirtis atskiroje ląstelėje. Kuo intensyvesnis ribosomų susidarymo procesas citoplazmoje, tuo aktyviau vyksta specifinių baltymų sintezė ribosomose. Šiuo atžvilgiu pažymėtina steroidiniai hormonai(SG) tikslinėse ląstelėse. SG patenka į branduolį ir suaktyvina rRNR sintezę. Dėl to didėja RNP kiekis ir dėl to citoplazmoje didėja ribosomų skaičius. Dėl to labai padidėja specialių baltymų sintezės lygis, kuris per daugybę biocheminių ir fiziologinės reakcijos suteikti tam tikrą farmakologinis poveikis(pvz., gimdoje auga liaukų epitelis).
Priklausomai nuo ląstelės ciklo fazės išvaizda branduolys ryškiai pasikeičia. Prasidėjus mitozei, branduolys mažėja, o tada visiškai išnyksta. Mitozės pabaigoje, atnaujinus rRNR sintezę, chromosomų regionuose, kuriuose yra rRNR genų, vėl atsiranda miniatiūriniai branduoliai.
2.8.5. branduolinė matrica
Chromosomos trimatėje branduolio erdvėje išsidėsčiusios ne atsitiktinai, o griežtai išdėstytos. Tai palengvina pastolių intrabranduolinė struktūra, vadinama branduoline matrica arba skeletu. Ši struktūra pagrįsta branduolio sluoksniu (žr. 19 pav.). Prie jo pritvirtintas vidinis baltyminis karkasas, užimantis visą branduolio tūrį. Chromosomos tarpfazėje prisitvirtina ir prie sluoksnio, ir prie vidinės baltymo matricos sričių.
Visi išvardyti komponentai yra ne sustingusios standžios konstrukcijos, o judrūs dariniai, kurių architektūra kinta priklausomai nuo funkcinė savybė ląstelės.
Branduolinė matrica atlieka svarbų vaidmenį chromosomų organizavime, DNR replikacijoje ir genų transkripcija. Replikacijos ir transkripcijos fermentai yra pritvirtinti prie branduolio matricos, o DNR grandinė yra „traukiama“ per šį fiksuotą kompleksą.
Paskutinį kartą lamina branduolinės matricos atkreipia dėmesį į ilgaamžiškumo problemą nagrinėjančius mokslininkus. Tyrimai parodė, kad lamina yra sudaryta iš kelių skirtingų baltymų, kuriuos koduoja genai. Šių genų (taigi ir laminato baltymų) struktūros pažeidimas drastiškai sutrumpina eksperimentinių gyvūnų gyvenimo trukmę.
2.9. Citoplazminiai intarpai
Inkliuzai yra nenuolatiniai ląstelės komponentai, kurie yra medžiagų, kurios šiuo metu nedalyvauja ląstelių metabolizme, nuosėdos. Dauguma inkliuzų matomi šviesos mikroskopu granulių pavidalu ir yra hialoplazmoje, organelėse arba vakuolėse. Yra kietų ir skystų intarpų. Priklausomai nuo ląstelės poreikių, jų gali būti dideliais kiekiais arba išnykti.
Bet kokie kieti intarpai ląstelėje, kaip taisyklė, yra maksimaliai dehidratuoti. Šis reiškinys ne tik sumažina inkliuzų tūrį, bet ir neleidžia juose vystytis bakterijoms. Be to, formavimosi procese kietieji inkliuzai tampa chemiškai inertiški, o tai neleidžia jiems dalyvauti cheminėse reakcijose su ląsteliniais junginiais.
Pagal cheminę sudėtį išskiriami angliavandenių, baltymų, lipidų ir mineralų intarpai. Pagal funkciją – trofinė, sekrecinė, specialioji ir kt.
A. Trofiniai (mitybiniai) intarpai
Tai apima baltymų, riebalų ir polisacharidų intarpus.
Baltymų intarpai . Ląstelėje yra junginių, kurių svarbą lemia tai, kad esant poreikiui jie gali tapti daugelio kitų ląstelei gyvybiškai svarbių medžiagų pirmtakais. Šie junginiai apima aminorūgštis. Jie gali būti naudojami ląstelėje kaip energijos šaltiniai angliavandenių, riebalų, hormonų ir kitų metabolitų sintezei. Todėl baltymų inkliuzai iš tikrųjų yra tam tikra ląstelių žaliava amino rūgščių gamybai.
Baltymų inkliuzų likimas visose ląstelėse yra maždaug vienodas. Pirmiausia jie susilieja su lizosoma, kur specialūs fermentai skaido baltymus į aminorūgštis. Pastarieji iš lizosomų patenka į citoplazmą. Kai kurios iš jų sąveikauja su tRNR citoplazmoje ir tokia forma yra pernešamos į ribosomas baltymų sintezei. Kita dalis patenka į specialius biocheminius ciklus, kur iš jų sintetinami riebalai, angliavandeniai, hormonai ir kiti metabolitai. Galiausiai aminorūgštys dalyvauja ląstelės energijos apykaitoje.
Polisacharidų intarpai . Gyvūnų ir grybelių ląstelėms glikogenas yra pagrindinė mitybos dalis. Augalams šis įtraukimas yra krakmolas.
Glikogenas žmogaus organizme daugiausia nusėda kepenų ląstelėse ir naudojamas ne tik pačios ląstelės poreikiams tenkinti, bet ir kaip viso organizmo energijos ištekliai. Pastaruoju atveju glikogenas ląstelėje suskaidomas į gliukozę, kuri iš ląstelės patenka į kraują ir pernešama po visą organizmą.
Glikogenas yra didelė šakota molekulė, sudaryta iš gliukozės likučių. Specialūs tarpląsteliniai procesai, jei reikia, atskiria gliukozės likučius iš glikogeno molekulės ir sintezuoja gliukozę. Pastarasis patenka į kraują ir išleidžiamas ląstelės poreikiams tenkinti. Atrodytų, kad pačią gliukozę būtų lengviau laikyti ląstelėje, jos nepavertus glikogenu, juolab kad gliukozės molekulė yra tirpi ir per plazminę membraną greitai patenka į ląstelę. Tačiau tam trukdo tai, kad gliukozė taip pat greitai, neužsilaikydama, palieka ląstelę. Laikykite ją narve gryna forma beveik neįmanoma. Be to, dideliais kiekiais gliukozės nusėdimas yra pavojingas, nes. tai gali lemti tokio koncentracijos gradiento susidarymą, kad pirmiausia ląstelė išsipučia dėl vandens antplūdžio, o paskui miršta. Todėl speciali fermentų sistema, šiek tiek modifikuojanti gliukozės molekulę, suriša ją su ta pačia molekule. Susidaro milžiniška šakota molekulė, susidedanti iš gliukozės likučių – glikogeno. Ši molekulė jau netirpi, kaip ir gliukozė, ir nepajėgi pakeisti ląstelės osmosinių savybių.
Riebalų intarpai . Šie intarpai hialoplazmoje gali būti lašų pavidalo. Daugelyje augalų yra aliejų, tokių kaip saulėgrąžų, žemės riešutų ir kt. Gausu riebalų intarpų riebalinis audinys asmuo, apsaugantis kūną nuo šilumos nuostolių, energijos saugykla ir kaip amortizatorius mechaninių poveikių metu.
Pažymėtina, kad vidutinio suaugusio žmogaus organizme glikogeno atsargų užtenka vienai normalios veiklos dienai, o riebalų atsargų – mėnesiui. Jei glikogenas, o ne riebalai būtų pagrindinis mūsų kūno energijos rezervas, kūno svoris padidėtų vidutiniškai 25 kg.
Kai kuriais atvejais riebalinių intarpų atsiradimas ląstelėje yra pavojaus signalas bėdų. Taigi, sergant difterija, mikroorganizmo toksinas blokuoja jo panaudojimą riebalų rūgštys ir jie dideliais kiekiais kaupiasi citoplazmoje. Tokiu atveju sutrinka medžiagų apykaita ir ląstelė žūva. Dažniausiai tokie sutrikimai atsiranda širdies raumens ląstelėse. Liga vadinama difteriniu miokarditu.
Visus maistinių medžiagų inkliuzus ląstelė naudoja intensyvios gyvybinės veiklos momentais. Embriogenezėje reikia daug maistinių medžiagų. Todėl kiaušinėlis net oogenezės stadijoje intarpų pavidalu intensyviai kaupia įvairias maistines medžiagas (trynį ir kt.), kurios užtikrina pirmųjų embriono vystymosi stadijų praėjimą.
b. Sekretoriniai intarpai
Įvairios gyvūnų liaukos ląstelėse susidarančios sekrecinės granulės yra įvairios cheminės prigimties ir gali būti atstovaujamos jonais, fermentais, hormonais, glikoproteinais ir kt., pavyzdžiui, virškinimo fermentais, kuriuos sintetina kasos ląstelės. Signalas apie sekrecinių inkliuzų susidarymą ir ištuštinimą kasoje yra maisto suvartojimas. Prieš valgant, intarpai kaupiasi citoplazmoje. Nustačius inkliuzų skaičių kasos ląstelėse, galima apytiksliai atspėti, kieno tai ląstelės – alkano ar gerai maitinančio žmogaus.
V. pigmento intarpai
Suteikti audinių ir organų spalvą. Melanino pigmentas dažnai kaupiamas gyvūnų ląstelėse. Augalai turi karotinoidų ir flavonoidų.
d) ekskreciniai intarpai
Augaluose, kurie neturi specialių šalinimo organų, vakuolėse dažnai kaupiasi galutiniai gyvybinės veiklos produktai kalcio oksalato arba kalcio karbonato druskų pavidalu. Ateityje augalas numeta lapus ar žievę kartu su tokiais susikaupusiais kristaliniais intarpais.
Branduolys yra esminė eukariotinių ląstelių dalis. Tai yra pagrindinis ląstelės reguliavimo komponentas. Ji atsakinga už paveldimos informacijos saugojimą ir perdavimą, viską kontroliuoja medžiagų apykaitos procesai narve . Ne organoidas, o ląstelės komponentas.
Šerdį sudaro:
1) branduolio apvalkalas (branduolinė membrana), per kurio poras vyksta mainai tarp ląstelės branduolio ir citoplazmos.
2) branduolio sultys, arba karioplazma, yra pusiau skysta, silpnai nudažyta plazmos masė, užpildanti visus ląstelės branduolius ir kurioje yra likę branduolio komponentai;
3) chromosomos, kurios matomos nesidalijančiame branduolyje tik padedant specialius metodus mikroskopija. Chromosomų rinkinys ląstelėje vadinamas ariotipas. Chromatinas ant nudažytų ląstelių preparatų yra plonų sruogelių (fibrilių), mažų granulių arba gumulėlių tinklas.
4) vienas ar daugiau sferinių kūnų – nukleolių, kurie yra specializuota ląstelės branduolio dalis ir yra susiję su ribonukleino rūgšties ir baltymų sinteze.
dvi branduolio būsenos:
1. tarpfazinis branduolys – turi branduolius. apvalkalas - kariolema.
2. branduolys ląstelių dalijimosi metu. tik chromatinas yra kitokioje būsenoje.
Branduolys apima dvi zonas:
1. vidinės-fibrilinės-baltymų molekulės ir pre-RNR
2. išorinis – granuliuotas – sudaro ribosomų subvienetus.
Branduolinis apvalkalas susideda iš dviejų membranų, atskirtų perinuklearine erdve. Abi jos yra persmelktos daugybe porų, kurių dėka galimas medžiagų mainai tarp branduolio ir citoplazmos.
Pagrindiniai branduolio komponentai yra chromosomos, susidarančios iš DNR molekulės ir įvairių baltymų. Šviesos mikroskopu jie aiškiai išsiskiria tik ląstelių dalijimosi (mitozės, mejozės) laikotarpiu. Nesidalančioje ląstelėje chromosomos atrodo kaip ilgi ploni siūlai, pasiskirstę visame branduolio tūryje.
Pagrindinės funkcijos ląstelės branduolys Sekantis:
- duomenų saugykla;
- informacijos perkėlimas į citoplazmą naudojant transkripciją, t.y., informaciją nešančios i-RNR sintezę;
- informacijos perdavimas dukterinėms ląstelėms replikacijos metu – ląstelių ir branduolių dalijimasis.
- reguliuoja biocheminius, fiziologinius ir morfologinius procesus ląstelėje.
vyksta branduolyje replikacija- DNR molekulių dubliavimas, taip pat transkripcija- RNR molekulių sintezė DNR šablone. Branduolyje susintetintos RNR molekulės patiria tam tikrų modifikacijų (pvz., per sujungimas nereikšmingi, beprasmiai regionai neįtraukiami į pasiuntinio RNR molekules), po to jie patenka į citoplazmą . Ribosomų surinkimas atsiranda ir branduolyje Specialusis ugdymas vadinami branduoliais. Branduolio skyrius - karioteka - susidaro išplečiant ir sujungiant vienas su kitu endoplazminio tinklo rezervuarus taip, kad branduolys turėtų dvigubas sieneles dėl jį supančių siaurų branduolio membranos skyrių. Branduolinio apvalkalo ertmė vadinama liumenas arba perinuklearinė erdvė. Branduolinio apvalkalo vidinį paviršių dengia branduolys lamina- standi baltymo struktūra, sudaryta iš laminų baltymų, prie kurių yra prijungtos chromosomų DNR grandinės. Kai kuriose vietose vidinė ir išorinė branduolio apvalkalo membranos susilieja ir suformuoja vadinamąsias branduolio poras, per kurias vyksta medžiagų mainai tarp branduolio ir citoplazmos.
12. Dviejų membranų organelės (mitochondrijos, plastidės). Jų struktūra ir funkcijos.
Mitochondrijos - tai apvalios arba strypo formos struktūros, dažnai šakojančios, 0,5 µm storio ir dažniausiai iki 5–10 µm ilgio.
Mitochondrijų apvalkalą sudaro dvi membranos, kurios skiriasi chemine sudėtimi, fermentų rinkiniu ir funkcijomis. Vidinė membrana formuoja lapų (cristae) arba vamzdelio (vamzdelių) formos invaginacijas. Vidinės membranos apribota erdvė yra matrica
organelės. Jame su elektroninis mikroskopas randami 20-40 nm skersmens grūdeliai. Jie kaupia kalcio ir magnio jonus, taip pat polisacharidus, tokius kaip glikogenas.
Matricoje yra savo organelių baltymų biosintezės aparatas. Ją vaizduoja 2–6 žiedinės ir be histonų (kaip ir prokariotuose) DNR molekulės kopijos, ribosomos, transportinės RNR (tRNR) rinkinys, DNR replikacijos, transkripcijos ir paveldimos informacijos vertimo fermentai. Pagrindinė funkcija
mitochondrijos susideda iš fermentinio energijos išgavimo iš tam tikrų cheminių medžiagų (jų oksidacijos būdu) ir energijos kaupimo biologiškai tinkama forma (sintezuojant adenozino trifosfato -ATP molekules). Apskritai šis procesas vadinamas oksidacinis fosforilinimas.
Tarp šalutinių funkcijų mitochondrijas galima vadinti dalyvavimu steroidinių hormonų ir kai kurių aminorūgščių (glutamino) sintezėje.
plastidai - tai pusiau autonominės (gali egzistuoti gana autonomiškai nuo ląstelės branduolinės DNR) dviejų membranų organelės, būdingos fotosintetiniams eukariotų organizmams. Yra trys pagrindiniai plastidų tipai: chloroplastai, chromoplastai ir leukoplastai.Plastidų visuma ląstelėje vadinamaplastidoma . Kiekvienas iš šių tipų tam tikromis sąlygomis gali pereiti vienas į kitą. Kaip ir mitochondrijose, plastiduose yra savo DNR molekulių. Todėl jie taip pat gali daugintis nepriklausomai nuo ląstelių dalijimosi. Plastidai yra unikalūs augalų ląstelės.
Chloroplastai. Chloroplastų ilgis svyruoja nuo 5 iki 10 mikronų, skersmuo – nuo 2 iki 4 mikronų. Chloroplastus riboja dvi membranos. Išorinė membrana lygi, vidinė sudėtingos sulankstytos struktūros. Mažiausia raukšlė vadinama t ilakoidas. Tylakoidų grupė, sukrauta kaip monetų krūva, vadinama g žaizda. Granulės yra sujungtos viena su kita plokščiais kanalais - lamelės. Tilakoidinėse membranose yra fotosintezės pigmentų ir fermentų, kurie užtikrina ATP sintezę. Pagrindinis fotosintezės pigmentas yra chlorofilas, kuris yra atsakingas už žalia spalva chloroplastai.
Vidinė chloroplastų erdvė yra užpildyta stroma. Stromoje yra apskrita plika DNR, ribosomos, Kalvino ciklo fermentai ir krakmolo grūdeliai. Kiekvieno tilakoido viduje yra protonų rezervuaras, yra H + kaupimasis. Chloroplastai, kaip ir mitochondrijos, dalijasi į dvi dalis, geba autonomiškai daugintis. Žemesniųjų augalų chloroplastai vadinami chromatoforai.
Leukoplastai. Išorinė membrana yra lygi, vidinė sudaro mažus tilakoidus. Stromoje yra žiedinė „nuoga“ DNR, ribosomos, fermentai rezervinių maistinių medžiagų sintezei ir hidrolizei. Pigmentų nėra. Ypač daug leukoplastų turi požeminių augalo organų (šaknų, gumbų, šakniastiebių ir kt.) ląstelių. .). Amiloplastai- sintetina ir sandėliuoja krakmolą , elaioplast- alyvos , proteinoplastai- baltymai. Tame pačiame leukoplaste gali kauptis skirtingos medžiagos.
Chromoplastai. Išorinė membrana lygi, vidinė arba taip pat lygi, arba sudaro pavienius tilakoidus. Stromoje yra žiedinė DNR ir pigmentai. - karotinoidai, suteikiant chromoplastams geltoną, raudoną arba oranžinę spalvą. Pigmentų kaupimosi forma yra skirtinga: kristalų pavidalo, ištirpusių lipidų lašeliuose ir tt Chromoplastai laikomi paskutiniu plastidų vystymosi etapu.
Plastidai gali tarpusavyje transformuotis vienas į kitą: leukoplastai – chloroplastai – chromoplastai.
Vieno membranos organelės (ER, Golgi aparatas, lizosomos). Jų struktūra ir funkcijos.
vamzdinis Ir vakuolinė sistema susidaro susisiekiančiomis arba atskiromis vamzdinėmis arba plokščiomis (cisterninėmis) ertmėmis, apribotomis membranomis ir išplitusiomis po visą ląstelės citoplazmą. Šioje sistemoje yra grubus Ir lygus citoplazminis tinklas. Grubus tinklo struktūros bruožas yra polisomų pritvirtinimas prie jo membranų. Dėl šios priežasties jis atlieka tam tikros kategorijos baltymų, kurie daugiausia pašalinami iš ląstelės, pavyzdžiui, išskiriami liaukos ląstelės, sintezės funkciją. Grubus tinklo srityje susidaro baltymų ir lipidų susidarymas. citoplazminės membranos ir jų surinkimas. Tankiai supakuotos į sluoksniuotą struktūrą grubaus tinklo cisternos yra aktyviausios baltymų sintezės vietos ir vadinamos ergastoplazma.
Lygiojo citoplazminio tinklo membranose nėra polisomų. Funkciniu požiūriu šis tinklas yra susijęs su angliavandenių, riebalų ir kitų nebaltyminių medžiagų, tokių kaip steroidiniai hormonai, metabolizmu (lytinėse liaukose, antinksčių žievėje). Per kanalėlius ir cisternas medžiagos, ypač liaukos ląstelės išskiriama medžiaga, iš sintezės vietos patenka į pakavimo vietą į granules. Kepenų ląstelių srityse, kuriose gausu lygių tinklo struktūrų, kenksmingos medžiagos sunaikinamos ir neutralizuojamos. toksiškos medžiagos kai kurie vaistai (barbitūratai). Lygiojo dryžuotų raumenų tinklo pūslelėse ir kanalėliuose kaupiasi (nusėda) kalcio jonai, kurie atlieka svarbų vaidmenį susitraukimo procese.
Golgi kompleksas-yra plokščių membraninių maišelių šūsnis, vadinamas cisternos. Cisternos yra visiškai izoliuotos viena nuo kitos ir nėra tarpusavyje sujungtos. Iš cisternų išilgai kraštų atsišakoja daugybė kanalėlių ir pūslelių. Iš EPS karts nuo karto suvarstomos vakuolės (pūslelės) su susintetintomis medžiagomis, kurios persikelia į Golgi kompleksą ir su juo susijungia. EPS sintezuojamos medžiagos tampa sudėtingesnės ir kaupiasi Golgi komplekse. Golgi komplekso funkcijos :1- „Golgi“ komplekso rezervuaruose vyksta tolesnė cheminių medžiagų, patekusių į jį iš EPS, transformacija ir komplikacijos. Pavyzdžiui, susidaro medžiagos, reikalingos ląstelių membranai atnaujinti (glikoproteinai, glikolipidai), polisacharidai.
2- Golgi komplekse kaupiasi medžiagos ir laikinas jų „saugojimas“
3- Susidariusios medžiagos „supakuojamos“ į pūsleles (vakuoles) ir tokia forma juda per ląstelę.
4- Golgi komplekse susidaro lizosomos (sferinės organelės su ardančiais fermentais).
Lizosomos- mažos sferinės organelės, kurių sieneles sudaro viena membrana; sudėtyje yra litikų(skaldymo) fermentai. Iš pradžių lizosomose, surištose iš Golgi komplekso, yra neaktyvių fermentų. Tam tikromis sąlygomis jų fermentai aktyvuojami. Lizosomai susiliejus su fagocitine ar pinocitine vakuole, susidaro virškinimo vakuolė, kurioje tarpląsteliniu būdu virškinamos įvairios medžiagos.
Lizosomų funkcijos :1- Atlikite fagocitozės ir pinocitozės metu absorbuotų medžiagų skaidymą. Biopolimerai suskaidomi į monomerus, kurie patenka į ląstelę ir yra naudojami jos poreikiams. Pavyzdžiui, jie gali būti naudojami naujų organinių medžiagų sintezei arba gali būti toliau skaidomi energijai gauti.
2- Sunaikinkite senus, pažeistus, perteklinius organelius. Organelių skilimas gali įvykti ir ląstelės badavimo metu.
Vakuolės- sferinės vienos membranos organelės, kurios yra vandens ir jame ištirpusių medžiagų rezervuarai. Vakuolės apima: fagocitinės ir pinocitinės vakuolės, virškinimo vakuolės, pūslelės, surištos iš EPS ir Golgi komplekso. Gyvūnų ląstelių vakuolės yra mažos, daug, tačiau jų tūris neviršija 5% viso ląstelių tūrio. Jų pagrindinė funkcija - medžiagų transportavimas per ląstelę, santykio tarp organelių įgyvendinimas.
Augalų ląstelėje vakuolės sudaro iki 90% tūrio. Subrendusio augalo ląstelėje yra tik viena vakuolė, ji užima centrinę vietą. Augalų ląstelės vakuolinė membrana yra tonoplastas, jos turinys – ląstelių sultys. Vakuolių funkcijos augalo ląstelėje: priežiūra ląstelių sienelės esant įtampai, įvairių medžiagų, įskaitant ląstelės atliekas, kaupimasis. Vakuolės tiekia vandenį fotosintezei. Gali apimti:
Atsarginės medžiagos, kurias gali panaudoti pati ląstelė ( organinės rūgštys, amino rūgštys, cukrus, baltymai). - medžiagos, kurios pasišalina iš ląstelės metabolizmo ir kaupiasi vakuolėje (fenoliai, taninai, alkaloidai ir kt.) - fitohormonai, fitoncidai,
Pigmentai (dažančiosios medžiagos), suteikiantys ląstelei purpurinę, raudoną, mėlyną, violetinę, o kartais geltoną ar kreminę sultis. Būtent ląstelių sulčių pigmentai dažo gėlių žiedlapius, vaisius, šakniavaisius.
14. Nemembraninės organelės (mikrotubulai, ląstelės centras, ribosomos). Jų struktūra ir funkcijos.Ribosoma - nemembraninė ląstelės organelė, kuri atlieka baltymų sintezę. Susideda iš dviejų subvienetų – mažo ir didelio. Ribosoma susideda iš 3-4 rRNR molekulių, kurios sudaro jos karkasą, ir keliasdešimties įvairių baltymų molekulių. Ribosomos sintetinamos branduolyje. Ląstelėje ribosomos gali būti granuliuoto ER paviršiuje arba ląstelės hialoplazmoje polisomų pavidalu. polisomas - tai i-RNR ir kelių ribosomų kompleksas, nuskaitantis iš jo informaciją. Funkcija ribosomos- baltymų biosintezė. Jei ribosomos yra ant ER, tai jų susintetinti baltymai naudojami viso organizmo poreikiams, hialoplazminės ribosomos sintetina baltymus pačios ląstelės poreikiams. Ribosomos prokariotinės ląstelės mažesnės nei eukariotinės ribosomos. Tos pačios mažos ribosomos randamos mitochondrijose ir plastiduose.
mikrovamzdeliai - tuščiavidurės cilindrinės ląstelės struktūros, susidedančios iš neredukuojamo baltymo tubulino. Mikrovamzdeliai negali susitraukti. Mikrotubulo sieneles sudaro 13 baltymo tubulino gijų. Mikrotubulai yra ląstelių hialoplazmos storyje.
Blakstienos ir žvyneliai - judėjimo organelės. Pagrindinė funkcija - ląstelių judėjimas arba juos supančio skysčio ar dalelių judėjimas išilgai ląstelių. Daugialąsčiame organizme blakstienėlės būdingos epiteliui. kvėpavimo takų, kiaušintakiai, o žvyneliai – spermatozoidams. Blakstienos ir žvyneliai skiriasi tik dydžiu – žvyneliai yra ilgesni. Jų pagrindas yra mikrovamzdeliai, išdėstyti 9(2) + 2 sistema.Tai reiškia, kad 9 dvigubi mikrovamzdeliai (dubletai) sudaro cilindro sienelę, kurios centre yra 2 pavieniai mikrovamzdeliai. Blakstienas ir žvynelius palaiko baziniai kūnai. Bazinis korpusas yra cilindro formos, sudarytas iš 9 mikrovamzdelių tripletų (tripletų), pagrindinio kūno centre mikrovamzdelių nėra.
kl e tikslus centras - mitozinis centras, nuolatinė struktūra beveik visose gyvūnų ir kai kurių augalų ląstelėse, lemia besidalijančios ląstelės polius (žr. Mitozė) . Ląstelės centras dažniausiai susideda iš dviejų centriolių – tankių 0,2-0,8 dydžio granulių mikronas, esantys stačiu kampu vienas kito atžvilgiu. Formuojantis mitoziniam aparatui centrioliai nukrypsta link ląstelės polių, nulemdami ląstelių dalijimosi veleno orientaciją. Todėl teisingiau K. c. skambinti mitozinis centras, tai atspindi jo funkcinę reikšmę, juolab kad tik kai kuriose ląstelėse K. c. esantis jos centre. Organizmo vystymosi eigoje jie kinta, nes padėtis K. c. ląstelėse, taip pat ir jo forma. Kai ląstelė dalijasi, kiekviena iš dukterinių ląstelių gauna centriolių porą. Jų dubliavimosi procesas dažniau vyksta ankstesnio ląstelių dalijimosi pabaigoje. Serialo atsiradimas patologinės formos ląstelės dalijimasis yra susijęs su nenormaliu dalijimusi. c.