ישנן שלוש השערות למקור תאים איקריוטיים:
- השערה סימביוטית, או סימביוגנזה,
- פולשני,
- כימרי.
עד כה, בעולם המדעי, מוכרת ההשערה העיקרית של מקור האיקריוטים סימביוגנזה.
על פי סימביוגנזה, אברוני תאים אוקריוטיים כגון כלורופלסטים ודגלים התרחשו על ידי החדרת פרוקריוט אחד לתא פרוקריוטי אחר, גדול יותר, שמילא את תפקיד התא המארח.
בהשערה הסימביוטית יש קשיים בהסבר מקור הגרעין של תאים אוקריוטיים ובשאלה איזה פרוקריוט היה המאכסן בכל זאת. נתוני הניתוח המולקולרי של הגנום והחלבונים האיקריוטיים מראים שמצד אחד זה היה אורגניזם קרוב לארכיאה (בעבר הם היו שייכים לחיידקים, ואז הם הופרדו לענף נפרד). מצד שני, באוקריוטים יש חלבונים (וגנים האחראים לסינתזה שלהם) האופייניים לקבוצות אחרות של פרוקריוטות מושלמות.
על פי השערת האינוואגינציה של מקורם של תאים אוקריוטיים, האברונים שלהם נוצרו על ידי פלישה של ציטו קרום פלזמהעם ההפרדה שלאחר מכן של מבנים אלה. נוצרו משהו כמו כדורים, מוקפים בממברנה ומכילים בתוך הציטופלזמה תרכובות ומבנים שנלכדו כאן. בהתאם למה שנכנס פנימה, נוצרו אברונים שונים.
לפרוקריוטים אין אברונים אמיתיים, תפקידיהם מבוצעים על ידי פלישות ממברנות. לכן, קל לדמיין שרוך כזה. כמו כן, לטובת השערת האינוואגינציה עומד הדמיון בין הממברנה הציטופלזמית והממברנות הכפולות של האברונים.
מנקודת מבט של פלישה, מקורו של הגרעין מוסבר בקלות, אך לא ניתן להסביר מדוע הגנום והריבוזומים של הקומפלקס הגרעיני-ציטופלסמי שונים מאלה שבכלורופלסטים ובמיטוכונדריה (נזכיר שהם מכילים גם DNA וריבוזומים). יתרה מכך, באברונים אלו, מערכת הביוסינתזה של חלבונים (DNA, RNA, ריבוזומים) דומה לזו של פרוקריוטים.
ההבדל הזה מוסבר היטב רק מנקודת המבט של ההשערה הראשונה, הסימביוטית. לדבריה, כך או אחרת, פרוקריוט אירובי נכנס לפרוקריוט אנאירובי. הוא לא התעכל, אלא התחיל, להיפך, לאכול על חשבון התא המארח. בתורו, הוא השתמש בחמצן כדי להשיג אנרגיה, ושיטה זו של חמצון היא הרבה יותר יעילה, הוא נתן את האנרגיה העודפת למארח הפרוקריוטי, שבמקרה זה גם הועיל. נוצרה סימביוזה. לאחר מכן, הפרוקריוט שהוכנס הפך לפשוט יותר, חלק מהגנום שלו היגר לתוך התא המארח, הוא לא יכול היה להתקיים יותר באופן עצמאי.
באופן דומה, סימביוגנזה מסבירה את מקור הכלורופלסטים. רק פרוקריוטות המסוגלות לפוטוסינתזה (בדומה לאצות כחולות ירוקות) כבר הוצגו.
האיקריוטים הראשונים ללא סימביונים פוטוסינתטיים הולידו את החיות שבהן הופיעו - צמחים.
נכון להיום, ישנם פרוטוזואים (אורגניזמים אוקריוטיים חד-תאיים) שאין להם מיטוכונדריה או כלורופלסטים. אבל במקום אותם, פרוקריוטים סימביונים מתיישבים בציטופלזמה, ומבצעים את הפונקציות המתאימות. עובדה זו, כמו גם הדמיון של מערכת הביוסינתזה של חלבונים של מיטוכונדריה ופלסטידים עם פרוקריוטים, נחשבים כעדות לסימביוגנזה. ההוכחה היא גם שהמיטוכונדריה והכלורופלסטים מתרבים בעצמם, הם אף פעם לא נבנים על ידי התא מאפס.
לטובת ההשערה השלישית, הכימרית, של מקורם של תאים אוקריוטיים, אומר מידה גדולההגנום שלהם, העולה על החיידק פי אלפי או יותר, כמו גם מגוון החלבונים המסונתזים שנמצאים ב קבוצות שונותפרוקריוטים. ברור שבמהלך האבולוציה של האוקריוטים, הגנום שלהם הסתבך, הוא הוכפל, הופיעו בו גנים מווסתים רבים. אך עדיין, הגידול הראשוני בגודל הגנום יכול היה להתרחש עקב שילוב הגנום של מספר פרוקריוטים.
אולי בימי קדם, פרוקריוט מסוים רכש את יכולת הפגוציטוזיס ובאכילה כך ספג, בין השאר, פרוקריוטים אחרים, שלא תמיד התעכלו. הגנום שלהם הכיל גנים שימושיים למארח, והוא כלל אותם בגנום שלו. אולי חלק מהפרוקריוטים שהיו בפנים הפכו לאברונים, מה שמשלב את ההשערה הכימרית עם סימביוגנזה.
מכיוון שלכל ההשערות לעיל יש חזק ו צדדים חלשים, וגם במידה רבה לא שוללים את עמדותיו של זה, אז לדעתנו, שילוב של גורמים רבים המתוארים על ידי השערות שונות יכול לשחק תפקיד במקורם של המבנים של תאים אוקריוטיים.
כמו כן, יש לציין כי על פי ההשערה הסימביוטית, ניתן להתייחס למקורם של אברוני ממברנה אחרים כגון ואקוולים, קומפלקס גולגי וכו' כפישוט נוסף, למשל, של המיטוכונדריה.
כמו כן, אנו מציינים כי השילוב של חלבונים מפרוקריוטים שונים באאוקריוטים עשוי כלל לא להצביע על כימריזם של הראשונים. ייתכן שתפקודים אנזימטיים מסוימים יכולים להתבצע רק על ידי חלבונים בעלי מבנה מסוים. והאבולוציה של האיקריוטים הגיעה לזה שוב ושוב, ללא קשר לאבולוציה של קבוצות פרוקריוטות שאינן אבותיהם. נניח שהייתה התכנסות ברמה המולקולרית.
הופעת האוקריוטים היא אירוע מרכזי. זה שינה את מבנה הביוספרה ופתח הזדמנויות חדשות ביסודו לאבולוציה מתקדמת. התא האוקריוטי הוא תוצאה של אבולוציה ארוכה של עולם הפרוקריוטים, עולם שבו חיידקים מגוונים הסתגלו זה לזה וחיפשו דרכים לשתף פעולה ביעילות.
מתאר ציר זמן (חזרה)

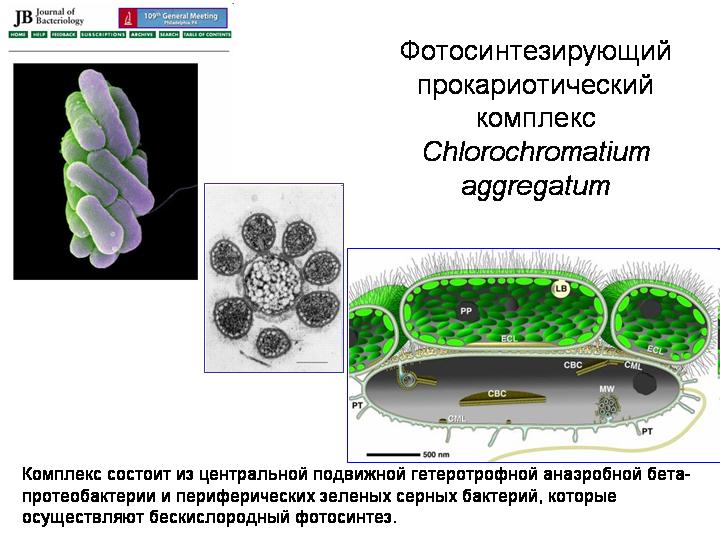
קומפלקס פרוקריוטי פוטוסינתטי Chlorochromatium aggregatum.
אוקריוטים התעוררו כתוצאה מסימביוזה של מספר סוגים של פרוקריוטים. פרוקריוטים באופן כללי נוטים למדי לסימביוזה (ראה פרק 3 בהולדת המורכבות). הנה מערכת סימביוטית מעניינת המכונה Chlorochromatium aggregatum. חי באגמים עמוקים, שבהם יש תנאים אנוקסיים בעומק. המרכיב המרכזי הוא בטא-פרוטאובקטריה הטרוטרופית ניידת. סביבו, ערימות הן בין 10 ל-60 חיידקי גופרית ירוקה פוטוסינתטיים. כל הרכיבים מחוברים על ידי יציאות קרום חיצוניחיידק מרכזי. משמעות הקהילה היא שהבטא-פרוטאובקטריה הניידת גוררת את החברה כולה למקומות נוחים לחייהם של חיידקי הגופרית החביבים, וחיידקי הגופרית עוסקים בפוטוסינתזה ומספקים מזון לעצמם ולבטא-פרוטאובקטריה. אולי כמה אסוציאציות מיקרוביאליות עתיקות מהסוג הזה היו אבותיהם הקדמונים של האוקריוטים.
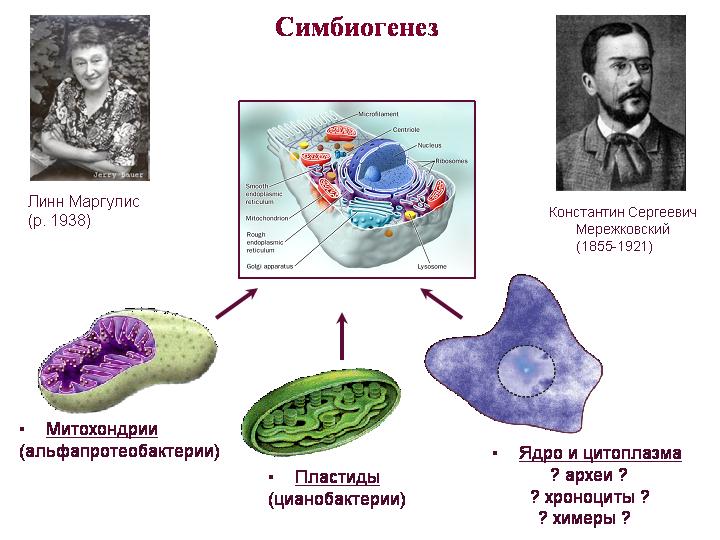
תורת הסימביוגנזה. מרז'קובסקי, מרגוליס. מיטוכונדריה הם צאצאים של אלפא-פרוטאובקטריה, פלסטידים הם צאצאים של ציאנובקטריה. קשה יותר להבין מי היה האב הקדמון של כל השאר, כלומר הציטופלזמה והגרעין. הגרעין והציטופלזמה של האוקריוטים משלבים תכונות של ארכאים וחיידקים, ויש להם גם תכונות ייחודיות רבות.
לגבי המיטוכונדריה. אולי זו הייתה רכישת המיטוכונדריה (ולא הגרעין) שהייתה רגע המפתח בהתפתחות האיקריוטים. רוב הגנים המיטוכונדריים הקדומים הועברו לגרעין, שם הם הגיעו לשליטת מערכות ויסות גרעיניות. גנים גרעיניים אלה ממקור מיטוכונדריאלי מקודדים לא רק לחלבונים מיטוכונדריאליים, אלא גם לחלבונים רבים הפועלים בציטופלזמה. זה מצביע על כך שהסימביונטה המיטוכונדריאלית מילאה תפקיד חשוב יותר ביצירת התא האוקריוטי מהצפוי.
קיום משותף של שני גנומים שונים בתא אחד הצריך פיתוח של מערכת יעילה לוויסות שלהם. וכדי לנהל ביעילות את עבודתו של גנום גדול, יש צורך לבודד את הגנום מהציטופלזמה, בה מתרחש חילוף חומרים ואלפי תגובה כימית. המעטפת הגרעינית רק מפרידה בין הגנום לתהליכים הכימיים הסוערים של הציטופלזמה. רכישת סימביונים (מיטוכונדריה) עשויה להפוך לגירוי חשוב לפיתוח מערכות הוויסות של הגרעין והגנים.
כך גם לגבי רבייה מינית. אתה יכול לחיות ללא רבייה מינית כל עוד הגנום שלך קטן מספיק. אורגניזמים בעלי גנום גדול, אך נטולי רבייה מינית, נידונים להכחדה מהירה, למעט חריגים נדירים.
![]()
Alphaproteobacteria - קבוצה זו כללה את אבות המיטוכונדריה.
Rhodospirillum הוא מיקרואורגניזם מדהים שיכול לחיות עקב פוטוסינתזה, כולל בתנאים אנאירוביים, וכהטרוטרוף אירובי, ואפילו ככימואוטוטרוף אירובי. הוא יכול, למשל, לגדול עקב חמצון פחמן חד חמצני CO ללא שימוש במקורות אנרגיה אחרים. בנוסף לכל זה, הוא גם יודע לתקן חנקן אטמוספרי. כלומר, זה בפנים התואר הגבוה ביותראורגניזם אוניברסלי.
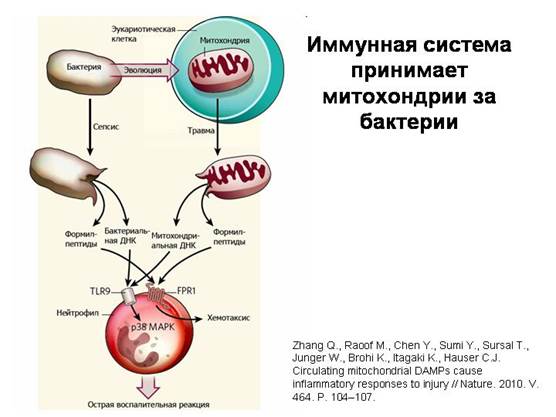
מערכת החיסון טועה במיטוכונדריה כחיידקים. כאשר מיטוכונדריה פגומות חודרות לזרם הדם במהלך פציעה, הן משחררות מולקולות אופייניות שנמצאות רק בחיידקים ובמיטוכונדריה (DNA מעגלי מסוג חיידקים וחלבונים הנושאים חומצת אמינו שונה מיוחדת פורמילמתיונין באחד מקצוותיהם). זה נובע מהעובדה שמנגנון סינתזת החלבון במיטוכונדריה נשאר זהה לזה של חיידקים. תאים מערכת החיסון- נויטרופילים - מגיבים לחומרים המיטוכונדריאליים הללו באותו אופן כמו לחיידקים, ובעזרת אותם קולטנים. זהו האישור הברור ביותר לאופי החיידקי של המיטוכונדריה.
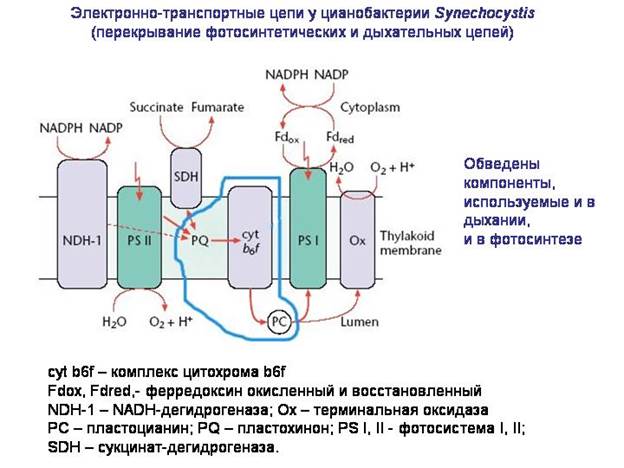
תפקיד עיקרימיטוכונדריה - נשימה חמצן. ככל הנראה, הגירוי לקשר של האב הקדמון האנאירובי של הגרעין והציטופלזמה עם ה"פרוטומיטוכונדריה" היה הצורך להגן מפני פעולה רעילהחַמצָן.
מהיכן קיבלו חיידקים, כולל אלפאפרוטובקטריה, את המערכות המולקולריות הדרושות נשימה חמצן? נראה שהם התבססו על מערכות מולקולריות של פוטוסינתזה. שרשרת העברת האלקטרונים, שנוצרה בחיידקים כחלק מהמנגנון הפוטוסינתטי, הותאמה לנשימת חמצן. בחלק מהחיידקים, חלקים משרשראות הובלה של האלקטרונים עדיין משמשים בו זמנית הן בפוטוסינתזה והן בנשימה. ככל הנראה, אבות המיטוכונדריה היו הטרוטרופיים אלפא-פרוטאובקטריות אירוביות, אשר, בתורן, צאצאי אלפא-פרוטאובקטריות פוטוסינתטיות, כגון rhodospirillum.
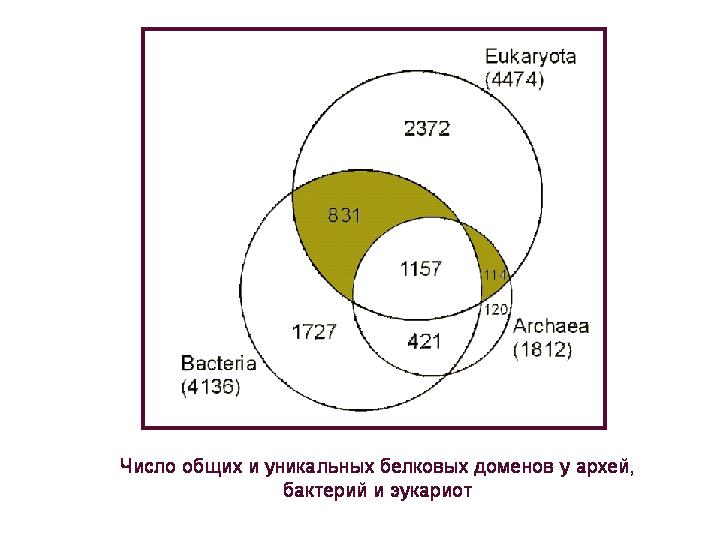
מספר תחומי החלבון הנפוצים והייחודיים בארכיאה, חיידקים ואיקריוטים. תחום חלבון הוא חלק ממולקולת חלבון שיש לה תפקיד ספציפי ומבנה אופייני, כלומר רצף של חומצות אמינו. כל חלבון, ככלל, מכיל אחד או יותר מהיחידות או התחומים המבניים והתפקודיים הללו.
ניתן לחלק 4.5 אלף תחומי חלבון שיש לאאוקריוטים ל-4 קבוצות: 1) זמינים רק באוקריוטים, 2) משותפים לכל שלוש ממלכות העל, 3) משותף לאאוקריוטים וחיידקים, אך נעדר בארכאה; 4) נפוץ לאאוקריוטים ולארכיאה, אך נעדר בחיידקים. נשקול שניים קבוצות אחרונות(הם מודגשים בצבע באיור), שכן לגבי חלבונים אלה ניתן לדבר בוודאות מסוימת על מקורם: חיידקי או ארכאלי, בהתאמה.
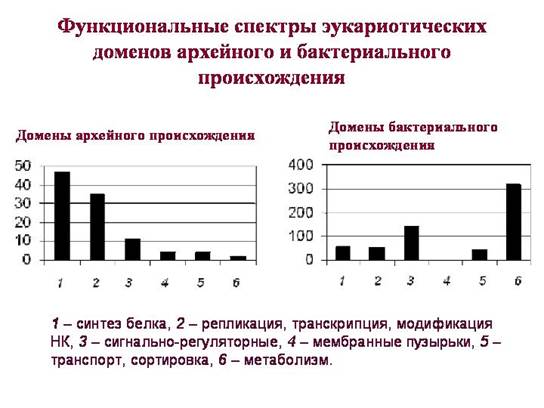
רגע מפתחבכך שלתחומים האוקריוטיים שהועברו כביכול מחיידקים ומארכאים יש תפקידים שונים באופן משמעותי. תחומים שעברו בירושה מארכיאה (הספקטרום התפקודי שלהם מוצג בגרף השמאלי) ממלאים תפקיד מפתח בחייו של תא איקריוטי. ביניהם שולטים תחומים הקשורים לאחסון, רבייה, ארגון וקריאה של מידע גנטי. רוב התחומים ה"ארכאיים" שייכים לאותן קבוצות פונקציונליות שבהן חילופי גנים אופקיים בפרוקריוטים מתרחשים בתדירות נמוכה יותר. ככל הנראה, אוקריוטים קיבלו את התסביך הזה בירושה ישירה (אנכית) מארכיאה.
בין התחומים שמקורם בחיידק, ישנם גם חלבונים הקשורים לתהליכי מידע, אך הם מעטים. רובם עובדים רק במיטוכונדריה או פלסטידים. הריבוזומים האיקריוטיים של הציטופלזמה הם ממקור ארכאי, הריבוזומים של המיטוכונדריה והפלסטידים הם ממקור חיידקי.
בין תחומי החיידקים של אוקריוטים, חלקם של חלבונים מווסתים אותות גבוה בהרבה. מחיידקים, איקריוטים ירשו חלבונים רבים האחראים למנגנוני תגובת התא לגורמים סביבה חיצונית. וגם - חלבונים רבים הקשורים לחילוף חומרים (לפרטים נוספים, ראה פרק 3, "הולדת המורכבות").
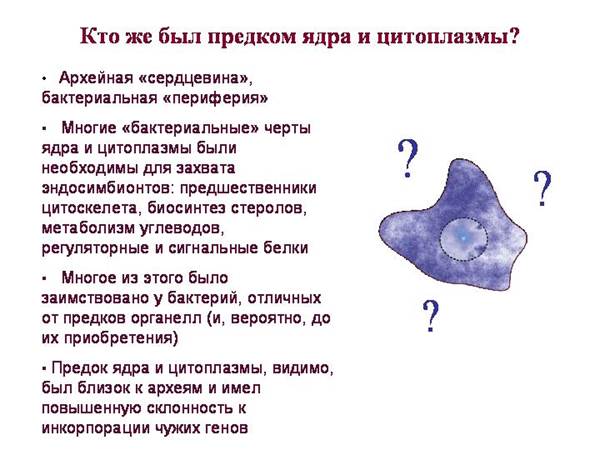
לאוקריוטים יש:
"הליבה" הארכאית (מנגנונים לעבודה עם מידע גנטי וסינתזת חלבונים)
"פריפריה" חיידקית (מטבוליזם ומערכות ויסות אותות)
התרחיש הפשוט ביותר: ARCHEIA בלעה BACTERIA (אבותיהם של המיטוכונדריה והפלסטידים) ואת כל סימנים חיידקייםנרכש מהם.
· תרחיש זה פשוט מדי מכיוון שלאוקריוטים יש חלבונים חיידקיים רבים שלא ניתן היה להשאיל מאבות קדמונים מיטוכונדריה או פלסטידית.
לאוקריוטים יש תחומים "בקטריאליים" רבים שאינם אופייניים לא לציאנובקטריה (אבות של פלסטידים) או לאלפאפרוטובקטריה (אבות של מיטוכונדריה). הם התקבלו מחיידקים אחרים.
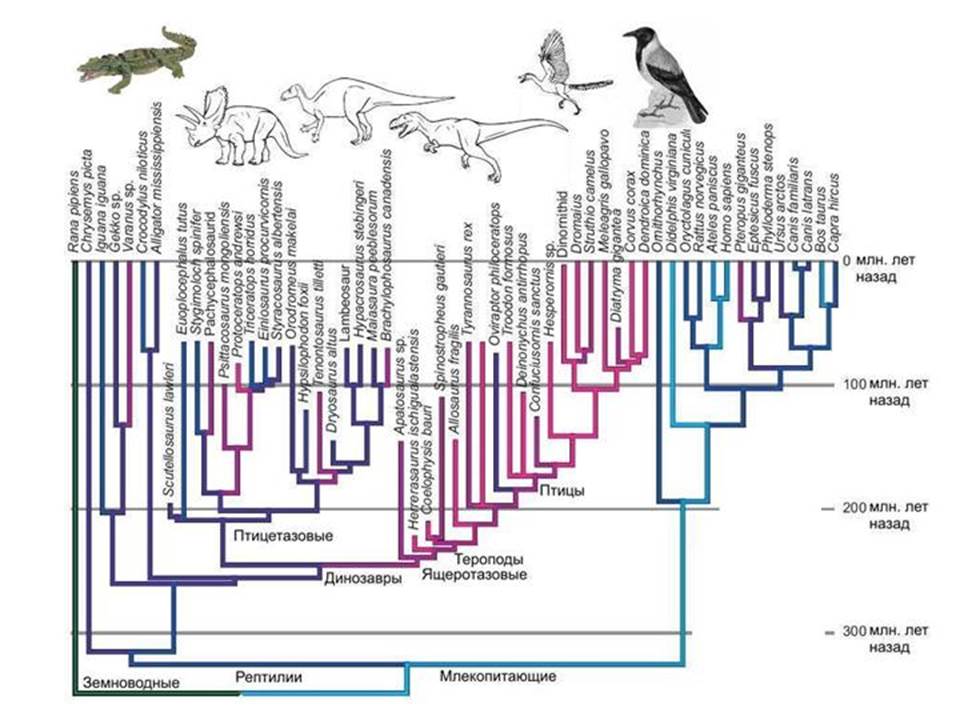
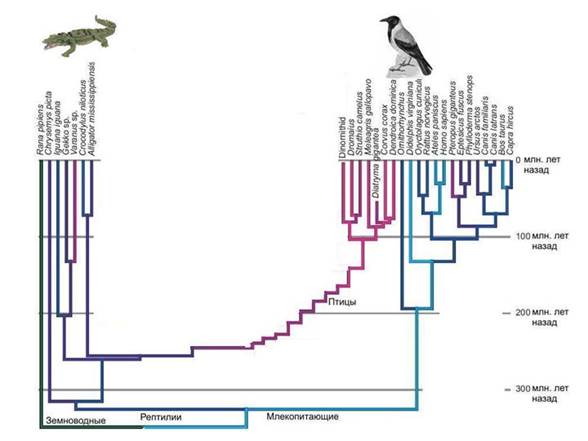
ציפורים ודינוזאורים. שחזור פרוטו-אאוקריוטים הוא קשה. ברור שלקבוצת הפרוקריוטים הקדומים שהולידו את הגרעין והציטופלזמה היו מספר תכונות ייחודיות שאין לפרוקריוטים ששרדו עד היום. וכאשר אנו מנסים לשחזר את הופעתו של האב הקדמון הזה, אנו עומדים בפני העובדה שהיקף ההשערות מתברר כגדול מדי.
אֲנָלוֹגִיָה. ידוע שציפורים צאצאי דינוזאורים, ולא מכמה דינוזאורים לא ידועים, אלא לגמרי קבוצה מסוימת- דינוזאורים מניראפטורים, השייכים לתרופודים, ותרופודים, בתורם, הם אחת מקבוצות הדינוזאורים הלטאות. נמצאו צורות מעבר רבות בין דינוזאורים חסרי מעוף לציפורים.
אבל מה נוכל לומר על אבותיהם של הציפורים אם לא היה תיעוד מאובנים? IN המקרה הטוב ביותרהיינו מגלים שהקרובים הקרובים ביותר של ציפורים הם תנינים. אבל האם נוכל לשחזר את המראה של אבותיהם הישירים של ציפורים, כלומר דינוזאורים? בְּקוֹשִׁי. אבל בדיוק בעמדה הזו אנו מוצאים את עצמנו כאשר אנו מנסים לשחזר את מראה האב הקדמון של הגרעין והציטופלזמה. ברור שזו הייתה קבוצה של כמה דינוזאורים פרוקריוטיים, קבוצה שנכחדה, שבניגוד לדינוזאורים אמיתיים, לא הותירה עקבות ברורים בתיעוד הגיאולוגי. ארכאה מודרנית ביחס לאאוקריוטים היא כמו תנינים מודרנייםכלפי ציפורים. נסו לשחזר את המבנה של דינוזאורים בהכירו רק ציפורים ותנינים.
טיעון בעד העובדה שחיידקים רבים חיו בפרקמבריון, לא דומים לאלה הנוכחיים. סטרומטוליטים פרוטרוזואיים היו הרבה יותר מורכבים ומגוונים מאלה המודרניים. סטרומטוליטים הם תוצר של פעילות חיונית של קהילות מיקרוביאליות. האם זה לא אומר שגם חיידקים פרוטרוזואיקונים היו מגוונים יותר מהמודרניים, ושקבוצות רבות של חיידקים פרוטרוזואיקונים פשוט לא שרדו עד היום?

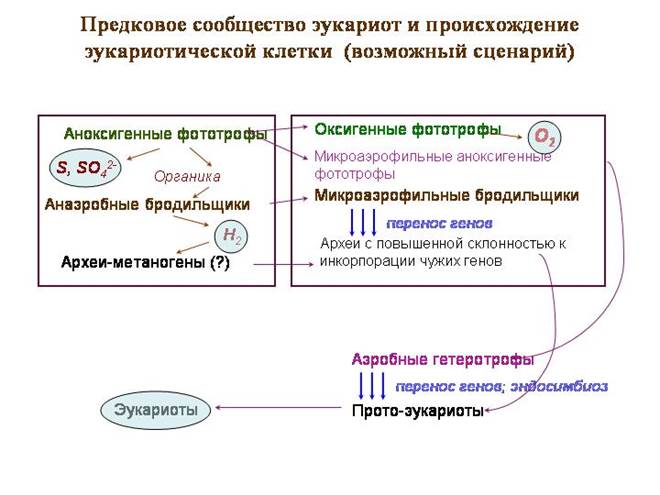
קהילת האבות של האיקריוטים ומקור התא האיקריוטי (תרחיש אפשרי)
"קהילת האבות" ההיפותטית היא מחצלת חיידקים טיפוסית, רק שבחלקו העליון חיו אבותיהם של הציאנובקטריות, שעדיין לא עברו לפוטוסינתזה חמצנית. הם עסקו בפוטוסינתזה נוגדת חמצון. תורם האלקטרונים לא היה מים, אלא מימן גופרתי. כפי ש מוּצָר לְוָאִישוחררו גופרית וסולפטים.
בשכבה השנייה גרו חיידקים פוטוסינתטיים סגולים, כולל אלפאפרוטובקטריה, אבות המיטוכונדריה. חיידקים סגולים משתמשים באור באורך גל ארוך (אדום ואינפרא אדום). לגלים אלה יש את עוצמת החדירה הטובה ביותר. חיידקים סגולים עדיין חיים לעתים קרובות מתחת לשכבה של ציאנובקטריה. אלפאפרוטובקטריה הסגולה משתמשת גם במימן גופרתי כתורם אלקטרונים.
בשכבה השלישית היו חיידקים מתסיסים שעיבדו חומר אורגני; חלקם פלטו מימן כפסולת. זה יצר בסיס לחיידקים מפחיתי סולפט. יכול להיות גם ארכיאה מתנוגנית. בין הארכאים שחיו כאן היו אבותיהם של הגרעין והציטופלזמה.
אירועי משבר החלו עם המעבר של ציאנובקטריה לפוטוסינתזה של חמצן. כתורם אלקטרונים החלו להשתמש בציאנובקטריה במקום מימן גופרתי מים רגילים. זה פתח הזדמנויות גדולות, אבל גם השלכות שליליות. במקום גופרית וסולפטים החלו להשתחרר חמצן במהלך הפוטוסינתזה - חומר רעיל ביותר לכל תושבי כדור הארץ הקדומים.
הראשונים שנתקלו ברעל זה היו מפיקיו, ציאנובקטריה. הם כנראה היו הראשונים שפיתחו אמצעי הגנה מפניו. שרשראות העברת האלקטרונים ששימשו לפוטוסינתזה שונו והחלו לשמש לנשימה אירובית. המטרה המקורית, ככל הנראה, לא הייתה להשיג אנרגיה, אלא רק לנטרל חמצן.
עד מהרה נאלצו תושבי השכבה השנייה של הקהילה - חיידקים סגולים - לפתח מערכות הגנה דומות. בדיוק כמו ציאנובקטריה, הם פיתחו מערכות נשימה אירובית המבוססות על מערכות פוטוסינתטיות. אלו היו אלפאפרוטובקטריה הסגולה שפיתחה את שרשרת הנשימה המושלמת ביותר, שמתפקדת כעת במיטוכונדריה של האיקריוטים.
בשכבה השלישית של הקהילה, הופעת החמצן החופשי בוודאי גרמה למשבר. מתנוגנים ומפחיתי סולפט רבים משתמשים מימן מולקולריבעזרת אנזימי הידרוגנז. חיידקים כאלה אינם יכולים לחיות בתנאים אירוביים מכיוון שחמצן מעכב הידרוגנזים. חיידקים רבים המייצרים מימן, בתורם, אינם גדלים בסביבה שבה אין מיקרואורגניזמים המנצלים אותו. מבין המתסיסים, ככל הנראה הקהילה שמרה על צורות שפולטות תרכובות אורגניות נמוכות (פירובט, לקטט, אצטט וכו') כמוצרים סופיים. מתסיסים אלה פיתחו אמצעי הגנה משלהם מפני חמצן, פחות יעילים. בין הניצולים היו ארכאים - אבות הגרעין והציטופלזמה.
אולי, ברגע זה של משבר, התרחש אירוע מרכזי - היחלשות הבידוד הגנטי באבותיהם של האיקריוטים ותחילת ההשאלה הפעילה של גנים זרים. פרוטו-אאוקריוטים שילבו את הגנים של מתססים שונים עד שהפכו בעצמם לתסיסים מיקרו-אירופיליים, התסיסו פחמימות לפירובט וחומצת חלב.
תושבי השכבה השלישית - אבותיהם של האוקריוטים - היו כעת במגע ישיר עם התושבים החדשים של השכבה השנייה - אלפאפרוטובקטריה האירובית, שלמדו להשתמש בחמצן לאנרגיה. חילוף החומרים של פרוטו-אאוקריוטים ואלפאפרוטובקטריה הפך למשלים, מה שיצר את התנאים המוקדמים לסימביוזה. ועצם מיקומם של אלפאפרוטובקטריה בקהילה (בין השכבה העליונה, המשחררת חמצן, לשכבה התחתונה) קבע מראש את תפקידם כ"מגינים" של אבות קדמונים אוקריוטיים מעודף חמצן.
סביר להניח שפרוטו-אוקריוטים בלעו ורכשו חיידקים רבים ושונים כאנדוסימביונטים. ניסויים מסוג זה עדיין נמשכים. אוקריוטים חד תאיים, שיש להם מגוון עצום של סימביונים תוך תאיים. מבין הניסויים הללו, הברית עם אלפאפרוטובקטריה האירובית התבררה כמוצלחת ביותר.
ביולוגיה וגנטיקה
על פי ההשערה הסימביוטית של אברונים גופניים של התא האיקריוטי הפופולרי כיום בעלי גנום משלהם מאופיינים במקור עצמאי ומקורם תאים פרוקריוטייםסימביונים. בתחילה, כמות המידע בגנום של התא המארח, מחד גיסא, ובסימביונטים של מיטוכונדריה משוערת של צנטריולים וכלורופלסטים, מאידך גיסא, הייתה ניתנת להשוואה. בעתיד, הגנום של הסימביונטים עלול לאבד חלק מתפקודיהם הגנטיים עם תנועת בלוקים של גנים לתוך הגנום...
השערות למקורם של תאים אוקריוטיים.
1. סימביוטי.
על פי ההשערה הסימביוטית, הפופולרית כיום, האברונים הגופניים של תא אוקריוטי, בעלי גנום משלהם, מאופיינים במקור עצמאי ומקורם בתאי סימביון פרוקריוטיים. ככל הנראה, התא המארח היה פרוקריוט אנאירובי, המסוגל רק לתנועת אמבואידים. מיטוכונדריה ופלגלה התעוררו באמצעות שינויים בסימביונטים של תאי המארח, פרוקריוטים אירוביים וחיידקים, הדומים לספירושטים מודרניים. כלורופלסטים של תאי צמחים ירוקים צמחו מסימביונטים - תאים פרוקריוטיים של אצות כחולות-ירוקות. חשיבות מיוחדת הייתה הרכישה על ידי התא במהלך האבולוציה של דגלים עם גופים בסיסיים קרובים לצנטריולים. זה איפשר להתעצם פעילות מוטוריתוליצור מנגנון למיטוזה.
שאלת מקור הגרעין קשה. בתחילה, כמות המידע בגנום של התא המארח, מצד אחד, ובסימביונטים של מיטוכונדריה משוערת, צנטריולים וכלורופלסטים, מצד שני, הייתה כנראה דומה. בעתיד, הגנום של הסימביונטים עלול לאבד חלק מתפקודיהם הגנטיים עם העברת בלוקים של גנים לגנום של התא המארח. זה לווה בגידול בנפח הגנום הגרעיני, ריכוזיות הוויסות הגנטי של תפקודי התא, ואובדן האוטונומיה על ידי המיטוכונדריה ואברונים גופניים אחרים. עם זאת, לא ניתן לשלול שהגרעין הוא גם נגזרת של סימביון תוך תאי. מקורם של הממברנות התוך-פלזמיות של הרשת הציטופלזמית המחוספסת והחלקה, הקומפלקס הלמלרי והמבנים שלפוחית-ואקואולרים מוסבר על ידי טרנספורמציות אבולוציוניות של הממברנה החיצונית מעטפת גרעין. ההשערה הסימביוטית אינה מסבירה מספר עובדות. לפיכך, החלבון bacillin, המרכיב את הריסים והדגלים של פרוקריוטים מודרניים, שונה מחלבון הטובולין של אוקריוטים. לחיידקים לא היו מבנים בהרכב של מיקרוטובולים "9 + 2" או "9 + 0" האופייניים לדגלים, ריסים, גופים בזאליים או צנטריולים של תאים אוקריוטיים.
2. פלישה.
השערת האינוואגינציה של מקור התא האוקריוטי נובעת מהעובדה שצורת האבות הייתה פרוקריוט אירובי. הוא הכיל כמה גנומים, כל אחד מחובר אליו קרום תא. אברונים גופניים עם דנ"א, כמו גם גרעין, התעוררו כתוצאה מהפלישה ושרוכים של שברי הקליפה יחד עם הגנום, ולאחר מכן התמחות תפקודית לגרעין, המיטוכונדריה, הכלורופלסטים, הסיבוך של הגנום הגרעיני וה פיתוח ממברנות ציטופלזמיות. השערה זו מסבירה באופן משביע רצון את נוכחותם של 2 ממברנות במעטפת הגרעין, המיטוכונדריה והכפורופלסטים. היא נתקלת בקשיים בהסבר ההבדלים בפרטי תהליך הביוסינתזה של חלבונים באברוני הגופיות ובציטופלזמה של התא האוקריוטי. במיטוכונדריה ובכלורופלסטים, תהליך זה תואם בדיוק לזה שבתאים פרוקריוטים מודרניים. ההשערות הסימביוטיות וההסתגלות אינן ממצות את כל נקודות המבט על מקור הארגון הסלולרי מהסוג האוקריוטי.
3. הופעת הרב-תאית.
הצורות הקולוניאליות של פרוטוזואה הטרוטרופית היו האב הקדמון של אורגניזמים רב-תאיים.
שלב 1: ליד כל דגל הייתה פועל יוצא של הציטופלזמה צווארון או כיס שאליו הונעו חלקיקי מזון (פגוציטוזיס).
שלב 2: פגוציטים התמחות תאים (תפקוד פגוציטי-עיכול) וקינוציטים (תנועת פונקציה; נשארו בפריפריה). Int. שכבת phagocytoblast + ext. kinocytoblast = phagocytella או parenchymella (Mechnikov).
שלב 3: נוצר פה בקוטב אחד. חור => הוליד ספוגים, coelenterates וכו'.
לפי האקל: כדור. מושבה של ים, על ידי מילוי הנוזל, נוצר אורגניזם דמוי בועה (blastea), פלישה היא יצור דו-שכבתי (גסטרה); (האקל האמין שהראשון היה עיכול בטני).
4. תכונות של רב תאי.
בעלי חיים רב-תאיים מהווים את הקבוצה הגדולה ביותר של אורגניזמים חיים על פני כדור הארץ, המונה יותר מ-1.5 מיליון מינים. מובילים את מוצאם מהפשוטים ביותר, הם עברו טרנספורמציות משמעותיות בתהליך האבולוציה הקשור לסיבוך הארגון.
אחד המאפיינים החשובים ביותר בארגון של אורגניזמים רב-תאיים הוא ההבדל המורפולוגי והתפקודי בין תאי גופם. במהלך האבולוציה, תאים דומים בגוף של בעלי חיים רב-תאיים התמחו בביצוע פונקציות מסוימות, מה שהוביל להיווצרותבדים.
בדים שונים בשילובלאיברים, ולאיברים לתוך מערכות איברים.ליישם את הקשר ביניהם ולתאם את עבודתם,מערכות רגולטוריותעצבני ואנדוקרינית. תודה לעצבניים ויסות הומורליפעילויות של כל המערכות, אורגניזם רב תאי מתפקד כמערכת ביולוגית אינטגרלית.
השגשוג של קבוצה של בעלי חיים רב-תאיים קשור לסיבוך מבנה אנטומיו פונקציות פיזיולוגיות. אז, עלייה בגודל הגוף הובילה להתפתחות תעלת עיכול, מה שאפשר להם לאכול חומר מזון גדול, המספק כמות גדולה של אנרגיה ליישום כל תהליכי החיים. מערכות השרירים והשלד המפותחות הבטיחו תנועת אורגניזמים, שמירה על מבנה גוף מסוים, הגנה ותמיכה באיברים. יכולת ל תנועה אקטיביתאפשרו לבעלי חיים לחפש מזון, למצוא מחסה ולהתיישב.
עם הגידול בגודל גוף החיות, נוצר צורך במראה החיצונימערכות זרימת דם תוך תחבורה,מתן תמיכת חיים לרקמות המרוחקות מפני השטח של הגוף" ואיברים חומרים מזינים, חמצן, כמו גם הסרת תוצרי קצה מטבוליים.
מערכת הובלה מחזורית כזו הפכה לדם רקמה נוזלית.
התעצמות הפעילות הנשימתית הלכה במקביל להתפתחות המתקדמתמערכת עצביםואיברי חישה. היה מהלך מחלקות מרכזיותמערכת העצבים לקצה הקדמי של גוף החיה, וכתוצאה מכך קטע הראש מבודד. מבנה זה של החלק הקדמי של גוף החיה אפשר לו לקבל מידע על שינויים ב סביבהולהגיב להם כראוי.
בעלי חיים רב-תאיים מגוונים ביותר במבנה, בתכונות החיים, שונות בגודל, במשקל גוף וכו'.
השיעור מתאים לתכנית ולספר הלימוד מאת I.N. Ponomareva, המספקים לימוד ציטולוגיה בכיתה י"א. השיעור מתקיים בצורה של דיון מדעי ומיועד ל-90 דקות.
מטרות השיעור: היכרות עם ההשערות החשובות ביותרעל מקור התא האוקריוטי ואורגניזמים רב-תאיים; פיתוח היכולת להתווכח ולהגן על נקודת המבט; טיפוח תרבות של דיון ציבורי.
צִיוּד: טבלאות המתארות את מבנה התא ואת האברונים הבודדים שלו, דיאגרמה של הופעת תא איקריוטי, ציורים המתארים טריכופלקס וצורות שונות של אצות.
במהלך השיעורים
מוֹרֶה.עם התפתחות מדע התא - ציטולוגיה והופעת תוצאות מחקר חדשות של יסודי זה מערכת ביולוגיתויכוחים סוערים התעוררו בין מדענים, שהפכו לדיונים מדעיים רציניים. לאורך ההיסטוריה של הציטולוגיה כמדע, כמעט כל השערה חדשה זכתה גם לתומכיה וגם ליריביה.
דיון מדעי הוא דיון ציבורי בשאלה או בעיה. היום בשיעור נקיים דיון במהלכו נדון בשניים בעיות קריטיותציטולוגיה: בעיית מקורם של האיקריוטים הראשונים ובעיית מקורם של האורגניזמים הרב-תאיים הראשונים.
אתה יודע שהכיתה חולקה בעבר לקבוצות שיציגו ויגנו על נקודות מבט שונות על שני הנושאים הללו שאנו דנים בהם. הרשו לי להזכיר לכם את שמות הקבוצות הללו.
1. תומכים בהשערה האוטוגנית של מקור התא האוקריוטי ואברוניו.
2. תומכים בהשערה הסימביוגנית של מקור התא האוקריוטי ואברוניו.
3. תומכים בהשערת המקור הקולוניאלי של האורגניזמים החד-תאיים הראשונים.
4. תומכים בהשערת מקורם של אורגניזמים רב-תאיים מריסיות על ידי תאי תאי של התאים שלהם.
נציגי כל קבוצה במצגת קצרה יציינו את המהות של נקודת מבט כזו או אחרת על כל אחת מהבעיות הנדונות, ולאחר מכן יענו על שאלות, הערות, התנגדויות הן של "יריביהם" המדעיים והן של כל הנוכחים בכיתה.
בעיית מקור אברוני ממברנהוגרעינים של תאים אוקריוטיים הוא אחד מנושא הדיון האקטואלי ביותר של הציטולוגיה המודרנית. כדי לפתור בעיה זו, הוצעו כמה השערות שונות במדע. בואו נסתכל על שניים מהחשובים שבהם.
השערה אוטוגנית של מקור התא האוקריוטי
נאום של נציגי קבוצת התלמידים הראשונה
האיקריוטים הראשונים הופיעו על פני כדור הארץ לפני 1.1-1.4 מיליארד שנים (בעידן הפרוטרוזואיקון). זו הייתה אבן הדרך הגדולה הבאה באבולוציה הקדם-קמברית לאחר הופעת הפוטוסינתזה.
ההשערה האוטוגנית, או הרציפה, קובעת שתא איקריוטי מורכב התפתח ישירות מתא פרוקריוטי: כתוצאה מהפלישה של קרום הפלזמה, ולאחר מכן סידורו מחדש לביצוע פונקציות מסוימות, קמו האברונים החשובים ביותר. ואברונים מורכבים כמו מיטוכונדריה ופלסידים מקורם במבני הממברנה התוך פלזמית של המבנה הצינורי הקיים בפרוקריוטים.
שאלות
1. מה תומך בנקודת מבט זו? האם יש צורות מעבר, כלומר. תאים פרוקריוטיים עם "בסיסים" של אברונים אוקריוטיים?
תשובה.אין צורות שריד כאלה בקרב אורגניזמים חיים. אבל הבה נזכיר את נוכחותם של מזוזומים בתאי חיידקים, כלומר. מבני ממברנה בעלי צורה צינורית ושלפוחית (כדורית), הנוצרים על ידי פלישה של קרום הפלזמה לתוך הציטופלזמה. ההנחה היא שמזוזומים מעורבים ביצירת דפנות תאים, שכפול DNA ותהליכים נוספים. בהחלט ייתכן שעל בסיס מזוזומים התעורר בהדרגה קומפלקס הממברנה והתמיין, והוליד אברונים שונים של התא האוקריוטי.
2. ידוע כי ההרכב הביוכימי של מספר אברונים, בפרט מיטוכונדריה וכלורופלסטים, שונה מאוד מהרכב קרום הפלזמה של הציטופלזמה. האם זה לא מעיד שמקורם אינו מהמקור קרום תא- ממברנות פלזמה?
תשובה.ואכן, הרכב החלבון של המיטוכונדריה והכלורופלסטים הוא ייחודי. אבל מקוריות זו יכולה להירכש בפעם השנייה, יכולה להיות תוצאה של הסתגלות לביצוע פונקציות מסוימות על ידי האברונים הללו.
מוֹרֶה.בואו נקשיב לטיעונים של תומכי המקור הסימביוגני של האיקריוטים ונקבע איזו השערה פופולרית יותר בכיתה.
השערה סימביוגנית של מקור התא האוקריוטי
נאום של נציגי קבוצת התלמידים השנייה
ההשערה הסימביוגנית (כיום נקראת לעתים קרובות תיאוריה) נובעת מהעובדה שאאוקריוטים הם תוצאה של סימביוזה בין פרוקריוטים שונים.
כבר בתחילת המאה ה-20. הבוטנאים הרוסים א.ס. פמינסין, ב.מ. קוזו-פוליאנסקי וק.ס. מרז'קובסקי העלה השערה לפיה תא הצמחים הירוקים (אוקריוטים) קיבל פלסטידים כתוצאה מסימביוזה של תא נטול כלורופיל עם תאים של אצות כחולות. השערה זו של מקורו הסימביוגני של התא האוקריוטי הקדימה את זמנה, נשכחה, ושוב משכה תשומת לב באמצע המאה ה-20.
ההשערה ששמה פותחה על ידי Lynn Sagan-Margulis (1983). על פי השערה זו, התא הראשוני של חיידק פרוקריוטי גדול, לאחר שנכנס לסימביוזה עם תאי האצות הכחולות-ירוקות, רכש פלסטידים. סימביוזה עם תאים פרוקריוטים הטרוטרופיים הובילה להפיכתם למיטוכונדריה. כמה תאים, בהיותם הטרוטרופים, לכדו חיידקים אחרים, קטנים יותר, שהם סיבות לא ידועותלא עיכל. תאים פרוקריוטים קטנים שנלכדו הצליחו לספוג חמצן. תכונה זו הייתה מועילה לתא שבלע את החיידק, שכן היא העניקה לו הרבה יותר אנרגיה שנאגרה במולקולות ATP.
שאלות
1. אילו עדויות ניתנות לטובת המקור הסימביוגני של פלסטידים ומיטוכונדריה?
תשובה.הראיות לדעה זו הן כדלקמן.
1. מיטוכונדריה וכלורופלסטים מוקפים בקרום כפול.
2. אברונים אלו מתרבים בחלוקה, ואינם נושרים מאברוני קרום אחרים.
3. למיטוכונדריה ולכלורופלסטים יש חומר גנטי משלהם, המקודד רק לחלבונים שלהם.
4. ל-DNA של האברונים הללו יש מבנה מעגלי, כמו בפרוקריוטים, ולא ליניארי, כמו באיקריוטים.
5. למיטוכונדריה ולכלורופלסטים יש מנגנון סינתזת RNA וחלבונים משלהם, והריבוזומים שלהם פרוקריוטיים יותר מאשר אוקריוטיים.
6. חלק מהחלבונים של האברונים הללו דומים במבנה הראשוני שלהם לחלבונים חיידקיים דומים ואינם דומים לחלבונים הציטופלזמיים המתאימים.
2. ידוע שלא כל החלבונים של המיטוכונדריה והכלורופלסטים מסונתזים על פי ה-DNA שלהם; חלק מהחלבונים של האברונים הללו מקודדים ב-DNA של הגרעין. איך אתה יכול להסביר עובדה זו?
תשובה.ואכן, רק חלק מהחלבונים שלהם מקודדים ב-DNA הפרטי של המיטוכונדריה והכלורופלסטים, בעוד השאר מקודדים ב-DNA של גרעין התא. ככל הנראה, במהלך האבולוציה, חלק מהחומר הגנטי "זרם" מהגנומים של המיטוכונדריה והכלורופלסטים אל הגנום הגרעיני. הגנום של הכלורופלסט הוא נפחי יותר מהגנום המיטוכונדריאלי, והגנום המיטוכונדריאלי של אוקריוטים נמוכים יותר (לדוגמה, שמרים) נפחי יותר מזה של יונקים, מה שמאשר בעקיפין את ההנחה לעיל.
3. אנו יודעים שבנוסף למיטוכונדריה ולכלורופלסטים, כמות קטנה של DNA בתא האאוקריוטי מצויה גם בצנטריולים ובבסיס הפלגלה. האם הם עלו גם באופן סימביוגני?
תשובה.כן. סימביוזה עם חיידקים דמויי ספירוצ'טים עלולה להוביל להופעת דגלים, מצד אחד, ולהפיכת חלק מהתאים דמויי הספירוצ'טים הללו לצנטריולים, מצד שני. התפיסה המודרנית של סימביוגנזה קובעת שהתא האוקריוטי נוצר כתוצאה מכמה פעולות עוקבות של סימביוגנזה.
4. כיצד נוצרו שאר האברונים של התא האוקריוטי (קומפלקס גולגי, EPS, ליזוזומים)?
תשובה.ההנחה היא שהרשת האנדופלזמית, הליזוזומים ומנגנון הגולגי יכלו להתעורר על ידי פלישה של קרום הפלזמה.
5. כיצד, לפי ההשערה הסימביוגנית, יכול היה להיווצר גרעין מוקף בקרום כפול?
תשובה.ההנחה היא שהגרעין התפתח גם כתוצאה מבידוד ה-DNA מהציטופלזמה על ידי פלישה של קרום הפלזמה ועוטפת החומר הגרעיני.
6. מהתשובות שלך לשתי השאלות האחרונות, עולה כי הופעתו של תא אוקריוטי אינה רק תוצאה של סימביוזה תורשתית של מספר פרוקריוטים. מה אתה אומר על זה?
תשובה.כן זה כן. תומכי ההשערה הסימביוגנית מודים שכמה מבנים של התא האוקריוטי התעוררו כתוצאה מהתפתחותו העצמית והסיבוך שלו.
7. הבאת נתונים ביוכימיים, גנטיים, מיקרוסקופיים אלקטרונים שנים האחרונותלטובת המקור הסימביוגנטי של התא האוקריוטי. אבל האם יש שרידים אבולוציוניים, מה שנקרא מאובנים חיים, המצביעים על קשרים בין פרוקריוטים לאאוקריוטים?
תשובה.יחסים כאלה נצפים בין אורגניזמים של צמחים ובעלי חיים. למשל, האמבה החיה חופשית Pelomyxa palustris(Pelomyx marsh) אין מיטוכונדריה, אך מכיל חיידקים סימביוטיים המספקים לו נשימה.
מוֹרֶה.נראה לי שנשאלו מספיק שאלות לנציגי הקבוצה השנייה. תומכים באיזו השערה הביעו, לדעתך, טיעונים משכנעים יותר ואיזו השערה מוכרת יותר במדע?
ואכן, ההשערה הסימביוגנית מוצאת מספר רב של תומכים. אך יש לזכור כי לשיקולים המובעים לטובת ההשערה הסימביוגנית אין אישור ניסיוני, ולכן לא ניתן להתייחס אליה כתיאוריה. לדעתי, חשוב גם לזכור שלסימביוזה היה תפקיד בולט, אך לא היחיד, בהופעתו של התא האיקריוטי. חֲשִׁיבוּתביצירת איקריוטים, היו גם תהליכים של התפתחות עצמית של תאים פרוקריוטים.
בואו נעבור ל הבעיה הבאה- בעיית מקורם של אורגניזמים רב-תאיים.
השערות למקור הקולוניאלי של המטאזואים הראשונים
מוֹרֶה.ההתקדמות בהתפתחות הציטולוגיה והביולוגיה המולקולרית הוכיחה את הקשר המבני והביוכימי של אורגניזמים חד-תאיים ורב-תאיים. כל זה אישר את ההשערה של מקורם של בעלי חיים רב-תאיים מ צורות חד תאיות. המחלוקות המדעיות העיקריות נוגעות לשאלות מאיזה פרוטוזואה הגיעו בעלי חיים רב-תאיים ואיך נראו החיות הרב-תאיות הראשונות? ישנן השערות רבות לגבי מקורם של בעלי חיים רב-תאיים מפרוטוזואה. בואו נשקול כמה מהם.
נאום של נציגי קבוצת התלמידים השלישית
נצפה בצדק שיש השערות רבות לגבי מקורם של בעלי חיים רב-תאיים מפרוטוזואה. המנומק מביניהם, לדעתנו, צריך להיחשב כהשערה הקולוניאלית.
ההשערה הקולוניאלית הראשונה למקורם של אורגניזמים רב-תאיים הוצעה על ידי ארנסט האקל. הוא פיתח את "תיאוריית הגסטרה" שלו, לפיה האב הקדמון המשותף של כל בעלי החיים הרב-תאיים דומה לזחל שוחה חופשי של coelenterates - אלמוגים בעלי שתי שכבות של תאים.
הביולוג הרוסי איליה איליץ' מכניקוב הציע השערה נוספת: בעלי חיים רב-תאיים צמחו מפרוטוזואה קולוניאלית פלגלרית המסוגלת לפאגוציטוזיס, כלומר. לכידת מזון על ידי תאים עם עיכול תוך תאי. תיאוריה זו מניחה שתאי פני השטח שלכדו מזון שוקעים עמוק בגוף המושבה לצורך עיכול, ומפנים מקום לתאים רעבים. התוצאה היא אורגניזם שבו תאי השכבה הפנימית מעכלים מזון, והשכבה החיצונית, המורכבת מתאים רעבים, לוכדת אותו. הוא גם מבצע את הפונקציות של קליטה, תנועה והגנה. מכניקוב כינה את האורגניזם המוצע phagocetella (אורגניזם המורכב מתאים העוסקים בפאגוציטוזה).
לבסוף, ההשערה השלישית הועלתה על ידי הזואולוג הגרמני O. Byuchli. לדבריה, האורגניזם הרב-תאי המקורי היה מורכב משתי שכבות של תאים, יתרה מכך, עם צידו התחתון הוא זחל לאורך הקרקע, ו הצד העליוןהיו לו פונקציות הגנה ורגישות. לאחר שפגש מזון בגדלים גדולים, אורגניזם כזה התעטף סביבו ועכל אותו בשכבה התחתונה. בוטשלי קרא לאורגניזם המשוער הזה פלאקולה (בעל חיים בצורת עוגה עוטפת).
שאלות
1. איזו משלוש ההשערות הללו אתה מעדיף? מי מהם הוא המוכר ביותר במדע?
תשובה.קשה לתת עדיפות להשערה כלשהי, שכן אין נתונים פליאונטולוגיים על הצעדים הראשונים באבולוציה של בעלי חיים רב-תאיים. מדענים מנסים למצוא אישור להשערות המקור הקולוניאלי של בעלי חיים רב-תאיים על ידי השוואת אורגניזמים היפותטיים עם צורות פרימיטיביות חיות של בעלי חיים רב-תאיים.
בעל החיים הרב-תאי הפרימיטיבי ביותר הידוע כיום הוא Trichoplax (איור 1). זה תואר על ידי ארטמי ואסילביץ' איבנוב (1973). טריכופלקס נמצא בחוף האירופי של האוקיינוס האטלנטי. לחיה זו יש מראה של צלחת דקה של שתי שכבות של תאים עם דגלים. בחלל גופו ישנם תאי עיכול נפרדים. בין שתי שכבות התאים יש תאים שנראים כמו אמבות.
לאחר שלמד את Trichoplax, A.V. איבנוב הגיע למסקנה שהוא דומה מאוד לפאגוצטלה ההיפותטית I.I. Mechnikov והוא, למעשה, המודל החי שלה. מצד שני, טריכופלקס גם נראה כמו פלאקולה. לפיכך, הגילוי של Trichoplax, בדומה לזחל של coelenterates, מפגיש את ההשערות של האקל (1866), מכניקוב (1877) ובוקלי (1884).
2. במשך זמן רב, מקורם של בעלי חיים רב-תאיים פרימיטיביים היה קשור לדגלים קולוניאליים ירוקים כגון אודורינה, וולבוקס, פנדורינה. האם הם לא נחשבים כעת כצורות מעבר לרב-תאיות?
תשובה.אכן, מקורם של בעלי חיים רב-תאיים אינו קשור עוד לדגלים קולוניאליים ירוקים. נקודת מבט זו מבוססת על כך שהתפתחותם ורבייתם האינדיבידואלית קרובים לאצות, ולא לבעלי חיים רב-תאיים פרימיטיביים.
3. אילו בעלי חיים רב-תאיים פרימיטיביים הולידו אורגניזם קולוניאלי היפותטי? האם הם היו ספוגים, מגניבים, או נציגים של סוג אחר של בעל חיים?
תשובה.ההנחה היא שכמה סוגים של בעלי חיים רב-תאיים ברמות ארגון שונות יצאו מאבות קדמונים דמויי פגוצטלה בבת אחת: ספוגים, בעלי חיים תלת-שכבתיים פרימיטיביים הקרובים לפלנרים שאינם מעיים השייכים לתולעים שטוחות. ספוגים ו-coelenterates הם ענפים ללא מוצא של האבולוציה. ההתמחות הצרה של טיפוסים אלו, הסתגלותם לאורח חיים לא תנועתי או פסיבי סגרה את האפשרויות להמשך התפתחותם המתקדמת.
4. אתה ממשיך לדבר על מקורם של החיות הרב-תאיות הראשונות, אבל הרב-תאיות אופיינית לא רק לנציגי ממלכת החיות. איך יכול להיות פטריות רב תאיות, צמחים?
תשובה.המדע המודרני מאמין שלצמחים אין קו חד בין חד-תאי לרב-תאי. הרב-תאי התעורר שוב ושוב ללא תלות זה בזה במחלקות שונות של צמחים. אז, באצות מוזהבות, מרובות הלקות, פירופיות, ירוקות, נצפים מינים בעלי מבנה חוטי, קולוניאלי-רב-תאי וסיפון (הגוף מורכב מתא ענק אחד בעל גרעינים). נוכחותן של צורות כאלה מעידה, לדעתנו, על ניסיונות לפרוץ לדרגת הארגון הבאה - רב-תאית, שבה תאים שונים מבצעים פונקציות שונות(איור 2).
אורז. 2. צורות שונותאצות (1 - חד תאיות;
2 - קולוניאלי; 3 - רב ליבות; 4 - חוטי)
המידע הזמין למדע המודרני על התפתחותן של צורות רב-תאיות בפרקמבריון הוא דל. זה נכון במיוחד עבור פטריות. הסיבה היא שימור לקוי של שרידי האורגניזמים הללו. כמו במספר צמחים, פטריות עוברות מעבר ממבנה לא תאי (סיפונל, כמו למשל בפטריית העובש mucor) לרב תאי. כך, לדעתנו, בשלב מסוים בהתפתחות פטריות (לפני כ-1.3 מיליארד שנה), בחלקן, הפך התפטיר הסיפונל להיווצרות רב-תאית.
מוֹרֶה.בואו נסכם את מה שנאמר. אורגניזמים רב-תאיים, לפי מדע מודרני, הופיע קודם לכן בין צמחים ופטריות. בבעלי חיים, הראשון צורות רב תאיותמקורו לפני כ-700 מיליון שנה. תחילתן של בעלי חיים רב-תאיים ניתנה על ידי אורגניזמים דגלים קולוניאליים הטרוטרופיים עתיקים.
השערה של מוצא לא קולוניאלי של המטאזואים הראשונים
נאום של נציגי קבוצת התלמידים הרביעית
נציגי הקבוצה הקודמת מתעקשים כי אבותיהם הרחוקים של בעלי חיים רב-תאיים היו מושבות של פרוטוזואה. אמנם זו נקודת מבט נפוצה, אבל לא היחידה.
אנו מציגים השערה המצביעה על כך שבתהליך האבולוציה, פרוטוזואה בודדות הפכו לחלוטין ליצורים רב-תאיים. רעיון זה הועלה על ידי הזואולוג הסרבי המפורסם I. Hadji. לדעתו, בעלי חיים רב-תאיים התפתחו מריסים רב-גרעיניים.
לסילאטים יש מבנה מורכב למדי. הציטופלזמה שלהם מיוצגת על ידי שתי שכבות - היקפית ומרכזית, שבהן מתרחש עיכול תוך תאי. לריסיות יש פה תאי, לוע, ואקואולים מתכווציםעם תעלות adductor. כל החלקים הללו המובחנים באופן שונה אורגניזם חד תאי- אברונים - האדג'י מחשיב את אב הטיפוס של האיברים של בעל חיים רב תאי. כן, הוא חושב כך עורמקורם של יצורים רב-תאיים מהשכבה החיצונית של הציטופלזמה (אקטופלזמה), ומהמעיים שלהם - מהשכבה הפנימית של הציטופלזמה (אנטופלזמה).
המעבר ממצב חד-תאי לרב-תאי יכול להתרחש בגוף הריסת מיד על ידי היווצרות גבולות תאים סביב גרעינים בודדים ומקטעים סמוכים של הציטופלזמה. תהליך זה כביכול נקרא "תלולריזציה" (מהמילה הלטינית cellula- תא), וההשערה עצמה נקראת תורת התאיות.
שאלות
1. האם להשערה של I. Hadji יש תמיכה כלשהי באמבריולוגיה של אורגניזמים רב-תאיים נמוכים יותר, כמו, למשל, ההשערה של I.I. מכניקוב?
תשובה.למרבה הצער, אין נתונים כאלה של אמבריולוגיה השוואתית. איש מעולם לא צפה בתהליך של תאי תאי של ריצות בטבע.
2. אתה לא חושב שההשערה של האדג'י לא עומדת בביקורת, בעיקר בגלל שעצם העיקרון העומד בבסיסה הוא מרושע. השערת התאיות, לדעתנו, סותרת את ההוראות העיקריות של התיאוריה התאית, כלומר: היא משווה חלקים של תא בודד לרקמות ואיברים של אורגניזמים רב-תאיים. מה אתה יכול להגיד על זה?
תשובה.לא נהיה כל כך קטגוריים בשיפוטים. ההשערה של האדג'י אינה משווה בין אברוני הריסים לרקמות ואיברים של אורגניזמים רב-תאיים. היא רק סופרת חינוך אפשריאיברים דומים המבוססים על אברוני ריסים.
3. על מה עוד מבוססת ההשערה של האדג'י, מלבד העובדה שיש ריסיות מבנה מורכבוהמבנים שלהם יכולים להפוך לאיברים של בעלי חיים רב-תאיים?
תשובה.אישור עקיף להשערה הוא דמיון מסוים בין ריצות לטורבלריות - פרימיטיביות תולעים שטוחות: גדלים קרובים, אותה צורת גוף, מיקומם של מספר מבנים וכו'. (איור 3 ו-4).
מוֹרֶה. הקשבנו לטיעונים בעד המוצא הקולוניאלי והלא קולוניאלי של בעלי החיים הרב-תאיים הראשונים, התוודענו לדעות על מקורם של צמחים ופטריות רב-תאיים. מה, לדעתך, השערה - המקור הקולוניאלי או הלא-קולוניאלי של בעלי חיים רב-תאיים - מנומקת ומוכחת יותר?
בהחלט, ההשערה של המקור הקולוניאלי של החיות הרב-תאיות הראשונות. השערה זו זוכה להכרה הרחבה ביותר בעולם המדעי. אבל צריך לזכור שההשערה השנייה כל הזמן מתפתחת ומשתנה.
מסכם את השיעור
1. ציון לשיעור.
2. שיעורי בית:
- ללמוד את הפסקה "בעיות הניתנות לוויכוח של ציטולוגיה";
ו – להכיר בבית את אחת מבעיות הציטולוגיה שנדונו במאה ה-18, שהתבצעה בין תומכים שונים בפרפורמיזם; להכין מערכת טיעונים לטובת נקודות מבט שונות.