שאלה 1. מהם ההבדלים במבנה של תאים אאוקריוטיים ופרוקריוטיים?
לפרוקריוטים אין גרעין בצורת אמיתית (קריון ביוונית - גרעין). ה-DNA שלהם הוא מולקולה עגולה אחת, הממוקמת בחופשיות בציטופלזמה ואינה מוקפת בממברנה. בְּ תאים פרוקריוטייםאין פלסטידים, מיטוכונדריה, רטיקולום אנדופלזמי, מנגנון גולגי, ליזוזומים. גם לפרוקריוטים וגם לאאוקריוטים יש ריבוזומים (לגרעיניים יש גדולים יותר). הדגל של תא פרוקריוטי דק יותר ופועל על עיקרון שונה מהדגל של האוקריוטים. אורגניזמים אוקריוטיים הם פטריות, צמחים, בעלי חיים - חד-תאיים ורב-תאיים; פרוקריוטים - חיידקים ואצות כחולות ירוקות (ציאנובקטריה).
שאלה 2. ספר לנו על פינו - ופגוציטוזיס. במה שונים תהליכים אלו?
קרום התא הוא תצורה ניידת המסוגלת ללכוד אובייקטים מהסביבה החיצונית על ידי יצירת פלישות ויציאות. תהליך זה נקרא אנדוציטוזיס. הגורם לאנדוציטוזיס הוא תגובות ביוכימיות מורכבות המתרחשות בציטופלזמה וקשורות בעיקר לשינויים במבנה השלישוני של חלבונים תוך תאיים. אם התא לוכד טיפת נוזל - זו פינוציטוזיס, אם חלקיק מוצק - פגוציטוזיס. כתוצאה מכך נוצרים ואקוולים פינוציטיים או פגוציטים (שלפוחית ממברנה). התהליך, הפוך לאנדוציטוזיס (שחרור תכולת ה-vacuoles מהתא), נקרא אקסוציטוזיס.
שאלה 3. הרחב את הקשר בין המבנה והתפקודים של קרום התא.
ידוע שהבסיס של כל ממברנה הוא דו-שכבה (שכבה כפולה) של פוספוליפידים, שבה ה"ראשים" ההידרופיליים של מולקולות (גליצרול) מופנים כלפי חוץ, והשאריות ההידרופוביות. חומצות שומן- בפנים. מולקולות של חלבונים קשורות לדו-שכבת השומנים, שיכולה לצמוד לממברנה משני הצדדים, לשקוע לתוכה או אפילו לחדור אליה. מיקומו של קרום התא בגבול התא ו סביבהמגדיר את תפקידיו העיקריים. דו-שכבה חזקה, אלסטית, המתחדשת בקלות, היא מחסום המבטיח את קביעות הסביבה התוך-תאית ומגן על הציטופלזמה מפני חדירת חומרים זרים. פונקציית ההובלה של הממברנה היא סלקטיבית. מולקולות קטנות לא טעונות (02, N2) חודרות בקלות ישירות דרך הדו-שכבה. חלקיקים גדולים יותר ו/או טעונים (Na+, K+, חלק מההורמונים) עוברים דרך נקבוביות חלבון מיוחדות (תעלות) או מועברים על ידי חלבוני נשא. בהיותו מבנה נייד, קרום התא יכול לבצע גם הובלה של חומרים באמצעות אנדו - ואקסוציטוזיס.
שאלה 4. אילו אברוני תאים נמצאים בציטופלזמה?
ניתן לחלק את האברונים הממוקמים בציטופלזמה של תאים אוקריוטיים לשלוש קבוצות: ממברנה אחת, ממברנה כפולה ולא ממברנה. אברונים חד-ממברניים כוללים את הרשת האנדופלזמית (חלק ומחוספס), את מנגנון הגולגי, ליזוזומים ו-vacuoles. שתיים אברונים קרומייםהם פלסטידים ומיטוכונדריה; ללא ממברנה - ריבוזומים, ציטושלד ו מרכז תאים.
שאלה 5. תאר את האברונים של הציטופלזמה וחשיבותם בחיי התא.
הרטיקולום האנדופלזמי (ER) הוא אוסף של ואקוולים, תעלות וצינוריות. הוא יוצר רשת אחת בתוך הציטופלזמה, בשילוב עם הממברנה החיצונית. מעטפת גרעין. הבדיל בין EPS חלק למחוספס. ER חלק מעורב בסינתזה של שומנים ופחמימות, וגם מנטרל חומרים רעילים. ריבוזומים ממוקמים על פני הממברנות של ה-ER המחוספס.
מנגנון הגולגי הוא אברון בעל ממברנה אחת המהווה חלק מרשת ממברנה אחת של התא ומהווה ערימה של מיכלים שטוחים. זה המקום שבו מתבצע המיון והאריזה הסופיים של מוצרי פסולת תאים לתוך שלפוחיות ממברנות (ואקואולות). בין היתר, מנגנון גולגי יוצר ליזוזומים ומספק אקסוציטוזיס.
ליזוזומים הם שלפוחיות קרומיות קטנות המכילות אנזימים לעיכול חומרים מזינים. ליזוזומים מתמזגים עם ה-Vacuole האנדוציטי ליצירת ה-Vacuole של העיכול. אם תוכן הליזוזומים משתחרר בתוך התא עצמו, מתרחשת האוטוליזה שלו (עיכול עצמי של התא).
מיטוכונדריה מסווגים כאברונים דו-ממברניים. הקרום החיצוני שלהם חלק, והפנימי יוצר קפלים (קריסטה). המיטוכונדריה הן תחנות האנרגיה של התא, תפקידן העיקרי הוא סינתזה של ATP.
פלסטידים הם אברונים דו-ממברניים של תאי צמחים. ישנם שלושה סוגים של פלסטידים: כלורופלסטים, כרומופלסטים ולקופלסטים. לוקופלסטים חסרי צבע מאחסנים עמילן; כלורופלסטים ירוקים מבצעים פוטוסינתזה; כרומופלסטים כתומים, צהובים ואדומים מספקים צבע פירות ופרחים (מושכים מאביקים ומפזרים זרעים). נחשב למבוסס שבעבר הרחוק המיטוכונדריה והפלסטידים מקורם בפרוקריוטים, "נבלעו" על ידי תא איקריוטי ונכנסו איתו לסימביוזה. למיטוכונדריה ולפלסטידים יש DNA מעגלי, המסנתזים חלק מהחלבונים באופן עצמאי, והריבוזומים שלהם קטנים יותר מהאיקריוטים.
ריבוזומים הם אברונים קטנים רבים שאינם קרומיים הנוצרים על ידי שתי יחידות משנה - גדולות וקטנות. יחידות משנה מורכבות מחלבון ו-RNA ריבוזומי. תפקיד הריבוזומים הוא סינתזת חלבון. חלק מהריבוזומים ממוקמים ישירות בציטופלזמה, וחלק נמצאים על הממברנות של ה-ER המחוספס.
מרכז תאים - אורגנואיד של מבנה לא קרומי של תאי בעלי חיים, פטריות ו צמחים נמוכים יותר. הוא מורכב משני צנטריולים, הדומים בצורתם לגלילים ומורכבים מצינורות החלבון הקטנים ביותר; משתתף ביצירת ציר הביקוע.
ואקוולה היא שלפוחית קרומית מלאה במוהל תאים. זה חייב להיות קיים בתא הצמח. תפקידו של ה-vacuole הוא הצטברות של מים, מלחים וחומרי הזנה. הוא עשוי גם להכיל פיגמנטים (כחול, סגול) ולצבור תוצרי פסולת של חילוף חומרים.
שלד הציטו הוא אברון שאינו ממברנה, שהוא צינור חבל חלבון הממוקם הן ליד הממברנה והן בציטופלזמה. תפקידם לשמור על צורת התא, להבטיח את ההובלה התוך תאית של חומרים, כמו גם את התנועה הפעילה של התא (אמבה, phagocyte). שלפוחית ממברנה המכילה חומרים כלשהם יכולה לנוע לאורך הצינורות של שלד הציטו, כמו על מסילות.
איך מורידים חיבור בחינם? . וקישור לחיבור הזה; תא איקריוטי. ציטופלזמה. אברוניםכבר בסימניות שלך.חיבורים נוספים בנושא
שאלה 1. היכן נוצר הליזוזום? ליזוזומים הם מבני ממברנה המכילים אנזימים פעילים רבים המעורבים בפירוק תרכובות מקרומולקולריות: חלבונים, שומנים, פחמימות. ליזוזומים נוצרים במתחם גולגי, שם הם נכנסים ושם מצטברים אנזימים. שאלה 2. מה תפקיד המיטוכונדריה? מיטוכונדריה - מבני תאיםמכוסה בקרום כפול. על הממברנה הפנימית, שיש לו שפעים רבים, ממוקם כמות גדולהאנזימים המעורבים בסינתזת ATP. לָכֵן, פונקציה עיקריתמיטוכונדריה - מספקת לתאים אנרגיה באמצעות סינתזה של ATP. שאלה 3. מה
שאלה 1. אילו סימנים לפרימיטיביות של פרוקריוטים בהשוואה לאאוקריוטים תוכל למנות? לפרוקריוטים חסרים גרעין וכרומוזומים. המידע התורשתי של פרוקריוטים מיוצג על ידי מולקולת DNA בודדת, אשר סגורה בצורה של טבעת וממוקמת בציטופלזמה. הריבוזומים בפרוקריוטים קטנים מאוד. L אברונים מוקפים ממברנות (רשתית אנדופלזמית, מנגנון Golgi, מיטוכונדריה, פלסטידים), לפרוקריוטים אין בכלל. בתא איקריוטי, מבנים אלו מיועדים לסינתזה של תרכובות שונות. בשל היעדרם בפרוקריוטים של התגובה
שאלה 1. ממה נוצרים דפנות הרשת האנדופלזמית ומכלול הגולגי? הקירות של הרשת האנדופלזמית וקומפלקס גולגי נוצרים על ידי קרום חד-שכבתי. שאלה 2. שם את הפונקציות של הרשת האנדופלזמית. הרטיקולום האנדופלזמי (ER) יוצר את מערכת ההובלה של התא. ב-ER החלק מתבצעת סינתזה של שומנים ופחמימות. ב-ER המחוספס (גרגירי), חלבונים מסונתזים עקב עבודת הריבוזומים המחוברים לממברנות ה-ER. שאלה 3. מה תפקידם של הריבוזומים? תפקידם העיקרי של הריבוזומים הוא סינתזת חלבון. שאלה 4. מדוע רוב הריבוזומים ממוקמים על הערוצים של האנדופלזמה
מבחן בנושא "חזזיות. אצות 1. פוטוסינתזה באצות מתרחשת ב: א) הגרעין ג) הציטופלזמה ב) כרומטפורים ד) כלורופלסטים 2. חזזית כמו אורגניזם בודדשיטת ההתרבות היא אינהרנטית: א) מינית ג) פרתנוגנזה ב) א-מינית ד) אין תשובה נכונה 3. חזזיות נקראות מדדים של טוהר האוויר, שכן הן: א) מטהרות את האוויר ב) מזהמות את האוויר ג) יוצרות בית גידול עבור צמחים ובעלי חיים אחרים ד) גדלים רק במקומות עם אוויר נקי 4. האם הם מסוגלים תנועה אקטיביתאַצָה? א) כל האצות מסוגלות לנוע ב) לזוז
שאלה 1. מהן הפונקציות קרום חיצוניתאים? קרום התא החיצוני מורכב משכבת שומנים כפולה וממולקולות חלבון, שחלקן ממוקמות על פני השטח, וחלקן חודרות לשתי שכבות השומנים דרך ודרכן. קרום התא החיצוני מבצע תפקיד מגן, המפריד את התא מהסביבה החיצונית ומונע פגיעה בתכולתו. בנוסף, קרום התא החיצוני מספק הובלה של חומרים אל תוך התא וממנו, מאפשר לתאים ליצור אינטראקציה זה עם זה. שאלה 2. באילו דרכים חומרים שונים יכולים להיכנס לתא? דרך החיצוני
שאלה 1. מהם השלבים העיקריים בהופעתם והתפתחותם של החיים על פני כדור הארץ? ישנן השערות רבות המנסות להסביר את מקורם והתפתחותם של החיים על הפלנטה שלנו. ולמרות שהם מציעים גישות שונות לפתרון בעיה זו, רובם מניחים את קיומם של שלושה שלבים אבולוציוניים: אבולוציה כימית, פרה-ביולוגית וביולוגית. בשלב האבולוציה הכימית התרחשה סינתזה אביוגנית של פולימרים אורגניים. בשלב השני נוצרו קומפלקסים של חלבון-חומצה גרעין-ליפואיד (מדענים קראו להם אחרת: קואצרבטים, היפר-מחזורים, פרוביונים, אבות
שאלה 1. מהם תפקידיו של מרכז התא? מרכז התא מבצע את הפונקציה של יצירת השלד הפנימי של התא (ציטושלד). שלד הציטו הוא רשת של מיקרוטובולים החודרים לציטופלזמה, שומרים על צורת התא, מבטיחים את התנועה של אברוני התא, כמו גם עבודה של אברוני תנועה מיוחדים - ריסים ודגלים. מרכז התא גם מבטיח חלוקת תאים תקינה. הצנטריולים של מרכז התא מתפצלים לקטבים של התא המתחלק ויוצרים ציר חלוקה, שבגללו נוצרים לאחר מכן שני תאי בת מתא אם אחד. צנטרולים מורכבים מצילינדרים
ציטופלזמה- חלק חובה של התא, מוקף בין קרום הפלזמה לגרעין; הוא מחולק להיאלופלזמה (החומר העיקרי של הציטופלזמה), אברונים (מרכיבים קבועים של הציטופלזמה) ותכלילים (מרכיבים זמניים של הציטופלזמה). ההרכב הכימי של הציטופלזמה: הבסיס הוא מים (60-90% מהמסה הכוללת של הציטופלזמה), תרכובות אורגניות ואי-אורגניות שונות. הציטופלזמה היא בסיסית. תכונה אופיינית של הציטופלזמה של תא איקריוטי היא תנועה מתמדת ( ציקלוזיס). הוא מזוהה בעיקר על ידי תנועה של אברוני תאים, כגון כלורופלסטים. אם התנועה של הציטופלזמה נעצרת, התא מת, כי, רק להיות בפנים בתנועה מתמדת, הוא יכול לבצע את תפקידיו.
היאלופלזמה ( ציטוסול) היא תמיסה קולואידית חסרת צבע, רירית, סמיכה ושקופה. בו מתרחשים כל התהליכים המטבוליים, הוא מספק את החיבור בין הגרעין לבין כל האברונים. בהתאם לדומיננטיות של החלק הנוזלי או מולקולות גדולות בהיאלופלזמה, מבחינים בין שתי צורות של היאלופלזמה: סוֹל- היאלופלזמה נוזלית יותר ו לְהַגלִיד- היאלופלזמה צפופה יותר. מעברים הדדיים אפשריים ביניהם: הג'ל הופך לסול ולהיפך.
תפקידי הציטופלזמה:
- שילוב של כל מרכיבי התא למערכת אחת,
- סביבה למעבר של תהליכים ביוכימיים ופיזיולוגיים רבים,
- סביבה לקיומם ולתפקודם של אברונים.
קירות תאים
קירות תאיםלהגביל תאים איקריוטיים. ניתן להבחין לפחות בשתי שכבות בכל קרום תא. השכבה הפנימית צמודה לציטופלזמה ומיוצגת על ידי קרום פלזמה(מילים נרדפות - פלזמהלמה, קרום התא, קרום ציטופלזמי), שמעליו נוצרת השכבה החיצונית. IN כלוב חיותהוא דק ונקרא גליקוקליקס(נוצר על ידי גליקופרוטאין, גליקוליפידים, ליפופרוטאינים), בתא צמחי - עבה, הנקרא דופן תא(נוצר על ידי תאית).
את כל ממברנות ביולוגיותבעלי מאפיינים ומאפיינים מבניים משותפים. כרגע מקובל באופן כללי דגם פסיפס נוזלי של מבנה הממברנה. הבסיס של הממברנה הוא דו-שכבה שומנית, שנוצרת בעיקר על ידי פוספוליפידים. פוספוליפידים הם טריגליצרידים שבהם שארית חומצת שומן אחת מוחלפת בשארית חומצה זרחתית; הקטע של המולקולה שבו נמצאת שארית החומצה הזרחתית נקרא הראש ההידרופילי, הקטעים שבהם נמצאים שאריות חומצות השומן נקראים זנבות הידרופוביים. בממברנה, הפוספוליפידים מסודרים בצורה מסודרת בקפדנות: הזנבות ההידרופוביים של המולקולות פונים זה לזה, והראשים ההידרופיליים פונים החוצה, לכיוון המים.
בנוסף לליפידים, הממברנה מכילה חלבונים (בממוצע ≈ 60%). הם קובעים את רוב הפונקציות הספציפיות של הממברנה (הובלה של מולקולות מסוימות, קטליזה של תגובות, קליטה והמרת אותות מהסביבה וכו'). הבחנה: 1) חלבונים היקפיים(ממוקם על פני השטח החיצוניים או הפנימיים של דו-שכבת השומנים), 2) חלבונים חצי אינטגרליים(שקוע בדו-שכבת השומנים לעומקים שונים), 3) חלבונים אינטגרליים או טרנסממברניים(מחלחלים לממברנה דרך ודרך, תוך כדי מגע עם הסביבה החיצונית והפנימית של התא כאחד). חלבונים אינטגרליים נקראים במקרים מסוימים יוצרי תעלות, או תעלות, מכיוון שהם יכולים להיחשב כתעלות הידרופיליות שדרכן עוברות מולקולות קוטביות לתוך התא (המרכיב השומני של הממברנה לא יאפשר להם לעבור).
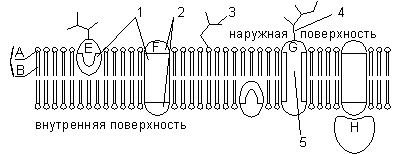
A - ראש הידרופילי של הפוספוליפיד; C, זנבות הידרופוביים של הפוספוליפיד; 1 - אזורים הידרופוביים של חלבונים E ו-F; 2, אזורים הידרופיליים של חלבון F; 3 - שרשרת אוליגוסכריד מסועפת המחוברת לליפיד במולקולת גליקוליפידית (גליקוליפידים שכיחים פחות מגליקופרוטאין); 4 - שרשרת אוליגוסכריד מסועפת המחוברת לחלבון במולקולת גליקופרוטאין; 5 - תעלה הידרופלית (מתפקדת כנקבובית שדרכה יכולים לעבור יונים וכמה מולקולות קוטביות).
הממברנה עשויה להכיל פחמימות (עד 10%). מרכיב הפחמימות של הממברנות מיוצג על ידי שרשראות אוליגוסכרידים או פוליסכרידים הקשורות למולקולות חלבון (גליקופרוטאינים) או שומנים (גליקוליפידים). בעיקרון, פחמימות ממוקמות על פני השטח החיצוניים של הממברנה. פחמימות מספקות פונקציות קולטן של הממברנה. בתאים של בעלי חיים, גליקופרוטאין יוצרים קומפלקס אפיממברנה, הגליקוקאליקס, בעובי של כמה עשרות ננומטרים. קולטני תאים רבים ממוקמים בו, בעזרתו מתרחשת הידבקות תאים.
מולקולות של חלבונים, פחמימות ושומנים ניידות, מסוגלות לנוע במישור הממברנה. עובי ממברנת הפלזמה הוא כ-7.5 ננומטר.
תפקודי ממברנה
הממברנות מבצעות את הפונקציות הבאות:
- הפרדה של תוכן סלולרי מהסביבה החיצונית,
- ויסות חילוף החומרים בין התא לסביבה,
- חלוקת התא לתאים ("תאים"),
- מיקום "מסועים אנזימטיים",
- מתן תקשורת בין תאים ברקמות אורגניזמים רב תאיים(הַדבָּקָה),
- זיהוי אות.
החשוב ביותר תכונת הממברנה- חדירות סלקטיבית, כלומר. ממברנות הן חדירות מאוד לחומרים או מולקולות מסוימות, וחדירות גרוע (או אטומות לחלוטין) לאחרים. תכונה זו עומדת בבסיס התפקוד הרגולטורי של הממברנות, המבטיח חילופי חומרים בין התא לסביבה החיצונית. התהליך שבו חומרים עוברים דרך קרום התא נקרא הובלה של חומרים. הבחנה: 1) הובלה פסיבית- תהליך העברת חומרים, הולך ללא אנרגיה; 2) מעבר פעיל- תהליך העברת חומרים, הולך עם עלות האנרגיה.
בְּ הובלה פסיביתחומרים נעים מאזור שיש בו יותר ריכוז גבוהלאזור עם נמוך יותר, כלומר. לאורך שיפוע הריכוז. בכל תמיסה יש מולקולות של הממס והמומס. תהליך התנועה של מולקולות מומסות נקרא דיפוזיה, תנועת מולקולות הממס נקראת אוסמוזה. אם המולקולה טעונה, אז ההובלה שלה מושפעת מהשיפוע החשמלי. לכן, לעתים קרובות מדברים על גרדיאנט אלקטרוכימי, המשלב את שני הגרדיאנטים יחד. מהירות התחבורה תלויה בגודל השיפוע.
ניתן להבחין הסוגים הבאיםהובלה פסיבית: 1) דיפוזיה פשוטה- הובלה של חומרים ישירות דרך דו-שכבת השומנים (חמצן, פחמן דו חמצני); 2) דיפוזיה דרך תעלות ממברנות- הובלה דרך חלבונים יוצרי תעלה (Na +, K +, Ca 2+, Cl -); 3) הקלה על דיפוזיה- הובלה של חומרים באמצעות חלבוני הובלה מיוחדים, שכל אחד מהם אחראי לתנועה של מולקולות מסוימות או קבוצות של מולקולות קשורות (גלוקוז, חומצות אמינו, נוקלאוטידים); 4) סְפִיגָה- הובלה של מולקולות מים (בסך הכל מערכות ביולוגיותהממס הוא מים).
כּוֹרַח מעבר פעילמתרחש כאשר יש צורך להבטיח העברת מולקולות דרך הממברנה כנגד השיפוע האלקטרוכימי. הובלה זו מתבצעת על ידי חלבוני נשא מיוחדים, שפעילותם דורשת הוצאה אנרגטית. מקור האנרגיה הוא מולקולות ATP. הובלה פעילה כוללת: 1) Na + /K + -משאבה (משאבת נתרן-אשלגן), 2) אנדוציטוזיס, 3) אקסוציטוזיס.
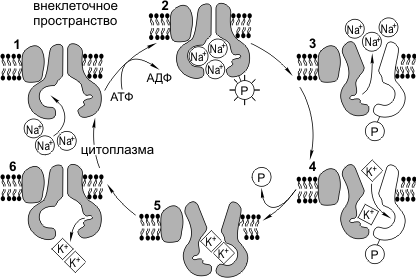
עבוד Na + /K + -משאבה. ל תפקוד רגילהתא חייב לשמור על יחס מסוים של יוני K + ו- Na + בציטופלזמה ובסביבה החיצונית. ריכוז K + בתוך התא צריך להיות גבוה משמעותית מאשר מחוצה לו, ו- Na + - להיפך. יש לציין כי Na + ו- K + יכולים להתפזר בחופשיות דרך נקבוביות הממברנה. משאבת Na+/K+ נוגדת את השוויון של ריכוזי יונים אלו ומשאבת באופן אקטיבי את Na+ מהתא ואת K+ אל התא. משאבת Na + /K + - היא חלבון טרנסממברני המסוגל לבצע שינויים קונפורמטיביים, כך שהוא יכול לחבר גם K + וגם Na +. ניתן לחלק את מחזור הפעולה של Na + /K + -משאבה לשלבים הבאים: 1) הוספת Na + עם בְּתוֹךממברנות, 2) זרחון של חלבון המשאבה, 3) שחרור Na + בחלל החוץ תאי, 4) התקשרות של K + עם בחוץממברנות, 5) דה-פוספורילציה של חלבון המשאבה, 6) שחרור K+ בחלל התוך תאי. משאבת הנתרן-אשלגן צורכת כמעט שליש מכל האנרגיה הדרושה לחיי התא. במהלך מחזור פעולה אחד, המשאבה שואבת 3Na + מהתא ומשאבת 2K +.
אנדוציטוזיס- תהליך הספיגה על ידי התא של חלקיקים גדולים ומקרומולקולות. ישנם שני סוגים של אנדוציטוזיס: 1) פגוציטוזיס- לכידה וספיגה של חלקיקים גדולים (תאים, חלקי תאים, מקרומולקולות) ו-2) פינוציטוזה- לכידה וספיגה של חומר נוזלי (תמיסה, תמיסה קולואידית, תרחיף). תופעת הפאגוציטוזיס התגלתה על ידי I.I. Mechnikov בשנת 1882. במהלך אנדוציטוזיס, קרום הפלזמה יוצר פלישה, קצוותיו מתמזגים, והמבנים התוחמים מהציטופלזמה על ידי ממברנה אחת נשרכים לתוך הציטופלזמה. פרוטוזואה רבים וכמה לויקוציטים מסוגלים לפגוציטוזיס. פינוציטוזה נצפה בתאי האפיתל של המעי, באנדותל של נימי הדם.
אקסוציטוזיס- תהליך הפוך לאנדוציטוזיס: הפרשה חומרים שוניםמהתא. במהלך האקסוציטוזיס, קרום השלפוחית מתמזג עם הממברנה הציטופלזמית החיצונית, תוכן השלפוחית מוסר מחוץ לתא, והממברנה שלו נכללת בממברנה הציטופלזמית החיצונית. בדרך זו, מתאי הבלוטות הפרשה פנימיתהורמונים מופרשים, בפרוטוזואה - שאריות לא מעוכלותמזון.
לך ל הרצאות מספר 5"תורת התא. סוגי ארגון סלולרי»
לך ל הרצאות מספר 7"תא אוקריוטי: מבנה ותפקודים של אברונים"
תוכן מופשט
2.
הציטופלזמה של תא איקריוטי
4. ממברנות ציטופלזמיות
4.1. כימיה ומאפיינים ממברנות תאים
5. קרום פלזמה
5.1. צמיחת קרום פלזמה
6.1. פונקציות תחבורה
6.2. פונקציות הקולטן של קרום הפלזמה
6.3. אנשי קשר בין תאיים
6.4. מגע פשוט. חיבור מסוג מנעול
6.6. מגע ביניים
6.7. מגע מרווח
בִּיבּלִיוֹגְרָפִיָה
1. תפקידם של מדענים בגילוי קרומי התא
עד סוף המאה ה-18 ההנחה הייתה שהתאים מופרדים בקיר משותף ולא ניתן לבודד זה מזה. בפיתוח נוסף של מדע זה, תפקיד חשוב מילאו מחקריו של הבוטנאי הצרפתי בריסו דה מירבל (1776-1854), שבהם התבטא לראשונה הרעיון של מחקר השוואתי של רקמות צמחיות. מירבל היה הראשון שהציע שתאים סמוכים מופרדים על ידי מחיצה משותפת - קרום.
חוקר הטבע הגרמני רודולפי הגיע למסקנה שתאים מבודדים, שיש להם ממברנות משלהם המקיפות אותם מכל עבר. כל ספק כי ניתן לבודד ולבודד תאים נעלמו לבסוף לאחר עבודתו של הבוטנאי I. Moldengauer (1766-1827), שממנה נובע כי התא הוא מבנה עצמאי המכוסה בקרום. אז נוצר רעיון חדש של התא.
זה נוסח בצורה הברורה ביותר מאוחר יותר על ידי פ. מיין. הגדרתו היא כדלקמן: "תא של אורגניזם צמחי הוא חלל מוקף לחלוטין בקרום וגטטיבי". פ.מיין ציין בבירור דעה זו בתחילת המאה התשע-עשרה. שהחליף את התצוגה המקורית של התא כזמן. אבל התוכן הספציפי של האנטומיה של הצמח עדיין היה שונה מהותית מהישן. החוקרים המשיכו לתאר באופן שיטתי את המבנה גופים שוניםצמחים, כמו גם לפרט ולהבהיר את הנלמד. תקופה זו נחשבת בצדק לתקופת איסוף החומר, תקופת הצטברות של מידע רב על המבנה הטוב ביותר של צמחים.
אז, עצם נוכחותם של תאים ב בדים שוניםלא העלה ספקות. אבל האם כל מגוון המבנים של אורגניזמים צמחיים מותש רק על ידי תאים? השאלה עדיין לא הייתה ברורה לגמרי. יתר על כן, רוב המדענים ענו על שאלה זו בשלילה, והצביעו, יחד עם תאים, על הכלים והסיבים של הצמחים כמבנים מוזרים למדי. לסיכום כל הידע בתחום זה, מיין כתב ישירות שרקמות הצמח מורכבות מתאי, סיבים וכלי דם. הופעתם של אלה האחרונים מתאי מטמורפוזה התבססה לבסוף רק מעט מאוחר יותר - בשנות ה-30. המאה הקודמת.
2. ציטופלזמה של תא איקריוטי
ציטופלזמההוא מנגנון העבודה המטבולי של התא. אברונים כלליים ומיוחדים מרוכזים בו, התהליכים המטבוליים העיקריים מתרחשים בציטופלזמה. בעשורים האחרונים חלה התקדמות גדולה במיוחד בניתוח מורפו-פונקציונלי דיסקרטי של אברונים ומערכות ממברנות בודדות של המנגנון המטבולי של הציטופלזמה. זה מאפשר לדמיין את עבודתו של המנגנון המטבולי כמערכת אינטגרלית.
במחקרים מודרניים של המנגנון המטבולי של התא, בולטת במיוחד הנטייה לחקור את המשמעות התפקודית של מבנים מורפולוגיים - לנתח את התפקוד באמצעות הארגון המבני והביוכימי של מבנים אלה. ההצלחות החשובות ביותר בהקשר זה הושגו בחקר האברונים העיקריים של סינתזת חלבון (ריבוזומים) ומערכות המספקות חילוף חומרים אנרגטי (מיטוכונדריה, כלורופלסטים וממברנות מצמידות של תאים פרוקריוטים). הכללה עיקרית בחקר המנגנון המטבולי של הציטופלזמה היא הרעיון של זה כמערכת תלת פאזית מובחנת פלסטית המורכבת מהציטופלזמה הראשית, ממברנות תוך תאיות ותכולת מערכת הממברנות. לפיכך, המנגנון המטבולי של הציטופלזמה, מצד אחד, מורכב עם מבנים מיוחדים המבצעים פונקציות מסוימות, ומצד שני, זוהי מערכת אינטגרלית אחת שנמצאת בקשר הדוק הן עם המנגנון פני השטח והן עם המנגנונים הגרעיניים. תָא.
גישה זו החלה בשנות ה-60. דה דוב בתורת מערכות הממברנות האקסופלזמיות. לאחר מכן, גישה זו פותחה הן בתורת הליזוזומים והן ב רעיונות עכשווייםעל מנגנון גולגי והרשת האנדופלזמית, וכן על האינטראקציה הדינמית של מערכות ממברנות תוך-תאיות. מבטיח מאוד, לדעתנו, גם הרעיון שהתגבש לאחרונה לגבי הציטופלזמה העיקרית - ההיאלופלזמה. בניגוד לתפיסה המוקדמת יותר של ההיאלופלזמה כמערכת קולואידית חיה הומוגנית, היא מפרשת את הציטופלזמה הראשית כשלב מובחן והטרוגני של הציטופלזמה המסוגל ליצור מבנים מורכבים. עם תכונה זו של ההיאלופלזמה, כבר הכרנו חלקית את הדוגמה של מערכת התת-ממברנה של מנגנון פני השטח עם המבנים המכנו-כימיים והתומכים הדינמיים שלה.
נכון לעכשיו, מתברר כי מערכות כאלה הקשורות לממברנה של היאלופלזמה נוצרות לא רק באזור ממברנות פלזמה, אבל גם באזור כל הממברנות, ולפעמים אפילו לא-ממברניות (מרכז התא) של הציטופלזמה. חלק זה של הציטופלזמה מוצע לבודד לתוך תת-מערכת מיוחדת של היאלופלזמה, המציינת אותו במונח "ציטוסול". באופן טבעי, התפקודים והארגון המורפוביוכימי של ציטוזול כזה באזור הרשת האנדופלזמית המחוספסת והחלקה (ER), הממברנות של מנגנון הגולגי, המיטוכונדריה, מרכז התא, המעטפה הגרעינית וכו' יהיו שונים וספציפיים עבור אלה. מבנים שסביבם נוצרים החלקים המתאימים של הציטוזול. .
בהתחשב במגוון המבני והתפקודי של האברונים המספקים מטבוליזם תוך תאי בציטופלזמה של תאים, התפקוד של חלקי הציטוזול התואמים לכל אורגנואיד ספציפי ונכללים למעשה בהרכבו לא יוגבל רק לתפקוד התמיכה-התכווצות . המבנים והאנזימים המרכיבים את הציטוזול הם ספציפיים לכל אברון ומעורבים ישירות בהבטחת התהליכים המתרחשים באורגנואיד זה. תהליכים ביוכימיים.
שיקול של הציטופלזמה מומלץ להתחיל בתיאור הבעיה של ארגון מבני וביוכימי של ריבוזומים - מכונות מולקולריות של סינתזת חלבון. במקרה זה, המשמעות של הארגון המבני של תהליכים ביוכימיים במערכות סלולריות באה לידי ביטוי בבירור במיוחד, והושגה הצלחה רבה במחקרו.
התחום השני של ביולוגיה של התא, שבו גם הארגון המבני של תהליכים ביוכימיים נחקר בפירוט רב, הוא הניתוח של מה שנקרא ממברנות מצומדות - מנגנון חילוף החומרים של אנרגיית התא. נשקול אותם בעת אפיון המיטוכונדריה, הפלסטידים ומבנים אחרים של הציטופלזמה האחראים על חילוף החומרים האנרגטי בתא. לאחר מכן, נדון בתוצאות של ניתוח דיסקרטי של הממברנות העיקריות.
גוף התא בפועל ותכולתו מופרדים מהסביבה החיצונית או מיסודות שכנים באורגניזמים רב-תאיים על ידי קרום פלזמה. מחוץ לממברנת הפלזמה, חוץ תאי, ממוקם דופן תא, או קיר, מתבטא היטב בצמחים ובאורגניזמים פרוקריוטיים; בתאי בעלי חיים זה נעדר או מתבטא בצורה חלשה מאוד. כל פנים התא, למעט הגרעין, נקרא ציטופלזמה. זהו מונח כללי המדגיש את חלוקת התא לשני מרכיבים עיקריים: הציטופלזמה והגרעין.
הציטופלזמה של תאים אוקריוטייםהטרוגני במבנהו ובהרכבו וכולל הילופלזמה, ממברנה ורכיבים שאינם ממברניים. מרכיבי הממברנה כוללים את המערכת הוואקואולרית (רטיקולום אנדופלזמי, מנגנון גולגי, ליזוזומים, וואקוולים צמחיים), אברוני ממברנה (מיטוכונדריה ופלסטידים). רכיבים שאינם ממברניים כוללים צנטריולים, האופייניים לתאי בעלי חיים, וקומפלקסים ומבנים מקרו-מולקולריים שאינם ממברניים, כגון ריבוזומים, מיקרוטובולים ומיקרופילמנטים. חלוקה כזו של הציטופלזמה למרכיבים נפרדים אין פירושה בידוד המבני והתפקודי שלהם זה מזה.
3. Hyaloplasm - הסביבה הפנימית של התא
המונחים "היאלופלזמה" (מ-hyaline - שקוף, שקוף), "פלזמה בסיסית", או "מטריקס ציטופלזמה" מציינים חלק חשוב מאוד בתא, סביבתו האמיתית, הפנימית. IN מיקרוסקופ אלקטרונילמטריצה הציטופלזמית יש מראה של חומר הומוגני או דק עם צפיפות אלקטרונים נמוכה. מתצפיות ישנות על התכונות הפיזיקוכימיות של הציטופלזמה, התקבל הרעיון שההיאלופלזמה היא מערכת קולואידית מורכבת הכוללת ביו-פולימרים שונים: חלבונים, חומצות גרעין, פוליסכרידים ועוד. מערכת זו מסוגלת לעבור מחומר דמוי סול (נוזל) מצב לג'ל ובחזרה. לדוגמה, בלחצים הידרוסטטיים גבוהים ציטופלזמהאינו מתעבה, אלא מתנזל באופן הפיך. תופעה זו מוסברת בשיבוש הקשרים בין מולקולות בהרכב ההיאלופלזמה.
הודות לשיטות מחקר אלקטרוניות-מיקרוסקופיות ופיזיו-כימיות, עלו רעיונות לגבי הפלזמה הראשית כמערכת מאורגנת ומסודרת מרובת רכיבים. אזורים נפרדים של היאלופלזמה יכולים לשנות את מצב הצבירה שלהם - בהתאם לתנאים או למשימה התפקודית. לפיכך, ידוע שניתן לפזר מולקולות חלבון טובולין בודדות בהיאלופלזמה, אך ברגעים מסוימים הן מתחילות להרכיב ולבנות מבנים צינוריים ארוכים - מיקרוטובולים.
תהליך זה של הרכבה עצמית של מיקרוטובולים הוא הפיך: כאשר תנאי החיים של התא משתנים (עלייה בלחץ או שינוי בחדירות קרומי התא), המיקרוטובוליות מתפרקות למולקולות טובולין מונומריות. באותו אופן, בהיאלופלזמה חסרת מבנה לכאורה, יכולים להתעורר ולהתפורר קומפלקסים פיבריליים וחוטים שונים של מולקולות חלבון. הפלזמה העיקרית יכולה לקחת חלק ביצירת קרומי תאים, חוטים ומיקרופילמנטים. באמבות, השכבות החיצוניות של הציטופלזמה (אקטופלזמה) או אזור הציטופלזמה באזור הפסאודופודיה מורכבות למעשה מהיאלופלזמה אחת, ממברנות, מיטוכונדריה ומבנים אחרים נמצאים שם רק לעתים רחוקות. עם זאת, לאחר חשיפה ליונים המעודדים נפיחות תאים, מתרחשים סידורים מחדש מהירים ופתאומיים של מקטעים אלו של הציטופלזמה - מופיעים בהם ואקוולים קטנים של ממברנה. לכן, הפלזמה הראשית חייבת להכיל את כל אבני הבניין (ליפידים וחלבונים) הנחוצים ליצירת ממברנות.
מבנים ציטופלזמה פיברילרית יכולים להיווצר גם כתוצאה מתהליכי צבירה ספציפיים בהיאלופלזמה. אם גרעינים גדולים, מבני ממברנה ואז ריבוזומים משוקעים מהומוגניות של תאים, אזי הסופרנטנט שנותר יכיל את המרכיבים הכימיים העיקריים של ההיאלופלזמה. הרכב ההיאלופלזמה של מיקרומולקולות כולל בעיקר חלבונים כדוריים שונים ואנזימים של המטריצה הציטופלזמית. הם מהווים 20-25% תוכן כללי חלבונים בתא איקריוטי.
IN תאים חיידקיים, דלים ביסודות ממברנה, חלבונים היאלופלסמים יכולים להוות כ-50% מכלל החלבונים. אנזימי המטריצה החשובים ביותר כוללים אנזימי גליקוליזה, אנזימים לחילוף חומרים של סוכרים, בסיסים חנקן, חומצות אמינו, שומנים ותרכובות חשובות אחרות. המטריצה מכילה אנזימים להפעלת חומצות אמינו במהלך סינתזת חלבון, העברה RNA. המאפיינים האוסמוטיים והמאגרים של התא נקבעים במידה רבה על ידי הרכב ומבנה ההיאלופלזמה.
התפקיד החשוב ביותר של ההיאלופלזמה הוא שהמדיום החצי-נוזלי הזה מאחד את כל המבנים התאיים ומבטיח את האינטראקציה הכימית שלהם זה עם זה. רוב תהליכי ההובלה התוך תאית מתבצעים דרך ההיאלופלזמה: העברת חומצות אמינו, חומצות שומן, נוקלאוטידים וסוכרים. בהיאלופלזמה קיימת זרימה קבועה של יונים אל וממברנת הפלזמה, אל המיטוכונדריה, הגרעין וה-vacuoles. ההיאלופלזמה היא כלי הקיבול ואזור התנועה העיקריים של מסת מולקולות ה-ATP. בו מתרחשת שקיעה של מוצרי חילוף: גליקוגן, טיפות שומן. מבחינה מבנית ומורפולוגית, ההיאלופלסמה עדיין לא מובנת.
4. ממברנות ציטופלזמיות
תכונה משותפת של כל ממברנות התא, קרום הפלזמה החיצוני וכל הממברנות התוך-תאיות ואברוני הממברנה היא שהם שכבות דקות (6-10 ננומטר) בעלות אופי ליפופרוטאין (ליפידים בקומפלקס עם חלבונים), סגורות על עצמן. אין ממברנות פתוחות עם קצוות חופשיים בתא. קרומי תאים תמיד מגבילים חללים או אזורים, סוגרים אותם מכל עבר ובכך מפרידים את התוכן של חללים כאלה מסביבתם. אז, קרום הפלזמה, המכסה את כל פני התא, בעל צורה מורכבת וצמחים רבים, אינו מופרע בשום מקום, הוא סגור. הוא מפריד את תוכן הציטופלזמה מהסביבה המקיפה את התא.
ממברנות סגורות תוך תאיות יוצרות שלפוחיות - ואקואולות בעלות צורה כדורית או שטוחה. במקרה האחרון, שקיות קרום שטוח, או טנקים, נוצרים. לעתים קרובות, לחללים המוגדרים על ידי ממברנות יש צורה מורכבת הדומה לספוג או לרשת, אך במקרה זה, חללים כאלה מתוחמים ללא הפרעות על ידי ממברנה. בווריאציות כאלה, הממברנות מפרידות גם בין שני שלבים מבניים של הציטופלזמה: היאלופלזמה מהתוכן של ואקוולים ובורות מים. לממברנות המיטוכונדריה והפלסטידים יש את אותה תכונה: הם מפרידים את התוכן הפנימי מהחללים הבין-ממברניים ומההיאלופלזמה.
הממברנה הגרעינית יכולה להיות מיוצגת גם כשקית ממברנה כפולה מחוררת בעלת צורה כדורית. הממברנות של המעטפת הגרעינית תוחמות, מפרידות את הקריופלזמה והכרומוזומים זה מזה מחלל החלל הפרי-גרעיני ומההיאלופלזמה. תכונות מורפולוגיות כלליות אלו של ממברנות התא נקבעות על פי שלהן תרכובת כימית, אופי הליפופרוטאין שלהם.
4.1. כימיה ומאפיינים של ממברנות התא
רָאשִׁי רכיבים כימייםקרומי התא הם שומנים (40%) וחלבונים (60%), בנוסף, פחמימות נמצאות בממברנות רבות. ליפידים הם קבוצה גדולה חומר אורגניבעל מסיסות ירודה במים (הידרופוביות) ומסיסות בממיסים ושומנים אורגניים (ליפופיליות).
הרכב השומנים בממברנות התא מגוון מאוד. נציגים אופייניים של שומנים הנמצאים בממברנות התא הם פוספוליפידים, ספינגומיליינים, ומהשומנים הסטרואידים, כולסטרול. גליצרוליפידים הם אסטרים של אלכוהול תלת-הידרי, גליצרול עם שתי חומצות שומן וחומצה זרחתית, אשר, בתורה, יכולים להיות קשורים לקבוצות כימיות שונות (כולין, סרין, אינוזיטול, אתנולמין וכו').
קבוצה נוספת של שומני ממברנה הם ספינגומיליינים, כאשר הגליצרול מוחלף באלכוהול האמינו ספינגוזין. מבין השומנים הקשורים לסטרואידים, יותר מכל בקרומי הכולסטרול. IN תאי צמחיםכולסטרול לא נמצא, הוא מוחלף בפיטסטרולים. לחיידקים אין סטרולים. תכונה אופייניתשומני הממברנה היא חלוקת המולקולות שלהם לשני חלקים שונים בתפקוד: זנבות לא קוטביים שאינם נושאים מטענים ומורכבים מחומצות שומן, וראשים קוטביים טעונים. ראשי קוטב נושאים מטענים שליליים או יכולים להיות ניטרליים (אם יש להם מטענים חיוביים ושליליים כאחד).
נמצא שממברנות התא שונות מאוד זו מזו מבחינת הרכב השומנים. לפיכך, ממברנות הפלזמה של תאי בעלי חיים עשירות בכולסטרול (עד 30%), יש בהן מעט לציטין, ואילו ממברנות מיטוכונדריה, להיפך, עשירות בפוספוליפידים ועניות בכולסטרול. מ סה"כתכולת השומנים של לציטין בשברים של הרשת האנדופלזמית היא 60-70% מכלל הפוספוליפידים, בעוד שבממברנת הפלזמה זה יכול להיות 25-35%.
באופן כללי, קרום הפלזמה מאופיין תוכן גבוהכולסטרול וספינגוליפידים, כמו גם הדומיננטיות של חומצות שומן רוויות וחד בלתי רוויות בהרכב הפוספוליפידים, בעוד שהמיטוכונדריה, הרשת האנדופלזמית וממברנות ציטופלזמיות רבות אחרות מכילות מעט כולסטרול וספינגוליפידים והרבה יחסית חומצות שומן רב בלתי רוויות. ככל הנראה, בקשר לכך, הממברנות הציטופלזמיות פחות נוקשות ממברנת הפלזמה, הן מתמזגות יותר.
גם כמות החלבונים בממברנות התא שונה. אז במיטוכונדריה, בהשוואה לממברנות אחרות, יש הכי הרבה חלבונים. הממברנות שונות מאוד במערך מולקולות החלבון. חלקם קשורים לראשי שומנים באמצעות קשרים יוניים ולכן מופקים בקלות ממברנות עם תמיסות מלח. אחרים יוצרים קשרי מלח עם האזורים הקוטביים של שומנים באמצעות אינטראקציה עם יוני Mg2+ או Ca2+. חלבונים מופקים גם באמצעות תרכובות קלאטיות כגון ורסן (EDTA). רוב החלבונים מקיימים אינטראקציה עם שומנים בהרכב הממברנות המבוססות על קשרים הידרופוביים.
נודע כי נראה כי חלבוני ממברנה רבים מורכבים משני חלקים: מאזורים עשירים בחומצות אמינו קוטביות (נושאות מטען), ואזורים המועשרים בחומצות אמינו לא קוטביות (גליצין, אלנין, ואליון, לאוצין). חלבונים כאלה בשכבות השומנים של הממברנות ממוקמים בצורה כזו שהאזורים הלא קוטביים שלהם שקועים, כביכול, בחלק ה"שומן" של הממברנה, שם נמצאים האזורים ההידרופוביים של השומנים. החלק הקוטבי (הידרופילי) של חלבונים כאלה יוצר אינטראקציה עם ראשי השומנים ומופנה לכיוון הפאזה המימית. לכן, חלבונים כאלה הקשורים לליפידים על ידי אינטראקציות הידרופוביות כמעט ואינם מופקים בשלבים מימיים. ניתן לבודד אותם רק על ידי הרס הממברנה, מיצוי שומנים ממנו בעזרת ממיסים או דטרגנטים אורגניים.
על ידי תפקיד ביולוגיניתן לחלק את חלבוני הממברנה לשלוש קבוצות: אנזימים, חלבוני קולטן וחלבונים מבניים. מערך האנזימים בממברנות יכול להיות גדול מאוד ומגוון (לדוגמה, לפחות 24 אנזימים שונים נמצאו בקרום הפלזמה של תאי הכבד).
לממברנות שונות יש קבוצה אופיינית של אנזימים. במילים אחרות, ההטרוגניות הכימית של ממברנות נקבעת לא רק על ידי שומנים, אלא גם על ידי חלבונים. המשותף לכל ממברנות התא הוא מבנה הליפופרוטאין שלהם; השוני טמון בהבדלים הכמותיים והאיכותיים בשומנים ובליפידים הרכב חלבון. לדוגמה, הרכב וכמות השומנים והחלבונים יהיו שונים בתכלית בקרומים של הרשת האנדופלזמית ובממברנה הפנימית של המיטוכונדריה.
מרכיב הפחמימות של הממברנות מיוצג בעיקר על ידי גליקופרוטאין - מולקולות חלבון באופן קוולנטי (בניגוד לנוקלאופרוטאין וליפופרוטאינים) הקשורות לשרשראות פחמימות.
4.2. מבנה ממברנות התא
הרעיון שממברנות תאים מבוססות על שכבת שומנים כפולה התקבל עוד בשנות ה-20. בשנת 1925 פורסמה עבודה שהצביעה על היווצרות שכבה ביליפידית רציפה של קרום הפלזמה. למרות מספר שגיאות, הרעיון הכללי של קיומה של שכבת ביליפיד התברר כנכון. התברר שאם שומנים מופקים מממברנת האריתרוציטים ואז מניחים אותם על פני המיניסקוס של המים, אזי ניתן לחשב את השטח שתפוס על ידי השכבה החד-שכבתית המתקבלת של השומנים. התברר ששטח זה הוא פי שניים מהשטח שנכבש על ידי פני השטח של אריתרוציטים, מהם שומנים שומנים.
מדענים הציעו שבממברנות של אריתרוציטים, שומנים מסודרים בשתי שכבות. בנוסף, התברר כי מתח הפנים של התא נמוך משמעותית ממתח הפנים של שכבת השומנים המלאכותית. יתרה מכך, החוקרים מצאו שכאשר מוסיפים חלבון לשמן, מתח הפנים מופחת לערך המאפיין את מתח הפנים של התאים. בהתבסס על זה, הסיק כי הממברנה מכילה חלבון.
כך עלתה השערה לגבי מבנה הממברנה, לפיה מדובר ב"סנדוויץ'" תלת שכבתי (חלבון-ליפיד-חלבון). שכבת השומנים במרכז היא שכבה דו-מולקולרית שבה הקצוות ההידרופוביים של מולקולות השומנים מכוונים זה לזה, והקצוות ההידרופיליים הם כלפי שכבות החלבון המונחות משני צידי שכבת השומנים. השערה זו אוששה ישירות כאשר ניתן היה להשתמש במיקרוסקופ אלקטרונים כדי לחקור את ממברנות התא. בזכותה אפשר היה לראות טוב במיוחד על ממברנות מעטפת המיאלין של העצבים שהממברנה היא באמת תלת-שכבתית: שתי שכבות כהות של 2.5 ננומטר כל אחת ושכבה בהירה רחבה יותר ביניהן.
ניתן להראות את המבנה התלת-שכבתי של ממברנות התא במיקרוסקופ אלקטרוני בשיטת פיצול-הקפאה, שהיא חשובה במיוחד, מכיוון שבמקרה זה ניתן לצפות באנזימים ספציפיים או אפילו במערכות אנזימים. בתחילת שנות ה-60. רוברטסון ניסח את ההשערה של ממברנה ביולוגית "אלמנטרית" יחידה ויחידה. שאלת המקור, היווצרות ממברנות בתא חי עדיין לא נחקרה מספיק. ידוע שמספר וגודלם של יסודות הממברנה בתאים יכולים להגדיל ולהקטין, כלומר, ניתן לדבר על ממברנוגנזה והרס של ממברנות.
5. קרום פלזמה
ממברנת הפלזמה, או פלזמהלמה, תופסת מקום מיוחד בין ממברנות תאים שונות. זהו מבנה היקפי שטחי המגביל את התא מבחוץ, מה שקובע את הקשר הישיר שלו עם הסביבה החוץ-תאית, וכתוצאה מכך עם כל החומרים והגירויים הפועלים על התא. מבחינה כימית, קרום הפלזמה הוא קומפלקס ליפופרוטאין.
העקרונות העיקריים של ארגון הממברנה כבר נדונו בסעיף הקודם. עוביו כ-10 ננומטר והוא העבה ביותר מבין קרומי התא. המרכיבים העיקריים של ממברנות הפלזמה הם שומנים (כ-40%), חלבונים (יותר מ-60%) ופחמימות (כ-1%). כפי שכבר הוזכר, קרום הפלזמה עשיר יותר בכולסטרול בהשוואה לממברנות אחרות, וחומצות שומן רוויות שולטות בפוספוליפידים שלה.
הרכב החלבונים המרכיבים את קרום הפלזמה הוא מגוון מאוד, אשר נקבע על ידי מגוון העומסים התפקודיים שלו. בין האנזימים נמצאו 5 "- נוקלאוטידאז, ATPase תלוי Mg המופעל על ידי יוני Na ו-K, פוספטאז אלקליין וחומצי, אדנילט ציקלאז, RNase, פעילות פרוטאוליטית. אנזימים של שרשרת הנשימה ואנזימים גליקוליטים נעדרים לחלוטין בממברנת הפלזמה. של תאים אוקריוטים.אבל בתאים פרוקריוטיים זה בהרכב של קרום הפלזמה כי האלמנטים של שרשרת הובלה אלקטרונים וזרחון חמצוני הם מקומיים.
יש להדגיש כי ייתכן שחלק מהחלבונים המצויים בשברים של קרום הפלזמה אינם קשורים ישירות לשכבת הביליפיד. אנזימים רבים הפעילים בחלקים של קרום הפלזמה יכולים להיות מקומיים ל-glycocalyx, קומפלקס גליקופרוטאין הקשור לממברנת הפלזמה. הרכב הגליקוקליקס כולל פחמימות שונות, שיכולות להוות כ-1% מהמשקל היבש של הממברנות.
פחמימות יוצרות שרשראות ארוכות ומסועפות של פוליסכרידים המחוברות לשכבת חלבונים המגדירה לרוב את צורת התא. שקי ממברנה יכולים לצמוד לממברנת הפלזמה כאן מבפנים; במקרה זה, ישנן שלוש שכבות ממברנות ליד פני התאים: קרום הפלזמה עצמו ושתי ממברנות של alveoli pellicular.
בסיליאטים של הנעל, הגלעין יוצר עיבויים, הממוקמים בצורה של משושים, שבמרכזם יוצאות ריסים. הקשיחות של תצורות קליקולריות יכולה להיות קשורה גם לאלמנטים של הציטופלזמה הנמצאת בבסיס קרום הפלזמה, עם השכבה הקורטיקלית. כך, בפסגות של euglena pellicle ליד הממברנה, בנוסף ל-vacuoles של הממברנה, נמצאות צרורות מקבילות של מיקרו-צינוריות ומיקרו-פילמנטים. חיזוק היקפי פיברילרי זה, יחד עם היקפי הממברנה הרב-שכבתית המקופלת, יוצר מבנה פלקל קשיח.
5.1. צמיחת קרום פלזמה
לאחר חלוקת התא, יש עלייה בנפח תאי הבת הגדלים, ובקשר לכך, יש עלייה בשטח התא, עלייה בשטח קרום הפלזמה. אבל זו לא הדוגמה היחידה. גידול מהירנפח ומשטח. פני השטח של תאים שגדלים במהירות בחוטי האבקנים של דגנים יכולים לגדול פי 65 בשעה, כלומר, בכל דקה שהפלזמה גדלה בערך המקורי שלה. קצב גדילה כה גבוה של קרום הפלזמה יכול להיות מוסבר רק על ידי שילוב מהיר של מבשרי ממברנה מוכנים, שומנים וחלבונים, בשכבת הליפופרוטאין הישנה. דרך דומהצמיחה אפשרית בהחלט, שכן ידוע שבתא מתרחש סידור מחדש מתמיד של ממברנות - החלפת שומנים מסוימים באחרים, שינוי בכמותם, שינוי בהרכב החלבונים וכו'. לכן, ממברנות מאוד לאביליות. תרכובות.
אבל תואר גם מקרה נוסף של היווצרות קרום פלזמה חדש. הפלסמודיום של ה-Myxomycete Physarum polycephalum נקב בנימי זכוכית, וגליל הפלזמה שנוצר, נטול קרום הפלזמה, תוקן מיד ולאחר מכן לאחר 1, 2, 3 ו-6 שניות לאחר תחילת הניסוי, ולאחר מכן נחקר. תחת מיקרוסקופ אלקטרונים. הציטופלזמה של פטרייה זו מאופיינת בנוכחות של מספר רב של שלפוחיות קטנות המכילות ריר, המכסה את הפלסמודיום כולו מבחוץ. ניתן היה לראות כי שלפוחיות כאלה, המפוזרות בפלזמה הראשית, נאספו בקצה הבלתי מוגן של הפלזמה, ויצרו ואקואולים שטוחים, הממוקמים במקביל לפני השטח החיצוני של החלק החשוף. ואקואולים שטוחים כאלה התמזגו ויצרו ממברנה רציפה - קרום פלזמה חדש המבודד רכיבי פלזמה מהסביבה החיצונית.
ככל הנראה, תהליך כזה של חידוש של קרום הפלזמה מתרחש לא רק כאשר הוא ניזוק קשות, אלא גם כל הזמן, במהלך חיי התא. אז, כתוצאה מאקסוציטוזיס, נראה כי ממברנות ה-vacuole, המתמזגות עם קרום הפלזמה, מוטמעות בה, ובכך מגדילות את שטח הפנים הכולל של התא. תצפיות על תהליך ההפרשה (אחת מצורות האקסוציטוזיס) הובילו חוקרים רבים לרעיון שיש זרימות קבועות של יסודות ממברנה בתא: מקרום הפלזמה לתוך הציטופלזמה (אנדוציטוזה) ולהפך, זרימה של מבני ממברנה. מהציטופלזמה אל פני התא (אקסוציטוזיס). במחזור זה של ממברנות, התפקיד המוביל מוקצה למערכת ה-vacuoles של הממברנה של מנגנון גולגי.
6. פונקציות של קרום הפלזמה
קרום הפלזמה כן שורה שלמההתפקודים התאיים החשובים ביותר, המובילים שבהם הם הפונקציה של תיחום חומרי הציטופלזמה מהסביבה החיצונית ותפקוד הובלת חומרים שונים הן לתוך התא והן מחוצה לו. פונקציות ההובלה כוללות הובלה פסיבית של מים, יונים, חומרים בעלי משקל מולקולרי נמוך והובלה אקטיבית של חומרים אלו כנגד שיפוע ריכוז, וכן צורות שונות של הובלה של תרכובות וקומפלקסים במשקל מולקולרי גבוה (אנדוציטוזה).
קרום הפלזמה, בנוסף, מעורב בסילוק מהתאים של התוצרים הנוצרים בה. הוא גם משתתף בתהליכי המחשוף החוץ תאי של ביופולימרים. על פני ממברנת הפלזמה ישנם מבנים שונים של קולטנים המקיימים אינטראקציה ספציפית עם גורמים תאיים ועם תאים שכנים. לפיכך, קרום התא מעורב בהעברת אותות לתא. קרום הפלזמה לוקח חלק באינטראקציות בין-תאיות באורגניזמים רב-תאיים. מקטעים נפרדים של קרום הפלזמה בתאי בעלי חיים מיוחדים מעורבים בבניית תהליכים מיוחדים של תאים, כגון מיקרוווילי, ריסים, תולדות קולטן וכו'. לבסוף, קרום הפלזמה ממלא תפקיד חשוב בחלוקת התא.
ספירה קצרה זו של תפקידי קרום הפלזמה רחוקה מלהיות מלאה, אך היא מראה את החשיבות המופלגת של מבנה היקפי זה של כל תא, בין אם מדובר בתא של חיידקים או באורגניזמים אוקריוטיים.
6.1. פונקציות תחבורה
קרום הפלזמה, כמו ממברנות ליפופרוטאין תאיות אחרות, חדיר למחצה. המשמעות היא שמולקולות שונות עוברות דרכו במהירויות שונות, וככל שגודלן של המולקולות גדול יותר, כך מהירות המעבר שלהן דרך הממברנה נמוכה יותר. תכונה זו מגדירה את קרום הפלזמה כמחסום אוסמוטי.
למים וגזים המומסים בו יש את יכולת החדירה המקסימלית, יונים חודרים לממברנה הרבה יותר לאט (כפי 104 לאט יותר). לכן, אם תא, למשל, אריתרוציט, ממוקם בסביבה שבה ריכוז המלחים נמוך יותר מאשר בתא (יתר לחץ דם), אז מים מבחוץ יזרזו לתוך התא, מה שיוביל לעלייה של נפחו ועד לקרע של קרום הפלזמה ("הלם" היפוטוני).
להיפך, כאשר מניחים אריתרוציט בתמיסות מלח בריכוז גבוה יותר מאשר בתא, ייצאו ממנו מים לתוך סביבה חיצונית. במקביל, התא יתקמט, יקטן בנפח. הובלה פסיבית כזו של מים אל מחוץ לתא ואל התא עדיין מתנהלת בקצב נמוך. כתוצאה מכך, בקרום התא, בשכבת הליפופרוטאין שלו, יש נקבוביות מיוחדות לחדירת מים ויונים.
6.2. פונקציות הקולטן של קרום הפלזמה
פונקציות אלה קשורות ללוקליזציה על קרום הפלזמה של מבנים מיוחדים המכוונים להכרה ספציפית של גורמים כימיים או פיזיקליים. לפני השטח של התא יש קבוצה גדולה של רכיבים - קולטנים הקובעים אפשרות לתגובות ספציפיות עם סוכנים שונים. חלבוני ממברנה או אלמנטים גליקוקליקסים (פוליסכרידים, גליקופרוטאין) יכולים לפעול כקולטנים כאלה על פני התא.
הוא האמין שאתרים כאלה הרגישים לחומרים בודדים יכולים להיות מפוזרים על פני התא או לאסוף באזורים קטנים. לדוגמה, על פני השטח של תאי חיידקים או בעלי חיים, יש מספר מוגבל של אתרים שחלקיקים ויראליים יכולים להיקשר אליהם. מעניין שנגיפים שונים נקשרים בדרך כלל לחלקים שונים של פריפריית התא, ולאותו תא חיידקי יכולים להיות כמה סוגים שוניםקולטנים.
השימוש בשיטות אימונולוגיות שונות הראה כי אנטיגנית מקומית על פני התא רכיבים פעיליםשמגיבים ספציפית עם נוגדנים או תאי חיסון. רוב האנטיגנים המשטחים הללו מכילים קבוצות פחמימות (גליקוליפידים, גליקופרוטאינים). המרכיבים האנטיגנים של דופן החיידק הם גם לרוב ליפופוליסכרידים וגליקוליפידים. על פני התא, לרוב ממש בממברנת הליפופרוטאין שלו, ישנם קולטנים שונים הנקשרים פיזיולוגית חומרים פעיליםכמו הורמונים שונים, נוירוטרנסמיטורים וכו'.
תאים שוניםלאורגניזמים של בעלי חיים עשויים להיות סטים שונים של קולטנים או רגישות שונה של אותו קולטן. תפקידם של קולטני תאים רבים הוא להעביר אותות מהמשטח אל התא. נכון להיום, מערכת העברת הגירוי לתאים בעזרת הורמונים מסוימים, הכוללים שרשראות פפטידים, נחקרה היטב. הורמונים אלו נמצאו נקשרים לקולטנים ספציפיים על פני קרום הפלזמה של התא.
הגיוון והספציפיות של קבוצות הקולטנים על פני התאים מובילים ליצירת מערכת סמנים מורכבת מאוד המאפשרת להבחין בין התאים של האדם עצמו (מאותו פרט או מאותו מין) מאלה של אחרים. תאים דומים נכנסים לאינטראקציות זה עם זה, מה שמוביל להידבקות של משטחים (צימוד בפרוטוזואה ובחיידקים, היווצרות קומפלקסים של תאי רקמה). במקרה זה, תאים הנבדלים במערך הסמנים הקובעים או שאינם תופסים אותם אינם נכללים מאינטראקציה כזו, או נהרסים בבעלי חיים גבוהים יותר כתוצאה מתגובות אימונולוגיות.
קרום הפלזמה קשור ללוקליזציה של קולטנים ספציפיים המגיבים אליהם גורמים פיזיים. אז, בממברנת הפלזמה או בנגזרות שלה בחיידקים פוטוסינתטיים ובאצות כחולות-ירוקות, חלבוני קולטן (כלורופילים) המקיימים אינטראקציה עם קוונטות האור הם מקומיים. בממברנת הפלזמה של תאי בעלי חיים רגישים לאור, קיימת מערכת מיוחדת של חלבוני פוטורצפטורים (רודופסין), שבעזרתם הופך אות האור לכימי, אשר בתורו מוביל ליצירת חומר חשמלי. דַחַף.
6.3. אנשי קשר בין תאיים
קרום הפלזמה, כפי שכבר הוזכר, מעורב באופן פעיל במגעים בין-תאיים הקשורים לצימוד. אורגניזמים חד תאיים. באורגניזמים רב-תאיים, עקב אינטראקציות בין-תאיות, נוצרים הרכבים תאיים מורכבים, שניתן לבצע את תחזוקתם דרכים שונות. ברקמות נבטיות, עובריות, במיוחד על שלבים מוקדמיםהתפתחות, תאים נשארים מחוברים זה לזה בגלל היכולת של המשטחים שלהם להיצמד זה לזה. מאפיין זה של חיבור תאים יכול להיקבע על ידי המאפיינים של פני השטח שלהם, המקיימים אינטראקציה ספציפית זה עם זה.
המנגנון של קשרים אלה עדיין לא נחקר מספיק, אבל, ככל הנראה, הוא מסופק על ידי האינטראקציה בין ליפופרוטאינים לבין הגליקוקאליקס של ממברנות הפלזמה. עם אינטראקציה בין-תאית כזו של תאים עובריים בין ממברנות פלזמה, תמיד נשאר פער ברוחב של כ-20 ננומטר, מלא ב-glycocalyx. טיפול ברקמה באמצעות אנזימים המפרים את שלמות הגליקוקאליקס (ריריות הפועלות בצורה הידרוליטית על מוצינים, מוקופוליסכרידים) או פגיעה בממברנת הפלזמה (פרוטאזות) מובילים לבידוד התאים זה מזה, להתנתקותם. עם זאת, אם גורם הדיסוציאציה מוסר, התאים יכולים להרכיב מחדש ולהצטבר מחדש.
אז אפשר לנתק תאים של ספוגים בצבעים שונים - כתום וצהוב. התברר כי בתערובת התאים הללו נוצרים שני סוגי אגרגטים: אלו המורכבים מצהובים בלבד ורק מתאי כתומים. במקרה זה, השעיות תאים מעורבות מתארגנות בעצמן, ומשחזרות את המבנה הרב-תאי המקורי. תוצאות דומות התקבלו עם השעיות תאים מופרדות של עוברי דו-חיים; במקרה זה, קיימת הפרדה מרחבית סלקטיבית של תאי אקטודרם מהאנדודרם ומהמזנכימה.
יתר על כן, אם משתמשים ברקמות לצבירה מחדש שלבים מאוחריםהתפתחות של עוברים, ואז הרכבים שונים של תאים עם סגוליות רקמה ואיברים מורכבים באופן עצמאי במבחנה, נוצרים אגרגטים אפיתליליים הדומים לאבוביות הכליות וכו'. קשרים בין תאים ברקמות ובאיברים של אורגניזמים בעלי חיים רב-תאיים יכולים להיווצר על ידי מורכבות מיוחדת. מבנים שקראו למגעים בין-תאיים תקינים. המגעים הבין-תאיים המובנים הללו בולטים במיוחד ברקמות הגבול האינטגמנטרי, באפיתל.
ייתכן שהבידוד הראשוני של שכבת תאים המחוברים זה לזה בעזרת מגעים בין-תאיים מובנים מיוחדים הבטיח היווצרות והתפתחות של רקמות ואיברים בפילוגנזה של בעלי חיים. הודות למיקרוסקופ אלקטרוני, הצטברו הרבה נתונים על מבנה האולטרה של תצורות חיבור אלה. למרבה הצער, שלהם הרכב ביוכימיוהמבנה המולקולרי עדיין לא מובנים היטב.
על ידי לימוד הקשרים של תאים בשכבות אפיתל, ניתן למצוא את המבנים הבאים המחברים תאים זה לזה: מגע פשוט, חיבור מסוג "נעילה", מגע הדוק, מגע ביניים, או אזור הידבקות, מגע דסמוזומלי. , מגע דמוי חריץ.
6.4. מגע פשוט. חיבור מסוג מנעול
מגע פשוט נמצא בין רוב התאים ממקורות שונים הסמוכים זה לזה. רוב פני השטח של תאי האפיתל המגעים מחוברים גם הם במגע פשוט, כאשר קרומי הפלזמה של התאים המתקשרים מופרדים במרווח של 15-20 ננומטר. כפי שכבר הוזכר, חלל זה מייצג את הרכיבים העל-ממברניים של משטחי התא. רוחב הפער בין ממברנות התא יכול להיות יותר מ-20 ננומטר, ויוצרים הרחבות, חללים, אך לא פחות מ-10 ננומטר. מהצד של הציטופלזמה, אין מבנים נוספים מיוחדים סמוכים לאזור זה של קרום הפלזמה.
חיבור מסוג "נעילה" הוא בליטה של קרום הפלזמה של תא אחד לתוך אינטוסוסספציה (בליטה) של אחר. על החתך, סוג זה של חיבור דומה לתפר של נגר. למרחב הבין-ממברני ולציטופלזמה באזור ה"טירות" יש את אותם מאפיינים כמו באזורים של מגע פשוט.
6.5. קשר סגירה הדוק
זהו אזור שבו השכבות החיצוניות של שני ממברנות הפלזמה קרובות ככל האפשר. הממברנה התלת-שכבתית נראית לעתים קרובות במגע זה: שתי השכבות האוסמופיליות החיצוניות של שתי הממברנות מתמזגות לשכבה משותפת בעובי 2-3 ננומטר. היתוך ממברנה אינו מתרחש על פני כל אזור המגע ההדוק, אלא הוא סדרה של חיבורים נקודתיים. מהצד של הציטופלזמה באזור זה, נמצאים לעתים קרובות פיברילים רבים בקוטר של כ-8 ננומטר, הממוקמים במקביל לפני השטח של הפלזמה.
מגעים מסוג זה נמצאו בין פיברובלסטים בתרבית רקמה, בין אפיתל עוברי ותאי מזנכימליים.
6.6. מגע ביניים (או אזור הידבקות)
במקום זה, המרחק הבין-ממברני מורחב במקצת (עד 25-30 ננומטר) ובניגוד למגע פשוט, הוא מלא בתוכן צפוף, ככל הנראה בעל אופי חלבוני.
6.7. מגע מרווח
זהו אזור של 0.5-3 מיקרומטר באורך, שבו ממברנות הפלזמה מופרדות בפער של 2-3 ננומטר, מה שאחרי האוסמציה מעניק למבנה כולו מראה של שבע שכבות. מהצד של הציטופלזמה, לא מוצאים מבני ממברנה מיוחדים. סוג זה של חיבור נמצא בכל סוגי הרקמות. נראה שהתפקיד התפקודי של צומת הפער הוא בהעברת יונים ומולקולות מתא לתא. לדוגמה, בשריר הלב, העברת פוטנציאל הפעולה מתא לתא מתרחשת באמצעות סוג של מגע שבו יונים יכולים לעבור בחופשיות דרך צמתים בין-תאיים. שמירה על קשר יוני זה בין תאים תלויה באנרגיה המתקבלת באמצעות זרחון חמצוני.
בִּיבּלִיוֹגְרָפִיָה
1. ורמל ע"מ תולדות תורת התא. מ.: נאוקה, 1970. 259 עמ'.
2. Gulyaev G. V., Malchenko V. V. מילון מונחים בגנטיקה, ציטולוגיה, גידול, ייצור זרעים ומדעי הזרעים. מ.: Rosselkhozizdat, 1983. 240 עמ'.
3. De Duve K. מסע לעולם של תא חי. מ.: מיר, 1987. 253 עמ'.
4. Zavarzin A. A., Kharazova A. D. יסודות הציטולוגיה הכללית: ספר לימוד. ל.: בית ההוצאה לנינגרד. un-ta, 1982. 240 עמ'.
5. ממולקולות לאדם. / מתחת. ed. Naumova N.P.M.: Education, 1973. 480 p.
6. פאושבע ז.פ. סדנה בנושא ציטולוגיה של צמחים. M.: Agropromizdat, 1988. 271 עמ'.
7. Troshin A. S., Braun A. D., Vakhtin Yu. B., Zhilkin L. N., Sukhanova K. M. Cytology. מוסקבה: חינוך, 1970. 304 עמ'.
8. Chentsov Yu. S. כללי ציטולוגיה. מ.: MGU, 1978. 344 עמ'.
מופשט פלוס
פלזמהלמה(דופן התא) של תאי בעלי חיים נוצר על ידי קרום המכוסה מבחוץ בשכבת גליקוקליקס בעובי 10-20 ננומטר. פלזמהלמהמבצע פונקציות תוחמות, מחסום, הובלה וקולטן. בשל המאפיין של חדירות סלקטיבית, הפלזמהלמה מסדירה את ההרכב הכימי של הסביבה הפנימית של התא. הפלזמלמה מכילה מולקולות קולטן המזהות באופן סלקטיבי חומרים פעילים ביולוגית (הורמונים). בשכבות ושכבות, תאים שכנים נשמרים עקב נוכחותם של סוג אחראנשי קשר, המיוצגים על ידי קטעים של הפלזמה בעלי מבנה מיוחד. מבפנים, השכבה הקורטיקלית (קורטיקלית) צמודה לממברנה ציטופלזמהעובי 0.1-0.5 מיקרומטר.
ציטופלזמה.בציטופלזמה יש מספר מבנים פורמליים בעלי מאפיינים קבועים של המבנה וההתנהגות בתקופות שונות של חיי התא. לכל אחד מהמבנים הללו יש תפקיד ספציפי. מכאן נבעה השוואתם לאיברי האורגניזם כולו, שבקשר אליו קיבלו את השם אברונים, או אברונים. בציטופלזמה מופקדים חומרים שונים - תכלילים (גליקוגן, טיפות שומן, פיגמנטים). הציטופלזמה חדורת ממברנות רטיקולום אנדופלזמי.
רשת אנדופלזמית (EMF). הרשת האנדופלזמית היא רשת מסועפת של תעלות וחללים בציטופלזמה של תא, שנוצרה על ידי ממברנות. על ממברנות התעלות ישנם אנזימים רבים המבטיחים את הפעילות החיונית של התא. ישנם 2 סוגים ממברנות EMF- חלק ומחוספס. על ממברנות רטיקולום אנדופלזמי חלקיש מערכות אנזימים המעורבות בשומן ו חילוף חומרים של פחמימות. פונקציה עיקרית רטיקולום אנדופלזמי מחוספס- סינתזת חלבון, המתבצעת בריבוזומים המחוברים לממברנות. רשת אנדופלזמית- זוהי מערכת זרימת דם תוך תאית נפוצה, דרך ערוציה מועברים חומרים בתוך התא ומתא לתא.
ריבוזומיםלבצע את הפונקציה של סינתזת חלבון. ריבוזומים הם חלקיקים כדוריים בקוטר של 15-35 ננומטר, המורכבים מ-2 תת יחידות בגדלים לא שווים ומכילים כמות שווה בערך של חלבוני mRNA. ריבוזומים בציטופלזמה ממוקמים או מחוברים למשטח החיצוני של הממברנות של הרשת האנדופלזמית. בהתאם לסוג החלבון המסונתז, ניתן לשלב ריבוזומים לקומפלקסים - פוליריבוזומים. ריבוזומים קיימים בכל סוגי התאים.
מתחם גולגי.האלמנט המבני העיקרי מתחם גולגיהוא קרום חלק היוצר חבילות של בורות מים שטוחים, או ואקוולים גדולים, או שלפוחיות קטנות. בורות המים של מתחם גולגי מחוברים לערוצים של הרשת האנדופלזמית. חלבונים, פוליסכרידים, שומנים המסונתזים על גבי ממברנות הרשת האנדופלזמית מועברים אל המכלול, מתעבים בתוך מבניו ו"ארוזים" בצורה של סוד מוכן לשחרור, או משמשים בתא עצמו במהלך חייו.
מיטוכונדריה.ההפצה הכללית של המיטוכונדריה בבעלי החיים ו צוֹמֵחַלהצביע על התפקיד החשוב ש מיטוכונדריהלשחק בכלוב. מיטוכונדריהבעלי צורה של גופים כדוריים, סגלגלים וגופים גליליים, הם יכולים להיות חוטיים. גודל המיטוכונדריה הוא 0.2-1 מיקרומטר בקוטר, עד 5-7 מיקרומטר באורך. אורכם של צורות חוטיות מגיע ל-15-20 מיקרון. מספר המיטוכונדריות בתאים של רקמות שונות אינו זהה, יש יותר מהן בהן התהליכים הסינתטיים אינטנסיביים (כבד) או שעלויות האנרגיה גבוהות. דופן המיטוכונדריה מורכבת מ-2 ממברנות - חיצוניות ופנימיות. הממברנה החיצונית חלקה, ומחיצות - רכסים, או cristae, יוצאות מהפנים הפנימי של האורגנואיד. על הממברנות של הקריסטה ישנם אנזימים רבים המעורבים בחילוף החומרים באנרגיה. הפונקציה העיקרית של המיטוכונדריה - סינתזה של ATP.
ליזוזומים- גופים סגלגלים קטנים בקוטר של כ-0.4 מיקרון, מוקפים בקרום תלת-שכבתי אחד. ליזוזומים מכילים כ-30 אנזימים המסוגלים לפרק חלבונים, חומצות גרעין, פוליסכרידים, שומנים וחומרים נוספים. פירוק חומרים על ידי אנזימים נקרא תְמוּגָה, לכן נקרא האורגנואיד ליזוזום. מאמינים שליזוזומים נוצרים מהמבנים של קומפלקס גולגי או ישירות מהרשת האנדופלזמית. פונקציות של ליזוזומים : עיכול תוך תאי של חומרי הזנה, הרס של מבנה התא עצמו במהלך מותו במהלך התפתחות עוברית, כאשר הרקמות העובריות מוחלפות ברקמות קבועות, ובמספר מקרים נוספים.
סנטריולים.מרכז התא מורכב מ-2 גופים גליליים קטנים מאוד הממוקמים בזוויות ישרות זה לזה. גופים אלו נקראים צנטריולים. דופן הצנטרול מורכבת מ-9 זוגות של מיקרוטובולים. סנטריולים מסוגלים להרכיב את עצמם והם אברונים מתרבים בעצמם של הציטופלזמה. לצנטרולים תפקיד חשוב בחלוקת תאים: הם מתחילים את הצמיחה של מיקרוטובולים היוצרים את ציר החלוקה.
הליבה.הליבה היא החשובה ביותר רְכִיבתאים. הוא מכיל מולקולות DNA ולכן מבצע שתי תפקידים עיקריים: 1) אחסון ושחזור מידע גנטי, 2) ויסות תהליכים מטבוליים המתרחשים בתא. תא שאבד הליבה, לא יכול להתקיים. הגרעין גם אינו מסוגל להתקיים עצמאי. לרוב התאים יש גרעין אחד, אך ניתן לראות 2-3 גרעינים בתא אחד, למשל, בתאי כבד. תאים מרובי גרעינים ידועים עם מספר הגרעינים בכמה עשרות. צורת הגרעינים תלויה בצורת התא. הגרעינים הם כדוריים, מרובי אונות. הגרעין מוקף בקרום המורכב משני ממברנות בעלות המבנה התלת-שכבתי הרגיל. הממברנה הגרעינית החיצונית מכוסה בריבוזומים, הממברנה הפנימית חלקה. את התפקיד העיקרי בפעילות החיונית של הגרעין ממלא המטבוליזם בין הגרעין לציטופלזמה. התוכן של הגרעין כולל מיץ גרעיני, או קריופלזמה, כרומטין וגרעין. הרכב המיץ הגרעיני כולל חלבונים שונים, כולל רוב האנזימים הגרעיניים, נוקלאוטידים חופשיים, חומצות אמינו, תוצרי הגרעין והכרומטין, הנעים מהגרעין לציטופלזמה. כרומטיןמכיל DNA, חלבונים ומהווה חלקים ספירליים ודחוסים של כרומוזומים. גרעיןהוא גוף מעוגל צפוף הממוקם במיץ הגרעיני. מספר הנוקלאולי משתנה בין 1 ל-5-7 או יותר. יש גרעינים רק בגרעינים שאינם מתחלקים, בזמן מיטוזה הם נעלמים, ולאחר השלמת החלוקה הם נוצרים שוב. הגרעין אינו אברון תא עצמאי; הוא נטול ממברנה ונוצר סביב אזור הכרומוזום שבו מקודד מבנה ה-rRNA. בגרעין נוצרים ריבוזומים, אשר עוברים לאחר מכן לציטופלזמה. כרומטיןהנקראים גושים, גרגירים ומבנים דמויי רשת של הגרעין, מוכתמים בעוצמה בצבעים מסוימים ושונים בצורתם מהגרעין.
2)1. תורת התא
תורת התא היא רעיון כללי של מבנה התאים כיחידות חיים, רבייתם ותפקידם ביצירת אורגניזמים רב-תאיים.
הופעתן וניסוחן של הוראות מסוימות תורת התאקדמה תקופה ארוכה למדי של הצטברות של תצפיות על המבנה של אורגניזמים חד-תאיים ורב-תאיים שונים של צמחים ובעלי חיים. תקופה זו הייתה קשורה בפיתוח היישום והשיפור של שיטות מחקר אופטיות שונות.
רוברט הוק היה הראשון שצפה, באמצעות עדשות מגדילות, את חלוקת רקמת הפקק ל"תאים", או "תאים". תיאוריו הולידו מחקרים שיטתיים של אנטומית הצמחים, אשר אישרו את תצפיותיו של רוברט הוק והראו כי חלקי הצמחים השונים מורכבים מ"שלפוחיות" או "שקיות" מרווחות. מאוחר יותר גילה א' לוונהוק את עולם האורגניזמים החד-תאיים וראה לראשונה תאים של בעלי חיים. מאוחר יותר, תאי בעלי חיים תוארו על ידי פ. פונטנה; אך מחקרים רבים אלה ואחרים לא הובילו באותה תקופה להבנת האוניברסליות של המבנה התא, לרעיונות ברורים לגבי מהו תא. התקדמות בחקר המיקרואנטומיה והתאים קשורה להתפתחות המיקרוסקופיה במאה ה-19. בשלב זה, הרעיונות לגבי מבנה התאים השתנו: לא דופן התא, אלא התוכן האמיתי שלו, הפרוטופלזמה, החל להיחשב לדבר העיקרי בארגון התא. בפרוטופלזמה התגלה מרכיב קבוע בתא, הגרעין. כל התצפיות הרבות הללו אפשרו ל-T. Schwann בשנת 1838 לעשות מספר הכללות. הוא הראה שתאי צמחים ובעלי חיים דומים זה לזה ביסודו. "הכשרון של טי שוואן לא היה בכך שהוא גילה תאים ככאלה, אלא שהוא לימד חוקרים להבין את משמעותם". רעיונות אלו פותחו עוד בעבודותיו של ר' וירצ'וב. יצירת תורת התא הפכה לאירוע מרכזי בביולוגיה, אחת ההוכחות המכריעות לאחדות כל הטבע החי. לתיאוריית התא הייתה השפעה משמעותית על התפתחות הביולוגיה, שימשה את הבסיס העיקרי לפיתוח דיסציפלינות כמו אמבריולוגיה, היסטולוגיה ופיזיולוגיה. הוא סיפק את היסודות להבנת החיים, להסבר היחסים בין אורגניזמים, להבנת התפתחות הפרט.
ההוראות העיקריות של תורת התאים שמרו על משמעותן עד היום, אם כי במשך יותר ממאה וחמישים שנה התקבל מידע חדש על מבנה, פעילות חיונית והתפתחות של תאים. תורת התא מניחה כיום:
1) התא הוא היחידה היסודית של החיים: - אין חיים מחוץ לתא.
2) תא הוא מערכת אחת המורכבת מאלמנטים רבים הקשורים זה לזה באופן טבעי, המייצגים היווצרות אינטגרלית מסוימת, המורכבת מיחידות פונקציונליות מצומדות - אברונים או אורגנואידים.
3) תאים דומים - הומולוגיים - במבנה ובתכונות בסיסיות.
4) מספר התאים גדל על ידי חלוקת התא המקורי לאחר הכפלת החומר הגנטי שלו: תא אחר תא.
5) אורגניזם רב תאי הוא מערכת חדשה, אנסמבל מורכב של תאים רבים, מאוחדים ומשולבים במערכות של רקמות ואיברים, המחוברים זה לזה בעזרת גורמים כימיים, הומוראליים ועצבניים.
6) תאים של אורגניזמים רב-תאיים הם טוטיפוטנטיים, כלומר. בעלי העוצמה הגנטית של כל התאים של אורגניזם נתון, שוות ערך מבחינת מידע גנטי, אך נבדלות זו מזו בביטוי שונה של גנים שונים, מה שמוביל לגיוון המורפולוגי והתפקודי שלהם - להתמיינות.
הרעיון של תא כיחידה חיונית עצמאית ניתן בעבודותיו של T. Schwann. ר' וירצ'וב גם האמין שכל תא נושא את מלוא מאפייני החיים: "התא הוא האלמנט המורפולוגי האחרון של כל הגופים החיים, ואין לנו זכות לחפש פעילות חיים אמיתית מחוצה לו".
המדע המודרני הוכיח במלואו הצעה זו. בספרות הפופולרית תא נקרא לרוב "אטום חיים", "קוואנטום של חיים", ובכך מדגיש שתא הוא היחידה הקטנה ביותר של חומר חי, שמחוץ לה אין חיים.
מאפיין כללי כזה של התא צריך, בתורו, להתבסס על הגדרת החי - מה חי, מה זה חיים. קשה מאוד לתת הגדרה סופית של החיים, של החיים.
M.V. וולקנשטיין נותן את ההגדרה הבאה לחיים: "אורגניזמים חיים הם מערכות פתוחות, מווסתות את עצמן ומתרבות בעצמן, שהחומרים המתפקדים החשובים ביותר שלהן הם חלבונים וחומצות גרעין". יצורים חיים מאופיינים במספר תכונות מצטברות, כמו יכולת רבייה, שימוש והמרה של אנרגיה, חילוף חומרים, רגישות ושונות. ושילוב כזה של הסימנים הללו ניתן למצוא על ברמה התאית. אין יחידת חיים קטנה יותר מתא. אנו יכולים לבודד רכיבים בודדים או אפילו מולקולות מתא ולוודא שלרבים מהם יש תכונות פונקציונליות ספציפיות. לפיכך, סיבורי אקטומיוזין מבודדים יכולים להתכווץ בתגובה לתוספת של ATP; מחוץ לתא, אנזימים רבים המשתתפים בסינתזה או פירוק של מולקולות ביו-אורגניות מורכבות "פועלות" בצורה מושלמת; ריבוזומים מבודדים בנוכחות הגורמים הדרושים יכולים לסנתז חלבון, פותחו מערכות לא תאיות לסינתזה אנזימטית של חומצות גרעין וכו'. האם כל הרכיבים התאיים, המבנים, האנזימים, המולקולות הללו יכולים להיחשב חיים? האם קומפלקס האקטומיוזין יכול להיחשב חי? נראה שלא, ולו רק בגלל שיש לו רק חלק ממכלול הנכסים של החיים. כך גם לגבי שאר הדוגמאות. רק התא כשלעצמו הוא היחידה הקטנה ביותר שיש לה את כל המאפיינים ביחד העונים להגדרה של "חי".
3) הבסיס של מנגנון פני השטח של תאים (PAC) הוא בָּחוּץ קרום התא, או פלזמהלמה.בנוסף לפלזמה, ל-PAC יש קומפלקס אפיממברני, בעוד לאאוקריוטים יש גם קומפלקס תת-ממברני. המרכיבים הביוכימיים העיקריים של הפלזמהלמה (מהפלזמה היוונית - היווצרות ולמה - קליפה, קרום) הם שומנים וחלבונים. היחס הכמותי שלהם ברוב האיקריוטים הוא 1:1, ובפרוקריוטים, החלבונים שולטים בפלסמה. כמות קטנה של פחמימות נמצאת בקרום התא החיצוני וניתן למצוא תרכובות דמויות שומן (ביונקים - כולסטרול, ויטמינים מסיסים בשומן). בשנת 1925 הציעו E. Gorter ו-F. Grendel (הולנד) שהבסיס של הממברנה הוא שכבה כפולה של שומנים - השכבה הביליפידית. בשנת 1935, ג'יי דניאלי וג'י דוסון הציעו את המודל המרחבי הראשון של ארגון הממברנה, שנקרא מודל "סנדוויץ'" או "סנדוויץ'". לדעתם, בסיס הממברנה הוא השכבה הביליפידית, ושני משטחי השכבה מכוסים בשכבות רצופות של חלבונים. מחקר נוסף של ממברנות התא, כולל הפלזמהלמה, הראה שכמעט בכל המקרים יש להם מבנה דומה. בשנת 1972, S. Singer and G. Nicholson (ארה"ב) גיבשו את המושג של מבנה נוזלי-פסיפסממברנות תאים (איור). על פי מודל זה, ממברנות מבוססות על שכבת שומנים,אבל החלבונים שבו ממוקמים במולקולות ובמתחמים נפרדים, כלומר. פסיפס (מצרפתית. פסיפס - פסיפס; תמונה המורכבת מחתיכות נפרדות). בפרט, מולקולות של חלבונים אינטגרליים (מלטינית אינטגרר - שלם) יכולות לחצות את השכבה הביליפידית, חצי אינטגרלית - לטבול בה חלקית, והיקפית (מהפריפריה היוונית - עיגול) - הממוקמת על פני השטח שלה (איור). ביולוגיה מולקולרית מודרנית אישרה את תוקפו של מודל הפסיפס הנוזל, אם כי התגלו גרסאות אחרות של קרומי תאים. בפרט, בארכיבקטריה, הבסיס של הממברנה הוא חד-שכבה של שומנים מורכבים, וחלק מהחיידקים מכילים שלפוחיות ממברנות בציטופלזמה, שקירותיה מיוצגים על ידי חד-שכבה חלבונית. קומפלקס סופרממברנימנגנון פני השטח של תאים מאופיין במגוון של מבנים (איור). בפרוקריוטים, קומפלקס האפיממברנה ברוב המקרים מיוצג על ידי דופן תא בעוביים שונים, שבסיסו הוא המורכב גליקופרוטאין murein (בארכיבקטריה, פסאודומוראין). במספר אובקטריות, החלק החיצוני של קומפלקס האפיממברנה מורכב מממברנה נוספת בעלת תכולה גבוהה של ליפופוליסכרידים, באאוקריוטים, המרכיב האוניברסלי של הקומפלקס העל-ממברני הוא פחמימות - מרכיבים של גליקוליפידים וגליקופרוטאין של הפלזמהלמה. בשל כך, הוא נקרא במקור glycocalyx (מהיוונית גליקוס - מתוק, פחמימה וקאלום הלטינית - עור עבה, קליפה). בנוסף לפחמימות, חלבונים היקפיים מעל השכבה הביליפידית כלולים בגליקוקאליקס. גרסאות מורכבות יותר של קומפלקס האפיממברנה נמצאות בצמחים (דופן התא עשוי מתאית), פטריות ופרוקי רגליים (כיסוי חיצוני עשוי מכיטין). תת-ממברנה(מ-lat. sub - under) הקומפלקס אופייני רק לתאים איקריוטיים. הוא מורכב ממגוון של מבנים חוטי חלבון: סיבים דקים (מפיבריל לטיני - סיבים, חוט), מיקרו-סיביים (ממיקרוים יווניים - קטנים), סיבים שלד (מהשלד היווני - מיובשים) ומיקרוטובולים. הם מחוברים זה לזה על ידי חלבונים ויוצרים את מנגנון השריר והשלד של התא. הקומפלקס התת-ממברני מקיים אינטראקציה עם חלבוני הממברנה הפלזמה, אשר, בתורם, קשורים לקומפלקס העל-ממברני. כתוצאה מכך, PAH היא מערכת אינטגרלית מבחינה מבנית. זה מאפשר לו לבצע פונקציות חשובות עבור התא: בידוד, הובלה, קטליטי, קולטן איתות ומגע.
4) הממברנות מכילות גם גליקוליפידים וכולסטרול. גליקוליפידיםהם שומנים עם פחמימות מחוברות אליהם. כמו פוספוליפידים, גליקוליפידיםיש ראשים קוטביים וזנבות לא קוטביים. הכולסטרול קרוב לשומנים; למולקולה שלה יש גם חלק קוטבי.
1. היוצר של תורת התא הוא:
ר' וירקוב
IN AND. ורנדסקי
T.Schwann
2. תורת התא נוצרה ב:
1839
3. קבוצה של אברוני ממברנה של הציטופלזמה של תאים אוקריוטיים:
ריבוזומים, מיטוכונדריה, פלסטידים, ליזוזומים
ספרוזומים, וואקוולים, פלסטידים
מיקרוטובולים, מיקרופילמנטים, ריבוזומים, צנטרוזום
EPS, ריבוזומים, ליזוזומים, מנגנון גולגי
מיטוכונדריה, ER, מנגנון גולגי
4. אברונים למטרות כלליות:
מיופיברילים
ריבוזומים, מיטוכונדריה
EPS, מנגנון גולגי
דגלים, ריבוזומים
פלאגלה, מיופיברילים
5. בגרעין הבין-פאזי, ישנם:
ריבוזומים
נוקלאולי
קריולמה
מיץ גרעיני
כרומטין
6. סוגי כרומוזומים בהתאם למיקום הצנטרומר:
תת-מרכזי
טלוצנטרי
אקרוצנטרי
מטאצנטרי
כתף לא אחידה
7. כל כרומוזום במטאפאזה של מיטוזה מורכב מ:
DNA, חלבוני היסטון, חלבונים שאינם היסטונים
2x מיקרופיברילים
2x כרומטידות
4x semichromatids
כרומונם
8. לתאים סומטיים יש קבוצה של כרומוזומים:
פוליפלואיד
דיפלואיד
הפלואיד
טטרפלואיד
9. סינתזה מתבצעת על EPS חלק:
פחמימות
10. כרומופלסטים מכילים פיגמנט:
הֵמוֹגלוֹבִּין
ליקופן
קרוטן
קסנטופיל
כלורופיל
11. פונקציות של ריבוזומים:
הורמונים
סינתזה של פחמימות
חומצות גרעין
סינתזת חלבון
סינתזה של שומנים
12. קבוצות הכללה:
ויטמין
טרופי
מינרל
הפרשה, הפרשה
פיגמנט
13. לתאי מין יש קבוצה של כרומוזומים:
דיפלואיד
פוליפלואיד
פלואיד
14. איקריוטים כוללים:
ציאנובקטריה
בעלי חיים
חזזיות
צמחים
15. קבוצה של אברונים לא ממברניים של הציטופלזמה של תאים אוקריוטיים:
ריבוזומים, ואקוולים, מיקרוטובולים
ריבוזומים, צנטרוזום
ER, מיטוכונדריה, ריבוזומים
microtubules, microfibrils
מיקרופילמנטים, פלסטידים, ליזוזומים
16. פונקציות של היאלופלזמה:
השתתפות בחילוף החומרים
מְלָאכוּתִי
תוחם
הומיאוסטטי
הובלה
17. תפקידי הנוקלאולי:
סינתזת חלבון
יצירת תת-יחידות ריבוזום
סינתזת RNA לתחבורה
היווצרות מיקרופיברילים
סינתזה של RNA ריבוזומלי
18. פונקציות EPS:
קוֹלֵט
הובלה
תוחם
מָגֵן
סינטטי
19. פונקציות של ליזוזומים:
סינתזה והצטברות של פחמימות
הרס של איברים זמניים של עוברים וזחלים
סינתזה של חלבונים, שומנים, פחמימות
אוטוליזה, הרס של חלקיקים זרים
עיכול תוך תאי
20. Vacuoles הם:
אברונים שאינם קרומיים של תאי בעלי חיים
אברוני ממברנה של תאי צמחים
אברוני ממברנה של תאי חיידקים
אברוני ממברנה של בעלי חיים חד-תאיים
אברונים לא קרומיים של תאי צמחים
21. פונקציות של ואקוולים:
להסיר עודפי מים
לשמור על הלחץ האוסמוטי של התא
לבצע עיכול תוך תאי
לשמור על לחץ טורגור בתאי הצמח
לבצע סינתזה והצטברות שומנים בתאי הצמח
22. כלורופלסטים מכילים פיגמנט:
קסנתופיל
כלורופיל
23. ריבוזומים כוללים:
T-RNA, מגנזיום
חלבונים, DNA, מגנזיום
רק סנאים
חלבונים, i-RNA
חלבונים, r-RNA, מגנזיום
24. הרכב המיקרופילמנטים כולל חלבון:
25. מיקרוטובולים מכילים חלבון:
טובולין
26. מיקרופיברילים מכילים חלבון:
קולגן
קרטין
27. הגרעין אופייני לתאים:
כל היצורים החיים
אוקריוט
צמחים
פרוקריוט