^ קרום פלזמה , או פלזמהלמה, תופס מקום מיוחד בין ממברנות תאים שונות. זהו מבנה היקפי שטחי המגביל את התא מבחוץ, מה שקובע את הקשר הישיר שלו עם הסביבה החוץ-תאית, וכתוצאה מכך עם כל החומרים והגירויים הפועלים על התא. לכן, תפקידה של קרום הפלזמה הוא להוות מחסום, מחסום בין התוכן התוך תאי המאורגן בצורה מורכבת לבין הסביבה החיצונית. במקרה זה, הפלסמולמה מבצעת לא רק את התפקיד של מחסום מכני, אלא, והכי חשוב, היא מגבילה את הזרימה החופשית של חומרים נמוכים וגבוהים מולקולריים בשני הכיוונים דרך הממברנה. יתרה מכך, הפלסמולמה פועלת כמבנה ש"מזהה", קולטנים, חומרים כימיים שונים ומווסתת באופן סלקטיבי את ההובלה של חומרים אלו אל תוך התא וממנו. במילים אחרות, קרום הפלזמה מבצע פונקציות הקשורות להובלה טרנסממברנית סלקטיבית מווסתת של חומרים ופועל כמנתח תאים ראשוני. בהקשר זה, ניתן לשקול את הפלזמהלמה אורגנואיד התאנכלל במערכת הוואקואולרית של התא. כמו ממברנות אחרות של מערכת זו (הממברנות של הליזוזומים, האנדוזומים, מנגנון הגולגי וכו'), היא נוצרת ומתעדכנת עקב הפעילות הסינתטית של הרשת האנדופלזמית ובעלת הרכב דומה. למרבה הפלא, ניתן להשוות את קרום הפלזמה לממברנה של ואקוול תוך תאי, אך הופכת מבפנים החוצה: היא אינה מוקפת בהיאלופלזמה, אלא מקיפה אותה.
תפקיד הובלת מחסום של הפלזמה
מקיף את התא מכל צדדיו, קרום הפלזמה פועל כמחסום מכני. על מנת לנקב אותו במיקרו מחטים או מיקרופיפטות, נדרש מאמץ רב למדי. עם הלחץ של מחט מיקרו עליה, היא תחילה מתכופפת חזק, ורק אז פורצת דרך. ממברנות שומנים מלאכותיות פחות יציבות. יציבות מכנית זו של ממברנת הפלזמה עשויה להיקבע על ידי רכיבים נוספים כגון הגליקוקאליקס והשכבה הקורטיקלית של הציטופלזמה (איור 127).
גליקוקליקסהיא שכבה חיצונית לממברנת הליפופרוטאין המכילה שרשראות פוליסכרידים של חלבונים אינטגרליים של הממברנה - גליקופרוטאין. שרשראות אלו מכילות פחמימות כמו מנוז, גלוקוז, N-אצטיל גלוקוזאמין, חומצה סיאלית וכו'. הטרופולימרים פחמימות כאלה יוצרים שרשראות מסועפות, ביניהן ניתן לאתר גליקוליפידים ופרוטאוגליקנים המבודדים מהתא. שכבת ה-glycocalyx מושקת בכבדות, בעלת עקביות דמוית ג'לי, מה שמפחית משמעותית את קצב הדיפוזיה באזור זה. חומרים שונים. אנזימים הידרוליטיים המופרשים מהתא, המעורבים בביקוע החוץ-תאי של פולימרים (עיכול חוץ-תאי) למולקולות מונומר, אשר מועברות לאחר מכן אל הציטופלזמה דרך קרום הפלזמה, יכולים גם "להיתקע" כאן.
IN מיקרוסקופ אלקטרוני, במיוחד כש שיטות מיוחדותבניגוד לפוליסכרידים, ל-glycocalyx יש צורה של שכבה סיבית רופפת, בעובי 3-4 ננומטר, המכסה את כל פני התא. הגליקוקאליקס מתבטא בצורה טובה במיוחד בגבול המברשת של תאי אפיתל המעי הסופג (אנטרוציטים), עם זאת, הוא נמצא כמעט בכל תאי החי, אך דרגת חומרתו שונה (איור 128).
היציבות המכנית של קרום הפלזמה, בנוסף, מסופקת על ידי מבנה השכבה הקורטיקלית הסמוכה לה מצד הציטופלזמה ומבנים פיברילריים תוך תאיים. קליפת המוח(מהמילה - קורטקס - לנבוח, לקלף) שִׁכבָהציטופלזמה שוכבת במגע הדוק עם ליפופרוטאינים קרום חיצוני, יש מספר תכונות. כאן, בעובי של 0.1-0.5 מיקרון, אין ריבוזומים ושלפוחיות ממברנות, אלא ב במספרים גדוליםישנם אלמנטים פיברילרים של הציטופלזמה - מיקרופילמנטים ולעתים קרובות מיקרוטובולים. המרכיב הפיברילרי העיקרי בשכבת הקורטיקלית הוא רשת של מיקרו-סיבי אקטין. כאן ממוקמים גם מספר חלבוני עזר, הנחוצים לתנועת קטעים של הציטופלזמה (לפרטים נוספים על מערכת השלד-מוטורית של התאים, ראה להלן). תפקידם של חלבונים הקשורים לאקטין אלו חשוב מאוד, שכן הוא מסביר את השתתפותם בחיבור, ב"עיגון" החלבונים האינטגרליים של קרום הפלזמה.
בפרוטוזואה רבים, במיוחד ריסים, קרום הפלזמה לוקח חלק בהיווצרות כדורים, שכבה קשיחה שקובעת לרוב את צורת התא. שקי ממברנה יכולים לצמוד לממברנת הפלזמה כאן מבפנים; במקרה זה, ישנן שלוש שכבות קרום ליד פני התאים: קרום הפלזמה עצמו ושתי ממברנות של alveoli pellicular. בסיליה של הנעל, הגלעין יוצר עיבויים, הממוקמים בצורה של משושים, שבמרכזם יוצאות ריסים (איור 129). הקשיחות של תצורות קליקולריות יכולה להיות קשורה גם לאלמנטים של הציטופלזמה הנמצאת בבסיס קרום הפלזמה, עם השכבה הקורטיקלית. כך, בפסגות של euglena pellicle ליד הממברנה, בנוסף ל-vacuoles של הממברנה, נמצאות צרורות מקבילות של מיקרו-צינוריות ומיקרו-פילמנטים. חיזוק היקפי פיברילרי זה, יחד עם היקפי הממברנה הרב-שכבתית המקופלת, יוצר מבנה פלקל קשיח.
תפקיד המחסום של הפלזמלמה מורכב גם בהגבלת הדיפוזיה החופשית של חומרים. ניסויי מודל על ממברנות שומנים מלאכותיות הראו שהן חדירות למים, גזים, מולקולות קטנות לא קוטביות של חומרים מסיסים בשומן, אך אטומות לחלוטין למולקולות טעונות (יונים) ולמולקולות גדולות ולא טעונות (סוכרים) (איור 130).
ממברנות טבעיות גם מגבילות את קצב החדירה של תרכובות במשקל מולקולרי נמוך לתא.
הובלה טרנסממברנית של יונים ותרכובות במשקל מולקולרי נמוך
קרום הפלזמה, כמו ממברנות תאי ליפופרוטאין אחרות, חדיר למחצה. המשמעות היא שמולקולות שונות עוברות דרכו במהירויות שונות ו מידה גדולה יותרמולקולות, ככל שהמעבר שלהן דרך הממברנה יהיה איטי יותר. תכונה זו מגדירה את קרום הפלזמה כמחסום אוסמוטי. למים וגזים המומסים בו יש את יכולת החדירה המקסימלית, יונים חודרים לממברנה הרבה יותר לאט (בערך פי 10 4 לאט יותר). לכן, אם תא, למשל, אריתרוציט, ממוקם בסביבה שבה ריכוז המלחים נמוך יותר מאשר בתא (יתר לחץ דם), אז מים מבחוץ יזרזו לתוך התא, מה שיוביל לעלייה של נפח התא ועד לקרע של קרום הפלזמה ("הלם היפוטוני"). להיפך, כאשר מניחים אריתרוציט בתמיסות מלח, יותר ריכוז גבוהמאשר בתא, תהיה יציאה של מים מהתא לסביבה החיצונית. במקביל, התא יתקמט, יקטן בנפח.
הובלה פסיבית כזו של מים אל מחוץ לתא ואל התא עדיין מתנהלת בקצב נמוך. קצב חדירת המים דרך הממברנה הוא כ-10 -4 ס"מ לשנייה, שהם פי 100,000 פחות מקצב הדיפוזיה של מולקולות המים בשכבה מימית בעובי 7.5 ננומטר. הגיע למסקנה כי ב קרום תא, בשכבת הליפופרוטאין שלו יש "נקבוביות" מיוחדות לחדירת מים ויונים. מספרם אינו כה גדול: השטח הכולל עם גודל של "נקבובית" בודדת של כ-0.3-0.8 ננומטר צריך להיות רק 0.06% משטח התא כולו.
שלא כמו ממברנות שומנים מלאכותיות דו-שכבתיות, ממברנות טבעיות, ובעיקר קרום הפלזמה, עדיין מסוגלות להעביר יונים ומונומרים רבים, כגון סוכרים, חומצות אמינו וכו'. החדירות ליונים נמוכה, וקצב המעבר של יונים שונים הוא לא אותו הדבר. קצב מעבר גבוה יותר עבור קטיונים (K + , Na +) ונמוך בהרבה עבור אניונים (Cl -).
הובלת יונים דרך הפלזמה מתרחשת עקב ההשתתפות בתהליך זה של חלבוני הובלה ממברנה - permeases. חלבונים אלו יכולים להעביר חומר אחד לכיוון אחד (אוניפורט) או מספר חומרים בו זמנית (סימפורט), או יחד עם יבוא של חומר אחד, להוציא אחר מהתא (אנטיפורט). אז, למשל, גלוקוז יכול להיכנס לתאים באופן סימפורטלי יחד עם יון Na +.
הובלת יונים יכולה להתבצע לאורך שיפוע הריכוז - באופן פסיביללא צריכת אנרגיה נוספת. לדוגמה, יון Na + נכנס לתא סביבה חיצוניתכאשר הריכוז שלו גבוה יותר מאשר בציטופלזמה. במקרה של הובלה פסיבית, חלק מחלבוני הובלת הממברנה יוצרים קומפלקסים מולקולריים, ערוציםדרכם עוברות מולקולות מומסות דרך הממברנה על ידי דיפוזיה פשוטה לאורך שיפוע ריכוז. חלק מהערוצים הללו פתוחים לצמיתות, בעוד שהחלק השני יכול להיסגר או להיפתח בתגובה לקשירה למולקולות איתות או לשינויים בריכוז היונים התוך-תאיים. במקרים אחרים, חלבוני ממברנה מיוחדים - מוביליםנקשרים באופן סלקטיבי ליון כזה או אחר ומעבירים אותו דרך הממברנה (דיפוזיה מסייעת) (איור 131).
נראה שנוכחותם של תעלות הובלה ונשאים של חלבונים כאלה אמורה להוביל לשיווי משקל בריכוזי יונים וחומרים בעלי משקל מולקולרי נמוך משני צידי הממברנה. למעשה, זה לא כך: ריכוז היונים בציטופלזמה של תאים שונה באופן חד לא רק מזה שבסביבה החיצונית, אלא אפילו מפלסמת הדם השוטפת את התאים בגוף החי. על השולחן 14 מציג את ריכוזי היונים בתוך התא ומחוצה לו.
טבלה 14
| והוא | ריכוז תוך תאי, mM | ריכוז חוץ תאי, mM |
| Na+ | 5-15 | 145 |
| K+ | 140 | 5 |
| Mg2+ | 30 | 1-2 |
| *Ca2+ | 1-2 | 2,5-5 |
| Cl- | 4 | 110 |
*ריכוז Ca 2+ במצב חופשי בציטוזול של תאים אוקריוטיים הוא 10 -7 M, ומחוץ 10 -3 M.
כפי שניתן לראות, במקרה זה, הריכוז הכולל של קטיונים חד ערכיים בתוך ומחוץ לתאים כמעט זהה (150 מ"מ), איזוטוני. אבל מסתבר שבציטופלזמה הריכוז של K+ גבוה כמעט פי 50, ו-Na+ נמוך יותר מאשר בפלזמה בדם. יתרה מכך, הבדל זה נשמר רק בתא חי: אם התא נהרג או מדוכאים התהליכים המטבוליים בו, אזי לאחר זמן מה ההבדלים היוניים משני צידי קרום הפלזמה ייעלמו. אתה יכול פשוט לקרר את התאים ל-+2 0 C, ולאחר זמן מה הריכוז של K + ו- Na + משני צידי הממברנה יהפוך להיות זהה. כאשר התאים מחוממים, ההבדל הזה משוחזר. תופעה זו נובעת מכך שיש בתאים נשאי חלבון ממברנה הפועלים כנגד שיפוע הריכוז, תוך הוצאת אנרגיה עקב הידרוליזה של ATP. סוג זה של עבודה נקרא מעבר פעיל, והוא מתבצע בעזרת חלבון משאבות יונים. קרום הפלזמה מכיל משאבת מולקולה בת שתי תת-יחידות (K + + Na +), שהיא גם ATPase. במהלך הפעולה, משאבה זו שואבת 3 יוני Na + במחזור אחד ושואבת 2 יוני K + לתוך התא כנגד שיפוע הריכוז. במקרה זה, מולקלת מולקולת ATP אחת, אשר עוברת לזרחון ATPase, וכתוצאה מכך Na+ מועבר דרך הממברנה מהתא, ו-K+ מקבל את ההזדמנות להיקשר למולקולת החלבון ולאחר מכן מועבר לתוך תא (איור 132). כתוצאה מהובלה פעילה בעזרת משאבות ממברנות, מוסדר בתא גם ריכוז הקטיונים הדו ערכיים Mg 2+ ו- Ca 2+, גם בצריכת ATP.
כגון משרה מלאה permease ומשאבות יוצר ריכוז קבוע של יונים וחומרים במשקל מולקולרי נמוך בתא, יוצר את מה שנקרא. הומאוסטזיס, קביעות ריכוזים באופן אוסמוטי חומרים פעילים. יש לציין שכ-80% מסך ה-ATP של התא מושקע בשמירה על הומאוסטזיס.
בשילוב עם הובלה פעילה של יונים על פני ממברנת הפלזמה, מועברים סוכרים, נוקלאוטידים וחומצות אמינו שונות.
לפיכך, ההובלה הפעילה של גלוקוז, אשר באופן סימפטי (במקביל) נכנס לתא יחד עם זרימת יון ה-Na+ המועבר באופן פסיבי, תהיה תלויה בפעילות המשאבה (K + + Na +). אם משאבה זו (K + -Na +) תיחסם, אז בקרוב ההבדל בריכוז של Na + משני צידי הממברנה ייעלם, בעוד שהדיפוזיה של Na + לתוך התא תקטן, ובמקביל זרימת הגלוקוז לתא תיפסק. ברגע שהעבודה של (K + -Na +)-ATPase משוחזרת ונוצר הבדל בריכוז היונים, מיד עולה הזרימה המפוזרת של Na + ובמקביל גם הובלת הגלוקוז. באופן דומה, דרך הממברנה וזרימת חומצות האמינו, המועברות על ידי חלבוני נשא מיוחדים הפועלים כמערכות סימפורט, ומעבירות יונים בו-זמנית.
ההובלה הפעילה של סוכרים וחומצות אמינו בתאי חיידקים נובעת משיפוע של יוני מימן.
כשלעצמו, השתתפותם של חלבוני ממברנה מיוחדים המעורבים בהובלה פסיבית או אקטיבית של תרכובות במשקל מולקולרי נמוך מעידה על הספציפיות הגבוהה של תהליך זה. אפילו במקרה של הובלת יונים פסיבית, חלבונים "מזהים" יון נתון, מקיימים איתו אינטראקציה, נקשרים באופן ספציפי, משנים את המבנה שלהם ותפקודם. לכן, כבר על הדוגמה של תחבורה חומרים פשוטיםממברנות פועלות כמנתחים, כקולטנים. תפקיד קולטן זה בא לידי ביטוי במיוחד כאשר ביופולימרים נספגים בתא.
הובלה שלפוחית: אנדוציטוזיס ואקסוציטוזיס
מקרומולקולות כגון חלבונים, חומצות גרעין, פוליסכרידים, קומפלקסים ליפופרוטאינים ואחרים אינם עוברים דרך ממברנות התא, בניגוד לאופן שבו יונים ומונומרים מועברים. הובלת מיקרומולקולות, הקומפלקסים שלהן, חלקיקים אל תוך התא ומחוצה לו מתרחשת בצורה שונה לחלוטין - באמצעות העברה שלפוחית. מונח זה אומר שמקרומולקולות שונות, ביו-פולימרים או קומפלקסים שלהם אינם יכולים להיכנס לתא דרך קרום הפלזמה. ולא רק דרכו: ממברנות תא כלשהן אינן מסוגלות להעביר טרנס-ממברנה של ביופולימרים, למעט ממברנות שיש להן נשאי קומפלקס חלבון מיוחד - פורינים (ממברנות של מיטוכונדריה, פלסטידים, פרוקסיסומים). מקרומולקולות נכנסות לתא או מתא ממברנה אחד למשנהו סגורה בתוך ואקואולות או שלפוחיות. כגון העברה שלפוחיתניתן לחלק לשני סוגים: אקסוציטוזיס- הסרה של מוצרים מקרומולקולריים מהתא, ו אנדוציטוזיס- ספיגה של מקרומולקולות על ידי התא (איור 133).
במהלך אנדוציטוזיס, קטע מסוים של הפלזמהלמה לוכד, כביכול, את החומר החוץ-תאי, עוטף אותו ב-vacuole של הממברנה שנוצר עקב פלישה של קרום הפלזמה. בוואקום ראשוני כזה, או ב אנדוזום, כל ביופולימרים, קומפלקסים מקרומולקולריים, חלקי תאים או אפילו תאים שלמים יכולים להיכנס, שם הם מתפרקים, מתפרקים למונומרים, הנכנסים להיאלופלזמה על ידי העברה טרנסממברנית. המשמעות הביולוגית העיקרית של אנדוציטוזיס היא רכישת אבני בניין באמצעות עיכול תוך תאי, המתבצע בשלב השני של האנדוציטוזיס לאחר היתוך של האנדוזום הראשוני עם הליזוזום, וואקואול המכיל קבוצה של אנזימים הידרוליטיים (ראה להלן).
אנדוציטוזה מחולקת רשמית ל פינוציטוזהו פגוציטוזיס(איור 134). Phagocytosis - לכידה וספיגה של חלקיקים גדולים על ידי תא (לפעמים אפילו תאים או חלקיהם) - תוארה לראשונה על ידי I.I. Mechnikov. פגוציטוזיס, היכולת ללכוד חלקיקים גדולים על ידי תא, נמצאת בין תאים של בעלי חיים, הן חד-תאיים (לדוגמה, אמבה, כמה ריצות טורפות) והן תאים מיוחדים של בעלי חיים רב-תאיים. תאים מיוחדים, פגוציטים, אופייניים הן לחסרי חוליות (אמובוציטים של דם או נוזל חלל) והן לבעלי חוליות (נויטרופילים ומקרופאגים). פינוציטוזיס הוגדר במקור כספיגת מים או תמיסות מימיות חומרים שונים. כיום ידוע שגם פגוציטוזיס וגם פינוציטוזיס מתנהלים בצורה דומה מאוד, ולכן השימוש במונחים אלו יכול לשקף רק הבדלים בנפחים ובמסה של חומרים נספגים. המשותף לתהליכים הללו הוא שהחומרים הנספגים על פני קרום הפלזמה מוקפים בממברנה בצורת וואקוולה - אנדוזום, שנע בתוך התא.
אנדוציטוזיס, לרבות פינוציטוזיס ופגוציטוזיס, יכול להיות לא ספציפי או מכונן, קבוע וספציפי, בתיווך קולטנים (רצפטור). אנדוציטו לא ספציפי h (פינוציטוזיס ופאגוציטוזיס), מה שנקרא מכיוון שהוא ממשיך כאילו אוטומטית ולעתים קרובות יכול להוביל ללכידה וספיגה של חומרים זרים לחלוטין או אדישים לתא, למשל, חלקיקי פיח או צבעים.
אנדוציטוזיס לא ספציפי מלווה לעתים קרובות בספיגה ראשונית של החומר הלוכד על ידי גליקוקליקס של ממברנת הפלזמה. הגליקוקאליקס, בשל הקבוצות החומציות של הרב-סוכרים שלו, הוא בעל מטען שלילי ונקשר היטב לקבוצות שונות של חלבונים טעונים חיובית. עם ספיחה כזו אנדוציטוזיס לא ספציפי, מקרומולקולות וחלקיקים קטנים (חלבונים חומציים, פריטין, נוגדנים, ויריון, חלקיקים קולואידים) נספגים. פינוציטוזיס בפאזה נוזלית מוביל לספיגה יחד עם המדיום הנוזלי של מולקולות מסיסות שאינן נקשרות לפלסמה.
עַל השלב הבאיש שינוי במורפולוגיה של פני התא: זה או הופעה של פלישות קטנות של קרום הפלזמה, פלישה, או שזה הופעה על פני התא של יציאות, קפלים או "סלסולים" (rafl - באנגלית) , אשר, כביכול, חופפים, מקפלים, מפרידים נפחים קטנים של מדיום נוזלי (איור 135, 136). הסוג הראשון של התרחשות של שלפוחית פינוציטית, פינוזומים, אופייני לתאי אפיתל המעי, אנדותל, לאמבות, השני - לפגוציטים ופיברובלסטים. תהליכים אלו תלויים באספקת אנרגיה: מעכבי נשימה חוסמים תהליכים אלו.
בעקבות סידור מחדש זה של פני השטח, מתבצע תהליך ההידבקות וההתמזגות של הממברנות המתקשרות, מה שמוביל להיווצרות שלפוחית פניציטית (פינוזום), המתנתקת משטח התא ונכנסת עמוק לתוך הציטופלזמה. גם אנדוציטוזיס לא ספציפי וגם קולטן, המוביל לביקוע שלפוחיות הממברנה, מתרחש באזורים מיוחדים של קרום הפלזמה. אלה הם מה שנקרא בורות גובלים. הם נקראים כך מכיוון שמצד הציטופלזמה, קרום הפלזמה מכוסה, לבוש, בשכבה סיבית דקה (כ-20 ננומטר), אשר בחתכים אולטרה-דקים, כביכול, גובלת, מכסה בליטות קטנות, בורות (איור. 137). כמעט לכל תאי בעלי החיים יש בורות אלה; הם תופסים כ-2% משטח התא. השכבה המקיפה מורכבת בעיקר מחלבון קלתריןקשור למספר חלבונים נוספים. שלוש מולקולות של קלתרין, יחד עם שלוש מולקולות של חלבון בעל משקל מולקולרי נמוך, יוצרות מבנה של טריסקליון, הדומה לצלב קרס בעל שלוש קרס (איור 138). טריסקליונים של קלתרין על פני השטח הפנימיים של הבורות של קרום הפלזמה יוצרים רשת רופפת המורכבת מחומשים ומשושים, המזכירים בדרך כלל סל. שכבת הקלתרין מכסה את כל היקף ה-vacuoles האנדוציטיים הראשוניים המפרידים, הגובלים על ידי שלפוחיות.
קלתרין שייך לאחד מהמינים שנקראים. "הלבשת" חלבונים (COP - חלבונים מצופים). חלבונים אלו נקשרים לחלבוני קולטן אינטגרליים מהצד של הציטופלזמה ויוצרים שכבת חבישה סביב היקף הפינוזום העולה, השלפוחית האנדוזומלית הראשונית - השלפוחית "התוחמת". בהפרדה של האנדוזום הראשוני מעורבים גם חלבונים - דינמינים, המתפלמרים סביב צוואר השלפוחית המפרידה (איור 139).
לאחר שהשלפוחית התחומה נפרדת מהפלסמולמה ומתחילה לעבור לעומק הציטופלזמה, שכבת הקלתרין מתפרקת, מתפרקת, קרום האנדוזום (פינוזומים) מקבל את צורתו הרגילה. לאחר אובדן שכבת הקלתרין, האנדוזומים מתחילים להתמזג זה עם זה.
נמצא כי הממברנות של הבורות התוחמים מכילים מעט כולסטרול יחסית, מה שיכול לקבוע את הירידה בנוקשות הממברנה ולתרום ליצירת בועות. המשמעות הביולוגית של הופעת "מעיל" קלתרין לאורך הפריפריה של השלפוחית עשויה להיות שהיא מספקת הידבקות של השלפוחיות התוחמות ליסודות שלד הציטוש והובלתם לאחר מכן בתא, ומונעת מהם להתמזג זה עם זה. .
עוצמת פינוציטוזיס לא ספציפי בשלב נוזלי יכולה להיות גבוהה מאוד. אז תא האפיתל מעי דקיוצר עד 1000 פינוזומים בשנייה, ומקרופאגים יוצרים כ-125 פינוזומים בדקה. גודל הפינוזומים קטן, הגבול התחתון שלהם הוא 60-130 ננומטר, אבל השפע שלהם מוביל לעובדה שבמהלך אנדוציטוזיס, הפלסמולמה מוחלפת במהירות, כאילו "בזבזתי" על היווצרות של וואקוולים קטנים רבים. אז במקרופאגים, כל קרום הפלזמה מוחלף תוך 30 דקות, בפיברובלסטים - תוך שעתיים.
גורלם הנוסף של האנדוזומים עשוי להיות שונה, חלקם עשויים לחזור אל פני התא ולהתמזג איתו, אך רובנכנס לתהליך של עיכול תוך תאי. אנדוזומים ראשוניים מכילים בעיקר מולקולות זרות הכלואות בתווך הנוזלי ואינם מכילים אנזימים הידרוליטיים. אנדוזומים יכולים להתמזג אחד עם השני תוך הגדלת גודלם. לאחר מכן הם מתמזגים עם ליזוזומים ראשוניים (ראה להלן), שמכניסים לחלל האנדוזום אנזימים המעבירים הידרוליזה של ביו-פולימרים שונים. פעולתם של הידרולאזות ליזוזומליות אלו גורמת לעיכול תוך תאי - פירוק פולימרים למונומרים.
כפי שכבר הוזכר, במהלך phagocytosis ו-pinocytosis, תאים מאבדים שטח גדול של הפלסמולה (ראה מקרופאגים), אשר, עם זאת, משוחזר במהירות במהלך מיחזור הממברנה, עקב החזרה של vacuoles ושילובם בפלסמולמה. זה נובע מהעובדה שלפוחיות קטנות יכולות להיפרד מאנדוזומים או וואקוולים, כמו גם מליזוזומים, אשר שוב מתמזגים עם קרום הפלזמה. עם מיחזור כזה, מתרחשת מעין העברת "מעבורת" של ממברנות: פלסמולמה - פינוזום - וואקוולה - פלסמולמה. זה מוביל לשיקום האזור המקורי של קרום הפלזמה. נמצא שעם החזר כזה, מיחזור ממברנה, כל החומר הנספג נשמר באנדוזום הנותר.
ספֵּצִיפִיאוֹ בתיווך קולטןלאנדוציטוזיס יש מספר הבדלים מבלתי ספציפיים. העיקר שנספגות מולקולות שלגביהן יש קולטנים ספציפיים על קרום הפלזמה שקשורים רק לסוג זה של מולקולות. לעתים קרובות נקראות מולקולות כאלה הנקשרות לחלבוני קולטן על פני התאים ליגנדים.
אנדוציטוזיס בתיווך קולטן תואר לראשונה בהצטברות חלבונים בביציות של עופות. חלבוני גרגירי חלמון, ויטלוגנינים, מסונתזים פנימה בדים שונים, אבל אז הם נכנסים לשחלות עם זרימת הדם, שם הם נקשרים לקולטני ממברנה מיוחדים של ביציות ואז, בעזרת אנדוציטוזיס, נכנסים לתא, שבו מתרחשת שקיעת גרגירי חלמון.
דוגמה נוספת לאנדוציטוזיס סלקטיבי היא הובלת כולסטרול לתוך התא. שומן זה מסונתז בכבד ובשילוב עם פוספוליפידים אחרים ומולקולת חלבון, יוצר את מה שנקרא. ליפופרוטאין בצפיפות נמוכה (LDL), המופרש על ידי הכבד ו מערכת דםמתפשט בכל הגוף (איור 140). קולטני ממברנת פלזמה ייעודיים הממוקמים בצורה מפוזרת על פני השטח תאים שונים, מזהים את מרכיב החלבון של LDL, ויוצרים קומפלקס קולטן-ליגנד ספציפי. בעקבות זאת, מתחם כזה עובר לאזור הבורות הגובלים ומופנם - מוקף בממברנה וטבול עמוק לתוך הציטופלזמה. הוכח כי קולטנים מוטנטים יכולים לקשור ל-LDL, אך אינם מצטברים באזור הבורות הגבולים. בנוסף לקולטני LDL, נמצאו יותר משני תריסר חומרים נוספים המעורבים באנדוציטוזיס קולטן של חומרים שונים, שכולם משתמשים באותו מסלול הפנמה דרך הבורות הגובלים. כנראה, תפקידם הוא בהצטברות של קולטנים: בור אחד ויחיד יכול לאסוף כ-1000 קולטנים ממעמדות שונים. עם זאת, בפיברובלסטים, צבירי קולטני LDL ממוקמים באזור הבורות הגבולים גם בהיעדר ליגנד במדיום.
גורלו הנוסף של חלקיק ה-LDL הנספג הוא שהוא עובר ריקבון בהרכב ליזוזום משני. לאחר טבילה בציטופלזמה של שלפוחית גבול עמוסה ב-LDL, יש אובדן מהיר של שכבת הקלתרין, שלפוחיות הממברנה מתחילות להתמזג זו בזו, ויוצרות אנדוזום - וואקוולה המכילה חלקיקי LDL נספגים שעדיין קשורים לקולטנים על פני הממברנה. . לאחר מכן מתרחשת ההתנתקות של קומפלקס הליגנד-קולטן, ואקואולים קטנים מתפצלים מהאנדוזום, שהממברנות שלו מכילות קולטנים חופשיים. שלפוחיות אלו ממוחזרות, משולבות בממברנת הפלזמה, וכך חוזרים הקולטנים אל פני התא. גורלו של LDL הוא שלאחר איחוי עם ליזוזומים, הם עוברים הידרוליזה לכולסטרול חופשי, אותו ניתן לשלב בקרום התא.
אנדוזומים מאופיינים בערך pH נמוך יותר (pH 4-5), סביבה חומצית יותר מאשר ואקואולים אחרים של תאים. הסיבה לכך היא נוכחות של חלבונים בממברנות שלהם. משאבת פרוטון, שאיבת יוני מימן עם צריכה בו-זמנית של ATP (ATPase תלוי H+). הסביבה החומצית בתוך האנדוזומים ממלאת תפקיד קריטי בניתוק הקולטנים והליגנדים. חוץ מזה, סביבה חומציתהוא אופטימלי להפעלה של אנזימים הידרוליטים בהרכב של ליזוזומים, המופעלים עם היתוך של ליזוזומים עם אנדוזומים ומובילים להיווצרות אנדוליזומים, שבו מתרחש פיצול של ביו-פולימרים נספגים.
במקרים מסוימים, גורלם של ליגנדים מפורקים אינו קשור להידרוליזה ליזוזומלית. אז בתאים מסוימים, לאחר התקשרות של קולטני פלסמולמה לחלבונים מסוימים, ואקואולים מצופים קלתרין שוקעים לתוך הציטופלזמה ומועברים לאזור אחר של התא, שם הם מתמזגים שוב עם קרום הפלזמה, והחלבונים הקשורים מתנתקים מהתאים. קולטנים. כך מתבצעת העברה, טרנסציטוזיס, של חלק מהחלבונים דרך דופן תא האנדותל מפלסמת הדם אל הסביבה הבין-תאית (איור 141). דוגמה נוספת לטרנסציטוזיס היא העברת נוגדנים. אז ביונקים, הנוגדנים של האם יכולים להיות מועברים לגור דרך חלב. במקרה זה, קומפלקס הקולטן-נוגדנים נותר ללא שינוי באנדוזום.
פגוציטוזיס
כפי שכבר הוזכר, phagocytosis הוא וריאנט של אנדוציטוזיס וקשור לספיגה על ידי התא של אגרגטים גדולים של מקרומולקולות עד לתאים חיים או מתים. כמו גם פינוציטוזיס, פגוציטוזיס יכול להיות לא ספציפי (לדוגמה, ספיגת חלקיקי זהב קולואידי או פולימר דקסטרן על ידי פיברובלסטים או מקרופאגים) וספציפית, בתיווך קולטנים על פני ממברנת הפלזמה של תאים פגוציטים. במהלך פגוציטוזיס, נוצרים ואקואולים אנדוציטיים גדולים - פאגוזום, אשר לאחר מכן מתמזגים עם ליזוזומים ליצירת פאגוליזוזומים.
על פני התאים המסוגלים לפגוציטוזיס (ביונקים, אלה נויטרופילים ומקרופאגים), ישנה קבוצה של קולטנים המקיימים אינטראקציה עם חלבוני ליגנד. אז ב זיהומים חיידקייםנוגדנים לחלבונים חיידקיים נקשרים למשטחים תאים חיידקיים, ויוצרים שכבה שבה אזור ה-F c של הנוגדנים מסתכל החוצה. שכבה זו מזוהה על ידי קולטנים ספציפיים על פני המקרופאגים והנויטרופילים, ובאתרי הקישור שלהם מתחילה ספיגת החיידק במעטפתו בקרום הפלזמה של התא (איור 142).
אקסוציטוזיס
קרום הפלזמה מעורב בסילוק חומרים מהתא בעזרת אקסוציטוזיס- התהליך ההפוך של אנדוציטוזיס (ראה איור 133).
במקרה של אקסוציטוזיס, מוצרים תוך תאיים סגורים בוואקווולים או שלפוחיות ומופרדים מההיאלופלזמה על ידי ממברנה מתקרבים אל קרום הפלזמה. בנקודות המגע שלהם, קרום הפלזמה וממברנת הוואקואול מתמזגים, והבועה מתרוקנת אל הסביבה. בעזרת אקסוציטוזיס מתרחש תהליך המיחזור של ממברנות המעורבות באנדוציטוזיס.
אקוציטוזה קשורה לשחרור של חומרים שונים המסונתזים בתא. הפרשת, שחרור חומרים לסביבה, תאים יכולים לייצר ולשחרר תרכובות במשקל מולקולרי נמוך (אצטילכולין, אמינים ביוגניים וכו'), כמו גם, ברוב המקרים, מקרומולקולות (פפטידים, חלבונים, ליפופרוטאינים, פפטידוגליקנים ועוד). אקסוציטוזיס או הפרשה ברוב המקרים מתרחשת בתגובה לאות חיצוני (דחף עצבי, הורמונים, מתווכים וכו'). למרות שבמקרים מסוימים מתרחשת אקוציטוזיס ללא הרף (הפרשה של פיברונקטין וקולגן על ידי פיברובלסטים). באופן דומה, חלק מהפוליסכרידים (המיצלולוזות) המעורבים ביצירת דפנות התא מוסרים מהציטופלזמה של תאי הצמח.
רוב החומרים המופרשים משמשים תאים אחרים של אורגניזמים רב-תאיים (הפרשת חלב, מיצי עיכול, הורמונים וכו'). אבל לעתים קרובות תאים מפרישים חומרים לצרכיהם. כך למשל, הצמיחה של קרום הפלזמה מתבצעת על ידי הטבעת קטעים של הממברנה כחלק מ-vacuole exocytic, חלק מיסודות הגליקוקליקס מופרשים על ידי התא בצורה של מולקולות גליקופרוטאין וכו'.
אנזימים הידרוליטים המבודדים מתאי על ידי אקסוציטוזיס יכולים להיספג בשכבת הגליקוקאליקס ולספק חישוף חוץ-תאי הקשור לממברנה של ביו-פולימרים שונים ומולקולות אורגניות. לעיכול לא תאי ממברנה יש חשיבות רבה עבור בעלי חיים. נמצא כי ב אפיתל מעינמצא יונקים באזור מה שנקרא גבול המברשת של אפיתל היניקה, העשיר במיוחד בגליקוקליקס. כמות גדולהמגוון אנזימים. חלק מהאנזימים הללו הם ממקור לבלב (עמילאז, ליפאז, פרוטאנזים שונים וכו'), וחלקם מופרשים על ידי תאי האפיתל עצמם (אקסוהידרולאזים, המפרקים בעיקר אוליגומרים ודימרים תוך יצירת מוצרים מובלים).
תפקיד הקולטן של הפלזמה
כבר נפגשנו עם תכונה זו של קרום הפלזמה בעת היכרות עם פונקציות התחבורה שלה. חלבוני נשאים ומשאבות הם גם קולטנים המזהים ומקיימים אינטראקציה עם יונים מסוימים. חלבוני קולטנים נקשרים לליגנדים ומשתתפים בבחירת המולקולות הנכנסות לתאים.
חלבוני ממברנה או אלמנטים גליקוקליקסים - גליקופרוטאין יכולים לפעול כקולטנים כאלה על פני התא. אתרים רגישים כאלה לחומרים בודדים יכולים להיות מפוזרים על פני התא או לאסוף באזורים קטנים.
לתאים שונים של אורגניזמים של בעלי חיים עשויים להיות סטים שונים של קולטנים או רגישות שונה של אותו קולטן.
תפקידם של קולטנים תאיים רבים אינו רק הקישור של חומרים ספציפיים או היכולת להגיב אליהם גורמים פיזייםאלא גם בהעברת אותות בין-תאיים מהמשטח לתוך התא. נכון להיום, מערכת העברת האותות לתאים בעזרת הורמונים מסוימים, הכוללים שרשראות פפטידים, נחקרה היטב. הורמונים אלו נמצאו נקשרים לקולטנים ספציפיים על פני קרום הפלזמה של התא. קולטנים, לאחר התקשרות להורמון, מפעילים חלבון נוסף, שנמצא כבר בחלק הציטופלזמי של קרום הפלזמה, אדנילט ציקלאז. אנזים זה מסנתז את מולקולת ה-AMP המחזורית מ-ATP. תפקידו של AMP מחזורי (cAMP) הוא בכך שהוא שליח משני – מפעיל של אנזימים – קינאזות הגורמים לשינויים בחלבוני אנזים אחרים. לכן, כאשר הורמון הלבלב גלוקגון, המיוצר על ידי תאי A של האיים של לנגרהנס, פועל על תא הכבד, ההורמון נקשר לקולטן ספציפי, הממריץ את ההפעלה של אדנילט ציקלאז. cAMP מסונתז מפעיל חלבון קינאז A, אשר בתורו מפעיל מפל של אנזימים שבסופו של דבר מפרקים גליקוגן (פוליסכריד אחסון בעלי חיים) לגלוקוז. פעולת האינסולין הפוכה - הוא ממריץ את כניסת הגלוקוז לתאי הכבד ואת שקיעתו בצורת גליקוגן.
באופן כללי, שרשרת האירועים מתפתחת באופן הבא: ההורמון יוצר אינטראקציה ספציפית עם החלק הקולטני של מערכת זו, ומבלי לחדור לתא, מפעיל את adenylate cyclase, אשר מסנתז cAMP, אשר מפעיל או מעכב אנזים תוך תאי או קבוצת אנזימים. . לפיכך, הפקודה, האות מממברנת הפלזמה מועבר בתוך התא. היעילות של מערכת אדנילט ציקלאז זו גבוהה מאוד. לפיכך, האינטראקציה של מולקולת הורמון אחת או כמה יכולה להוביל, עקב סינתזה של מולקולות cAMP רבות, להגברת אותות אלפי פעמים. במקרה זה, מערכת ה-adenylate cyclase משמשת כממיר של אותות חיצוניים.
יש עוד דרך שבה משתמשים בשליחים משניים אחרים - זה מה שנקרא. מסלול phosphatidylinositol. תחת פעולת אות מתאים (חלק מתווכי עצבים וחלבונים), מופעל האנזים פוספוליפאז C, המבקע את הפוספוליפיד פוספאטידילינוזיטול דיפוספט, שהוא חלק מממברנת הפלזמה. תוצרי ההידרוליזה של שומנים זה מפעילים מחד גיסא חלבון קינאז C אשר מפעיל את מפל הקינאז אשר מוביל לתגובות תאיות מסוימות ומאידך גיסא מוביל לשחרור יוני סידן המווסתים שורה שלמהתהליכים תאיים.
דוגמה נוספת לפעילות הקולטנים היא הקולטנים לאצטילכולין, מוליך עצבי חשוב. אצטילכולין משתחרר מ סוף עצבים, נקשר לרצפטור שעל סיב השריר, גורם לזרימה אימפולסיבית של Na + לתוך התא (דפולריזציה של הממברנה), פתיחה מיידית של כ-2000 תעלות יונים באזור הסיום הנוירו-שרירי.
הגיוון והספציפיות של קבוצות הקולטנים על פני התאים מובילים ליצירת מערכת סמנים מורכבת מאוד המאפשרת להבחין בין התאים של האדם עצמו (מאותו פרט או מאותו מין) מאלה של אחרים. תאים דומים נכנסים לאינטראקציות זה עם זה, מה שמוביל להידבקות של משטחים (צימוד בפרוטוזואה ובחיידקים, היווצרות קומפלקסים של תאי רקמה). במקרה זה, תאים הנבדלים במערך הסמנים הקובעים או שאינם תופסים אותם אינם נכללים מאינטראקציה כזו, או נהרסים בבעלי חיים גבוהים יותר כתוצאה מתגובות אימונולוגיות (ראה להלן).
קרום הפלזמה קשור ללוקליזציה של קולטנים ספציפיים המגיבים לגורמים פיזיקליים. לכן, בקרום הפלזמה או בנגזרותיו בחיידקים פוטוסינתטיים ובאצות כחולות-ירוקות, חלבוני קולטן (כלורופילים) המקיימים אינטראקציה עם קוונטות האור הינם מקומיים. בממברנת הפלזמה של תאי בעלי חיים רגישים לאור קיימת מערכת מיוחדת של חלבוני פוטורצפטורים (רודופסין), שבעזרתם הופך אות האור לכימי, אשר בתורו מביא ליצירת דחף חשמלי.
זיהוי בין תאי
באורגניזמים רב-תאיים, עקב אינטראקציות בין-תאיות, נוצרים הרכבים תאיים מורכבים, שניתן לבצע את תחזוקתם דרכים שונות. ברקמות נבטיות, עובריות, במיוחד על שלבים מוקדמיםהתפתחות, תאים נשארים מחוברים זה לזה בגלל היכולת של המשטחים שלהם להיצמד זה לזה. הנכס הזה הַדבָּקָה(חיבור, הידבקות) של תאים ניתן לקבוע על ידי המאפיינים של פני השטח שלהם, אשר אינטראקציה ספציפית אחד עם השני. המנגנון של קשרים אלה נחקר היטב, הוא מסופק על ידי האינטראקציה בין גליקופרוטאין של ממברנות פלזמה. עם אינטראקציה בין-תאית כזו של תאים בין ממברנות פלזמה, תמיד נשאר פער ברוחב של כ-20 ננומטר, מלא ב-glycocalyx. טיפול ברקמה באמצעות אנזימים המפרים את שלמות הגליקוקאליקס (ריריות הפועלות בצורה הידרוליטית על מוצינים, מוקופוליסכרידים) או פגיעה בממברנת הפלזמה (פרוטאזות) מובילים לבידוד התאים זה מזה, להתנתקותם. עם זאת, אם גורם הדיסוציאציה מוסר, התאים יכולים להרכיב מחדש ולהצטבר מחדש. אז אפשר לנתק תאים של ספוגים בצבעים שונים, כתום וצהוב. התברר כי בתערובת התאים הללו נוצרים שני סוגי אגרגטים: אלו המורכבים מצהובים בלבד ורק מתאי כתומים. במקרה זה, השעיות תאים מעורבות מתארגנות בעצמן, ומשחזרות את המבנה הרב-תאי המקורי. תוצאות דומות התקבלו עם השעיות תאים מופרדות של עוברי דו-חיים; במקרה זה, קיימת הפרדה מרחבית סלקטיבית של תאי אקטודרם מהאנדודרם ומהמזנכימה. יתר על כן, אם משתמשים ברקמות לצבירה מחדש שלבים מאוחריםהתפתחות של עוברים, ואז הרכבי תאים שונים עם סגוליות רקמה ואיברים מתאספים באופן עצמאי במבחנה, נוצרים אגרגטים של אפיתל, בדומה ל צינוריות כליה, וכו.
נמצא שגליקופרוטאינים טרנסממברניים אחראים להצטברות של תאים הומוגניים. ישירות לחיבור, הידבקות, תאים אחראים על המולקולות של מה שנקרא. חלבוני CAM (מולקולות הדבקה בתאים). חלקם מחברים תאים זה עם זה עקב אינטראקציות בין-מולקולריות, אחרים יוצרים קשרים או מגעים בין-תאיים מיוחדים.
אינטראקציות בין חלבונים דביקים יכולים להיות הומופיליכאשר תאים שכנים נקשרים זה לזה בעזרת מולקולות הומוגניות, הטרופיליכאשר סוגים שונים של CAMs על תאים שכנים מעורבים בהדבקה. קישור בין תאי מתרחש דרך מולקולות קישור נוספות.
ישנן מספר סוגים של חלבוני CAM. אלו הם קדהרינים, N-CAM דמויי אימונוגלובולינים (מולקולות הדבקה של תאי עצב), סלקטינים, אינטגרינים.
קדהריניםהם חלבוני קרום פיברילרי אינטגרלי היוצרים הומודימרים מקבילים. תחומים נפרדים של חלבונים אלו קשורים ליוני Ca 2+, מה שמקנה להם קשיחות מסוימת. ישנם יותר מ-40 מינים של קדרינים. לפיכך, E-cadherin אופייני לתאים של עוברים מושתלים מראש ותאי אפיתל של אורגניזמים בוגרים. P-cadherin אופייני לתאי טרופובלסט, שליה ואפידרמיס; N-cadherin ממוקם על פני השטח של תאי עצב, תאי עדשה ועל שרירי הלב והשלד.
^ מולקולות הידבקות של תאי עצב (N-CAM) שייכים למשפחת העל של האימונוגלובולינים, הם יוצרים קשרים בין תאי עצב. חלק מה-N-CAMs מעורבים בחיבור של סינפסות כמו גם היצמדות תאים. מערכת החיסון.
selectinsכמו כן, חלבונים אינטגרליים של קרום הפלזמה מעורבים בהדבקה של תאי אנדותל, בקשירה של טסיות דם, לויקוציטים.
אינטגריניםהם הטרודימרים, עם שרשראות ו- . אינטגרינים מחברים בעיקר תאים עם מצעים חוץ-תאיים, אך הם יכולים גם להשתתף בהיצמדות תאים זה לזה.
זיהוי של חלבונים זרים
כפי שכבר הוזכר, תגובה מורכבת מתפתחת על מקרומולקולות (אנטיגנים) זרות שנכנסו לגוף - תגובה חיסונית. מהותו נעוצה בעובדה שחלק מהלימפוציטים מייצרים חלבונים מיוחדים - נוגדנים הנקשרים באופן ספציפי לאנטיגנים. לדוגמה, מקרופאגים מזהים קומפלקסים של אנטיגן-נוגדנים עם קולטני פני השטח שלהם וסופגים אותם (לדוגמה, ספיגת חיידקים במהלך phagocytosis).
בגוף של כל בעלי החולייתנים, בנוסף, קיימת מערכת של קליטה של תאים זרים או משלהם, אך עם חלבונים ממברנת פלזמה שהשתנו, למשל, כאשר זיהום ויראליאו עם מוטציות, הקשורות לרוב לניוון גידולי של תאים.
חלבונים ממוקמים על פני השטח של כל תאי החולייתנים, מה שנקרא. תסביך היסטורי תאימות גדול(תסביך היסטו-תאימות מרכזי - MHC). אלו הם חלבונים אינטגרליים, גליקופרוטאינים, הטרודמרים. חשוב מאוד לזכור שלכל אדם יש סט שונה של חלבוני MHC אלו. זה נובע מהעובדה שהם מאוד פולימורפיים, כי לכל פרט יש מספר רב של צורות מתחלפות של אותו גן (יותר מ-100), בנוסף, ישנם 7-8 לוקוסים המקודדים למולקולות MHC. התוצאה היא בכל תא אורגניזם נתון, בעל קבוצה של חלבוני MHC, יהיו שונים מתאי פרט מאותו מין. צורה מיוחדתלימפוציטים, T-לימפוציטים, מזהים את ה-MHC של גופם, אבל השינוי הקטן ביותרבמבנה ה-MHC (לדוגמה, קשר עם וירוס, או תוצאה של מוטציה בתאים בודדים), מוביל לכך שלימפוציטים מסוג T מזהים תאים שהשתנו כאלה והורסים אותם, אך לא על ידי פגוציטוזיס. הם מפרישים חלבוני פרפורין ספציפיים מ-vacuoles הפרשה, המוטבעים בקרום הציטופלזמי של התא המשתנה, יוצרים בו תעלות טרנס-ממברניות, מה שהופך את קרום הפלזמה לחדיר, מה שמוביל למוות של התא שהשתנה (איור 143, 144).
קשרים בין תאיים מיוחדים
בנוסף לקשרים הדביקים הפשוטים יחסית (אך הספציפיים) הללו (איור 145), ישנם מספר מבנים בין-תאיים מיוחדים, מגעים או חיבורים המבצעים פונקציות מסוימות. מדובר בחיבורי נעילה, עיגון ותקשורת (איור 146).
נְעִילָהאוֹ חיבור הדוקמאפיין אפיתל חד-שכבתי. זהו האזור שבו השכבות החיצוניות של שני ממברנות הפלזמה קרובות ככל האפשר. הממברנה התלת-שכבתית נראית לעתים קרובות במגע זה: שתי השכבות האוסמופיליות החיצוניות של שתי הממברנות נראות מתמזגות לשכבה אחת משותפת בעובי 2-3 ננומטר. היתוך הממברנות אינו מתרחש על פני כל אזור המגע ההדוק, אלא הוא סדרה של התכנסות נקודתית של ממברנות (איור 147a, 148).
בהכנות מישוריות של שברי קרום פלזמה באזור המגע ההדוק, בשיטת ההקפאה והשבץ, נמצא כי נקודות המגע של הממברנות הן שורות של כדוריות. אלו הם החלבונים אוקלודין וקלאודין, חלבונים אינטגרליים מיוחדים של קרום הפלזמה, הבנויים בשורות. שורות כאלה של כדוריות או רצועות יכולות להצטלב בצורה כזו שהן יוצרות, כביכול, סריג או רשת על פני המחשוף. מבנה זה אופייני מאוד לאפיתליה, במיוחד בלוטות ומעי. במקרה האחרון, מגע חזק יוצר אזור רציף של איחוי של קרומי פלזמה, המקיף את התא בחלקו האפיקלי (העליון, מסתכל לתוך לומן המעי) (איור 148). לפיכך, כל תא של השכבה מוקף, כביכול, בסרט של מגע זה. מבנים כאלה ניתן לראות גם עם כתמים מיוחדים במיקרוסקופ אור. הם קיבלו את השם ממורפולוגים צלחות קצה. התברר שבמקרה זה תפקידו של המגע ההדוק הסוגר אינו רק בחיבור מכני של תאים זה עם זה. אזור מגע זה אינו חדיר בצורה גרועה למקרומולקולות ויונים, וכך הוא נועל, חוסם את החללים הבין-תאיים, מבודד אותם (ועמם את הסביבה הפנימית של הגוף) מהסביבה החיצונית (במקרה זה, לומן המעי).
ניתן להדגים זאת באמצעות ניגודים צפופים באלקטרונים כגון תמיסת לנטנום הידרוקסיד. אם לומן המעי או הצינור של בלוטה כלשהי מלאים בתמיסה של לנטנום הידרוקסיד, אז על מקטעים תחת מיקרוסקופ אלקטרונים, לאזורים שבהם נמצא חומר זה יש צפיפות אלקטרונים גבוהה והם יהיו כהים. התברר שגם אזור המגע ההדוק וגם החללים הבין-תאיים שמתחתיו לא מתכהים. אם הצמתים ההדוקים נפגעים (על ידי טיפול אנזימטי קל או הסרה של יוני Ca++), אז הלנתנום חודר גם לאזורים הבין-תאיים. באופן דומה, צמתים הדוקים הוכחו כבלתי חדירים להמוגלובין ולפריטין בצינוריות הכליות.
לפיכך, צמתים הדוקים הם מחסומים לא רק עבור מקרומולקולות, אלא גם אטומים לנוזלים ויונים.
מגע סגירה, או הדוק, מתרחש בין כל סוגי האפיתל חד-שכבתי (אנדותל, מזותל, אפנדימה).
עֲגִינָהאוֹ חיבורים שלוביםאו אנשי קשר נקראים כך מכיוון שהם מחברים לא רק את ממברנות הפלזמה של תאים שכנים, אלא גם נקשרים לאלמנטים הפיברילריים של השלד הציטוניים (איור 149). סוג זה של תרכובות מאופיין בנוכחות של שני סוגים של חלבונים. אחד מהם הוא חלבוני מקשר (קושר) טרנסממברני, המעורבים או בחיבור הבין-תאי בפועל או בחיבור של הפלסמולמה עם מרכיבי המטריצה החוץ-תאית (ממברנה בסיסית של אפיתליה, חלבונים מבניים חוץ-תאיים של רקמת חיבור).
השני הוא חלבונים תוך תאיים המחברים או מעגנים את הסיפונים הציטופלזמיים של השלד הציטופלזמי מאחורי מרכיבי הממברנה של מגע כזה.
חיבורי עיגון כוללים חיבורי נקודות עיגון בין-תאיים, פסי עיגון, חיבורים מוקדים או לוחות עיגון, כולם נקשרים תוך תאיים למיקרופילמנטים של אקטין.
קבוצה נוספת של צמתים בין-תאיים מעגנים, דסמוזומים והמידסמוזומים, נקשרות לאלמנטים אחרים של השלד הציטוניים, כלומר לחוטי ביניים.
חיבורים נקודתיים בין-תאיים נמצאו ברקמות רבות שאינן אפיתל, אך המבנה תואר בצורה ברורה יותר. סרטים מיוחדים (דביקים).באפיתל חד-שכבתי (איור 150). מבנה זה מקיף את כל היקף תא האפיתל, בדומה למה שקורה במקרה של צומת הדוק. לרוב, חגורה או סרט כזה נמצאים מתחת לחיבור הדוק (ראה איור 146). במקום זה, ממברנות הפלזמה אינן מוכנסות זו לזו, אלא אפילו מתרחקות במקצת במרחק של 25-30 ננומטר, ונראה ביניהן אזור של צפיפות מוגברת. זהו לא יותר מאשר אתרי האינטראקציה של גליקופרוטאין טרנסממברני, הנצמדים זה לזה באופן ספציפי ומספקים חיבור מכני בין הממברנות של שני תאים שכנים. חלבוני קישור אלו שייכים ל-E-cadherins, חלבונים המספקים הכרה ספציפית של ממברנות הומוגניות על ידי תאים. הרס שכבת הגליקופרוטאין הזו מוביל לבידוד של תאים בודדים ולהרס של שכבת האפיתל. בצד הציטופלזמי ליד הממברנה, נראה הצטברות של חומר צפוף כלשהו, אליו צמודה שכבה של חוטים דקים (6-7 ננומטר) השוכנים לאורך קרום הפלזמה בצורת צרור העובר לאורך כל ההיקף של התא. חוטים דקים הם ספירלי אקטין; הם נקשרים לממברנת הפלזמה באמצעות חלבון הקטנין, היוצר שכבה צפופה ליד הממברנה.
המשמעות התפקודית של חיבור סרט כזה טמונה רק בהיצמדות מכנית של תאים זה לזה: עם התכווצות חוטי האקטין בסרט, צורת התא יכולה להשתנות. מאמינים כי התכווצות קואופרטיבית של ספירלי אקטין בכל תאי שכבת האפיתל יכולה לגרום לשינוי בגיאומטריה שלה, למשל, התקפלות לצינור, בדומה למה שמתרחש במהלך היווצרות הצינור העצבי בעוברי חוליות.
^ אנשי קשר מוקדים אוֹ לוחות הדבקהמתרחשים בתאים רבים ונחקרים היטב בפיברובלסטים. הם בנויים לפי תוכנית כלליתעם רצועות הדבקה, אך מתבטאים כאזורים קטנים - לוחות על הפלזמלמה. במקרה זה, חלבוני אינטגרין מקשר טרנסממברני נקשרים באופן ספציפי לחלבוני מטריקס חוץ-תאיים (לדוגמה, פיברונקטין) (איור 151). מהצד של הציטופלזמה, אותם גליקופרוטאין קשורים לחלבוני ממברנה, הכוללים גם וינקולין, אשר בתורו קשור לצרור חוטי אקטין. המשמעות התפקודית של מגעים מוקדיים טמונה הן בעיגון התא למבנים חוץ-תאיים והן ביצירת מנגנון המאפשר לתאים לנוע.
דסמוזומים, מבנים בצורת לוחות או כפתורים גם מחברים תאים זה לזה (איור 152, 153a). במרחב הבין-תאי נראית כאן גם שכבה צפופה המיוצגת על ידי קדהרינים ממברנה אינטגרלית - דסמגליינים, המקשרים תאים זה לזה. בצד הציטופלזמי, צמודה לפלסמולמה שכבה של חלבון desmoplakin, שאיתה קשורים חוטי הביניים של השלד הציטוניים. Desmosomes נמצאים לרוב באפיתליה, ובמקרה זה חוטי הביניים מכילים קרטין. בתאי שריר הלב, קרדיומיוציטים, מכילים סיבים של דסמין כחלק מדסמוזומים. באנדותל כלי הדם, דסמוזומים מכילים חוטי ביניים של וימנטין.
המידסמוזומים- באופן עקרוני, הם דומים במבנה לדסמוזום, אבל הם חיבור של תאים עם מבנים בין-תאיים. אז באפיתל, הגליקופרוטאינים המקשרים (אינטגרינים) של הדסמוזומים מקיימים אינטראקציה עם החלבונים של מה שנקרא. קרום בסיס, הכולל קולגן, למינין, פרוטאוגליקנים וכו'.
התפקיד הפונקציונלי של דסמוזומים והמידסמוזומים הוא מכני בלבד - הם דבקים בחוזקה תאים זה לזה ולמטריקס החוץ-תאי הבסיסי, מה שמאפשר לשכבות האפיתל לעמוד בעומסים מכניים כבדים. באופן דומה, דסמוזומים קושרים בחוזקה את תאי שריר הלב זה לזה, מה שמאפשר להם לבצע עומס מכני עצום תוך שהם נשארים קשורים למבנה מתכווץ אחד.
שלא כמו מגע הדוק, כל סוגי מגעי ההדבקה חדירים לתמיסות מימיות ואינם ממלאים תפקיד בהגבלת הדיפוזיה.
^ פער אנשי קשר נחשבים לחיבורי תקשורת של תאים; אלו המבנים המעורבים בהעברה ישירה חומרים כימייםמתא לתא, שיכול למלא תפקיד פיזיולוגי מרכזי לא רק בתפקודם של תאים מיוחדים, אלא גם לספק אינטראקציות בין-תאיות במהלך התפתחות האורגניזם, במהלך ההתמיינות של התאים שלו. מאפיין של סוג זה של מגעים הוא התכנסות של ממברנות הפלזמה של שני תאים שכנים במרחק של 2-3 ננומטר (איור 147b, 153b). זה הנסיבות האלה במשך זמן רבלא אפשרו להבחין בחתכים דקים במיוחד המין הזהמגע ממגע מבודד (סוגר) הדוק. בעת שימוש ב-lanthanum hydroxide, נצפה שחלק מהמגעים ההדוקים דולפים את הניגוד. במקרה זה, הלנתנום מילא פער דק ברוחב של כ-3 ננומטר בין ממברנות הפלזמה הסמוכות של תאים שכנים. זה היה מקור המונח - מגע פער. התקדמות נוספת בפענוח המבנה שלו הושגה בשיטת הקפאה-שבבים. התברר כי אזורי צומת פערים (מ-0.5 עד 5 מיקרומטר בגודל) על מחשופים של ממברנות מנוקדים בחלקיקים בקוטר 7-8 ננומטר המסודרים בצורה משושה בפרק זמן של 8-10 ננומטר, בעלי תעלה ברוחב של כ-2 ננומטר. מֶרְכָּז. חלקיקים אלה נקראים קשרים(איור 154). באזורים של מגע מרווח, יכולים להיות בין 10-20 לכמה אלפי קשרים, תלוי ב תכונות פונקציונליותתאים. קונקסונים בודדו באופן הכנה, הם מורכבים משש יחידות משנה של קונקטין, חלבון במשקל מולקולרי של כ-30 אלף. בשילוב זה עם זה יוצרים קונקסונים אגרגט גלילי - קונקסון, שבמרכזו יש תעלה. קונקסונים בודדים משובצים בקרום הפלזמה בצורה כזו שהם חודרים דרכו. קונקסון אחד על ממברנת הפלזמה של התא מנוגד בדיוק לקונקסון על קרום הפלזמה של התא השכן, כך שהתעלות של שני הקונקקסונים יוצרות יחידה אחת. קונקסונים ממלאים את התפקיד של תעלות בין-תאיות ישירות שדרכן יכולים להתפזר יונים וחומרים בעלי משקל מולקולרי נמוך מתא לתא. נמצא כי קונקסונים יכולים להיסגר, לשנות את קוטר התעלה הפנימית, ובכך להשתתף בוויסות ההובלה של מולקולות בין תאים.
המשמעות התפקודית של צמתים מרווחים הובנה במחקר של תאים ענקיים בלוטות הרוקדיפטרה. בשל גודלן, ניתן להכניס בקלות מיקרואלקטרודות לתאים כאלה על מנת לחקור את המוליכות החשמלית של הממברנות שלהם. התברר שאם מכניסים אלקטרודות לשני תאים סמוכים, ממברנות הפלזמה שלהם מפגינות התנגדות חשמלית נמוכה, וזרם זורם בין התאים. יתרה מכך, התברר שכאשר מוזרק צבע ניאון לתא אחד, התווית מתגלה במהירות בתאים שכנים. באמצעות פלואורכרומים שונים, נמצא על תאי תרבית רקמת יונקים כי חומרים בעלי משקל מולקולרי של לא יותר מ-1-1.5 אלף וגודל של לא יותר מ-1.5 ננומטר יכולים להיות מועברים דרך צומת פערים (בחרקים, חומרים בעלי משקל מולקולרי במשקל של עד 2,000). בין החומרים הללו היו יונים שונים, חומצות אמינו, נוקלאוטידים, סוכרים, ויטמינים, סטרואידים, הורמונים, cAMP. לא חלבונים ולא חומצות גרעין יכולים לעבור דרך צומת פער.
היכולת הזו של צמתים מרווחים לשמש מקום להובלה של תרכובות במשקל מולקולרי נמוך משמשת באלה מערכות תאים, שבו יש צורך בשידור מהיר של דחף חשמלי (גל עירור) מתא לתא ללא השתתפות של מתווך עצבי. כך, כל תאי השריר של שריר הלב של הלב מחוברים באמצעות חיבורים מרווחים (בנוסף, התאים שם מחוברים גם בצמתים דביקים) (איור 147ב). זה יוצר תנאי להפחתה סינכרונית של מספר עצום של תאים. עם צמיחת התרבות של תאי שריר לב עובריים (שריר הלב), חלק מהתאים בשכבה מתחילים להתכווץ באופן ספונטני ללא תלות זה בזה. תדר שונה, ורק לאחר היווצרותם של צמתים מרווחים ביניהם הם מתחילים לפעום באופן סינכרוני כשכבה אחת מתכווצת של תאים. באותו אופן מובטחת כיווץ מפרק של תאי שריר חלקים בדופן הרחם.
גאפ junctions יכולים לשרת את המטרה של שיתוף פעולה מטבולי בין תאים על ידי החלפת מולקולות שונות, הורמונים, cAMP או מטבוליטים. דוגמה לכך היא טיפוח משותף של תאים מוטנטיים תימידין קינאז עם תאים נורמליים: כאשר מתרחשים חיבורי פער בין סוגי תאים אלו, תאים מוטנטיים מקבלים תימידין טריפוספט מתאי נורמלי דרך חיבורי פער ויכולים להשתתף בסינתזת ה-DNA.
בעוברים מוקדמים של חוליות, החל משלב 8 התאים, רוב התאים מחוברים זה לזה על ידי חיבורים מרווחים. כאשר העובר מתמיין, חיבורי הרווח בין כל התאים נעלמים ונשארים רק בין קבוצות של תאים מיוחדים. לדוגמה, במהלך היווצרות הצינור העצבי, החיבור של התאים של מבנה זה עם שאר האפידרמיס מופרע, מנותק.
השלמות והתפקוד של צמתים מרווחים תלויים מאוד ברמת יוני Ca 2+ בתוך התא. בדרך כלל, ריכוז הסידן בציטופלזמה נמוך מאוד. אם Ca 2+ מוזרק לאחד מתאי שכבת תרבית הרקמה, אזי אין עלייה ברמת Ca 2+ בציטופלזמה בתאים שכנים; תאים, כביכול, מנותקים משכניהם, מפסיקים להתנהל חַשְׁמַלוצבעים. לאחר זמן מה, לאחר שהסידן המוכנס נצבר על ידי המיטוכונדריה, המבנה והתפקודים של צומת הפער משוחזרים. תכונה זו חשובה מאוד לשמירה על שלמות ותפעול כל שכבת התאים, שכן פגיעה באחד מהם אינה מועברת לשכנה דרך צמתים מרווחים, המפסיקים לפעול כערוצי דיפוזיה בין-תאיים.
^ מגע סינפטי (סינפסות). סוג זה של מגע אופייני עבור רקמת עצביםומתרחשת הן בין שני נוירונים והן בין נוירון לאלמנט אחר - קולטן או אפקטור (לדוגמה, סוף עצבי-שרירי). סינפסות הן אזורי מגע בין שני תאים המתמחים להעברה חד-כיוונית של עירור או עיכוב מאלמנט אחד למשנהו (איור 155). בעיקרון הסוג הזהעומס תפקודי, העברת דחפים יכולה להתבצע גם על ידי סוגים אחרים של מגע (לדוגמה, מגע מרווח בשריר הלב), עם זאת, בחיבור הסינפטי, יעילות גבוההבמימוש דחף עצבי. סינפסות נוצרות על תהליכים של תאי עצב - אלו הם החלקים הסופיים של דנדריטים ואקסונים. לסינפסות פנימיות יש בדרך כלל שלוחות בצורת אגס, פלאקים בסוף התהליך של תא העצב. הרחבה סופנית כזו של תהליך של אחד מתאי העצב יכולה ליצור קשר וליצור קשר סינפטי הן עם הגוף של תא עצב אחר והן עם התהליכים שלו. תהליכים היקפיים של תאי עצב (אקסונים) יוצרים מגעים ספציפיים עם תאי אפקטור או קולטן. לכן, סינפסה היא מבנה שנוצר בין אזורים של שני תאים (כמו גם דסמוזום). הממברנות של תאים אלו מופרדות על ידי חלל בין תאי - שסע סינפטי ברוחב של כ-20-30 ננומטר. לעתים קרובות בלומן של חריץ זה נראה חומר דק סיבים בניצב לממברנות. הממברנה באזור המגע הסינפטי של תא אחד נקראת פרה-סינפטית, השני, התופס את הדחף, נקרא פוסט-סינפטי. במיקרוסקופ אלקטרוני, שתי הממברנות נראות צפופות ועבות. ליד הממברנה הפרה-סינפטית, מתגלה מספר עצום של ואקוולים קטנים, שלפוחיות סינפטיות מלאות בנירוטרנסמיטורים. שלפוחיות סינפטיות בזמן מעבר הדחף העצבי מוציאות את תוכנן לתוך השסע הסינפטי. הממברנה הפוסט-סינפטית נראית לרוב עבה יותר ממברנות רגילות בשל הצטברות של סיבים דקים רבים מסביבו מהצד של הציטופלזמה.
Plasmodesmata. סוג זה של תקשורת בין-תאית נמצא בצמחים. Plasmodesmata הם תעלות ציטופלזמה צינוריות דקות המחברים בין שני תאים סמוכים. הקוטר של ערוצים אלה הוא בדרך כלל 20-40 ננומטר. הממברנה המגבילה את הערוצים הללו עוברת ישירות לתוך ממברנות הפלזמה של תאים שכנים. Plasmodesmata עוברים דרך דופן התא המפרידה בין התאים (איור 156, 157). לפיכך, בחלק מתאי הצמח, פלסמודסמטה מחברת את ההיאלופלזמה של תאים שכנים, כך שבאופן פורמלי אין הבחנה מוחלטת, הפרדה של גוף של תא אחד למשנהו, זה דווקא סינציטיום: איחוד של טריטוריות תאים רבות בעזרת ציטופלזמה גשרים. אלמנטים צינוריים של ממברנה יכולים לחדור לתוך הפלסמודסמטה, לחבר את בורות המים של הרשת האנדופלזמית של תאים שכנים. Plasmodesmata נוצרות במהלך חלוקת התא, כאשר דופן התא הראשוני נבנה (ראה להלן). בתאים שחולקו לאחרונה, מספר הפלסמודסמטה יכול להיות גדול מאוד (עד 1000 לתא), עם הזדקנות התאים מספרם יורד עקב קרעים בעובי הולך וגדל. דופן תא.
התפקיד הפונקציונלי של plasmodesmata גדול מאוד: בעזרתם, זרימה בין-תאית של תמיסות המכילות חומרים מזינים, יונים ותרכובות אחרות. טיפות שומנים יכולות לנוע לאורך פלסמודסמטה תאים נגועים בנגיפים צמחיים דרך פלסמודסמטה. עם זאת, ניסויים מראים שתחבורה חופשית דרך פלסמודסמטה מוגבלת לחלקיקים בעלי מסה של לא יותר מ-800 דלטון.
> פונקציות של קרום הפלזמה. מנגנוני הובלה של חומרים דרך הפלזמה. תפקוד הקולטן של הפלזמה
קרום הפלזמה מבצע מספר פונקציות חשובות:
1) מחסום.תפקיד המחסום של קרום הפלזמה הוא להגביל את הדיפוזיה החופשית של חומרים מתא לתא, כדי למנוע דליפה של תוכן מסיס במים של התא. אך מאחר והתא חייב לקבל את חומרי ההזנה הדרושים, לשחרר את תוצרי חילוף החומרים הסופיים ולווסת את ריכוזי היונים התוך-תאיים, נוצרו בו מנגנונים מיוחדים להעברת חומרים דרך קרום התא.
2) הובלה.פונקציית ההובלה היא הבטחת כניסה ויציאה של חומרים שונים לתא ומחוצה לו. נכס חשובממברנות - החדירות הסלקטיבית, או חדירות למחצה.הוא מעביר בקלות מים וגזים מסיסים במים ודוחה מולקולות קוטביות כמו גלוקוז או חומצות אמינו.
ישנם מספר מנגנונים להובלת חומרים על פני הממברנה:
הובלה פסיבית;
מעבר פעיל;
הובלה באריזת ממברנה.
הובלה פסיבית.ריכוך -זוהי תנועה של חלקיקי התווך, המובילה להעברת חומר מאזור שבו ריכוזו גבוה לאזור עם ריכוז נמוך. במהלך הובלת דיפוזיה, הממברנה מתפקדת כמחסום אוסמוטי. קצב הדיפוזיה תלוי בגודל המולקולות ובמסיסותן היחסית בשומנים. אֵיך גדלים קטנים יותרמולקולות וככל שהן מסיסות יותר בשומן (ליפופיליות), כך הן יעברו מהר יותר דרך דו-שכבת השומנים. דיפוזיה יכולה להיות ניטראלי(העברת מולקולות לא טעונות) ו קל(בעזרת חלבוני נשא מיוחדים). דיפוזיה קלה מהירה יותר מדיפוזיה ניטרלית. למים יש את עוצמת החדירה המקסימלית, מכיוון שהמולקולות שלהם קטנות ולא טעונות. דיפוזיה של מים על פני קרום התא נקראת סְפִיגָה.ההנחה היא שבממברנת התא קיימות "נקבוביות" מיוחדות לחדירת מים וכמה יונים. מספרם קטן, והקוטר הוא בערך 0.3-0.8 ננומטר. מולקולות מסיסות בקלות בשכבת הדו-שומנים, כגון O, ומולקולות קוטביות לא טעונות בקוטר קטן (CO, אוריאה) מתפזרות במהירות הגבוהה ביותר דרך הממברנה.
העברת מולקולות קוטביות (סוכרים, חומצות אמינו) המתבצעת בעזרת חלבונים מיוחדים להובלת ממברנה נקראת הקלה על דיפוזיה.חלבונים כאלה נמצאים בכל סוגי הממברנות הביולוגיות, וכל חלבון ספציפי נועד לשאת מולקולות ממעמד מסוים. חלבוני התחבורה הם טרנסממברניים; שרשרת הפוליפפטיד שלהם חוצה את דו-שכבת השומנים מספר פעמים, ונוצרת דרך מעברים בה. זה מבטיח העברת חומרים ספציפיים דרך הממברנה ללא מגע ישיר איתה. ישנם שני סוגים עיקריים של חלבוני תחבורה: חלבונים נשאים (טרנספורטרים)ו יצירת ערוציםחלבונים (תעלות חלבון). חלבוני נשא נושאים מולקולות על פני הממברנה על ידי שינוי תצורה תחילה. חלבונים יוצרי תעלה יוצרים נקבוביות מלאות מים בממברנה. כאשר הנקבוביות פתוחות, עוברות דרכן מולקולות של חומרים ספציפיים (בדרך כלל יונים אנאורגניים בגודל ובמטען הנכון). אם למולקולה של החומר המועבר אין מטען, אזי כיוון ההובלה נקבע על ידי שיפוע הריכוז. אם המולקולה טעונה, אז ההובלה שלה, בנוסף לשיפוע הריכוז, מושפעת גם מהמטען החשמלי של הממברנה (פוטנציאל הממברנה). צד פנימיהפלזמלמה בדרך כלל טעונה שלילי ביחס לזה החיצוני. פוטנציאל הממברנה מקל על חדירת יונים בעלי מטען חיובי לתא ומונע מעבר של יונים בעלי מטען שלילי.
מעבר פעיל.הובלה פעילה היא תנועה של חומרים כנגד שיפוע אלקטרוכימי. זה תמיד מתבצע על ידי חלבוני טרנספורטר והוא קשור קשר הדוק למקור אנרגיה. לחלבוני הנשא יש אתרי קישור עם החומר המועבר. ככל שיותר אתרים כאלה קשורים לחומר, כך קצב ההובלה גבוה יותר. העברה סלקטיבית של חומר אחד נקראת יוניפורט.העברת מספר חומרים מתבצעת מערכות תחבורה משותפות.אם ההעברה הולכת בכיוון אחד, היא כן סימן,אם מול נגד נמל.לדוגמה, גלוקוז מועבר מהנוזל החוץ-תאי לתוך התא בצורה חד-פורטלית. העברת הגלוקוז וה-Na 4 מחלל המעי או האבובות של הכליות, בהתאמה, לתאי המעי או הדם מתבצעת באופן סימפורטלי, וההעברה של C1 ~ ו- HCO "היא אנטי-פורט. .
דוגמה לחלבון נשא שמשתמש באנרגיה המשתחררת במהלך הידרוליזה של ATP כדי להעביר חומרים היא לא + -ל + לִשְׁאוֹב,נמצא בממברנת הפלזמה של כל התאים. משאבת Na + -K פועלת על עיקרון האנטי-פורט, שואבת Na "מהתא ו-K t לתא כנגד השיפועים האלקטרוכימיים שלהם. שיפוע ה-Na + יוצר לחץ אוסמוטי, שומר על נפח התא ומבטיח הובלה של סוכרים וחומצות אמינו ב נוכחות של Na + ו-ATP, תחת פעולת ATPase, פוספט סופני מופרד מ-ATP ומחובר לשאריות חומצה אספרטיתעל מולקולת ATPase. מולקולת ATPase עוברת זרחון, משנה את תצורתה, ו-Na+ מופרש מהתא. לאחר הסרת ה-Na מהתא, K "מועבר תמיד לתוך התא. לשם כך, הפוספט המחובר בעבר מבוקע מ-ATPase בנוכחות K. האנזים עובר דה-פוספורילציה, משחזר את תצורתו, ו-K 1 הוא " נשאב" לתוך התא.
ATPase נוצר על ידי שתי תת יחידות, גדולות וקטנות. תת-היחידה הגדולה מורכבת מאלפי שאריות חומצות אמינו שחוצות את הדו-שכבה מספר פעמים. יש לו פעילות קטליטית והוא יכול להיות פוספורילציה ודה-פוספורילציה הפיכה. ליחידת המשנה הגדולה בצד הציטופלזמי יש אתרים לקשירת Na+ ו-ATP, וכן הלאה בחוץ-אתרים לקישור K + ו-ouabain. תת-היחידה הקטנה היא גליקופרוטאין ותפקידה אינו ידוע עדיין.
למשאבת Na + -K יש אפקט אלקטרוגני. הוא מוציא מהתא שלושה יוני Na f טעונים חיובית ומכניס לתוכו שני יוני K. כתוצאה מכך זורם זרם דרך הממברנה ויוצר פוטנציאל חשמלי בעל ערך שלילי בחלק הפנימי של התא ביחס לפני השטח החיצוניים שלו. . משאבת Na "-K + מווסתת את נפח התא, שולטת בריכוז החומרים בתוך התא, שומרת על לחץ אוסמוטי ומשתתפת ביצירת פוטנציאל הממברנה.
הובלה באריזת ממברנה. העברת מקרומולקולות (חלבונים, חומצות גרעין, פוליסכרידים, ליפופרוטאינים) וחלקיקים אחרים דרך הממברנה מתבצעת באמצעות היווצרות והיתוך עוקבים של שלפוחיות (שלפוחיות) המוקפות בממברנה. תהליך הובלה שלפוחית מתרחש בשני שלבים. בתחילה, קרום השלפוחית והפלזמה נדבקים זה לזה ואז מתמזגים. במהלך שלב 2, יש צורך לעקור מולקולות מים על ידי דו-שכבות שומנים באינטראקציה, המתקרבות זו לזו עד למרחק של 1-5 ננומטר. הוא האמין כי תהליך זה מופעל על ידי מיוחד חלבוני היתוך(הם בודדו עד כה רק בווירוסים). הובלה שלפוחית יש תכונה חשובה- מקרומולקולות נספגות או מופרשות בשלפוחיות בדרך כלל אינן מתערבבות עם מקרומולקולות או אברונים אחרים של התא. בועות יכולות להתמזג עם ממברנות ספציפיות, מה שמבטיח חילופי מקרומולקולות בין החלל החוץ-תאי לתוכן התא. באופן דומה, מקרומולקולות מועברות מתא תא אחד למשנהו.
הובלה של מקרומולקולות וחלקיקים לתא נקראת אנדוציטוזיס.במקרה זה, החומרים המובלים עטופים על ידי חלק מממברנת הפלזמה, נוצרת בועה (vacuole) אשר נעה בתוך התא. בהתאם לגודל השלפוחיות שנוצרו, מבחינים בין שני סוגים של אנדוציטוזיס - פינוציטוזה ופגוציטוזיס.
פינוציטוזהמספק ספיגה של חומרים נוזליים ומומסים בצורה של בועות קטנות (d=150 ננומטר). פגוציטוזיס -זוהי ספיגה של חלקיקים גדולים, מיקרואורגניזמים או שברי אברונים, תאים. במקרה זה, נוצרות שלפוחיות גדולות, פגוזומים או ואקוולים (d-250 ננומטר או יותר). בפרוטוזואה, הפונקציה הפאגוציטית היא סוג של תזונה. אצל יונקים, הפונקציה הפאגוציטית מתבצעת על ידי מקרופאגים ונויטרופילים, המגנים על הגוף מפני זיהום על ידי בליעת חיידקים פולשים. מקרופאגים מעורבים גם בסילוק תאים ישנים או פגומים ושבריהם (בגוף האדם, מקרופאגים סופגים יותר מ-100 תאי דם אדומים ישנים מדי יום). פגוציטוזיס מתחיל רק כאשר החלקיק הנספג נקשר לפני השטח של הפגוציט ומפעיל תאי קולטן מיוחדים. הקישור של חלקיקים לקולטני ממברנה ספציפיים גורמת להיווצרות פסאודופודיה, העוטפת את החלקיק ומתמזגת בקצוות, ויוצרות בועה - פאגוזום.היווצרות של פגוזום ופגוציטוזיס תקין מתרחשת רק אם, במהלך תהליך המעטפת, החלקיק נמצא במגע מתמיד עם קולטני הפלזמהלמה, כאילו "מתכווץ".
חלק ניכר מהחומר הנספג בתא באנדוציטוזיס מגיע לליזוזומים. חלקיקים גדולים כלולים ב פאגוזומיםאשר לאחר מכן מתמזגים עם ליזוזומים כדי ליצור פאגוליזוזומים.נוזלים ומקרומולקולות הנקלטות במהלך פינוציטוזיס מועברות בתחילה לאנדוזומים, אשר מתמזגים גם עם ליזוזומים ויוצרים אנדוליזומים. אנזימים הידרוליטיים שונים הקיימים בליזוזומים הורסים במהירות מקרומולקולות. תוצרי הידרוליזה (חומצות אמינו, סוכרים, נוקלאוטידים) מועברים מהליזוזומים אל הציטוזול, שם הם משמשים את התא. רוב מרכיבי הממברנה של שלפוחיות אנדוציטיות מפאגוזומים ואנדוזומים מוחזרים על ידי אקסוציטוזיס לממברנת הפלזמה ומנוצלים שם מחדש. רָאשִׁי משמעות ביולוגיתאנדוציטוזיס היא רכישת אבני בניין באמצעות עיכול תוך תאי של מקרומולקולות בליזוזומים.
ספיגת חומרים ב תאים איקריוטייםמתחיל באזורים מיוחדים של קרום הפלזמה, מה שנקרא בורות גובלים.במיקרוגרפי אלקטרונים, הבורות נראים כמו פלישות של קרום הפלזמה, שצדו הציטופלזמי מכוסה בשכבה סיבית. השכבה, כביכול, גובלת בבורות קטנים של הפלזמה. הבורות תופסים כ-2% משטח השטח הכולל של קרום התא האוקריוטי. תוך דקה הבורות גדלים, חודרים עמוק יותר ויותר, נמשכים לתוך התא ואז, מצטמצמים בבסיסם, מתפצלים ויוצרים שלפוחיות גבולות. הוכח שכרבע מהממברנה בצורת שלפוחית גבול מתפצלת מקרום הפלזמה של פיברובלסטים תוך דקה אחת. השלפוחיות מאבדות במהירות את גבולן ורוכשות את היכולת להתמזג עם הליזוזום.
יתכן אנדוציטוזיס לא ספציפי(מכונן) ו ספֵּצִיפִי(קוֹלֵט). בְּ אנדוציטוזיס לא ספציפיהתא לוכד וסופג חומרים זרים לו לחלוטין, למשל, חלקיקי פיח, צבעים. בתחילה, חלקיקים מופקדים על הגליקוקאליקס של הפלזמה. קבוצות חלבון טעונות חיוביות משוקעות (נספגות) טוב במיוחד, שכן הגליקוקאליקס נושא מטען שלילי. ואז המורפולוגיה של קרום התא משתנה. זה יכול לשקוע, ליצור פלישות (אינוואגינציות), או להיפך, ליצור צמחים שנראים מתקפלים, המפרידים בין נפחים קטנים של המדיום הנוזלי. היווצרות האינוואגינציות אופיינית יותר לתאי אפיתל המעי, אמבות וצמחים - לפגוציטים ופיברובלסטים. תהליכים אלו יכולים להיחסם על ידי מעכבי נשימה. השלפוחיות המתקבלות - אנדוזומים ראשוניים - יכולות להתמזג זה עם זה, ולהגדיל את גודלן. לאחר מכן, הם משתלבים עם ליזוזומים, הופכים לאנדוליזום - ואקוול עיכול. עוצמת פינוציטוזיס לא ספציפי בשלב נוזלי היא גבוהה למדי. מקרופאגים יוצרים עד 125, ותאי אפיתל של המעי הדק עד אלף פינוזומים לדקה. שפע הפינוזומים מוביל לעובדה שהפלזמלמה מתבזבזת במהירות על היווצרות של ואקוולים קטנים רבים. שיקום הממברנה מתרחש די מהר במהלך המיחזור במהלך אקסוציטוזיס עקב החזרת ה-vacuoles ושילובם בפלסמה. במקרופאגים, כל קרום הפלזמה מוחלף תוך 30 דקות, ובפיברובלסטים תוך שעתיים.
יותר דרך יעילהספיגה מהנוזל החוץ תאי של מקרומולקולות ספציפיות היא אנדוציטוזיס ספציפי(מתווכת על ידי קולטנים). במקרה זה, מקרומולקולות נקשרות לקולטנים משלימים על פני התא, מצטברות בפוסה הגובלת, ולאחר מכן, יוצרות אנדוזום, טובלות בציטוזול. אנדוציטוזה של הרצפטור מבטיחה הצטברות של מקרומולקולות ספציפיות בקולטן שלו. מולקולות הנקשרות לקולטן על פני הפלזמהלמה נקראות ליגנדים.בעזרת אנדוציטוזיס קולטן בתאי בעלי חיים רבים, הכולסטרול נספג מהסביבה החוץ-תאית.
קרום הפלזמה לוקח חלק בסילוק חומרים מהתא (אקסוציטוזיס). במקרה זה, ה-vacuoles מתקרבים לפלסמה. בנקודות המגע, הפלסמולמה וממברנת ה-vacuole מתמזגים ותכולת ה-vacuole נכנסת לסביבה. בחלק מהפרוטוזואה, אתרים על קרום התא לאקסוציטוזיס נקבעים מראש. אז בקרום הפלזמה של כמה ריצות יש אזורים מסוימים עם סידור נכון של כדוריות גדולות של חלבונים אינטגרליים. ל- Mucocysts ו-trichocysts של ciliates המוכנים לחלוטין להפרשה יש הילה של כדוריות חלבון אינטגרליות בחלק העליון של הפלזמה. חלקים אלה של הממברנה של המוקוציסטים והטריכוציסטים נמצאים במגע עם פני התא. אקסוציטוזיס מוזר נצפתה בנויטרופילים. הם מסוגלים, בתנאים מסוימים, לשחרר את הליזוזומים שלהם לסביבה. במקרים מסוימים, נוצרות יציאות קטנות של הפלזמה המכיל ליזוזומים, אשר לאחר מכן מתפרקים ועוברים לסביבה. במקרים אחרים, יש פלישה של הפלזמה לעומק התא ולכידת הליזוזומים שלו הממוקמים רחוק משטח התא.
התהליכים של אנדוציטוזיס ואקסוציטוזיס מבוצעים בהשתתפות מערכת המרכיבים הפיברילריים של הציטופלזמה הקשורה לפלסמולמה.
תפקוד הקולטן של הפלזמה.זהו אחד העיקריים, האוניברסליים עבור כל התאים, הוא תפקוד הקולטן של הפלזמה. הוא קובע את האינטראקציה של תאים זה עם זה ועם הסביבה החיצונית.
ניתן לייצג באופן סכמטי את כל מגוון האינטראקציות הבין-תאיות המידעיות כשרשרת של תגובות עוקבות אות-קולטן-תגובה שליח משני (מושג אות-תגובה).העברת המידע מתא לתא מתבצעת על ידי מולקולות איתות המיוצרות בחלק מהתאים ומשפיעות באופן ספציפי על אחרים הרגישים לאות (תאי מטרה). מולקולת אות - מתווך ראשינקשר לקולטנים הנמצאים על תאי מטרה המגיבים רק לאותות מסוימים. מולקולות איתות - ליגנדים -מתקרבים לקולטן שלהם כמו מפתח למנעול. ליגנדים לקולטני ממברנה (קולטני פלזמלמה) הם מולקולות הידרופיליות, הורמונים פפטידים, נוירוטרנסמיטורים, ציטוקינים, נוגדנים, ולקולטנים גרעיניים - מולקולות מסיסות בשומן, הורמונים סטרואידים ובלוטת התריס, ויטמין D. חלבוני ממברנה או רכיבי קולטן גליקוקליקס יכולים לשמש כאלמנטים של קולטני glycocalyx. משטח התא - פוליסכרידים וגליקופרוטאין. הוא האמין כי אזורים רגישים לחומרים בודדים מפוזרים על פני התא או נאספים באזורים קטנים. אז, על פני השטח של תאים פרוקריוטים ותאי בעלי חיים יש מספר מוגבל של מקומות שאיתם חלקיקים ויראליים יכולים להיקשר. חלבוני ממברנה (נשאים ותעלות) מזהים, מקיימים אינטראקציה ונושאים רק חומרים מסוימים. קולטני התא מעורבים בהעברת אותות מפני השטח של התא לתוכו. הגיוון והספציפיות של קבוצות הקולטנים על פני התא מובילים ליצירת מערכת מורכבת מאוד של סמנים המאפשרים להבחין בין התאים של עצמו מאלה של אחרים. תאים דומים מקיימים אינטראקציה זה עם זה, המשטחים שלהם יכולים להיצמד זה לזה (צימוד בפרוטוזואה, היווצרות רקמה ברב-תא). תאים שאינם קולטים סמנים, כמו גם כאלה הנבדלים במערך הסמנים הקובעים, נהרסים או נדחים. כאשר נוצר קומפלקס הקולטן-ליגנד, מופעלים חלבונים טרנסממברניים: חלבון ממיר, חלבון מגבר. כתוצאה מכך, הקולטן משנה את המבנה שלו ומקיים אינטראקציה עם המבשר של השליח השני שנמצא בתא - שָׁלִיחַ.שליחים יכולים להיות סידן מיונן, פוספוליפאז C, אדנילט ציקלאז, גואנילט ציקלאז. בהשפעת השליח, הפעלת אנזימים המעורבים בסינתזה מונופוספטים מחזוריים - AMPאוֹ HMF.האחרונים משנים את הפעילות של שני סוגים של אנזימי חלבון קינאז בציטופלזמה של התא, מה שמוביל לזרחון של חלבונים תוך-תאיים רבים.
ההיווצרות השכיחה ביותר של cAMP, שבהשפעתו גוברת הפרשת מספר הורמונים - תירוקסין, קורטיזון, פרוגסטרון, פירוק הגליקוגן בכבד ובשרירים, תדירות וחוזק של התכווצויות לב, הרס אוסטאוד והיפוך. ספיגת מים באבוביות הנפרון עולה.
הפעילות של מערכת ה-adenylate cyclase גבוהה מאוד - סינתזה של cAMP מובילה לעלייה של אלפית האות.
תחת פעולת cGMP, הפרשת אינסולין על ידי הלבלב, היסטמין על ידי תאי פיטום, סרוטונין על ידי טסיות עולה, ורקמת שריר חלק מופחתת.
במקרים רבים, במהלך היווצרות קומפלקס הקולטן-ליגנד, מתרחש שינוי בפוטנציאל הממברנה, אשר בתורו מוביל לשינוי בחדירות הפלזמה. תהליכים מטבולייםבכלוב.
על ממברנת הפלזמה ישנם קולטנים ספציפיים המגיבים לגורמים פיזיקליים. אז, בחיידקים פוטוסינתטיים, כלורופילים ממוקמים על פני התא המגיבים לאור. בבעלי חיים רגישים לאור, קרום הפלזמה מכיל כל המערכתחלבוני fogoreceptor-rhodopsins, בעזרתם הופך גירוי האור לאות כימי, ולאחר מכן דחף חשמלי.
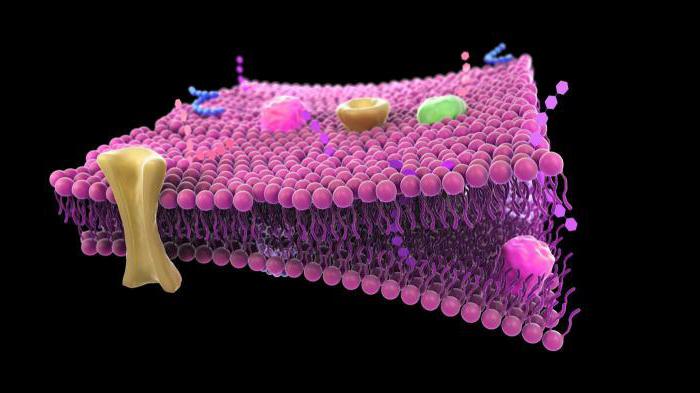
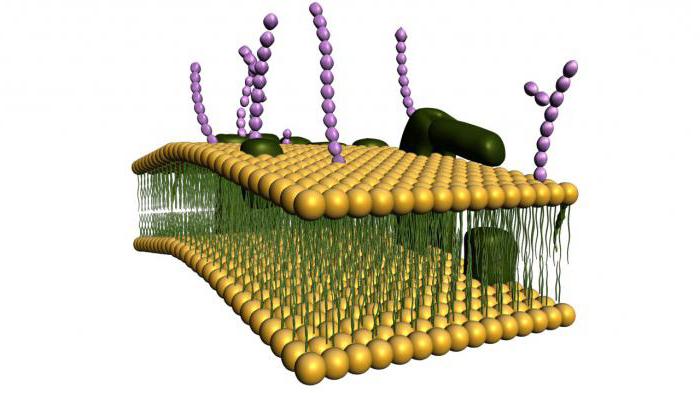
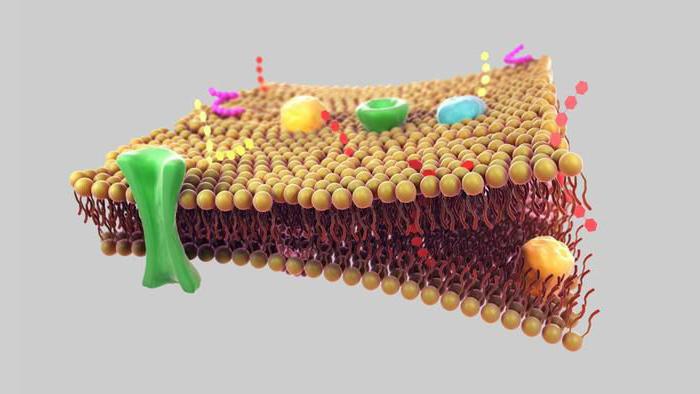
קרום התא (פלזמה) הוא מחסום חצי חדיר המפריד בין הציטופלזמה של תאים מהסביבה. כיום, מודל הפסיפס הנוזל של הממברנה, שהוצע בשנות ה-70 על ידי ג'יי זינגר וג'י ניקולסון, נלקח כבסיס בביולוגיה.
מודל זה מבוסס על מספר עקרונות בסיסיים:
1. הממברנה מורכבת משכבה כפולה של מולקולות שומנים. חלקים הידרופיליים, קוטביים של מולקולות (ראשים) ממוקמים מחוץ לממברנה, חלקים הידרופוביים, לא קוטביים (זנב) - בפנים.
2. חלבוני ממברנה משובצים בפסיפס בשכבת הדו-שכבה השומנית. חלקם עוברים דרך הממברנה (הם נקראים אינטגרלים), אחרים ממוקמים על פני השטח החיצוניים או הפנימיים של הממברנה (הם נקראים היקפיים).
3. לבסיס הליפיד של הממברנה יש תכונות של נוזל (כגון שמן נוזלי) ויכול לשנות את הצפיפות שלו. צמיגות הממברנה תלויה בהרכב השומנים ובטמפרטורה. בהקשר זה, חלבוני הממברנה והשומנים עצמם יכולים לנוע בחופשיות לאורך הממברנה ובתוכה. זוהי תכונה חשובה מאוד של הממברנה, המבטיחה שהממברנה מבצעת פונקציות רבות (ראה להלן)
4. ממברנות של רוב התוך תאיים אברוני ממברנהדומים ביסודם לממברנת הפלזמה.
5. למרות המבנה המשותף של הממברנות של כל התאים, הרכב החלבונים והשומנים בכל סוג תא ובתוך התא שונה. גם ההרכב של שכבות השומנים החיצוניות והפנימיות שונה.
פונקציות של קרום הפלזמה
הממברנה מבצעת מגוון שלם של פונקציות בתא הקשורות לשמירה על שלמות התא, הבטחת חילופי חומרים ואנרגיה בין התא לבין סביבהואינטראקציה עם תאים אחרים.
פונקציית תחבורה
לממברנת הפלזמה יש תכונות של מחסום חצי חדיר המאפשר ואינו מאפשר מעבר לחומרים ומולקולות מסוימות בלבד. ישנם מספר סוגי הובלה דרך הממברנה: הובלה פסיבית, הובלה אקטיבית, הובלת יונים, הובלה באריזת ממברנה (אנדו ואקסוציטוזיס).
הובלה פסיביתהוא הובלה של חומרים על פני הממברנה מאזור בריכוז גבוה לאזור בריכוז נמוך. הוא מתבצע בשתי צורות: בצורה של דיפוזיה פשוטה ובצורה של דיפוזיה "קלה". שני התהליכים הללו אינם דורשים אנרגיה, הם איטיים יחסית ונפסקים כאשר ריכוז החומרים משני צידי הממברנה משתווה. קצב הדיפוזיה ועצם האפשרות לשנע חומרים דרך הממברנה תלויים (בנוסף לריכוז) במספר גורמים: טמפרטורה, גודל מולקולרי ויכולת התמוססות בשומנים. חומרים מסיסים בשומן עוברים בשכבות השומנים בקלות, חומרים מסיסים במים בקושי. ישנן "נקבוביות" ליפידים וחלבונים מיוחדות בקרום שדרכן עוברת הדיפוזיה. דיפוזיה פשוטה היא תהליך איטי יחסית והטבע התאים חלבונים נושאי ממברנה מיוחדים כדי להאיץ את ההובלה. הם מתחברים עם החומר המועבר ומעבירים אותו מצד אחד של הממברנה לצד השני. עבור כל קבוצת חומרים, הממברנה חייבת להיות בעלת נשאים משלה. תהליך כזה נקרא "דיפוזיה מוקלת", המתקדם מהר פי עשרה מאשר דיפוזיה פשוטה.
הובלה יונית היא מעין הובלה פסיבית ליונים טעונים, אך יש לה מאפיינים משלה. חלבונים בציטופלזמה של תא נושאים בדרך כלל מטען שלילי על פני השטח שלהם, ויוצרים רקע חשמלי מסוים בתא. אם זרימות של יונים חיוביים (קטיונים) או שליליים (אניונים) מופנות לתוך התא, אז יונים חיובייםיותר יכנסו, מאחר וחלק מהקטיונים ייקשרו בחלבונים ציטופלזמיים, ינטרלו וייצרו הבדל נוסף בריכוזי יונים בעלי מטען שונה בתוך התא ומחוצה לו. להובלת יונים בממברנה, ישנן נקבוביות יונים מיוחדות או נשאים.
מעבר פעיל- רוב צורה חשובהתחבורה עבור התא. דיפוזיה, אם היא נמשכת מספיק זמן, מובילה לכך שמשני צידי הממברנה נוצר שיווי משקל, דבר שאינו מקובל על התא - זהו מוות עבורו. הרכב הציטופלזמה ו נוזל חוץ תאישונה מאוד. לכן קיימת בתא מערכת "הובלה פעילה". במקרה זה, הובלת מולקולות מתבצעת כנגד שיפוע הריכוז (מאזור הריכוז הנמוך לאזור הגבוה). לשם כך, ישנם קומפלקסים מיוחדים של ממברנות חלבון (תעלות יוניות ומולקולריות) הפועלות עם צריכת אנרגיה. עד 40% מכלל האנרגיה המיוצר על ידי תא הולך לעלויות הובלה!
הובלה באריזת ממברנה (אנדו - ואקסוציטוזיס). במקרים רבים, התא חייב להעביר דרך הממברנה לא מולקולות בודדות או יונים, אלא קומפלקסים מולקולריים שלמים ואפילו חלקיקים. הם אינם עוברים דרך נקבוביות הממברנה. בהקשר זה קיים סוג מיוחד של הובלה בתא - הובלה באריזת ממברנה. במקרים מסוימים מתרחשת בליטה של קרום הפלזמה החיצוני המכסה חלקיק זר (לדוגמה: חלקיק מזון או חיידק), סוגרת אותו לטבעת ממברנה וטבילה לעומק הציטופלזמה של התא. תהליך כזה התגלה על ידי המדען הרוסי המפורסם, חתן פרס נובל I.I. Mechnikov ושמו פאגוציטוזיס. תופעה זו התגלתה תוך כדי לימוד תכונות הגנהתאי דם הלוכדים חיידקים ומנטרלים אותם. התא לוכד חלקיקים קטנים יותר וטיפות נוזל על ידי יצירת פלישות של הממברנה - תהליך זה נקרא פינוציטוזיס (פינוס ביוונית - נוזל).
למעשה, פגוציטוזיס ופינוציטוזיס הם זנים של תופעה שכיחה האופיינית לרוב התאים - אנדוציטוזיס. התהליך ההפוך של אנדוציטוזיס - הסרת כל החומרים והמוצרים מהתאים נקראת אקסוציטוזיס. על בסיס הובלת הממברנה, נבנה כל תהליך ההפרשה והספיגה של סודות והורמונים על ידי תאים. חשוב להדגיש שאנדו ואקסוציטוזיס הם תהליכים שצורכים אנרגיה ולכן הם סוגים של הובלה פעילה.
תפקוד קולטן הממברנה
עקרונות הפעולה של קולטני הממברנה.
התא מקבל כל הזמן אותות מהסביבה החיצונית על נוכחותן של מולקולות איתות שונות ועליו להגיב בצורה נאותה לאותות אלו, תוך העברת מידע מהן אל התא. לשם כך, מורכבים קולטנים מיוחדים מובנים בממברנת הפלזמה. ככלל, מדובר בתצורות מורכבות של מספר מולקולות חלבון (הרכב הקולטנים עשוי לכלול גם שומנים ממברניים ופחמימות).
לכל קולטני ממברנת התא השונים יש מספר תכונות משותפות:
1. קולטנים הם ספציפיים - כלומר. נקשרים רק לחומרים מסוימים. הספציפיות של קולטן נקבעת על פי מבנה ה"אתר הפעיל" במולקולה שלו והיכולת של מולקולות אחרות להיקשר לאתר פעיל זה.
2. תהליך הקליטה והעברת האותות לממברנה או לעומק התא מתרחש בהוצאה משמעותית של אנרגיה.
3. על פי עקרון הפעולה ניתן לחלק את כל הקולטנים לשלוש קבוצות: קולטנים פעולה ישירה; קולטנים עקיפים ורצפטורים קטליטיים. במקרה הראשון, המולקולה נקשרת לחלק הקולטני של הקומפלקס ומעבירה את האות ישירות לתעלת היונים. בגרסה השנייה, החלק הקולטני של הקומפלקס מעביר אות לתעלת יונים בממברנה או עמוק לתוך התא באמצעות מערכת חלבוני עזר הנקראים "שליחים שניים". בגרסה השלישית, החלק הקולטני של הקומפלקס, לאחר אינטראקציה עם מולקולת האות, מופעל ומבצע את תפקידיו של אנזים, ובכך משפיע על תפקוד התא.
תפקיד הבידוד של הממברנה. אנשי קשר בין תאיים
IN אורגניזם רב תאיתאים נמצאים בקשר מתמיד אחד עם השני, אשר לפעמים רוכש תכונות של אינטראקציה קרובה. זה בולט במיוחד ברקמות הגובלות בסביבה החיצונית (איטמנטים, מעיים, כיווני אוויר) או כאשר יש צורך במגע קרוב של תאים לעבודה מתואמת (שרירים חלקים ולב, תאי עצביםוסיבים).
ישנם ארבעה סוגים של צמתים בין-תאיים: צמתים מבודדים (צומת tight junction), צמתים הדוקים (גap junction), דסמוזומים (דסמוזום) וצמתים דביקים (צומת הידבקות).
בידוד אנשי קשר עומדים בשמם - מתחמי חלבוןבממברנות של תאים שכנים מחוברים בצורה כזו שאף מולקולות ויונים לא עוברות דרך מגע זה. צמתים הדוקים מאפשרים למולקולות קטנות ויונים לעבור דרכם, וחלפה בין תאים יכולה להתרחש דרכם. בפרט, צמתים הדוקים מותאמים על ידי עצבים ו תאי שרירלהעברת אותות חשמליים. דסמוזומים נועדו להחזיק קבוצת תאים זה לזה או למצע אחר.
הם יכולים להיעלם במהירות ולהופיע שוב. מגעי הדבקה נועדו גם לחבר תאים זה לזה. הם רגישים יותר ויכולים להיווצר במהירות ולהיעלם לפי הצורך.
הגרעין אחראי על אחסון החומר הגנטי שנרשם ב-DNA, וגם שולט בכל התהליכים של התא. הציטופלזמה מכילה אברונים, שלכל אחד מהם יש תפקידים משלו, כמו למשל סינתזה חומר אורגני, עיכול וכו 'ועל המרכיב האחרון נדבר ביתר פירוט במאמר זה.
בביולוגיה?
שִׂיחָה שפה פשוטה, זו קליפה. עם זאת, זה לא תמיד בלתי חדיר לחלוטין. הובלה של חומרים מסוימים על פני הממברנה מותרת כמעט תמיד.
בציטולוגיה ניתן לחלק את הממברנות לשני סוגים עיקריים. הראשון הוא קרום הפלזמה המכסה את התא. השני הוא הממברנות של האברונים. ישנם אברונים בעלי קרום אחד או שניים. תאים עם ממברנה בודדת כוללים את הרשת האנדופלזמית, ואקואולים וליזוזומים. פלסטידים ומיטוכונדריה שייכים לשתי הממברנות.
כמו כן, ממברנות יכולות להיות בתוך אברונים. בדרך כלל מדובר בנגזרות של הממברנה הפנימית של אברונים דו-ממברניים.
כיצד מסודרים הממברנות של אברונים דו-ממברניים?
לפסטידים ולמיטוכונדריה יש שתי קונכיות. הקרום החיצוני של שני האברונים חלק, אך הפנימי יוצר את המבנים הדרושים לתפקוד האורגנואיד.
אז, למעטפת המיטוכונדריה יש בליטות פנימה - קריסטות או רכסים. עליהם מתרחש מחזור התגובות הכימיות הנחוצות לנשימה התאית.
נגזרות של הממברנה הפנימית של הכלורופלסטים הן שקיות בצורת דיסק - thylakoids. הם נאספים בערימות - דגנים. גרנה נפרדת משולבת זו בזו בעזרת למלות - מבנים ארוכים הנוצרים גם ממברנות.
מבנה הממברנות של אברונים חד-ממברניים
לאברונים אלה יש רק קרום אחד. לרוב מדובר בקרום חלק המורכב משומנים וחלבונים.
תכונות של מבנה קרום הפלזמה של התא
הממברנה מורכבת מחומרים כמו שומנים וחלבונים. המבנה של קרום הפלזמה מספק את עוביו של 7-11 ננומטר. עיקר הממברנה מורכב משומנים. 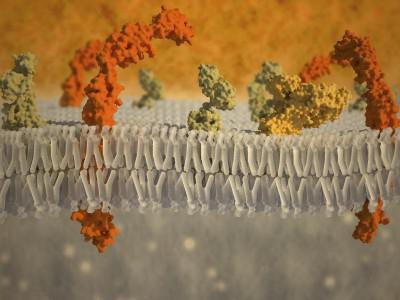
המבנה של קרום הפלזמה מספק נוכחות של שתי שכבות בו. הראשונה היא שכבה כפולה של פוספוליפידים, והשנייה היא שכבה של חלבונים.
שומנים בממברנת פלזמה
הליפידים המרכיבים את קרום הפלזמה מחולקים לשלוש קבוצות: סטרואידים, ספינגופוספוליפידים וגליצרופוספוליפידים. המולקולה של האחרון מכילה בהרכבה את השארית של האלכוהול התלת-הידרי גליצרול, שבו אטומי המימן של שתי קבוצות הידרוקסיל מוחלפים בשרשראות של חומצות שומן, ואטום המימן של קבוצת ההידרוקסיל השלישית מוחלף בשארית חומצה זרחתית. , שאליו מצמידים, בתורו, שאריות של אחד הבסיסים החנקניים.
ניתן לחלק את מולקולת הגליצרופוספוליפיד לשני חלקים: הראש והזנבות. הראש הידרופילי (כלומר, הוא מתמוסס במים), והזנבות הידרופוביים (הם דוחים מים, אבל מתמוססים בממיסים אורגניים). בשל מבנה זה, ניתן לקרוא למולקולת הגליצרופוספוליפידים אמפיפילית, כלומר גם הידרופובי וגם הידרופילי בו-זמנית.
ספינגופוספוליפידים דומים ב מבנה כימיעבור גליצרופוספוליפידים. אבל הם שונים מאלה שהוזכרו לעיל בכך שבהרכב שלהם, במקום שאריות גליצרול, יש להם שאריות אלכוהול ספינגוזין. למולקולות שלהם יש גם ראשים וזנבות.
התמונה למטה מציגה בבירור את המבנה של קרום הפלזמה.
חלבוני ממברנת פלזמה
באשר לחלבונים המרכיבים את מבנה קרום הפלזמה, מדובר בעיקר בגליקופרוטאין.
בהתאם למיקומם בקליפה, ניתן לחלק אותם לשתי קבוצות: פריפריאלי ואינטגרלי. הראשון הם אלו שנמצאים על פני הממברנה, והשניים הם אלו שחודרים את כל עובי הממברנה ונמצאים בתוך שכבת השומנים.
בהתאם לתפקודים שמבצעים חלבונים, ניתן לחלק אותם לארבע קבוצות: אנזימים, מבניים, תחבורה וקולטן.
כל החלבונים שנמצאים במבנה של קרום הפלזמה אינם קשורים כימית לפוספוליפידים. לכן, הם יכולים לנוע בחופשיות בשכבה הראשית של הממברנה, להתאסף בקבוצות וכו'. לכן לא ניתן לקרוא למבנה של קרום הפלזמה של התא סטטי. זה דינמי, מכיוון שהוא משתנה כל הזמן.
מה תפקידו של קרום התא?
המבנה של קרום הפלזמה מאפשר לו להתמודד עם חמש פונקציות.
הראשון והעיקרי הוא הגבלת הציטופלזמה. כתוצאה מכך, לתא יש צורה קבועהוגודל. פונקציה זו מובטחת על ידי העובדה כי קרום הפלזמה הוא חזק ואלסטי.
התפקיד השני הוא אספקה בשל הגמישות שלהם, קרומי פלזמה יכולים ליצור יציאות וקפלים בצמתים שלהם.
הפונקציה הבאה דופן תא- הובלה. זה מסופק על ידי חלבונים מיוחדים. תודות להם החומרים הנכוניםניתן להעביר אל הכלוב, וניתן להיפטר ממנו מיותרים.
בנוסף, קרום הפלזמה מבצע פונקציה אנזימטית. זה מתבצע גם הודות לחלבונים.
והפונקציה האחרונה היא איתות. בשל העובדה שחלבונים בהשפעת תנאים מסוימים יכולים לשנות את המבנה המרחבי שלהם, קרום הפלזמה יכול לשלוח אותות לתאים.
עכשיו אתה יודע הכל על ממברנות: מהי קרום בביולוגיה, מה הם, איך מסודרים קרום הפלזמה והממברנות האורגניות, אילו פונקציות הם מבצעים.
הרצאה מספר 4.
מספר שעות: 2
קרום פלזמה
1.
2.
3. אנשי קשר בין תאיים.
1. מבנה קרום הפלזמה
קרום הפלזמה, או פלזמהלמה, הוא מבנה היקפי משטח המגבילהתא מבחוץ ומספק את הקשר שלו עם תאים אחרים והסביבה החוץ תאית. יש לו עוביבערך 10 ננומטר. בין שאר קרומי התא, הפלזמהלמה היא העבה ביותר. מבחינה כימית, קרום הפלזמה הוא קומפלקס ליפופרוטאין.המרכיבים העיקריים הם שומנים (כ-40%), חלבונים (יותר מ-60%) ופחמימות (כ-2-10%).
הליפידים הם קבוצה גדולהחומרים אורגניים בעלי מסיסות ירודה במים (הידרופוביות) ומסיסות טובה בממיסים ושומנים אורגניים (ליפופיליות).שומנים מייצגים המצויים בממברנת הפלזמה הם פוספוליפידים, ספינגומיאלינים וכולסטרול. IN תאי צמחיםהכולסטרול מוחלף בפיטסטרול. על ידי תפקיד ביולוגיניתן לחלק חלבוני ממברנת פלזמה ל חלבוני אנזים, קולטן וחלבונים מבניים.פחמימות פלזמה הן חלק מהפלזמה במצב קשור (גליקוליפידים וגליקופרוטאינים).
כיום זה מקובל באופן כללי מודל נוזל-פסיפס של מבנה ממברנה ביולוגית.לפי מודל זה, הבסיס המבני של הממברנה נוצר על ידי שכבה כפולה של פוספוליפידים עטופה בחלבונים. זנבות המולקולות פונים זה לזה בשכבה כפולה, בעוד ראשי הקוטב נשארים בחוץ ויוצרים משטחים הידרופיליים. מולקולות החלבון אינן יוצרות שכבה רציפה, הן ממוקמות בשכבת השומנים, צוללות לעומקים שונים (יש חלבונים היקפיים, חלק מהחלבונים חודרים דרך הממברנה, חלקם שקועים בשכבת השומנים). רוב החלבונים אינם קשורים לשומני ממברנה; נראה שהם צפים ב"אגם שומנים". לכן, מולקולות חלבון מסוגלות לנוע לאורך הממברנה, להתאסף בקבוצות, או להיפך, להתפזר על פני הממברנה. זה מצביע על כך שקרום הפלזמה אינו יצירה סטטית וקפואה.
מחוץ לפלזמה נמצאת שכבת האפיממברנה - גליקוקליקס. העובי של שכבה זו הוא כ-3-4 ננומטר. Glycocalyx נמצא כמעט בכל תאי בעלי החיים. זה קשור לממברנת הפלזמה קומפלקס גליקופרוטאין.פחמימות יוצרות שרשראות ארוכות ומסועפות של פוליסכרידים הקשורות לחלבונים ולשומנים של קרום הפלזמה. הגליקוקליקס יכול להכיל חלבוני אנזים המעורבים בפירוק חוץ תאי של חומרים שונים. מוצרים של פעילות אנזימטית (חומצות אמינו, נוקלאוטידים, חומצת שומןוכו') מועברים דרך קרום הפלזמה ונספגים על ידי תאים.
קרום הפלזמה מתחדש כל הזמן. זה מתרחש על ידי שרוך בועות קטנות מפני השטח שלו לתוך התא והטבעת vacuoles מתוך התא לתוך הממברנה. לפיכך, בתא יש זרימה מתמדת של יסודות ממברנה: ממברנת הפלזמה לתוך הציטופלזמה. (אנדוציטוזה)וזרימת מבני הממברנה מהציטופלזמה אל פני התא (אקסוציטוזיס).במחזור של ממברנות, התפקיד המוביל מוקצה למערכת הוואקווולים של הממברנה של קומפלקס גולגי.
4. פונקציות של קרום הפלזמה. מנגנוני הובלה של חומרים דרך הפלזמה. תפקוד הקולטן של הפלזמה
קרום הפלזמה מבצע מספר פונקציות חשובות:
1) מַחסוֹם.תפקיד המחסום של קרום הפלזמה הוא להגבלת הדיפוזיה החופשית של חומרים מתא לתא, מניעתדליפת סיבוב של התוכן המסיס במים של התא. אך מאזאיך התא חייב לקבל את החומרים המזינים הדרושים, אתהלחלק תוצרי קצה מטבוליים, לווסת תוך תאיריכוזי יונים, ואז הוא יצר מנגנונים מיוחדים להעברת חומרים דרך קרום התא.
2) תַחְבּוּרָה.פונקציית ההובלה היא הבטחת כניסה ויציאה של חומרים שונים לתא ומחוצה לו. תכונה חשובה של הממברנה היא החדירות הסלקטיבית, או חדירות למחצה. זה עובר בקלות מים ומסיס במיםגזים ודוחה מולקולות קוטביות כגון גלוקוז אוחומצות אמינו.
ישנם מספר מנגנונים להובלת חומרים על פני הממברנה:
הובלה פסיבית;
מעבר פעיל;
הובלה באריזת ממברנה.
הובלה פסיבית. ריכוך -זוהי התנועה של חלקיקים של המדיום, המובילה להעברתחומרים מאזור שבו ריכוזו גבוה לאזור עם ריכוז נמוךtion. במהלך הובלת דיפוזיה, הממברנה מתפקדת כמחסום אוסמוטי. קצב הדיפוזיה תלוי בערךמולקולות ומסיסותן היחסית בשומנים. כמה שפחות פעמיםמידות של מולקולות וככל שהן מסיסות יותר בשומן (ליפופיליות), כך הן יעברו מהר יותר בשכבת הדו-שומנים.דיפוזיה יכולה להיות ניטראלי(העברה של לא טעוןמולקולות) ו קל(בעזרת חלבונים מיוחדיםאפים). דיפוזיה קלה מהירה יותר מדיפוזיה ניטרלית.מקסימום חודרלמים יש את היכולתכיצד המולקולות שלו קטנות ולא טעונות. פיזור מים דרך תאיםקרום נקרא osmo שפמנוןההנחה היא כי בתאממברנה לחדירהמים וכמה יונים בעצםיש "נקבוביות" מיוחדות. המספר שלהםקטן, והקוטר הואבערך 0.3-0.8 ננומטר. פיזור המהיר ביותר דרך הממברנה ובכן, מסיס בקלות בשומניםדו-שכבה של מולקולה, למשל O, ו מולקולות קוטביות לא טעונותly בקוטר קטן (SO, mo chevin).
הובלה של מולקולות קוטביות (עםסוכרים, חומצות אמינו), מיושםנמסר בעזרת הובלת ממברנה מיוחדתחלבונים נקראים הקלה על דיפוזיה.חלבונים כאלה הםנמצאים בכל סוגי הממברנות הביולוגיות, וכל אחד ספציפי חלבון ny נועד להעביר מולקולות ממעמד מסוים sa. חלבוני הובלה הם טרנסממברניים, שרשרת הפוליפפטיד שלהם חוצה את דו-שכבת השומנים מספר פעמים, ויוצרת יש לו מעברים. זה מבטיח העברת ספציפיחומרים דרך הממברנה ללא מגע ישיר איתו.ישנם שני סוגים עיקריים של חלבוני תחבורה: סנאים- מובילים (מובילים) ו יצירת ערוציםחלבונים (לבןערוצי ki). חלבוני נשא נושאים מולקולות על פני הממברנה על ידי שינוי תצורה תחילה.חלבונים יוצרי תעלה יוצרים ממברנות מלאות מים נקבוביות. כאשר הנקבוביות פתוחות, מולקולות של חומרים ספציפיים(בדרך כלל יונים אנאורגניים בגודל ומטען מתאימים) עוברים דרכם. אם למולקולת החומר המועבר אין מטען, אז כיוון ההובלה נקבע על ידי שיפוע הריכוז.אם המולקולה טעונה, אז על ההובלה שלה, בנוסף לשיפוע, con ריכוז, המטען החשמלי של הממברנה משפיע גם על (ממברנהפוטנציאל). הצד הפנימי של הפלזמה נטען בדרך כללשלילי ביחס כלפי חוץ. פוטנציאל הממברנה מקל על חדירת יונים בעלי מטען חיובי לתא ו מונע מעבר של יונים בעלי מטען שלילי.
מעבר פעיל. הובלה פעילה היא תנועה של חומרים כנגד שיפוע אלקטרוכימי. זה תמיד מתבצע על ידי חלבוני טראנס.סבלים וקרובים זאן עם מקור אנרגיהוואי. בחלבונים-העברתלצ'יקי יש עלילות מחייב עם הובלהחומר ראוי.כמה שיותר כאלה tkov מתקשר עם דבריםככל שהמהירות גבוהה יותרגידול התחבורה. העברה סלקטיבית של חומר אחד נקראת יוניפורט. העברת מספר חומרים מתבצעת קוטראן מערכות ספורט. אם ההעברה עוברת לכיוון אחד -זֶה סימן,אם מול נגד נמל.כך,לדוגמה, גלוקוז מועבר מהנוזל החוץ-תאי לתוך התא בצורה חד-פורטלית. העברת גלוקוז ונא 4 ממערכת המעיים אוצינוריות של הכליות, בהתאמה, לתוך תאי המעי או הדם מתבצעת באופן סימפורטלי, והעברה של C1 ~ ו- HCO "הוא אנטי-פורט. ההנחה היא כי במהלך ההעברה, קונפורמציה הפיכהשינויים במסוע, המאפשרים לנוע לחומרים המחוברים אליו.
דוגמה לחלבון נשא המשמש להובלהחומרים, האנרגיה המשתחררת במהלך ההידרוליזה של ATP היאלא + -K + משאבה, נמצא בממברנת הפלזמה של כל התאים. Na+-K המשאבה פועלת על העיקרון של אנטי-פורט, שאיבהוואי נא "מהתא ואל פנים התא כנגד האלקטרוכימיים שלהם שיפועים. מִדרוֹן Na+ יוצר לחץ אוסמוטי, שומר על נפח התא ומבטיח הובלה של סוכרים וחומצות אמינוחומצות. פעולת המשאבה הזו צורכת שליש מכל האנרגיה הדרושה לפעילות החיונית של התאים.כאשר לומדים את מנגנון הפעולה Na + - K + משאבה הותקנהידוע שזהו אנזים ATPase ואנזים טרנסממברני חלבון אינטגרלי. בנוכחות Na+ ו-ATP תחת הפעולה של ATP-פוספט סופני מופרד מ-ATP ומחובר לשאריותחומצה אספרטית על מולקולת ATPase. מולקולת Phos ATPaseמפוריל, משנה את תצורתו ו Na+ נגזר מ תאים. בעקבות הנסיגהלא K" מועבר תמיד מהתא לתוך התא. לשם כך, הפוספט שהתחבר קודם לכן מבוקע מ-ATPase בנוכחות K. האנזים עובר דה-פוספורילציה, משחזר את תצורתו ו-K 1 "נשאב" לתא.
ATPase נוצר על ידי שתי תת יחידות, גדולות וקטנות.תת-היחידה הגדולה מורכבת מאלפי שאריות חומצות אמינו,חציית הדו-שכבה מספר פעמים. יש לו קטליטי פעילות ומסוגלת להיות זרחן ודפוספו באופן הפיךמרופט. תת-יחידה גדולה בצד הציטופלזמיאין אתרי קישור Na+ ו-ATP, ובחוץ -אתרים לקישור K + ו-ouabain. תת-היחידה הקטנה היאגליקופרוטאין ותפקודו עדיין לא ידוע.
Na+-K למשאבה יש אפקט אלקטרוגני. הוא מסיר שלושהיון טעון חיובילא פ מחוץ לתא ומציג שנייםיון K כתוצאה מכך, זרם זורם דרך הממברנה ויוצר אלקטרוןפוטנציאל ric בעל ערך שלילי בחלק הפנימי של התא ביחס לפני השטח החיצוניים שלו. Na "-K + המשאבה מווסתת את נפח התא, שולטת בריכוז החומריםבתוך התא, שומר על לחץ אוסמוטי, משתתף ביצירת פוטנציאל הממברנה.
הובלה באריזת ממברנה. העברה על פני הממברנה של מקרומולקולות (חלבונים, חומצות גרעין)הרבה, פוליסכרידים, ליפופרוטאינים) וחלקיקים אחרים מתבצעים באמצעות היווצרות והיתוך עוקבים של מוקףשלפוחיות הקשורות לממברנה (שלפוחיות). תהליך הובלה שלפוחיתזה עובר שני שלבים. בהתחלהממברנת שלפוחית ופלסמהלהיצמד יחד ואז להתמזג.למעבר של שלב 2, זה הכרחידימו כך שמולקולות המים הן אתהצפופים על ידי דו-שכבות שומנים באינטראקציה, המתקרבות זו לזו עד למרחק של 1-5 ננומטר.שוקל שהתהליך הזה מופעלמיוחד חלבוני היתוך(הֵםמבודד עד כה רק מוירוסים). הובלה שלפוחית ישתכונה חשובה - מקרומולקולות נספגות או מופרשות,כלול שלפוחית, בדרך כלל לאניתן לערבב עם מקרומולים אחריםculae או אברוני תאים. פושלפוחיות יכולות להתמזג עם ספציפיות ממברנות, המספקותחילופי chivaet של מקרומולקולות ביןdu מרחב תאיים והתוכן של התא. באופן דומהמקרומולקולות מועברות מתא תא אחד למשנהו.
הובלה של מקרומולקולות וחלקיקים לתא נקראת אנדו ציטוזה.במקרה זה, החומרים המובלים עטופים בתהאת קרום הפלזמה, נוצרת שלפוחית (vacuole), אשרשעובר לתוך התא. תלוי בגודל התמונהבועות, ישנם שני סוגים של אנדוציטוזיס - פינוציטוזה ופגוציטוזיס.
פינוציטוזהמספק ספיגה של נוזל ומומסחומרים בצורת בועות קטנות (ד =150 ננומטר). פגוציטוזיס -הוא קליטה של חלקיקים גדולים, מיקרואורגניזםקריאה או שברי אברונים, תאים. במקביל הם נוצריםשיה שלפוחיות גדולות, פאגוזומים או וואקוולים ( d -250 ננומטר או יותר). בְּ תפקוד phagocytic protozoan - סוג של תזונה. אצל יונקים, הפונקציה הפאגוציטית מתבצעת על ידי מקרופאגים וסירוספרופילים המגנים על הגוף מפני זיהום על ידי קליטת חיידקים פולשים. גם מקרופאגים מעורבים בניצוליצירת תאים ישנים או פגומים ושבריהם (בגוףמקרופאגים אנושיים בולעים יותר מ-100 אריתריטידים ישנים מדי יוםרוציטים). Phagocytosis מתחיל רק כאשר החלקיק שנבלענקשר לפני השטח של הפגוציט ומפעיל מיוחדתאים קולטן ניי. שיוך חלקיקים עם re ספציפיקולטני ממברנה גורמים להיווצרות פסאודופודיה, אשרשיפון עוטף את החלקיק ומתמזג בקצוות ויוצר בועה -פאגוזום.היווצרות פאגוזומים ופגוציטוזיס תקיןהולך רק אם בתהליך של עטוף את החלקיקנמצא כל הזמן במגע עם קולטני פלזמהלמה, כאילו "קפאוןברק מהבהב."
חלק ניכר מהחומר שנספג בתא על ידי enדוציטוזה, מסתיימת בליזוזומים. חלקיקים גדולים כוללמקווה פנימה פאגוזומיםאשר לאחר מכן מתמזגים עם ליזוזומים כדי ליצור פאגוליזוזומים.נוזל ומקרומולקולות שנספגו במהלךפינוציטוזיס, מועברים בתחילה לאנדוזומים, אשרמתמזגים עם ליזוזומים ליצירת אנדוליזומים. אני נוכח אנזימים הידרוליטיים שונים הנמצאים בליזוזומיםro להרוס מקרומולקולות. מוצרי הידרוליזה (חומצת אמינוהמון, סוכרים, נוקלאוטידים) מועברים מהליזוזומים לציטוזול,איפה הם משמשים את התא. רוב מרכיבי הממברנה שלפוחיות אנדוציטיות מפאגוזומים ואנדוזומים חוזרות באקסוציטוזיס לממברנת הפלזמה ושם נעלמות מחדשlyse. המשמעות הביולוגית העיקרית של אנדוציטוזיס היאיש קבלה של אבני בניין עקב תוך תאי עיכול של מקרומולקולות בליזוזומים.
ספיגת חומרים בתאים איקריוטים מתחילה באזורים cialized של קרום הפלזמה, מה שנקראאנחנו X בורות גובלים.על מיקרוגרפי אלקטרוניםבורות נראים כמו פלישות של קרום הפלזמה, ציטופלזמהשצד המחצלת שלו מכוסה בשכבה סיבית. שכבה כמוהיה גובל בכיכר בורות קטנים malemas. הבורות תופסים כ-2% מהמעל פני השטח של קרום התאאנחנו אוקריוטים. תוך דקה הפוסות גדלות יותר ויותרשיה, נמשכים לתוך הכלוב ואז, מצטמצם בבסיס, מתפצל,יוצרים שלפוחיות שוליות.נקבע כי מפיברובלסט ממברנה מטיתטוב תוך דקה פתיתכרבע מממברנות בצורת pu עם גבולות zyrkov. בועות מאבדות מהר הגבול שלהם ולרכוש דרךהיכולת להתמזג עם הליזוזום.
יתכן אנדוציטוזיס לא ספציפי(מכונן)ו ספֵּצִיפִי(קוֹלֵט).בְּ אנדוציטוזיס לא ספציפיהתא משתלטסופג חומרים זרים לו לחלוטין, למשל, חלקיקי פיח,צבעים. ראשית, חלקיקים מופקדים על הגליקוקאליקס פלזמהלמה. טוב במיוחד משקעים (נספג) עלקבוצות טעונות חיובית של חלבונים, שכן הגליקוקאליקס נושא מטען שלילי. אז משתנה המורפולוגיה של התאממברנות. זה יכול לשקוע וליצור פלישות(פלישה), או להיפך, יוצרים תולדות,שנראה שמצטברים ומפרידים בין נפחים קטנים סביבה נוזלית. היווצרות האינוואגינציות אופיינית יותרעבור תאים של אפיתל המעי, אמבות וצמחים - עבור פגוציטים ו פיברובלסטים. תהליכים אלה יכולים להיחסם על ידי מעכביםנְשִׁימָה. השלפוחיות המתקבלות הן אנדוזומים ראשוניים שיכולים להתמזגאחד את השני, הולך וגדל בגודלם. בעתיד הם יצטרפו מתערבבים עם ליזוזומים, הופכים לאנדוליזום - מעכלואקואול חדש. עוצמת פינוציטוזיס לא ספציפי בשלב נוזלי עדגבוה בטירוף. מקרופאגים יוצרים עד 125, ותאי אפיתל דקהמעי עד אלף פינוזום לדקה. שפע הפינוזומים מוביל לעובדה שהפלזמלמה מתבזבזת במהירות על היווצרות של מספר רב שלנוכחות של ואקוולים קטנים. התאוששות הממברנה די מהירה.tro במהלך מיחזור בתהליך של אקסוציטוזיס עקב החזרה של wakuoles ושילובם בפלסמהלמה. במקרופאגים, כולם פלזמההממברנה הקלאסית מוחלפת תוך 30 דקות, ובפיברובלסטים תוך שעתיים.
דרך יעילה יותר לספוג מנוזל חוץ תאימקרומולקולות ספציפיות לעצם הוא ספציפי en דוציטוזה(מתווכת על ידי קולטנים). המקרומולקולות בזהנקשרים לקולטנים משלימים על פני השטחתאים מצטברים בפוסה הגובלת, ולאחר מכן, יוצרים אנדוזום, שקועים בציטוזול. אנדוציטוזה של הרצפטור מבטיחה הצטברות של מקרומולקולות ספציפיות בקולטן שלו.מולקולות הנקשרות על פני הפלזמהלמה לקולטןטורוס נקראים ליגנדים. בעזרת הקולטן אנדוציטוזיס בתאי בעלי חיים רבים היא הספיגהכולסטרול חוץ תאיסביבה.
קרום הפלזמה לוקח חלק בסילוק חומרים מהתא (אקסוציטוזיס). במקרה זה, ה-vacuoles מתקרבים לפלסמה. בנקודות המגע, הפלסמולמה וממברנת ה-vacuole מתמזגים ותכולת ה-vacuole נכנסת לסביבה.בחלק מהפרוטוזואה, אתרים על קרום התא לאקסוציטוזיס נקבעים מראש. אז, בממברנת הפלזמה לכמה ריצות יש אזורים מסוימים עם סידור נכון של כדוריות גדולות של חלבונים אינטגרליים. בְּמוקוציסטות וטריכוציסטות של ריצות מוכנות לחלוטין להפרשה, בחלק העליון של הפלזמה יש קורולה של כדוריות אינטגרליותחלבונים. קטעים אלה של הממברנה של ריריות וטריכוציסטות של סופריקהלהיצמד לפני השטח של התא.אקסוציטוזיס מוזר נצפתה בנויטרופילים. הם ספומסוגל להשתחרר לסביבה בתנאים מסוימיםלפוצץ את הליזוזומים שלך. במקרים מסוימים, נוצרות יציאות קטנות של הפלזמה המכיל ליזוזומים, אשר לאחר מכן מתפרקים ועוברים לסביבה. במקרים אחרים, יש פלישה של הפלזמלמה עמוק לתוך התא ולכידת הליזוזומים שלו, הממוקמת nyh רחוק משטח התא.
התהליכים של אנדוציטוזיס ואקסוציטוזיס מבוצעים בהשתתפות מערכת המרכיבים הפיברילריים של הציטופלזמה הקשורה לפלסמולמה.
תפקוד הקולטן של הפלזמה. זֶה מהעיקר, האוניברסלי עבור כל התאים, הוא reתפקוד הקולטן של הפלזמה. זה מגדיר אינטראקציהתאים זה עם זה ועם הסביבה.
ניתן לייצג באופן סכמטי את כל מגוון האינטראקציות הבין-תאיות המידעיות כשרשרת של עוקביםתגובות אות-קולטן-שליח שני-תגובה (מוּשָׂג אות-תגובת).אותות מעבירים מידע מתא לתאניו מולקולות המיוצרות בחלק מהתאים ומיוחדותלהשפיע פיזית על אחרים הרגישים לאות (תאים-מי שני). מולקולת אות - מתווך ראשיכריכה etsya עם קולטנים הממוקמים על תאי מטרה, מגיבים רק עבור אותות מסוימים. מולקולות איתות - ליגנדים-מתקרבים לקולטן שלהם כמו מפתח למנעול. ליגנד-mi עבור קולטני ממברנה (קולטני פלסמלמה) הואמולקולות הידרופיליות, הורמונים פפטידים, נוירומדיהטורי, ציטוקינים, נוגדנים, ולקולטנים גרעיניים - שומן מולקולות מסיסות, סטרואידים והורמוני בלוטת התריס, ויטמין Dכקולטנים למעלהתאים יכולים לפעול כחלבוניםממברנות או אלמנטים גליקוקליטיםca - פוליסכרידים וגליקופרוטאינים.הוא האמין כי רגישלהפריד אזורי חומרים, לפזרsan לאורך פני התא אוברינס לרצועות קטנות. כן, עלפני השטח של תאים פרוקריוטיםותאי בעלי חיים יש גבולותמספר קבוע של מקומות שאיתם הם יכוליםקושרים חלקיקים ויראליים. אני אניסנאים (נשאים וקאנהly) ללמוד, ליצור אינטראקציה ולהעבירללבוש רק חומרים מסוימים.קולטני תאים מעורביםהעברת אותות מפני השטח של התא פנימה.גיוון וספציפיותתעלה של קולטנים על פני התאמוביל למערכת מורכבת מאודאנחנו סמנים להבדילמשלו תאים מזרים. תאים דומיםמתקשרים זה עם זה, המשטחים שלהם יכולים להיצמד זה לזה (צימוד בפרוטוזואה, היווצרות רקמות באורגניזמים רב-תאיים). תאים לא קולטיםסמנים נפוצים, כמו גם אלה הנבדלים ביניהםבורון של סמנים קובעיםנקרעים או נדחים.עם היווצרות קומפלקס הקולטן-ליגנד,חלבונים טרנסממברניים: ממיר חלבון, משפר חלבון.כתוצאה מכך, הקולטן משנה את הקונפורמציה והאינטראקציה שלולא עם מבשר סלולרי של השליח השני ka - שָׁלִיחַ.שליחים יכולים להיות סידן מיונן, פוספוליפעבור C, adenylate cyclase, guanylate cyclase. מושפע מסנג'רהפעלה של אנזימים המעורבים בסינתזה מונופוספטים מחזוריים - AMPאוֹ HMF.האחרונים משנים את הנכסנוכחותם של שני סוגים של אנזימי חלבון קינאז בציטופלזמה של התא, מה שמוביל לזרחון של חלבונים תוך תאיים רבים.
ההיווצרות השכיחה ביותר של cAMP, תחת הפעולה שלהפרשת מספר הורמונים - תירוקסין, קורטיזון, פרוגסטרון, עולה, פירוק הגליקוגן בכבד ובשרירים עולה,קצב לב וכוח, אוסטאוהרס, הפוך ספיגת מים בצינוריות של הנפרון.
הפעילות של מערכת ה-adenylate cyclase גבוהה מאוד - סינתזה של cAMP מובילה לעלייה של אלפית האות.
תחת פעולת cGMP, הפרשת אינסולין על ידי הלבלב, היסטמין על ידי תאי פיטום, סרוטוניןבוציטים, רקמת שריר חלק מופחתת.
במקרים רבים, עם היווצרות קומפלקס הקולטן-ליגנדיש שינוי בפוטנציאל הממברנה, אשר בתורו מוביל לשינוי בחדירות הפלזמה ולמטבוליתכמה תהליכים בתא.
קרום הפלזמה מכיל קולטנים ספציפיים טורי מגיבה לגורמים פיזיים. אז, בחיידקים פוטוסינתטיים, כלורופילים ממוקמים על פני התא,מגיבה לאור. בחיות רגישות לאור בפלזמהישנה מערכת שלמה של חלבוני פוגורצפטורים בממברנה הקוסטית -רודופסינים, בעזרתם הופך גירוי האורהופך לאות כימי ולאחר מכן לדחף חשמלי.
3. אנשי קשר בין תאיים
בבעלי חיים רב-תאיים, הפלסמולמה לוקחת חלק בהיווצרות קשרים בין תאייםמתן אינטראקציות בין-תאיות. ישנם מספר סוגים של מבנים כאלה.
§ מגע פשוט.מגע פשוט נמצא בין רוב התאים ממקורות שונים הסמוכים זה לזה. מייצג את ההתכנסות של ממברנות הפלזמה של תאים שכנים במרחק של 15-20 ננומטר. במקרה זה, מתרחשת האינטראקציה של שכבות הגליקוקאליקס של תאים שכנים.
§ מגע הדוק (סוגר). עם חיבור כזה, השכבות החיצוניות של שני ממברנות הפלזמה קרובות ככל האפשר. ההתקרבות כה צפופה שיש מעין מיזוג של חלקים של ממברנות הפלזמה של שני תאים שכנים. היתוך הממברנות אינו מתרחש על פני כל אזור המגע הדוק, אלא הוא סדרה של התכנסות נקודתית של ממברנות. תפקידו של מגע הדוק הוא לחבר תאים זה לזה באופן מכני. אזור זה בלתי חדיר עבור מקרומולקולות ויונים, ולכן הוא נועל, תוחם את הפערים הבין-תאיים (ויחד איתם, את הסביבה הפנימית של הגוף) מהסביבה החיצונית.
§ כתם ההדבקה, או דסמוזום. הדסמוזום הוא אזור קטן בקוטר של עד 0.5 מיקרומטר. באזור הדסמוזום בצד הציטופלזמה, יש אזור של סיבים דקים. התפקיד התפקודי של הדסמוזומים הוא בעיקר בחיבור המכני בין התאים.
§ קשר מרווח, או קשר. עם סוג זה של מגע, קרומי הפלזמה של תאים שכנים מופרדים בפער של 2-3 ננומטר על פני מרחק של 0.5-3 מיקרון. במבנה של פלסמולים ממוקמים מתחמי חלבון מיוחדים (קונקסונים). קונקסון אחד על ממברנת הפלזמה של התא מנוגד בדיוק לקונקסון על קרום הפלזמה של התא הסמוך. כתוצאה מכך נוצרת ערוץ מתא אחד למשנהו. קונקסונים יכולים להתכווץ, לשנות את קוטר התעלה הפנימית, ובכך להשתתף בוויסות ההובלה של מולקולות בין תאים. סוג זה של חיבור נמצא בכל קבוצות הרקמות. התפקיד הפונקציונלי של צומת הפער הוא לשאת יונים ומולקולות קטנות מתא לתא. אז, בשריר הלב, עירור, המבוסס על תהליך של שינוי חדירות יונים, מועבר מתא לתא דרך הקשר.
§ מגע סינפטי, או סינפסה. סינפסות הן אזורי מגע בין שני תאים המתמחים להעברה חד-כיוונית של עירור או עיכוב מאלמנט אחד למשנהו. סוג זה של חיבור אופייני לרקמת עצב ומתרחש הן בין שני נוירונים והן בין נוירון לבין אלמנט אחר. הממברנות של תאים אלו מופרדות על ידי חלל בין תאי - שסע סינפטי ברוחב של כ-20-30 ננומטר. הממברנה באזור המגע הסינפטי של תא אחד נקראת פרה-סינפטית, השנייה - פוסט-סינפטית. ליד הממברנה הפרה-סינפטית, מתגלה מספר עצום של ואקוולים קטנים (שלפוחיות סינפטיות) המכילות את הנוירוטרנסמיטר. בזמן המעבר של הדחף העצבי, שלפוחיות סינפטיות מוציאות את הנוירוטרנסמיטר לתוך השסע הסינפטי. המתווך מקיים אינטראקציה עם אתרי הקולטנים של הממברנה הפוסט-סינפטית, מה שמוביל בסופו של דבר להעברת דחף עצבי. בנוסף להעברת דחף עצבי, סינפסות מספקות קשר נוקשה בין המשטחים של שני תאים המקיימים אינטראקציה.
§ פלסמודזמה.סוג זה של תקשורת בין-תאית נמצא בצמחים. Plasmodesmata הן תעלות צינוריות דקות המקשרות בין שני תאים סמוכים. הקוטר של ערוצים אלה הוא בדרך כלל 40-50 ננומטר. Plasmodesmata עוברים דרך דופן התא המפרידה בין התאים. בתאים צעירים, מספר הפלסמודסמטה יכול להיות גבוה מאוד (עד 1000 לתא). עם הזדקנות התאים, מספרם יורד עקב קרעים עם עלייה בעובי דופן התא. התפקיד הפונקציונלי של plasmodesmata הוא להבטיח זרימה בין-תאית של תמיסות המכילות חומרים מזינים, יונים ותרכובות אחרות. Plasmodesmata מדביקים תאים בנגיפים צמחיים.
מבנים מיוחדים של קרום הפלזמה
הפלזמה של תאי בעלי חיים רבים יוצרת תולדות של מבנים שונים (microvilli, cilia, flagella). לרוב נמצא על פני השטח של תאי בעלי חיים רבים microvilli.ליציאות אלה של הציטופלזמה, התחום על ידי הפלזמה, יש צורה של גליל עם חלק עליון מעוגל. Microvilli אופייניים לתאי אפיתל, אך נמצאים גם בתאים של רקמות אחרות. קוטר המיקרוווילי הוא כ-100 ננומטר. מספרם ואורכם שונים עבור סוגים שוניםתאים. המשמעות של microvilli טמונה בגידול משמעותי בשטח פני התא. זה חשוב במיוחד עבור תאים המעורבים בספיגה. אז, באפיתל המעי, יש עד 2x10 8 microvilli לכל 1 מ"מ 2 של פני השטח.
[הרצאה קודמת] [תוכן עניינים] [הרצאה הבאה]