מבנה תא הצמח
תא הצמח מורכב מקשיחים יותר או פחות דופן תאופרוטופלסט. דופן התא הוא דופן התא ו ממברנה ציטופלזמית. המונח פרוטופלסט מגיע מהמילה פרוטופלסמה, אשר במשך זמן רבנהג להתייחס לכל היצורים החיים. הפרוטופלסט הוא הפרוטופלסמה של תא בודד.
הפרוטופלסט מורכב מהציטופלזמה והגרעין. הציטופלזמה מכילה אברונים (ריבוזומים, מיקרוטובולים, פלסטידים, מיטוכונדריה) ומערכות ממברנות (רטיקולום אנדופלזמי, דיקטיוזומים). הציטופלזמה כוללת גם את המטריצה הציטופלזמית (חומר בסיסי) בה טבולים אברונים ומערכות ממברנות. הציטופלזמה מופרדת מדופן התא קרום פלזמה, שהוא קרום אלמנטרי. שלא כמו רוב תאי בעלי החיים, תאי צמחים מכילים ואקואול אחד או יותר. אלו שלפוחיות מלאות בנוזל ומוקפות בקרום אלמנטרי (טונופלסט).
בתא צמחי חי, החומר העיקרי נמצא ב בתנועה מתמדת. אברונים מעורבים בתנועה, הנקראת זרימת הציטופלזמה או ציקלוזה. ציקלוזיס מקל על תנועת החומרים בתא והחלפתם בין התא לסביבה.
קרום פלזמה. זהו מבנה פוספוליפיד דו-שכבתי. תאי צמחים מאופיינים בפלישות של קרום הפלזמה.
קרום הפלזמה מבצע את הפונקציות הבאות:
משתתף בחילופי חומרים בין התא לסביבה;
מתאם את הסינתזה וההרכבה של מיקרו-סיבי תאית של דופן התא;
מעביר אותות הורמונליים וחיצוניים השולטים בצמיחת תאים והתמיינות.
הליבה. זהו המבנה הבולט ביותר בציטופלזמה של תא איקריוטי. הקרנל מבצע שתי פונקציות חשובות:
שולט בפעילות החיונית של התא, וקובע אילו חלבונים, ובאיזו שעה יש לסנתז;
מאחסן מידע גנטי ומעביר אותו לתאי בת במהלך חלוקת התא.
הגרעין של תא איקריוטי מוקף בשתי ממברנות יסוד היוצרות את המעטפת הגרעינית. הוא חדור לנקבוביות רבות בקוטר של 30 עד 100 ננומטר, הנראות רק במיקרוסקופ אלקטרונים. לנקבוביות מבנה מורכב. קרום חיצוניהממברנה הגרעינית במקומות מסוימים משולבת עם הרשת האנדופלזמית. ניתן לראות את המעטפת הגרעינית כחלק מיוחד, מובחן מקומי, של הרשת האנדופלזמית (ER).
בגרעין מוכתם בצבעים מיוחדים, אפשר להבחין חוטים דקיםוגושים של כרומטין ונוקלאופלזמה (החומר העיקרי של הגרעין). הכרומטין מורכב מ-DNA הקשור לחלבונים מיוחדים הנקראים היסטונים. בתהליך חלוקת התא, הכרומטין הופך ליותר ויותר דחוס ומורכב לכרומוזומים. DNA מקודד מידע גנטי.
אורגניזמים שונים במספר הכרומוזומים בתאים סומטיים. לדוגמה, לכרוב יש - 20 כרומוזומים; חמניות - 34; חיטה - 42; בני אדם - 46, ואחד ממיני השרך אופיוגלוסום - 1250. לתאי מין (גמטות) יש רק מחצית ממספר הכרומוזומים האופייני לתאים סומטיים בגוף. מספר הכרומוזומים בגמטות נקרא הפלואיד (יחיד), בתאים סומטיים - דיפלואיד (כפול). תאים שיש להם יותר משתי קבוצות של כרומוזומים נקראים פוליפלואידים.
במיקרוסקופ אור ניתן לראות מבנים כדוריים - נוקלאולים. לכל גרעין יש גרעינים אחד או יותר, הנראים בגרעינים שאינם מתחלקים. RNA ריבוזומלי מסונתז בגרעין. בדרך כלל בגרעינים של אורגניזמים דיפלואידים יש שני גרעינים, אחד לכל אחד סט הפלואידיכרומוזומים. לנוקלאולים אין קרום משלהם. מבחינה ביוכימית, הנוקלאולי מאופיינים ריכוז גבוה RNA, שקשור כאן לפוספופרוטאינים. גודל הנוקלאולי תלוי ב מצב תפקודיתאים. הבחינו כי בתא הצומח במהירות, שבו מתרחשים תהליכים אינטנסיביים של סינתזת חלבון, נוקלאולי מתרבים בגודלם. בנוקלאולים מיוצרים mRNA וריבוזומים, הממלאים תפקיד סינתטי רק בגרעין.
נוקלאופלזמה (קריופלזמה) מיוצגת על ידי נוזל הומוגני שבו מומסים חלבונים שונים, כולל אנזימים.
פלסטידים. ואקוולים, קירות תאי תאית ופלסידים הם מרכיבים אופייניים לתאי צמחים. לכל פלסטיד יש מעטפת משלו, המורכבת משני ממברנות יסודיות. בתוך הפלסטיד, מערכת ממברנות מובחנת ו מעלות משתנותחומר הומוגני - סטרומה. פלסטידים בוגרים מסווגים על סמך הפיגמנטים שהם מכילים.
כלורופלסטים, שבהם מתרחשת פוטוסינתזה, מכילים כלורופילים וקרוטנואידים. בדרך כלל יש צורה של דיסק בקוטר של 4 - 5 מיקרון. בתא אחד של המזופיל (אמצע העלה) יכולים להיות 40 - 50 כלורופלסטים; ביריעות מ"מ 2 - כ-500,000. בציטופלזמה, כלורופלסטים ממוקמים בדרך כלל במקביל לממברנת התא.
המבנה הפנימי של הכלורופלסט מורכב. לסטרומה מחלחלת מערכת ממברנות מפותחת בצורת שלפוחיות - thylakoids. כל תילקואיד מורכב משני ממברנות. נוצרים תילקואידים מערכת בודדת. ככלל, הם נאספים בערימות - דגנים, הדומים לעמודי מטבעות. התילקואידים של גרנאים בודדים מחוברים ביניהם על ידי תילקואידים סטרומה, או תילקואידים בין-גרניים. כלורופיל וקרוטנואידים משובצים בממברנות התילקואיד. כלורופלסטים בצמחים ירוקים ובאצות מכילים לרוב גרגרי עמילן וטיפות שומנים קטנות (שומן). גרגרי עמילן הם מאגרים זמניים של מוצרי פוטוסינתזה. הם יכולים להיעלם מהכלורופלסטים של צמח שהיה בחושך רק 24 שעות, ולהופיע שוב 3–4 שעות לאחר העברת הצמח לאור.
כלורופלסטים הם אברונים חצי אוטונומיים ודומים לחיידקים. לדוגמה, לריבוזומים של חיידקים וכלורופלסטים יש דמיון גבוה למדי. הם קטנים יותר מריבוזומים אוקריוטיים. סינתזת חלבון על הריבוזומים של חיידקים וכלורופלסטים מעוכבת על ידי כלורמפניקול, שאין לו השפעה על תאים אוקריוטיים. בנוסף, גם לחיידקים וגם לכלורופלסטים יש סוג דומה של נוקלואיד, המאורגן בצורה דומה. למרות העובדה שיצירת כלורופלסטים וסינתזה של פיגמנטים הכלולים בהם נשלטת במידה רבה על ידי ה-DNA הכרומוזומלי של התא, בכל זאת, בהיעדר DNA משלו, כלורופלסטים אינם נוצרים.
כלורופלסטים יכולים להיחשב לאברוני התא העיקריים, שכן הם הראשונים בשרשרת הטרנספורמציה של אנרגיית השמש, וכתוצאה מכך האנושות מקבלת מזון ודלק. בכלורופלסטים מתרחשת לא רק פוטוסינתזה. הם גם מעורבים בסינתזה של חומצות אמינו ו חומצות שומן, משמשים כאחסון של עתודות זמניות של עמילן.
כרומופלסטים הם פלסטידים פיגמנטיים. מגוונים בצורתם, אין להם כלורופיל, אלא מסנתזים וצוברים קרוטנואידים, המעניקים צבע צהוב, כתום, אדום לפרחים, עלים ישנים, לפירות ולשורשים. כרומופלסטים יכולים להתפתח מכלורופלסטים, שבמקרה זה מאבדים כלורופיל ומבני קרום פנימיים, צוברים קרוטנואידים. זה מתרחש במהלך הבשלת פירות רבים. כרומופלסטים מושכים חרקים ובעלי חיים אחרים שאיתם הם התפתחו יחד.
לוקופלסטים הם פלסטידים שאינם פיגמנטים. חלקם מסנתזים עמילן (עמילופלסטים), אחרים מסוגלים להיווצר חומרים שונים, כולל שומנים וחלבונים. באור, לויקופלסטים הופכים לכלורופלסטים.
פרופלסטידים הם פלסטידים קטנים, חסרי צבע או ירוקים חיוורים, לא מובחנים המצויים בתאים המריסטמטיים (המתחלקים) של שורשים ויורים. הם המבשרים של פלסטידים אחרים, מובחנים יותר - כלורופלסטים, כרומופלסטים ואמינופלסטים. אם התפתחות פרוטופלסטידים מתעכבת בגלל חוסר אור, עלול להופיע בהם גוף פרולמלרי אחד או יותר, שהם הצטברויות חצי גבישיות של ממברנות צינוריות. פלסטידים המכילים גופים פרולמלרים נקראים אטיופלסטים. באור, האטיופלסטים הופכים לכלורופלסטים, בעוד שהקרומים של הגופים הפרולמלרים יוצרים תילקואידים. אטיופלסטים נוצרים בעלים של צמחים הנמצאים בחושך. פרוטופלסטים של עוברי זרעים הופכים תחילה לאטיופלסטים, שמהם מתפתחים כלורופלסטים באור. פלסטידים מאופיינים במעברים קלים יחסית מסוג אחד לאחר. פלסטידים, כמו חיידקים, מתרבים על ידי חלוקה לשניים. בתאים מריסטמטיים, זמן החלוקה של פרוטופלסטידים עולה בקנה אחד עם זמן חלוקת התא. עם זאת, בתאים בוגרים רובהפלסטיד נוצר כתוצאה מחלוקת פלסטידים בוגרים.
מיטוכונדריה. כמו כלורופלסטים, המיטוכונדריה מוקפת בשני ממברנות יסוד. הקרום הפנימי יוצר קפלים ובליטות רבות - cristae, אשר מגדילים משמעותית את פני השטח הפנימיים של המיטוכונדריה. הם קטנים משמעותית מפלסידים, בקוטר של כ-0.5 מיקרומטר, ומשתנים באורך ובצורה.
במיטוכונדריה מתרחש תהליך הנשימה, כתוצאה ממנו מתפרקות מולקולות אורגניות עם שחרור האנרגיה והעברתה למולקולות ATP, מאגר האנרגיה העיקרי של כל התאים האוקריוטיים. רוב תאי הצמח מכילים מאות או אלפי מיטוכונדריות. מספרם בתא אחד נקבע לפי הצורך של התא ב-ATP. המיטוכונדריה נמצאות בתנועה מתמדת, נעות מחלק אחד של התא לאחר, מתמזגות זו עם זו ומתחלקות. מיטוכונדריה בדרך כלל מתאספת במקום שבו יש צורך באנרגיה. אם קרום הפלזמה מעבירה חומרים באופן פעיל מתא לתא, אז המיטוכונדריה ממוקמות לאורך פני הממברנה. באצות חד-תאיות תנועתיות, מיטוכונדריה מצטברות בבסיס הדגל, ומספקות את האנרגיה הדרושה לתנועתן.
מיטוכונדריה, כמו פלסטידים, הם אברונים חצי אוטונומיים המכילים את הרכיבים הדרושים לסינתזה של החלבונים שלהם. הממברנה הפנימית מקיפה מטריצה נוזלית המכילה חלבונים, RNA, DNA, ריבוזומים דמויי חיידקים ומומסים שונים. DNA קיים בצורה של מולקולות מעגליות הממוקמות בנוקלואיד אחד או יותר.
בהתבסס על הדמיון של חיידקים למיטוכונדריה ולכלורופלסטים של תאים אוקריוטיים, ניתן להניח שהמיטוכונדריה והכלורופלסטים מקורם בחיידקים שמצאו "מפלט" בתאים הטרוטרופיים גדולים יותר - מבשרי האוקריוטים.
מיקרוגופים. שלא כמו פלסטידים ומיטוכונדריה, שמתוחמים בשתי ממברנות, גופי מיקרו הם אברונים כדוריים המוקפים בקרום אחד. למיקרוגופים יש תוכן גרגירי (גרגירי), לפעמים הם מכילים גם תכלילי חלבון גבישי. מיקרוגופים קשורים לאזור אחד או שניים של הרשת האנדופלזמית.
חלק מהמיקרוגופים, הנקראים פרוקסיזומים, משחקים תפקיד חשובבחילוף החומרים של חומצה גליקולית, הקשורה ישירות ל-photorespiration. בעלים ירוקים הם קשורים למיטוכונדריה ולכלורופלסטים. גופים מיקרוניים אחרים, הנקראים גליאוקסיזומים, מכילים את האנזימים הדרושים להמרת שומנים לפחמימות. זה קורה בהרבה זרעים במהלך הנביטה.
Vacuoles הם אזורים של התא התחום על ידי ממברנה, מלאים בנוזל - מוהל תאים. הם מוקפים בטונופלסט (ממברנה ואקוולרית).
תא צמחי צעיר מכיל מספר רב של ואקואולים קטנים, המתמזגים לכדי ואקואול אחד גדול ככל שהתא מתבגר. בתא בוגר, עד 90% מנפחו יכול להיות תפוס על ידי וואקואול. במקרה זה, הציטופלזמה נלחצת בצורה של שכבה היקפית דקה לממברנת התא. הגידול בגודל התא נובע בעיקר מצמיחת ה-vacuole. כתוצאה מכך, מתעורר לחץ טורגור וגמישות הרקמה נשמרת. זהו אחד התפקידים העיקריים של הוואקוול והטונופלסט.
המרכיב העיקרי של המיץ הוא מים, השאר משתנים בהתאם לסוג הצמח ומצבו הפיזיולוגי. Vacuoles מכילים מלחים, סוכרים, לעתים רחוקות יותר חלבונים. הטונופלסט ממלא תפקיד פעיל בהובלה והצטברות של יונים מסוימים ב-vacuole. ריכוז היונים במוהל התא יכול לעלות משמעותית על ריכוזו בסביבה. בְּ תוכן גבוהחלק מהחומרים יוצרים גבישים בוואקוולים. גבישי הסידן אוקסלט הנפוצים ביותר הם צורה שונה.
Vacuoles הם מקומות הצטברות של מוצרים מטבוליים (מטבוליזם). אלו יכולים להיות חלבונים, חומצות ואפילו חומרים רעילים לבני אדם (אלקלואידים). לעתים קרובות מופקדים פיגמנטים. כחול, סגול, סגול, אדום כהה, ארגמן נותנים לתאי הצמח פיגמנטים מקבוצת האנתוציאנין. בניגוד לפיגמנטים אחרים, הם מתמוססים היטב במים ונמצאים במוהל תאים. הם קובעים את הצבע האדום והכחול של ירקות רבים (צנוניות, לפת, כרוב), פירות (ענבים, שזיפים, דובדבנים), פרחים (פרחים, גרניום, דלפיניום, ורדים, אדמוניות). לפעמים פיגמנטים אלו מסווים את הכלורופיל בעלים, למשל במייפל האדום הדקורטיבי. אנתוציאנינים צובעים את עלי הסתיו צבע אדום בוהק. הם נוצרים במזג אוויר שמש קר, כאשר סינתזה של כלורופיל נעצרת בעלים. בעלים, כאשר לא נוצרים אנתוציאנינים, לאחר הרס הכלורופיל, הופכים קרוטנואידים צהובים-כתומים של כלורופלסטים בולטים. העלים בעלי צבע עז ביותר בסתיו קר וצלול.
ואקוולים מעורבים בהרס של מקרומולקולות, במחזור המרכיבים שלהן בתא. ריבוזומים, מיטוכונדריה, פלסטידים, נופלים ל-vacuoles, נהרסים. על פי פעילות עיכול זו, ניתן להשוות אותם לליזוזומים - אברונים של תאי בעלי חיים.
ואקוולים נוצרים מהרשת האנדופלזמית (רטיקולום)
ריבוזומים. חלקיקים קטנים (17 - 23 ננומטר), המורכבים מכמויות שוות בערך של חלבון ו-RNA. בריבוזומים, חומצות אמינו מתחברות ליצירת חלבונים. יש יותר מהם בתאים עם חילוף חומרים פעיל. ריבוזומים ממוקמים בציטופלזמה של התא באופן חופשי או מחוברים לרשת האנדופלזמית (80S). הם נמצאים גם בגרעין (80S), במיטוכונדריה (70S), בפלסטידים (70S).
ריבוזומים יכולים ליצור קומפלקס שעליו יש סינתזה סימולטנית של פוליפפטידים זהים, שמידע לגביו נלקח ממולקולה אחת ומ-RNA. קומפלקס כזה נקרא פוליריבוזומים (פוליזומים). לתאים המסנתזים חלבונים בכמויות גדולות יש מערכת נרחבת של פוליזומים המחוברים לרוב משטח חיצוניקונכיות של הגרעין.
רשת אנדופלזמית. זוהי מערכת ממברנות תלת מימדית מורכבת בהיקף בלתי מוגבל. בהקשר של ER נראה כמו שני ממברנות יסודיות עם רווח שקוף צר ביניהם. צורתו ואורכו של ER תלויים בסוג התא, בפעילותו המטבולית ובשלב ההתמיינות. בתאים המפרישים או מאחסנים חלבונים, ה-ER מעוצב כמו שקים שטוחים או בורות מים, עם ריבוזומים רבים המחוברים אל פני השטח החיצוניים שלו. רשת זו נקראת הרשת האנדופלזמית המחוספסת. ER חלק הוא בדרך כלל בצורת צינורית. רטיקולום אנדופלזמי מחוספס וחלק יכול להיות קיים באותו תא. ככלל, ישנם קשרים רבים ביניהם.
הרשת האנדופלזמית מתפקדת כמערכת התקשורת של התא. הוא קשור ל מעטפת חיצוניתגרעינים. למעשה, שני המבנים הללו יוצרים מערכת ממברנה אחת. כאשר המעטפת הגרעינית נקרעת במהלך חלוקת התא, השברים שלה דומים לשברי ER. הרשת האנדופלזמית היא מערכת להעברת חומרים: חלבונים, שומנים, פחמימות, לחלקים שונים של התא. רשת אנדופלזמה של תאים שכנים מחוברים דרך גדילים ציטופלזמיים - פלסמודסמטה - העוברים דרך ממברנות התא.
הרטיקולום האנדופלזמי הוא האתר העיקרי לסינתזה ממברנות תאים. בחלק מתאי הצמח, ממברנות של ואקוולים ומיקרוגופים, נוצרים כאן בורות של דיקטיוזומים.
מערכת גולג'י. מונח זה משמש להתייחס לכל הדיקטיוזומים, או גופי הגולגי, בתא. דיקטיוזומים הם קבוצות של שלפוחיות שטוחות בצורת דיסק או בורות מים המסתעפים לתוכם מערכת מורכבתצינורות. דיקטיוזומים בצמחים גבוהים מורכבים מ-4 - 8 בורות, שנאספו יחד.
בדרך כלל, בחפיסת טנקים, מבחינים בצד מתפתח ומתבגר. הממברנות של הבורות היוצרים דומים במבנה לממברנות ה-ER, והממברנות של הבורות המתבגרים דומים לממברנת הפלזמה.
דיקטיוזומים מעורבים בהפרשה, וברוב הצמחים הגבוהים, ביצירת קרומי התא. פוליסכרידים של דופן התא המסונתזים על ידי דיקטיוזומים מצטברים בשלפוחיות, הנפרדות לאחר מכן מבורות מתבגרים. שלפוחיות הפרשה אלו נודדות ומתמזגות עם ה-CMP; בעוד שהפוליסכרידים הכלולים בהם מובנים בתוך קרום התא. חלק מהחומרים המצטברים בדיקטיוזומים נוצרים במבנים אחרים, למשל ב-ER, ואז מועברים לדיקטיוזומים, שם הם משתנים (משונים) לפני ההפרשה. לדוגמה, גליקופרוטאינים הם מרכיב חשוב חומר בנייהקרום תא. חלק החלבון מסונתז על ידי פוליזומים של ה-ER המחוספס, חלק הפחמימות מסונתז בדיקטיוזומים, כאשר שני החלקים מתאחדים ליצירת גליקופרוטאין.
ממברנות הן מבנים דינמיים וניידים המשנים כל הזמן את צורתם ושטחם. הרעיון של המערכת האנדופלזמית מבוסס על ניידות של ממברנות. על פי תפיסה זו, הממברנות הפנימיות של הציטופלזמה, למעט ממברנות המיטוכונדריה והפלסטידים, הן שלם יחיד ומקורן ברטיקולום האנדופלזמי. בורות מים חדשים של דיקטיוזומים נוצרים מהרשת האנדופלזמית דרך שלב השלפוחית הביניימית, ושלפוחיות ההפרשה הנפרדות מהדיקטיוזומים תורמים בסופו של דבר להיווצרות קרום הפלזמה. לפיכך, הרטיקולום האנדופלזמי והדיקטיוזומים יוצרים שלם פונקציונלי שבו הדיקטיוזומים ממלאים תפקיד מבני בינייםבתהליך של המרת ממברנות הדומות לרשת האנדופלזמית לממברנות הדומות לפלזמה. ברקמות שהתאים שלהן גדלים ומתחלקים בצורה חלשה, מרכיבי הממברנה מתעדכנים כל הזמן.
מיקרוטובולים נמצאים כמעט בכל התאים האיקריוטיים. הם מבנים גליליים בקוטר של כ-24 ננומטר. אורכם משתנה. כל צינורית מורכבת מתת-יחידות של חלבון הנקרא טובולין. יחידות המשנה יוצרות 13 חוטים אורכיים המקיפים את החלל המרכזי. מיקרוטובולים הם מבנים דינמיים, הם נהרסים באופן קבוע ונוצרים בשלבים מסוימים של מחזור התא. הם מורכבים פנימה מקומות מיוחדיםהנקראים מרכזי ארגון מיקרוטובולים. בתאי צמחים, יש להם מבנה אמורפי בעל ביטוי חלש.
פונקציות של microtubules: להשתתף ביצירת קרום התא; להפנות בועות של דיקטיוזומים לממברנה היוצרות, כמו חוטי ציר שנוצרים בתא מתחלק; ממלאים תפקיד ביצירת לוחית התא (הגבול הראשוני בין תאי הבת). בנוסף, microtubules מרכיב חשוב flagella ו cilia, בתנועתם הם ממלאים תפקיד חשוב.
מיקרופילמנטים, כמו מיקרוטובולים, נמצאים כמעט בכל התאים האוקריוטיים. הם חוטים ארוכים בעובי 5-7 ננומטר, המורכבים מהחלבון המתכווץ אקטין. צרורות של מיקרופילמנטים נמצאים בתאים רבים של צמחים גבוהים יותר. ככל הנראה, הם ממלאים תפקיד חשוב בזרמי הציטופלזמה. מיקרופילמנטים, יחד עם מיקרוטובולים, יוצרים רשת גמישה הנקראת שלד ציטו.
החומר הטחון נחשב זה מכבר לתמיסה הומוגנית (הומוגנית) עשירה בחלבון עם מעט או ללא מבנים כלל. אולם, נכון להיום, באמצעות מיקרוסקופ אלקטרונים במתח גבוה, נקבע כי החומר הקרקע הוא סריג תלת מימדי הבנוי מחוטים דקים (קוטר 3-6 ננומטר) הממלאים את התא כולו. מרכיבים אחרים של הציטופלזמה, כולל מיקרוטובולים ומיקרופילמנטים, תלויים מהסריג המיקרו-טראבקולרי הזה.
המבנה המיקרוטרבקולרי הוא סריג של גדילי חלבון, שהחלל ביניהם מלא במים. יחד עם מים, לסריג יש עקביות של ג'ל; הג'ל נראה כמו גופים ג'לטינים.
אברונים מחוברים לסריג המיקרוטרבקולרי. הסריג מספק קישור בין חלקים נפרדיםתאים ומכוון הובלה תוך תאית.
טיפות שומנים הן מבנים כדוריים שנותנים גרנולריות לציטופלזמה תא צמחיתחת מיקרוסקופ אור. במיקרוגרפים של אלקטרונים, הם נראים אמורפיים. טיפות דומות מאוד אך קטנות יותר נמצאות בפלסידים.
טיפות ליפידים, בטעות שהן אברונים, קראו להן ספרוזומים והאמינו שהן מוקפות בקרום שכבה בודדת או כפולה. עם זאת, נתונים עדכניים מראים שלטיפות שומנים אין ממברנות, אך עשויות להיות מצופות בחלבון.
חומרים ארגסטיים הם "המוצרים הפסיביים" של הפרוטופלסט: חומרי רזרבה או פסולת. הם עשויים להופיע ולהיעלם פנימה תקופות שונותמחזור התא. מלבד גרגרי עמילן, גבישים, פיגמנטים אנתוציאנין וטיפות שומנים. אלה כוללים שרפים, מסטיקים, טאנינים וחלבונים. חומרים ארגאסטיים הם חלק מממברנת התא, החומר העיקרי של הציטופלזמה והאברונים, כולל ואקוולים.
Flagella ו-cilia הם מבנים דקים דמויי שיער המשתרעים מפני השטח של תאים אוקריוטיים רבים. יש להם קוטר קבוע, אבל האורך משתנה בין 2 ל 150 מיקרון. באופן קונבנציונלי, הארוכים והמעטים מהם נקראים דגלים, והקצרים והרבים יותר נקראים ריסים. אין הבדלים ברורים בין שני סוגי המבנים הללו, ולכן המונח flagellum משמש להתייחס לשניהם.
בחלק מהאצות והפטריות, הדגלים הם איברי תנועה, בעזרתם הם נעים במים. בצמחים (לדוגמה, טחבים, עצי כבד, שרכים, כמה ג'מנוספרמים), רק לתאי נבט (גמטות) יש דגלים.
לכל דגל יש ארגון ספציפי. טבעת חיצונית של 9 זוגות של מיקרו-צינוריות מקיפה שתי מיקרו-צינוריות נוספות הממוקמות במרכז הדגל. "ידיות" המכילות אנזים משתרעות ממיקרו-צינורית אחת של כל אחד מהזוגות החיצוניים. זהו דפוס הארגון הבסיסי של 9+2 שנמצא בכל הדגלים באורגניזמים איקריוטיים. מאמינים כי תנועת הפלגלה מבוססת על החלקה של מיקרוטובולים, בעוד שהזוגות החיצוניים של המיקרוטובולים נעים אחד לאורך השני ללא התכווצות. החלקה של זוגות מיקרוטובולים זה לזה גורמת לכיפוף מקומי של הדגל.
דגלים "צומחים" ממבנים גליליים ציטופלזמיים הנקראים גופים בזאליים, המהווים גם את החלק הבסיסי של הדגל. לגופים הבסיסיים יש מבנה פנימי, הדומה למבנה של דגלון, אלא שהצינורות החיצוניים נאספים בשלישיות, ולא בזוגות, והצינורות המרכזיים נעדרים.
דופן תא. דופן התא תוחמת את גודל הפרוטופלסט ומונעת את קריעתו עקב ספיגת המים על ידי הוואקואול.
לדופן התא יש תפקידים ספציפיים שחשובים לא רק לתא ולרקמה שבה נמצא התא, אלא גם לצמח כולו. לדפנות התא תפקיד חיוני בספיגה, הובלה והפרשה של חומרים, ובנוסף, ניתן לרכז בהם פעילות ליזוזומלית, או עיכול.
רכיבי דופן התא. המרכיב האופייני ביותר של דופן התא הוא תאית, שקובעת במידה רבה את הארכיטקטורה שלו. מולקולות תאית מורכבות ממולקולות גלוקוז חוזרות המחוברות מקצה לקצה. מולקולות תאית דקות ארוכות משולבות למיקרו-סיביים בעובי של 10 - 25 ננומטר. מיקרופיברילים משתלבים זה בזה ויוצרים חוטים דקים, שבתורם ניתן ללפף אחד סביב השני, כמו חוטים בחבל. לכל "חבל", או מקרופיבריל כזה, עובי של כ-0.5 מיקרון, מגיע לאורך של 4 מיקרון. מקרופיברילים חזקים כמו חוטי פלדה בגודל שווה.
מסגרת התאית של דופן התא מלאה במולקולות מטריצת תאית השזורות בה. הוא מורכב מפוליסכרידים הנקראים hemicelluloses, ו-pectins, או pectins, כימית קרוב מאוד hemicelluloses.
מרכיב נוסף בדופן התא, ליגנין, הוא הפולימר הנפוץ ביותר בתאי הצמח לאחר תאית. לגנין מגביר את קשיחות הדופן ונמצא בדרך כלל בתאים המבצעים פונקציה תומכת או מכנית.
קוטין, סוברין, שעווה - בדרך כלל מופקדים בקונכיות של רקמות המגן של צמחים. קוטין, למשל, נמצא בקרומי התא של האפידרמיס, וסוברין נמצא ברקמת ההגנה המשנית, הפקק. שני החומרים מצויים בשילוב עם שעווה ומונעים איבוד יתר של מים מהצמח.
שכבות של דופן התא. עובי הדופן של תאי הצמח משתנה מאוד, בהתאם לתפקידם של התאים במבנה הצמח וגיל התא עצמו. תַחַת מיקרוסקופ אלקטרונישתי שכבות נראות בדופן התא הצמחי: הלמינה החציונית (המכונה גם החומר הבין-תאי), וקיר התא הראשוני. תאים רבים מניחים שכבה נוספת - המשנית דופן תא. הצלחת החציונית ממוקמת בין הקירות הראשוניים של תאים סמוכים. הקיר המשני, אם בכלל, מופקד על ידי פרוטופלסט התא על פני השטח הפנימיים של דופן התא הראשוני.
צלחת אמצעית. הלמינה החציונית מורכבת בעיקר מפקטין. במקום שבו צריך להיווצר דופן תא, בין שני תאים שזה עתה נוצרו, מציינים לראשונה מקלעת צפופה של צינוריות של הרשת האנדופלזמית והבור של מנגנון הגולגי (דיקטיוזומים). ואז מופיעות בועות במקום הזה, מלאות בפקטין (מפוליסכרידים). הבועות הללו מופרדות מהמיכלים של מנגנון גולגי. דופן התא המוקדם מכיל פוליסכרידים שונים, כאשר העיקריים שבהם הם פקטין והמיצלולוז. מאוחר יותר, הוא מורכב מחומרים צפופים יותר - תאית וליגנין.
דופן תא ראשונית. זוהי שכבה של מעטפת תאית המונחת לפני או במהלך צמיחת התא. בנוסף לתאית, המיצלולוזה ופקטין, הממברנות הראשוניות מכילות גליקופרוטאין. קונכיות ראשוניות יכולות להתגרות. רכיב הפקטין מקנה פלסטיות, המאפשרת לקליפה הראשונית להתרחב ככל שהשורש, הגבעול או העלה מתארכים.
לתאים המתחלקים באופן פעיל (רוב התאים הבוגרים המעורבים בתהליכי הפוטוסינתזה, הנשימה וההפרשה) יש ממברנות ראשוניות. תאים כאלה עם ממברנה ראשונית ופרוטופלסט חי מסוגלים לאבד צורה אופיינית, מחלקים ומתמיינים לסוג תא חדש. הם מעורבים בריפוי פצעים והתחדשות רקמות בצמחים.
קרומי תאים ראשוניים אינם אחידים בעובי לכל אורכם, אך יש להם חתכים דקים, הנקראים שדות נקבוביות ראשוניים. גדילי הציטופלזמה, או plasmodesmata, המחברים את הפרוטופלסטים של תאים שכנים בדרך כלל עוברים דרך שדות הנקבוביות הראשוניים.
דופן תא משני. למרות העובדה שלתאים צמחיים רבים יש רק דופן ראשונית, בחלקם, הפרוטופלסט מפקיד דופן משנית לכיוון מרכז התא. זה מתרחש בדרך כלל לאחר הפסקת צמיחת התאים ושטח הממברנה הראשונית כבר לא גדל. מסיבה זו, הקליפה המשנית שונה מהראשונית. ממברנות משניות נחוצות במיוחד על ידי תאים מיוחדים המחזקים את הצמח ומוליכים מים. לאחר שקיעת הממברנה המשנית, הפרוטופלסט של תאים אלה, ככלל, מת. יש יותר תאית בקליפות המשניות מאשר בראשוניות, ונעדרים בהן חומרי פקטין וגליקופרוטאין. קשה למתוח את הממברנה המשנית, המטריצה שלו מורכבת מהמיצלולוזה.
במעטפת המשנית ניתן להבחין בשלוש שכבות - חיצוניות, אמצעיות ופנימיות (S 1, S 2, S 3). מבנה השכבות של הקונכיות המשניות מגדיל משמעותית את חוזקן. מיקרו-סיבי תאית בממברנה המשנית מופקדים צפופים יותר מאשר בראשי. ליגנין - רכיב רגילקונכיות משניות של עץ.
הנקבוביות בממברנות של תאים מגע ממוקמות זו מול זו. שתי נקבוביות מנוגדות וקרום נקבוביות יוצרות זוג נקבוביות. בתאים עם ממברנות משניות, ישנם שני סוגים עיקריים של נקבוביות: פשוטות ותוחמות. בנקבוביות תוחמות, הקיר המשני תלוי מעל חלל הנקבוביות. בנקבוביות פשוטות, זה לא המקרה.
צמיחת דופן התא. ככל שהתא גדל, העובי והשטח של קרום התא גדלים. מתיחת מעטפת היא תהליך מורכב. הוא נשלט על ידי הפרוטופלסט ומווסת על ידי הורמון אוקסין.
בתאים הגדלים באופן אחיד לכל הכיוונים, שקיעת המיופיברילים היא אקראית. מיופיברילים אלה יוצרים רשת לא סדירה. תאים כאלה נמצאו בליבת הגזע, ברקמות האחסון ובמהלך גידול תאים במבחנה. בתאים מתארכים, המיופיברילים של המעטפת הצדדית מופקדים בזווית ישרה לציר ההתארכות.
חומרים מטריקס - פקטין, hemicelluloses וגליקופרוטאינים מועברים לממברנה בשלפוחיות של dictyosomes. יחד עם זאת, פקטין אופייניים יותר לתאים גדלים, בעוד שהמיצלולוזות שולטות בתאים שאינם גדלים.
מיקרו-סיבי תאית מסונתזים על פני התא בעזרת קומפלקס אנזים הקשור לממברנת הפלזמה. הכיוון של מיקרופיברילים נשלט על ידי מיקרוטובולים הממוקמים ב משטח פנימיקרום פלזמה.
פלסמודזמה. אלו הם חוטים דקים של הציטופלזמה המחברים את הפרוטופלסטים של תאים שכנים. Plasmodesmata עוברים דרך דופן התא בכל מקום, או מרוכזים בשדות נקבוביות ראשוניים או בממברנות בין זוגות של נקבוביות. תחת מיקרוסקופ אלקטרונים, פלסמודסמטה מופיעות כתעלות צרות המרופדות בקרום הפלזמה. לאורך ציר התעלה משתרע מתא אחד למשנהו צינור גלילי קטן יותר, ה-desmotubule, המתקשר עם הרשת האנדופלזמית של שני התאים הסמוכים. פלסמודסמטה רבות נוצרות במהלך חלוקת התא, כאשר הרטיקולום האנדופלזמי הצינורי נקלט על ידי לוחית התא המתפתחת. Plasmodesmata יכול להיווצר גם בממברנות של תאים שאינם מתחלקים. מבנים אלה מספקים העברה יעילה של חומרים מסוימים מתא לתא.
חלוקת תא. בְּ אורגניזמים רב תאייםחלוקת תאים יחד עם עלייה בגודלם היא דרך לצמיחה של האורגניזם כולו. תאים חדשים שנוצרים במהלך החלוקה דומים במבנה ובתפקוד, הן לתא האב והן זה לזה. ניתן לחלק את תהליך החלוקה באאוקריוטים לשני שלבים חופפים חלקית: מיטוזה וציטוקינזיס.
מיטוזיס היא היווצרות של שני גרעיני בת מגרעין אחד, שוות ערך זה לזה מבחינה מורפולוגית וגנטית. ציטוקינזיס היא חלוקת החלק הציטופלזמי של התא עם היווצרות תאי בת.
מחזור התא. תא חי עובר סדרה של אירועים עוקבים המרכיבים את מחזור התא. משך המחזור עצמו משתנה בהתאם לסוג התא ו גורמים חיצוניים, למשל, על טמפרטורה או אבטחה חומרים מזינים. בדרך כלל המחזור מחולק ל-interphase ולארבעה שלבים של מיטוזה.
Interphase. התקופה שבין חלוקות מיטוטיות עוקבות.
האינטרפאזה מחולקת לשלוש תקופות, המסומנות כ-G 1, S, G 2.
במהלך תקופת G 1, שמתחילה לאחר מיטוזה. במהלך תקופה זו, כמות הציטופלזמה עולה, כולל אברונים שונים. בנוסף, על פי ההשערה המודרנית, במהלך תקופת G 1, מסונתזים חומרים שמעוררים או מעכבים את תקופת S ואת שאר המחזור, ובכך קובעים את תהליך החלוקה.
תקופת S עוקבת אחר תקופת G 1, שבה החומר הגנטי (DNA) משוכפל.
במהלך תקופת G2, העוקבת אחרי S, נוצרים המבנים המעורבים ישירות במיטוזה, כגון רכיבי ציר.
חלק מהתאים עוברים מספר בלתי מוגבל של מחזורי תאים. זֶה אורגניזמים חד תאייםוכמה תאים של אזורי גדילה פעילים (מריסטמים). כמה תאים מיוחדים לאחר התבגרות מאבדים את יכולתם להתרבות. הקבוצה השלישית של תאים, למשל, אלו היוצרים רקמת פצע (קלוס), שומרת על היכולת להתחלק רק בתנאים מיוחדים.
מיטוזה, או ביקוע גרעיני. זהו תהליך מתמשך המחולק לארבעה שלבים: פרופאזה, מטאפאזה, אנפאזה, טלופזה. כתוצאה מהמיטוזה, החומר הגנטי שהכפיל את עצמו ב-interphase מחולק שווה בשווה בין שני גרעיני הבת.
אחד ה סימנים מוקדמיםהמעבר של תא לחלוקה הוא הופעתה של חגורה צרה בצורת טבעת של מיקרוטובולים ישירות מתחת לקרום הפלזמה. חגורה צפופה יחסית זו מקיפה את הגרעין במישור המשווני של הציר המיטוטי העתידי. מכיוון שהיא מופיעה לפני הפרופזה, היא נקראת חגורת הקדם-פרופאזה. הוא נעלם לאחר הציר המיטוטי, הרבה לפני הופעתו בטלופזה המאוחר של לוחית התא, הגדל מהמרכז לפריפריה ומתמזג עם קרום תא האם באזור שנכבש בעבר על ידי חגורת הקדם-פרופאזה.
פרופאזה. בתחילת הפרופזה, הכרומוזומים דומים לחוטים ארוכים המפוזרים בתוך הגרעין. לאחר מכן, כשהגדילים מתקצרים ומתעבים, אפשר לראות שכל כרומוזום מורכב לא אחד, אלא משני גדילים שלובים זה בזה, הנקראים כרומטידים. בפרופאזה המאוחרת, שתי הכרומטידות הזוגיות המקוצרות של כל כרומוזום שוכבות זו לצד זו במקביל, מחוברות בקטע צר הנקרא צנטרומר. יש לו מיקום ספציפי על כל כרומוזום והוא מחלק את הכרומוזום לשתי זרועות באורכים שונים.
מיקרו-צינוריות ממוקמות במקביל לפני השטח של הגרעין לאורך ציר הציר. זהו הביטוי המוקדם ביותר של הרכבת ציר מיטוטי.
בסוף הפרופזה, הגרעין מאבד בהדרגה את קווי המתאר הברורים שלו ולבסוף נעלם. זמן קצר לאחר מכן מתפרקת גם המעטפת הגרעינית.
מטאפאזה. בתחילת המטאפאזה, הציר, שהוא מבנה תלת מימדי, הרחב ביותר באמצע ומצטמצם לכיוון הקטבים, תופס את המקום שתפס בעבר הגרעין. סיבי הציר הם צרורות של microtubules. במהלך המטאפאזה, הכרומוזומים, שכל אחד מהם מורכב משתי כרומטידות, מסודרים כך שהצנטרומרים שלהם נמצאים במישור המשווני של הציר. עם הצנטרומר שלו, כל כרומוזום מחובר לחוטי הציר. עם זאת, כמה גדילים עוברים מקוטב אחד למשנהו מבלי להיות מחוברים לכרומוזומים.
כאשר כל הכרומוזומים ממוקמים במישור המשווני, המטאפאזה הושלמה. כרומוזומים מוכנים להתחלק.
אנפאזה. הכרומטידות של כל כרומוזום נפרדות. עכשיו אלה כרומוזומי הבת. קודם כל, הצנטרומר מתחלק, ושני כרומוזומי הבת נמשכים לקטבים מנוגדים. במקרה זה, הצנטרומרים נעים קדימה, וזרועות הכרומוזומים נמתחות מאחור. חוטי הציר המחוברים לכרומוזומים מתקצרים, ותורמים להפרדת הכרומטידים ולתנועת כרומוזומי הבת בכיוונים מנוגדים.
טלופאז. בטלופאזה, ההפרדה של שתי קבוצות זהות של כרומוזומים הושלמה, בעוד שסביב כל אחת מהן נוצר קרום גרעיני. הרשת המחוספסת לוקחת חלק פעיל בכך. מנגנון הציר נעלם. במהלך הטלופאז, הכרומוזומים מאבדים את בהירות המתאר שלהם, נמתחים, הופכים בחזרה לחוטים דקים. הנוקלאולים משוחזרים. כאשר הכרומוזומים הופכים בלתי נראים, המיטוזה הושלמה. שני גרעיני הבת נכנסים לשלבים. הם שקולים מבחינה גנטית זה לזה ולגרעין האב. זה חשוב מאוד, שכן יש להעביר את התוכנית הגנטית, ואיתה כל הסימנים, לאורגניזמים הבתים.
משך המיטוזה משתנה אורגניזמים שוניםוזה תלוי בסוג הבד. עם זאת, הפרופזה היא הארוכה ביותר והאנפאזה היא הקצרה ביותר. בתאי קצה השורש, משך הפרופזה הוא 1-2 שעות; מטאפאזה - 5 - 15 דקות; אנפאזה - 2 - 10 דקות; טלופאז - 10 - 30 דקות. משך האינטרפאז הוא בין 12 ל-30 שעות.
בתאים איקריוטים רבים, מרכזי ארגון המיקרוטובוליים האחראים על היווצרות הציר המיטוטי קשורים לצנטריולים.
ציטוקינזיס. זהו תהליך החלוקה של הציטופלזמה. ברוב האורגניזמים, התאים מתחלקים על ידי נסיגת דופן התא ויצירת תלם ביקוע, אשר מעמיק בהדרגה, ודוחס את שאר החוטים של הציר המיטוטי. בכל הצמחים (בריופיטים וצמחי כלי דם) ובחלק מהאצות, תאים מתחלקים עקב היווצרות לוחית תא.
בטלופאז המוקדם נוצרת מערכת סיבים בצורת חבית הנקראת פרגמופלסט בין שני גרעיני הבת. סיבי הפרגמופלסט, כמו סיבי הציר המיטוטי, מורכבים ממיקרוטובולים. טיפות קטנות מופיעות במישור המשווני של הפרגמופלסט. הם מתמזגים ויוצרים לוחית תא שגדלה עד שהיא מגיעה לדופן התא המתחלק. זה משלים את ההפרדה של שני תאי הבת. טיפות מתמזגות הן בועות המתנתקות ממנגנון גולגי. בעיקרון, הם מכילים חומרי פקטין, שמהם נוצרת הצלחת האמצעית. ממברנות שלפוחית מעורבות בבניית קרום הפלזמה משני צידי הצלחת. במקביל, plasmodesmata נוצרים משברים של הרשת האנדופלזמית הצינורית.
לאחר היווצרות הצלחת החציונית, כל פרוטופלסט מפקיד עליו קרום ראשוני. בנוסף, כל תא בת מפקיד שכבה חדשה של ממברנה סביב הפרוטופלסט כולו, שממשיכה את הממברנה שעלתה מלוחית התא. המעטפת המקורית של תא האב נהרסת ככל שתאי הבת גדלים.
סוגים שונים של מיטוזה איקריוטית
חלוקת התא שתוארה לעיל בצמחים, וגם בבעלי חיים, אינה הצורה היחידה של חלוקת תאים עקיפה. הסוג הפשוט ביותר של מיטוזה הוא pleuromitosis. זה דומה לחלוקה בינארית תאים פרוקריוטיים, שבו הנוקלואידים לאחר שכפול נשארים קשורים לממברנת הפלזמה. הממברנה מתחילה לצמוח בין נקודות הקישור של ה-DNA ובכך מפיצה את הכרומוזומים לתוך אזורים שוניםתאים. לאחר מכן, במהלך היווצרות התכווצות התא, כל אחת ממולקולות ה-DNA תהיה בתא נפרד חדש.
אופייני לחלוקה של תאים אוקריוטיים הוא היווצרות של ציר הבנוי ממיקרוטובוליות. עם pleuromitosis סגור (זה נקרא סגור מכיוון שההתבדלות של הכרומוזומים מתרחשת מבלי לשבור את המעטפת הגרעינית), לא צנטריולים, אלא מבנים אחרים הממוקמים על בְּתוֹךממברנה גרעינית. אלה הם מה שנקרא גופים קוטביים של מורפולוגיה בלתי מוגדרת, שמהם משתרעים מיקרוטובולים. יש שניים מהגופים האלה. הם נפרדים זה מזה מבלי לאבד את הקשר איתם מעטפת גרעין. כתוצאה מכך, נוצרים שני חצאי צירים הקשורים לכרומוזומים. כל התהליך של יצירת ציר מיטוטי והפרדת כרומוזומים במקרה זה מתרחש מתחת למעטפת הגרעינית. סוג זה של מיטוזה מתרחש בקרב פרוטוזואה ומופץ באופן נרחב בפטריות (chytridia, zygomycetes, שמרים, oomycetes, ascomycetes, myxomycetes, וכו'). יש צורות של pleuromitosis סגורה למחצה, כאשר המעטפת הגרעינית נהרסת בקטבים של הציר שנוצר.
הצורה הבאה של מיטוזה היא אורתומיטוזיס. במקרה זה, ה-COMTs ממוקמים בציטופלזמה; מההתחלה, לא נוצרים חצאי צירים, אלא ציר דו-קוטבי. ישנן שלוש צורות של אורתומיטוזיס (מיטוזה רגילה), חצי סגורה וסגורה. באורתומיטוזיס סגורה למחצה, נוצר ציר ביסימטרי בעזרת TsOMT הממוקם בציטופלזמה, המעטפת הגרעינית נשמרת לאורך כל המיטוזה, למעט אזורי הקוטב. מסות של חומר גרגירי, או אפילו צנטריולים, עשויות להתגלות כ-COMT. צורה זו של מיטוזה נמצאת ב-zoospores של אצות ירוקות, חומות, אדומות, בחלקן פטריות תחתונותוגרגרינה. עם אורתומיטוזיס סגור, הממברנה הגרעינית נשמרת לחלוטין, שבה נוצר ציר אמיתי. מיקרוטובולים נוצרים בקאריופלזמה, לעתים רחוקות יותר הם גדלים מה-TsOMT הפנימי, שאינו קשור (בניגוד לפלורומיטוזיס) עם הממברנה הגרעינית. סוג זה של מיטוזה אופייני לחלוקה של מיקרו-גרעיני ריסים, אך יכול להתרחש גם בפרוטוזואה. באורתומיטוזיס פתוח, המעטפת הגרעינית מתפרקת לחלוטין. סוג זה של חלוקת תאים אופייני לאורגניזמים של בעלי חיים, כמה פרוטוזואה ותאים של צמחים גבוהים יותר. צורה זו של מיטוזה, בתורה, מיוצגת על ידי טיפוסים אסטרליים ואנסטרליים.
מהסקירה הקצרה, ברור ש תכונה עיקריתמיטוזיס באופן כללי היא הופעת מבנים של ציר הביקוע, שנוצר בקשר עם TsOMT, שהוא מגוון במבנה.
מורפולוגיה של הדמות המיטוטית
המנגנון המיטוטי בא לידי ביטוי טוב במיוחד בשלב המטאפאזה של המיטוזה. במטאפאזה, במישור המשווני של התא, ממוקמים כרומוזומים, שמהם נמשכים חוטי הציר המכונים בכיוונים מנוגדים, ומתכנסים בשני קטבים שונים של הדמות המיטוטית. לפיכך, הציר המיטוטי הוא אוסף של כרומוזומים, קטבים וסיבים. סיבי הציר הם מיקרוטובולים בודדים או צרורותיהם. מיקרו-צינוריות מתחילות מקטבי הציר, וחלקן מגיעות אל הצנטרומרים, שם נמצאים ה-kinetochores של הכרומוזומים (kinetochore microtubules), חלקם הולכים יותר לכיוון הקוטב הנגדי, אך אינם מגיעים אליו. הם נקראים "מיקרוטובולים בין קוטביים". מהקטבים יוצאת קבוצה של מיקרוטובולים רדיאליים, ויוצרת סביבם מבנה הדומה ל"זוהר קורן" – אלו הן מיקרוטובולים אסטרליים.
לפי מורפולוגיה, דמויות מיטוטיות מחולקות לסוגים אסטרליים ואנסטרליים.
סוג הציר האסטרלי, או מתכנס, מאופיין בכך שהקטבים שלו מיוצגים על ידי אזור קטן שאליו מתכנסות (מתכנסות) המיקרוטובולים. בדרך כלל, צנטרוזומים המכילים צנטרולים ממוקמים בקטבים של הצירים האסטרליים. עם זאת, ידועים מקרים של מיטוזות אסטרליות צנטרולריות (במהלך מיוזה של חלק מחסרי חוליות). בנוסף, מציינים מיקרוטובולים רדיאליים המקרינים מהקטבים, שאינם חלק מהציר, אלא יוצרים אזורי כוכבים - ציסטרים. סוג זה של חלוקה מיטוטית דומה למשקולת. לסוג האנסטריאלי של הדמות המיטוטית אין ציסטרים בקטבים. האזורים הקוטביים של הציר כאן רחבים, הם נקראים כובעים קוטביים, הם כוללים centrioles. במקרה זה, סיבי הציר אינם יוצאים מנקודה אחת, אלא מתפצלים בחזית רחבה (מתפצלים) מכל אזור כובעי הקוטב. סוג זה של ציר מאפיין תאים מתחלקים של צמחים גבוהים יותר, אך ניתן למצוא אותו גם בבעלי חיים גבוהים יותר. בעוברות מוקדמת של יונקים, מיטוזות צנטרולריות (מתפצלות) נצפות במהלך חלוקת הבשלת הביציות ובחלוקות I ו-II של הזיגוטה. אבל כבר בחלוקת התא השלישי ובכל התאים שלאחר מכן, הם מתחלקים בהשתתפות צירים אסטרליים, שבקטבים שלהם נמצאים תמיד צנטרולים. באופן כללי, עבור כל צורות המיטוזה, הכרומוזומים עם הקינטופורים שלהם, הגופים הקוטביים (צנטרוזומים) וסיבי הציר נשארים מבנים נפוצים.
צנטרומרים וקינטוכורים
ייתכן שיש לצנטרומרים לוקליזציה שונהלאורך הכרומוזומים. צנטרומרים הולוצנטריים מתרחשים כאשר מיקרוטובולים מקושרים לאורך כל הכרומוזום (חלק מהחרקים, נמטודות, חלק מהצמחים). צנטרומרים מונוצנטריים - כאשר מיקרוטובולים קשורים לכרומוזומים באתר אחד. צנטרומרים מונוצנטריים יכולים להיות מנוקדים (לדוגמה, בחלק מהשמרים הניצנים), כאשר רק מיקרו-צינורית אחת מתקרבת לקינטוכור, וזונאלית, שבה צרור מיקרו-צינוריות מתקרב לקינטוכור המורכב. למרות המגוון של אזורי הצנטרומרים, כולם קשורים אליהם מבנה מורכב kinetochore, שיש לו דמיון בסיסי במבנה ובתפקוד בכל האיקריוטים. Kinetochores הם מבני חלבון מיוחדים לרובממוקם בצנטרומרים של הכרומוזומים. אלו קומפלקסים מורכבים המורכבים מחלבונים רבים. מבחינה מורפולוגית, הם דומים מאוד, בעלי אותו מבנה, החל מדיאטומים לבני אדם. הם מבנים תלת-שכבתיים: השכבה הצפופה הפנימית הצמודה לגוף הכרומוזום, השכבה האמצעית הרפויה והשכבה הצפופה החיצונית. סיבים רבים משתרעים מהשכבה החיצונית, ויוצרים את מה שנקרא הכתר הסיבי של הקינטוכור. IN צורה כלליתלקינטוכורים יש צורה של לוחות או דיסקים השוכבים באזור ההתכווצות הראשונית של הכרומוזום, בצנטרומר. בדרך כלל יש קינטוכור אחד לכל כרומוזום או כרומטיד. לפני אנפאזה, קינטוכורים על כל כרומטיד אחות ממוקמים, כל אחד מתקשר עם צרור המיקרוטובולים שלו. בצמחים, הקינטוכור אינו נראה כמו לוחות, אלא כחצי כדור. התפקיד הפונקציונלי של קינטוכורים הוא לקשור כרומטידות אחיות זו לזו, לקבע מיקרוטובולים מיטוטיים, לווסת הפרדת כרומוזומים ולמעשה להזיז כרומוזומים במהלך מיטוזה בהשתתפות מיקרוטובולים. באופן כללי, מבני חלבון, קינטוכורים, מוכפלים בתקופת S, במקביל להכפלת הכרומוזומים. אבל החלבונים שלהם נמצאים על הכרומוזומים בכל תקופות מחזור התא.
רקמת צמחים
בדים חינוכיים(מריסטמים)
רקמות חינוכיות בגוף הצמחים ממוקמות ב מקומות שונים, אז הם מחולקים לקבוצות הבאות (איור 0; 1).
1. מריסטמות אפיקליות (אפיקליות) ממוקמות בחלק העליון, או הקודקודים, של האיברים הציריים - גזע, שורש. בעזרת המריסטמים הללו, האיברים הצומחיים של הצמחים מבצעים את צמיחתם לאורך.
2. מריסטמים רוחביים אופייניים לאיברים צירים. שם הם ממוקמים באופן קונצנטרי, בצורה של מצמד.
3. מריסטמים בין קלוריות, או בין קלוריות, מקורם במריסטמים האפיקליים. אלו קבוצות של תאים שעדיין לא מסוגלות להתרבות, אבל יצאו לדרך של התמיינות. אין ביניהם תאים ראשוניים, אבל יש הרבה תאים מיוחדים.
4. מריסטמים פצעים מספקים שיקום של החלק הפגוע בגוף. התחדשות מתחילה עם דה-דיפרנציאציה, כלומר, התפתחות הפוכה מתאי מיוחדים לתאים מריסטמטיים. הם הופכים לפלוגן, היוצר פקק המכסה את פני הפצע. תאים לא מובחנים, מתחלקים, יכולים ליצור רקמה פרנכימלית רופפת - קאלוס. ממנו, בתנאים מסוימים, נוצרים איברי צמחים.
רקמות אינטגמנטרי
הם פועלים כמחסום גבול, המפריד בין הרקמות הבסיסיות סביבה. המרכיבים העיקריים של צמח מורכבים רק מתאי חיים. אינטגמנט משני ושלישוני - בעיקר מהמתים עם דפנות תאים עבות.
התפקודים העיקריים של רקמות המוח:
הגנה על הצמח מפני ייבוש;
・הגנה מפני כניסה מיקרואורגניזמים מזיקים;
· הגנה מפני כֶּלֶף;
· הגנה מפני נזק מכני;
ויסות חילוף החומרים בין הצמח לסביבה;
תפיסה של גירוי.
הרקמה העיקרית של האפידרמיס היא האפידרמיס, האפידרמיס. מורכב מתאי חיים. נוצר ממריסטמים אפיקליים. מכסה גבעולים ועלים צעירים.
האפידרמיס נוצר בצמחים בקשר ליציאה מבית הגידול המימי ליבשה על מנת למנוע את התייבשותו. בנוסף לסטומטה, כל תאי האפידרמיס מחוברים זה לזה. הקירות החיצוניים של התאים הראשיים עבים יותר מהשאר. כל פני השטח מכוסים בשכבת קאטין ושעוות ירקות. שכבה זו נקראת הקוטיקולה (עור). הוא נעדר על השורשים הגדלים וחלקים תת-מימיים של צמחים. כאשר הוא יבש, החדירות של הציפורן נחלשת באופן משמעותי.
בנוסף לתאים העיקריים, ישנם אחרים באפידרמיס, במיוחד שערות או טריכומות. הם חד-תאיים ורב-תאיים (איור 2). מבחינה פונקציונלית, הם מגדילים את פני האפידרמיס, למשל, באזור הצמיחה של השורשים, משמשים כהגנה מכנית, נצמדים לתמיכה ומפחיתים את איבוד המים. למספר צמחים יש שערות בלוטות, כמו סרפדים.
רק לצמחים גבוהים יותר יש סטמטות באפידרמיס, המווסתות את חילופי המים והגזים. אם אין לציפורן, אז אין צורך בסטומטה. סטומות הן קבוצת תאים היוצרים את המנגנון הסטומטלי, המורכב משני תאים נגררים ותאי אפידרמיס סמוכים - תאי צד. הם שונים מתאי האפידרמיס העיקריים (איור 3). תאי שמירה שונים מהתאים הסובבים אותם בצורתם ובנוכחותם של מספר רב של כלורופלסטים וקירות מעובים בצורה לא אחידה. אלה הפונים זה לזה עבים יותר מהשאר (איור 4). בין תאי השמירה נוצר מרווח סטמאטלי המוביל לחלל תת-סטומטי הנקרא חלל התת-סטומטי. תאי שמירה הם בעלי פוטוסינתזה גבוהה. הם מכילים מספר גדול שלחילוף עמילן ומיטוכונדריה רבים.
המספר והתפוצה של סטומטות, סוגי מנגנון הסטומטלי משתנים מאוד בצמחים שונים. סטמאטה נעדרת אצל בריופיטים מודרניים. הפוטוסינתזה בהם מתבצעת על ידי דור הגמטופיטים, והספורופיטים אינם מסוגלים להתקיים עצמאי.
בדרך כלל סטמטות ממוקמות בחלק התחתון של העלה. בצמחים הצפים על פני המים - במשטח העליון. בעלים של דגנים, העגבניות מפוזרות לרוב באופן שווה משני הצדדים. עלים כאלה מוארים באופן שווה יחסית. על 1 מ"מ 2 של פני השטח, מ 100 עד 700 stomata ניתן לאתר.
רקמה משנית (periderm). רקמה זו מחליפה את האפידרמיס כאשר צבע ירוקיורה שנתיים מוחלפים בחום. הוא רב-שכבתי ומורכב משכבה מרכזית של תאים קמביאליים - פלוגן. תאי פלוגן, מתחלקים, מניחים שכבת פלם כלפי חוץ, ופלודרם פנימה (איור 5).
פלמה, או פקק. בתחילה הוא מורכב מתאי חיים בעלי דופן דקה. עם הזמן, הקירות שלהם ספוגים בשעוות סוברין ושעוות ירקות ומתים. תוכן התא מתמלא באוויר.
פונקציות פלמה:
מונע אובדן לחות
מגן על הצמח מפני נזק מכני;
מגן מפני פתוגנים
מספק בידוד תרמי, שכן התאים מתמלאים באוויר.
תאים של הפלוגן, הממוקמים באפידרמיס עצמו, השכבה התת אפידרמיסית הבסיסית, לעתים רחוקות יותר בשכבות העמוקות של הקורטקס הראשוני, הם הבסיס היוצר של הקורטקס הראשוני.
שכבת הפקק אינה קבועה. מתרחשות בו הפסקות, המתקשרות עם החללים הבין-תאיים הנמצאים בקרבת מקום. במקביל, נוצרות על פני השטח פקעות קטנות - עדשים, המתקשרות את חללי החללים הבין-תאיים עם אוויר אטמוספרי (איור 6.7).
בסתיו, הפלוגן מתחת לעדשים מניח שכבה של תאים פקוקים, אשר מפחיתים מאוד את הטרנספירציה, אך אינם שוללים אותה לחלוטין. באביב, שכבה זו קורסת מבפנים. על קליפת ליבנה בהירה, עדשים נראות בבירור בצורה של קווים כהים.
גם הרקמה השלישונית (הקרום) אופיינית רק לצורות עץ של צמחים.
הפלוגן מונח שוב ושוב בשכבות העמוקות של הקורטקס. הרקמות שנמצאות מחוצה לו בסופו של דבר מתות ויוצרות קרום. התאים שלו מתים ואינם מסוגלים להימתח. עם זאת, תאים חיים הממוקמים עמוק יותר מתחלקים, מה שמוביל לעלייה בגודל הרוחבי של הגבעול. עם הזמן, השכבה החיצונית של הקרום נשברת. זמן התרחשות של פער כזה הוא ערך קבוע למדי עבור צמחים בודדים. בעץ תפוח זה קורה בשנה השביעית לחיים, בקרן - בחמישים. במינים מסוימים הוא אינו מופיע כלל. תפקידו העיקרי של הפילינג הוא הגנה מפני נזקים מכניים ותרמיים.
פרנכימה
זוהי קבוצה של רקמות מיוחדות הממלאות את החללים בתוך גוף הצמח בין רקמות מוליכות ומכניות (איור 8). לעתים קרובות יותר לתאי פרנכימה יש צורה מעוגלת, לעתים רחוקות יותר צורה מוארכת. נוכחותם של חללים בין-תאיים מפותחים אופיינית. המרווחים בין התאים יוצרים יחד מערכת הובלה - האפופלסט. בנוסף, החללים הבין-תאיים מהווים את "מערכת האוורור" של הצמח. דרך סטמטות, או עדשים, הן מחוברות לאוויר אטמוספרי ומספקות אופטימליות הרכב הגזבתוך הצמח. חללים בין-תאיים מפותחים נחוצים במיוחד עבור צמחים הגדלים על אדמה ספוגת מים, שבה חילופי גזים רגילים קשה. פרנכימה זו נקראת aerenchyma (איור 9).
אלמנטים של הפרנכימה, הממלאים את הפערים בין רקמות אחרות, מבצעים גם את הפונקציה של תמיכה. תאי פרנכימה חיים, אין להם דפנות תאים עבות, כמו סקלרנכימה. לכן, תכונות מכניות מסופקות על ידי טורגור. אם תכולת המים יורדת, מה שמוביל לפסמוליזה ונבילה של הצמח.
פרנכימה ההטמעה נוצרת על ידי תאים בעלי דופן דקה עם חללים בין-תאיים רבים. התאים של מבנה זה מכילים כלורופלסטים רבים, ולכן זה נקרא כלורנכימה. כלורופלסטים ממוקמים לאורך הקיר ללא הצללה זה על זה. בפרנכימה ההטמעה מתרחשות תגובות פוטוסינתזה, המספקות לצמח חומרים אורגניים ואנרגיה. התוצאה של תהליכי פוטוסינתזה היא האפשרות לקיומם של כל היצורים החיים על פני כדור הארץ.
רקמות הטמעה קיימות רק בחלקים המוארים של הצמח; הן מופרדות מהסביבה על ידי אפידרמיס שקוף. אם האפידרמיס מוחלף ברקמות אינגומנטריות משניות אטומות, פרנכימה ההטמעה נעלמת.
פרנכימת האגירה משמשת כלי קיבול לחומרים אורגניים שאינם בשימוש זמנית על ידי האורגניזם הצמחי. בעצם לדחות חומר אורגניכפי ש סוגים שוניםתכלילים מסוגלים לכל תא עם פרוטופלסט חי, אך חלק מהתאים מתמחים בכך (איור 10, 11). תרכובות עשירות באנרגיה מופקדות רק בעונת הגידול, נצרך בתקופה הרדומה ולקראת עונת הגידול הבאה. לכן, חומרי רזרבה מופקדים באיברים הצומחים רק בצמחים רב-שנתיים.
כלי הקיבול של המניות יכול להיות איברים רגילים (בריחה, שורש), כמו גם מתמחים (קני שורש, פקעות, פקעות). כל צמחי הזרע אוגרים חומרים בעלי ערך אנרגטי בזרעים (קוטילונים, אנדוספרם). צמחים רבים של אקלים צחיח אוגרים לא רק חומרים אורגניים, אלא גם מים (איור 12). לדוגמה, אלוורה אוגרת מים בעלים בשרניים, קקטוסים בנצרים.
בדים מכניים
התכונות המכניות של תאי צמחים מסופקות על ידי:
הקליפה הקשה של התא
עכירות, כלומר, מצב הטורגור של תאים.
למרות העובדה שכמעט לכל תאי הרקמה יש תכונות מכניות, עם זאת, יש רקמות בצמח שתכונות מכניות הן בסיסיות עבורן. אלו הן קולנכימה וסקלנכימה. בדרך כלל הם מתפקדים באינטראקציה עם רקמות אחרות. בתוך הגוף יוצרים הצמחים מעין מסגרת. לכן, הם נקראים חיזוק.
לא כל הצמחים באים לידי ביטוי באותה מידה בדים מכניים. במידה הרבה פחות, צמחים החיים ב סביבה מימיתמאשר ארציים. הסיבה היא זאת צמחי מיםצריך פחות תמיכה פנימית. גופם נתמך במידה רבה על ידי המים שמסביב. אוויר ביבשה אינו יוצר תמיכה כזו, שכן יש לו צפיפות נמוכה יותר מאשר מים. מסיבה זו נוכחותם של רקמות מכניות מיוחדות הופכת לרלוונטית.
שיפור של מבני תמיכה פנימיים התרחש בתהליך האבולוציה.
קולנכימה. הוא נוצר רק על ידי תאים חיים, מוארכים לאורך ציר האיבר. סוג זה של רקמה מכנית נוצר מוקדם מאוד, במהלך תקופת הצמיחה הראשונית. לכן, חשוב שהתאים יישארו בחיים וישמרו על יכולת המתיחה יחד עם תאי המתיחה שנמצאים בקרבת מקום.
תכונות של תאי קולנכימה:
עיבוי לא אחיד של הקליפה, וכתוצאה מכך חלקים מסוימים ממנה נשארים דקים, בעוד שאחרים מתעבים;
קונכיות אינן מתגרות.
תאי קולנכימה ממוקמים באופן שונה זה מזה. בתאים סמוכים נוצרות עיבויים בפינות הפונות זו לזו (איור 13). קולנכימה כזו נקראת קולנכימה פינתית. במקרה אחר, התאים מסודרים בשכבות מקבילות. קרומי התא הפונים לשכבות אלו מעובות מאוד. זוהי קולנכימה למלרית. תאים יכולים להיות ממוקמים בצורה רופפת, עם מרווחים בין-תאיים בשפע - זוהי קולנכימה רופפת. קולנכימה כזו נמצאת לעתים קרובות בצמחים על קרקעות ספוגות מים.
לקולנכימה חשיבות מיוחדת בצמחים צעירים, בצורות עשבוניות, וגם בחלקי צמחים שבהם לא מתרחשת צמיחה משנית, כגון עלים. במקרה זה, הוא מונח קרוב מאוד לפני השטח, לפעמים ממש מתחת לאפידרמיס. אם לאיבר יש פרצופים, אז שכבות עבות של קולנכימה נמצאות לאורך הפסגות שלהן.
תאי קולנכימה מתפקדים רק בנוכחות טורגור. מחסור במים מפחית את יעילות הקולנכימה והצמח קמל באופן זמני, למשל, עלי מלפפון נפולים ביום חם. לאחר מילוי התאים במים, הפונקציות של הקולנכימה משוחזרות.
סקלרנכימה. הסוג השני של רקמות מכניות. בניגוד לקולנכימה, שבה כל התאים חיים, תאי סקלנכימה מתים. הקירות שלהם עבים מאוד. הם מבצעים פונקציה מכנית. עיבוי חזק של הממברנה מוביל לשיבוש הובלת חומרים, וכתוצאה מכך הפרוטופלסט מת (איור 14). הליגניפיקציה של ממברנות התא של סקלרנכימה מתרחשת כאשר איבר הצמח כבר השלים את צמיחתו. לכן, הם כבר לא מפריעים למתיחה של הרקמות הסובבות.
בהתאם לצורה, מבחינים בין שני סוגים של תאי סקלרנכימה - סיבים וסקלרים.
לסיבים צורה מוארכת מאוד עם קירות עבים מאוד וחלל קטן. הם קצת יותר קטנים מסיבי עץ. לעתים קרובות מתחת לאפידרמיס יוצרים שכבות אורך וגדילים. בפלואם או בקסלם ניתן למצוא אותם בנפרד או בקבוצות. בפלומה הם נקראים סיבי באסט (איור 15), ובקסילם הם נקראים סיבי ליבריפורם (איור 16).
Sclereids, או תאים אבנים, מיוצגים על ידי תאים מעוגלים או מסועפים עם ממברנות חזקות. בגופו של צמח ניתן למצוא אותם בנפרד (תאים תומכים) (איור 17) או בקבוצות (איור 18). יש לציין כי המאפיינים המכניים תלויים מאוד במיקום הסקלרידים. חלק מהסקלרים יוצרים שכבות רצופות, כמו למשל בקליפת האגוזים או באבני הפירות (פירות האבן).
רקמות הפרשה
כל תא חיבתחילה יש את הפונקציה של סילוק חומרים, בעוד שההובלה יכולה לעבור הן לאורך שיפוע הריכוז והן כנגד השיפוע. ניתן להסיר חומרים שעברו סינתזה בתא ובהמשך ישפיעו על פעילותם של תאים אחרים (פיטו-הורמונים, אנזימים). תהליך זה נקרא הפרשה. במקרה שבו מופרשים חומרים שהם תוצרי פסולת של התא, התהליך נקרא הפרשה. למרות העובדה שלהפרשת חומרים בתאי צמחים ובעלי חיים יש דמיון מהותי, לצמחים יש מספר תכונות הנובעות מהבדלים מהותיים בפעילות החיונית. רמת התגובות המטבוליות בצמחים נמוכה בהרבה מאשר בבעלי חיים. לכן, כמויות קטנות יותר של פסולת נפלטים באופן יחסי. תכונה נוספת היא כי, באופן עצמאי סינתזה כמעט כל הדרוש תרכובות אורגניות, צמחים לעולם אינם יוצרים עתודות מוגזמות. החומרים המשתחררים יכולים לשמש כחומר מוצא לתגובות אחרות (לדוגמה, CO 2 ו- H 2 O).
אם אצל בעלי חיים תהליך ההפרשה של רעלים קשור להסרתם מהגוף, ייתכן שהצמחים לא יעשו זאת, ויבודדו חומרים מיותרים בפרוטופלסט החי (הסרה של חומרים שונים לתוך הוואקואול), בתאים מתים (רוב הרקמות). צמח רב שנתימורכב מתאים כאלה), בחללים בין-תאיים.
ניתן לחלק את החומרים המשתחררים לשניים קבוצות גדולות. הקבוצה הראשונה היא חומרים אורגניים המסונתזים ישירות על ידי התא (אנזימים, פוליסכרידים, ליגנינים, טרפנים, האחרונים הם יסודות מרכיבים שמנים חיונייםושרף). הקבוצה השנייה - חומרים הנכנסים לתא בתחילה מבחוץ בעזרת קסלם או הובלת פלואם (מים, מלח מינרלי, חומצות אמינו, חד סוכרים וכו'). הסוד המופרש לעיתים רחוקות הוא הומוגני ובדרך כלל מורכב מתערובת שבה חומר אחד הוא בעל הריכוז הגבוה ביותר.
לצמחים, בניגוד לבעלי חיים, אין שלם מערכת ההפרשה. ישנם רק מבנים מיוחדים הפזורים ברחבי הצמח - אידיובלסטים. התאים היוצרים אותם קטנים יותר מתאי הפרנכימה השוכנים בקרבת מקום. יש להם ציטופלזמה צפופה באלקטרונים, עם אלמנטים מפותחים של הרשת האנדופלזמית וקומפלקס גולגי. לרוב לא בא לידי ביטוי ואקואול מרכזי. בינם לבין תאים חיים אחרים, תאים אלה מחוברים על ידי פלסמודסמטה רבים.
בהתאם למיקום, רקמות ההפרשה יכולות להיות חיצוניות ופנימיות.
רקמות הפרשה חיצוניות. שערות בלוטות הן נגזרות של האפידרמיס. מבחינה מורפולוגית הם משתנים - הם יכולים להיות בעלי ראש רב תאי, להיות מוארכים, בצורת מגן על רגל וכו'. (איור 19). אלה כוללים שערות סרפד צורבות.
הידתודות הן מבנים המשחררים עודפי מים בתנאים של טרנספירציה נמוכה ולחות גבוהה - גטציה. מבנים אלה נוצרים על ידי קבוצות של תאים חיים חסרי צבע עם קירות דקים - התמה. רקמה זו צמודה לצרור המוליך (איור 20). מים משתחררים דרך סטמטות מים מיוחדות, הנבדלות מהסטומטות רגילות על ידי חוסר תנועה ופער פתוח כל הזמן. הרכב נוזל הביוב משתנה מאוד מכמעט מים נקייםלתערובת מורכבת מאוד של חומרים.
נקטריות. צמחים רבים מתאפיינים בשחרור נוזל המכיל בין 7 ל-87% חד- ואוליגוסכרידים. תהליך זה מתבצע על ידי מבנים מיוחדים - נקטריות. בהתאם למיקום, פרח או פרחוני, נבדלים נקטריונים (איור 21), וכן צמחים הממוקמים על גבעולים ועלים - נקטרי חוץ פרחוני, או חוץ פרחוני. צוף יכול להיות מיץ פלואם ללא שינוי, אשר מועבר אל פני השטח דרך החללים הבין-תאיים ומופרש דרך הסטומטה. נקטרים מורכבים יותר נוצרים על ידי פרנכימה בלוטית, מכוסה באפידרמיס עם שערות בלוטות. הצוף מופרש על ידי תאי אפידרמיס או שערות בלוטות. במקרה זה, הצוף המופרש שונה ממוהל הפלואם. מכיוון שהוא נשלט על ידי גלוקוז, פרוקטוז, סוכרוז, ובמיץ פלואם - גלוקוז. יונים נמצאים בריכוז נמוך. יונים נמצאים בריכוז נמוך. כדי למשוך מאביקים, צוף עשוי להכיל הורמוני סטרואידיםשחיוניים לחרקים.
בלוטות עיכול קיימות בצמחים אוכלי חרקים (טל שמש, nepenthys וכו'). בתגובה למגע של חסרי חוליות קטנים, הם מפרישים סוד המכיל אנזימים הידרוליטיים, חומצות פורמיות, בוטיריות ופרופיוניות. כתוצאה מכך, ביופולימרים מתפרקים למונומרים, המשמשים את המפעל לצרכיו.
בלוטות מלח מתפתחות בצמחים הגדלים על קרקעות מלוחות. בלוטות אלה ממוקמות על העלים ובצורה מומסת, יונים עודפים מועלים אל פני השטח. מלח מופקד תחילה על הציפורן, ואז הוא נשטף בגשם (איור 22). בנוסף, לצמחים רבים יש שערות מלח על העלים. כל שערה מורכבת משני תאים: האחד יוצר את הראש, השני - הגבעול. מלחים מצטברים ב-vacuole של תא הראש, וכאשר ריכוזם הופך גבוה, הראש נופל. במקומו צומח תא חדש שצובר גם מלחים. במהלך צמיחת העלה, הראש עלול ליפול ולהיווצר מספר פעמים. היתרון של שערות מלח הוא שעל ידי שחרור מלחים הן מאבדות מעט מים. תופעה זו חשובה מאוד לצמחים הגדלים על קרקעות מלוחות, בהן מים מתוקים מועטים.
רקמות הפרשה פנימית. מבני הפרשה פנימיים, המפוזרים בכל גוף הצמח בצורה של אידיובלסטים, אינם מסירים חומרים מחוץ לגוף, צוברים אותם בעצמם. אידיובלסטים הם תאים צמחיים השונים בצורתם, במבנה או בתוכן מתאי אחרים באותה רקמה. בהתאם למקור, מבחינים בכלי קיבול סכיזוגניים (מיוונית schiso - פיצול) וליזוגניים (ליזיס יווני - פירוק) של חומרים מופרשים (איור 23).
כלי קיבול סכיזוגניים. אלו חללים בין-תאיים נרחבים יותר או פחות מלאים בחומרים המופרשים. לדוגמה, מעברי שרף של עצי מחט, ארליה, מטרייה ואחרים. מאמינים שיש לשרף תכונות קוטל חיידקים, דוחה אוכלי עשב, מה שהופך צמחים לבלתי אכילים עבורם.
כלי קיבול ליסיגניים נוצרים במקום תאים חיים, שמתים ונהרסים לאחר הצטברות חומרים בהם. ניתן לראות תצורות כאלה בקליפת פירות הדר.
בנוסף לסוגים העיקריים, יש צורות ביניים. חולבים הם משני סוגים: מפרקי ולא מפרקי. הראשונים נוצרים מתאי חיים רבים המסודרים בשרשראות. בנקודת המגע מושמדים הקליפות, הפרוטופלסטים מתמזגים וכתוצאה מכך נוצרת רשת אחת. חלביים כאלה נמצאים ב-Compositae. תאים חלביים לא מפולחים נוצרים על ידי תא ענק מרובה גרעינים. הוא מתעורר בשלבי ההתפתחות המוקדמים, מתארך ומסתעף ככל שהצמח גדל ויוצר רשת מורכבת, למשל בצמחי אופורביה (איור 25).
קירות התא של lactifers מושקים בכבדות. הם אינם מתכלים ולכן הם פלסטיים. החלק המרכזי של הלקטיס תפוס על ידי ואקואול עם לטקס, והפרוטופלסט תופס מיקום קרוב לקיר. לא תמיד ניתן לקבוע את הגבול בין הפרוטופלסט ל-vacuole. הלקטיפרים חודרים למריסטם וגדלים יחד עם איבר הצמח.
רקמות מוליכות
סוג זה מתייחס לרקמות מורכבות, מורכב מתאים מובחנים בצורה שונה. בנוסף לאלמנטים המוליכים בפועל, הרקמה מכילה אלמנטים מכניים, הפרשה ואגירה (איור 26). רקמות מוליכות מאחדות את כל איברי הצמח למערכת אחת. ישנם שני סוגים של רקמות מוליכות: קסלם ופלואם (קסילון יווני - עץ; phloios - קליפה, באסט). יש להם הבדלים מבניים ותפקודיים כאחד.
האלמנטים המוליכים של הקסלם נוצרים על ידי תאים מתים. הם נושאים הובלה למרחקים של מים וחומרים מומסים בהם מהשורש לעלים. האלמנטים המוליכים של הפלואם שומרים על הפרוטופלסט בחיים. הם נושאים הובלה למרחקים ארוכים מעלים פוטוסינתטיים לשורש.
בדרך כלל קסלם ופלואם ממוקמים בגוף הצמח ב סדר מסוים, יוצרות שכבות או צרורות מוליכות. בהתאם למבנה, נבדלים מספר סוגים של צרורות מוליכים, האופייניים להם קבוצות מסוימותצמחים. בצרור הפתוח הביטחוני בין הקסילם והפלואם, יש קמביום, המספק צמיחה משנית (איור 27-A, 28). בצרור הפתוח הדו-צדדי, הפלואם ממוקם משני צידי הקסילם (איור 27-B, 29). צרורות סגורים אינם מכילים קמביום, ולכן אינם מסוגלים להתעבות משני (איור 27-B, 27-D, 30.31). ניתן למצוא שני סוגים נוספים של צרורות קונצנטריים, כאשר הפלואם מקיף את הקסילם (איור 27-E, 32), או שהקסילם מקיף את הפלואם (איור 27-F).
Xylem (עץ). התפתחות הקסילם בצמחים גבוהים יותר קשורה באספקת חילופי מים. מכיוון שהמים מופרשים כל הזמן דרך האפידרמיס, אותה כמות לחות חייבת להיספג על ידי הצמח ולהוסיף לאיברים המבצעים טרנספירציה. יש לקחת בחשבון שנוכחות של פרוטופלסט חי בתאים מוליכים מים תאט מאוד את ההובלה, ותאים מתים מתפקדים כאן יותר. עם זאת, לתא מת אין עמידות, ולכן הממברנה חייבת להיות בעלת תכונות מכניות. הערה: התפרצות היא מצבם של תאי צמחים, רקמות ואיברים, שבו הם הופכים אלסטיים עקב הלחץ של תכולת התאים על הממברנות האלסטיות שלהם. ואכן, האלמנטים המוליכים של הקסלם מורכבים מתאי מת הנמתחים לאורך ציר האיבר עם קונכיות עבות.
בתחילה, הקסילם נוצר מהמריסטם הראשוני - פרוקמביום, הממוקם בחלק העליון של האיברים הציריים. הפרוטוקסילם מבדיל ראשון, ואחריו המטקסילם. ידועים שלושה סוגים של היווצרות קסלם. בסוג exarch, אלמנטים של הפרוטוקסילם מופיעים תחילה על הפריפריה של הצרור הפרוקמביאלי, ואז אלמנטים של המטקסילם מופיעים במרכז. אם התהליך הולך בכיוון ההפוך (כלומר מהמרכז לפריפריה), אז זה טיפוס אנדרכי. בסוג המסרכי, הקסילם מונח במרכז הצרור הפרוקמביאלי, ולאחר מכן הוא מופקד הן לכיוון המרכז והן לכיוון הפריפריה.
השורש מאופיין בסוג אקסרק של היווצרות קסלם, ואילו לגבעולים יש סוג אנדרך. בצמחים מאורגנים נמוכים, שיטות היווצרות הקסיל מגוונות מאוד ויכולות לשמש כמאפיינים שיטתיים.
בצמחים מסוימים (לדוגמה, חד-צוגיים), כל התאים הפרוקמביאליים מתמיינים לרקמות מוליכות שאינן מסוגלות להתעבות משני. בצורות אחרות (למשל, וודי), מריסטמים לרוחב (קמביום) נשארים בין הקסילם והפלואם. תאים אלו מסוגלים להתחלק, לחדש את הקסילם והפלואם. תהליך זה נקרא צמיחה משנית. צמחים רבים הגדלים בתנאי אקלים יציבים יחסית גדלים ללא הרף. בצורות המותאמות לשינויי אקלים עונתיים - מעת לעת. כתוצאה מכך נוצרות טבעות צמיחה שנתיות מוגדרות היטב.
השלבים העיקריים של התמיינות של תאים פרוקמביאליים. לתאיו יש ממברנות דקות שאינן מונעות מהם להימתח במהלך צמיחת האיבר. לאחר מכן, הפרוטופלסט מתחיל להניח את הממברנה המשנית. אבל לתהליך הזה יש תכונות מובהקות. הממברנה המשנית אינה מופקדת בשכבה רציפה, שלא תאפשר לתא להימתח, אלא בצורת טבעות או בספירלה. התארכות התא אינה קשה. בתאים צעירים, הטבעות או הסלילים של הסליל ממוקמים קרוב זה לזה. בתאים בוגרים הם מתפצלים כתוצאה ממתיחה של התאים (איור 33). עיבויים טבעתיים וספירליים של הקליפה אינם מונעים צמיחה, אולם מבחינה מכנית הם נחותים מקונכיות, כאשר העיבוי המשני יוצר שכבה רציפה. לכן, לאחר הפסקת הגדילה בקסילם נוצרים אלמנטים בעלי מעטפת מתמשכת (metaxylem). יש לציין שהעיבוי המשני כאן אינו טבעתי או ספיראלי, אלא מנוקד, דמוי מדרגות, מרושת (איור 34). התאים שלו אינם מסוגלים להימתח, ומתים תוך מספר שעות. תהליך זה בתאים סמוכים מתרחש באופן מתואם. מספר רב של ליזוזומים מופיעים בציטופלזמה. ואז הם מתפרקים, והאנזימים שבהם הורסים את הפרוטופלסט. כאשר הקירות הרוחביים נהרסים, התאים הממוקמים בשרשרת זה מעל זה יוצרים כלי חלול (איור 35). רוֹב אנגיוספריםולכמה שרכים יש כלים.
תא מוליך שאינו נוצר דרך נקבים בדופן נקרא טרכייד. תנועת המים דרך הטראכאידים איטית יותר מאשר דרך הכלים. העובדה היא שבטרכאידים הממברנה הראשונית אינה נקטעת בשום מקום. הטרכאידים מתקשרים זה עם זה דרך נקבוביות. יובהר כי בצמחים הנקבובית היא רק שקע בממברנה המשנית עד לקרום הראשוני, ואין נקבים דרך בין הטרכאידים.
הנפוצות ביותר הן נקבוביות עם גבולות (איור 35-1). הערוץ שלהם, הפונה לחלל התא, יוצר שלוחה - תא הנקבוביות. לנקבוביות של רוב הצמחים המחטניים על הקליפה הראשונית יש עיבוי - טורוס, שהוא סוג של שסתום ומסוגל לווסת את עוצמת הובלת המים. על ידי תזוזה, הטורוס חוסם את זרימת המים דרך הנקבובית, אך לאחר מכן הוא אינו יכול לחזור למקומו הקודם, מבצע פעולה חד פעמית.
הנקבוביות פחות או יותר מעוגלות, מוארכות בניצב לציר המוארך (קבוצה של נקבוביות אלו מזכירה סולם, ולכן נקבוביות זו נקראת נקבוביות סולם). הובלה דרך הנקבוביות מתבצעת הן בכיוון האורך והן בכיוון הרוחבי. נקבוביות קיימות לא רק בטרכאידים, אלא גם בתאי כלי דם בודדים היוצרים כלי.
מנקודת המבט של תורת האבולוציה, טרכאידים הם המבנה הראשון והעיקרי שמוליך מים בגוף של צמחים גבוהים יותר. מאמינים שהכלים מקורם בטרכאידים עקב תמוגה של הקירות הרוחביים ביניהם (איור 36). לרוב השרכים והג'מנוספרמים אין. הם מעבירים מים דרך טראכאידים.
במהלך ההתפתחות האבולוציונית, הופיעו כלי דם קבוצות שונותצמחים שוב ושוב, אבל הם רכשו את המשמעות התפקודית החשובה ביותר באנגיוספרמים, שיש להם אותם יחד עם טרכיידים. מאמינים שהחזקת מנגנון תחבורה מתקדם יותר סייעה להם לא רק לשרוד, אלא גם להשיג מגוון משמעותי של צורות.
Xylem היא רקמה מורכבת, בנוסף לאלמנטים מוליכים מים, היא מכילה אחרים. פונקציות מכניות מבוצעות על ידי סיבים ליבריפורמים (lat. liber - bast, forma - טופס). נוכחותם של מבנים מכניים נוספים חשובה מכיוון שלמרות העיבוי, הקירות של האלמנטים נושאי המים עדיין דקים מדי. הם לא מסוגלים לקיים את עצמם מסה גדולהצמח רב שנתי. הסיבים התפתחו מטראכאידים. הם מאופיינים בגדלים קטנים יותר, פגזים lignified (lignified) וחללים צרים. על הקיר ניתן למצוא נקבוביות נטולות שוליים. סיבים אלו אינם יכולים להוליך מים, תפקידם העיקרי הוא תמיכה.
הקסילם מכיל גם תאים חיים. המסה שלהם יכולה להגיע ל-25% מנפח העץ הכולל. מכיוון שלתאים אלה יש צורה מעוגלת, הם נקראים פרנכימה עץ. בגוף הצמח, הפרנכימה ממוקמת בשתי דרכים. במקרה הראשון, התאים מסודרים בצורה של מיתרים אנכיים - זוהי פרנכימה החבל. במקרה אחר, הפרנכימה יוצרת קרניים אופקיות. הן נקראות קרני הליבה, מכיוון שהן מחברות את הליבה והקורטקס. הליבה מבצעת מספר פונקציות, כולל אחסון חומרים.
פלואם (באסט). זוהי רקמה מורכבת, שכן היא נוצרת על ידי תאים הטרוגניים. התאים המוליכים העיקריים נקראים יסודות מסננת (איור 37). האלמנטים המוליכים של הקסלם נוצרים על ידי תאים מתים, בעוד שבפלואם הם שומרים על פרוטופלסט חי, אם כי שונה מאוד, במהלך תקופת התפקוד. הפלואם מבצע את יציאתם של חומרים פלסטיים מהאיברים הפוטוסינתטיים. לכל התאים החיים של הצמחים יש את היכולת להוביל חומרים אורגניים. ומכאן שאם ניתן למצוא את הקסילם רק בצמחים גבוהים יותר, אז ההובלה של חומרים אורגניים בין תאים מתבצעת גם ב צמחים נמוכים יותר.
קסילם ופלואם מתפתחים ממריסטמים אפיקליים. בשלב הראשון נוצר פרוטופלואם בחבל הפרוקמביאלי. כשהרקמות שמסביב גדלות, היא נמתחת, וכאשר הצמיחה מסתיימת, נוצר מטפלואם במקום פרוטופלואם.
בְּ קבוצות שונותבצמחים גבוהים יותר, ניתן למצוא שני סוגים של יסודות מסננת. בשרכים וגימנוספרמים, הוא מיוצג על ידי תאי מסננת. שדות המסננת בתאים מפוזרים לאורך הקירות הצדדיים. הפרוטופלסט שומר על גרעין הרוס מעט.
באנגיוספרמים, יסודות מסננת נקראים צינורות מסננת. הם מתקשרים זה עם זה דרך צלחות מסננת. לתאים בוגרים חסרים גרעינים. עם זאת, ליד צינור המסננת ממוקם תא נלווה, שנוצר יחד עם צינור המסננת כתוצאה מחלוקה מיטוטית של תא האם המשותף (איור 38). לתא הנלווה יש ציטופלזמה צפופה יותר עם מספר רב של מיטוכונדריה פעילות, כמו גם גרעין מתפקד במלואו, כמות גדולה plasmodesmata (פי עשרה יותר מאשר תאים אחרים). תאים נלווים משפיעים על הפעילות התפקודית של תאי צינורות מסננת לא גרעינית.
למבנה של תאי מסננת בוגרים יש כמה מוזרויות. אין ואקוול, ולכן הציטופלזמה נוזלית מאוד. הגרעין עשוי להיעדר (באנגיוספרמים) או להיות במצב מקומט, לא פעיל מבחינה תפקודית. גם ריבוזומים וקומפלקס גולגי נעדרים, אך הרשת האנדופלזמית מפותחת היטב, אשר לא רק חודרת לציטופלזמה, אלא גם עוברת לתאים שכנים דרך הנקבוביות של שדות המסננת. מיטוכונדריה ופלסטידים מפותחים היטב נמצאים בשפע.
בין תאים, הובלת חומרים עוברת דרך חורים הממוקמים על ממברנות התא. פתחים כאלה נקראים נקבוביות, אך בניגוד לנקבוביות הטראכייד, הם עוברים. ההנחה היא שהם פלסמודסמטה מורחבת מאוד, שעל קירותיה מופקד הפוליסכריד הקאלוז. הנקבוביות מסודרות בקבוצות, ויוצרות שדות מסננות. בצורות פרימיטיביות, שדות מסננת מפוזרים באקראי על פני כל פני הקונכייה, באנגיוספרמים מתקדמים יותר הם ממוקמים בקצוות של תאים שכנים הסמוכים זה לזה, ויוצרים צלחת מסננת (איור 39). אם יש עליו שדה מסננת אחד, זה נקרא פשוט, אם יש כמה - מורכבים.
מהירות תנועת התמיסות דרך יסודות המסננת היא עד 150 ס"מ ∕ שעה. זהו פי אלף מקצב הדיפוזיה החופשית. ככל הנראה מתרחשת הובלה פעילה, ומיטוכונדריות רבות של יסודות מסננת ותאי לוויין מספקים את ה-ATP הדרוש לכך.
החיים של יסודות המסננת של הפלואם תלויים בנוכחות של מריסטמים לרוחב. אם הם קיימים, אז אלמנטי המסננת פועלים לאורך כל חיי הצמח.
בנוסף ליסודות מסננת ותאי לוויין, הפלואם מכיל סיבי באסט, סקלרידים ופרנכימה.
שאלות לשליטה וחזרה
סימנים לצמחים גבוהים יותר? איזה דור של צמחים גבוהים יותר נקרא גמטופיט? ספורופיט?
מהן הסיבות התורמות להתפתחות רקמות מובחנות בצמחים יבשתיים.
רשום את סוגי הצמחים.
אילו קבוצות של מריסטמים אתה מכיר? היכן הם ממוקמים בגוף הצמח?
רקמות מרכיבים, תפקידיהן, משמעות.
אִרגוּן סוגים שוניםכיסוי רקמות.
המבנה של מנגנון הסטומטלי. רשום את סוגי מכשירי הסטומטלי המוכרים לך. כיצד מתפקדות הסטומטה?
תכונות של רקמות parenchymal, סוגי parenchyma.
סימנים של פרנכימה וסקלרנכימה.
רשום את סוגי מבני ההפרשה של צמח. מה המשמעות שלהם?
שיטות ליצירת כלי קיבול לחומרים המופרשים.
מה זה חולבים? לאילו סוגים הם מחולקים?
כיצד מאורגנות רקמות מוליכות?
מה זה צרורות מוליכים? הטיפוסים שלהם.
כיווני תנועה של חומרים דרך הקסילם? פלואם? האם הרכב החומרים המובלים שונה בקסלם ובפלואם?
המבנה והתפקודים של xylem, phloem.
ארגון האלמנטים המוליכים של הפלואם בצמחים ממגוון קבוצות שיטתיות.
כיצד מועברים חומרים בפלואם?
חלבונים נוספים הקשורים אליהם, מה שנקרא חלבוני MAP. חלבונים אלו, על ידי ייצוב המיקרוטובולים, מאיצים את תהליך פילמור הטובולין. מבנה ותפקוד של מיקרופילמנטים מיקרופילמנטים הם מבני חלבון חוטים דקים מאוד וארוכים הנמצאים בכל הציטופלזמה. מתחת לממברנת הפלזמה, מיקרופילמנטים יוצרים מקלעת רציפה, ויוצרים ...
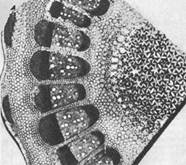

כמו, למשל, שמיר, גזר, לפת וכו', ברוב המקרים יש לו את אותו סוג של רקמה שציינתי זה עתה בפקק. כך, תא הצמח התגלה לראשונה. אבל הוק לא יכול היה להתמודד עם המיקרוסקופ שלו במשך זמן רב: רעיונות של המצאות אחרות רוחשים בראשו (שעוני קפיצים, מצפנים משופרים וכו'), והוא מסר ברצון את תחזוקה נוספת של ...
לפחות איזה מושג לגבי השיטות המתאימות למחקר תאים. מיקרוסקופ אור השיטה העתיקה ביותר ובו בזמן השכיחה ביותר לחקר תאים היא מיקרוסקופיה. אנו יכולים לומר שתחילת המחקר של התא הונחה על ידי המצאת המיקרוסקופ האופטי לאור. לא חמוש עין אנושיתבעל רזולוציה של כ-1/10 מ"מ. זֶה...
עם אברוני תאים רבים, הוא מספק חיבור בין מבנים תוך תאיים בודדים. ליזוזומים ליזוזומים (מ-lis- ומיוונית. סומה - גוף), מבנים בתאים של אורגניזמים של בעלי חיים וצמחים המכילים אנזימים שיכולים לפרק (כלומר lyse - ומכאן השם) חלבונים, פוליסכרידים, פפטידים, חומצות גרעין. זהו מחלקה מאוד מגוונת של בועות בגודל של 0.1-0.4 מיקרון, ...
המבנה של תא צמחי. רקמות צמחיות
מבנה תא הצמח
תא צמחי מורכב מדופן תא נוקשה פחות או יותר ומפרוטופלסט. דופן התא היא דופן התא והממברנה הציטופלזמית. המונח פרוטופלסט מגיע מהמילה פרוטופלסמה, אשר שימשה זה מכבר כדי להתייחס לכל היצורים החיים. הפרוטופלסט הוא הפרוטופלסמה של תא בודד.
הפרוטופלסט מורכב מהציטופלזמה והגרעין. הציטופלזמה מכילה אברונים (ריבוזומים, מיקרוטובולים, פלסטידים, מיטוכונדריה) ומערכות ממברנות (רטיקולום אנדופלזמי, דיקטיוזומים). הציטופלזמה כוללת גם את המטריצה הציטופלזמית (חומר בסיסי) בה טבולים אברונים ומערכות ממברנות. הציטופלזמה מופרדת מדופן התא על ידי קרום הפלזמה, שהוא קרום אלמנטרי. שלא כמו רוב תאי בעלי החיים, תאי צמחים מכילים ואקואול אחד או יותר. אלו שלפוחיות מלאות בנוזל ומוקפות בקרום אלמנטרי (טונופלסט).
בתא צמחי חי, החומר הקרקע נמצא בתנועה מתמדת. אברונים מעורבים בתנועה, הנקראת זרימת הציטופלזמה או ציקלוזה. ציקלוזיס מקל על תנועת החומרים בתא והחלפתם בין התא לסביבה.
קרום פלזמה.זהו מבנה פוספוליפיד דו-שכבתי. תאי צמחים מאופיינים בפלישות של קרום הפלזמה.
קרום הפלזמה מבצע את הפונקציות הבאות:
משתתף בחילופי חומרים בין התא לסביבה;
מתאם את הסינתזה וההרכבה של מיקרו-סיבי תאית של דופן התא;
מעביר אותות הורמונליים וחיצוניים השולטים בצמיחת תאים והתמיינות.
הליבה.זהו המבנה הבולט ביותר בציטופלזמה של תא איקריוטי. הקרנל מבצע שתי פונקציות חשובות:
שולט בפעילות החיונית של התא, וקובע אילו חלבונים, ובאיזו שעה יש לסנתז;
מאחסן מידע גנטי ומעביר אותו לתאי בת במהלך חלוקת התא.
הגרעין של תא איקריוטי מוקף בשתי ממברנות יסוד היוצרות את המעטפת הגרעינית. הוא חדור לנקבוביות רבות בקוטר של 30 עד 100 ננומטר, הנראות רק במיקרוסקופ אלקטרונים. לנקבוביות מבנה מורכב. הממברנה החיצונית של הממברנה הגרעינית במקומות מסוימים משולבת עם הרשת האנדופלזמית. ניתן לראות את המעטפת הגרעינית כחלק מיוחד, מובחן מקומי, של הרשת האנדופלזמית (ER).
בגרעין המוכתם בצבעים מיוחדים, ניתן להבחין בחוטים דקים וגושים של כרומטין ונוקלאופלזמה (החומר העיקרי של הגרעין). הכרומטין מורכב מ-DNA הקשור לחלבונים מיוחדים הנקראים היסטונים. בתהליך חלוקת התא, הכרומטין הופך ליותר ויותר דחוס ומורכב לכרומוזומים. DNA מקודד מידע גנטי.
אורגניזמים שונים במספר הכרומוזומים בתאים סומטיים. לדוגמה, לכרוב יש - 20 כרומוזומים; חמניות - 34; חיטה - 42; בני אדם - 46, ואחד ממיני השרך אופיוגלוסום - 1250. לתאי מין (גמטות) יש רק מחצית ממספר הכרומוזומים האופייני לתאים סומטיים בגוף. מספר הכרומוזומים בגמטות נקרא הפלואיד (יחיד), בתאים סומטיים - דיפלואיד (כפול). תאים שיש להם יותר משתי קבוצות של כרומוזומים נקראים פוליפלואידים.
במיקרוסקופ אור ניתן לראות מבנים כדוריים - נוקלאולים. לכל גרעין יש גרעינים אחד או יותר, הנראים בגרעינים שאינם מתחלקים. RNA ריבוזומלי מסונתז בגרעין. בדרך כלל, לגרעין של אורגניזמים דיפלואידים יש שני גרעינים, אחד לכל קבוצה הפלואידית של כרומוזומים. לנוקלאולים אין קרום משלהם. מבחינה ביוכימית, הנוקלאולי מאופיינים בריכוז גבוה של RNA, הקשור כאן לפוספופרוטאינים. גודל הגרעינים תלוי במצב התפקוד של התא. הבחינו כי בתא הצומח במהירות, שבו מתרחשים תהליכים אינטנסיביים של סינתזת חלבון, נוקלאולי מתרבים בגודלם. בנוקלאולים מיוצרים mRNA וריבוזומים, הממלאים תפקיד סינתטי רק בגרעין.
נוקלאופלזמה (קריופלזמה) מיוצגת על ידי נוזל הומוגני שבו מומסים חלבונים שונים, כולל אנזימים.
פלסטידים.ואקוולים, קירות תאי תאית ופלסידים הם מרכיבים אופייניים לתאי צמחים. לכל פלסטיד יש מעטפת משלו, המורכבת משני ממברנות יסודיות. בתוך הפלסטיד, מבדילים מערכת ממברנות ודרגה משתנה של חומר הומוגני, הסטרומה. פלסטידים בוגרים מסווגים על סמך הפיגמנטים שהם מכילים.
כלורופלסטים, שבהם מתרחשת פוטוסינתזה, מכילים כלורופילים וקרוטנואידים. בדרך כלל יש צורה של דיסק בקוטר של 4 - 5 מיקרון. בתא אחד של המזופיל (אמצע העלה) יכולים להיות 40 - 50 כלורופלסטים; בממ"ר של עלה - כ-500,000. בציטופלזמה, כלורופלסטים ממוקמים בדרך כלל במקביל לממברנת התא.
המבנה הפנימי של הכלורופלסט מורכב. לסטרומה מחלחלת מערכת ממברנות מפותחת בצורת שלפוחיות - thylakoids. כל תילקואיד מורכב משני ממברנות. תילקואידים יוצרים מערכת אחת. ככלל, הם נאספים בערימות - דגנים, הדומים לעמודי מטבעות. התילקואידים של גרנאים בודדים מחוברים ביניהם על ידי תילקואידים סטרומה, או תילקואידים בין-גרניים. כלורופיל וקרוטנואידים משובצים בממברנות התילקואיד. כלורופלסטים בצמחים ירוקים ובאצות מכילים לרוב גרגרי עמילן וטיפות שומנים קטנות (שומן). גרגרי עמילן הם מאגרים זמניים של מוצרי פוטוסינתזה. הם יכולים להיעלם מהכלורופלסטים של צמח שהיה בחושך רק 24 שעות, ולהופיע שוב 3–4 שעות לאחר העברת הצמח לאור.
כלורופלסטים הם אברונים חצי אוטונומיים ודומים לחיידקים. לדוגמה, לריבוזומים של חיידקים וכלורופלסטים יש דמיון גבוה למדי. הם קטנים יותר מריבוזומים אוקריוטיים. סינתזת חלבון על הריבוזומים של חיידקים וכלורופלסטים מעוכבת על ידי כלורמפניקול, שאין לו השפעה על תאים אוקריוטיים. בנוסף, גם לחיידקים וגם לכלורופלסטים יש סוג דומה של נוקלואיד, המאורגן בצורה דומה. למרות העובדה שיצירת כלורופלסטים וסינתזה של פיגמנטים הכלולים בהם נשלטת במידה רבה על ידי ה-DNA הכרומוזומלי של התא, בכל זאת, בהיעדר DNA משלו, כלורופלסטים אינם נוצרים.
כלורופלסטים יכולים להיחשב לאברוני התא העיקריים, שכן הם הראשונים בשרשרת הטרנספורמציה של אנרגיית השמש, וכתוצאה מכך האנושות מקבלת מזון ודלק. בכלורופלסטים מתרחשת לא רק פוטוסינתזה. הם גם מעורבים בסינתזה של חומצות אמינו וחומצות שומן, משמשים כמאגר של עתודות זמניות של עמילן.
כרומופלסטים הם פלסטידים פיגמנטיים. מגוונים בצורתם, אין להם כלורופיל, אלא מסנתזים וצוברים קרוטנואידים, המעניקים צבע צהוב, כתום, אדום לפרחים, עלים ישנים, לפירות ולשורשים. כרומופלסטים יכולים להתפתח מכלורופלסטים, שבמקרה זה מאבדים כלורופיל ומבני קרום פנימיים, צוברים קרוטנואידים. זה מתרחש במהלך הבשלת פירות רבים. כרומופלסטים מושכים חרקים ובעלי חיים אחרים שאיתם הם התפתחו יחד.
לוקופלסטים הם פלסטידים שאינם פיגמנטים. חלקם מסנתזים עמילן (עמילופלסטים), אחרים מסוגלים ליצור חומרים שונים, כולל שומנים וחלבונים. באור, לויקופלסטים הופכים לכלורופלסטים.
פרופלסטידים הם פלסטידים קטנים, חסרי צבע או ירוקים חיוורים, לא מובחנים המצויים בתאים המריסטמטיים (המתחלקים) של שורשים ויורים. הם המבשרים של פלסטידים אחרים, מובחנים יותר - כלורופלסטים, כרומופלסטים ואמינופלסטים. אם התפתחות פרוטופלסטידים מתעכבת בגלל חוסר אור, עלול להופיע בהם גוף פרולמלרי אחד או יותר, שהם הצטברויות חצי גבישיות של ממברנות צינוריות. פלסטידים המכילים גופים פרולמלרים נקראים אטיופלסטים. באור, האטיופלסטים הופכים לכלורופלסטים, בעוד שהקרומים של הגופים הפרולמלרים יוצרים תילקואידים. אטיופלסטים נוצרים בעלים של צמחים הנמצאים בחושך. פרוטופלסטים של עוברי זרעים הופכים תחילה לאטיופלסטים, שמהם מתפתחים כלורופלסטים באור. פלסטידים מאופיינים במעברים קלים יחסית מסוג אחד לאחר. פלסטידים, כמו חיידקים, מתרבים על ידי חלוקה לשניים. בתאים מריסטמטיים, זמן החלוקה של פרוטופלסטידים עולה בקנה אחד עם זמן חלוקת התא. עם זאת, בתאים בוגרים, רוב הפלסטידים נוצרים כתוצאה מחלוקת פלסטידים בוגרים.
מיטוכונדריה.כמו כלורופלסטים, המיטוכונדריה מוקפת בשני ממברנות יסוד. הקרום הפנימי יוצר קפלים ובליטות רבות - cristae, אשר מגדילים משמעותית את פני השטח הפנימיים של המיטוכונדריה. הם קטנים משמעותית מפלסידים, בקוטר של כ-0.5 מיקרומטר, ומשתנים באורך ובצורה.
במיטוכונדריה מתרחש תהליך הנשימה, כתוצאה ממנו מתפרקות מולקולות אורגניות עם שחרור האנרגיה והעברתה למולקולות ATP, מאגר האנרגיה העיקרי של כל התאים האוקריוטיים. רוב תאי הצמח מכילים מאות או אלפי מיטוכונדריות. מספרם בתא אחד נקבע לפי הצורך של התא ב-ATP. המיטוכונדריה נמצאות בתנועה מתמדת, נעות מחלק אחד של התא לאחר, מתמזגות זו עם זו ומתחלקות. מיטוכונדריה בדרך כלל מתאספת במקום שבו יש צורך באנרגיה. אם קרום הפלזמה מעבירה חומרים באופן פעיל מתא לתא, אז המיטוכונדריה ממוקמות לאורך פני הממברנה. באצות חד-תאיות תנועתיות, מיטוכונדריה מצטברות בבסיס הדגל, ומספקות את האנרגיה הדרושה לתנועתן.
מיטוכונדריה, כמו פלסטידים, הם אברונים חצי אוטונומיים המכילים את הרכיבים הדרושים לסינתזה של החלבונים שלהם. הממברנה הפנימית מקיפה מטריצה נוזלית המכילה חלבונים, RNA, DNA, ריבוזומים דמויי חיידקים ומומסים שונים. DNA קיים בצורה של מולקולות מעגליות הממוקמות בנוקלואיד אחד או יותר.
בהתבסס על הדמיון של חיידקים למיטוכונדריה ולכלורופלסטים של תאים אוקריוטיים, ניתן להניח שהמיטוכונדריה והכלורופלסטים מקורם בחיידקים שמצאו "מפלט" בתאים הטרוטרופיים גדולים יותר - מבשרי האוקריוטים.
מיקרוגופים.שלא כמו פלסטידים ומיטוכונדריה, שמתוחמים בשתי ממברנות, גופי מיקרו הם אברונים כדוריים המוקפים בקרום אחד. למיקרוגופים יש תוכן גרגירי (גרגירי), לפעמים הם מכילים גם תכלילי חלבון גבישי. מיקרוגופים קשורים לאזור אחד או שניים של הרשת האנדופלזמית.
חלק מהמיקרוגופים, הנקראים פרוקסיזומים, ממלאים תפקיד חשוב במטבוליזם של חומצה גליקולית, הקשורה ישירות לפוטו-נשימה. בעלים ירוקים הם קשורים למיטוכונדריה ולכלורופלסטים. גופים מיקרוניים אחרים, הנקראים גליאוקסיזומים, מכילים את האנזימים הדרושים להמרת שומנים לפחמימות. זה קורה בהרבה זרעים במהלך הנביטה.
ואקוולים -אלו אזורים בתא התחום על ידי הממברנה, מלאים במוהל תאים נוזלי. הם מוקפים בטונופלסט (ממברנה ואקוולרית).
תא צמחי צעיר מכיל מספר רב של ואקואולים קטנים, המתמזגים לכדי ואקואול אחד גדול ככל שהתא מתבגר. בתא בוגר, עד 90% מנפחו יכול להיות תפוס על ידי וואקואול. במקרה זה, הציטופלזמה נלחצת בצורה של שכבה היקפית דקה לממברנת התא. הגידול בגודל התא נובע בעיקר מצמיחת ה-vacuole. כתוצאה מכך, מתעורר לחץ טורגור וגמישות הרקמה נשמרת. זהו אחד התפקידים העיקריים של הוואקוול והטונופלסט.
המרכיב העיקרי של המיץ הוא מים, השאר משתנים בהתאם לסוג הצמח ומצבו הפיזיולוגי. Vacuoles מכילים מלחים, סוכרים, לעתים רחוקות יותר חלבונים. הטונופלסט ממלא תפקיד פעיל בהובלה והצטברות של יונים מסוימים ב-vacuole. ריכוז היונים במוהל התא יכול לעלות משמעותית על ריכוזו בסביבה. עם תכולה גבוהה של חומרים מסוימים נוצרים גבישים ב-vacuoles. הנפוצים ביותר הם גבישי סידן אוקסלט, בעלי צורה שונה.
Vacuoles הם מקומות הצטברות של מוצרים מטבוליים (מטבוליזם). אלו יכולים להיות חלבונים, חומצות ואפילו חומרים רעילים לבני אדם (אלקלואידים). לעתים קרובות מופקדים פיגמנטים. כחול, סגול, סגול, אדום כהה, ארגמן נותנים לתאי הצמח פיגמנטים מקבוצת האנתוציאנין. בניגוד לפיגמנטים אחרים, הם מתמוססים היטב במים ונמצאים במוהל תאים. הם קובעים את הצבע האדום והכחול של ירקות רבים (צנוניות, לפת, כרוב), פירות (ענבים, שזיפים, דובדבנים), פרחים (פרחים, גרניום, דלפיניום, ורדים, אדמוניות). לפעמים פיגמנטים אלו מסווים את הכלורופיל בעלים, למשל במייפל האדום הדקורטיבי. אנתוציאנינים צבע עלי הסתיו אדום בוהק. הם נוצרים במזג אוויר שמש קר, כאשר סינתזה של כלורופיל נעצרת בעלים. בעלים, כאשר לא נוצרים אנתוציאנינים, לאחר הרס הכלורופיל, הופכים קרוטנואידים צהובים-כתומים של כלורופלסטים בולטים. העלים בעלי צבע עז ביותר בסתיו קר וצלול.
ואקוולים מעורבים בהרס של מקרומולקולות, במחזור המרכיבים שלהן בתא. ריבוזומים, מיטוכונדריה, פלסטידים, נופלים ל-vacuoles, נהרסים. על פי פעילות עיכול זו, ניתן להשוות אותם לליזוזומים - אברונים של תאי בעלי חיים.
ואקוולים נוצרים מהרשת האנדופלזמית (רטיקולום)
ריבוזומים.חלקיקים קטנים (17 - 23 ננומטר), המורכבים מכמויות שוות בערך של חלבון ו-RNA. בריבוזומים, חומצות אמינו מתחברות ליצירת חלבונים. יש יותר מהם בתאים עם חילוף חומרים פעיל. ריבוזומים ממוקמים בציטופלזמה של התא באופן חופשי או מחוברים לרשת האנדופלזמית (80S). הם נמצאים גם בגרעין (80S), במיטוכונדריה (70S), בפלסטידים (70S).
ריבוזומים יכולים ליצור קומפלקס שעליו יש סינתזה סימולטנית של פוליפפטידים זהים, שמידע לגביו נלקח ממולקולה אחת ומ-RNA. קומפלקס כזה נקרא פוליריבוזומים (פוליזומים). לתאים המסנתזים חלבונים בכמויות גדולות יש מערכת נרחבת של פוליזומים, המחוברים לעתים קרובות למשטח החיצוני של המעטפת הגרעינית.
רשת אנדופלזמית.זוהי מערכת ממברנות תלת מימדית מורכבת בהיקף בלתי מוגבל. בהקשר של ER נראה כמו שני ממברנות יסודיות עם רווח שקוף צר ביניהם. צורתו ואורכו של ER תלויים בסוג התא, בפעילותו המטבולית ובשלב ההתמיינות. בתאים המפרישים או מאחסנים חלבונים, ה-ER מעוצב כמו שקים שטוחים או בורות מים, עם ריבוזומים רבים המחוברים אל פני השטח החיצוניים שלו. רשת זו נקראת הרשת האנדופלזמית המחוספסת. ER חלק הוא בדרך כלל בצורת צינורית. רטיקולום אנדופלזמי מחוספס וחלק יכול להיות קיים באותו תא. ככלל, ישנם קשרים רבים ביניהם.
הרשת האנדופלזמית מתפקדת כמערכת התקשורת של התא. זה קשור לקליפה החיצונית של הגרעין. למעשה, שני המבנים הללו יוצרים מערכת ממברנה אחת. כאשר המעטפת הגרעינית נקרעת במהלך חלוקת התא, השברים שלה דומים לשברי ER. הרשת האנדופלזמית היא מערכת להעברת חומרים: חלבונים, שומנים, פחמימות, לחלקים שונים של התא. רשת אנדופלזמה של תאים שכנים מחוברים דרך גדילים ציטופלזמיים - פלסמודסמטה - העוברים דרך ממברנות התא.
הרשת האנדופלזמית היא האתר העיקרי לסינתזה של ממברנות התא. בחלק מתאי הצמח, ממברנות של ואקוולים ומיקרוגופים, נוצרים כאן בורות של דיקטיוזומים.
מערכת גולג'י.מונח זה משמש להתייחס לכל הדיקטיוזומים, או גופי הגולגי, בתא. דיקטיוזומים הם קבוצות של שלפוחיות שטוחות בצורת דיסק, או בורות מים, המסתעפים לאורך הקצוות למערכת מורכבת של צינוריות. דיקטיוזומים בצמחים גבוהים מורכבים מ-4 - 8 בורות, שנאספו יחד.
בדרך כלל, בחפיסת טנקים, מבחינים בצד מתפתח ומתבגר. הממברנות של הבורות היוצרים דומים במבנה לממברנות ה-ER, והממברנות של הבורות המתבגרים דומים לממברנת הפלזמה.
דיקטיוזומים מעורבים בהפרשה, וברוב הצמחים הגבוהים, ביצירת קרומי התא. פוליסכרידים של דופן התא המסונתזים על ידי דיקטיוזומים מצטברים בשלפוחיות, הנפרדות לאחר מכן מבורות מתבגרים. שלפוחיות הפרשה אלו נודדות ומתמזגות עם ה-CMP; בעוד שהפוליסכרידים הכלולים בהם מובנים בתוך קרום התא. חלק מהחומרים המצטברים בדיקטיוזומים נוצרים במבנים אחרים, למשל ב-ER, ואז מועברים לדיקטיוזומים, שם הם משתנים (משונים) לפני ההפרשה. לדוגמה, גליקופרוטאין הם חומר בנייה חשוב של קרום התא. חלק החלבון מסונתז על ידי פוליזומים של ה-ER המחוספס, חלק הפחמימות מסונתז בדיקטיוזומים, כאשר שני החלקים מתאחדים ליצירת גליקופרוטאין.
ממברנות הן מבנים דינמיים וניידים המשנים כל הזמן את צורתם ושטחם. הרעיון של המערכת האנדופלזמית מבוסס על ניידות של ממברנות. על פי תפיסה זו, הממברנות הפנימיות של הציטופלזמה, למעט ממברנות המיטוכונדריה והפלסטידים, הן שלם יחיד ומקורן ברטיקולום האנדופלזמי. בורות מים חדשים של דיקטיוזומים נוצרים מהרשת האנדופלזמית דרך שלב השלפוחית הביניימית, ושלפוחיות ההפרשה הנפרדות מהדיקטיוזומים תורמים בסופו של דבר להיווצרות קרום הפלזמה. לפיכך, הרשת האנדופלזמית והדיקטיוזומים מהווים יחידה תפקודית שבה הדיקטיוזומים ממלאים את התפקיד של מבני ביניים בתהליך המרת ממברנות כמו הרשת האנדופלזמית לממברנות כמו הפלזמה. ברקמות שהתאים שלהן גדלים ומתחלקים בצורה חלשה, מרכיבי הממברנה מתעדכנים כל הזמן.
מיקרוטובוליםנמצא כמעט בכל התאים האיקריוטיים. הם מבנים גליליים בקוטר של כ-24 ננומטר. אורכם משתנה. כל צינורית מורכבת מתת-יחידות של חלבון הנקרא טובולין. יחידות המשנה יוצרות 13 חוטים אורכיים המקיפים את החלל המרכזי. מיקרוטובולים הם מבנים דינמיים, הם נהרסים באופן קבוע ונוצרים בשלבים מסוימים של מחזור התא. ההרכבה שלהם מתרחשת במקומות מיוחדים, הנקראים מרכזי ארגון מיקרוטובולי. בתאי צמחים, יש להם מבנה אמורפי בעל ביטוי חלש.
פונקציות של microtubules: להשתתף ביצירת קרום התא; להפנות בועות של דיקטיוזומים לממברנה היוצרות, כמו חוטי ציר שנוצרים בתא מתחלק; ממלאים תפקיד ביצירת לוחית התא (הגבול הראשוני בין תאי הבת). בנוסף, microtubules הם מרכיב חשוב של flagella ו cilia, בתנועתם הם ממלאים תפקיד חשוב.
מיקרופילמנטים,כמו מיקרוטובולים, הם נמצאים כמעט בכל התאים האיקריוטיים. הם חוטים ארוכים בעובי 5-7 ננומטר, המורכבים מהחלבון המתכווץ אקטין. צרורות של מיקרופילמנטים נמצאים בתאים רבים של צמחים גבוהים יותר. ככל הנראה, הם ממלאים תפקיד חשוב בזרמי הציטופלזמה. מיקרופילמנטים, יחד עם מיקרוטובולים, יוצרים רשת גמישה הנקראת שלד ציטו.
חומר בסיסבמשך זמן רב נחשב לתמיסה הומוגנית (הומוגנית) עשירה בחלבון עם מספר קטן של מבנים או אפילו חסרת מבנה. אולם, נכון להיום, באמצעות מיקרוסקופ אלקטרונים במתח גבוה, נקבע כי החומר הקרקע הוא סריג תלת מימדי הבנוי מחוטים דקים (קוטר 3-6 ננומטר) הממלאים את התא כולו. מרכיבים אחרים של הציטופלזמה, כולל מיקרוטובולים ומיקרופילמנטים, תלויים מהסריג המיקרו-טראבקולרי הזה.
המבנה המיקרוטרבקולרי הוא סריג של גדילי חלבון, שהחלל ביניהם מלא במים. יחד עם מים, לסריג יש עקביות של ג'ל; הג'ל נראה כמו גופים ג'לטינים.
אברונים מחוברים לסריג המיקרוטרבקולרי. הסריג מספק תקשורת בין חלקים בודדים של התא ומכוון את ההובלה התוך תאית.
טיפות שומנים- מבנים בעלי צורה כדורית, המעניקים גרעיניות לציטופלזמה של תא צמחי תחת מיקרוסקופ אור. במיקרוגרפים של אלקטרונים, הם נראים אמורפיים. טיפות דומות מאוד אך קטנות יותר נמצאות בפלסידים.
טיפות ליפידים, בטעות שהן אברונים, קראו להן ספרוזומים והאמינו שהן מוקפות בקרום שכבה בודדת או כפולה. עם זאת, נתונים עדכניים מראים שלטיפות שומנים אין ממברנות, אך עשויות להיות מצופות בחלבון.
חומרים ארגסטיים -אלו הם "המוצרים הפסיביים" של הפרוטופלסט: חומרי רזרבה או פסולת. הם יכולים להופיע ולהיעלם בתקופות שונות של מחזור התא. מלבד גרגרי עמילן, גבישים, פיגמנטים אנתוציאנין וטיפות שומנים. אלה כוללים שרפים, מסטיקים, טאנינים וחלבונים. חומרים ארגאסטיים הם חלק מממברנת התא, החומר העיקרי של הציטופלזמה והאברונים, כולל ואקוולים.
פלאגלה וריסים -הם מבנים דקים דמויי שיער המשתרעים מפני השטח של תאים איקריוטיים רבים. יש להם קוטר קבוע, אבל האורך משתנה בין 2 ל 150 מיקרון. באופן קונבנציונלי, הארוכים והמעטים מהם נקראים דגלים, והקצרים והרבים יותר נקראים ריסים. אין הבדלים ברורים בין שני סוגי המבנים הללו, ולכן המונח flagellum משמש להתייחס לשניהם.
בחלק מהאצות והפטריות, הדגלים הם איברי תנועה, בעזרתם הם נעים במים. בצמחים (לדוגמה, טחבים, עצי כבד, שרכים, כמה ג'מנוספרמים), רק לתאי נבט (גמטות) יש דגלים.
לכל דגל יש ארגון ספציפי. טבעת חיצונית של 9 זוגות של מיקרו-צינוריות מקיפה שתי מיקרו-צינוריות נוספות הממוקמות במרכז הדגל. "ידיות" המכילות אנזים משתרעות ממיקרו-צינורית אחת של כל אחד מהזוגות החיצוניים. זהו דפוס הארגון הבסיסי של 9+2 שנמצא בכל הדגלים באורגניזמים איקריוטיים. מאמינים כי תנועת הפלגלה מבוססת על החלקה של מיקרוטובולים, בעוד שהזוגות החיצוניים של המיקרוטובולים נעים אחד לאורך השני ללא התכווצות. החלקה של זוגות מיקרוטובולים זה לזה גורמת לכיפוף מקומי של הדגל.
דגלים "צומחים" ממבנים גליליים ציטופלזמיים הנקראים גופים בזאליים, המהווים גם את החלק הבסיסי של הדגל. לגופים הבסיסיים יש מבנה פנימי הדומה לזה של דגל, אלא שהצינוריות החיצוניות מסודרות בשלישיות ולא בזוגות, והצינוריות המרכזיות נעדרות.
דופן תא.דופן התא תוחמת את גודל הפרוטופלסט ומונעת את קריעתו עקב ספיגת המים על ידי הוואקואול.
לדופן התא יש תפקידים ספציפיים שחשובים לא רק לתא ולרקמה שבה נמצא התא, אלא גם לצמח כולו. לדפנות התא תפקיד חיוני בספיגה, הובלה והפרשה של חומרים, ובנוסף, ניתן לרכז בהם פעילות ליזוזומלית, או עיכול.
רכיבי דופן התא. המרכיב האופייני ביותר של דופן התא הוא תאית, שקובעת במידה רבה את הארכיטקטורה שלו. מולקולות תאית מורכבות ממולקולות גלוקוז חוזרות המחוברות מקצה לקצה. מולקולות תאית דקות ארוכות משולבות למיקרו-סיביים בעובי של 10 - 25 ננומטר. מיקרופיברילים משתלבים זה בזה ויוצרים חוטים דקים, שבתורם ניתן ללפף אחד סביב השני, כמו חוטים בחבל. לכל "חבל", או מקרופיבריל כזה, עובי של כ-0.5 מיקרון, מגיע לאורך של 4 מיקרון. מקרופיברילים חזקים כמו חוטי פלדה בגודל שווה.
מסגרת התאית של דופן התא מלאה במולקולות מטריצת תאית השזורות בה. הוא מורכב מפוליסכרידים הנקראים hemicelluloses, ו-pectins, או pectins, כימית קרוב מאוד hemicelluloses.
מרכיב נוסף בדופן התא, ליגנין, הוא הפולימר הנפוץ ביותר בתאי הצמח לאחר תאית. לגנין מגביר את קשיחות הדופן ונמצא בדרך כלל בתאים המבצעים פונקציה תומכת או מכנית.
קוטין, סוברין, שעווה - בדרך כלל מופקדים בקונכיות של רקמות המגן של צמחים. קוטין, למשל, נמצא בקרומי התא של האפידרמיס, וסוברין נמצא ברקמת ההגנה המשנית, הפקק. שני החומרים מצויים בשילוב עם שעווה ומונעים איבוד יתר של מים מהצמח.
שכבות של דופן התא. עובי הדופן של תאי הצמח משתנה מאוד, בהתאם לתפקידם של התאים במבנה הצמח וגיל התא עצמו. תחת מיקרוסקופ אלקטרונים, שתי שכבות נראות בדופן התא הצמחי: הלמינה החציונית (המכונה גם החומר הבין-תאי), ודופן התא הראשוני. תאים רבים מניחים שכבה נוספת, דופן התא המשני. הצלחת החציונית ממוקמת בין הקירות הראשוניים של תאים סמוכים. הקיר המשני, אם בכלל, מופקד על ידי פרוטופלסט התא על פני השטח הפנימיים של דופן התא הראשוני.
צלחת אמצעית. הלמינה החציונית מורכבת בעיקר מפקטין. במקום שבו צריך להיווצר דופן תא, בין שני תאים שזה עתה נוצרו, מציינים לראשונה מקלעת צפופה של צינוריות של הרשת האנדופלזמית והבור של מנגנון הגולגי (דיקטיוזומים). ואז מופיעות בועות במקום הזה, מלאות בפקטין (מפוליסכרידים). הבועות הללו מופרדות מהמיכלים של מנגנון גולגי. דופן התא המוקדם מכיל פוליסכרידים שונים, כאשר העיקריים שבהם הם פקטין והמיצלולוז. מאוחר יותר, הוא מורכב מחומרים צפופים יותר - תאית וליגנין.
דופן תא ראשונית. זוהי שכבה של מעטפת תאית המונחת לפני או במהלך צמיחת התא. בנוסף לתאית, המיצלולוזה ופקטין, הממברנות הראשוניות מכילות גליקופרוטאין. קונכיות ראשוניות יכולות להתגרות. רכיב הפקטין מקנה פלסטיות, המאפשרת לקליפה הראשונית להתרחב ככל שהשורש, הגבעול או העלה מתארכים.
לתאים המתחלקים באופן פעיל (רוב התאים הבוגרים המעורבים בתהליכי הפוטוסינתזה, הנשימה וההפרשה) יש ממברנות ראשוניות. תאים כאלה עם ממברנה ראשונית ופרוטופלסט חי מסוגלים לאבד את צורתם האופיינית, להתחלק ולהתמיין לסוג תא חדש. הם מעורבים בריפוי פצעים והתחדשות רקמות בצמחים.
קרומי תאים ראשוניים אינם אחידים בעובי לכל אורכם, אך יש להם חתכים דקים, הנקראים שדות נקבוביות ראשוניים. גדילי הציטופלזמה, או plasmodesmata, המחברים את הפרוטופלסטים של תאים שכנים בדרך כלל עוברים דרך שדות הנקבוביות הראשוניים.
דופן תא משני. למרות העובדה שלתאים צמחיים רבים יש רק דופן ראשונית, בחלקם, הפרוטופלסט מפקיד דופן משנית לכיוון מרכז התא. זה מתרחש בדרך כלל לאחר הפסקת צמיחת התאים ושטח הממברנה הראשונית כבר לא גדל. מסיבה זו, הקליפה המשנית שונה מהראשונית. ממברנות משניות נחוצות במיוחד על ידי תאים מיוחדים המחזקים את הצמח ומוליכים מים. לאחר שקיעת הממברנה המשנית, הפרוטופלסט של תאים אלה, ככלל, מת. יש יותר תאית בקליפות המשניות מאשר בראשוניות, ונעדרים בהן חומרי פקטין וגליקופרוטאין. קשה למתוח את הממברנה המשנית, המטריצה שלו מורכבת מהמיצלולוזה.
במעטפת המשנית ניתן להבחין בשלוש שכבות - חיצוניות, אמצעיות ופנימיות (S1, S2, S3). מבנה השכבות של הקונכיות המשניות מגדיל משמעותית את חוזקן. מיקרו-סיבי תאית בממברנה המשנית מופקדים צפופים יותר מאשר בראשי. לגנין הוא מרכיב נפוץ בעורות המשניים של העץ.
הנקבוביות בממברנות של תאים מגע ממוקמות זו מול זו. שתי נקבוביות מנוגדות וקרום נקבוביות יוצרות זוג נקבוביות. בתאים עם ממברנות משניות, ישנם שני סוגים עיקריים של נקבוביות: פשוטות ותוחמות. בנקבוביות תוחמות, הקיר המשני תלוי מעל חלל הנקבוביות. בנקבוביות פשוטות, זה לא המקרה.
צמיחת דופן התא. ככל שהתא גדל, העובי והשטח של קרום התא גדלים. מתיחת מעטפת היא תהליך מורכב. הוא נשלט על ידי הפרוטופלסט ומווסת על ידי הורמון אוקסין.
בתאים הגדלים באופן אחיד לכל הכיוונים, שקיעת המיופיברילים היא אקראית. מיופיברילים אלה יוצרים רשת לא סדירה. תאים כאלה נמצאו בליבת הגזע, ברקמות האחסון ובמהלך גידול תאים במבחנה. בתאים מתארכים, המיופיברילים של המעטפת הצדדית מופקדים בזווית ישרה לציר ההתארכות.
חומרים מטריקס - פקטין, hemicelluloses וגליקופרוטאינים מועברים לממברנה בשלפוחיות של dictyosomes. יחד עם זאת, פקטין אופייניים יותר לתאים גדלים, בעוד שהמיצלולוזות שולטות בתאים שאינם גדלים.
מיקרו-סיבי תאית מסונתזים על פני התא בעזרת קומפלקס אנזים הקשור לממברנת הפלזמה. הכיוון של מיקרופיברילים נשלט על ידי מיקרוטובולים הממוקמים ליד המשטח הפנימי של קרום הפלזמה.
פלסמודזמה.אלו הם חוטים דקים של הציטופלזמה המחברים את הפרוטופלסטים של תאים שכנים. Plasmodesmata עוברים דרך דופן התא בכל מקום, או מרוכזים בשדות נקבוביות ראשוניים או בממברנות בין זוגות של נקבוביות. תחת מיקרוסקופ אלקטרונים, פלסמודסמטה מופיעות כתעלות צרות המרופדות בקרום הפלזמה. לאורך ציר התעלה משתרע מתא אחד למשנהו צינור גלילי קטן יותר, ה-desmotubule, המתקשר עם הרשת האנדופלזמית של שני התאים הסמוכים. פלסמודסמטה רבות נוצרות במהלך חלוקת התא, כאשר הרטיקולום האנדופלזמי הצינורי נקלט על ידי לוחית התא המתפתחת. Plasmodesmata יכול להיווצר גם בממברנות של תאים שאינם מתחלקים. מבנים אלה מספקים העברה יעילה של חומרים מסוימים מתא לתא.
חלוקת תא.באורגניזמים רב-תאיים, חלוקת תאים, יחד עם עלייה בגודלם, היא דרך לצמיחה של האורגניזם כולו. תאים חדשים שנוצרים במהלך החלוקה דומים במבנה ובתפקוד, הן לתא האב והן זה לזה. ניתן לחלק את תהליך החלוקה באאוקריוטים לשני שלבים חופפים חלקית: מיטוזה וציטוקינזיס.
מיטוזיס היא היווצרות של שני גרעיני בת מגרעין אחד, שוות ערך זה לזה מבחינה מורפולוגית וגנטית. ציטוקינזיס היא חלוקת החלק הציטופלזמי של התא עם היווצרות תאי בת.
מחזור התא.תא חי עובר סדרה של אירועים עוקבים המרכיבים את מחזור התא. משך המחזור עצמו משתנה בהתאם לסוג התא ולגורמים חיצוניים כמו טמפרטורה או זמינות חומרי הזנה. בדרך כלל המחזור מחולק ל-interphase ולארבעה שלבים של מיטוזה.
Interphase.התקופה שבין חלוקות מיטוטיות עוקבות.
האינטרפאזה מחולקת לשלוש תקופות, המסומנות כ-G1, S, G2.
במהלך תקופת G1, שמתחילה לאחר מיטוזה. במהלך תקופה זו, כמות הציטופלזמה עולה, כולל אברונים שונים. בנוסף, על פי ההשערה הנוכחית, חומרים מסונתזים במהלך תקופת G1 המעוררים או מעכבים את תקופת S ואת שאר המחזור, ובכך קובעים את תהליך החלוקה.
תקופת S עוקבת אחר תקופת G1, שבה החומר הגנטי (DNA) משוכפל.
במהלך תקופת G2, העוקבת אחרי S, נוצרים מבנים המעורבים ישירות במיטוזה, כמו רכיבי ציר.
חלק מהתאים עוברים מספר בלתי מוגבל של מחזורי תאים. אלה הם אורגניזמים חד-תאיים וכמה תאים של אזורי צמיחה פעילים (מריסטמים). כמה תאים מיוחדים לאחר התבגרות מאבדים את יכולתם להתרבות. הקבוצה השלישית של תאים, למשל, אלו היוצרים רקמת פצע (קלוס), שומרת על היכולת להתחלק רק בתנאים מיוחדים.
מיטוזה,או ביקוע גרעיני. זהו תהליך מתמשך המחולק לארבעה שלבים: פרופאזה, מטאפאזה, אנפאזה, טלופזה. כתוצאה מהמיטוזה, החומר הגנטי שהכפיל את עצמו ב-interphase מחולק שווה בשווה בין שני גרעיני הבת.
אחד הסימנים המוקדמים ביותר למעבר של תא לחלוקה הוא הופעת פס צר וטבעתי של מיקרו-צינוריות ממש מתחת לממברנת הפלזמה. חגורה צפופה יחסית זו מקיפה את הגרעין במישור המשווני של הציר המיטוטי העתידי. מכיוון שהיא מופיעה לפני הפרופזה, היא נקראת חגורת הקדם-פרופאזה. הוא נעלם לאחר הציר המיטוטי, הרבה לפני הופעתו בטלופזה המאוחר של לוחית התא, הגדל מהמרכז לפריפריה ומתמזג עם קרום תא האם באזור שנכבש בעבר על ידי חגורת הקדם-פרופאזה.
פרופאזה. בתחילת הפרופזה, הכרומוזומים דומים לחוטים ארוכים המפוזרים בתוך הגרעין. לאחר מכן, כשהגדילים מתקצרים ומתעבים, אפשר לראות שכל כרומוזום מורכב לא אחד, אלא משני גדילים שלובים זה בזה, הנקראים כרומטידים. בפרופאזה המאוחרת, שתי הכרומטידות הזוגיות המקוצרות של כל כרומוזום שוכבות זו לצד זו במקביל, מחוברות בקטע צר הנקרא צנטרומר. יש לו מיקום ספציפי על כל כרומוזום והוא מחלק את הכרומוזום לשתי זרועות באורכים שונים.
מיקרו-צינוריות ממוקמות במקביל לפני השטח של הגרעין לאורך ציר הציר. זהו הביטוי המוקדם ביותר של הרכבת ציר מיטוטי.
בסוף הפרופזה, הגרעין מאבד בהדרגה את קווי המתאר הברורים שלו ולבסוף נעלם. זמן קצר לאחר מכן מתפרקת גם המעטפת הגרעינית.
מטאפאזה. בתחילת המטאפאזה, הציר, שהוא מבנה תלת מימדי, הרחב ביותר באמצע ומצטמצם לכיוון הקטבים, תופס את המקום שתפס בעבר הגרעין. סיבי הציר הם צרורות של microtubules. במהלך המטאפאזה, הכרומוזומים, שכל אחד מהם מורכב משתי כרומטידות, מסודרים כך שהצנטרומרים שלהם נמצאים במישור המשווני של הציר. עם הצנטרומר שלו, כל כרומוזום מחובר לחוטי הציר. עם זאת, כמה גדילים עוברים מקוטב אחד למשנהו מבלי להיות מחוברים לכרומוזומים.
כאשר כל הכרומוזומים ממוקמים במישור המשווני, המטאפאזה הושלמה. כרומוזומים מוכנים להתחלק.
אנפאזה. הכרומטידות של כל כרומוזום נפרדות. עכשיו אלה כרומוזומי הבת. קודם כל, הצנטרומר מתחלק, ושני כרומוזומי הבת נמשכים לקטבים מנוגדים. במקרה זה, הצנטרומרים נעים קדימה, וזרועות הכרומוזומים נמתחות מאחור. חוטי הציר המחוברים לכרומוזומים מתקצרים, ותורמים להפרדת הכרומטידים ולתנועת כרומוזומי הבת בכיוונים מנוגדים.
מבנה תא הצמח
תא צמחי מורכב מדופן תא נוקשה פחות או יותר ומפרוטופלסט. דופן התא היא דופן התא והממברנה הציטופלזמית. המונח פרוטופלסט מגיע מהמילה פרוטופלסמה, אשר שימשה זה מכבר כדי להתייחס לכל היצורים החיים. הפרוטופלסט הוא הפרוטופלסמה של תא בודד.
הפרוטופלסט מורכב מהציטופלזמה והגרעין. הציטופלזמה מכילה אברונים (ריבוזומים, מיקרוטובולים, פלסטידים, מיטוכונדריה) ומערכות ממברנות (רטיקולום אנדופלזמי, דיקטיוזומים). הציטופלזמה כוללת גם את המטריצה הציטופלזמית (חומר בסיסי) בה טבולים אברונים ומערכות ממברנות. הציטופלזמה מופרדת מדופן התא על ידי קרום הפלזמה, שהוא קרום אלמנטרי. שלא כמו רוב תאי בעלי החיים, תאי צמחים מכילים ואקואול אחד או יותר. אלו שלפוחיות מלאות בנוזל ומוקפות בקרום אלמנטרי (טונופלסט).
בתא צמחי חי, החומר הקרקע נמצא בתנועה מתמדת. אברונים מעורבים בתנועה, הנקראת זרימת הציטופלזמה או ציקלוזה. ציקלוזיס מקל על תנועת החומרים בתא והחלפתם בין התא לסביבה.
קרום פלזמה.זהו מבנה פוספוליפיד דו-שכבתי. תאי צמחים מאופיינים בפלישות של קרום הפלזמה.
קרום הפלזמה מבצע את הפונקציות הבאות:
משתתף בחילופי חומרים בין התא לסביבה;
מתאם את הסינתזה וההרכבה של מיקרו-סיבי תאית של דופן התא;
מעביר אותות הורמונליים וחיצוניים השולטים בצמיחת תאים והתמיינות.
הליבה.זהו המבנה הבולט ביותר בציטופלזמה של תא איקריוטי. הקרנל מבצע שתי פונקציות חשובות:
שולט בפעילות החיונית של התא, וקובע אילו חלבונים, ובאיזו שעה יש לסנתז;
מאחסן מידע גנטי ומעביר אותו לתאי בת במהלך חלוקת התא.
הגרעין של תא איקריוטי מוקף בשתי ממברנות יסוד היוצרות את המעטפת הגרעינית. הוא חדור לנקבוביות רבות בקוטר של 30 עד 100 ננומטר, הנראות רק במיקרוסקופ אלקטרונים. לנקבוביות מבנה מורכב. הממברנה החיצונית של הממברנה הגרעינית במקומות מסוימים משולבת עם הרשת האנדופלזמית. ניתן לראות את המעטפת הגרעינית כחלק מיוחד, מובחן מקומי, של הרשת האנדופלזמית (ER).
בגרעין המוכתם בצבעים מיוחדים, ניתן להבחין בחוטים דקים וגושים של כרומטין ונוקלאופלזמה (החומר העיקרי של הגרעין). הכרומטין מורכב מ-DNA הקשור לחלבונים מיוחדים הנקראים היסטונים. בתהליך חלוקת התא, הכרומטין הופך ליותר ויותר דחוס ומורכב לכרומוזומים. DNA מקודד מידע גנטי.
אורגניזמים שונים במספר הכרומוזומים בתאים סומטיים. לדוגמה, לכרוב יש - 20 כרומוזומים; חמניות - 34; חיטה - 42; בני אדם - 46, ואחד ממיני השרך אופיוגלוסום - 1250. לתאי מין (גמטות) יש רק מחצית ממספר הכרומוזומים האופייני לתאים סומטיים בגוף. מספר הכרומוזומים בגמטות נקרא הפלואיד (יחיד), בתאים סומטיים - דיפלואיד (כפול). תאים שיש להם יותר משתי קבוצות של כרומוזומים נקראים פוליפלואידים.
במיקרוסקופ אור ניתן לראות מבנים כדוריים - נוקלאולים. לכל גרעין יש גרעינים אחד או יותר, הנראים בגרעינים שאינם מתחלקים. RNA ריבוזומלי מסונתז בגרעין. בדרך כלל, לגרעין של אורגניזמים דיפלואידים יש שני גרעינים, אחד לכל קבוצה הפלואידית של כרומוזומים. לנוקלאולים אין קרום משלהם. מבחינה ביוכימית, הנוקלאולי מאופיינים בריכוז גבוה של RNA, הקשור כאן לפוספופרוטאינים. גודל הגרעינים תלוי במצב התפקוד של התא. הבחינו כי בתא הצומח במהירות, שבו מתרחשים תהליכים אינטנסיביים של סינתזת חלבון, נוקלאולי מתרבים בגודלם. בנוקלאולים מיוצרים mRNA וריבוזומים, הממלאים תפקיד סינתטי רק בגרעין.
נוקלאופלזמה (קריופלזמה) מיוצגת על ידי נוזל הומוגני שבו מומסים חלבונים שונים, כולל אנזימים.
פלסטידים.ואקוולים, קירות תאי תאית ופלסידים הם מרכיבים אופייניים לתאי צמחים. לכל פלסטיד יש מעטפת משלו, המורכבת משני ממברנות יסודיות. בתוך הפלסטיד, מבדילים מערכת ממברנות ודרגה משתנה של חומר הומוגני, הסטרומה. פלסטידים בוגרים מסווגים על סמך הפיגמנטים שהם מכילים.
כלורופלסטים, שבהם מתרחשת פוטוסינתזה, מכילים כלורופילים וקרוטנואידים. בדרך כלל יש צורה של דיסק בקוטר של 4 - 5 מיקרון. בתא אחד של המזופיל (אמצע העלה) יכולים להיות 40 - 50 כלורופלסטים; בממ"ר של עלה - כ-500,000. בציטופלזמה, כלורופלסטים ממוקמים בדרך כלל במקביל לממברנת התא.
המבנה הפנימי של הכלורופלסט מורכב. לסטרומה מחלחלת מערכת ממברנות מפותחת בצורת שלפוחיות - thylakoids. כל תילקואיד מורכב משני ממברנות. תילקואידים יוצרים מערכת אחת. ככלל, הם נאספים בערימות - דגנים, הדומים לעמודי מטבעות. התילקואידים של גרנאים בודדים מחוברים ביניהם על ידי תילקואידים סטרומה, או תילקואידים בין-גרניים. כלורופיל וקרוטנואידים משובצים בממברנות התילקואיד. כלורופלסטים בצמחים ירוקים ובאצות מכילים לרוב גרגרי עמילן וטיפות שומנים קטנות (שומן). גרגרי עמילן הם מאגרים זמניים של מוצרי פוטוסינתזה. הם יכולים להיעלם מהכלורופלסטים של צמח שהיה בחושך רק 24 שעות, ולהופיע שוב 3–4 שעות לאחר העברת הצמח לאור.
כלורופלסטים הם אברונים חצי אוטונומיים ודומים לחיידקים. לדוגמה, לריבוזומים של חיידקים וכלורופלסטים יש דמיון גבוה למדי. הם קטנים יותר מריבוזומים אוקריוטיים. סינתזת חלבון על הריבוזומים של חיידקים וכלורופלסטים מעוכבת על ידי כלורמפניקול, שאין לו השפעה על תאים אוקריוטיים. בנוסף, גם לחיידקים וגם לכלורופלסטים יש סוג דומה של נוקלואיד, המאורגן בצורה דומה. למרות העובדה שיצירת כלורופלסטים וסינתזה של פיגמנטים הכלולים בהם נשלטת במידה רבה על ידי ה-DNA הכרומוזומלי של התא, בכל זאת, בהיעדר DNA משלו, כלורופלסטים אינם נוצרים.
כלורופלסטים יכולים להיחשב לאברוני התא העיקריים, שכן הם הראשונים בשרשרת הטרנספורמציה של אנרגיית השמש, וכתוצאה מכך האנושות מקבלת מזון ודלק. בכלורופלסטים מתרחשת לא רק פוטוסינתזה. הם גם מעורבים בסינתזה של חומצות אמינו וחומצות שומן, משמשים כמאגר של עתודות זמניות של עמילן.
כרומופלסטים הם פלסטידים פיגמנטיים. מגוונים בצורתם, אין להם כלורופיל, אלא מסנתזים וצוברים קרוטנואידים, המעניקים צבע צהוב, כתום, אדום לפרחים, עלים ישנים, לפירות ולשורשים. כרומופלסטים יכולים להתפתח מכלורופלסטים, שבמקרה זה מאבדים כלורופיל ומבני קרום פנימיים, צוברים קרוטנואידים. זה מתרחש במהלך הבשלת פירות רבים. כרומופלסטים מושכים חרקים ובעלי חיים אחרים שאיתם הם התפתחו יחד.
לוקופלסטים הם פלסטידים שאינם פיגמנטים. חלקם מסנתזים עמילן (עמילופלסטים), אחרים מסוגלים ליצור חומרים שונים, כולל שומנים וחלבונים. באור, לויקופלסטים הופכים לכלורופלסטים.
פרופלסטידים הם פלסטידים קטנים, חסרי צבע או ירוקים חיוורים, לא מובחנים המצויים בתאים המריסטמטיים (המתחלקים) של שורשים ויורים. הם המבשרים של פלסטידים אחרים, מובחנים יותר - כלורופלסטים, כרומופלסטים ואמינופלסטים. אם התפתחות פרוטופלסטידים מתעכבת בגלל חוסר אור, עלול להופיע בהם גוף פרולמלרי אחד או יותר, שהם הצטברויות חצי גבישיות של ממברנות צינוריות. פלסטידים המכילים גופים פרולמלרים נקראים אטיופלסטים. באור, האטיופלסטים הופכים לכלורופלסטים, בעוד שהקרומים של הגופים הפרולמלרים יוצרים תילקואידים. אטיופלסטים נוצרים בעלים של צמחים הנמצאים בחושך. פרוטופלסטים של עוברי זרעים הופכים תחילה לאטיופלסטים, שמהם מתפתחים כלורופלסטים באור. פלסטידים מאופיינים במעברים קלים יחסית מסוג אחד לאחר. פלסטידים, כמו חיידקים, מתרבים על ידי חלוקה לשניים. בתאים מריסטמטיים, זמן החלוקה של פרוטופלסטידים עולה בקנה אחד עם זמן חלוקת התא. עם זאת, בתאים בוגרים, רוב הפלסטידים נוצרים כתוצאה מחלוקת פלסטידים בוגרים.
מיטוכונדריה.כמו כלורופלסטים, המיטוכונדריה מוקפת בשני ממברנות יסוד. הקרום הפנימי יוצר קפלים ובליטות רבות - cristae, אשר מגדילים משמעותית את פני השטח הפנימיים של המיטוכונדריה. הם קטנים משמעותית מפלסידים, בקוטר של כ-0.5 מיקרומטר, ומשתנים באורך ובצורה.
במיטוכונדריה מתרחש תהליך הנשימה, כתוצאה ממנו מתפרקות מולקולות אורגניות עם שחרור האנרגיה והעברתה למולקולות ATP, מאגר האנרגיה העיקרי של כל התאים האוקריוטיים. רוב תאי הצמח מכילים מאות או אלפי מיטוכונדריות. מספרם בתא אחד נקבע לפי הצורך של התא ב-ATP. המיטוכונדריה נמצאות בתנועה מתמדת, נעות מחלק אחד של התא לאחר, מתמזגות זו עם זו ומתחלקות. מיטוכונדריה בדרך כלל מתאספת במקום שבו יש צורך באנרגיה. אם קרום הפלזמה מעבירה חומרים באופן פעיל מתא לתא, אז המיטוכונדריה ממוקמות לאורך פני הממברנה. באצות חד-תאיות תנועתיות, מיטוכונדריה מצטברות בבסיס הדגל, ומספקות את האנרגיה הדרושה לתנועתן.
מיטוכונדריה, כמו פלסטידים, הם אברונים חצי אוטונומיים המכילים את הרכיבים הדרושים לסינתזה של החלבונים שלהם. הממברנה הפנימית מקיפה מטריצה נוזלית המכילה חלבונים, RNA, DNA, ריבוזומים דמויי חיידקים ומומסים שונים. DNA קיים בצורה של מולקולות מעגליות הממוקמות בנוקלואיד אחד או יותר.
בהתבסס על הדמיון של חיידקים למיטוכונדריה ולכלורופלסטים של תאים אוקריוטיים, ניתן להניח שהמיטוכונדריה והכלורופלסטים מקורם בחיידקים שמצאו "מפלט" בתאים הטרוטרופיים גדולים יותר - מבשרי האוקריוטים.
מיקרוגופים.שלא כמו פלסטידים ומיטוכונדריה, שמתוחמים בשתי ממברנות, גופי מיקרו הם אברונים כדוריים המוקפים בקרום אחד. למיקרוגופים יש תוכן גרגירי (גרגירי), לפעמים הם מכילים גם תכלילי חלבון גבישי. מיקרוגופים קשורים לאזור אחד או שניים של הרשת האנדופלזמית.
חלק מהמיקרוגופים, הנקראים פרוקסיזומים, ממלאים תפקיד חשוב במטבוליזם של חומצה גליקולית, הקשורה ישירות לפוטו-נשימה. בעלים ירוקים הם קשורים למיטוכונדריה ולכלורופלסטים. גופים מיקרוניים אחרים, הנקראים גליאוקסיזומים, מכילים את האנזימים הדרושים להמרת שומנים לפחמימות. זה קורה בהרבה זרעים במהלך הנביטה.
ואקוולים -אלו אזורים בתא התחום על ידי הממברנה, מלאים במוהל תאים נוזלי. הם מוקפים בטונופלסט (ממברנה ואקוולרית).
תא צמחי צעיר מכיל מספר רב של ואקואולים קטנים, המתמזגים לכדי ואקואול אחד גדול ככל שהתא מתבגר. בתא בוגר, עד 90% מנפחו יכול להיות תפוס על ידי וואקואול. במקרה זה, הציטופלזמה נלחצת בצורה של שכבה היקפית דקה לממברנת התא. הגידול בגודל התא נובע בעיקר מצמיחת ה-vacuole. כתוצאה מכך, מתעורר לחץ טורגור וגמישות הרקמה נשמרת. זהו אחד התפקידים העיקריים של הוואקוול והטונופלסט.
המרכיב העיקרי של המיץ הוא מים, השאר משתנים בהתאם לסוג הצמח ומצבו הפיזיולוגי. Vacuoles מכילים מלחים, סוכרים, לעתים רחוקות יותר חלבונים. הטונופלסט ממלא תפקיד פעיל בהובלה והצטברות של יונים מסוימים ב-vacuole. ריכוז היונים במוהל התא יכול לעלות משמעותית על ריכוזו בסביבה. עם תכולה גבוהה של חומרים מסוימים נוצרים גבישים ב-vacuoles. הנפוצים ביותר הם גבישי סידן אוקסלט, בעלי צורה שונה.
Vacuoles הם מקומות הצטברות של מוצרים מטבוליים (מטבוליזם). אלו יכולים להיות חלבונים, חומצות ואפילו חומרים רעילים לבני אדם (אלקלואידים). לעתים קרובות מופקדים פיגמנטים. כחול, סגול, סגול, אדום כהה, ארגמן נותנים לתאי הצמח פיגמנטים מקבוצת האנתוציאנין. בניגוד לפיגמנטים אחרים, הם מתמוססים היטב במים ונמצאים במוהל תאים. הם קובעים את הצבע האדום והכחול של ירקות רבים (צנוניות, לפת, כרוב), פירות (ענבים, שזיפים, דובדבנים), פרחים (פרחים, גרניום, דלפיניום, ורדים, אדמוניות). לפעמים פיגמנטים אלו מסווים את הכלורופיל בעלים, למשל במייפל האדום הדקורטיבי. אנתוציאנינים צבע עלי הסתיו אדום בוהק. הם נוצרים במזג אוויר שמש קר, כאשר סינתזה של כלורופיל נעצרת בעלים. בעלים, כאשר לא נוצרים אנתוציאנינים, לאחר הרס הכלורופיל, הופכים קרוטנואידים צהובים-כתומים של כלורופלסטים בולטים. העלים בעלי צבע עז ביותר בסתיו קר וצלול.
ואקוולים מעורבים בהרס של מקרומולקולות, במחזור המרכיבים שלהן בתא. ריבוזומים, מיטוכונדריה, פלסטידים, נופלים ל-vacuoles, נהרסים. על פי פעילות עיכול זו, ניתן להשוות אותם לליזוזומים - אברונים של תאי בעלי חיים.
ואקוולים נוצרים מהרשת האנדופלזמית (רטיקולום)
ריבוזומים.חלקיקים קטנים (17 - 23 ננומטר), המורכבים מכמויות שוות בערך של חלבון ו-RNA. בריבוזומים, חומצות אמינו מתחברות ליצירת חלבונים. יש יותר מהם בתאים עם חילוף חומרים פעיל. ריבוזומים ממוקמים בציטופלזמה של התא באופן חופשי או מחוברים לרשת האנדופלזמית (80S). הם נמצאים גם בגרעין (80S), במיטוכונדריה (70S), בפלסטידים (70S).
ריבוזומים יכולים ליצור קומפלקס שעליו יש סינתזה סימולטנית של פוליפפטידים זהים, שמידע לגביו נלקח ממולקולה אחת ומ-RNA. קומפלקס כזה נקרא פוליריבוזומים (פוליזומים). לתאים המסנתזים חלבונים בכמויות גדולות יש מערכת נרחבת של פוליזומים, המחוברים לעתים קרובות למשטח החיצוני של המעטפת הגרעינית.
רשת אנדופלזמית.זוהי מערכת ממברנות תלת מימדית מורכבת בהיקף בלתי מוגבל. בהקשר של ER נראה כמו שני ממברנות יסודיות עם רווח שקוף צר ביניהם. צורתו ואורכו של ER תלויים בסוג התא, בפעילותו המטבולית ובשלב ההתמיינות. בתאים המפרישים או מאחסנים חלבונים, ה-ER מעוצב כמו שקים שטוחים או בורות מים, עם ריבוזומים רבים המחוברים אל פני השטח החיצוניים שלו. רשת זו נקראת הרשת האנדופלזמית המחוספסת. ER חלק הוא בדרך כלל בצורת צינורית. רטיקולום אנדופלזמי מחוספס וחלק יכול להיות קיים באותו תא. ככלל, ישנם קשרים רבים ביניהם.
הרשת האנדופלזמית מתפקדת כמערכת התקשורת של התא. זה קשור לקליפה החיצונית של הגרעין. למעשה, שני המבנים הללו יוצרים מערכת ממברנה אחת. כאשר המעטפת הגרעינית נקרעת במהלך חלוקת התא, השברים שלה דומים לשברי ER. הרשת האנדופלזמית היא מערכת להעברת חומרים: חלבונים, שומנים, פחמימות, לחלקים שונים של התא. רשת אנדופלזמה של תאים שכנים מחוברים דרך גדילים ציטופלזמיים - פלסמודסמטה - העוברים דרך ממברנות התא.
הרשת האנדופלזמית היא האתר העיקרי לסינתזה של ממברנות התא. בחלק מתאי הצמח, ממברנות של ואקוולים ומיקרוגופים, נוצרים כאן בורות של דיקטיוזומים.
מערכת גולג'י.מונח זה משמש להתייחס לכל הדיקטיוזומים, או גופי הגולגי, בתא. דיקטיוזומים הם קבוצות של שלפוחיות שטוחות בצורת דיסק, או בורות מים, המסתעפים לאורך הקצוות למערכת מורכבת של צינוריות. דיקטיוזומים בצמחים גבוהים מורכבים מ-4 - 8 בורות, שנאספו יחד.
בדרך כלל, בחפיסת טנקים, מבחינים בצד מתפתח ומתבגר. הממברנות של הבורות היוצרים דומים במבנה לממברנות ה-ER, והממברנות של הבורות המתבגרים דומים לממברנת הפלזמה.
דיקטיוזומים מעורבים בהפרשה, וברוב הצמחים הגבוהים, ביצירת קרומי התא. פוליסכרידים של דופן התא המסונתזים על ידי דיקטיוזומים מצטברים בשלפוחיות, הנפרדות לאחר מכן מבורות מתבגרים. שלפוחיות הפרשה אלו נודדות ומתמזגות עם ה-CMP; בעוד שהפוליסכרידים הכלולים בהם מובנים בתוך קרום התא. חלק מהחומרים המצטברים בדיקטיוזומים נוצרים במבנים אחרים, למשל ב-ER, ואז מועברים לדיקטיוזומים, שם הם משתנים (משונים) לפני ההפרשה. לדוגמה, גליקופרוטאין הם חומר בנייה חשוב של קרום התא. חלק החלבון מסונתז על ידי פוליזומים של ה-ER המחוספס, חלק הפחמימות מסונתז בדיקטיוזומים, כאשר שני החלקים מתאחדים ליצירת גליקופרוטאין.
ממברנות הן מבנים דינמיים וניידים המשנים כל הזמן את צורתם ושטחם. הרעיון של המערכת האנדופלזמית מבוסס על ניידות של ממברנות. על פי תפיסה זו, הממברנות הפנימיות של הציטופלזמה, למעט ממברנות המיטוכונדריה והפלסטידים, הן שלם יחיד ומקורן ברטיקולום האנדופלזמי. בורות מים חדשים של דיקטיוזומים נוצרים מהרשת האנדופלזמית דרך שלב השלפוחית הביניימית, ושלפוחיות ההפרשה הנפרדות מהדיקטיוזומים תורמים בסופו של דבר להיווצרות קרום הפלזמה. לפיכך, הרשת האנדופלזמית והדיקטיוזומים מהווים יחידה תפקודית שבה הדיקטיוזומים ממלאים את התפקיד של מבני ביניים בתהליך המרת ממברנות כמו הרשת האנדופלזמית לממברנות כמו הפלזמה. ברקמות שהתאים שלהן גדלים ומתחלקים בצורה חלשה, מרכיבי הממברנה מתעדכנים כל הזמן.
מיקרוטובוליםנמצא כמעט בכל התאים האיקריוטיים. הם מבנים גליליים בקוטר של כ-24 ננומטר. אורכם משתנה. כל צינורית מורכבת מתת-יחידות של חלבון הנקרא טובולין. יחידות המשנה יוצרות 13 חוטים אורכיים המקיפים את החלל המרכזי. מיקרוטובולים הם מבנים דינמיים, הם נהרסים באופן קבוע ונוצרים בשלבים מסוימים של מחזור התא. ההרכבה שלהם מתרחשת במקומות מיוחדים, הנקראים מרכזי ארגון מיקרוטובולי. בתאי צמחים, יש להם מבנה אמורפי בעל ביטוי חלש.
פונקציות של microtubules: להשתתף ביצירת קרום התא; להפנות בועות של דיקטיוזומים לממברנה היוצרות, כמו חוטי ציר שנוצרים בתא מתחלק; ממלאים תפקיד ביצירת לוחית התא (הגבול הראשוני בין תאי הבת). בנוסף, microtubules הם מרכיב חשוב של flagella ו cilia, בתנועתם הם ממלאים תפקיד חשוב.
מיקרופילמנטים,כמו מיקרוטובולים, הם נמצאים כמעט בכל התאים האיקריוטיים. הם חוטים ארוכים בעובי 5-7 ננומטר, המורכבים מהחלבון המתכווץ אקטין. צרורות של מיקרופילמנטים נמצאים בתאים רבים של צמחים גבוהים יותר. ככל הנראה, הם ממלאים תפקיד חשוב בזרמי הציטופלזמה. מיקרופילמנטים, יחד עם מיקרוטובולים, יוצרים רשת גמישה הנקראת שלד ציטו.
חומר בסיסבמשך זמן רב נחשב לתמיסה הומוגנית (הומוגנית) עשירה בחלבון עם מספר קטן של מבנים או אפילו חסרת מבנה. אולם, נכון להיום, באמצעות מיקרוסקופ אלקטרונים במתח גבוה, נקבע כי החומר הקרקע הוא סריג תלת מימדי הבנוי מחוטים דקים (קוטר 3-6 ננומטר) הממלאים את התא כולו. מרכיבים אחרים של הציטופלזמה, כולל מיקרוטובולים ומיקרופילמנטים, תלויים מהסריג המיקרו-טראבקולרי הזה.
המבנה המיקרוטרבקולרי הוא סריג של גדילי חלבון, שהחלל ביניהם מלא במים. יחד עם מים, לסריג יש עקביות של ג'ל; הג'ל נראה כמו גופים ג'לטינים.
אברונים מחוברים לסריג המיקרוטרבקולרי. הסריג מספק תקשורת בין חלקים בודדים של התא ומכוון את ההובלה התוך תאית.
טיפות שומנים- מבנים בעלי צורה כדורית, המעניקים גרעיניות לציטופלזמה של תא צמחי תחת מיקרוסקופ אור. במיקרוגרפים של אלקטרונים, הם נראים אמורפיים. טיפות דומות מאוד אך קטנות יותר נמצאות בפלסידים.
טיפות ליפידים, בטעות שהן אברונים, קראו להן ספרוזומים והאמינו שהן מוקפות בקרום שכבה בודדת או כפולה. עם זאת, נתונים עדכניים מראים שלטיפות שומנים אין ממברנות, אך עשויות להיות מצופות בחלבון.
חומרים ארגסטיים -אלו הם "המוצרים הפסיביים" של הפרוטופלסט: חומרי רזרבה או פסולת. הם יכולים להופיע ולהיעלם בתקופות שונות של מחזור התא. מלבד גרגרי עמילן, גבישים, פיגמנטים אנתוציאנין וטיפות שומנים. אלה כוללים שרפים, מסטיקים, טאנינים וחלבונים. חומרים ארגאסטיים הם חלק מממברנת התא, החומר העיקרי של הציטופלזמה והאברונים, כולל ואקוולים.
פלאגלה וריסים -הם מבנים דקים דמויי שיער המשתרעים מפני השטח של תאים איקריוטיים רבים. יש להם קוטר קבוע, אבל האורך משתנה בין 2 ל 150 מיקרון. באופן קונבנציונלי, הארוכים והמעטים מהם נקראים דגלים, והקצרים והרבים יותר נקראים ריסים. אין הבדלים ברורים בין שני סוגי המבנים הללו, ולכן המונח flagellum משמש להתייחס לשניהם.
בחלק מהאצות והפטריות, הדגלים הם איברי תנועה, בעזרתם הם נעים במים. בצמחים (לדוגמה, טחבים, עצי כבד, שרכים, כמה ג'מנוספרמים), רק לתאי נבט (גמטות) יש דגלים.
לכל דגל יש ארגון ספציפי. טבעת חיצונית של 9 זוגות של מיקרו-צינוריות מקיפה שתי מיקרו-צינוריות נוספות הממוקמות במרכז הדגל. "ידיות" המכילות אנזים משתרעות ממיקרו-צינורית אחת של כל אחד מהזוגות החיצוניים. זהו דפוס הארגון הבסיסי של 9+2 שנמצא בכל הדגלים באורגניזמים איקריוטיים. מאמינים כי תנועת הפלגלה מבוססת על החלקה של מיקרוטובולים, בעוד שהזוגות החיצוניים של המיקרוטובולים נעים אחד לאורך השני ללא התכווצות. החלקה של זוגות מיקרוטובולים זה לזה גורמת לכיפוף מקומי של הדגל.
דגלים "צומחים" ממבנים גליליים ציטופלזמיים הנקראים גופים בזאליים, המהווים גם את החלק הבסיסי של הדגל. לגופים הבסיסיים יש מבנה פנימי הדומה לזה של דגל, אלא שהצינוריות החיצוניות מסודרות בשלישיות ולא בזוגות, והצינוריות המרכזיות נעדרות.
דופן תא.דופן התא תוחמת את גודל הפרוטופלסט ומונעת את קריעתו עקב ספיגת המים על ידי הוואקואול.
לדופן התא יש תפקידים ספציפיים שחשובים לא רק לתא ולרקמה שבה נמצא התא, אלא גם לצמח כולו. לדפנות התא תפקיד חיוני בספיגה, הובלה והפרשה של חומרים, ובנוסף, ניתן לרכז בהם פעילות ליזוזומלית, או עיכול.
רכיבי דופן התא. המרכיב האופייני ביותר של דופן התא הוא תאית, שקובעת במידה רבה את הארכיטקטורה שלו. מולקולות תאית מורכבות ממולקולות גלוקוז חוזרות המחוברות מקצה לקצה. מולקולות תאית דקות ארוכות משולבות למיקרו-סיביים בעובי של 10 - 25 ננומטר. מיקרופיברילים משתלבים זה בזה ויוצרים חוטים דקים, שבתורם ניתן ללפף אחד סביב השני, כמו חוטים בחבל. לכל "חבל", או מקרופיבריל כזה, עובי של כ-0.5 מיקרון, מגיע לאורך של 4 מיקרון. מקרופיברילים חזקים כמו חוטי פלדה בגודל שווה.
מסגרת התאית של דופן התא מלאה במולקולות מטריצת תאית השזורות בה. הוא מורכב מפוליסכרידים הנקראים hemicelluloses, ו-pectins, או pectins, כימית קרוב מאוד hemicelluloses.
מרכיב נוסף בדופן התא, ליגנין, הוא הפולימר הנפוץ ביותר בתאי הצמח לאחר תאית. לגנין מגביר את קשיחות הדופן ונמצא בדרך כלל בתאים המבצעים פונקציה תומכת או מכנית.
קוטין, סוברין, שעווה - בדרך כלל מופקדים בקונכיות של רקמות המגן של צמחים. קוטין, למשל, נמצא בקרומי התא של האפידרמיס, וסוברין נמצא ברקמת ההגנה המשנית, הפקק. שני החומרים מצויים בשילוב עם שעווה ומונעים איבוד יתר של מים מהצמח.
שכבות של דופן התא. עובי הדופן של תאי הצמח משתנה מאוד, בהתאם לתפקידם של התאים במבנה הצמח וגיל התא עצמו. תחת מיקרוסקופ אלקטרונים, שתי שכבות נראות בדופן התא הצמחי: הלמינה החציונית (המכונה גם החומר הבין-תאי), ודופן התא הראשוני. תאים רבים מניחים שכבה נוספת, דופן התא המשני. הצלחת החציונית ממוקמת בין הקירות הראשוניים של תאים סמוכים. הקיר המשני, אם בכלל, מופקד על ידי פרוטופלסט התא על פני השטח הפנימיים של דופן התא הראשוני.
צלחת אמצעית. הלמינה החציונית מורכבת בעיקר מפקטין. במקום שבו צריך להיווצר דופן תא, בין שני תאים שזה עתה נוצרו, מציינים לראשונה מקלעת צפופה של צינוריות של הרשת האנדופלזמית והבור של מנגנון הגולגי (דיקטיוזומים). ואז מופיעות בועות במקום הזה, מלאות בפקטין (מפוליסכרידים). הבועות הללו מופרדות מהמיכלים של מנגנון גולגי. דופן התא המוקדם מכיל פוליסכרידים שונים, כאשר העיקריים שבהם הם פקטין והמיצלולוז. מאוחר יותר, הוא מורכב מחומרים צפופים יותר - תאית וליגנין.
דופן תא ראשונית. זוהי שכבה של מעטפת תאית המונחת לפני או במהלך צמיחת התא. בנוסף לתאית, המיצלולוזה ופקטין, הממברנות הראשוניות מכילות גליקופרוטאין. קונכיות ראשוניות יכולות להתגרות. רכיב הפקטין מקנה פלסטיות, המאפשרת לקליפה הראשונית להתרחב ככל שהשורש, הגבעול או העלה מתארכים.
לתאים המתחלקים באופן פעיל (רוב התאים הבוגרים המעורבים בתהליכי הפוטוסינתזה, הנשימה וההפרשה) יש ממברנות ראשוניות. תאים כאלה עם ממברנה ראשונית ופרוטופלסט חי מסוגלים לאבד את צורתם האופיינית, להתחלק ולהתמיין לסוג תא חדש. הם מעורבים בריפוי פצעים והתחדשות רקמות בצמחים.
קרומי תאים ראשוניים אינם אחידים בעובי לכל אורכם, אך יש להם חתכים דקים, הנקראים שדות נקבוביות ראשוניים. גדילי הציטופלזמה, או plasmodesmata, המחברים את הפרוטופלסטים של תאים שכנים בדרך כלל עוברים דרך שדות הנקבוביות הראשוניים.
דופן תא משני. למרות העובדה שלתאים צמחיים רבים יש רק דופן ראשונית, בחלקם, הפרוטופלסט מפקיד דופן משנית לכיוון מרכז התא. זה מתרחש בדרך כלל לאחר הפסקת צמיחת התאים ושטח הממברנה הראשונית כבר לא גדל. מסיבה זו, הקליפה המשנית שונה מהראשונית. ממברנות משניות נחוצות במיוחד על ידי תאים מיוחדים המחזקים את הצמח ומוליכים מים. לאחר שקיעת הממברנה המשנית, הפרוטופלסט של תאים אלה, ככלל, מת. יש יותר תאית בקליפות המשניות מאשר בראשוניות, ונעדרים בהן חומרי פקטין וגליקופרוטאין. קשה למתוח את הממברנה המשנית, המטריצה שלו מורכבת מהמיצלולוזה.
במעטפת המשנית ניתן להבחין בשלוש שכבות - חיצוניות, אמצעיות ופנימיות (S1, S2, S3). מבנה השכבות של הקונכיות המשניות מגדיל משמעותית את חוזקן. מיקרו-סיבי תאית בממברנה המשנית מופקדים צפופים יותר מאשר בראשי. לגנין הוא מרכיב נפוץ בעורות המשניים של העץ.
הנקבוביות בממברנות של תאים מגע ממוקמות זו מול זו. שתי נקבוביות מנוגדות וקרום נקבוביות יוצרות זוג נקבוביות. בתאים עם ממברנות משניות, ישנם שני סוגים עיקריים של נקבוביות: פשוטות ותוחמות. בנקבוביות תוחמות, הקיר המשני תלוי מעל חלל הנקבוביות. בנקבוביות פשוטות, זה לא המקרה.
צמיחת דופן התא. ככל שהתא גדל, העובי והשטח של קרום התא גדלים. מתיחת מעטפת היא תהליך מורכב. הוא נשלט על ידי הפרוטופלסט ומווסת על ידי הורמון אוקסין.
בתאים הגדלים באופן אחיד לכל הכיוונים, שקיעת המיופיברילים היא אקראית. מיופיברילים אלה יוצרים רשת לא סדירה. תאים כאלה נמצאו בליבת הגזע, ברקמות האחסון ובמהלך גידול תאים במבחנה. בתאים מתארכים, המיופיברילים של המעטפת הצדדית מופקדים בזווית ישרה לציר ההתארכות.
חומרים מטריקס - פקטין, hemicelluloses וגליקופרוטאינים מועברים לממברנה בשלפוחיות של dictyosomes. יחד עם זאת, פקטין אופייניים יותר לתאים גדלים, בעוד שהמיצלולוזות שולטות בתאים שאינם גדלים.
מיקרו-סיבי תאית מסונתזים על פני התא בעזרת קומפלקס אנזים הקשור לממברנת הפלזמה. הכיוון של מיקרופיברילים נשלט על ידי מיקרוטובולים הממוקמים ליד המשטח הפנימי של קרום הפלזמה.
פלסמודזמה.אלו הם חוטים דקים של הציטופלזמה המחברים את הפרוטופלסטים של תאים שכנים. Plasmodesmata עוברים דרך דופן התא בכל מקום, או מרוכזים בשדות נקבוביות ראשוניים או בממברנות בין זוגות של נקבוביות. תחת מיקרוסקופ אלקטרונים, פלסמודסמטה מופיעות כתעלות צרות המרופדות בקרום הפלזמה. לאורך ציר התעלה משתרע מתא אחד למשנהו צינור גלילי קטן יותר, ה-desmotubule, המתקשר עם הרשת האנדופלזמית של שני התאים הסמוכים. פלסמודסמטה רבות נוצרות במהלך חלוקת התא, כאשר הרטיקולום האנדופלזמי הצינורי נקלט על ידי לוחית התא המתפתחת. Plasmodesmata יכול להיווצר גם בממברנות של תאים שאינם מתחלקים. מבנים אלה מספקים העברה יעילה של חומרים מסוימים מתא לתא.
חלוקת תא.באורגניזמים רב-תאיים, חלוקת תאים, יחד עם עלייה בגודלם, היא דרך לצמיחה של האורגניזם כולו. תאים חדשים שנוצרים במהלך החלוקה דומים במבנה ובתפקוד, הן לתא האב והן זה לזה. ניתן לחלק את תהליך החלוקה באאוקריוטים לשני שלבים חופפים חלקית: מיטוזה וציטוקינזיס.
מיטוזיס היא היווצרות של שני גרעיני בת מגרעין אחד, שוות ערך זה לזה מבחינה מורפולוגית וגנטית. ציטוקינזיס היא חלוקת החלק הציטופלזמי של התא עם היווצרות תאי בת.
מחזור התא.תא חי עובר סדרה של אירועים עוקבים המרכיבים את מחזור התא. משך המחזור עצמו משתנה בהתאם לסוג התא ולגורמים חיצוניים כמו טמפרטורה או זמינות חומרי הזנה. בדרך כלל המחזור מחולק ל-interphase ולארבעה שלבים של מיטוזה.
Interphase.התקופה שבין חלוקות מיטוטיות עוקבות.
האינטרפאזה מחולקת לשלוש תקופות, המסומנות כ-G1, S, G2.
במהלך תקופת G1, שמתחילה לאחר מיטוזה. במהלך תקופה זו, כמות הציטופלזמה עולה, כולל אברונים שונים. בנוסף, על פי ההשערה הנוכחית, חומרים מסונתזים במהלך תקופת G1 המעוררים או מעכבים את תקופת S ואת שאר המחזור, ובכך קובעים את תהליך החלוקה.
תקופת S עוקבת אחר תקופת G1, שבה החומר הגנטי (DNA) משוכפל.
במהלך תקופת G2, העוקבת אחרי S, נוצרים מבנים המעורבים ישירות במיטוזה, כמו רכיבי ציר.
חלק מהתאים עוברים מספר בלתי מוגבל של מחזורי תאים. אלה הם אורגניזמים חד-תאיים וכמה תאים של אזורי צמיחה פעילים (מריסטמים). כמה תאים מיוחדים לאחר התבגרות מאבדים את יכולתם להתרבות. הקבוצה השלישית של תאים, למשל, אלו היוצרים רקמת פצע (קלוס), שומרת על היכולת להתחלק רק בתנאים מיוחדים.
מיטוזה,או ביקוע גרעיני. זהו תהליך מתמשך המחולק לארבעה שלבים: פרופאזה, מטאפאזה, אנפאזה, טלופזה. כתוצאה מהמיטוזה, החומר הגנטי שהכפיל את עצמו ב-interphase מחולק שווה בשווה בין שני גרעיני הבת.
אחד הסימנים המוקדמים ביותר למעבר של תא לחלוקה הוא הופעת פס צר וטבעתי של מיקרו-צינוריות ממש מתחת לממברנת הפלזמה. חגורה צפופה יחסית זו מקיפה את הגרעין במישור המשווני של הציר המיטוטי העתידי. מכיוון שהיא מופיעה לפני הפרופזה, היא נקראת חגורת הקדם-פרופאזה. הוא נעלם לאחר הציר המיטוטי, הרבה לפני הופעתו בטלופזה המאוחר של לוחית התא, הגדל מהמרכז לפריפריה ומתמזג עם קרום תא האם באזור שנכבש בעבר על ידי חגורת הקדם-פרופאזה.
פרופאזה. בתחילת הפרופזה, הכרומוזומים דומים לחוטים ארוכים המפוזרים בתוך הגרעין. לאחר מכן, כשהגדילים מתקצרים ומתעבים, אפשר לראות שכל כרומוזום מורכב לא אחד, אלא משני גדילים שלובים זה בזה, הנקראים כרומטידים. בפרופאזה המאוחרת, שתי הכרומטידות הזוגיות המקוצרות של כל כרומוזום שוכבות זו לצד זו במקביל, מחוברות בקטע צר הנקרא צנטרומר. יש לו מיקום ספציפי על כל כרומוזום והוא מחלק את הכרומוזום לשתי זרועות באורכים שונים.
מיקרו-צינוריות ממוקמות במקביל לפני השטח של הגרעין לאורך ציר הציר. זהו הביטוי המוקדם ביותר של הרכבת ציר מיטוטי.
בסוף הפרופזה, הגרעין מאבד בהדרגה את קווי המתאר הברורים שלו ולבסוף נעלם. זמן קצר לאחר מכן מתפרקת גם המעטפת הגרעינית.
מטאפאזה. בתחילת המטאפאזה, הציר, שהוא מבנה תלת מימדי, הרחב ביותר באמצע ומצטמצם לכיוון הקטבים, תופס את המקום שתפס בעבר הגרעין. סיבי הציר הם צרורות של microtubules. במהלך המטאפאזה, הכרומוזומים, שכל אחד מהם מורכב משתי כרומטידות, מסודרים כך שהצנטרומרים שלהם נמצאים במישור המשווני של הציר. עם הצנטרומר שלו, כל כרומוזום מחובר לחוטי הציר. עם זאת, כמה גדילים עוברים מקוטב אחד למשנהו מבלי להיות מחוברים לכרומוזומים.
כאשר כל הכרומוזומים ממוקמים במישור המשווני, המטאפאזה הושלמה. כרומוזומים מוכנים להתחלק.
אנפאזה. הכרומטידות של כל כרומוזום נפרדות. עכשיו אלה כרומוזומי הבת. קודם כל, הצנטרומר מתחלק, ושני כרומוזומי הבת נמשכים לקטבים מנוגדים. במקרה זה, הצנטרומרים נעים קדימה, וזרועות הכרומוזומים נמתחות מאחור. חוטי הציר המחוברים לכרומוזומים מתקצרים, ותורמים להפרדת הכרומטידים ולתנועת כרומוזומי הבת בכיוונים מנוגדים.
טלופאז. בטלופאזה, ההפרדה של שתי קבוצות זהות של כרומוזומים הושלמה, בעוד שסביב כל אחת מהן נוצר קרום גרעיני. הרשת המחוספסת לוקחת חלק פעיל בכך. מנגנון הציר נעלם. במהלך הטלופאז, הכרומוזומים מאבדים את בהירות המתאר שלהם, נמתחים, הופכים בחזרה לחוטים דקים. הנוקלאולים משוחזרים. כאשר הכרומוזומים הופכים בלתי נראים, המיטוזה הושלמה. שני גרעיני הבת נכנסים לשלבים. הם שקולים מבחינה גנטית זה לזה ולגרעין האב. זה חשוב מאוד, שכן יש להעביר את התוכנית הגנטית, ואיתה כל הסימנים, לאורגניזמים הבתים.
משך המיטוזה משתנה באורגניזמים שונים ותלוי בסוג הרקמה. עם זאת, הפרופזה היא הארוכה ביותר והאנפאזה היא הקצרה ביותר. בתאי קצה השורש, משך הפרופזה הוא 1-2 שעות; מטאפאזה - 5 - 15 דקות; אנפאזה - 2 - 10 דקות; טלופאז - 10 - 30 דקות. משך האינטרפאז הוא בין 12 ל-30 שעות.
בתאים איקריוטים רבים, מרכזי ארגון המיקרוטובוליים האחראים על היווצרות הציר המיטוטי קשורים לצנטריולים.
ציטוקינזיס.זהו תהליך החלוקה של הציטופלזמה. ברוב האורגניזמים, התאים מתחלקים ע"י נסיגת קרום התא ויצירת תלם ביקוע, אשר מעמיק בהדרגה, ודוחס את שאר החוטים של הציר המיטוטי. בכל הצמחים (בריופיטים וכלי דם) ובחלק מהאצות, תאים מתחלקים עקב היווצרות של לוחית תא.
בטלופאז המוקדם נוצרת מערכת סיבים בצורת חבית הנקראת פרגמופלסט בין שני גרעיני הבת. סיבי הפרגמופלסט, כמו סיבי הציר המיטוטי, מורכבים ממיקרוטובולים. טיפות קטנות מופיעות במישור המשווני של הפרגמופלסט. הם מתמזגים ויוצרים לוחית תא שגדלה עד שהיא מגיעה לדופן התא המתחלק. זה משלים את ההפרדה של שני תאי הבת. טיפות מתמזגות הן בועות המתנתקות ממנגנון גולגי. בעיקרון, הם מכילים חומרי פקטין, שמהם נוצרת הצלחת האמצעית. ממברנות שלפוחית מעורבות בבניית קרום הפלזמה משני צידי הצלחת. במקביל, plasmodesmata נוצרים משברים של הרשת האנדופלזמית הצינורית.
לאחר היווצרות הצלחת החציונית, כל פרוטופלסט מפקיד עליו קרום ראשוני. בנוסף, כל תא בת מפקיד שכבה חדשה של ממברנה סביב הפרוטופלסט כולו, שממשיכה את הממברנה שעלתה מלוחית התא. המעטפת המקורית של תא האב נהרסת ככל שתאי הבת גדלים.
סוגים שונים של מיטוזה איקריוטית
חלוקת התא שתוארה לעיל בצמחים, וגם בבעלי חיים, אינה הצורה היחידה של חלוקת תאים עקיפה. הסוג הפשוט ביותר של מיטוזה הוא pleuromitosis.זה דומה לביקוע בינארי של תאים פרוקריוטים, שבהם הנוקלואידים לאחר השכפול נשארים קשורים לממברנת הפלזמה. הממברנה מתחילה לצמוח בין נקודות הקישור של ה-DNA ובכך מפזרת את הכרומוזומים לחלקים שונים של התא. לאחר מכן, במהלך היווצרות התכווצות התא, כל אחת ממולקולות ה-DNA תהיה בתא נפרד חדש.
אופייני לחלוקה של תאים אוקריוטיים הוא היווצרות של ציר הבנוי ממיקרוטובוליות. בְּ pleuromitosis סגור(זה נקרא סגור מכיוון שההתבדלות של הכרומוזומים מתרחשת מבלי לשבש את המעטפת הגרעינית) לא צנטריולים, אלא מבנים אחרים הממוקמים בצד הפנימי של הממברנה הגרעינית, משתתפים כמרכזי ארגון מיקרו-צינוריות (MCMT). אלה הם מה שנקרא גופים קוטביים של מורפולוגיה בלתי מוגדרת, שמהם משתרעים מיקרוטובולים. יש שניים מהגופים האלה. הם מתפצלים זה מזה מבלי לאבד קשר עם המעטפת הגרעינית. כתוצאה מכך, נוצרים שני חצאי צירים הקשורים לכרומוזומים. כל התהליך של יצירת ציר מיטוטי והפרדת כרומוזומים במקרה זה מתרחש מתחת למעטפת הגרעינית. סוג זה של מיטוזה מתרחש בקרב פרוטוזואה ומופץ באופן נרחב בפטריות (chytridia, zygomycetes, שמרים, oomycetes, ascomycetes, myxomycetes, וכו'). לפגוש טפסים pleuromitosis חצי סגורהכאשר המעטפת הגרעינית נהרסת בקטבים של הציר שנוצר.
הצורה הבאה של מיטוזה היא אורתומיטוזיס.במקרה זה, ה-COMTs ממוקמים בציטופלזמה; מההתחלה, לא נוצרים חצאי צירים, אלא ציר דו-קוטבי. יש שלוש צורות אורתומיטוזיס (מיטוזה רגילה) חצי סגורו סָגוּר. באורתומיטוזיס סגורה למחצה, נוצר ציר ביסימטרי בעזרת TsOMT הממוקם בציטופלזמה, המעטפת הגרעינית נשמרת לאורך כל המיטוזה, למעט אזורי הקוטב. מסות של חומר גרגירי, או אפילו צנטריולים, עשויות להתגלות כ-COMT. צורה זו של מיטוזה מצויה ב-zoospores של אצות ירוקות, חומות ואדומות, בחלק מהפטריות הנמוכות יותר ובגרגרינים. עם אורתומיטוזיס סגור, הממברנה הגרעינית נשמרת לחלוטין, שבה נוצר ציר אמיתי. מיקרוטובולים נוצרים בקאריופלזמה, לעתים רחוקות יותר הם גדלים מה-TsOMT הפנימי, שאינו קשור (בניגוד לפלורומיטוזיס) עם הממברנה הגרעינית. סוג זה של מיטוזה אופייני לחלוקה של מיקרו-גרעיני ריסים, אך יכול להתרחש גם בפרוטוזואה. באורתומיטוזיס פתוח, המעטפת הגרעינית מתפרקת לחלוטין. סוג זה של חלוקת תאים אופייני לאורגניזמים של בעלי חיים, כמה פרוטוזואה ותאים של צמחים גבוהים יותר. צורה זו של מיטוזה, בתורה, מיוצגת על ידי טיפוסים אסטרליים ואנסטרליים.
מהחומר שנסקר בקצרה, ניתן לראות כי המאפיין העיקרי של מיטוזה באופן כללי הוא הופעת מבני ציר ביקוע, הנוצרים בקשר עם ה-CMT, המגוונים במבנה שלהם.
מורפולוגיה של הדמות המיטוטית
המנגנון המיטוטי בא לידי ביטוי טוב במיוחד בשלב המטאפאזה של המיטוזה. במטאפאזה, במישור המשווני של התא, ממוקמים כרומוזומים, שמהם נמשכים חוטי הציר המכונים בכיוונים מנוגדים, ומתכנסים בשני קטבים שונים של הדמות המיטוטית. לפיכך, הציר המיטוטי הוא אוסף של כרומוזומים, קטבים וסיבים. סיבי הציר הם מיקרוטובולים בודדים או צרורותיהם. מיקרוטובולים מתחילים מעמודי הציר, וחלקם הולכים לצנטרומריםהיכן ממוקמים kinetochoresכרומוזומים (kinetochore microtubules), חלקם עוברים הלאה לכיוון הקוטב הנגדי, אך אינם מגיעים אליו. הם נקראים "מיקרוטובולים בין קוטביים". מהקטבים יוצאת קבוצה של מיקרוטובולים רדיאליים, ויוצרת סביבם מבנה הדומה ל"זוהר קורן" – אלו הן מיקרוטובולים אסטרליים.
לפי מורפולוגיה, דמויות מיטוטיות מחולקות לסוגים אסטרליים ואנסטרליים.
סוג הציר האסטרלי, או מתכנס, מאופיין בכך שהקטבים שלו מיוצגים על ידי אזור קטן שאליו מתכנסות (מתכנסות) המיקרוטובולים. בדרך כלל, צנטרוזומים המכילים צנטרולים ממוקמים בקטבים של הצירים האסטרליים. עם זאת, ידועים מקרים של מיטוזות אסטרליות צנטרולריות (במהלך מיוזה של חלק מחסרי חוליות). בנוסף, מציינים מיקרוטובולים רדיאליים המקרינים מהקטבים, שאינם חלק מהציר, אלא יוצרים אזורי כוכבים - ציסטרים. סוג זה של חלוקה מיטוטית דומה למשקולת. לסוג האנסטריאלי של הדמות המיטוטית אין ציסטרים בקטבים. האזורים הקוטביים של הציר כאן רחבים, הם נקראים כובעים קוטביים, הם כוללים centrioles. במקרה זה, סיבי הציר אינם יוצאים מנקודה אחת, אלא מתפצלים בחזית רחבה (מתפצלים) מכל אזור כובעי הקוטב. סוג זה של ציר מאפיין תאים מתחלקים של צמחים גבוהים יותר, אך ניתן למצוא אותו גם בבעלי חיים גבוהים יותר. בעוברות מוקדמת של יונקים, מיטוזות צנטרולריות (מתפצלות) נצפות במהלך חלוקת הבשלת הביציות ובחלוקות I ו-II של הזיגוטה. אבל כבר בחלוקת התא השלישי ובכל התאים שלאחר מכן, הם מתחלקים בהשתתפות צירים אסטרליים, שבקטבים שלהם נמצאים תמיד צנטרולים. באופן כללי, עבור כל צורות המיטוזה, הכרומוזומים עם הקינטופורים שלהם, הגופים הקוטביים (צנטרוזומים) וסיבי הציר נשארים מבנים נפוצים.
צנטרומרים וקינטוכורים
לצנטרומרים יכולה להיות לוקליזציה שונה לאורך הכרומוזומים. צנטרומרים הולוצנטריים מתרחשים כאשר מיקרוטובולים מקושרים לאורך כל הכרומוזום (חלק מהחרקים, נמטודות, חלק מהצמחים). צנטרומרים מונוצנטריים - כאשר מיקרוטובולים קשורים לכרומוזומים באתר אחד. צנטרומרים מונוצנטריים יכולים להיות מנוקדים (לדוגמה, בחלק מהשמרים הניצנים), כאשר רק מיקרו-צינורית אחת מתקרבת לקינטוכור, וזונאלית, שבה צרור מיקרו-צינוריות מתקרב לקינטוכור המורכב. למרות המגוון של אזורי הצנטרומרים, כולם קשורים למבנה המורכב של הקינטוכור, שיש לו דמיון בסיסי במבנה ובתפקודים בכל האיקריוטים. קינטוכורים הם מבני חלבון מיוחדים, הממוקמים בעיקר באזורי הצנטרומרים של הכרומוזומים. אלו קומפלקסים מורכבים המורכבים מחלבונים רבים. מבחינה מורפולוגית, הם דומים מאוד, בעלי אותו מבנה, החל מדיאטומים לבני אדם. הם מבנים תלת-שכבתיים: השכבה הצפופה הפנימית הצמודה לגוף הכרומוזום, השכבה האמצעית הרפויה והשכבה הצפופה החיצונית. סיבים רבים משתרעים מהשכבה החיצונית, ויוצרים את מה שנקרא הכתר הסיבי של הקינטוכור. בצורה הכללית, קינטוכורים נראים כמו לוחות או דיסקים השוכבים באזור ההתכווצות הראשונית של הכרומוזום, בצנטרומר. בדרך כלל יש קינטוכור אחד לכל כרומוזום או כרומטיד. לפני אנפאזה, קינטוכורים על כל כרומטיד אחות ממוקמים, כל אחד מתקשר עם צרור המיקרוטובולים שלו. בצמחים, הקינטוכור אינו נראה כמו לוחות, אלא כחצי כדור. התפקיד הפונקציונלי של קינטוכורים הוא לקשור כרומטידות אחיות זו לזו, לקבע מיקרוטובולים מיטוטיים, לווסת הפרדת כרומוזומים ולמעשה להזיז כרומוזומים במהלך מיטוזה בהשתתפות מיקרוטובולים. באופן כללי, מבני חלבון, קינטוכורים, מוכפלים בתקופת S, במקביל להכפלת הכרומוזומים. אבל החלבונים שלהם נמצאים על הכרומוזומים בכל תקופות מחזור התא.
רקמת צמחים
רקמות חינוכיות (מריסטמים)
רקמות חינוכיות בגוף הצמחים ממוקמות במקומות שונים, ולכן הן מחולקות לקבוצות הבאות (איור 0;1).
מריסטמות אפיקליות (אפיקליות) ממוקמות בחלק העליון, או הקודקודים, של האיברים הציריים - גזע, שורש. בעזרת המריסטמים הללו, האיברים הצומחיים של הצמחים מבצעים את צמיחתם לאורך.
מריסטמים לרוחב אופייניים לאיברים צירים. שם הם ממוקמים באופן קונצנטרי, בצורה של מצמד.
מריסטמים בין קלוריות, או בין קלוריות, מקורם במריסטמים אפיקליים. אלו קבוצות של תאים שעדיין לא מסוגלות להתרבות, אבל יצאו לדרך של התמיינות. אין ביניהם תאים ראשוניים, אבל יש הרבה תאים מיוחדים.
מריסטמים פצעים מספקים שיקום של החלק הפגוע בגוף. התחדשות מתחילה עם דה-דיפרנציאציה, כלומר, התפתחות הפוכה מתאי מיוחדים לתאים מריסטמטיים. הם הופכים לפלוגן, היוצר פקק המכסה את פני הפצע. תאים לא מובחנים, מתחלקים, יכולים ליצור רקמה פרנכימלית רופפת - קאלוס. ממנו, בתנאים מסוימים, נוצרים איברי צמחים.
רקמות אינטגמנטרי
הם פועלים כמחסום גבול, המפריד את הרקמות הבסיסיות מהסביבה. המרכיבים העיקריים של צמח מורכבים רק מתאי חיים. אינטגמנט משני ושלישוני - בעיקר מהמתים עם דפנות תאים עבות.
התפקודים העיקריים של רקמות המוח:
הגנה על הצמח מפני ייבוש;
הגנה מפני חדירת מיקרואורגניזמים מזיקים;
הגנה מפני כוויות שמש;
הגנה מפני נזק מכני;
ויסות חילוף החומרים בין הצמח לסביבה;
תפיסה של גירוי.
הרקמה העיקרית של האפידרמיס היא האפידרמיס, האפידרמיס. מורכב מתאי חיים. נוצר ממריסטמים אפיקליים. מכסה גבעולים ועלים צעירים.
האפידרמיס נוצר בצמחים בקשר ליציאה מבית הגידול המימי ליבשה על מנת למנוע את התייבשותו. בנוסף לסטומטה, כל תאי האפידרמיס מחוברים זה לזה. הקירות החיצוניים של התאים הראשיים עבים יותר מהשאר. כל פני השטח מכוסים בשכבת קאטין ושעוות ירקות. שכבה זו נקראת הקוטיקולה (עור). הוא נעדר על השורשים הגדלים וחלקים תת-מימיים של צמחים. כאשר הוא יבש, החדירות של הציפורן נחלשת באופן משמעותי.
בנוסף לתאים העיקריים, ישנם אחרים באפידרמיס, במיוחד שערות או טריכומות. הם חד-תאיים ורב-תאיים (איור 2).מבחינה פונקציונלית, הם מגדילים את פני האפידרמיס, למשל, באזור הצמיחה של השורשים, משמשים כהגנה מכנית, נצמדים לתמיכה ומפחיתים את איבוד המים. למספר צמחים יש שערות בלוטות, כמו סרפדים.
רק לצמחים גבוהים יותר יש סטמטות באפידרמיס, המווסתות את חילופי המים והגזים. אם אין לציפורן, אז אין צורך בסטומטה. סטומות הן קבוצת תאים היוצרים את המנגנון הסטומטלי, המורכב משני תאים נגררים ותאי אפידרמיס סמוכים - תאי צד. הם שונים מתאי האפידרמיס העיקריים (איור 3). תאי שמירה שונים מהתאים הסובבים אותם בצורתם ובנוכחותם של מספר רב של כלורופלסטים וקירות מעובים בצורה לא אחידה. אלה שניצבים זה מול זה עבים יותר מהשאר (איור 4). בין תאי השמירה נוצר מרווח סטמאטלי המוביל לחלל תת-סטומטי הנקרא חלל התת-סטומטי. תאי שמירה הם בעלי פוטוסינתזה גבוהה. הם מכילים כמות גדולה של עמילן רזרבה ומיטוכונדריה רבים.
המספר והתפוצה של סטומטות, סוגי מנגנון הסטומטלי משתנים מאוד בצמחים שונים. סטמאטה נעדרת אצל בריופיטים מודרניים. הפוטוסינתזה בהם מתבצעת על ידי דור הגמטופיטים, והספורופיטים אינם מסוגלים להתקיים עצמאי.
בדרך כלל סטמטות ממוקמות בחלק התחתון של העלה. בצמחים הצפים על פני המים - במשטח העליון. בעלים של דגנים, העגבניות מפוזרות לרוב באופן שווה משני הצדדים. עלים כאלה מוארים באופן שווה יחסית. על 1 מ"מ של פני השטח, מ 100 עד 700 stomata ניתן לאתר.
רקמה משנית (periderm). רקמה זו מחליפה את האפידרמיס כאשר הצבע הירוק של הנבטים השנתיים משתנה לחום. הוא רב-שכבתי ומורכב משכבה מרכזית של תאים קמביאליים - פלוגן. תאי פלוגן, המתחלקים, מניחים שכבת פלם מבחוץ ופלודרם מבפנים. (איור 5).
פלמה, או פקק. בתחילה הוא מורכב מתאי חיים בעלי דופן דקה. עם הזמן, הקירות שלהם ספוגים בשעוות סוברין ושעוות ירקות ומתים. תוכן התא מתמלא באוויר.
פונקציות פלמה:
מונע אובדן לחות;
מגן על הצמח מפני נזק מכני;
מגן מפני פתוגנים;
מספק בידוד תרמי, שכן התאים מתמלאים באוויר.
תאים של הפלוגן, הממוקמים באפידרמיס עצמו, השכבה התת אפידרמיסית הבסיסית, לעתים רחוקות יותר בשכבות העמוקות של הקורטקס הראשוני, הם הבסיס היוצר של הקורטקס הראשוני.
שכבת הפקק אינה קבועה. מתרחשות בו הפסקות, המתקשרות עם החללים הבין-תאיים הנמצאים בקרבת מקום. במקביל, נוצרות על פני השטח פקעות קטנות - עדשים, המתקשרות את החללים הבין-תאיים עם אוויר אטמוספרי. (איור 6,7).
בסתיו, הפלוגן מתחת לעדשים מניח שכבה של תאים פקוקים, אשר מפחיתים מאוד את הטרנספירציה, אך אינם שוללים אותה לחלוטין. באביב, שכבה זו קורסת מבפנים. על קליפת ליבנה בהירה, עדשים נראות בבירור בצורה של קווים כהים.
גם הרקמה השלישונית (הקרום) אופיינית רק לצורות עץ של צמחים.
הפלוגן מונח שוב ושוב בשכבות העמוקות של הקורטקס. הרקמות שנמצאות מחוצה לו בסופו של דבר מתות ויוצרות קרום. התאים שלו מתים ואינם מסוגלים להימתח. עם זאת, תאים חיים הממוקמים עמוק יותר מתחלקים, מה שמוביל לעלייה בגודל הרוחבי של הגבעול. עם הזמן, השכבה החיצונית של הקרום נשברת. זמן התרחשות של פער כזה הוא ערך קבוע למדי עבור צמחים בודדים. בעץ תפוח זה קורה בשנה השביעית לחיים, בקרן - בחמישים. במינים מסוימים הוא אינו מופיע כלל. תפקידו העיקרי של הפילינג הוא הגנה מפני נזקים מכניים ותרמיים.
פרנכימה
זוהי קבוצה של רקמות מיוחדות הממלאות את החללים בתוך גוף הצמח בין רקמות מוליכות למכניות. (איור 8).לעתים קרובות יותר לתאי פרנכימה יש צורה מעוגלת, לעתים רחוקות יותר צורה מוארכת. נוכחותם של חללים בין-תאיים מפותחים אופיינית. המרווחים בין התאים יוצרים יחד מערכת הובלה - האפופלסט. בנוסף, החללים הבין-תאיים מהווים את "מערכת האוורור" של הצמח. באמצעות סטמטות, או עדשים, הן מחוברות לאוויר אטמוספרי ומספקות הרכב גז אופטימלי בתוך הצמח. חללים בין-תאיים מפותחים נחוצים במיוחד עבור צמחים הגדלים על אדמה ספוגת מים, שבה חילופי גזים רגילים קשה. פרנכימה זו נקראת aerenchyma. (איור 9).
אלמנטים של הפרנכימה, הממלאים את הפערים בין רקמות אחרות, מבצעים גם את הפונקציה של תמיכה. תאי פרנכימה חיים, אין להם דפנות תאים עבות, כמו סקלרנכימה. לכן, תכונות מכניות מסופקות על ידי טורגור. אם תכולת המים יורדת, מה שמוביל לפסמוליזה ונבילה של הצמח.
פרנכימה ההטמעה נוצרת על ידי תאים בעלי דופן דקה עם חללים בין-תאיים רבים. התאים של מבנה זה מכילים כלורופלסטים רבים, ולכן זה נקרא כלורנכימה. כלורופלסטים ממוקמים לאורך הקיר ללא הצללה זה על זה. בפרנכימה ההטמעה מתרחשות תגובות פוטוסינתזה, המספקות לצמח חומרים אורגניים ואנרגיה. התוצאה של תהליכי פוטוסינתזה היא האפשרות לקיומם של כל היצורים החיים על פני כדור הארץ.
רקמות הטמעה קיימות רק בחלקים המוארים של הצמח; הן מופרדות מהסביבה על ידי אפידרמיס שקוף. אם האפידרמיס מוחלף ברקמות אינגומנטריות משניות אטומות, פרנכימה ההטמעה נעלמת.
פרנכימת האגירה משמשת כלי קיבול לחומרים אורגניים שאינם בשימוש זמנית על ידי האורגניזם הצמחי. באופן עקרוני, כל תא עם פרוטופלסט חי מסוגל להפקיד חומר אורגני בצורה של סוגים שונים של תכלילים, אך חלק מהתאים מתמחים בכך. (איור 10, 11).תרכובות עשירות באנרגיה מופקדות רק בעונת הגידול, נצרך בתקופה הרדומה ולקראת עונת הגידול הבאה. לכן, חומרי רזרבה מופקדים באיברים הצומחים רק בצמחים רב-שנתיים.
כלי הקיבול של המניות יכול להיות איברים רגילים (בריחה, שורש), כמו גם מתמחים (קני שורש, פקעות, פקעות). כל צמחי הזרע אוגרים חומרים בעלי ערך אנרגטי בזרעים (קוטילונים, אנדוספרם). צמחים רבים באקלים צחיח אוגרים לא רק חומרים אורגניים, אלא גם מים. (איור 12).לדוגמה, אלוורה אוגרת מים בעלים בשרניים, קקטוסים בנצרים.
בדים מכניים
התכונות המכניות של תאי צמחים מסופקות על ידי:
קליפה קשה של התא;
עכירות, כלומר, מצב הטורגור של תאים.
למרות העובדה שכמעט לכל תאי הרקמה יש תכונות מכניות, עם זאת, יש רקמות בצמח שתכונות מכניות הן בסיסיות עבורן. אלו הן קולנכימה וסקלנכימה. בדרך כלל הם מתפקדים באינטראקציה עם רקמות אחרות. בתוך הגוף יוצרים הצמחים מעין מסגרת. לכן, הם נקראים חיזוק.
לא לכל הצמחים יש רקמות מכניות מוגדרות היטב. במידה הרבה פחות, צמחים החיים בסביבה המימית זקוקים לתמיכה פנימית מאשר יבשתיים. הסיבה היא שצמחי מים זקוקים פחות לתמיכה פנימית. גופם נתמך במידה רבה על ידי המים שמסביב. אוויר ביבשה אינו יוצר תמיכה כזו, שכן יש לו צפיפות נמוכה יותר מאשר מים. מסיבה זו נוכחותם של רקמות מכניות מיוחדות הופכת לרלוונטית.
שיפור של מבני תמיכה פנימיים התרחש בתהליך האבולוציה.
קולנכימה. הוא נוצר רק על ידי תאים חיים, מוארכים לאורך ציר האיבר. סוג זה של רקמה מכנית נוצר מוקדם מאוד, במהלך תקופת הצמיחה הראשונית. לכן, חשוב שהתאים יישארו בחיים וישמרו על יכולת המתיחה יחד עם תאי המתיחה שנמצאים בקרבת מקום.
תכונות של תאי קולנכימה:
עיבוי לא אחיד של הקליפה, וכתוצאה מכך חלקים מסוימים ממנה נשארים דקים, בעוד שאחרים מתעבים;
קונכיות אינן מתגרות.
תאי קולנכימה ממוקמים באופן שונה זה מזה. בתאים סמוכים נוצרים עיבויים בפינות הפונות זו לזו ( אורז. 13). קולנכימה כזו נקראת קולנכימה פינתית. במקרה אחר, התאים מסודרים בשכבות מקבילות. קרומי התא הפונים לשכבות אלו מעובות מאוד. זוהי קולנכימה למלרית. תאים יכולים להיות ממוקמים בצורה רופפת, עם מרווחים בין-תאיים בשפע - זוהי קולנכימה רופפת. קולנכימה כזו נמצאת לעתים קרובות בצמחים על קרקעות ספוגות מים.
לקולנכימה חשיבות מיוחדת בצמחים צעירים, בצורות עשבוניות, וגם בחלקי צמחים שבהם לא מתרחשת צמיחה משנית, כגון עלים. במקרה זה, הוא מונח קרוב מאוד לפני השטח, לפעמים ממש מתחת לאפידרמיס. אם לאיבר יש פרצופים, אז שכבות עבות של קולנכימה נמצאות לאורך הפסגות שלהן.
תאי קולנכימה מתפקדים רק בנוכחות טורגור. מחסור במים מפחית את יעילות הקולנכימה והצמח קמל באופן זמני, למשל, עלי מלפפון נפולים ביום חם. לאחר מילוי התאים במים, הפונקציות של הקולנכימה משוחזרות.
סקלרנכימה. הסוג השני של רקמות מכניות. בניגוד לקולנכימה, שבה כל התאים חיים, תאי סקלנכימה מתים. הקירות שלהם עבים מאוד. הם מבצעים פונקציה מכנית. עיבוי חזק של הממברנה מוביל להפרה של הובלה של חומרים, וכתוצאה מכך הפרוטופלסט מת ( אורז. 14). הליגניפיקציה של ממברנות התא של סקלרנכימה מתרחשת כאשר איבר הצמח כבר השלים את צמיחתו. לכן, הם כבר לא מפריעים למתיחה של הרקמות הסובבות.
בהתאם לצורה, מבחינים בין שני סוגים של תאי סקלרנכימה - סיבים וסקלרים.
לסיבים צורה מוארכת מאוד עם קירות עבים מאוד וחלל קטן. הם קצת יותר קטנים מסיבי עץ. לעתים קרובות מתחת לאפידרמיס יוצרים שכבות אורך וגדילים. בפלואם או בקסלם ניתן למצוא אותם בנפרד או בקבוצות. בפלומה הם נקראים סיבי באסט ( אורז. 15), ובקסילם - סיבים ליבריפורמים ( אורז. 16).
Sclereids, או תאים אבנים, מיוצגים על ידי תאים מעוגלים או מסועפים עם ממברנות חזקות. בגופו של צמח ניתן למצוא אותם בנפרד (תאים תומכים) ( איור.17) או קבוצות (איור 18). יש לציין כי המאפיינים המכניים תלויים מאוד במיקום הסקלרידים. חלק מהסקלרים יוצרים שכבות רצופות, כמו למשל בקליפת האגוזים או באבני הפירות (פירות האבן).
רקמות הפרשה
לכל תא חי יש בתחילה את הפונקציה של סילוק חומרים, בעוד שההובלה יכולה לעבור הן לאורך שיפוע הריכוז והן כנגד השיפוע. ניתן להסיר חומרים שעברו סינתזה בתא ובהמשך ישפיעו על פעילותם של תאים אחרים (פיטו-הורמונים, אנזימים). תהליך זה נקרא הפרשה. במקרה שבו מופרשים חומרים שהם תוצרי פסולת של התא, התהליך נקרא הפרשה. למרות העובדה שלהפרשת חומרים בתאי צמחים ובעלי חיים יש דמיון מהותי, לצמחים יש מספר תכונות הנובעות מהבדלים מהותיים בפעילות החיונית. רמת התגובות המטבוליות בצמחים נמוכה בהרבה מאשר בבעלי חיים. לכן, כמויות קטנות יותר של פסולת נפלטים באופן יחסי. תכונה נוספת היא שבסינתזה עצמאית של כמעט כל התרכובות האורגניות הדרושות, צמחים לעולם אינם יוצרים עתודות מוגזמות שלהם. החומרים המשתחררים יכולים לשמש כחומר מוצא לתגובות אחרות (לדוגמה, CO2 ו- H2O).
אם אצל בעלי חיים תהליך הפרשת הרעלים קשור להסרתם מהגוף, ייתכן שהצמחים לא יעשו זאת, ויבודדו חומרים מיותרים בפרוטופלסט החי (הסרה של חומרים שונים לתוך הוואקואול), בתאים מתים (רוב הרקמות של צמחים רב שנתי). הצמח מורכב מתאים כאלה), בחללים בין-תאיים.
ניתן לחלק את החומרים הנפלטים לשתי קבוצות גדולות. הקבוצה הראשונה היא חומרים אורגניים המסונתזים ישירות על ידי התא (אנזימים, פוליסכרידים, ליגנינים, טרפנים, האחרונים הם מרכיבים מרכיבים של שמנים אתריים ושרף). הקבוצה השנייה - חומרים הנכנסים בתחילה לתא מבחוץ בעזרת שינוע קסילם או פלואם (מים, מלחים מינרלים, חומצות אמינו, חד סוכרים ועוד). הסוד המופרש לעיתים רחוקות הוא הומוגני ובדרך כלל מורכב מתערובת שבה חומר אחד הוא בעל הריכוז הגבוה ביותר.
לצמחים, בניגוד לבעלי חיים, אין מערכת הפרשה מלאה. ישנם רק מבנים מיוחדים הפזורים ברחבי הצמח - אידיובלסטים. התאים היוצרים אותם קטנים יותר מתאי הפרנכימה השוכנים בקרבת מקום. יש להם ציטופלזמה צפופה באלקטרונים, עם אלמנטים מפותחים של הרשת האנדופלזמית וקומפלקס גולגי. לרוב לא בא לידי ביטוי, יש וואקוול מרכזי. בינם לבין תאים חיים אחרים, תאים אלה מחוברים על ידי פלסמודסמטה רבים.
בהתאם למיקום, רקמות ההפרשה יכולות להיות חיצוניות ופנימיות.
רקמות הפרשה חיצוניות.שערות בלוטות הן נגזרות של האפידרמיס. מבחינה מורפולוגית הם משתנים - הם יכולים להיות בעלי ראש רב תאי, להיות מוארכים, בצורת מגן על רגל וכו'. (איור 19). אלה כוללים שערות סרפד צורבות.
הידתודות הן מבנים המשחררים עודפי מים בתנאים של טרנספירציה נמוכה ולחות גבוהה - גטציה. מבנים אלה נוצרים על ידי קבוצות של תאים חיים חסרי צבע עם קירות דקים - התמה. רקמה זו צמודה לצרור המוליך (איור 20). מים משתחררים דרך סטמטות מים מיוחדות, הנבדלות מהסטומטות רגילות על ידי חוסר תנועה ופער פתוח כל הזמן. הרכב נוזל הביוב משתנה מאוד ממים כמעט טהורים ועד לתערובת מורכבת מאוד של חומרים.
נקטריות. צמחים רבים מתאפיינים בשחרור נוזל המכיל בין 7 ל-87% חד- ואוליגוסכרידים. תהליך זה מתבצע על ידי מבנים מיוחדים - נקטריות. בהתאם למיקום, פרח או פרחוני, נבדלים נקטריונים (איור 21), וכן צמחים הממוקמים על גבעולים ועלים - נקטרי חוץ פרחוני, או חוץ פרחוני. צוף יכול להיות מיץ פלואם ללא שינוי, אשר מועבר אל פני השטח דרך החללים הבין-תאיים ומופרש דרך הסטומטה. נקטרים מורכבים יותר נוצרים על ידי פרנכימה בלוטית, מכוסה באפידרמיס עם שערות בלוטות. הצוף מופרש על ידי תאי אפידרמיס או שערות בלוטות. במקרה זה, הצוף המופרש שונה ממוהל הפלואם. מכיוון שהוא נשלט על ידי גלוקוז, פרוקטוז, סוכרוז, ובמיץ פלואם - גלוקוז. יונים נמצאים בריכוז נמוך. יונים נמצאים בריכוז נמוך. כדי למשוך מאביקים, הצוף עשוי להכיל הורמונים סטרואידים, הנחוצים לחרקים.
בלוטות עיכול קיימות בצמחים אוכלי חרקים (טל שמש, nepenthys וכו'). בתגובה למגע של חסרי חוליות קטנים, הם מפרישים סוד המכיל אנזימים הידרוליטיים, חומצות פורמיות, בוטיריות ופרופיוניות. כתוצאה מכך, ביופולימרים מתפרקים למונומרים, המשמשים את המפעל לצרכיו.
בלוטות מלח מתפתחות בצמחים הגדלים על קרקעות מלוחות. בלוטות אלה ממוקמות על העלים ובצורה מומסת, יונים עודפים מועלים אל פני השטח. מלח מופקד תחילה על הציפורן, ואז הוא נשטף בגשם (איור 22). בנוסף, לצמחים רבים יש שערות מלח על העלים. כל שערה מורכבת משני תאים: האחד יוצר את הראש, השני - הגבעול. מלחים מצטברים ב-vacuole של תא הראש, וכאשר ריכוזם הופך גבוה, הראש נופל. במקומו צומח תא חדש שצובר גם מלחים. במהלך צמיחת העלה, הראש עלול ליפול ולהיווצר מספר פעמים. היתרון של שערות מלח הוא שעל ידי שחרור מלחים הן מאבדות מעט מים. תופעה זו חשובה מאוד לצמחים הגדלים על קרקעות מלוחות, בהן מים מתוקים מועטים.
רקמות הפרשה פנימית.מבני הפרשה פנימיים, המפוזרים בכל גוף הצמח בצורה של אידיובלסטים, אינם מסירים חומרים מחוץ לגוף, צוברים אותם בעצמם. אידיובלסטים הם תאים צמחיים השונים בצורתם, במבנה או בתוכן מתאי אחרים באותה רקמה. בהתאם למקור, מבחינים בכלי קיבול סכיזוגניים (מיוונית schiso - פיצול) וליזוגניים (ליזיס יווני - פירוק) של חומרים מופרשים (איור 23).
כלי קיבול סכיזוגניים. אלו חללים בין-תאיים נרחבים יותר או פחות מלאים בחומרים המופרשים. לדוגמה, מעברי שרף של עצי מחט, ארליה, מטרייה ואחרים. הוא האמין כי לשרף יש תכונות קוטל חיידקים, דוחה אוכלי עשב, מה שהופך צמחים בלתי אכילים עבורם.
כלי קיבול ליסיגניים נוצרים במקום תאים חיים, שמתים ונהרסים לאחר הצטברות חומרים בהם. ניתן לראות תצורות כאלה בקליפה
מבנה תא הצמח
תא הצמח מורכב מקשיחים יותר או פחות דופן תאו פרוטופלסט.דופן התא היא דופן התא והממברנה הציטופלזמית. המונח פרוטופלסט בא מהמילה פּרוֹטוֹפּלָסמָה,אשר שימש זמן רב להתייחס לכל היצורים החיים. הפרוטופלסט הוא הפרוטופלסמה של תא בודד.
הפרוטופלסט מורכב מ ציטופלזמהו גרעינים.הציטופלזמה מכילה אברונים (ריבוזומים, מיקרוטובולים, פלסטידים, מיטוכונדריה) ומערכות ממברנות (רטיקולום אנדופלזמי, דיקטיוזומים). הציטופלזמה כוללת גם את המטריצה הציטופלזמית ( חומר בסיס) שבהם טבולים אברונים ומערכות ממברנות. הציטופלזמה מופרדת מדופן התא קרום פלזמה, שהוא קרום אלמנטרי. שלא כמו רוב תאי בעלי החיים, תאי צמחים מכילים אחד או יותר ואקוולים. אלו שלפוחיות מלאות נוזל המוקפות בממברנה יסודית ( טונופלסט).
בתא צמחי חי, החומר הקרקע נמצא בתנועה מתמדת. בתנועה הנקראת זרם ציטופלזמי או ציקלוזיס,אברונים מעורבים. ציקלוזיס מקל על תנועת החומרים בתא והחלפתם בין התא לסביבה.
קרום פלזמה.זהו מבנה פוספוליפיד דו-שכבתי. תאי צמחים מאופיינים בפלישות של קרום הפלזמה.
קרום הפלזמה מבצע את הפונקציות הבאות:
משתתף בחילופי חומרים בין התא לסביבה;
מתאם את הסינתזה וההרכבה של מיקרו-סיבי תאית של דופן התא;
מעביר אותות הורמונליים וחיצוניים השולטים בצמיחת תאים והתמיינות.
הליבה.זהו המבנה הבולט ביותר בציטופלזמה של תא איקריוטי. הקרנל מבצע שתי פונקציות חשובות:
שולט בפעילות החיונית של התא, וקובע אילו חלבונים, ובאיזו שעה יש לסנתז;
מאחסן מידע גנטי ומעביר אותו לתאי בת במהלך חלוקת התא.
הגרעין של תא אוקריוטי מוקף בשתי ממברנות יסודיות שנוצרות מעטפת גרעין.הוא חדור לנקבוביות רבות בקוטר של 30 עד 100 ננומטר, הנראות רק במיקרוסקופ אלקטרונים. לנקבוביות מבנה מורכב. הממברנה החיצונית של הממברנה הגרעינית במקומות מסוימים משולבת עם הרשת האנדופלזמית. ניתן לראות את המעטפת הגרעינית כחלק מיוחד, מובחן מקומי, של הרשת האנדופלזמית (ER).
בליבה הצבועה בצבעים מיוחדים ניתן להבחין בחוטים וגושים דקים כרומטיןו נוקלאופלזמה(החומר העיקרי של הגרעין). הכרומטין מורכב מ-DNA הקשור לחלבונים מיוחדים הנקראים היסטונים. בתהליך חלוקת התא, הכרומטין הופך צפוף יותר ויותר ומתאסף לתוך כרומוזומים. DNA מקודד מידע גנטי.
אורגניזמים שונים במספר הכרומוזומים בתאים סומטיים. לדוגמה, לכרוב יש - 20 כרומוזומים; חמניות - 34; חיטה - 42; אנשים - 46, ואחד ממיני השרך אופיוגלוסום – 1250. לתאי מין (גמטות) יש רק מחצית ממספר הכרומוזומים האופייניים לתאים סומטיים בגוף. מספר הכרומוזומים בגמטות נקרא הפלואיד(יחיד), בתאים סומטיים - דיפלואיד(לְהַכפִּיל). תאים עם יותר משתי קבוצות של כרומוזומים נקראים פוליפלואיד.
תחת מיקרוסקופ אור, ניתן לראות מבנים כדוריים - נוקלאולי. לכל גרעין יש גרעינים אחד או יותר, הנראים בגרעינים שאינם מתחלקים. RNA ריבוזומלי מסונתז בגרעין. בדרך כלל, לגרעין של אורגניזמים דיפלואידים יש שני גרעינים, אחד לכל קבוצה הפלואידית של כרומוזומים. לנוקלאולים אין קרום משלהם. מבחינה ביוכימית, הנוקלאולי מאופיינים בריכוז גבוה של RNA, הקשור כאן לפוספופרוטאינים. גודל הגרעינים תלוי במצב התפקוד של התא. הבחינו כי בתא הצומח במהירות, שבו מתרחשים תהליכים אינטנסיביים של סינתזת חלבון, נוקלאולי מתרבים בגודלם. בנוקלאולים מיוצרים mRNA וריבוזומים, הממלאים תפקיד סינתטי רק בגרעין.
נוקלאופלזמה (קריופלזמה) מיוצגת על ידי נוזל הומוגני שבו מומסים חלבונים שונים, כולל אנזימים.
פלסטידים. Vacuoles, דופן תא תאית ו פלסטידיםהם מרכיבים אופייניים לתאי צמחים. לכל פלסטיד יש מעטפת משלו, המורכבת משני ממברנות יסודיות. בתוך הפלסטיד מבחינים במערכת ממברנות ובדרגה משתנה של חומר הומוגני - סטרומה.פלסטידים בוגרים מסווגים על סמך הפיגמנטים שהם מכילים.
כלורופלסטים,בהם מתרחשת פוטוסינתזה, מכילים כלורופילים וקרוטנואידים. בדרך כלל יש צורה של דיסק בקוטר של 4 - 5 מיקרון. בתא אחד של המזופיל (אמצע העלה) יכולים להיות 40 - 50 כלורופלסטים; ביריעות מ"מ 2 - כ-500,000. בציטופלזמה, כלורופלסטים ממוקמים בדרך כלל במקביל לממברנת התא.
המבנה הפנימי של הכלורופלסט מורכב. הסטרומה מחלחלת במערכת מפותחת של ממברנות בצורת בועות - thylakoids.כל תילקואיד מורכב משני ממברנות. תילקואידים יוצרים מערכת אחת. ככלל, הם נאספים בערימות - דגניםהדומים לעמודים של מטבעות. התילקואידים של גרנאים בודדים מחוברים ביניהם על ידי תילקואידים סטרומה, או תילקואידים בין-גרניים. כלורופיל וקרוטנואידים משובצים בממברנות התילקואיד. כלורופלסטים בצמחים ירוקים ובאצות מכילים לרוב גרגרי עמילן וטיפות שומנים קטנות (שומן). גרגרי עמילן הם מאגרים זמניים של מוצרי פוטוסינתזה. הם יכולים להיעלם מהכלורופלסטים של צמח שהיה בחושך רק 24 שעות, ולהופיע שוב 3–4 שעות לאחר העברת הצמח לאור.
כלורופלסטים הם אברונים חצי אוטונומיים ודומים לחיידקים. לדוגמה, לריבוזומים של חיידקים וכלורופלסטים יש דמיון גבוה למדי. הם קטנים יותר מריבוזומים אוקריוטיים. סינתזת חלבון על הריבוזומים של חיידקים וכלורופלסטים מעוכבת על ידי כלורמפניקול, שאין לו השפעה על תאים אוקריוטיים. בנוסף, גם לחיידקים וגם לכלורופלסטים יש סוג דומה של נוקלואיד, המאורגן בצורה דומה. למרות העובדה שיצירת כלורופלסטים וסינתזה של פיגמנטים הכלולים בהם נשלטת במידה רבה על ידי ה-DNA הכרומוזומלי של התא, בכל זאת, בהיעדר DNA משלו, כלורופלסטים אינם נוצרים.
כלורופלסטים יכולים להיחשב לאברוני התא העיקריים, שכן הם הראשונים בשרשרת הטרנספורמציה של אנרגיית השמש, וכתוצאה מכך האנושות מקבלת מזון ודלק. בכלורופלסטים מתרחשת לא רק פוטוסינתזה. הם גם מעורבים בסינתזה של חומצות אמינו וחומצות שומן, משמשים כמאגר של עתודות זמניות של עמילן.
כרומופלסטים -פלסטידים פיגמנטים. מגוונים בצורתם, אין להם כלורופיל, אלא מסנתזים וצוברים קרוטנואידים, המעניקים צבע צהוב, כתום, אדום לפרחים, עלים ישנים, לפירות ולשורשים. כרומופלסטים יכולים להתפתח מכלורופלסטים, שבמקרה זה מאבדים כלורופיל ומבני קרום פנימיים, צוברים קרוטנואידים. זה מתרחש במהלך הבשלת פירות רבים. כרומופלסטים מושכים חרקים ובעלי חיים אחרים שאיתם הם התפתחו יחד.
לוקופלסטים -פלסטידים לא פיגמנטים. חלקם מסנתזים עמילן ( עמילופלסטים), אחרים מסוגלים ליצור חומרים שונים, כולל שומנים וחלבונים. באור, לויקופלסטים הופכים לכלורופלסטים.
פרופלסטידים -פלסטידים קטנים חסרי צבע או ירוקים חיוורים, הנמצאים בתאים מריסטמטיים (מתחלקים) של שורשים ונצרים. הם המבשרים של פלסטידים אחרים, מובחנים יותר - כלורופלסטים, כרומופלסטים ואמינופלסטים. אם התפתחות פרוטופלסטידים מתעכבת בגלל חוסר אור, אחד או יותר גופים פרולמלרים, שהם הצטברויות חצי גבישיות של ממברנות צינוריות. פלסטידים המכילים גופים פרולמלרים נקראים אטיופלסטים.באור, האטיופלסטים הופכים לכלורופלסטים, בעוד שהקרומים של הגופים הפרולמלרים יוצרים תילקואידים. אטיופלסטים נוצרים בעלים של צמחים הנמצאים בחושך. פרוטופלסטים של עוברי זרעים הופכים תחילה לאטיופלסטים, שמהם מתפתחים כלורופלסטים באור. פלסטידים מאופיינים במעברים קלים יחסית מסוג אחד לאחר. פלסטידים, כמו חיידקים, מתרבים על ידי חלוקה לשניים. בתאים מריסטמטיים, זמן החלוקה של פרוטופלסטידים עולה בקנה אחד עם זמן חלוקת התא. עם זאת, בתאים בוגרים, רוב הפלסטידים נוצרים כתוצאה מחלוקת פלסטידים בוגרים.
מיטוכונדריה.כמו כלורופלסטים, המיטוכונדריה מוקפת בשני ממברנות יסוד. הממברנה הפנימית יוצרת קפלים ובליטות רבות - כריסט,אשר מגדילים באופן משמעותי את פני השטח הפנימיים של המיטוכונדריה. הם קטנים משמעותית מפלסידים, בקוטר של כ-0.5 מיקרומטר, ומשתנים באורך ובצורה.
במיטוכונדריה מתרחש תהליך הנשימה, כתוצאה ממנו מתפרקות מולקולות אורגניות עם שחרור האנרגיה והעברתה למולקולות ATP, מאגר האנרגיה העיקרי של כל התאים האוקריוטיים. רוב תאי הצמח מכילים מאות או אלפי מיטוכונדריות. מספרם בתא אחד נקבע לפי הצורך של התא ב-ATP. המיטוכונדריה נמצאות בתנועה מתמדת, נעות מחלק אחד של התא לאחר, מתמזגות זו עם זו ומתחלקות. מיטוכונדריה בדרך כלל מתאספת במקום שבו יש צורך באנרגיה. אם קרום הפלזמה מעבירה חומרים באופן פעיל מתא לתא, אז המיטוכונדריה ממוקמות לאורך פני הממברנה. באצות חד-תאיות תנועתיות, מיטוכונדריה מצטברות בבסיס הדגל, ומספקות את האנרגיה הדרושה לתנועתן.
מיטוכונדריה, כמו פלסטידים, הם אברונים חצי אוטונומיים המכילים את הרכיבים הדרושים לסינתזה של החלבונים שלהם. הממברנה הפנימית מקיפה מטריצה נוזלית המכילה חלבונים, RNA, DNA, ריבוזומים דמויי חיידקים ומומסים שונים. DNA קיים בצורה של מולקולות מעגליות הממוקמות בנוקלואיד אחד או יותר.
בהתבסס על הדמיון של חיידקים למיטוכונדריה ולכלורופלסטים של תאים אוקריוטיים, ניתן להניח שהמיטוכונדריה והכלורופלסטים מקורם בחיידקים שמצאו "מפלט" בתאים הטרוטרופיים גדולים יותר - מבשרי האוקריוטים.
מיקרוגופים.שלא כמו פלסטידים ומיטוכונדריה, שמתוחמים על ידי שני ממברנות, מיקרוגוףהם אברונים כדוריים המוקפים בקרום אחד. למיקרוגופים יש תוכן גרגירי (גרגירי), לפעמים הם מכילים גם תכלילי חלבון גבישי. מיקרוגופים קשורים לאזור אחד או שניים של הרשת האנדופלזמית.
כמה מיקרוגופים קראו פרוקסיזומים,ממלאים תפקיד חשוב בחילוף החומרים של חומצה גליקולית, הקשורה ישירות ל-photorespiration. בעלים ירוקים הם קשורים למיטוכונדריה ולכלורופלסטים. מיקרוגופים אחרים, הנקראים גליאוקסיזומים,מכילים את האנזימים הדרושים להמרת שומנים לפחמימות. זה קורה בהרבה זרעים במהלך הנביטה.
ואקוולים -אלו הם אזורים בתא התחום על ידי הממברנה, מלאים בנוזל - מוהל תאים.הם מוקפים טונופלסט(ממברנה ואקוולרית).
תא צמחי צעיר מכיל מספר רב של ואקואולים קטנים, המתמזגים לכדי ואקואול אחד גדול ככל שהתא מתבגר. בתא בוגר, עד 90% מנפחו יכול להיות תפוס על ידי וואקואול. במקרה זה, הציטופלזמה נלחצת בצורה של שכבה היקפית דקה לממברנת התא. הגידול בגודל התא נובע בעיקר מצמיחת ה-vacuole. כתוצאה מכך, מתעורר לחץ טורגור וגמישות הרקמה נשמרת. זהו אחד התפקידים העיקריים של הוואקוול והטונופלסט.
המרכיב העיקרי של המיץ הוא מים, השאר משתנים בהתאם לסוג הצמח ומצבו הפיזיולוגי. Vacuoles מכילים מלחים, סוכרים, לעתים רחוקות יותר חלבונים. הטונופלסט ממלא תפקיד פעיל בהובלה והצטברות של יונים מסוימים ב-vacuole. ריכוז היונים במוהל התא יכול לעלות משמעותית על ריכוזו בסביבה. עם תכולה גבוהה של חומרים מסוימים נוצרים גבישים ב-vacuoles. הנפוצים ביותר הם גבישי סידן אוקסלט, בעלי צורה שונה.
Vacuoles הם מקומות הצטברות של מוצרים מטבוליים (מטבוליזם). אלו יכולים להיות חלבונים, חומצות ואפילו חומרים רעילים לבני אדם (אלקלואידים). לעתים קרובות מופקדים פיגמנטים. כחול, סגול, סגול, אדום כהה, ארגמן נותנים לתאי הצמח פיגמנטים מקבוצת האנתוציאנין. בניגוד לפיגמנטים אחרים, הם מתמוססים היטב במים ונמצאים במוהל תאים. הם קובעים את הצבע האדום והכחול של ירקות רבים (צנוניות, לפת, כרוב), פירות (ענבים, שזיפים, דובדבנים), פרחים (פרחים, גרניום, דלפיניום, ורדים, אדמוניות). לפעמים פיגמנטים אלו מסווים את הכלורופיל בעלים, למשל במייפל האדום הדקורטיבי. אנתוציאנינים צבע עלי הסתיו אדום בוהק. הם נוצרים במזג אוויר שמש קר, כאשר סינתזה של כלורופיל נעצרת בעלים. בעלים, כאשר לא נוצרים אנתוציאנינים, לאחר הרס הכלורופיל, הופכים קרוטנואידים צהובים-כתומים של כלורופלסטים בולטים. העלים בעלי צבע עז ביותר בסתיו קר וצלול.
ואקוולים מעורבים בהרס של מקרומולקולות, במחזור המרכיבים שלהן בתא. ריבוזומים, מיטוכונדריה, פלסטידים, נופלים ל-vacuoles, נהרסים. על ידי פעילות עיכול זו ניתן להשוות איתן ליזוזומים- אברונים של תאי בעלי חיים.
ואקוולים נוצרים מהרשת האנדופלזמית (רטיקולום)
ריבוזומים.חלקיקים קטנים (17 - 23 ננומטר), המורכבים מכמויות שוות בערך של חלבון ו-RNA. בריבוזומים, חומצות אמינו מתחברות ליצירת חלבונים. יש יותר מהם בתאים עם חילוף חומרים פעיל. ריבוזומים ממוקמים בציטופלזמה של התא באופן חופשי או מחוברים לרשת האנדופלזמית (80S). הם נמצאים גם בגרעין (80S), במיטוכונדריה (70S), בפלסטידים (70S).
ריבוזומים יכולים ליצור קומפלקס שעליו יש סינתזה סימולטנית של פוליפפטידים זהים, שמידע לגביו נלקח ממולקולה אחת ומ-RNA. תסביך כזה נקרא פוליריבוזומים (פוליזומים).לתאים המסנתזים חלבונים בכמויות גדולות יש מערכת נרחבת של פוליזומים, המחוברים לעתים קרובות למשטח החיצוני של המעטפת הגרעינית.
רשת אנדופלזמית.זוהי מערכת ממברנות תלת מימדית מורכבת בהיקף בלתי מוגבל. בהקשר של ER נראה כמו שני ממברנות יסודיות עם רווח שקוף צר ביניהם. צורתו ואורכו של ER תלויים בסוג התא, בפעילותו המטבולית ובשלב ההתמיינות. בתאים המפרישים או מאחסנים חלבונים, ה-ER הוא בצורה של שקים שטוחים או בורות מים,עם ריבוזומים רבים המחוברים אל פני השטח החיצוניים שלו. רשת זו נקראת רטיקולום אנדופלזמי מחוספס. ER חלק הוא בדרך כלל בצורת צינורית. רטיקולום אנדופלזמי מחוספס וחלק יכול להיות קיים באותו תא. ככלל, ישנם קשרים רבים ביניהם.
הרשת האנדופלזמית מתפקדת כמערכת התקשורת של התא. זה קשור לקליפה החיצונית של הגרעין. למעשה, שני המבנים הללו יוצרים מערכת ממברנה אחת. כאשר המעטפת הגרעינית נקרעת במהלך חלוקת התא, השברים שלה דומים לשברי ER. הרשת האנדופלזמית היא מערכת להעברת חומרים: חלבונים, שומנים, פחמימות, לחלקים שונים של התא. רטיקולום אנדופלזמי של תאים שכנים מחובר דרך גדילים ציטופלזמיים - plasmodesmata -שעוברים דרך קירות התא.
הרשת האנדופלזמית היא האתר העיקרי לסינתזה של ממברנות התא. בחלק מתאי הצמח, ממברנות של ואקוולים ומיקרוגופים, בורות מים דיקטיוזומים.
מערכת גולג'י.מונח זה משמש להתייחס לכל דיקטיוזום, או גוף גולג'י, בכלוב. דיקטיוזומים הם קבוצות של שלפוחיות שטוחות בצורת דיסק, או בורות מים, המסתעפים לאורך הקצוות למערכת מורכבת של צינוריות. דיקטיוזומים בצמחים גבוהים מורכבים מ-4 - 8 בורות, שנאספו יחד.
בדרך כלל, בחפיסת טנקים, מבחינים בצד מתפתח ומתבגר. הממברנות של הבורות היוצרים דומים במבנה לממברנות ה-ER, והממברנות של הבורות המתבגרים דומים לממברנת הפלזמה.
דיקטיוזומים מעורבים בהפרשה, וברוב הצמחים הגבוהים, ביצירת קרומי התא. פוליסכרידים של דופן התא המסונתזים על ידי דיקטיוזומים מצטברים בשלפוחיות, הנפרדות לאחר מכן מבורות מתבגרים. שלפוחיות הפרשה אלו נודדות ומתמזגות עם ה-CMP; בעוד שהפוליסכרידים הכלולים בהם מובנים בתוך קרום התא. חלק מהחומרים המצטברים בדיקטיוזומים נוצרים במבנים אחרים, למשל ב-ER, ואז מועברים לדיקטיוזומים, שם הם משתנים (משונים) לפני ההפרשה. לדוגמה, גליקופרוטאין הם חומר בנייה חשוב של קרום התא. חלק החלבון מסונתז על ידי פוליזומים של ה-ER המחוספס, חלק הפחמימות מסונתז בדיקטיוזומים, כאשר שני החלקים מתאחדים ליצירת גליקופרוטאין.
ממברנות הן מבנים דינמיים וניידים המשנים כל הזמן את צורתם ושטחם. הרעיון מבוסס על ניידות ממברנה מערכת אנדופלזמית.על פי תפיסה זו, הממברנות הפנימיות של הציטופלזמה, למעט ממברנות המיטוכונדריה והפלסטידים, הן שלם יחיד ומקורן ברטיקולום האנדופלזמי. בורות מים חדשים של דיקטיוזומים נוצרים מהרשת האנדופלזמית דרך שלב השלפוחית הביניימית, ושלפוחיות ההפרשה הנפרדות מהדיקטיוזומים תורמים בסופו של דבר להיווצרות קרום הפלזמה. לפיכך, הרשת האנדופלזמית והדיקטיוזומים מהווים יחידה תפקודית שבה הדיקטיוזומים ממלאים את התפקיד של מבני ביניים בתהליך המרת ממברנות כמו הרשת האנדופלזמית לממברנות כמו הפלזמה. ברקמות שהתאים שלהן גדלים ומתחלקים בצורה חלשה, מרכיבי הממברנה מתעדכנים כל הזמן.
מיקרוטובוליםנמצא כמעט בכל התאים האיקריוטיים. הם מבנים גליליים בקוטר של כ-24 ננומטר. אורכם משתנה. כל צינורית מורכבת מתת-יחידות של חלבון הנקרא טובולין.יחידות המשנה יוצרות 13 חוטים אורכיים המקיפים את החלל המרכזי. מיקרוטובולים הם מבנים דינמיים, הם נהרסים באופן קבוע ונוצרים בשלבים מסוימים של מחזור התא. ההרכבה שלהם מתרחשת במקומות מיוחדים, הנקראים מרכזי ארגון מיקרוטובולי. בתאי צמחים, יש להם מבנה אמורפי בעל ביטוי חלש.
פונקציות של microtubules: להשתתף ביצירת קרום התא; להפנות בועות של דיקטיוזומים לממברנה היוצרות, כמו חוטי ציר שנוצרים בתא מתחלק; ממלאים תפקיד ביצירת לוחית התא (הגבול הראשוני בין תאי הבת). בנוסף, microtubules הם מרכיב חשוב של flagella ו cilia, בתנועתם הם ממלאים תפקיד חשוב.
מיקרופילמנטים,כמו מיקרוטובולים, הם נמצאים כמעט בכל התאים האיקריוטיים. הם חוטים ארוכים בעובי 5-7 ננומטר, המורכבים מהחלבון המתכווץ אקטין. צרורות של מיקרופילמנטים נמצאים בתאים רבים של צמחים גבוהים יותר. ככל הנראה, הם ממלאים תפקיד חשוב בזרמי הציטופלזמה. מיקרופילמנטים, יחד עם מיקרוטובולים, יוצרים רשת גמישה הנקראת ציטושלד.
חומר בסיסבמשך זמן רב נחשב לתמיסה הומוגנית (הומוגנית) עשירה בחלבון עם מספר קטן של מבנים או אפילו חסרת מבנה. אולם, נכון להיום, באמצעות מיקרוסקופ אלקטרונים במתח גבוה, נקבע כי החומר הקרקע הוא סריג תלת מימדי הבנוי מחוטים דקים (קוטר 3-6 ננומטר) הממלאים את התא כולו. מרכיבים אחרים של הציטופלזמה, כולל מיקרוטובולים ומיקרופילמנטים, מושעים מזה רשת מיקרוטרבקולרית.
המבנה המיקרוטרבקולרי הוא סריג של גדילי חלבון, שהחלל ביניהם מלא במים. יחד עם מים, לסריג יש עקביות של ג'ל; הג'ל נראה כמו גופים ג'לטינים.
אברונים מחוברים לסריג המיקרוטרבקולרי. הסריג מספק תקשורת בין חלקים בודדים של התא ומכוון את ההובלה התוך תאית.
טיפות שומנים- מבנים בעלי צורה כדורית, המעניקים גרעיניות לציטופלזמה של תא צמחי תחת מיקרוסקופ אור. במיקרוגרפים של אלקטרונים, הם נראים אמורפיים. טיפות דומות מאוד אך קטנות יותר נמצאות בפלסידים.
טיפות ליפידים, בטעות שהן אברונים, קראו להן ספרוזומים והאמינו שהן מוקפות בקרום שכבה בודדת או כפולה. עם זאת, נתונים עדכניים מראים שלטיפות שומנים אין ממברנות, אך עשויות להיות מצופות בחלבון.
חומרים ארגסטיים -אלו הם "המוצרים הפסיביים" של הפרוטופלסט: חומרי רזרבה או פסולת. הם יכולים להופיע ולהיעלם בתקופות שונות של מחזור התא. מלבד גרגרי עמילן, גבישים, פיגמנטים אנתוציאנין וטיפות שומנים. אלה כוללים שרפים, מסטיקים, טאנינים וחלבונים. חומרים ארגאסטיים הם חלק מממברנת התא, החומר העיקרי של הציטופלזמה והאברונים, כולל ואקוולים.
פלאגלה וריסים -הם מבנים דקים דמויי שיער המשתרעים מפני השטח של תאים איקריוטיים רבים. יש להם קוטר קבוע, אבל האורך משתנה בין 2 ל 150 מיקרון. באופן קונבנציונלי, הארוכים והמעטים מהם נקראים דגלים, והקצרים והרבים יותר נקראים ריסים. אין הבדלים ברורים בין שני סוגי המבנים הללו, ולכן המונח משמש להתייחס לשניהם. flagellum.
בחלק מהאצות והפטריות, הדגלים הם איברי תנועה, בעזרתם הם נעים במים. בצמחים (לדוגמה, טחבים, עצי כבד, שרכים, כמה ג'מנוספרמים), רק לתאי נבט (גמטות) יש דגלים.
לכל דגל יש ארגון ספציפי. טבעת חיצונית של 9 זוגות של מיקרו-צינוריות מקיפה שתי מיקרו-צינוריות נוספות הממוקמות במרכז הדגל. "ידיות" המכילות אנזים משתרעות ממיקרו-צינורית אחת של כל אחד מהזוגות החיצוניים. זהו דפוס הארגון הבסיסי של 9+2 שנמצא בכל הדגלים באורגניזמים איקריוטיים. מאמינים כי תנועת הפלגלה מבוססת על החלקה של מיקרוטובולים, בעוד שהזוגות החיצוניים של המיקרוטובולים נעים אחד לאורך השני ללא התכווצות. החלקה של זוגות מיקרוטובולים זה לזה גורמת לכיפוף מקומי של הדגל.
Flagella "צומח" ממבנים גליליים ציטופלזמיים הנקראים גופים בסיסיים,נוצר והחלק הבסיסי של הדגל. לגופים הבסיסיים יש מבנה פנימי הדומה לזה של דגל, אלא שהצינוריות החיצוניות מסודרות בשלישיות ולא בזוגות, והצינוריות המרכזיות נעדרות.
דופן תא.דופן התא תוחמת את גודל הפרוטופלסט ומונעת את קריעתו עקב ספיגת המים על ידי הוואקואול.
לדופן התא יש תפקידים ספציפיים שחשובים לא רק לתא ולרקמה שבה נמצא התא, אלא גם לצמח כולו. לדפנות התא תפקיד חיוני בספיגה, הובלה והפרשה של חומרים, ובנוסף, ניתן לרכז בהם פעילות ליזוזומלית, או עיכול.
רכיבי דופן התא.המרכיב האופייני ביותר של דופן התא הוא תָאִית,מה שקובע במידה רבה את הארכיטקטורה שלו. מולקולות תאית מורכבות ממולקולות גלוקוז חוזרות המחוברות מקצה לקצה. מולקולות תאית דקות ארוכות משולבות לתוך מיקרופיבריליםעובי 10 - 25 ננומטר. מיקרופיברילים משתלבים זה בזה ויוצרים חוטים דקים, שבתורם ניתן ללפף אחד סביב השני, כמו חוטים בחבל. לכל "חבל", או מקרופיבריל כזה, עובי של כ-0.5 מיקרון, מגיע לאורך של 4 מיקרון. מקרופיברילים חזקים כמו חוטי פלדה בגודל שווה.
מסגרת התאית של דופן התא מלאה במולקולות מטריצת תאית השזורות בה. הוא מכיל פוליסכרידים הנקראים hemicelluloses,ו פקטין,אוֹ פקטין,כימית קרוב מאוד להמיצלולוז.
מרכיב נוסף של דופן התא הוא ליגנין -הוא הפולימר השכיח ביותר בתאי הצמח לאחר תאית. לגנין מגביר את קשיחות הדופן ונמצא בדרך כלל בתאים המבצעים פונקציה תומכת או מכנית.
קוטין, סוברין, שעווה- מופקדים בדרך כלל בקונכיות של רקמות הגנה של צמחים. קוטין, למשל, נמצא בקרומי התא של האפידרמיס, וסוברין נמצא ברקמת ההגנה המשנית, הפקק. שני החומרים מצויים בשילוב עם שעווה ומונעים איבוד יתר של מים מהצמח.
שכבות של דופן התא.עובי הדופן של תאי הצמח משתנה מאוד, בהתאם לתפקידם של התאים במבנה הצמח וגיל התא עצמו. תחת מיקרוסקופ אלקטרונים, שתי שכבות נראות בדופן תא הצמח: צלחת אמצעית(נקרא גם חומר בין תאי), ו דופן תא ראשונית.תאים רבים מניחים שכבה נוספת - דופן תא משני.הצלחת החציונית ממוקמת בין הקירות הראשוניים של תאים סמוכים. הקיר המשני, אם בכלל, מופקד על ידי פרוטופלסט התא על פני השטח הפנימיים של דופן התא הראשוני.
צלחת אמצעית.הלמינה החציונית מורכבת בעיקר מפקטין. במקום שבו צריך להיווצר דופן תא, בין שני תאים שזה עתה נוצרו, מציינים לראשונה מקלעת צפופה של צינוריות של הרשת האנדופלזמית והבור של מנגנון הגולגי (דיקטיוזומים). ואז מופיעות בועות במקום הזה, מלאות בפקטין (מפוליסכרידים). הבועות הללו מופרדות מהמיכלים של מנגנון גולגי. דופן התא המוקדם מכיל פוליסכרידים שונים, כאשר העיקריים שבהם הם פקטין והמיצלולוז. מאוחר יותר, הוא מורכב מחומרים צפופים יותר - תאית וליגנין.
דופן תא ראשונית.זוהי שכבה של מעטפת תאית המונחת לפני או במהלך צמיחת התא. בנוסף לתאית, המיצלולוזה ופקטין, הממברנות הראשוניות מכילות גליקופרוטאין. קונכיות ראשוניות יכולות להתגרות. רכיב הפקטין מקנה פלסטיות, המאפשרת לקליפה הראשונית להתרחב ככל שהשורש, הגבעול או העלה מתארכים.
לתאים המתחלקים באופן פעיל (רוב התאים הבוגרים המעורבים בתהליכי הפוטוסינתזה, הנשימה וההפרשה) יש ממברנות ראשוניות. תאים כאלה עם ממברנה ראשונית ופרוטופלסט חי מסוגלים לאבד את צורתם האופיינית, להתחלק ולהתמיין לסוג תא חדש. הם מעורבים בריפוי פצעים והתחדשות רקמות בצמחים.
קרומי תאים ראשוניים אינם אחידים בעובי לכל אורכם, אך יש להם חלקים דקים הנקראים שדות נקבוביות ראשוניים.גדילי הציטופלזמה, או plasmodesmata, המחברים את הפרוטופלסטים של תאים שכנים בדרך כלל עוברים דרך שדות הנקבוביות הראשוניים.
דופן תא משני.למרות העובדה שלתאים צמחיים רבים יש רק דופן ראשונית, בחלקם, הפרוטופלסט מפקיד דופן משנית לכיוון מרכז התא. זה מתרחש בדרך כלל לאחר הפסקת צמיחת התאים ושטח הממברנה הראשונית כבר לא גדל. מסיבה זו, הקליפה המשנית שונה מהראשונית. ממברנות משניות נחוצות במיוחד על ידי תאים מיוחדים המחזקים את הצמח ומוליכים מים. לאחר שקיעת הממברנה המשנית, הפרוטופלסט של תאים אלה, ככלל, מת. יש יותר תאית בקליפות המשניות מאשר בראשוניות, ונעדרים בהן חומרי פקטין וגליקופרוטאין. קשה למתוח את הממברנה המשנית, המטריצה שלו מורכבת מהמיצלולוזה.
במעטפת המשנית ניתן להבחין בשלוש שכבות - חיצוניות, אמצעיות ופנימיות (S 1, S 2, S 3). מבנה השכבות של הקונכיות המשניות מגדיל משמעותית את חוזקן. מיקרו-סיבי תאית בממברנה המשנית מופקדים צפופים יותר מאשר בראשי. לגנין הוא מרכיב נפוץ בעורות המשניים של העץ.
הנקבוביות בממברנות של תאים מגע ממוקמות זו מול זו. נוצרות שתי נקבוביות מנוגדות וקרום נקבוביות מספר פעמים.בתאים עם ממברנות משניות, ישנם שני סוגים עיקריים של נקבוביות: פשוט ותוחם.בנקבוביות תוחמות, הממברנה המשנית תלויה מעל חלל נקבוביות.בנקבוביות פשוטות, זה לא המקרה.
צמיחת דופן התא.ככל שהתא גדל, העובי והשטח של קרום התא גדלים. מתיחת מעטפת היא תהליך מורכב. הוא נשלט על ידי הפרוטופלסט ומווסת על ידי הורמון אוקסין.
בתאים הגדלים באופן אחיד לכל הכיוונים, שקיעת המיופיברילים היא אקראית. מיופיברילים אלה יוצרים רשת לא סדירה. תאים כאלה נמצאים בליבת הגזע, ברקמות האחסון, ובמהלך גידול תאים במבחנה. בתאים מתארכים, המיופיברילים של המעטפת הצדדית מופקדים בזווית ישרה לציר ההתארכות.
חומרים מטריקס - פקטין, hemicelluloses וגליקופרוטאינים מועברים לממברנה בשלפוחיות של dictyosomes. יחד עם זאת, פקטין אופייניים יותר לתאים גדלים, בעוד שהמיצלולוזות שולטות בתאים שאינם גדלים.
מיקרו-סיבי תאית מסונתזים על פני התא בעזרת קומפלקס אנזים הקשור לממברנת הפלזמה. הכיוון של מיקרופיברילים נשלט על ידי מיקרוטובולים הממוקמים ליד המשטח הפנימי של קרום הפלזמה.
פלסמודזמה.אלו הם חוטים דקים של הציטופלזמה המחברים את הפרוטופלסטים של תאים שכנים. Plasmodesmata עוברים דרך דופן התא בכל מקום, או מרוכזים בשדות נקבוביות ראשוניים או בממברנות בין זוגות של נקבוביות. תחת מיקרוסקופ אלקטרונים, פלסמודסמטה מופיעות כתעלות צרות המרופדות בקרום הפלזמה. צינור גלילי קטן יותר נמתח לאורך ציר התעלה מתא אחד למשנהו - desmotube,אשר מתקשר עם הרשת האנדופלזמית של שני התאים הסמוכים. פלסמודסמטה רבות נוצרות במהלך חלוקת התא, כאשר הרטיקולום האנדופלזמי הצינורי נקלט על ידי לוחית התא המתפתחת. Plasmodesmata יכול להיווצר גם בממברנות של תאים שאינם מתחלקים. מבנים אלה מספקים העברה יעילה של חומרים מסוימים מתא לתא.
חלוקת תא.באורגניזמים רב-תאיים, חלוקת תאים, יחד עם עלייה בגודלם, היא דרך לצמיחה של האורגניזם כולו. תאים חדשים שנוצרים במהלך החלוקה דומים במבנה ובתפקוד, הן לתא האב והן זה לזה. ניתן לחלק את תהליך החלוקה באוקריוטים לשני שלבים חופפים חלקית: מיטוזהו ציטוקינזיס.
מיטוזה- זוהי היווצרות של שני גרעיני בת מגרעין אחד, שוות ערך זה לזה מבחינה מורפולוגית וגנטית. ציטוקינזיס -זוהי חלוקת החלק הציטופלזמי של התא עם היווצרות תאי בת.
מחזור התא.תא חי עובר סדרה של אירועים עוקבים המרכיבים את מחזור התא. משך המחזור עצמו משתנה בהתאם לסוג התא ולגורמים חיצוניים כמו טמפרטורה או זמינות חומרי הזנה. בדרך כלל המחזור מחולק ל interphaseוארבעה שלבים מיטוזה.
Interphase.התקופה שבין חלוקות מיטוטיות עוקבות.
האינטרפאזה מחולקת לשלוש תקופות, המסומנות כ-G 1, S, G 2.
במהלך תקופת G 1, שמתחילה לאחר מיטוזה. במהלך תקופה זו, כמות הציטופלזמה עולה, כולל אברונים שונים. בנוסף, על פי ההשערה המודרנית, במהלך תקופת G 1, מסונתזים חומרים שמעוררים או מעכבים את תקופת S ואת שאר המחזור, ובכך קובעים את תהליך החלוקה.
תקופת S עוקבת אחר תקופת G 1, שבה החומר הגנטי (DNA) משוכפל.
במהלך תקופת G2, העוקבת אחרי S, נוצרים מבנים המעורבים ישירות במיטוזה, למשל, רכיבי ציר.
חלק מהתאים עוברים מספר בלתי מוגבל של מחזורי תאים. אלה הם אורגניזמים חד-תאיים וכמה תאים של אזורי צמיחה פעילים (מריסטמים). כמה תאים מיוחדים לאחר התבגרות מאבדים את יכולתם להתרבות. הקבוצה השלישית של תאים, למשל, אלו היוצרים רקמת פצע (קלוס), שומרת על היכולת להתחלק רק בתנאים מיוחדים.
מיטוזה,או ביקוע גרעיני. זהו תהליך מתמשך המחולק לארבעה שלבים: פרופאזה, מטאפאזה, אנפאזה, טלופזה.כתוצאה מהמיטוזה, החומר הגנטי שהכפיל את עצמו ב-interphase מחולק שווה בשווה בין שני גרעיני הבת.
אחד הסימנים המוקדמים ביותר למעבר של תא לחלוקה הוא הופעת פס צר וטבעתי של מיקרו-צינוריות ממש מתחת לממברנת הפלזמה. חגורה צפופה יחסית זו מקיפה את הגרעין במישור המשווני של הציר המיטוטי העתידי. מכיוון שהוא מופיע לפני הנבואה, הוא נקרא חגורת preprophase.הוא נעלם לאחר הציר המיטוטי, הרבה לפני הופעתו בטלופזה המאוחר של לוחית התא, הגדל מהמרכז לפריפריה ומתמזג עם קרום תא האם באזור שנכבש בעבר על ידי חגורת הקדם-פרופאזה.
פרופאזה.בתחילת הפרופזה, הכרומוזומים דומים לחוטים ארוכים המפוזרים בתוך הגרעין. לאחר מכן, כאשר הגדילים מתקצרים ומתעבים, ניתן לראות שכל כרומוזום מורכב לא מאחד, אלא משני גדילים שלובים זה בזה, הנקראים כרומטידות.בנבואה מאוחרת, שתי הכרומטידות המקוצרות של כל כרומוזום שוכנות זו לצד זו במקביל, מחוברות בקטע צר הנקרא צנטרומר.יש לו מיקום ספציפי על כל כרומוזום והוא מחלק את הכרומוזום לשתי זרועות באורכים שונים.
מיקרו-צינוריות ממוקמות במקביל לפני השטח של הגרעין לאורך ציר הציר. זהו הביטוי המוקדם ביותר של הרכבת ציר מיטוטי.
בסוף הפרופזה, הגרעין מאבד בהדרגה את קווי המתאר הברורים שלו ולבסוף נעלם. זמן קצר לאחר מכן מתפרקת גם המעטפת הגרעינית.
מטאפאזה.בתחילה ציר מטאפאזה,המייצג מבנה תלת מימדי, הרחב ביותר באמצע ומצטמצם לכיוון הקטבים, תופס את המקום שבו תפוס הגרעין בעבר. סיבי הציר הם צרורות של microtubules. במהלך המטאפאזה, הכרומוזומים, שכל אחד מהם מורכב משתי כרומטידות, מסודרים כך שהצנטרומרים שלהם נמצאים במישור המשווני של הציר. עם הצנטרומר שלו, כל כרומוזום מחובר לחוטי הציר. עם זאת, כמה גדילים עוברים מקוטב אחד למשנהו מבלי להיות מחוברים לכרומוזומים.
כאשר כל הכרומוזומים ממוקמים במישור המשווני, המטאפאזה הושלמה. כרומוזומים מוכנים להתחלק.
אנפאזה.הכרומטידות של כל כרומוזום נפרדות. עכשיו זה יֶלֶדכרומוזומים. קודם כל, הצנטרומר מתחלק, ושני כרומוזומי הבת נמשכים לקטבים מנוגדים. במקרה זה, הצנטרומרים נעים קדימה, וזרועות הכרומוזומים נמתחות מאחור. חוטי הציר המחוברים לכרומוזומים מתקצרים, ותורמים להפרדת הכרומטידים ולתנועת כרומוזומי הבת בכיוונים מנוגדים.
טלופאז.בטלופאזה, ההפרדה של שתי קבוצות זהות של כרומוזומים הושלמה, בעוד שסביב כל אחת מהן נוצר קרום גרעיני. הרשת המחוספסת לוקחת חלק פעיל בכך. מנגנון הציר נעלם. במהלך הטלופאז, הכרומוזומים מאבדים את בהירות המתאר שלהם, נמתחים, הופכים בחזרה לחוטים דקים. הנוקלאולים משוחזרים. כאשר הכרומוזומים הופכים בלתי נראים, המיטוזה הושלמה. שני גרעיני הבת נכנסים לשלבים. הם שקולים מבחינה גנטית זה לזה ולגרעין האב. זה חשוב מאוד, שכן יש להעביר את התוכנית הגנטית, ואיתה כל הסימנים, לאורגניזמים הבתים.
משך המיטוזה משתנה באורגניזמים שונים ותלוי בסוג הרקמה. עם זאת, הפרופזה היא הארוכה ביותר והאנפאזה היא הקצרה ביותר. בתאי קצה השורש, משך הפרופזה הוא 1-2 שעות; מטאפאזה - 5 - 15 דקות; אנפאזה - 2 - 10 דקות; טלופאז - 10 - 30 דקות. משך האינטרפאז הוא בין 12 ל-30 שעות.
בתאים איקריוטים רבים, מרכזי ארגון המיקרוטובוליות האחראים על היווצרות הציר המיטוטי קשורים צנטריולים.
ציטוקינזיס.זהו תהליך החלוקה של הציטופלזמה. ברוב האורגניזמים, התאים מתחלקים ע"י נסיגת קרום התא ויצירת תלם ביקוע, אשר מעמיק בהדרגה, ודוחס את שאר החוטים של הציר המיטוטי. בכל הצמחים (בריופיטים וכלי דם) ובחלק מהאצות, התאים מתחלקים עקב היווצרותם. לוחית התא.
בטלופאז מוקדם נוצרת מערכת סיבים בצורת חבית בין שני גרעיני הבת, הנקראים פרגמופלסט.סיבי הפרגמופלסט, כמו סיבי הציר המיטוטי, מורכבים ממיקרוטובולים. טיפות קטנות מופיעות במישור המשווני של הפרגמופלסט. הם מתמזגים ויוצרים לוחית תא שגדלה עד שהיא מגיעה לדופן התא המתחלק. זה משלים את ההפרדה של שני תאי הבת. טיפות מתמזגות הן בועות המתנתקות ממנגנון גולגי. בעיקרון, הם מכילים חומרי פקטין, שמהם נוצרת הצלחת האמצעית. ממברנות שלפוחית מעורבות בבניית קרום הפלזמה משני צידי הצלחת. במקביל, plasmodesmata נוצרים משברים של הרשת האנדופלזמית הצינורית.
לאחר היווצרות הצלחת החציונית, כל פרוטופלסט מפקיד עליו קרום ראשוני. בנוסף, כל תא בת מפקיד שכבה חדשה של ממברנה סביב הפרוטופלסט כולו, שממשיכה את הממברנה שעלתה מלוחית התא. המעטפת המקורית של תא האב נהרסת ככל שתאי הבת גדלים.
סוגים שונים של מיטוזה איקריוטית
חלוקת התא שתוארה לעיל בצמחים, וגם בבעלי חיים, אינה הצורה היחידה של חלוקת תאים עקיפה. הסוג הפשוט ביותר של מיטוזה הוא pleuromitosis.זה דומה לחלוקה בינארית תאים פרוקריוטיים, שבו הנוקלואידים לאחר שכפול נשארים קשורים לממברנת הפלזמה. הממברנה מתחילה לצמוח בין נקודות הקישור של ה-DNA ובכך מפזרת את הכרומוזומים לחלקים שונים של התא. לאחר מכן, במהלך היווצרות התכווצות התא, כל אחת ממולקולות ה-DNA תהיה בתא נפרד חדש.
מאפיין לחלוקה תאים איקריוטייםהוא היווצרות של ציר הבנוי ממיקרוטובולים. בְּ pleuromitosis סגור(זה נקרא סגור מכיוון שההתבדלות של הכרומוזומים מתרחשת מבלי לשבש את המעטפת הגרעינית) לא צנטריולים, אלא מבנים אחרים הממוקמים בצד הפנימי של הממברנה הגרעינית, משתתפים כמרכזי ארגון מיקרו-צינוריות (MCMT). אלה הם מה שנקרא גופים קוטביים של מורפולוגיה בלתי מוגדרת, שמהם משתרעים מיקרוטובולים. יש שניים מהגופים האלה. הם מתפצלים זה מזה מבלי לאבד קשר עם המעטפת הגרעינית. כתוצאה מכך, נוצרים שני חצאי צירים הקשורים לכרומוזומים. כל התהליך של יצירת ציר מיטוטי והפרדת כרומוזומים במקרה זה מתרחש מתחת למעטפת הגרעינית. סוג זה של מיטוזה מתרחש בקרב פרוטוזואה ומופץ באופן נרחב בפטריות (chytridia, zygomycetes, שמרים, oomycetes, ascomycetes, myxomycetes, וכו'). לפגוש טפסים pleuromitosis חצי סגורהכאשר המעטפת הגרעינית נהרסת בקטבים של הציר שנוצר.
הצורה הבאה של מיטוזה היא אורתומיטוזיס.במקרה זה, ה-COMTs ממוקמים בציטופלזמה; מההתחלה, לא נוצרים חצאי צירים, אלא ציר דו-קוטבי. יש שלוש צורות אורתומיטוזיס (מיטוזה רגילה) חצי סגורו סָגוּר. באורתומיטוזיס סגורה למחצה, נוצר ציר ביסימטרי בעזרת TsOMT הממוקם בציטופלזמה, המעטפת הגרעינית נשמרת לאורך כל המיטוזה, למעט אזורי הקוטב. מסות של חומר גרגירי, או אפילו צנטריולים, עשויות להתגלות כ-COMT. צורה זו של מיטוזה מצויה ב-zoospores של אצות ירוקות, חומות ואדומות, בחלק מהפטריות הנמוכות יותר ובגרגרינים. עם אורתומיטוזיס סגור, הממברנה הגרעינית נשמרת לחלוטין, שבה נוצר ציר אמיתי. מיקרוטובולים נוצרים בקאריופלזמה, לעתים רחוקות יותר הם גדלים מה-TsOMT הפנימי, שאינו קשור (בניגוד לפלורומיטוזיס) עם הממברנה הגרעינית. סוג זה של מיטוזה אופייני לחלוקה של מיקרו-גרעיני ריסים, אך יכול להתרחש גם בפרוטוזואה. באורתומיטוזיס פתוח, המעטפת הגרעינית מתפרקת לחלוטין. סוג זה של חלוקת תאים אופייני לאורגניזמים של בעלי חיים, כמה פרוטוזואה ותאים של צמחים גבוהים יותר. צורה זו של מיטוזה, בתורה, מיוצגת על ידי טיפוסים אסטרליים ואנסטרליים.
מהחומר שנסקר בקצרה, ניתן לראות כי המאפיין העיקרי של מיטוזה באופן כללי הוא הופעת מבני ציר ביקוע, הנוצרים בקשר עם ה-CMT, המגוונים במבנה שלהם.
מורפולוגיה של הדמות המיטוטית
המנגנון המיטוטי בא לידי ביטוי טוב במיוחד בשלב המטאפאזה של המיטוזה. במטאפאזה, במישור המשווני של התא, ממוקמים כרומוזומים, שמהם מה שנקרא חוטי ציר,מתכנס בשני קטבים שונים של הדמות המיטוטית. לפיכך, הציר המיטוטי הוא אוסף של כרומוזומים, קטבים וסיבים. סיבי הציר הם מיקרוטובולים בודדים או צרורותיהם. מיקרוטובולים מתחילים מעמודי הציר, וחלקם הולכים לצנטרומרים היכן ממוקמים kinetochores כרומוזומים (kinetochore microtubules), חלקם עוברים הלאה לכיוון הקוטב הנגדי, אך אינם מגיעים אליו. הם נקראים "מיקרוטובולים בין קוטביים". מהקטבים יוצאת קבוצה של מיקרוטובולים רדיאליים, ויוצרת סביבם מבנה הדומה ל"זוהר קורן" – אלו הן מיקרוטובולים אסטרליים.
לפי מורפולוגיה, דמויות מיטוטיות מחולקות לסוגים אסטרליים ואנסטרליים.
סוג הציר האסטרלי, או מתכנס, מאופיין בכך שהקטבים שלו מיוצגים על ידי אזור קטן שאליו מתכנסות (מתכנסות) המיקרוטובולים. בדרך כלל, צנטרוזומים המכילים צנטרולים ממוקמים בקטבים של הצירים האסטרליים. עם זאת, ידועים מקרים של מיטוזות אסטרליות צנטרולריות (במהלך מיוזה של חלק מחסרי חוליות). בנוסף, מציינים מיקרוטובולים רדיאליים המקרינים מהקטבים, שאינם חלק מהציר, אלא יוצרים אזורי כוכבים - ציסטרים. סוג זה של חלוקה מיטוטית דומה למשקולת. לסוג האנסטריאלי של הדמות המיטוטית אין ציסטרים בקטבים. האזורים הקוטביים של הציר כאן רחבים, הם נקראים כובעים קוטביים, הם כוללים centrioles. במקרה זה, סיבי הציר אינם יוצאים מנקודה אחת, אלא מתפצלים בחזית רחבה (מתפצלים) מכל אזור כובעי הקוטב. סוג זה של ציר מאפיין תאים מתחלקים של צמחים גבוהים יותר, אך ניתן למצוא אותו גם בבעלי חיים גבוהים יותר. בעוברות מוקדמת של יונקים, מיטוזות צנטרולריות (מתפצלות) נצפות במהלך חלוקת הבשלת הביציות ובחלוקות I ו-II של הזיגוטה. אבל כבר בחלוקת התא השלישי ובכל התאים שלאחר מכן, הם מתחלקים בהשתתפות צירים אסטרליים, שבקטבים שלהם נמצאים תמיד צנטרולים. באופן כללי, עבור כל צורות המיטוזה, הכרומוזומים עם הקינטופורים שלהם, הגופים הקוטביים (צנטרוזומים) וסיבי הציר נשארים מבנים נפוצים.
צנטרומרים וקינטוכורים
לצנטרומרים יכולה להיות לוקליזציה שונה לאורך הכרומוזומים. הולוצנטריצנטרומרים מתרחשים כאשר מיקרוטובולים מקושרים לאורך כל הכרומוזום (חלק מהחרקים, נמטודות, חלק מהצמחים). מונוצנטריצנטרומרים - כאשר מיקרוטובולים קשורים לכרומוזומים באזור אחד. צנטרומרים מונוצנטריים יכולים להיות לְאַתֵר בִּמְדוּיָק(לדוגמה, בשמרים ניצנים מסוימים), כאשר רק מיקרו-צינורית אחת מתקרבת לקינטוכור, ו אזורי, שבו צרור של מיקרוטובולים מתקרב לקינטוכור המורכב. למרות מגוון אזורי הצנטרומרים, כולם קשורים למבנה מורכב. kinetochore, שיש לו דמיון מהותי במבנה ובתפקוד בכל האיקריוטים. קינטוכורים הם מבני חלבון מיוחדים, הממוקמים בעיקר באזורי הצנטרומרים של הכרומוזומים. אלו קומפלקסים מורכבים המורכבים מחלבונים רבים. מבחינה מורפולוגית, הם דומים מאוד, בעלי אותו מבנה, החל מדיאטומים לבני אדם. הם מבנים תלת-שכבתיים: השכבה הצפופה הפנימית הצמודה לגוף הכרומוזום, השכבה האמצעית הרפויה והשכבה הצפופה החיצונית. סיבים רבים משתרעים מהשכבה החיצונית, ויוצרים את מה שנקרא הכתר הסיבי של הקינטוכור. בצורה הכללית, קינטוכורים נראים כמו לוחות או דיסקים השוכבים באזור ההתכווצות הראשונית של הכרומוזום, בצנטרומר. בדרך כלל יש קינטוכור אחד לכל כרומוזום או כרומטיד. לפני אנפאזה, קינטוכורים על כל כרומטיד אחות ממוקמים, כל אחד מתקשר עם צרור המיקרוטובולים שלו. בצמחים, הקינטוכור אינו נראה כמו לוחות, אלא כחצי כדור. התפקיד הפונקציונלי של קינטוכורים הוא לקשור כרומטידות אחיות זו לזו, לקבע מיקרוטובולים מיטוטיים, לווסת הפרדת כרומוזומים ולמעשה להזיז כרומוזומים במהלך מיטוזה בהשתתפות מיקרוטובולים. באופן כללי, מבני חלבון, קינטוכורים, מוכפלים בתקופת S, במקביל להכפלת הכרומוזומים. אבל החלבונים שלהם נמצאים על הכרומוזומים בכל תקופות מחזור התא.
רקמת צמחים
רקמות חינוכיות (מריסטמים)
רקמות חינוכיות בגוף הצמחים ממוקמות במקומות שונים, ולכן הן מחולקות לקבוצות הבאות (איור 0;1).
מריסטמות אפיקליות (אפיקליות).ממוקם על החלק העליון, או הקודקודים, של האיברים הציריים - הגבעול, השורש. בעזרת המריסטמים הללו, האיברים הצומחיים של הצמחים מבצעים את צמיחתם לאורך.
מריסטמים לרוחבאופייני לאיברים צירים. שם הם ממוקמים באופן קונצנטרי, בצורה של מצמד.
בין קלוריות,אוֹ intercalary, meristemsמקורם במריסטמים האפיקיים. אלו קבוצות של תאים שעדיין לא מסוגלות להתרבות, אבל יצאו לדרך של התמיינות. אין ביניהם תאים ראשוניים, אבל יש הרבה תאים מיוחדים.
מריסטמים פצעיםלספק שיקום של החלק הפגוע של הגוף. התחדשות מתחילה עם דה-דיפרנציאציה, כלומר, התפתחות הפוכה מתאי מיוחדים לתאים מריסטמטיים. הם הופכים ל felogen,אילו צורות פְּקָקמכסה את פני הפצע. תאים לא מובחנים, מתחלקים, יכולים ליצור רקמה פרנכימית רופפת - יַבֶּלֶת.ממנו, בתנאים מסוימים, נוצרים איברי צמחים.
רקמות אינטגמנטרי
הם פועלים כמחסום גבול, המפריד את הרקמות הבסיסיות מהסביבה. המרכיבים העיקריים של צמח מורכבים רק מתאי חיים. אינטגמנט משני ושלישוני - בעיקר מהמתים עם דפנות תאים עבות.
התפקודים העיקריים של רקמות המוח:
הגנה על הצמח מפני ייבוש;
הגנה מפני חדירת מיקרואורגניזמים מזיקים;
הגנה מפני כוויות שמש;
הגנה מפני נזק מכני;
ויסות חילוף החומרים בין הצמח לסביבה;
תפיסה של גירוי.
רקמת מערכת ראשונית - אפידרמיס, אפידרמיס . מורכב מתאי חיים. נוצר ממריסטמים אפיקליים. מכסה גבעולים ועלים צעירים.
האפידרמיס נוצר בצמחים בקשר ליציאה מבית הגידול המימי ליבשה על מנת למנוע את התייבשותו. בנוסף לסטומטה, כל תאי האפידרמיס מחוברים זה לזה. הקירות החיצוניים של התאים הראשיים עבים יותר מהשאר. כל פני השטח מכוסים בשכבת קאטין ושעוות ירקות. שכבה זו נקראת לציפורן(עור). הוא נעדר על השורשים הגדלים וחלקים תת-מימיים של צמחים. כאשר הוא יבש, החדירות של הציפורן נחלשת באופן משמעותי.
מלבד תאים בסיסיים, ישנם אחרים באפידרמיס, במיוחד שערות, או טריכומות. הם חד-תאיים ורב-תאיים (איור 2). מבחינה פונקציונלית, הם מגדילים את פני האפידרמיס, למשל, באזור הצמיחה של השורשים, משמשים כהגנה מכנית, נצמדים לתמיכה ומפחיתים את איבוד המים. למספר צמחים יש שערות בלוטותלמשל סרפד.
רק בצמחים גבוהים יותר באפידרמיס יש סטומטההמווסתים את חילופי המים והגזים. אם אין לציפורן, אז אין צורך בסטומטה. סטומות הן קבוצה של תאים שנוצרים מנגנון הסטמטאלי,המורכב משניים תאי שמירהותאי אפידרמיס סמוכים תאי צד. הם שונים מתאי האפידרמיס העיקריים (איור 3 ). תאי שמירה שונים מהתאים הסובבים אותם בצורתם ובנוכחותם של מספר רב של כלורופלסטים וקירות מעובים בצורה לא אחידה. אלה שניצבים זה מול זה עבים יותר מהשאר (איור 4) . בין תאי השמירה נוצרים פתח סטמאטאלי, שמוביל ל חלל תת-סטומטישקוראים לו חלל תת-סטומטי.תאי שמירה הם בעלי פוטוסינתזה גבוהה. הם מכילים כמות גדולה של עמילן רזרבה ומיטוכונדריה רבים.
המספר והתפוצה של סטומטות, סוגי מנגנון הסטומטלי משתנים מאוד בצמחים שונים. סטמאטה נעדרת אצל בריופיטים מודרניים. הפוטוסינתזה בהם מתבצעת על ידי דור הגמטופיטים, והספורופיטים אינם מסוגלים להתקיים עצמאי.
בדרך כלל סטמטות ממוקמות בחלק התחתון של העלה. בצמחים הצפים על פני המים - במשטח העליון. בעלים של דגנים, העגבניות מפוזרות לרוב באופן שווה משני הצדדים. עלים כאלה מוארים באופן שווה יחסית. על 1 מ"מ 2 של פני השטח, מ 100 עד 700 stomata ניתן לאתר.
רקמה משנית (periderm). רקמה זו מחליפה את האפידרמיס כאשר הצבע הירוק של הנבטים השנתיים משתנה לחום. הוא רב שכבתי ומורכב משכבה מרכזית של תאים קמביים - פלוגן.תאי פלוגן, מתחלקים, מניחים שכבה בחוץ בחורים,ובפנים - פלודרמה(איור 5).
פלמה, או פְּקָק. בתחילה הוא מורכב מתאי חיים בעלי דופן דקה. עם הזמן, הקירות שלהם ספוגים בשעוות סוברין ושעוות ירקות ומתים. תוכן התא מתמלא באוויר.
פונקציות פלמה:
מונע אובדן לחות;
מגן על הצמח מפני נזק מכני;
מגן מפני פתוגנים;
מספק בידוד תרמי, שכן התאים מתמלאים באוויר.
תאים של הפלוגן, הממוקמים באפידרמיס עצמו, השכבה התת אפידרמיסית הבסיסית, לעתים רחוקות יותר בשכבות העמוקות של הקורטקס הראשוני, הם הבסיס היוצר של הקורטקס הראשוני.
שכבת הפקק אינה קבועה. מתרחשות בו הפסקות, המתקשרות עם החללים הבין-תאיים הנמצאים בקרבת מקום. במקביל נוצרות פקעות קטנות על פני השטח - עדשים, המתקשרים בין חללים בין-תאיים לאוויר אטמוספרי (איור 6,7).
בסתיו, הפלוגן מתחת לעדשים מניח שכבה של תאים פקוקים, אשר מפחיתים מאוד את הטרנספירציה, אך אינם שוללים אותה לחלוטין. באביב, שכבה זו קורסת מבפנים. על קליפת ליבנה בהירה, עדשים נראות בבירור בצורה של קווים כהים.
רקמה שלישונית (קרום),הוא גם אופייני רק לצורות עץ של צמחים.
הפלוגן מונח שוב ושוב בשכבות העמוקות של הקורטקס. הרקמות שנמצאות מחוצה לו בסופו של דבר מתות ויוצרות קרום. התאים שלו מתים ואינם מסוגלים להימתח. עם זאת, תאים חיים הממוקמים עמוק יותר מתחלקים, מה שמוביל לעלייה בגודל הרוחבי של הגבעול. עם הזמן, השכבה החיצונית של הקרום נשברת. זמן התרחשות של פער כזה הוא ערך קבוע למדי עבור צמחים בודדים. בעץ תפוח זה קורה בשנה השביעית לחיים, בקרן - בחמישים. במינים מסוימים הוא אינו מופיע כלל. תפקידו העיקרי של הפילינג הוא הגנה מפני נזקים מכניים ותרמיים.
פרנכימה
זוהי קבוצה של רקמות מיוחדות הממלאות את החללים בתוך גוף הצמח בין רקמות מוליכות למכניות. (איור 8). לעתים קרובות יותר לתאי פרנכימה יש צורה מעוגלת, לעתים רחוקות יותר צורה מוארכת. נוכחותם של חללים בין-תאיים מפותחים אופיינית. המרווחים בין התאים יוצרים יחד מערכת הובלה - אפופלסטבנוסף, החללים הבין-תאיים מהווים את "מערכת האוורור" של הצמח. באמצעות סטמטות, או עדשים, הן מחוברות לאוויר אטמוספרי ומספקות הרכב גז אופטימלי בתוך הצמח. חללים בין-תאיים מפותחים נחוצים במיוחד עבור צמחים הגדלים על אדמה ספוגת מים, שבה חילופי גזים רגילים קשה. פרנכימה זו נקראת אירנכימה(איור 9).
אלמנטים של הפרנכימה, הממלאים את הפערים בין רקמות אחרות, מבצעים גם את הפונקציה של תמיכה. תאי פרנכימה חיים, אין להם דפנות תאים עבות, כמו סקלרנכימה. לכן, תכונות מכניות מסופקות על ידי טורגור. אם תכולת המים יורדת, מה שמוביל לפסמוליזה ונבילה של הצמח.
פרנכימה הטמעהנוצר על ידי תאים דקים עם רווחים בין-תאיים רבים. התאים של מבנה זה מכילים כלורופלסטים רבים, כך זה נקרא כלורנכימה. כלורופלסטים ממוקמים לאורך הקיר ללא הצללה זה על זה. בפרנכימה ההטמעה מתרחשות תגובות פוטוסינתזה, המספקות לצמח חומרים אורגניים ואנרגיה. התוצאה של תהליכי פוטוסינתזה היא האפשרות לקיומם של כל היצורים החיים על פני כדור הארץ.
רקמות הטמעה קיימות רק בחלקים המוארים של הצמח; הן מופרדות מהסביבה על ידי אפידרמיס שקוף. אם האפידרמיס מוחלף ברקמות אינגומנטריות משניות אטומות, פרנכימה ההטמעה נעלמת.
פרנכימה של אחסוןמשמש כמאגר של חומרים אורגניים שאינם משמשים באופן זמני את האורגניזם הצמחי. באופן עקרוני, כל תא עם פרוטופלסט חי מסוגל להפקיד חומר אורגני בצורה של סוגים שונים של תכלילים, אך חלק מהתאים מתמחים בכך. (איור 10, 11). תרכובות עשירות באנרגיה מופקדות רק בעונת הגידול, נצרך בתקופה הרדומה ולקראת עונת הגידול הבאה. לכן, חומרי רזרבה מופקדים באיברים הצומחים רק בצמחים רב-שנתיים.
כלי הקיבול של המניות יכול להיות איברים רגילים (בריחה, שורש), כמו גם מתמחים (קני שורש, פקעות, פקעות). כל צמחי הזרע אוגרים חומרים בעלי ערך אנרגטי בזרעים (קוטילונים, אנדוספרם). צמחים רבים באקלים צחיח אוגרים לא רק חומרים אורגניים, אלא גם מים. (איור 12). לדוגמה, אלוורה אוגרת מים בעלים בשרניים, קקטוסים בנצרים.
בדים מכניים
התכונות המכניות של תאי צמחים מסופקות על ידי:
קליפה קשה של התא;
עכירות, כלומר, מצב הטורגור של תאים.
למרות העובדה שכמעט לכל תאי הרקמה יש תכונות מכניות, עם זאת, יש רקמות בצמח שתכונות מכניות הן בסיסיות עבורן. זֶה קולנכימהו סקלרנכימה. בדרך כלל הם מתפקדים באינטראקציה עם רקמות אחרות. בתוך הגוף יוצרים הצמחים מעין מסגרת. לכן הם נקראים חיזוק.
לא לכל הצמחים יש רקמות מכניות מוגדרות היטב. במידה הרבה פחות, צמחים החיים בסביבה המימית זקוקים לתמיכה פנימית מאשר יבשתיים. הסיבה היא שצמחי מים זקוקים פחות לתמיכה פנימית. גופם נתמך במידה רבה על ידי המים שמסביב. אוויר ביבשה אינו יוצר תמיכה כזו, שכן יש לו צפיפות נמוכה יותר מאשר מים. מסיבה זו נוכחותם של רקמות מכניות מיוחדות הופכת לרלוונטית.
שיפור של מבני תמיכה פנימיים התרחש בתהליך האבולוציה.
קולנכימה.הוא נוצר רק על ידי תאים חיים, מוארכים לאורך ציר האיבר. סוג זה של רקמה מכנית נוצר מוקדם מאוד, במהלך תקופת הצמיחה הראשונית. לכן, חשוב שהתאים יישארו בחיים וישמרו על יכולת המתיחה יחד עם תאי המתיחה שנמצאים בקרבת מקום.
תכונות של תאי קולנכימה:
עיבוי לא אחיד של הקליפה, וכתוצאה מכך חלקים מסוימים ממנה נשארים דקים, בעוד שאחרים מתעבים;
קונכיות אינן מתגרות.
תאי קולנכימה ממוקמים באופן שונה זה מזה. בתאים סמוכים נוצרים עיבויים בפינות הפונות זו לזו ( אורז. 13 ). קולנכימה זו נקראת פינה.במקרה אחר, התאים מסודרים בשכבות מקבילות. קרומי התא הפונים לשכבות אלו מעובות מאוד. זֶה קולנכימה למלרית.ניתן לסדר תאים בצורה רופפת, עם שפע של רווחים בין-תאיים הוא קולנכימה רופפת.קולנכימה כזו נמצאת לעתים קרובות בצמחים על קרקעות ספוגות מים.
לקולנכימה חשיבות מיוחדת בצמחים צעירים, בצורות עשבוניות, וגם בחלקי צמחים שבהם לא מתרחשת צמיחה משנית, כגון עלים. במקרה זה, הוא מונח קרוב מאוד לפני השטח, לפעמים ממש מתחת לאפידרמיס. אם לאיבר יש פרצופים, אז שכבות עבות של קולנכימה נמצאות לאורך הפסגות שלהן.
תאי קולנכימה מתפקדים רק בנוכחות טורגור. מחסור במים מפחית את יעילות הקולנכימה והצמח קמל באופן זמני, למשל, עלי מלפפון נפולים ביום חם. לאחר מילוי התאים במים, הפונקציות של הקולנכימה משוחזרות.
סקלרנכימה.הסוג השני של רקמות מכניות. בניגוד לקולנכימה, שבה כל התאים חיים, תאי סקלנכימה מתים. הקירות שלהם עבים מאוד. הם מבצעים פונקציה מכנית. עיבוי חזק של הממברנה מוביל להפרה של הובלה של חומרים, וכתוצאה מכך הפרוטופלסט מת ( אורז. 14 ). הליגניפיקציה של ממברנות התא של סקלרנכימה מתרחשת כאשר איבר הצמח כבר השלים את צמיחתו. לכן, הם כבר לא מפריעים למתיחה של הרקמות הסובבות.
בהתאם לצורה, מבחינים בין שני סוגים של תאי סקלרנכימה - סיבים וסקלרים.
סיביםיש צורה מוארכת מאוד עם קירות עבים מאוד חלל קטן. הם קצת יותר קטנים מסיבי עץ. לעתים קרובות מתחת לאפידרמיס יוצרים שכבות אורך וגדילים. בפלואם או בקסלם ניתן למצוא אותם בנפרד או בקבוצות. בפלומה הם נקראים סיבי באסט(אורז. 15 ), ובקסילם סיבים ליבריפורמים(אורז. 16 ).
סקלרידס,אוֹ תאים אבנים, מיוצגים על ידי תאים מעוגלים או מסועפים עם קליפות חזקות. בגופו של צמח ניתן למצוא אותם בנפרד (תאים תומכים) ( איור.17 ) או קבוצות (איור 18). יש לציין כי המאפיינים המכניים תלויים מאוד במיקום הסקלרידים. חלק מהסקלרים יוצרים שכבות רצופות, כמו למשל בקליפת האגוזים או באבני הפירות (פירות האבן).
רקמות הפרשה
לכל תא חי יש בתחילה את הפונקציה של סילוק חומרים, בעוד שההובלה יכולה לעבור הן לאורך שיפוע הריכוז והן כנגד השיפוע. ניתן להסיר חומרים שעברו סינתזה בתא ובהמשך ישפיעו על פעילותם של תאים אחרים (פיטו-הורמונים, אנזימים). תהליך כזה נקרא הַפרָשָׁה. במקרה שבו מופרשים חומרים שהם תוצרי פסולת של התא, התהליך נקרא הַפרָשָׁה. למרות העובדה שלהפרשת חומרים בתאי צמחים ובעלי חיים יש דמיון מהותי, לצמחים יש מספר תכונות הנובעות מהבדלים מהותיים בפעילות החיונית. רמת התגובות המטבוליות בצמחים נמוכה בהרבה מאשר בבעלי חיים. לכן, כמויות קטנות יותר של פסולת נפלטים באופן יחסי. תכונה נוספת היא שבסינתזה עצמאית של כמעט כל התרכובות האורגניות הדרושות, צמחים לעולם אינם יוצרים עתודות מוגזמות שלהם. החומרים המשתחררים יכולים לשמש כחומר מוצא לתגובות אחרות (לדוגמה, CO 2 ו- H 2 O).
אם אצל בעלי חיים תהליך הפרשת הרעלים קשור להסרתם מהגוף, ייתכן שהצמחים לא יעשו זאת, ויבודדו חומרים מיותרים בפרוטופלסט החי (הסרה של חומרים שונים לתוך הוואקואול), בתאים מתים (רוב הרקמות של צמחים רב שנתי). הצמח מורכב מתאים כאלה), בחללים בין-תאיים.
ניתן לחלק את החומרים הנפלטים לשתי קבוצות גדולות. הקבוצה הראשונה היא חומרים אורגניים המסונתזים ישירות על ידי התא (אנזימים, פוליסכרידים, ליגנינים, טרפנים, האחרונים הם מרכיבים מרכיבים של שמנים אתריים ושרף). הקבוצה השנייה - חומרים הנכנסים בתחילה לתא מבחוץ בעזרת שינוע קסילם או פלואם (מים, מלחים מינרלים, חומצות אמינו, חד סוכרים ועוד). הסוד המופרש לעיתים רחוקות הוא הומוגני ובדרך כלל מורכב מתערובת שבה חומר אחד הוא בעל הריכוז הגבוה ביותר.
לצמחים, בניגוד לבעלי חיים, אין מערכת הפרשה מלאה. ישנם רק מבנים מיוחדים הפזורים ברחבי הצמח - אידיובלסטים.התאים היוצרים אותם קטנים יותר מתאי הפרנכימה השוכנים בקרבת מקום. יש להם ציטופלזמה צפופה באלקטרונים, עם אלמנטים מפותחים של הרשת האנדופלזמית וקומפלקס גולגי. לרוב לא בא לידי ביטוי, יש וואקוול מרכזי. בינם לבין תאים חיים אחרים, תאים אלה מחוברים על ידי פלסמודסמטה רבים.
בהתאם למיקום, רקמות הפרשה יכולות להיות חיצוני ופנימי.
רקמות הפרשה חיצוניות.שערות בלוטותהם נגזרות עִלִית הָעוֹר. מבחינה מורפולוגית הם משתנים - הם יכולים להיות בעלי ראש רב תאי, להיות מוארכים, בצורת מגן על רגל וכו'. (איור 19). אלה כוללים שערות סרפד צורבות.
הידתודות- אלו הם מבנים המשחררים עודפי מים בתנאים של טרנספירציה נמוכה ולחות גבוהה - guttation. מבנים אלה נוצרים על ידי קבוצות של תאים חיים חסרי צבע עם קירות דקים - התואר.רקמה זו צמודה לצרור המוליך (איור 20). מים משתחררים דרך סטמטות מים מיוחדות, הנבדלות מהסטומטות רגילות על ידי חוסר תנועה ופער פתוח כל הזמן. הרכב נוזל הביוב משתנה מאוד ממים כמעט טהורים ועד לתערובת מורכבת מאוד של חומרים.
נקטריות.צמחים רבים מתאפיינים בשחרור נוזל המכיל בין 7 ל-87% חד- ואוליגוסכרידים. תהליך זה מתבצע על ידי מבנים מיוחדים - נקטריות. תלוי במיקום, יש פֶּרַח,אוֹ פִּרחִי, נקטריונים (איור 21), כמו גם צמחים הממוקמים על גבעולים, עלים - אקסטרהפלורה, או נקטריות חוץ פרחוניות.נקטר עשוי לייצג זהו מיץ פלואם ללא שינוי, אשר מועבר אל פני השטח דרך החללים הבין-תאיים ומופרש דרך הסטומטה. נקטרים מורכבים יותר נוצרים על ידי פרנכימה בלוטית, מכוסה באפידרמיס עם שערות בלוטות. הצוף מופרש על ידי תאי אפידרמיס או שערות בלוטות. במקרה זה, הצוף המופרש שונה ממוהל הפלואם. מכיוון שהוא נשלט על ידי גלוקוז, פרוקטוז, סוכרוז, ובמיץ פלואם - גלוקוז. יונים נמצאים בריכוז נמוך. יונים נמצאים בריכוז נמוך. כדי למשוך מאביקים, הצוף עשוי להכיל הורמונים סטרואידים, הנחוצים לחרקים.
בלוטות עיכולקיים בצמחים אוכלי חרקים (טל, nepentis וכו'). בתגובה למגע של חסרי חוליות קטנים, הם מפרישים סוד המכיל אנזימים הידרוליטיים, חומצות פורמיות, בוטיריות ופרופיוניות. כתוצאה מכך, ביופולימרים מתפרקים למונומרים, המשמשים את המפעל לצרכיו.
בלוטות מלחלהתפתח בצמחים הגדלים על קרקעות מלוחות. בלוטות אלה ממוקמות על העלים ובצורה מומסת, יונים עודפים מועלים אל פני השטח. מלח מופקד תחילה על הציפורן, ואז הוא נשטף בגשם (איור 22). בנוסף, צמחים רבים יש על העלים שערות מלח.כל שערה מורכבת משני תאים: האחד יוצר את הראש, השני - הגבעול. מלחים מצטברים ב-vacuole של תא הראש, וכאשר ריכוזם הופך גבוה, הראש נופל. במקומו צומח תא חדש שצובר גם מלחים. במהלך צמיחת העלה, הראש עלול ליפול ולהיווצר מספר פעמים. היתרון של שערות מלח הוא שעל ידי שחרור מלחים הן מאבדות מעט מים. תופעה זו חשובה מאוד לצמחים הגדלים על קרקעות מלוחות, בהן מים מתוקים מועטים.
רקמות הפרשה פנימית.מבני הפרשה פנימיים, המפוזרים בכל גוף הצמח בצורה של אידיובלסטים, אינם מסירים חומרים מחוץ לגוף, צוברים אותם בעצמם. אידיובלסטים הם תאים צמחיים השונים בצורתם, במבנה או בתוכן מתאי אחרים באותה רקמה.בהתאם למקור, מבחינים בכלי קיבול סכיזוגניים (מיוונית schiso - פיצול) וליזוגניים (ליזיס יווני - פירוק) של חומרים מופרשים (איור 23).
כלי קיבול סכיזוגניים.אלו חללים בין-תאיים נרחבים יותר או פחות מלאים בחומרים המופרשים. לדוגמה, מעברי שרף של עצי מחט, ארליה, מטרייה ואחרים. הוא האמין כי לשרף יש תכונות קוטל חיידקים, דוחה אוכלי עשב, מה שהופך צמחים בלתי אכילים עבורם.
כלי קיבול ליסיגנייםנוצרים במקום תאים חיים שמתים ונהרסים לאחר הצטברות חומרים בהם. ניתן לראות תצורות כאלה בקליפת פירות הדר.
בנוסף לסוגים העיקריים, יש צורות ביניים. חֲלָבִי- ישנם שני סוגים: מפרקיםו לא מפולח. הראשונים נוצרים מתאי חיים רבים המסודרים בשרשראות. בנקודת המגע מושמדים הקליפות, הפרוטופלסטים מתמזגים וכתוצאה מכך נוצרת רשת אחת. חלביים כאלה נמצאים ב-Compositae. לא מפולח lactifers נוצרים על ידי תא ענק מרובה גרעינים. הוא מתעורר בשלבי ההתפתחות המוקדמים, מתארך ומסתעף ככל שהצמח גדל ויוצר רשת מורכבת, למשל בצמחי אופורביה (איור 25).
קירות התא של lactifers מושקים בכבדות. הם אינם מתכלים ולכן הם פלסטיים. החלק המרכזי של הלקטיס תפוס על ידי ואקואול עם לטקס, והפרוטופלסט תופס מיקום קרוב לקיר. לא תמיד ניתן לקבוע את הגבול בין הפרוטופלסט ל-vacuole. הלקטיפרים חודרים למריסטם וגדלים יחד עם איבר הצמח.
רקמות מוליכות
סוג זה שייך ל קָשֶׁהרקמות, מורכב מתאים מובחנים בצורה שונה. בנוסף לאלמנטים המוליכים בפועל, הרקמה מכילה אלמנטים מכניים, הפרשה ואגירה (איור 26). רקמות מוליכות מאחדות את כל איברי הצמח למערכת אחת. ישנם שני סוגים של רקמות מוליכות: עֵצָהו פלואם(קסילון יווני - עץ; phloios - קליפה, באסט). יש להם הבדלים מבניים ותפקודיים כאחד.
האלמנטים המוליכים של הקסלם נוצרים על ידי תאים מתים. הם נושאים הובלה למרחקים של מים וחומרים מומסים בהם מהשורש לעלים. האלמנטים המוליכים של הפלואם שומרים על הפרוטופלסט בחיים. הם נושאים הובלה למרחקים ארוכים מעלים פוטוסינתטיים לשורש.
בדרך כלל קסילם ופלואם ממוקמים בגוף הצמח בסדר מסוים, יוצרים שכבות או צרורות מוליכים. בהתאם למבנה, נבדלים מספר סוגים של צרורות מוליכים, האופייניים לקבוצות מסוימות של צמחים. בצרור פתוח בטחונותבין הקסילם והפלואם נמצא הקמביום, המספק צמיחה משנית (איור 27-A, 28). בצרור פתוח דו-צדדיהפלואם ממוקם ביחס לקסילם משני הצדדים (איור 27-B, 29). צרורות סגורותאינם מכילים קמביום, ולכן אינם מסוגלים להתעבות משני (איור 27-B, 27-D, 30.31). יש עוד שני סוגים חבילות קונצנטריות,כאשר הפלואם מקיף את הקסם (איור 27-E, 32), או שהקסילם מקיף את הקסם (איור 27-E).
Xylem (עץ). התפתחות הקסילם בצמחים גבוהים יותר קשורה באספקת חילופי מים. מכיוון שהמים מופרשים כל הזמן דרך האפידרמיס, אותה כמות לחות חייבת להיספג על ידי הצמח ולהוסיף לאיברים המבצעים טרנספירציה. יש לקחת בחשבון שנוכחות של פרוטופלסט חי בתאים מוליכים מים תאט מאוד את ההובלה, ותאים מתים מתפקדים כאן יותר. עם זאת, לתא מת אין עכירות לכן, לקליפה חייבת להיות תכונות מכניות. הערה: התפרצות - מצבם של תאי הצמח, הרקמות והאיברים, בהם הם הופכים אלסטיים עקב הלחץ של תכולת התאים על הממברנות האלסטיות שלהם.ואכן, האלמנטים המוליכים של הקסלם מורכבים מתאי מת הנמתחים לאורך ציר האיבר עם קונכיות עבות.
בתחילה, הקסילם נוצר מהמריסטם הראשוני - פרוקמביום, הממוקם בחלק העליון של האיברים הציריים. ראשית מבדיל פרוטוקסילם,לאחר מכן מטקסילם.ידועים שלושה סוגים של היווצרות קסלם. בְּ exarchסוג, אלמנטים פרוטוקסילם מופיעים תחילה על הפריפריה של הצרור הפרוקמביאלי, ואז אלמנטים מטאקסילם מופיעים במרכז. אם התהליך הולך בכיוון ההפוך (כלומר מהמרכז לפריפריה), אז זה אנדרכיסוּג. בְּ סוג mesarchהקסיל מונח במרכז הצרור הפרוקמביאלי, ולאחר מכן הוא מופקד הן לכיוון המרכז והן לכיוון הפריפריה.
השורש מאופיין בסוג אקסרק של היווצרות קסלם, ואילו לגבעולים יש סוג אנדרך. בצמחים מאורגנים נמוכים, שיטות היווצרות הקסיל מגוונות מאוד ויכולות לשמש כמאפיינים שיטתיים.
בצמחים מסוימים (לדוגמה, חד-צוגיים), כל התאים הפרוקמביאליים מתמיינים לרקמות מוליכות שאינן מסוגלות להתעבות משני. בצורות אחרות (למשל, וודי), מריסטמים לרוחב (קמביום) נשארים בין הקסילם והפלואם. תאים אלו מסוגלים להתחלק, לחדש את הקסילם והפלואם. תהליך כזה נקרא צמיחה משנית.צמחים רבים הגדלים בתנאי אקלים יציבים יחסית גדלים ללא הרף. בצורות המותאמות לשינויי אקלים עונתיים - מעת לעת. כתוצאה מכך נוצרות טבעות צמיחה שנתיות מוגדרות היטב.
השלבים העיקריים של התמיינות של תאים פרוקמביאליים. לתאיו יש ממברנות דקות שאינן מונעות מהם להימתח במהלך צמיחת האיבר. לאחר מכן, הפרוטופלסט מתחיל להניח את הממברנה המשנית. אבל לתהליך הזה יש תכונות מובהקות. הממברנה המשנית אינה מופקדת בשכבה רציפה, שלא תאפשר לתא להימתח, אלא בצורת טבעות או בספירלה. התארכות התא אינה קשה. בתאים צעירים, הטבעות או הסלילים של הסליל ממוקמים קרוב זה לזה. בתאים בוגרים הם מתפצלים כתוצאה ממתיחה של התאים (איור 33). עיבויים טבעתיים וספירליים של הקליפה אינם מונעים צמיחה, אולם מבחינה מכנית הם נחותים מקונכיות, כאשר העיבוי המשני יוצר שכבה רציפה. לכן, לאחר הפסקת הגדילה בקסלם נוצרים אלמנטים בעלי מעטפת מתמשכת מתמשכת ( מטקסילם). יש לציין שהעיבוי המשני כאן אינו טבעתי או ספיראלי, אלא מנוקד, דמוי מדרגות, מרושת (איור 34). התאים שלו אינם מסוגלים להימתח, ומתים תוך מספר שעות. תהליך זה בתאים סמוכים מתרחש באופן מתואם. מספר רב של ליזוזומים מופיעים בציטופלזמה. ואז הם מתפרקים, והאנזימים שבהם הורסים את הפרוטופלסט. כאשר הקירות הרוחביים נהרסים, התאים הממוקמים בשרשרת זה מעל זה יוצרים כלי חלול (איור 35). לרוב האנגיוספרמים ולחלק מהשרכים יש כלי דם.
תא מוליך שאינו נוצר דרך נקבים בדופן נקרא tracheid.תנועת המים דרך הטראכאידים איטית יותר מאשר דרך הכלים. העובדה היא שבטרכאידים הממברנה הראשונית אינה נקטעת בשום מקום. הטרכאידים מתקשרים זה עם זה באמצעות מאז.יובהר כי בצמחים הנקבובית היא רק שקע בממברנה המשנית עד לקרום הראשוני, ואין נקבים דרך בין הטרכאידים.
הנפוצות ביותר הן נקבוביות עם גבולות (איור 35-1). הערוץ שלהם, הפונה לחלל התא, יוצר שלוחה - תא נקבוביות.לנקבוביות של רוב הצמחים המחטניים על הקליפה הראשונית יש עיבוי - בֶּלֶט,שהוא מעין שסתום ומסוגל לווסת את עוצמת הובלת המים. על ידי תזוזה, הטורוס חוסם את זרימת המים דרך הנקבובית, אך לאחר מכן הוא אינו יכול לחזור למקומו הקודם, מבצע פעולה חד פעמית.
הנקבוביות פחות או יותר מעוגלות, מוארכות בניצב לציר המוארך (קבוצה של נקבוביות אלו מזכירה סולם, ולכן נקבוביות זו נקראת נקבוביות סולם). הובלה דרך הנקבוביות מתבצעת הן בכיוון האורך והן בכיוון הרוחבי. נקבוביות קיימות לא רק בטרכאידים, אלא גם בתאי כלי דם בודדים היוצרים כלי.
מנקודת המבט של תורת האבולוציה, טרכאידים הם המבנה הראשון והעיקרי שמוליך מים בגוף של צמחים גבוהים יותר. מאמינים שהכלים מקורם בטרכאידים עקב תמוגה של הקירות הרוחביים ביניהם (איור 36). לרוב השרכים והג'מנוספרמים אין. הם מעבירים מים דרך טראכאידים.
בתהליך של התפתחות אבולוציונית, כלי דם התעוררו שוב ושוב בקבוצות שונות של צמחים, אך הם קיבלו את המשמעות התפקודית החשובה ביותר באנגיוספרמים, שיש להם אותם יחד עם טרכיידים. מאמינים שהחזקת מנגנון תחבורה מתקדם יותר סייעה להם לא רק לשרוד, אלא גם להשיג מגוון משמעותי של צורות.
Xylem היא רקמה מורכבת, בנוסף לאלמנטים מוליכים מים, היא מכילה אחרים. פונקציות מכניות מבוצעות על ידי סיבים ליבריפורמים ( La T. liber - lub, פורמה - צורה). נוכחותם של מבנים מכניים נוספים חשובה מכיוון שלמרות העיבוי, הקירות של האלמנטים נושאי המים עדיין דקים מדי. הם אינם מסוגלים להחזיק באופן עצמאי מסה גדולה של צמח רב שנתי. הסיבים התפתחו מטראכאידים. הם מאופיינים בגדלים קטנים יותר, פגזים lignified (lignified) וחללים צרים. על הקיר ניתן למצוא נקבוביות נטולות שוליים. סיבים אלו אינם יכולים להוליך מים, תפקידם העיקרי הוא תמיכה.
הקסילם מכיל גם תאים חיים. המסה שלהם יכולה להגיע ל-25% מנפח העץ הכולל. מכיוון שלתאים אלה יש צורה מעוגלת, הם נקראים פרנכימה עץ. בגוף הצמח, הפרנכימה ממוקמת בשתי דרכים. במקרה הראשון, התאים מסודרים בצורה של גדילים אנכיים - זהו פרנכימה של גדיל. במקרה אחר, הפרנכימה יוצרת קרניים אופקיות. קוראים להם קרני ליבה, כשהם מחברים את הליבה והקליפה. הליבה מבצעת מספר פונקציות, כולל אחסון חומרים.
פלואם (באסט). זוהי רקמה מורכבת, שכן היא נוצרת על ידי תאים הטרוגניים. התאים המוליכים העיקריים נקראים אלמנטים מסננת(איור 37). האלמנטים המוליכים של הקסלם נוצרים על ידי תאים מתים, בעוד שבפלואם הם שומרים על פרוטופלסט חי, אם כי שונה מאוד, במהלך תקופת התפקוד. הפלואם מבצע את יציאתם של חומרים פלסטיים מהאיברים הפוטוסינתטיים. לכל התאים החיים של הצמחים יש את היכולת להוביל חומרים אורגניים. ומכאן שאם ניתן למצוא את הקסילם רק בצמחים גבוהים יותר, אז הובלת חומרים אורגניים בין תאים מתבצעת גם בצמחים נמוכים יותר.
קסילם ופלואם מתפתחים ממריסטמים אפיקליים. בשלב הראשון, בחבל הפרוקמביאלי, א פרוטופלואם.כשהרקמות שמסביב גדלות, הוא נמתח, וכאשר הצמיחה מסתיימת, במקום פרוטופלואם, הוא נוצר מטפלואם.
בקבוצות שונות של צמחים גבוהים יותר ניתן למצוא שני סוגים של יסודות מסננת. בשרכים וגימנוספרמים, הוא מיוצג תאי מסננת.שדות המסננת בתאים מפוזרים לאורך הקירות הצדדיים. הפרוטופלסט שומר על גרעין הרוס מעט.
באנגיוספרמים, יסודות מסננת נקראים צינורות מסננת.הם מתקשרים זה עם זה דרך צלחות מסננת. לתאים בוגרים חסרים גרעינים. עם זאת, ליד צינור המסננת ממוקם תא לוויין, הנוצר יחד עם צינור המסננת כתוצאה מחלוקה מיטוטית של תא האם המשותף (איור 38). לתא הנלווה יש ציטופלזמה צפופה יותר עם מספר רב של מיטוכונדריה פעילות, כמו גם גרעין מתפקד במלואו, מספר עצום של פלסמודסמטה (פי עשרה מתאים אחרים). תאים נלווים משפיעים על הפעילות התפקודית של תאי צינורות מסננת לא גרעינית.
למבנה של תאי מסננת בוגרים יש כמה מוזרויות. אין ואקוול, ולכן הציטופלזמה נוזלית מאוד. הגרעין עשוי להיעדר (באנגיוספרמים) או להיות במצב מקומט, לא פעיל מבחינה תפקודית. גם ריבוזומים וקומפלקס גולגי נעדרים, אך הרשת האנדופלזמית מפותחת היטב, אשר לא רק חודרת לציטופלזמה, אלא גם עוברת לתאים שכנים דרך הנקבוביות של שדות המסננת. מיטוכונדריה ופלסטידים מפותחים היטב נמצאים בשפע.
בין תאים, הובלת חומרים עוברת דרך חורים הממוקמים על ממברנות התא. פתחים כאלה נקראים נקבוביות, אך בניגוד לנקבוביות הטראכייד, הם עוברים. ההנחה היא שהם פלסמודסמטה מורחבת מאוד, שעל קירותיה מופקד הפוליסכריד הקאלוז. הנקבוביות מסודרות בקבוצות, נוצרות שדות מסננת. בצורות פרימיטיביות, שדות מסננות מפוזרים באופן אקראי על פני כל פני המעטפת; באנגיוספרמים מתקדמים יותר, הם ממוקמים בקצוות של תאים שכנים הסמוכים זה לזה, נוצרים צלחת מסננת(איור 39). אם יש עליו שדה מסננת אחד, זה נקרא פשוט, אם יש כמה - מורכבים.
מהירות תנועת התמיסות דרך יסודות המסננת היא עד 150 ס"מ ∕ שעה. זהו פי אלף מקצב הדיפוזיה החופשית. ככל הנראה מתרחשת הובלה פעילה, ומיטוכונדריות רבות של יסודות מסננת ותאי לוויין מספקים את ה-ATP הדרוש לכך.
החיים של יסודות המסננת של הפלואם תלויים בנוכחות של מריסטמים לרוחב. אם הם קיימים, אז אלמנטי המסננת פועלים לאורך כל חיי הצמח.
בנוסף ליסודות מסננת ותאי לוויין, הפלואם מכיל סיבי באסט, סקלרידים ופרנכימה.
שאלות לשליטה וחזרה
סימנים לצמחים גבוהים יותר? איזה דור של צמחים גבוהים יותר נקרא גמטופיט? ספורופיט?
מהן הסיבות התורמות להתפתחות רקמות מובחנות בצמחים יבשתיים.
רשום את סוגי הצמחים.
אילו קבוצות של מריסטמים אתה מכיר? היכן הם ממוקמים בגוף הצמח?
רקמות מרכיבים, תפקידיהן, משמעות.
ארגון סוגים שונים של רקמות אינטגמנטריות.
המבנה של מנגנון הסטומטלי. רשום את סוגי מכשירי הסטומטלי המוכרים לך. כיצד מתפקדות הסטומטה?
תכונות של רקמות parenchymal, סוגי parenchyma.
סימנים של פרנכימה וסקלרנכימה.
רשום את סוגי מבני ההפרשה של צמח. מה המשמעות שלהם?
שיטות ליצירת כלי קיבול לחומרים המופרשים.
מה זה חולבים? לאילו סוגים הם מחולקים?
כיצד מאורגנות רקמות מוליכות?
מה זה צרורות מוליכים? הטיפוסים שלהם.
כיווני תנועה של חומרים דרך הקסילם? פלואם? האם הרכב החומרים המובלים שונה בקסלם ובפלואם?
המבנה והתפקודים של xylem, phloem.
ארגון אלמנטים מוליכים של פלואם בצמחים מקבוצות שיטתיות שונות.
מתמחה וחיידקי, בעלי דופן תא חיצונית. בתאים גבוהים יותר מבנה ותפקודים של אברונים תאים. חוקים של ג' מנדל עבודה >>
סוּג בדים. קרובינו הקרובים ביותר, יונקים אחרים, מִבְנֶהו... , המורכב מאחד תאים. צמחיםמורכב גם ממגוון ... microtubules, centrioles מרכז תאים. IN ירקות תאיםכמו כן מופרשים כלורופלסטים, שבהם ...
צבע אדום. IN רקמות צמחיםניתן למצוא נגזרות אנתרצין ... נגזרות אנתרצין כמו ברפואה ירקותחומרי גלם, ו... מוקפים ב-2-3, לעתים רחוקות יותר 4 תאיםעִלִית הָעוֹר. שערות חד-תאיות, ... תרכובות, מבוססות מבניםמתוכם המבנה של אנתראקן...