ייחודי בדרכו שלו מספק את הגיוון ותכונות החיים שלהם.
המיקרוביולוגיה המודרנית למחקר ולסיווג משתמשת בדברים הבאים תכונות ייחודיותבַּקטֶרִיָה:
- מבנה תאי מורפולוגי:
- "קוקי" כדוריים;
- "בצילים" בצורת מוט;
- ספירלה "ספירושטים";
- צורה מפותלת "ויבריוס".
- נְשִׁימָה:
- לנשום חמצן "אירובי";
- אנוקסי "אנאירובי".
- שיטת האכלה:
- אוטוטרופי "עצמאי" - מסוגל לסנתז חומר אורגנימאנאורגנית בעזרת אנרגיה תגובה כימיתאו פוטוסינתזה (לדוגמה, אצות כחולות-ירקות);
- הטרוטרופי - להמיר ולעבד חומרים אורגניים מוכנים המתקבלים כתוצאה מפעילות חיונית של אורגניזמים אחרים, זה כולל מיקרואורגניזמים פתוגניים (פתוגניים) וסימביוטיים (מועילים), כמו גם חיידקים החיים עקב תהליכי תסיסה, ריקבון וכו' .
- בית גידול:
- סביבה חיצונית (אדמה, מים וכו');
- בתוך אורגניזמים חיים;
- חומרים אורגניים מתים (גופות, משקעים, פסולת).
- שיטת הפצה והישרדות (בין אם נוצרים נבגי תאים, מעטפות, ריר).
- תגובה למבחן גראם (הפרדה לפי הרכב ומבנה הקירות, סיווג מפתח בהקשר לנושא המבנה דופן תא):
- גרם חיובי - מוכתם במהלך הליך צביעה גראם (הקליפה חדירה לצבע אנילין (סגול קריסטל, סגול מתיל וכו'));
- גרם שלילי - אין לצבוע במהלך הליך כתם גראם (דופן התא אטומה לצבע).
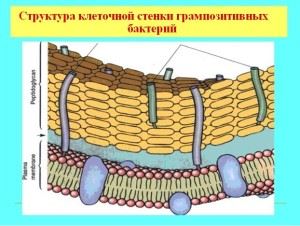
תכונות של דופן התא של חיידקים גרם חיוביים
מהות הבדיקה (הוצעה על ידי G.K. Gram בשנת 1884) היא ההליך לעיבוד דגימה של תרבית חיידקים עם צבע אנילין (סגול קריסטל, סגול קריסטל). לאחר כביסה עם אלכוהול, מיני גראם (+) של מיקרואורגניזמים שומרים על צבעם הכחול, בעוד שאחרים הופכים לדהויים. היכולת של דופן התא לקבע את הצבע נקבעת על פי הרכב ומבנה דופן התא. הסיווג של אורגניזמים חד-תאיים לפי סוג מבנה דופן התא שניתן לצבוע או לא בשיטת גראם הוא אחד המפתחים שבהם. בפועל, תוצאות בדיקת הגרם עוזרות לאבחן מחלות מדבקותלימוד תכונות ביוכימיותמיקרואורגניזמים.
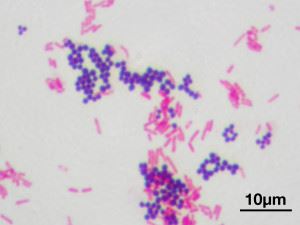
צביעת גראם. Cocci (כדורי) - גרם חיובי ו bacilli (מוטות) - גרם שלילי
לרוב החיידקים החיוביים לגרם יש קרום מסיבי חד-שכבתי החדיר לצבע אנילין. הרכב הקיר כולל מולקולות אורגניות מורכבות, הבסיס של קשיחות מכנית ניתן על ידי murein (הטרופולימר), גליקופפטידים, mucopeptides. מבנה הקיר קובע את צורת המיקרואורגניזם והוא משהו כמו שלד חיצוני, בעל מבנה נקבובי, בעובי של כ-40 מולקולות גליקופפטידים. בנוסף לבסיס הפפטידוגליקן, הקליפה מכילה חומצות טייכואיות ופוליסכרידים.
למרות תגובה כלליתעל הצבע, בין אורגניזמים חיוביים לגרם אפשר לפגוש אורגניזמים השייכים למגוון תת-מינים, צורות שונות, גודל, בית גידול ואופן האכלה.
רוב החיידקים הפתוגניים לבני אדם הם גרא חיוביים, ביניהם, למשל, הגורמים הגורמים למחלות זיהומיות כאלה:
- טטנוס (Clostridium tetani),
- בוטוליזם (Clostridium botulinum),
- אנתרקס (),
- סטרפטוקוקים,
- staphylococci (Staphylococcus aureus), אשר עמידים מאוד לטמפרטורה ולפעולה תרופות.
עמיד לחומצה, בשל המבנה הרב-שכבתי המיוחד של הקליפה (היא כוללת שעווה, פוליסכרידים וחלבונים), ככלל, הם אינם מוכתמים בשיטת גראם. עבורם משתמשים בשיטת זיל-נלסן מיוחדת. אבל חלק נותנים תוצאה חיוביתלפי שיטת גראם במקרה של ריכוז גבוה מאוד של הצבע או טמפרטורה גבוההבמהלך ההליך.
אורגניזמים חד-תאיים מהירי חומצה שורדים בחומצה ו סביבות אלקליות, למרות שהסביבה הנורמלית להתפתחותם ולגדילתם היא ניטרלית. בדרך כלל מסווגים כגראם חיוביים. נציגי קבוצה זו המסוכנים לאנשים ובעלי חיים הם הגורמים הגורמים למחלות כמו שחפת וצרעת. כמו כן עמידים לחומצה כמה חיידקי אדמה המסוגלים לקבע חנקן אטמוספרי ולהיות סימביונים צמחיים.
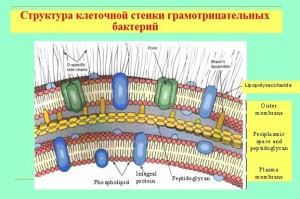
תכונות של הקליפה של חיידקים גרם שליליים
למרות שעובי דופן המוקופפטיד שלהם דק בהרבה, הדופן אטומה לצבע סגול קריסטל. זה הכל על מבנה והרכב כימי שונה מהותית של התא. למעטפת מבנה רב שכבתי ומורכב מ:
- המעטפת החיצונית (מבנים), ההרכב העיקרי שלה הוא ליפוסכרידים וחלבונים;
- חלל פריפלסמי;
- השכבה הפנימית (murein peptidoglycan), ההרכב שלה דומה לזה של הקירות של חיידקים גרם חיוביים, אבל, ככלל, יותר מפעמיים דק יותר.
תודה לעוד מבנה מורכבלחלק מחברי הקבוצה יש עמידות מוגברת לנוגדנים ואנטיביוטיקה.
עם הסיבוך של דופן התא, הופיעו פונקציות ואפשרויות חדשות. סוגים מסוימים של חיידקים החיים משטחים קשים, רכשה את היכולת לשנות את צורת התא (דפנות ה"שלד" הפכו לפלסטיק). זה חשוב בעת החלקה ותנועה על חוסר אחידות של בית גידול מוצק.
החלל הפריפלסמי רכש את הפונקציה של מקום אחסון מבודד לכמה אנזימים מידרוליזים הנחוצים לחיי התא, אך יחד עם זאת מסוגל לפרק את מולקולות הפולימר של החיידק עצמו. עקב הידרוליזה של מולקולות פולימר הנכנסות מבחוץ תא חימרחיב את מגוון החומרים המתאימים לתזונה, במקביל, הממברנה הפנימית, הבלתי חדירה לאנזימים, מונעת את "העיכול העצמי" של תא החיידק.
מבנה הממברנה החיצונית כולל חלבונים שיכולים ליצור נקבוביות הידרופיליות שדרכן יכולות לעבור לתא כמה מקרומולקולות (סוכרים וחומצות אמינו) הנחוצות להזנת חיידקים.
סוג זה של חיידקים כולל חיידקים רבים שיכולים להשתנות תרכובת כימיתבית גידול (חיידקים של תסיסה אצטית ואלכוהולית); סימביוטי - Escherichia coli (E coli), אורגניזמים פתוגניים (גורמי מחלות) רבים, כגון מנינגוקוקוס (Neisseria meningitidis); לגיונלה (Legionella pneumophila), והמפורסמים הליקובקטר פילורימתריס כיב פפטיועמידות גבוהה לחומצות.

שימוש אנושי בתכונות המבניות של חיידקים
המיקרואורגניזמים הראשונים ששימשו את האדם לצרכיהם היו חיידקי תסיסה לקטית ואלכוהולית. הם אלה שבישלו לנו ועדיין מבשלים גבינה, לחם ויין. יתרה מכך, אנשים החלו להשתמש בתוצר של עבודתם הרבה לפני גילוי החיידקים ותחילת לימוד המבנה וההרכב של המוצרים המטבוליים שלהם.
כיום, שיטות ביולוגיות לטיפול בפסולת ומי צואה הפכו לזמינות לא רק מפעלים עירונייםאלא גם לבעלי בתים פרטיים. תרבויות רדומות הן חלק תרופות מודרניותעבור בורות שופכין וביוב מקומי. חוותמשמש לעתים קרובות עמיד לחומצה לעיבוד מהיר של קומפוסט ופסולת בעלי חיים לדשנים ולמטרות רבות אחרות.
כמו כן, חיידקים מהונדסים גנטית רבים מעורבים בייצור תרופות, בסינתזה של סוגים חדשים של פולימרים וחומרים נוספים בעלי תכונות ייחודיות. תכונות כימיותומבנה מורכב.
המרכיבים הכימיים של דופן התא מחולקים בדרך כלל לבסיסיים, צבועים ומושקעים על פניו (adcrusting). לעיקר רכיבים כימייםכוללים תאית, hemicellulose, פקטין, חלבונים. המרכיבים המרכיבים את דופן התא הם ליגנין וסוברין. קוטין ושעווה יכולים להיות מופקדים על פני דופן התא. בנוסף, דפנות התא מכילות סיליקטים וסידן פחמתי.
הרכיבים הנפוצים ביותר של חומרים צמחיים מתחדשים הם תאית, ליגנין והמיצלולוזות (פוליוזות), המהווים 40, בהתאמה; 30 ו-26% ממנו משקל כולל. תאית היא חומר מחזק של דפנות תאים, היוצר מבנים חוטים מסודרים (סיבים). ליגנין והמיצלולוז הם ביו-פולימרים אמורפיים בעלי מבנה רשת או מסועף וממלאים את התפקיד של חומרי מילוי.
המבנה של חומרים אלה, האינטראקציה הבין-מולקולרית שלהם קובעים את תכונות מצעי הצמחים בכללותם ואת התנהגותם במהלך העיבוד.
תָאִית
תאית הוא החומר הנפוץ ביותר בכדור הארץ, הוא השלד של דופן התא של צמחים. מבנה התאית של סיבי כותנה וסיבים המסונתזים על ידי כמה חיידקים, אצות ורכיכות נחקר בצורה הטובה ביותר, שכן תאית נמצאת בצורה הטהורה ביותר בהם. בעץ הוא מתרחש רק בצורת קומפוזיט עם ביופולימרים אחרים, משם קשה לחלץ ללא הרס חלקי.
מבחינה כימית, תאית היא פוליסכריד, שהמאקרומולקולות שלו בנויות משאריות ß-D-גלוקוז ומייצגות שרשרת ליניארית של יחידות anhydroglucopyranose המחוברות בקשרים 1,4-גלוקוזידיים (איור 9).
איור.9. מבנה כימיתָאִית
זהו פולימר סטריאו-רגיל, בשרשרת שלו שייר צלוביוז משמש כחולייה חוזרת. הנוסחה הכוללת של תאית יש את הצורה - (C 6 H 10 O 5) n או [C 6 H 7 O 2 (OH) 3] n. יחידת המונומר של תאית מכילה שלוש קבוצות הידרוקסיל אלכוהול, מהן אחת ראשונית - CH 2 OH, ושתיים ב-C 2 ו- C 3 הן משניות - CHOH. חוליות הקצה שונות משאר חוליות השרשרת. לקישור קצה אחד (ב-C 4) יש הידרוקסיל אלכוהול משני חופשי נוסף. הקישור הסופי (ב-C 1) מכיל הידרוקסיל גליקוזידי (חצי-אצטלי) חופשי ויכול להתקיים בשתי צורות טאוטומריות - מחזורית (חצי-אצטלי) ופתוחה (אלדהיד). קבוצת האלדהיד הסופית מעניקה לתאית יכולת הפחתה (שיקום). ניתן להשתמש ביכולת של תאית לעבור תגובת הפחתה כדי לאפיין את אורך השרשראות שלה, האופייני ל המין הזהצמחים.
בשארית הגלוקוז נהוג להבחין במישורים הידרופיליים והידרופוביים, תלוי כמה קבוצות OH ו-CH מתאימות לו. זה חשוב כאשר לומדים את האינטראקציה של שייר גלוקוז עם שאריות חומצות אמינו ארומטיות במהלך הביקוע האנזימטי של מולקולת תאית במרכז הפעיל של האנזים.
בשל העובדה שקבוצות ההידרוקסיל באטום הגלוקוז הראשון והרביעי נבדלות במיקומן ביחס למישור הטבעת, מסתבר ששאריות הגלוקוז השכנות בשרשרת הפולימר מסובבות ב-180 0 זו לזו. שרשראות פולימרים מיוצבות על ידי קשרי מימן הנוצרים בין שיירי גלוקוז במהלך האינטראקציה של קבוצות הידרוקסיל ב-6,2,3 אטומי פחמן וחמצן טבעת. שרשראות פולימריות ליניאריות מחוברות זו לזו באמצעות קשרי מימן בין-מולקולריים הנוצרים בין אטומי חמצן של שרשרת אחת וקבוצות הידרוקסיל ב-C 6 של גלוקוז של שרשרת אחרת (איור 10). כתוצאה מאינטראקציות כאלה, נוצרים סיבים חזקים. הגמישות של מולקולת התאית נקבעת על ידי האפשרות של סיבוב של שאריות האנהידרוגלוקוז בקישור הצלוביוז ביחס לקשר הגליקוזידי. היציבות של הקונפורמציה הליניארית של תאית נקבעת, במיוחד, על ידי הדחייה המינימלית של C1-H ו-C4-H.
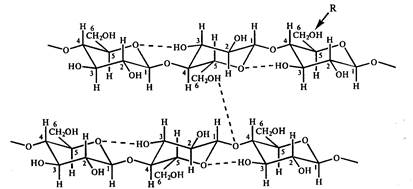
אורז. 10. ייצוג סכמטי של מערכת קשרי המימן בין שיירי גלוקוז בתאית. R הוא מרכז השינוי הכימי האפשרי של תאית. אז במתילצלולוזה, אצטילצלולוזה וקרבוקסיתילצלולוזה R \u003d CH 3, COCH 3, CH 2 COOHa, בהתאמה
מולקולת הגלוקוז מוצגת בקונפורמציה של הכיסא, שבה אטומי C2, C3, C5 ו-O5 נמצאים באותו מישור, והאטומים C1 ו-C4 נמצאים בצדדים מנוגדים שלו. שאריות הגלוקוז יכולות לקבל גם קונפורמציות פחות טובות מבחינה אנרגטית, שלפי מידת היציבות מסודרות לפי הסדר הבא: מבנה אלכסוני, מבנה סירה (או אמבטיה) ומבנה חצי כיסא הכי פחות נוח, שבו 5 אטומים של טבעת פירנוזה בת 6 איברים נופלים למישור אחד בבת אחת. היכולת של יחידת האנהידרוגלוקוז לקבל במולקולת התאית, בנוסף לקונפורמציות היציבות ביותר, הפחות יציבות (לדוגמה, מבנה של סירה או מבנה חצי כיסא) יש חֲשִׁיבוּתתוך הפרה של המבנה העל-מולקולרי המסודר של תאית, כמו גם בהידרוליזה אנזימטית של קשרים גליקוזידיים במולקולה שלו.
סיבים אלמנטריים (מיסלים) של תאית מורכבים מ-100-200 שרשראות פולימריות המחוברות באמצעות גשרי מימן. סיבים אלו בדופן התא מקבילים זה לזה ומקובצים בצורה כזו שנוצרות צרורות בצורת אליפסה. – מיקרופיברילים . כ-10 מיצלות יוצרים מיקרופיבריל בקוטר של 0.025 ננומטר (איור 11).
לכל צרור יש חלק גבישי מרכזי - הליבה, שבה ארוזות השרשראות בצורה כה הדוקה שאפילו מולקולות מים חודרות אליהן בקושי רב. מסביב לגרעין יש קליפת מוח פארא-גבישית עם מבנה פחות מסודר. אזורים פחות מסודרים אלה, הנקראים גם אזורים אמורפיים, מהווים בדרך כלל כ-15% ממבנה המיקרו של התאית. מיקרופיברילים יכולים להתלכד לתוך קבוצות בודדותאו שכבות – מקרופיברילים - עובי 0.4-0.5 מיקרון, נראים במיקרוסקופ אור. הקוטר ומידת הגבישיות של מיקרופיברילים משתנים מאוד בהתאם לסוג הצמח ולמצב הפיזיולוגי של התא.
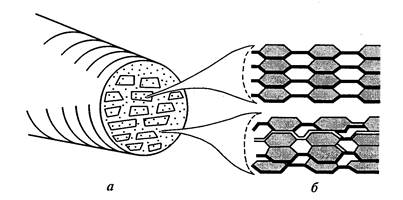
איור.11. דגם של מבנה מיקרו-פיבריל: א) חתך רוחב של מיקרו-פיבריל עם מיצלות ומרווחים בין-מי-תאיים; ב) פריסת תאית
עבור עיסת עץ עם אינדקס הזמנה של 50-70%, הממדים הרוחביים של הקריסטליט אינם עולים על 4x6 ננומטר. לפי מיקרוסקופ אלקטרונים, הממדים הממוצעים בקוטר יכולים להיות 1.5-2x2.5-3.5 ננומטר. בין 40 ל-80% מהמולקולות בגדלים כאלה ממוקמות על פני הגביש, שם תיתכן הידרציה גדולה יותר שלהן, הפרה של הסדר לטווח ארוך במבנה של קשרי מימן, וגם השפעה חזקה יותר של השפעות כימיות.
תאית טבעית היא polydisperse, מספר שאריות הגלוקוז במולקולת תאית, וכתוצאה מכך, אורך המולקולות עצמן יכול להשתנות באופן דרמטי ב קירות תאיםצמחים שונים. אז, עבור תאית שיער כותנה, מידת הפילמור של דופן התא הראשוני היא 2-6 אלף, המשנית - 13-14 אלף, עבור תאית Acetobacter xylinum - 2-6 אלף, עבור עץ - 8-10 אלף. המשקל המולקולרי המקסימלי שנקבע בניסוי של תאית הפשתן הוא 6,000,000. בשל העובדה שמידת הפילמור היא אינדיקטור ממוצע, המאפיין המדויק ביותר הוא עקומת ההתפלגות של מולקולות התאית לאורכה (התפלגות המשקל המולקולרית). עם זאת, מאפיין זה קשה להשגה ולכן נעשה שימוש במאפיינים ממוצעים: דרגות פילמור מספר ממוצע (DP n) וממוצע משקל (DP w). דרגת הפילמור הממוצעת במספר שווה למספר הכולל של שאריות אנהידרוגלוקוז בדגימת התאית חלקי מספר כוללמולקולות פולימר. כדי לקבוע זאת, הקצוות המצמצמים עוברים טיטרציה בדגימת הפוליסכריד. דרגת הפולימור הממוצעת במשקל של תאית נקבעת על ידי המסת דגימה של תאית בקומפלקס אלקליין של נחושת, אבץ, קובלט או קדמיום עם אתילן-דיאמין, או בקומפלקס של LiCl עם N,N'-dimethylacetamide. יחס DP w/DP נשווה לאחדות רק עבור פולימרים חד מפוזרים.
עבור עיסת עץ של אשוחית, אורן, ליבנה ואספן, דרגת הפילמור הממוצעת במשקל היא 12000, בעוד שאורכה של מולקולה כזו במצב המורחב מגיע לכמעט 6 מיקרון, שהם פי 6000 יותר מקוטרה. מולקולה כה ארוכה ובנויה באופן קבוע נערמת בצורה מסוימת. שיטת הערימה קובעת את המבנה העל-מולקולרי של הביופולימר, המבוסס על קשרי מימן תוך-מולקולריים הנוצרים על ידי יחידות צלוביוז. חקר שיטת הערימה וסוגי קשרי המימן במקרומולקולת התאית מתבצע באמצעות ניתוח קונפורמציה, דיפרקציה (פיזור קרני רנטגן בזווית קטנה, פיזור אלקטרונים), ספקטרוסקופיה IR ו-NMR. למרות התקדמות משמעותית בתחום זה, השאלה כיצד לערום מולקולות תאית לא נפתרה סופית.
תאית אינו מסיס במים ובממיסים אורגניים, אינרטי מבחינה כימית, חזק מאוד מבחינה מכנית. מים בין משטחים מרווחים, אך לא מקיימים אינטראקציה ישירה של גבישים, בנויים. הוצאת מי הידרציה, למשל, כתוצאה מייבוש, מקדמת התכנסות של מולקולות תאית שאינן מתקשרות בתחילה ויצירת סוגים שונים של קשרי מימן ביניהן, שאינם נהרסים במהלך ההידרציה. חוקרים מייחסים את העלייה במידת הגבישיות במהלך הרטיבות לחדירת מים מאזורים פחות מסודרים לאזורים מסודרים יותר. כתוצאה מכך, נצפית ירידה בגדלים האבסולוטיים של הגבישים ועלייה במידת הסדר הכללי. הוא האמין כי מים במקרה זה פועלים על פי העיקרון של "סיכה מולקולרית", מה שמקל על שברי מולקולות לא מסודרים להשלים את המבנה המסודר באמצעות אינטראקציה שיתופית עם "זרע" גבישי. החלפה של הקבוצות הקוטביות של תאית, למשל, ב-carboxymethyl, מובילה להפרה של הסדירות המולקולרית, מקדמת היווצרות של קשרי מימן רבים עם מים והופכת את התאית למסיסה.
העמידות של תאית להתפצלות, הן בתנאי טבע והן בתנאי מעבדה, נובעת לא כל כך מהמאפיינים של הקשר b-1,4-גלוקוזידי, אלא מהמבנה הגבישי של תאית ומה"אריזה" המיוחדת של המולקולות שלה. מבנים ביולוגיים.
אנזימים צלולוליטים אינם יכולים לחדור לתוך המבנים הפנימיים של סיבים תאית. הם פועלים על פני השטח שלהם. מספר הקשרים הגליקוזידיים הזמינים לפעולת אנזימים תלוי במידה רבה במידת ההתנפחות של תאית. ניתן להגיע לעלייה בדרגת הנפיחות באמצעות טיפול מקדים - אידוי, שחיקה, מכשירי קול וכו'. הנפיחות של כל הסיבים מוגברת על ידי חומצות מינרליות ואלקליות ב ריכוזים גבוהים, שכן הם מסוגלים לשבור קשרי מימן ולחדור לאזורים גבישיים. החלקים האמורפיים של תאית מושפעים בקלות יחסית מחומצות ואלקליות.
תחת פעולתן של תמיסות אלקליות מרוכזות על תאית, התכונות הפיזיקליות-כימיות והמבניות שלה משתנות: נפיחות עזה מורגשת ודרגת הגבישיות משתנה. חלק המשקל המולקולרי הגבוה של תאית, שאינו מסיס ב-17.5% NaOH, נקרא a-צלולוזה. כותנה מורכבת בעיקר מ-a-צלולוזה, בעוד שצמחים ועץ אחרים מכילים בעיקר b-צלולוזה, המסיסים בתמיסת אלקלית מרוכזת.
מיקרופיברילים טבולים במטריצת מעטפת אמורפית, שהיא תערובת מורכבת של פולימרים, שביניהם שולטים פוליסכרידים במשקלים מולקולריים שונים. ההנחה היא שעל פני כל המיקרופיברילים יש שכבה של מולקולות קסילוגלוקן הנספגות בקשרי מימן. ליגנין והמיצלולוזים ממלאים את החלל בין סיבים תאית אלמנטריים. לפיכך, החומר הליגני הוא מבנה פולימרי המורכב מחיזוק תאית הטבול במטריצה לינו-תאית.
תאית והמיצלולוז מאופיינים בקינטיקה שונה של היווצרות, מה שמצביע כנראה מקומות שוניםהסינתזה שלהם. נראה כי סינתזת תאית מתרחשת מחוץ לתא על פני השטח ממברנה ציטופלזמיתבהשתתפות GDF או UDF. התורמים של קבוצות גליקוזיל הם סוכר נוקלאוזיד דיפוספטים, הנוצרים מגלוקוז וסוכרוז. דרכים שונות. לדוגמה, מנוקלאוזיד טריפוספטים וסוכר-1-פוספטים בהשתתפות pyrophosphorylase. שאריות גליקוזיל מחוברות לשאריות סוכר בקצוות הלא-מצמצמים של השרשרת הגדלה. בין הפחמן הראשון של השייר "הנוסף" לקבוצת ההידרוקסיל של אטום הפחמן הרביעי של השייר הסופי, נוצר קשר גליקוזידי בהשתתפות הסינטטאז.
N (סוכר נוקלאוזיד דיפוספט) + (סוכר) n סינתטאז n (נוקלאוזיד דיפוספט) + (סוכר) n +1
בדופן התא נמצאו אנזימים של סינתזת תאית וגרגירים בקוטר 8 ננומטר, שהם, ככל הנראה, קומפלקסים מולטי-אנזימים הממוקמים על משטח חיצוניפלזמהלמה ובמטריקס הקיר. סינתזת תאית (E) בפלסמהלמה קשורה לחלבון המפעיל P, המסוגל לקשור ל-GTP ולעבור לצורה הפעילה עם היווצרות קומפלקס P-E-GTP.
בתהליך של סינתזת תאית נוצרות לא רק שרשראות תאית בודדות, אלא נוצרות גם מיקרו-סיבי תאית. סינתזה כזו מתבצעת ככל הנראה בהשתתפות מולקולות רבות של סינתזות תאית המרוכזות במקומות נפרדים של הפלזמה, וסינתזות תאית מורכבות מתת-יחידות רבות, שכל אחת מהן אחראית לסינתזה של שרשרת גלוקן אחת. עַל בְּתוֹךנמצאו ממברנות, "רוזטות" של שש כדוריות חלבון. רוזטות מופרשות לפלסמה דרך מערכת אנדוממברנית של שלפוחיות. בהתבסס על תכונות החיבור המבני של רוזטות, קומפלקסים סופניים ומיקרו-סיבי התאית שנוצרו, עלתה הנחה לגבי מנגנון היווצרותם של מיקרו-פיברילים במבנה טרנסממברני משלים: רוזטות - קומפלקס קצה - קצה גדל של המיקרו-פיבריל. קומפלקס טרמינלי מחובר לקצה המיקרופיבריל החדש שסונתז, ורוזטות הן אתר הקישור של מבשרי תאית והפרשת "נשאים" של שאריות גלוקוז.
רוזטות בודדות, יחד עם הקומפלקסים הסופיים המתאימים, אחראים לסינתזה של מיקרו-סיביים בעלי אוריינטציה אקראית של דופן התא הראשוני (איור 12), וקבוצה מכוונת של עד 175 רוזטות בחלקים נפרדים של הפלזמהלמה קובעת את היווצרותם של מקבילים. מיקרופיברילים של דופן התא המשני (איור 13).
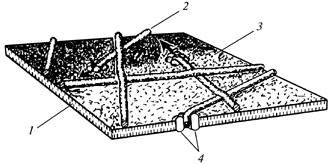
איור 12. מיקרו-סיבי תאית במהלך היווצרות
דופן תא ראשוני: 1
- שכבה כפולה של שומנים; 2
- מיקרופיבריל; 3
- פני השטח החיצוניים של הפלזמה; 4
- שקע
כל רוזטה יוצרת מיקרופיבריל בקוטר של 5 ננומטר. המרחק בין שורות רוזטות בממברנת הפלזמה מתאים למרחק בין מיקרו-סיביים בדופן התא. מיקרופיברילים דקים מכילים כ-50 שרשראות פוליגלוקניות. מיקרו-סיביים אלה יוצרים סיבים של דופן התא המשני, אשר קוטר יכול להיות עד 28 ננומטר. תאית, שלא כמו עמילן, היא תרכובת קשה לחילוף חומרים. העיכול שלו נקבע לפי המקור, התכולה בתזונה, אופי הטיפול המקדים ונע בין 6 ל-23% בממוצע. כן, עיכול תאית סובין חיטהבערך 15%.
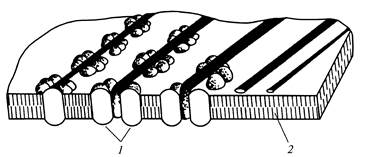
אורז. 13. מיקרו-סיבי תאית במהלך היווצרות
דופן תא משני: 1
- שקע; 2
- שכבה כפולה ליפידים
IN סיבים תזונתייםתאית מהווה כשליש. הפירוק היעיל שלו מתרחש כתוצאה מאינטראקציה סימביוטית של מיקרואורגניזמים שונים של המעי הגס והאנזימים הצלולוליטיים שלהם. IN מערכת עיכולתאית מגרה את פעילות המעי, מגבירה את הפריסטלטיקה שלו, מנרמלת את הפעילות מיקרופלורה של המעיים, סופג סטרולים, מונע את ספיגתם ומקדם את שחרור הכולסטרול. תאית מיקרו-גבישית (MCC), אבקה לא-סיבית של תאית טבעית, מצאה שימוש נרחב כתוסף מזון. הוא לא רק מפחית את תכולת הקלוריות של המזון, אלא הוא גם מעבה ומפזר שלו, מה שמשפר את איכות המזון.
לחיידקים גראם-שליליים יש מבנה דופן תא מורכב הרבה יותר מאשר לגראם-חיובי. הוא מכיל מספר גדול בהרבה של מקרומולקולות שונות סוג כימי. פפטידוגליקן יוצר רק את השכבה הפנימית של דופן התא, צמודה באופן רופף ל-CPM. ל סוגים שוניםאובקטריות גראם שליליות, התוכן של ההטרופולימר הזה משתנה מאוד. ברוב המינים הוא יוצר מבנה חד או דו-שכבתי, המאופיין בקשרים צולבים נדירים מאוד בין שרשראות הטרופולימרים.
כמה חיידקים גולשים (myxobacteria, flexibacteria) מסוגלים לשנות מעת לעת את צורת התאים בתהליך של תנועה לאורך מצע מוצק, למשל על ידי כיפוף, דבר המעיד על גמישות דופן התא שלהם, ובעיקר שכבת הפפטידוגליקן שלו. עם זאת, בדיקה מיקרוסקופית אלקטרונית גילתה שיש להם דופן תא אופייני לאובקטריה גראם-שלילית. ההסבר הסביר ביותר לגמישות דופן התא של חיידקים אלו הוא הצלבות נמוכה ביותר של מרכיב הפפטידוגליקן שלו.
לפרוקריוטים גראם שליליים יש קרום חיצוני, הכולל שומנים (22%), חלבונים, פוליסכרידים, ליפופרוטאינים.
ליפופוליסכרידים (LPS) הם הטרופולימרים בעלי מבנה מורכב בעלי מגוון פעילויות ביולוגיות. הקומפלקס הליפואידי גורם לרעילות (תגובות דלקתיות, חום, הלם אנדוטוקסין), מרכיב הפוליסכריד אחראי לסגוליות O-antigen. LPS משרה סינתזה של נוגדני Jg M ומשמשת באימונולוגיה כגורם אדג'ובנטי ומפעיל פוליקלונלי של תאי B.
דופן התא בחיידקים מבצעת בעיקר פונקציות עיצוב והגנה, מספקת קשיחות, יוצרת קפסולה וקובעת את יכולת התאים לספוח פאג'ים.
№15 צורות לא רצופות של חיידקים, פרוטופלסטים, ספירופלסטים, צורות L.
protoplasts, spheroplasts, צורות L הם צורות של חיידקים ללא KS
צורות L יכולות להתרחש בתנאים טבעיים בגוף האדם כתוצאה מטיפול ארוך טווח באנטיביוטיקה מסוימת, לרוב פניצילין.
ישנן צורות L לא יציבות ויציבות של חיידקים. הראשונים מסוגלים לחזור לצורתם המקורית כאשר הגורם שגרם להיווצרותם מתבטל. הם משחזרים את היכולת לסנתז פפטידוגליקן CS. אלה האחרונים, ככלל, אינם מסוגלים לחזור לאחור. צורות L של חיידקים שונים ממלאות תפקיד חיוני בפתוגנזה של מחלות זיהומיות רבות.
המאפיינים העיקריים של צורות L של חיידקים:
טרנספורמציה מתמדת מ-grampol-s ל- grammatrits-e.
שינוי בתכונות האנטיגניות
ירידה בארסיות
יכולת התמדה לאורך זמן
היכולת לחזור לצורתו המקורית במקרה של אובדן לא שלם של הסינצה של ה-COP.
№16 תכונות טינקטואליות של חיידקים. שיטות צביעה מתוחכמות. עקרון צביעה גראם של חיידקים.
טכניקת כתמי גראם
נייר סינון מונח על המריחה ויוצקים תמיסה קרבולית של סיגלית ג'נטיאן למשך 1-2 דקות.
מסירים את הנייר, מסננים את הצבע ובלי לשטוף את המריחה במים, יוצקים את התמיסה של לוגול למשך דקה.
תמיסת Lugol מרוקן והתכשיר מוחלש באלכוהול 96 0 למשך 30 שניות.
נשטף במים.
צבע 1-2 דקות תמיסה מימיתמַגֶנטָה.
עקרון הכיתור לפי גרם
גרם חיוביחיידקים שומרים על סגול ג'נטיאן בשילוב עם יוד - צביעה סגולהבַּקטֶרִיָה;
גראם שליליחיידקים לאחר חשיפה לאלכוהול מאבדים את הצבע, משתבחים וכאשר מטופלים בפוקסין, מוכתמים ב צבע אדום.
שיטות מורכבותכתמים משמשים לחקר מבנה התא והתמיינות של מיקרואורגניזמים. מריחות מוכתמות עוברות מיקרוסקופ במערכת טבילה. יש למרוח באופן עקבי על התכשיר צבעים מסוימים הנבדלים זה מזה בהרכב הכימי ובצבעם, חומרי חומרי מור, אלכוהולים, חומצות וכו'.
תכונות טינטקטוריותחיידקים (lat. tinctura, מטינגו - I color) יכולת הצבע: רגישות לצבע, עמידות לחומצה-אלכוהול-אלקלי, אחידות צבע, מטכרומטיות, יחס לצבע בשיטת גראם.
№17 תכונות של ההרכב הכימי של דופן התא של חיידקים עמידים לחומצה. עקרון הצביעה לפי שיטת Ziehl-Neelsen.
חיידקים עמידים לחומצה. דופן התא של חיידקים מסוימים מכילה כמות גדולה של שומנים ושעווה, מה שהופך אותם לעמידים בפני שינוי צבע לאחר צביעה בחומצות, אלקליות או אתנול (לדוגמה, מיני Mycobacterium או Nocardia). חיידקים כאלה נקראים מהירי חומצה וקשים להכתמתם (אם כי חיידקים מהירי חומצה נחשבים לחיובי גראם). לצביעה שלהם משתמשים בשיטת Ziehl-Neelsen.
מיקובקטריה ונוקרדיה מאופיינים במבנה מסובך של דופן התא. הם, כמו חיידקים גראם חיוביים, מבוססים על מסגרת Murein, אך האחרונה קשורה לפוליסכרידים ולשומנים. ליפידים מיוצגים על ידי חומצות מיקוליות, המעניקות למשטח התא הידרופוביות. הידרופוביות, מצד אחד, הופכת את התא לעמיד בפני פעולת כימיקלים שונים (חיידקים כאלה נקראים עמידים לחומצה), מצד שני, היא מעכבת את חילופי התא עם הסביבה ומאטה את צמיחתו. לכן, פעילי שטח מתווספים למדיה תזונתית לטיפוח Mycobacterium tuberculosis. עמידות החומצה של mycobacteria היא תכונה אבחנתית חשובה; לקביעתה, נעשה שימוש בכתם Ziehl-Neelsen.
טכניקת צביעה לחיידקים מהירי חומצה
לפי שיטת Ziehl-Neelsen
נייר סינון מונח על מריחה קבועה ויוצקים פוקסין קרבולי של Ziehl ומחממים בעדינות על מבער עד להופעת אדים. הפעולה חוזרת על עצמה 2-3 פעמים.
כשהתרופה התקררה, הסר את נייר הסינון, רוקן את הצבע ושטוף את התרופה במים.
התרופה נטבלה 2-3 פעמים בכוס עם חומצה גופרתית 5% למשך 1-2 שניות.
התכשיר נשטף היטב במים ומוכתם במתילן כחול אלקליין למשך 3-5 דקות.
נשטף במים ומיובש.
חיידקים עמידים לחומצה אינם מתעוותים עם חומצה גופרתית ושומרים על צבעם האדום, חיידקים שאינם עמידים לחומצה מאבדים את הצבע ונצבעים בכחול עם מתילן כחול.
№18 תכלילים של חיידקים, הרכבם ותפקידם הביולוגי. בציטופלזמה יש תכלילים שונים בצורת גרגירי גליקוגן, פוליסכרידים, חומצה בטא-הידרוקסי-בוטיר ופוליפוספטים (וולוטין). הם חומרי מילואים לצורכי התזונה והאנרגיה של חיידקים. לווליוטין יש זיקה לצבעים בסיסיים והוא מזוהה בקלות באמצעות שיטות מיוחדותצביעה (לדוגמה, לפי נייסר) בצורה של גרגירים מטאכרומטיים. הסידור האופייני של גרגירי וולוטין מתגלה בבצילוס דיפתריה בצורה של קטבים מוכתמים בעוצמה של התא. (לפי נייסר הוא בצבע כחול כהה)
№19 נבגי חיידקים, מבנהם חיידקים מסוימים בתום תקופת הצמיחה הפעילה מסוגלים ליצור נבגים. לפני זה דלדול הסביבה עם חומרים מזינים, שינוי ב-pH שלה והצטברות של מוצרים מטבוליים רעילים. ככלל, תא חיידק אחד יוצר נבג אחד - לוקליזציה של נבגים שונה (מרכזית, סופנית, תת-טרמינלית)
אם גודל הנבגים אינו עולה על הגודל הרוחבי של חיידק בצורת מוט, אז האחרון נקרא בזיל (גורם סיבתי). גַחֶלֶת). כאשר קוטר הנבגים גדול יותר, החיידקים הם בצורת ציר ונקראים קלוסטרידיום (גורמים הגורמים לזיהום אנאירובי). לטטנוס קלוסטרידיה יש נבגים עגולים ומזכירים מקלות תיפוף. Clostridia botulinum מאופיינים בנבגים סגלגלים גדולים, המעניקים להם מראה של מחבט טניס.
לפי ההרכב הכימי, ההבדל בין נבגים לתאים וגטטיביים הוא רק בתכולה הכמותית של תרכובות כימיות. נבגים מכילים פחות מים ויותר שומנים.
היווצרות נבגים קשורה לדחיסה ובידוד של אזור מסוים בציטופלזמה של תא וגטטיבי, ואחריו היווצרות גוף עגול או סגלגל בתוך החיידק, המכוסה בקרום רב-שכבתי צפוף, הרווי בגוף. כמות גדולה של שומנים, סידן וחומצה דיפיקולינית (איור 12).
פעם בגוף של בני אדם ובעלי חיים, נבגים נובטים לתאים וגטטיביים. תהליך נביטת הנבגים כולל שלושה שלבים: הפעלה, שלב ראשוניושלבי גדילה. הסוכנים המפעילים המפריעים למצב הרדום כוללים טמפרטורה גבוהה, תגובה חומצית של הסביבה, נזק מכניואחרים.הנבג מתחיל לספוג מים, משתחרר מסידן דיפיקולט, ומשמיד רבים מהמרכיבים המבניים שלו בעזרת אנזימים הידרוליטים. לאחר הרס השכבות החיצוניות, מתחילה תקופת היווצרות של תא וגטטיבי עם הפעלת הביוסינתזה, המסתיימת בחלוקת התא.
שיטת צביעת נבגים לפי Ozheshko
הנבגים מחזיקים בחוזקה פוקסין קרבולי והם נצבעים באדום, הציטופלזמה של החיידקים מוחלשת עם חומצה גופרתית 5% ולאחר צביעה בכחול מתילן, הופכת לכחול.
№20 כמוסה ומיקרוקפסולה של חיידקים קפסולה היא שכבה רירית של דופן התא של חיידקים, המורכבת מפוליסכרידים (פנאומוקוק) או פוליפפטידים (אנתרקס באצילוס). מיקרוקפסולה (עובי פחות מ-0.2 מיקרומטר) יכולה להיווצר על ידי רוב החיידקים, מקפסולה מוגדרת היטב (עובי של יותר מ-0.2 מיקרומטר) נוצרת על ידי פנאומוקוקוס, קלבסיאלה, אנתרקס ועוד כמה. בחיידקים פתוגניים, הקפסולה נוצרת במקרואורגניזם; על חומרי הזנה מלאכותיים היא בדרך כלל אובדת (למעט Klebsiella).
בבני אדם ובבעלי חיים, הקפסולה מגנה חיידקים פתוגנייםמ bacteriophage, phagocytosis וגורמי חסינות הומורלית, קובע את הספציפיות האנטיגנית של מיקרואורגניזמים.
כמוסות, בעלות עקביות ג'ל, אינן שומרות היטב על הצבע, ולרוב משתמשים בשיטות ניגודיות שליליות כדי לזהות אותן.
שיטת זיהוי הקפסולות של Burri-Gins
חיידקים מוכתמים באדום, כמוסות לא מוכתמות מנוגדות על הרקע הכהה של התרופה.
№21 דגלים חיידקיים פלאג'לה פועלים כאיבר תנועה המאפשר לחיידקים לנוע במהירות של 20-60 מיקרון לשנייה. לחיידקים יכולים להיות דגלים אחת (מונוטריכית) או מספר דגלים, הממוקמים על פני כל פני הגוף (פריטריכיים), או נאספים בצרורות (לופוטרי).
המיקום הפריטריכי של הפלגלה אופייני לאנטרובקטריות, פתוגנים של זיהומים אנאירוביים, טטנוס, בוטוליזם; Vibrio cholerae הוא מונוטריך ופסאודומונס הוא לופוטריק. לכמה מינים של ספירילה יש סידור אמפיטרי של דגלים. עובי הפגללה בממוצע 10-30 ננומטר, והאורך מגיע ל-10-20 מיקרון.
הבסיס של הדגלון הוא חוט ספירלי ארוך (פיבריל), אשר על פני דופן התא עובר למבנה מעוקל מעובה - קרס ומוצמד לגרגיר הבסיסי המוטבע בדופן התא וב-CPM (איור 10). .
הקוטר של הגרגירים הבסיסיים הוא כ-40 ננומטר והם מורכבים ממספר טבעות (זוג אחד בחיידקים גראם חיוביים, ארבעה בפרוקריוטים שליליים גראם). הסרת שכבת הפפטידוגליקן של דופן התא מובילה לאובדן יכולת התנועה של החיידקים, למרות שהדגלים נשארים שלמים.
הדגלים מורכבים כמעט לחלוטין מהחלבון פלגלין, עם תכולת פחמימות ו-RNA.
במיקרוסקופ ניתן לראות דגלים רק לאחר שיטות מיוחדות של תחריט והספגה במלחי כסף וכספית, ולאחר מכן צביעה בכחול מתילן (שיטת לפלר). יש לזכור שהדגלים רגישים מאוד להשפעות מכניות שונות. ניתן לשפוט בעקיפין את הנוכחות של דגלים לפי האופי הכיווני של התנועה בצניחה ה"תלויה" וה"מעוכה" במיקרוסקופים של שדה כהה וניגודיות פאזה, או במיקרוסקופ של שדה בהיר כשהמעבה מונמך ודיאפרגמת המיקרוסקופ חלקית. סמוי.
צביעת פלאגלום בשיטת לפלר
תחת מיקרוסקופיה מוצר מוגמרהדגלים נראים כמבנים דמויי חוט דקים.
#22 Pili פני השטח של enterobacteria וכמה מיקרואורגניזמים אחרים מכוסים מספר גדול(מ-10 עד כמה אלפים) villi - תצורות חוטיות בעלות אופי חלבוני. כמו פלגלה, הם בנויים מסוג אחד של חלבון - פילין, שתת-היחידות שלו מאורגנות בצורת חוט חלול בפנים ומקורן ב-CPM. הם קצרים ודקים יותר מהדגלים, רוחבם 10-12 ננומטר ואורכם עד 12 מיקרומטר.
Villi הם רב-פונקציונליים: הם מספקים העברה של גנים (צימוד), הם קולטני פאג, איבר לחיידקים להיצמד למצע מזין (הידבקות), ומשתתפים בהובלת מטבוליטים.
לסטרפטוקוקים יש שכבה חיצונית של שערות חלבון (פימבריה), אשר נקראות חלבון M (חלבון M). הסנאי הזה משחק תפקיד חשובבתהליכי האינטראקציה של חיידקים עם המאקרואורגניזם.
№23 Actinomycetes
הם מייצגים קבוצה מוזרה של חיידקים שנראים כמו חוטים מסועפים קטנים או ארוכים. הצטברות ההיפיות נקראת תפטיר. הדמיון לפטריות הוא חיצוני בלבד, שכן לאקטינומיציטים יש סוג תאים פרוקריוטי עם דופן תא שאינה מכילה כיטין ותאית. אקטינומיציטים הם גרם חיוביים, צורות רבות עמידות לחומצות, לחלק מהאקטינומיציטים יש קפסולה סביב החוטים.
תפטיר המצע נוצר כתוצאה מגידול התפטיר לתוך המצע התזונתי והאוויר, הגדל על פני המדיום (איור 19). ברקמות המושפעות (צורת רקמה), האקטינומיציטים יכולים ליצור גרגירי דרוזן, מחוטים שזורים בצפיפות בצורת קרניים הנמשכים מהמרכז ומסתיימים בעוביים בצורת צלוחית.
Actinomycetes מתרבים באופן א-מיני, יוצרים קונידיות או ספורופורים עם ספורנגיה בקצוות התפטיר האווירי. נושאי נבגים יכולים להיות ישרים, גליים, ספירליים. נבגים – סגלגלים, עגולים, גליליים, בעלי משטח חלק או קוצים, לעיתים ניידים עקב דגלים (זואספורים). הנבגים משמשים להתרבות אקטינומיציטים, הם אינם עמידים בחום, אך יכולים לעמוד בפני ייבוש. בנוסף, ניצנים ופירוק של התפטיר לצורות מוטות או קוקיות אפשריים.
Actinomycetes מופצים באופן נרחב בטבע, הם חיים במים, אדמה עשירה בחומוס. הם משתתפים במעגל החומרים בטבע. סוגים נפרדים actinomycetes משמשים כיצרנים של אנטיביוטיקה, ויטמינים, ליפידים, פרוטאזות, חומצות אמינו, סטרואידים.
Actinomycetes שייכים לסדר Actinomycetales, הכולל את המשפחות: Actinomycetaceae, Nocardiaceae, Streptomycetaceae, Mycobacteriaceae.
מינים פתוגניים לבני אדם נמצאים בקרב נציגי המשפחות Actinomycetaceae ו- Nocardiaceae. הראשונים נראים כמו מקלות מסועפים ארוכים או קצרים שאינם יוצרים תפטיר אווירי. הם הגורמים לאקטינומיקוזיס אנושיים ויוצרים דרוזן ברקמות המושפעות.
נציגים של משפחת Nocardiaceae דומים ל-mycobacteria, בעלי תאים חוטיים ויוצרים תפטיר אווירי ומצע על גבי חומרי הזנה. ההיפיות מפוצלות לתאים בצורת קוקואידים ותאים בצורת מוט. נוקרדיה פתוגנית גורמת לנוקרדיוזיס.
שיטות מחקר. מוכתם לפי גראם וזיהל-נלסן. הדרוזה מוסרת מהחומר הפתולוגי עם לולאה, מניחים בטיפת מים על שקף זכוכית, לוחצים קלות כלפי מטה עם כיסוי כיסוי, ואז מכניסים טיפה של תמיסה אלקלית של מתילן כחול מתחת לזכוכית ומעבירים מיקרוסקופ, ניגודיות פאזה. יכול לשמש.
№24 ספירושטים
חיידקים מפותלים עם תנועתיות פעילה. מידות הספירושטים נעות בעובי בין 0.1-0.3 מיקרון, באורך בין 7-500 מיקרון. תנועות שונות - מסליל ועד כפיפה. בדיקה מיקרוסקופית אלקטרונית אפשרה להבחין בספירושטים גליל פרוטופלזמי (גוף התא), חוט צירי (תומך) וקליפה חיצונית תלת-שכבתית. החוט הצירי ממוקם בחלל הפריפלסמי שבין המעטפת החיצונית לגליל הפרוטופלסמי ומורכב מסיבים בודדים (אנדופלגלות), שמספרם משתנה במינים שונים: בטרפונמה ולפטוספירה - 3-4; בבורליה - עד 30. כל אחד מהסיבים (endoflagella) מקובע באזור דיסקיות ההתקשרות בקצות הגליל הפרוטופלזמי ונמתח לקצה הנגדי שלו, עוטף אותו ומסתיים בחופשיות. ההרכב הכימי של סיבים דומה לזה של דגלים (איור 20).
הגליל הפרוטופלזמי מכיל: נוקלואיד, ריבוזומים, מזוזומים, תכלילים. המעטפת החיצונית (דופן התא) מכילה שכבה דקה של פפטידוגליקן, אלסטית ואין לה קשיחות. חיידקים אלו אינם יוצרים אנדוספורות, קפסולות ואקספליגלטים, הם גרם שליליים, הם מסודרים באופן אקראי במריחה.
ה-Spirochaetes שייכים לסדר Spirochaetales, משפחת Spirochaetaceae, הכוללת שלושה סוגים:
בורליה - בעלת 3-10 תלתלים משופעים לא אחידים, הקצוות מחודדים, באורך 10-30 מיקרון. התנועה קופצנית, לדברי רומנובסקי-גימסה, הם צבועים בכחול סָגוֹל(נציג Borreliarecurrentis - גורם לקדחת חוזרת מגיפה; Borreliaburgdorferi - גורם למחלת ליים).
Treponema - בעל 8-14 תלתלים מעוותים בחוזקה של משרעת שווה, אורך 5-15 מיקרון. התנועה חלקה, איטית עם סיבוב סביב ציר האורך, לפי רומנובסקי-גימסה, הם צבועים בחיוור צבע ורוד(נציג של Treponemapallidum - הגורם הסיבתי של עגבת).
לפטוספירה - יש עד שני תריסר תלתלים תכופים קטנים המסתיימים בקרס עם עיבוי דמוי כפתור, באורך 5-15 מיקרון. התנועה היא מאוד פעילה, תנועה מתרגלת קדימה, כיפוף וסיבוב סביב הציר. לדברי רומנובסקי-גימסה, הם מכתימים בצורה חלשה בצבע ורדרד-לילך (נציג של Leptospirainterrogans הוא הגורם הסיבתי של לפטוספירוזיס).
שיטות מחקר. במצב חי, ספירושטים נחקרים במיקרוסקופ ניגודיות פאזה ובמיקרוסקופ שדה אפל, תוך התבוננות בחומר הפעיל תנועה אופיינית spirochetes, תכונות של צורתם.
מכינים את ההכנות לפי בורי (חוטים מכווצים בהירים של ספירוצ'טים נראים על רקע כהה של התכשיר), מוכתמים לפי רומנובסקי-גימסה, לפי שיטת מורוזוב.
#25 ריקטסיה
מעמד "אלפאפרוטובקטריה"
להזמיןריקטסיאלס
מִשׁפָּחָה Rickettsiaceae
סוג ריקטסיה
מִשׁפָּחָה Ehrlichiaceae
ריקטסיה מגוונות בצורתן ומחולקות לסוגים הבאים:
coccoid חד גרגר (עד 0.5 מיקרון);
דו-גרגירי בצורת מוט (1-1.5 מיקרון);
בסיס שלושה-ארבעה גרגירים (3-4 מיקרון);
פיליפורמי רב גרגירי (10-40 מיקרון).
גרגרים (נוקלאופרוטאין) מתגלים על ידי צביעה לפי רומנובסקי-גימסה. כל הצורות הן הדדיות. מבחינה מבנית, יש להם את כל המרכיבים של תא חיידקי: דופן התא, קפסולת ליפואידים, ציטופלזמה, נוקלואיד, ריבוזומים, פילי. ריקטסיה מכילה גם DNA וגם RNA, יש תוכן גבוהפוספוליפידים, תכולת פחמימות נמוכה.
ברוב המקרים (למעט המין Rochalimaeaguintana), ריקטסיה אינה גדלה על חומרי הזנה מלאכותיים. מחזור החיים של ריקטסיה תלוי בפעילות החיונית של התא המארח ומורכב משני שלבים: וגטטיבי ומנוחה (גופים יסודיים). ריקטסי הנמצאים בשלב הווגטטיבי (איור 23) מתרבים באופן פעיל בביקוע בינארי ויש להם ניידות פעילה, ככל הנראה בשל מבנים דגלניים. ריקטזיה של שלב המנוחה (גופים יסודיים) הם כדוריים ולא פעילים.
שיטות מחקר.ריקטסיה מוכתמת היטב לפי רומנובסקי-גימסה בצבע לילך, לפי מורוזוב (בשיטת הכסוף) בשחור. כדי להבדיל בין ריקטסיה, שיטת הצביעה המוצעת על ידי P.F. זדרודובסקי:
ריקטסיה מכתים אודם באדום ומתגלים בקלות על רקע הציטופלזמה הכחולה וגרעין התא הכחול.
№26 כלמידיה
מעמדכלמידיה
להזמיןכלמידיאלס
פמיליה Chlamydiaceae
סוג: כלמידיה
1). גופים יסודיים הם מבנים כדוריים קטנים (0.2-0.5 מיקרומטר) צפופים באלקטרונים, נטולי פעילות מטבוליטים, בעלי נוקלואיד קומפקטי ודופן תא נוקשה, המסוננות באמצעות מסנני חיידקים. הם ההתחלה המדבקת של כלמידיה ומבטיחים את הישרדותם בסביבה החוץ-תאית וזיהום של תאים חדשים.
2). גופים רשתיים גדולים יותר (0.8-1.5 מיקרון), תצורות כדוריות בעלות מבנה רשת עם דופן תא דק ונוקלואיד פיברילרי. הם גדלים מגופים אלמנטריים בתוך תאים, נטולי זיהומים, ובעת חלוקה מבטיחים רבייה של כלמידיה. מכאן שם פרטי אחר, היסטורית של גופים רשתיים - "גוף ראשוני". גופים רשתיים הם הצורה הווגטטיבית של כלמידיה.
3). גופי ביניים הם שלב ביניים בין גופים אלמנטריים לרשתיים.
מחזור החיים של הכלמידיה מתחיל בעובדה שגופי יסוד עוברים פגוציטוזה על ידי התא המאכסן, ולאחר מכן תוך מספר שעות הם מתארגנים מחדש, גדלים בגודלם והופכים לצורות רשתיות שמתרבות בחלוקה רוחבית. מחזור החיים מסתיים כאשר צורות הביניים המתעוררות נדחסות, מצטמצמות בגודלן והופכות לגופים יסודיים. כלמידיה מתרבות בתוך ואקוולים ציטופלזמיים, ויוצרות מיקרוקולוניות (תכלילים) מוקפות בקרום. כל שלושת השלבים של התפתחות הכלמידיה נמצאים בהרכב המיקרוקולוניות. לאחר קריעת דופן ה-vacuole (שלפוחית) והממברנה של התא המאכסן, הכלמידיה החדשות שנוצרו משתחררות, והגופים היסודיים, מדביקים תאים אחרים, חוזרים על מחזור ההתפתחות. בתנאי גדילה אופטימליים בתאים איקריוטיים מעגל החייםכלמידיה היא 17-40 שעות (איור 24).
המוזרות של כלמידיה מתבטאת גם במבנה דופן התא שלהם. הוא נטול פפטידוגליקן והוא קרום דו-שכבתי, שקשיחותו נקבעת על ידי פפטידים המצולבים עם גשרים דיסולפידים. אחרת, הכלמידיה דומים לחיידקים גראם שליליים, מכיוון שהם מכילים גליקוליפידים הדומים לליפופוליסכרידים.
הסדר כלמידיה כולל משפחה אחת, כלמידיה, עם סוג יחיד, כלמידיה. עבור בני אדם, מינים פתוגניים הם C.trachomatis, C.psittaci, C.pneumoniae. כלמידיה גורמת לעין, לנשימה ול מערכות אורוגניטליותולהתאחד מתחת שם נפוץ"כלמידיה".
שיטות מחקר.לזיהוי מיקרוסקופי של גופי הכללה (מיקרוקולוניות) של כלמידיה בתאים נגועים (רקמות), נעשה שימוש בשיטות צביעה שונות: רומנובסקי-גימסה, מאצ'יאבלו ואחרות. כאשר מוכתמים על פי רומנובסקי-גימסה, הם מקבלים צבע כחול או סגול. בנוסף, כלמידיה נראות בבירור במצב לא מוכתם כאשר מיקרוסקופיה של תכשירים רטובים מתחת לזכוכית באמצעות מערכת אופטית ניגודיות פאזה. IN לָאַחֲרוֹנָההתגובה האימונופלואורסצנטית הישירה הנפוצה ביותר, כתם תפוז אקרידין.
№27 Mycoplasmas מעמדרכיכות
להזמין Mycoplasmatales
מִשׁפָּחָה Mycoplasmataceae
סוג Mycoplasma
סוג Ureaplasma
חמישה מינים הם פתוגניים לבני אדם: M . דלקת ריאות , M . הומיניס , M . איברי המין , M . גלישה בסתר ו U . urealyticum .
מיקופלזמות- הפרוקריוטים הקטנים ביותר (125-150 ננומטר) המסוגלים להתרבות באופן עצמאי. הוא האמין כי mycoplasmas הם הצאצאים הקרובים ביותר של התאים הפרוקריוטים המקוריים. הגנום של מיקופלזמות הוא מינימלי עבור תא, הוא קטן פי חמישה מהגנום coliוהוא 0.45 MD. המאפיין העיקרי של mycoplasmas הוא היעדר דופן התא. הם מוקפים בשכבה דמוית קפסולה, שמתחתיה יש רק קרום תלת-שכבתי דק בעובי 7.5-10 ננומטר, המכיל כמות משמעותית של כולסטרול. כתוצאה מכך, mycoplasmas מבודדים למחלקה מיוחדת Tenericutes, בכיתה Mollicutes ("עור עדין"), סדר Mycoplasmatales.
בשל היעדר דופן תא, מיקופלזמות (איור 21) הן רגישות אוסמוטיות ובעלות מגוון צורות:
א) תאים כדוריים או ביציים קטנים בגודל 0.2 מיקרומטר (גופים יסודיים) המסוננים באמצעות מסנני חיידקים;
ב) כדורי גדול יותר, בגודל של עד 1.5 מיקרון;
ג) תאים חוטיים, מסועפים בגודל של עד 150 מיקרון.
Mycoplasmas לא יוצרות נבגים, flagella, כמה מינים יש ניידות גלישה.
הם מתרבים על ידי ביקוע בינארי של תאים כדוריים וחוטים, ניצנים ושחרור של גופים יסודיים רבים שנוצרו בחוטים.
באשר לאנרגיה, מיקופלזמות משיגות אותה בדרך הרגילה עבור אנאירובים פקולטטיביים, על ידי תסיסה של פחמימות או חומצות אמינו. בשל הגנום הקטן של מיקופלזמות, יש להם יכולות ביוסינתטיות מוגבלות, ויש לגדל אותם על מדיה תזונתית המועשרת בשומנים, חלבונים, מבשרי חומצות גרעין. הם גדלים לאט, מושבות עם מרכז צפוף צומחות לתוך המדיום, הדומות ל"ביצה מטוגנת" (מרכז כהה ופריפריה פתוחה בהירה יותר). גודל המושבות קטן, אינו עולה על 600 מיקרון.
בפתולוגיה אנושית, מספר נציגים של הסוג Mycoplasma ממלאים את התפקיד הגדול ביותר: M.pneumoniae, M.hominis, M.anthritidis והמין היחיד של הסוג Ureaplasma–U.urealyticum (שנקרא כך בגלל פעילות urease). מיקופלזמות פתוגניות גורמות למחלות (מיקופלסמוזיס) של דרכי הנשימה, אורוגניטל ומפרקים עם מגוון ביטויים קליניים. כאשר מטפלים במחלות אלו, יש לזכור כי מיקופלזמות אינן רגישות לאנטיביוטיקה בטא-לקטם ולתרופות אחרות המעכבות את הסינתזה של דופן התא (בשל היעדרו בפתוגן).
שיטות מחקר. במיקרוסקופ אור נמצאות רק הצורות הגדולות ביותר של מיקופלזמות. במצב חי, הם נחקרים במיקרוסקופ של שדה כהה וניגודיות פאזה, רכיבים אולטרה-סטרוקטורליים מתגלים באמצעות מיקרוסקופ אלקטרונים.
№28 פטריות
כיתות (7): · Basidiomycetes(פטריות כובע). · זיגומיציטים (סוּג Mucor - מוקורמיקוזיס של בני אדם ובעלי חיים) Ascomycetes (חיות כיס, סוגים אספרגילוס , פניציליום , פטריות שמרים) Deuteromycetes - פטריות לא מושלמות, אינן מתרבות מינית ( קִמָחוֹן )
פטריות הן קבוצה גדולה ומתפתחת בהצלחה של אורגניזמים, הכוללת כ-80,000 מינים מזוהים. הגדלים שלהם נעים בין שמרים חד-תאיים ועד צלבים גדולים, פחזניות וקרניים. פטריות תופסות מגוון בתי גידול - הן במים והן ביבשה. בנוסף, הם חשובים בהקשר לתפקיד שהם ממלאים בביוספרה, ובקשר לעובדה שהם משמשים את בני האדם ברפואה ובכלכלה.
שיטות מחקר. גם תכשירים מקומיים (לא מוכתמים) וגם תכשירים מוכתמים מוכנים לבדיקה מיקרוסקופית.
מחקר של תכשירים לא מוכתמים
על מנת להבחין בצורה ברורה יותר בין מרכיבי הפטרייה, התכשיר מובהר, לשם כך מניחים את החומר הפתולוגי (קרום, פיסות ציפורניים, שיער, שריטות מהריריות, תוכן של מוקדים גרנולומטיים). זכוכית שעוןאו צלחת פטרי, שבה יוצקים תמיסה של 10-15% של נתרן קאוסטי או אשלגן ומניחים בתרמוסטט ב-37 0 C למשך 20-30 דקות. לאחר מכן מוציאים את החומר ומניחים אותו בטיפה של תמיסת גליצרול 50% על שקף זכוכית ומכסים אותו בכיסוי כיסוי, מיקרוסקופית במיקרוסקופ ניגודיות פאזה או אור. ניתן להשתמש בשיטה נוספת: טיפת גליצרין בתוספת 10% אשלגן קאוסטי נמרח על החומר הפתולוגי ונבדק לאחר 4-5 דקות, מכוסה בכיסוי כיסוי.
מוגלה ממורסות, תכולת כיבים, כיח מדוללים עם מי מלח פיזיולוגי או מים-אלכוהול (1:1) או תמיסה מימית של גליצרול 50%, מכינים תכשיר "טיפה מרוסקת" ונבדקים בהגדלה של x200, x400 באמצעות ניגודיות פאזה.
מחקר של תכשירים מוכתמים
מוגלה, דם, נוזל מוחי, משקעים של שטיפות סימפונות ושתן, מכינים מריחות דקות, המתוקנות בתערובת של Nikiforov, Carnoy, אלכוהול-פורמול, מיובשים ומוכתמים:
גִוּוּן לקטופוצ'סיןמכיל פוקסין חמוץ - 0.1 גרם, חומצה לקטית - 100 מ"ל. מוכתם למשך 3-5 דקות. הרקע של התרופה ורוד, התפטיר הוא אופלסנטי בכחול. פטריות מכתימות היטב עם mucormycosis ואספרגילוזיס.
№29 תאי שמרים ודמויי שמרים בעלי צורה מעוגלת, פיאל או מוארכת, בגודל 8-10 מיקרון, עם קרום דו-מעגלי. בציטופלזמה מצוינים תכלילים בצורה של גרגירי גליקוגן, וולוטין ושומנים. רבייה מתרחשת על ידי ניצנים ואסקוספורים. שמרים דמויי שמרים דומים לשמרים אמיתיים, ההבדל הוא היעדר אסקוספורים והיכולת ליצור פסאודומיצליום. עם היווצרות פסאודומיצליום, התאים מתארכים ונוגעים בבסיס צר. הם גורמים לקנדידה, המתפתחת אצל חולים עם ירידה חדה בעמידות הגוף ושימוש ממושך באנטיביוטיקה. שיטות ללימוד שמרים
התקופה המודרנית של חקר המגוון הביולוגי מאופיינת בפיתוח אינטנסיבי של שיטתיות פילוגנטית, שמטרתה לשחזר נתיבים ספציפיים של התפתחות היסטורית של אורגניזמים. במיקרוביולוגיה, הסיסטמטיקה הפילוגנטית קיבלה תנופה חזקה להתפתחות רק בסוף המאה ה-20. בקשר למחקר ההשוואתי של רצפי נוקלאוטידים שמורים ב-rRNA. בשמרים, טקסונומיה זו מבוססת בעיקר על מחקר של שני אזורים של rDNA באורך של כ-600 זוגות נוקלאוטידים: תחום D1/D2 בקצה 5' של הגן המקודד ל-26S rRNA ואזור ה-Internscribed Spacer (ITS) , הכולל את הגן rRNA 5.8S. מאמינים שבשל השמרנות של אזורים אלה, ההבדלים ביניהם עומדים ביחס ישר למרחק הפילוגנטי, למידת הקשר האבולוציוני. התברר כי רצף רצפי rDNA שאינם קלאוטידים הוא כלי רב עוצמה לבניית סיווג פילוגנטי של שמרים וקביעת מקומם במערכת הכללית של פטריות.
עד כה, רצפי נוקלאוטידים של rRNA פוענחו והונחו במאגרי מידע ממוחשבים הזמינים באינטרנט בנציגי כל מינים ידועיםשמרים. זה מאפשר בניית עצים פילוגנטיים המשקפים את התפתחות הגנים הריבוזומליים שלהם. התברר כי קיבוץ השמרים המבוסס על הדמיון של רצפי הנוקלאוטידים rRNA במקרים רבים אינו עולה בקנה אחד עם הקיבוץ לפי תכונות פנוטיפיות. רבות מהתכונות המסורתיות המשמשות בסיווג שמרים, כגון מאפייני ריבוי וגטטיביים, צורת אסקוספורה, תסיסה והטמעת סוכר, נחשבו כלא אמינות, לא מתאימות לקביעת קשרים פילוגנטיים. רצף של rRNA (rDNA) נחשב כעת חיוני בעת תיאור מיני שמרים חדשים.
השפעה חזקה במיוחד על חקר השמרים, כמו גם על רוב הקבוצות האחרות של מיקרואורגניזמים, הייתה להתפתחות מהירה בסוף המאה ה-20. ביולוגיה מולקולרית. בטקסונומיית השמרים המודרנית, נעשה שימוש נרחב בשיטות לשיטתיות גנים המבוססות על השוואה ישירה של גנומים ורצף של רצפי נוקלאוטידים. השימוש בשיטות ביולוגיות מולקולריות מאוחדות איפשר לקרב עוד יותר גישות לטקסונומיה של שמרים ופטריות חוטיות, לבסס קשרים בין אנמורפים שמרים לטלאומורפים חוטיים ולפתח קריטריונים חדשים ליצירת מערכת פילוגנטית מאוחדת עבור ממלכת מיקוטה כולה. . יחד עם זאת, ידע חדש הוליד חדש בעיות מדעיותבפרט, בעיית המתאם בין השיטות המולקולריות העדכניות ביותר לבין שיטות מסורתיות המבוססות על גישות מורפולוגיות ופיזיולוגיות לחקר השמרים. הגנום של Saccharomyces cerevisiae פוענח כמעט לחלוטין, מה שפותח סיכויים גדולים לגנומיקה של שמרים ואופקים חדשים לשימוש הביוטכנולוגי שלהם. לפיכך, מדע השמרים, לאחר שעבר יותר ממאה וחצי, ממשיך להתפתח במהירות במאה ה-21.
גם תכשירים מקומיים (לא מוכתמים) וגם תכשירים מוכתמים מוכנים לבדיקה מיקרוסקופית. כדי לחקור פטריות ברקמות, מתבצעת בדיקה פתואיסטולוגית.
תפטיר נבגי שמרים סלולריים
30. מורפולוגיה ושיטות מחקר של פטריות חוטיות. ישנם 2 סוגי גידול בפטריות: גידול היפל (hyphomycetes) וגידול שמרים (blastomycetes). בדרך כלל, הגוף הצומח של פטריות חוטיות מורכב מחוטים בעובי של כ-5 מיקרומטר, מסועפים מאוד ונקראים היפאים. להיפאות אין מחיצות רוחביות (בפטריות תחתונות) או שהן מופרדות על ידי מחיצות (מחיצות) לתאים (ב פטריות גבוהות יותר). דופן התא יכול להיות בעוביים שונים, לעתים קרובות המעקף נראה בבירור; בין התכלילים בציטופלזמה, המאפיינים ביותר הם גרגרי וולוטין, גליקוגן, פיגמנט מלנין. תאים פטרייתיים ישנים בוגרים עשירים בשומנים. הגרעין מכיל גרעין ורשת כרומטין; תאים יכולים להיות מרובי גרעינים. אוסף ההיפיות יוצר תפטיר (תפטיר). התפטיר יכול להיות מצע, הנוצר כתוצאה מגידולים של היפאים לתוך המדיום המזין, ואוויר, הגדל על פני המדיום. התפטיר מייצג צינורות מסועפים, הסתעפות מתבצעת על ידי יציאות לרוחב של היפאות. חוטי מיסיאלי מסודרים לפעמים בשורות מקבילות, צמודות זה לזה, הדומים ל"פתיל", ומכאן השם "קורמיה" (בדרמטופיטים).
31. עקרונות בסיסיים של סיווג ומורפולוגיה של פרוטוזואה. שיטות המחקר והמשמעות הרפואית שלהם.
פרוטוזואה הם אוקריוטים חד-תאיים, הדומים במבנה לתאים של בעלי חיים מאורגנים בצורה מורכבת. הצורה יכולה להיות בצורת אגס (Trichomonas, Giardia), ביצית (Balantidia), בצורת ציר (Trypanosoma, Leishmania), יכולה לקבל את התצורה המוזרה ביותר (אמבה)
רוב הפרוטוזואה הם ניידים והתנועה מתבצעת בעזרת פסאודופודיה (אמבה, מלריה פלסמודיום), פלגלה (גיארדיה, לישמניה), ריסים (בלנטידיה).
פסאודופודיה- בליטות זמניות של הציטופלזמה, שמשחררות אותן הפרוטוזואה משנות את צורת הגוף כל הזמן.
הפשוטים ביותר שייכים לממלכה פרוטוזואה (פרוטוס - הראשון, זואה - חיות). בעלי חשיבות רפואית הם:
Phylum Sarcomastigophora, subphylum Sarcodina (Sarcodidae). הגוף שלהם נטול כדורים, הם נעים בעזרת פסאודופודיה. שיעור זה כולל סוגים שוניםאמבה, כולל אמבה דיזנטרית (Entamoeba histolytica).
שיטות מחקר. לחקר פרוטוזואה מכינים תכשירים זמניים וקבועים (מוכתמים). מכינים תכשירים זמניים בשיטה של "טיפה מרוסקת" או "טיפה תלויה" בתוספת חמימה מי מלח פיזיולוגיאו צבעים תוך-חיוניים. מתכשירי הדם מכינים "טיפה עבה". לשם כך הופכים את האצבע שטופלה באתר בניקור מטה ואל הטיפות הבולטות מביאים שקף זכוכית שעליו לוקחים 2-3 טיפות דם, ואז מחלקים את הדם במחט או בפינה של שקופית זכוכית נוספת לקבלת סגלגל של כ-1 ס"מ, כדי להאיץ את ייבוש התרופה, ניתן לשים אותה בתרמוסטט בטמפרטורה של 35-37 0 C.
כדי לזהות ציסטות, נעשה שימוש בתמיסת Lugol חזקה, המכתימה את מבני הציסטות בצבע חום כהה.
מוכתם בהמטוקסילין ברזל לפי שיטת היידנהיין
מריחות לאחר הקיבוע מונחות בתמיסה של 2.5% של אלום ברזל למשך שעה.
לאחר שטיפה שלוש פעמים במים, צובעים בצבע (0.5 גרם המטוקסילין, 10 מ"ל אלכוהול 96 0, ולאחר הפירוק מוסיפים 90 מ"ל מים מזוקקים) למשך 5-10 דקות.
נשטף במים ומיובש.
צביעה היא הכרחית במקרים שבהם יש צורך לחשוף את הפרטים הקטנים ביותר של מבנה הגרעין והציטופלזמה של הפרוטוזואה.
לדפנות התאים של הצמחים יש חוזק יוצא דופן, ובתהליך הצמיחה הצמחים משנים את מבנהם והרכבם. המרכיבים העיקריים של קירות תאי הצמח הם פוליסכרידים, שביניהם שולטת תאית, מה שקובע במידה רבה את הארכיטקטורה של הקיר.
תָאִית. ההומפוליסכריד הזה הוא הפחמימה הנפוצה ביותר על פני כדור הארץ (צמחים יוצרים עד 1011 טון תאית בשנה). מונומרים של תאית הם שיירי גלוקוז המחוברים בשרשראות ארוכות (עד 10,000 שיירי גלוקוז בכל אחד מהם) באמצעות קשרים מסוג b (1 → 4) -גליקוזידים (איור 5.3). במולקולה כזו, אין חופש מוחלט של סיבוב סביב קשרי 1C-O- ו-O-4C, והפולימר מקבל קונפורמציה נוחה ליצירת קשרי מימן בין שרשרת כאשר השרשראות מסודרות אנטי מקבילות. כתוצאה מכך, מולקולות תאית משולבות לתוך מיקרופיבריליםעובי של 10 עד 25 ננומטר בערך. מיקרופיברילים משתלבים ויוצרים חוטים דקים, אשר, בתורו, ניתן ללפף אחד סביב השני, כמו גדילים בחבל, ויוצרים מקרופיברילים.כל מקרופיבריל הוא בעובי של כ-0.5 מיקרומטר ויכול להגיע לאורך של 6-8 מיקרומטר. חוזקם של מקרופיברילים דומה לחוזק של חוט פלדה שווה בעובי. חוץ מזה, סעיפים נפרדיםלמיקרופיברילים יש מבנה מסודר ומעניקים לדופן התא תכונות גבישיות. לפיכך, ניתן לציין את המורכבות והסדר הגבוה של תאית בהרכב דפנות התא, וזה לא מקרי: פולימר זה מבצע פונקציות הגנה ותומכות בצמח.
בצורה זו, פוליסכרידים אינם נגישים לפעולת האנזימים שלהם, ותאית אינה יכולה לשמש את הצמח כחומר רזרבה. רק לאורגניזמים בודדים (חלק מהחיידקים, פטריות, פרוטוזואה וחיות נדירות) יש מערכות אנזימים המסוגלות לפרק תאית.
מיקרו- ומקרו-פיברילים של תאית בדופן התא של צמחים שקועים בתוכם מַטרִיצָה, שגם מורכב בעיקר מפוליסכרידים ומשנה את מבנהו במהלך גידול הצמח. עַל שלבים מוקדמיםמטריצה התפתחותית מורכבת מ חומרים פקטין, ובהמשך מופיעים בו קסילניםופוליסכרידים ניטרליים שונים ("המיצלולוזה"). חומרי פקטין הם פולימרים של חומצה א-גלקטורונית, שבהם חלק מאטומי המימן מוחלפים בקבוצות מתיל (-CH 3) (איור 5.3). Xylanes הם פולימרים של xylose (איור 5.3).
בשלבים מאוחרים יותר של התפתחות, כאשר מתרחשת הלירות של דפנות התא, עֵצָן- מכיל פולימר עמיד כימית מספר גדולטבעות ארומטיות. בנוסף, כמויות קטנות של גליקופרוטאין, פולימרים שומנים בלתי מסיסים, נמצאות בהרכב דפנות תאי הצמח. מבנה שונהושעווה.
קירות התא של צמחים מסוימים מכילים פוליסכרידים נדירים בעלי מבנה יוצא דופן. למשל, בקירות ובבין תאיים
החומר של אצות אדום ים מכיל אגר הטרופוליסכריד מורכב, שהוא תערובת של פוליסכרידים סולפטים - אגרוז ואגרופקטין. Agarose בנוי משאריות מתחלפות של D-galactose ו-3,6-anhydro-L-lactose המקושרים לסירוגין על ידי קשרי b(1→4)- ו-a(1→3). באגרופקטין יש יותר מבנה מורכב: הוא מכיל D-galactose, 3,6-anhydrogalactose, חומצות אורוניות וסולפט. אגר משמש כדחס הנפוץ ביותר עבור מדיה מוצקה, הכרחי במיקרוביולוגיה, כמו גם ב תעשיית המזוןעבור מוצרי ג'לינג. יש לציין שרובם המוחלט של המיקרואורגניזמים אינם מסוגלים לפרק אגר, וזהו אחד היתרונות העיקריים שלו על פני מדחס חומרי תזונה אחר - ג'לטין. אגרוז מוצא יישום רחבבמחקר ביוכימי: היא סביבה מימיתיוצר ג'ל עם נקבוביות גדולות, שגודלו נקבע לפי ריכוזו. ג'לי אגרוז משמשים לפירוק חלבונים וחומצות גרעין, כמו גם לקיבוע תאים.
- 20.44 Kbדופן התא, מבנה, הרכב כימי. שינויים בדופן התא.
נוכחות של קליפה חזקה מאפייןתא צמחי המבדיל אותו מתא של בעלי חיים. הקליפה מעניקה לתא צורה וחוזק מסוימים ומגנה על התוכן החי - הפרוטופלסט ובעיקר הפלזמה, הנלחצים בחוזקה כנגד הקליפה מבפנים. התאים של צמחים גבוהים יותר, שבהם הממברנה מוסרת בשיטה האנזימטית, מעובדים על חומרי הזנה מיוחדים, מקבלים תמיד צורה כדורית. ללא קליפה, קיומו של תא צמחי בתנאים רגילים בלתי אפשרי, שכן לחץ הטורגור האופייני לו, מאוזן על ידי לחץ הנגד של הקליפה, יוביל בהכרח לקרע של הפלזמה ולהרס של הפרוטופלסט. מכלול קירות התא בהרכב גופו של צמח יבשתי, המתנשא מעל פני הקרקע ואין לו שלד פנימי, מהווה מעין מסגרת תומכת המעניקה לצמח חוזק מכני.
הקליפה, ככלל, חסרת צבע ושקופה, מעבירה בקלות אור שמש. מים וחומרים מומסים במשקל מולקולרי נמוך יכולים לנוע לאורכו. לכל תא יש מעטפת משלו; הקונכיות של תאים שכנים, כביכול, מוצקות על ידי חומרים בין-תאיים היוצרים את מה שמכונה הלמינה החציונית. כתוצאה מכך, תאים שכנים מופרדים זה מזה על ידי דופן המורכבת משתי קונכיות וצלחת אמצעית, מה שנותן סיבה לקרוא לקליפה גם דופן תא.
הקונכייה נבנית על ידי הפרוטופלסט של התא ולכן יכולה לגדול רק במגע איתו. לעתים קרובות מאוד, הקליפה נמשכת זמן רב יותר מהפרוטופלסט (כאשר התא מת מוקדם), אך לרוב הדבר אינו מוביל להפסקת קיומו של התא כיחידה מבנית, מכיוון שצורת התא אינה משתנה עקב חוזק הקליפה. לכן, גם לאחר המוות, תאי צמחים יכולים לבצע פונקציות חשובות של הובלת פתרונות או תמיכה מכנית. סוגים רבים של תאי צמחים - סיבים, טרכאידים, מקטעי כלי דם, תאי שעם - במצב בוגר הם קרום תא אחד. בעיקר עץ מורכב מקונכיות של תאים מתים.
ממברנות התא מגוונות כמו התאים עצמם. לעתים קרובות ניתן להשתמש בהרכב ובמבנה של הממברנה כדי לשפוט את המקור והתפקוד של תאים מסוגים שונים. לפיכך, המבנה של צמחי מאובנים נחקר בעיקר על ידי חקר הממברנות של התאים שלהם.
בתחילה, דופן התא הראשוני עולה החוצה מהפלזמה. הוא מורכב מפוליסכרידים - פקטין ותאית. קירות התא הראשוניים של תאים שכנים מחוברים על ידי למינה חציונית של פרוטופקטין. בדופן התא, מולקולות תאית ליניאריות, ארוכות מאוד, המורכבות מגלוקוז, מורכבות לצרורות - מיצלות, אשר, בתורן, משולבות לסיבים - הסיבים הדקים ביותר באורך בלתי מוגדר. תאית יוצר שלד רב מימדי, אשר טובל במטריצה אמורפית מושקת גבוהה של פחמימות שאינן תאית: פקטין, המיצלולוז וכו'. התאית היא שקובעת את חוזק דופן התא. מיקרופיברילים הם אלסטיים ובעלי חוזק מתיחה דומה לפלדה. פוליסכרידים מטריקס קובעים תכונות כאלה של הקיר כמו חדירות גבוהה למים, מולקולות קטנות ויונים מומסים ונפיחות חזקה. הודות למטריצה, מים וחומרים יכולים לעבור מתא לתא לאורך הקירות הסמוכים זה לזה. חלק מההמיצלולוזות יכולות להיות מופקדות בדפנות התא של זרעים כחומרי רזרבה.
חומר מעטפת, כולל תאית, נמצא בשימוש נרחב בתעשייה. תכונות העץ, הנייר, חומרי הטקסטיל תלויות במידה רבה במבנה המולקולרי ובהרכב הכימי של המעטפת, והידע בהם מסייע לשיפור טכנולוגיית הייצור ואיכות המוצר.
הרכב כימי וארגון מולקולרי של הקליפה. קרום התא בנוי בעיקר מפוליסכרידים, שהמונומרים שלהם - סוכרים - מחוברים ביניהם בקשר גליקוזידי (-O-) בצורת שרשרת. פוליסכרידים נקראים לפי הרכב המונומרים היוצרים אותם, בתוספת הסיום "an". לדוגמא, הפוליסכריד קסילן מורכב משאריות סוכר קסילוז, מנאן - מנוז, גלוקן - גלוקוז וכו'.
בנוסף לפוליסכרידים, הרכב הקליפה עשוי לכלול חלבונים, מלחים מינרליים, ליגנין, פיגמנטים, שומנים. בדרך כלל הפגזים ספוגים במים. פוליסכרידים מעטפת על פי תפקידם ניתן לחלק לחומרי שלד וחומרים מטריקס. החומר השלד של הקליפה צמחים גבוהים יותרהוא תאית (סיבים), המייצג כימית β-1, 4-D גלוקן. מספר שאריות הגלוקוז במולקולה, וכתוצאה מכך, אורך המולקולות עצמן עשוי להשתנות באופן דרמטי בדפנות התא של צמחים שונים, מה שמשפיע על תכונות התאית. המולקולות החוטיות שלו בקליפה מסודרות במקביל זו לזו ומקובצות בכמה עשרות באופן שמופיע סריג תלת מימדי מוארך, האופייני לגבישים. קבוצות גבישיות אלו, שבהיווצרותן מעורבים קשרים קוולנטיים ומימן, מהוות את הבסיס למיקרופיברילים - הסיבים הדקים ביותר היוצרים את המסגרת המבנית של המעטפת, את ה"חיזוק" שלה.
המצב הגבישי של מולקולות תאית במיקרו-סיביים קובע את השבירה הדו-פעמית של דופן התא, הנצפית במיקרוסקופ מקטב. במיקרו-פיברילים, ישנם אזורים פארא-גבישיים, כלומר אזורים עם סידור של מולקולות תאית רופף יותר, אך ללא אוריינטציה תלת-ממדית. מידת הגבישיות של מיקרו-סיביים, היחס בין אזורים גבישיים לפרא-גבישיים משתנים מאוד בהתאם לסוג הצמח, מצב התא וסוג הקליפה, מה שמשפיע על התאית המכנית והפיזיקלית כחומרי גלם. הקוטר של מיקרופיברילים משתנה גם בתאים שונים (בדרך כלל בתוך 10-30 ננומטר), האורך יכול להגיע לכמה מיקרומטרים. ניתן לשלב מיקרו-פיברילים לקבוצות או שכבות נפרדות - מקרו-פיברילים בעובי של 0.4-0.5 מיקרון, הנראים במיקרוסקופ אור.
איור 25
במהלך חיי התאים, במיוחד בהתאם לביצועים של כל תפקיד מיוחד, מתרחשים שינויים משניים בדופן התא. לעתים קרובות הם קשורים לשינויים קיצוניים בהרכב הכימי שלו, במבנה ובתכונות הפיזיקוכימיות שלו.
עצים. דופן התא עטופה (ספוגה) בחומר מיוחד - ליגנין, המגביר את הקשיות, תכולת הקלוריות, צפיפות דופן התא ומפחית את הפלסטיות שלו ואת יכולת הצמיחה שלו. דפנות תאים מגוונות אינן מאבדות את יכולתן להעביר מים ואוויר. הפרוטופלסט שלהם יכול להישאר בחיים, למרות שהוא בדרך כלל מת. עצים נפוץ מאוד בטבע. הוא מספק כוח לגזעים ולענפים של עצים. עץ מחטניים ועץ קשה מכיל עד 50% תאית ו-20...30% ליגנין. דפנות תאים מנומרות וצמחי מרפא רבים, במיוחד לקראת סוף עונת הגידול.
חיטוט, או תת-התרה. כתוצאה מרבדים רבים בדופן התא של חומר עמיד כימית - סוברין, מתרחשת הסוברין שלו. מטבעו הכימי, סוברין קרוב לשומנים. זוהי תרכובת הידרופוביה אמורפית, המורכבת מחומצות שומן רוויות פולימריים גבוהים וחומצות הידרוקסי מורכבות. סוברין מושקע בעיקר בדפנות התא של הרקמה המשנית המשנית - הפקק, שהוא מבודד מים ותרמי מצוין. בכמויות קטנות הוא מצטבר בדפנות התא של רקמות שונות. עם פקק מלא, הפרוטופלסט של התא מת, שכן lamellae suberin מונעות את חדירת המים והאוויר, עם חלקית - הפרוטופלסט נשמר לאורך זמן.
מינרליזציה. בתהליך החיים, דפנות תאי פני השטח של הנבטים יכולים לצבור חומרים מינרליים, במיוחד סיליקה וסידן פחמתי. בקבוצות מסוימות של צמחים, למשל, סוגים רבים של זרעים, עשבים, זנב סוס, תמיד יש תאים של רקמת המוח. , האפידרמיס, מכוסה בכבדות במלחים. יתר על כן, לא רק את דופן התא ניתן לחרוץ, אלא גם תולדות שונות של האפידרמיס - טריכומות. בנוסף, חומרים מינרליים מופקדים על פני התאים של רקמת האינטגמנטרית - האפידרמיס. כתוצאה ממינרליזציה, האיברים הציריים רוכשים חוזק מכני גדול יותר.
הרזיה. הפיכת תאית ופקטין לליחה ולחניכיים הקרובים אליהם, שהן פחמימות פולימריות, הנבדלות ביכולתן להתנפח חזק במגע עם מים. ריר נצפה בתאים התאיים של קליפת הזרע, למשל, בפשתן. להיווצרות הריר חשיבות אדפטיבית רבה. כאשר זרעים נובטים, הריר מקבע אותם במקום מסוים, סופג בקלות ושומר על לחות, מגן על הזרעים מפני התייבשות ומשפר את משטר המים של השתילים. הרירי של דפנות התא של וולוסק השורש מבטיח את הקשר החזק שלהם עם חלקיקי אדמה.
מריחה היא פירוק החומר הבין-תאי, המוביל להפרדה של תאים. מריחה טבעית מתרחשת בפירות בוגרים. היא מתבצעת באופן מלאכותי, למשל, כאשר פשתן מופרש כדי לשחרר את חומרי הגלם המסתחררים - קבוצות של תאים של סיבי באסט.
תיאור העבודה
נוכחות של קליפה חזקה היא תכונה אופיינית תא צמחישמבדיל אותו מתא חיה. הקליפה מעניקה לתא צורה וחוזק מסוימים ומגנה על התוכן החי - הפרוטופלסט ובעיקר הפלזמה, הנלחצים בחוזקה כנגד הקליפה מבפנים. התאים של צמחים גבוהים יותר, שבהם הממברנה מוסרת בשיטה האנזימטית, מעובדים על חומרי הזנה מיוחדים, מקבלים תמיד צורה כדורית.