Wielokrotnie wspominaliśmy, że jeden lub drugi z wymienionych badaczy zauważył jądra w komórkach.
Ponieważ w pracy Schleidena, do której przejdziemy dalej, jądro ma szczególne znaczenie, to odchodząc od chronologicznego ujęcia, rozważymy tutaj historię odkrycia tej najważniejszej części komórki. To jądro pomogło Schwannowi porównać komórki zwierzęce i roślinne, dlatego odkrycie jądra wyznacza najważniejszy etap w rozwoju teorii komórki.
Jądra zostały po raz pierwszy zaobserwowane w erytrocytach ryb przez Löwenhoeka w 1700 roku i przedstawił je na rysunku. Później na tym samym obiekcie - erytrocytach wielu kręgowców i bezkręgowców - Hewson (1777) naszkicował jądra. Znaczenie tej edukacji w tym wczesny okres Początków mikroskopii oczywiście nie mogli zrozumieć ani sami autorzy, ani ich współcześni. Fontana w swoich badaniach nad jadem żmii, przedstawiając komórki nabłonkowe naskórka i erytrocyty, odwołuje się do komórek jądra i krótko wspomina „je w tekście; ale i w tym czasie (praca Fontana ukazała się w 1781 r.), kiedy badanie mikroskopowe tkanek zwierzęcych, odkrycia Fontana nie można było zrozumieć.
W tym samym czasie niektórzy badacze obserwowali jądra w jajach. Cavolini (Filippo Cavolini, 1756-1810) widział jądra w jajach ryb (1787); i Poly (Poli, 1791) odnotowali jądra w jajach mięczaków. Ich obserwacje mijały bez śladu, nie zwracając na siebie uwagi.
W badaniu jaj ptasich (1825) Purkinė opisał „pęcherzyk embrionalny” (vesicula germinativa). To było jądro ptasiego jaja. Według opisu Purkine'a jest to „ściśnięta sferyczna bańka, ubrana w najcieńszą skorupę. Zawiera własną limfę, jest włączony do białego guzka piersiowego i jest pełen mocy generatywnej, dlatego nazwałem go „pęcherzykiem embrionalnym”. Purkyne przywiązał się do edukacji, którą odkrył bardzo ważne; po nim kolejni badacze nie ignorowali już owej tajemniczej "bańki". Odkrycie Purkineta nie przeszło więc bez śladu, podobnie jak obserwacje Cavoliniego i Poli, ale znaczenie „pęcherzyka zarodkowego” pozostawało przez długi czas niejasne, ponieważ w zrozumieniu części jaja, z punktu widzenia koncepcji „komórki, właściwy sposób został nakreślony dopiero po badaniach Schwanna.
U roślin pierwszy obraz jądra komórkowego wykonał Bauer w 1802 r., ale rysunek ten opublikowano dopiero w 1830 r. (J. Baker, 1949). Meyen (1830) przedstawia jądro na jednej figurze. Mirbel (1831-1832) w opracowaniu dotyczącym marchantii również przedstawia jądro, nadając mu nazwę kuli; widział go także francuski botanik Brognard (Adolphe Brogniart, 1801-1876). Ale te pierwsze obserwacje jąder w komórkach roślinnych nie spotkały się z uznaniem samych obserwatorów i też nie wzbudziły zainteresowania.
Uznanie jądra za nieodzowną część komórki roślinnej jest zasługą angielskiego botanika Roberta Browna (Robert Brown, 1773-1858).
Rozpoczynając swoją pracę botaniczną od opisu kolekcji zebranych podczas podróży do Australii, Brown przechodzi następnie do studium struktura anatomiczna rośliny. Nie stawiał w swojej pracy problemów czysto morfologicznych; studia anatomiczne są dla niego narzędziem do badania taksonomii roślin, ale w tych pracach Brown dokonuje wybitnych odkryć botanicznych dotyczących rozmnażania się roślin. W 1833 r. ukazała się praca Browna „O narządach i sposobie zapłodnienia storczyków” (zgłoszona do Linnean Society w Londynie już w listopadzie 1831 r.). Brown pisze w tym artykule, że w każdej komórce naskórka zaobserwował „pojedynczą zaokrągloną otoczkę, zwykle ciemniejszą niż błona komórkowa. Ta otoczka jest mniej lub bardziej ziarnista, lekko wypukła i chociaż wydaje się, że leży na powierzchni, w rzeczywistości jest pokryta zewnętrzną płytką komórki. Jego pozycja w komórce nie jest stała; często jednak w centrum lub w jego pobliżu” (s. 710). Tę otoczkę, czyli jądro (jądro) komórki, jak Brown inaczej określa tę formację, zaobserwował przez niego nie tylko w komórkach naskórka; widział jądro w miąższu, w wewnętrznych komórkach części roślin, „zwłaszcza gdy są one wolne od materii ziarnistej”. Brown jednak ostrożnie sugeruje, że jądro jest zwyczajne część integralna komórki. Nie ma kategorycznego stwierdzenia, że jądro jest obowiązkową organellą komórki; podobnie Brown nie podaje w swojej pracy obrazów jąder komórkowych. Niemniej jednak w badaniach Browna po raz pierwszy jądro nie jest wymieniane jako przypadkowa formacja w komórce, ale pojawia się jako pewnego rodzaju istotna część, która jest ważna dla życia komórki.
Meyen, autor „Fitotomii” – pracy, o której była mowa wcześniej – w późniejszym podręczniku” Nowy system Fizjologia roślin” (1837-1839) wymienia jądro jako stałą część komórki, której znaczenie pozostaje tajemnicze. Właściwie dopiero praca Negeli (S. Nageli, 1844) dowiodła uniwersalnego rozmieszczenia jąder komórkowych nie tylko w roślinach kwitnących, ale także w komórkach alg, grzybów, mchów i innych. niższe rośliny.
W histologii zwierząt termin „jądro” wprowadził Valentin. W wiadomości „O delikatna struktura narządy zmysłów” (1836) Valentin pisał o nabłonku spojówki: „Składa się z romboidalnych lub kwadratowych zaokrąglonych komórek leżących blisko siebie, których granice są utworzone przez proste włókniste linie; w każdej komórce, bez wyjątku, znajduje się nieco ciemniejsze i bardziej zwarte jądro (jądro) o okrągłym lub podłużnym zaokrąglonym kształcie. Przez większą część zajmuje środek komórki, składa się z drobnoziarnistej substancji, ale zawiera wewnątrz doskonale okrągłe ciało, które według tego samego wzoru tworzy w sobie coś w rodzaju drugiego jądra” (s. 143). Ten opis pokazuje, że Valentin wyraźnie obserwował jądra komórek nabłonkowych. Wewnątrz jądra Valentine zobaczył jąderko; był to najwyraźniej pierwszy opis tej struktury wewnątrzjądrowej.
Wyraźny opis i obraz jąder w komórkach nabłonkowych podał Henle (1837). Baker (1949) słusznie zauważa, że praca Valentina i Henlego rozpoczyna erę komórek jądrowych w histologii zwierząt.
W 1838 r. w Archiwum Müllera ukazał się artykuł młodego botanika Schleidena pod tytułem „Materiały do fitogenezy” (Beitrage zur Phytogenesis). Ta praca jest tradycyjnie uważana kamień milowy w rozwoju nauczania komórkowego, a jego autor jest uznawany wraz ze Schwannem za twórcę teoria komórki. Znaczenie Schleidena w historii teorii komórkowej jest bezdyskusyjne, ale w literaturze edukacyjnej, popularnej, a czasem historycznej znaczenie to jest ujęte powierzchownie i niepoprawnie. Schleiden jest czasami uznawany za prawie odkrywcę komórki roślinne, Dlatego. konieczne jest zrozumienie, jakie jest naprawdę znaczenie tego naukowca w historii nauczania komórkowego, gdzie jest prawda w legendzie, która rozwinęła się wokół jego pracy i tradycyjnie przechodzi z podręcznika do podręcznika.
Maciej Schleiden(Matthias Jacob Schleiden, 1804-1881) jest najwiekszy przedstawiciel Botanika niemiecka połowy ubiegłego wieku. Początkowo ukończył Wydział Prawa i praktykował adwokacką. Nie mając powodzenia w tej działalności, Schleiden w 1831 roku porzucił prawo i zaczął studiować medycynę i nauki przyrodnicze. Od 1840 był profesorem botaniki w Jenie, gdzie pozostał do 1862. Jest to główny okres aktywność twórcza Schleiden. W 1842 roku ukazało się jego główne dzieło „Podstawy botaniki naukowej”, które odegrało ważną rolę w kierunku dalszych badań botanicznych. Zamiast rozumowania przyrodniczo-filozoficznego Schleiden wymaga wprowadzenia do botaniki dokładne metody badania struktury i funkcji roślin; szczególnie podkreślał potrzebę zwrócenia uwagi na historię rozwoju, w której widział klucz do rozwiązania wielu kontrowersyjnych problemów. Stanowiska filozoficzne Schleidena, wyrażone przez niego w jego pismach, nie są oryginalne i ujawniają piętno filozofii kantowskiej. Od 1862 do 1864 Schleiden był profesorem antropologii w Dorpacie (obecnie Tartu, estońska SRR), w 1864 opuścił Dorpat w wyniku starć z kręgami kościelnymi i jednocześnie przestał działalność pedagogiczna. Bycie autorem nie tylko serii prace naukowe, ale także wielu popularnych utworów, Schleiden był szeroko znany.
„Materiały do fitogenezy” to drugie dzieło Schleidena, który był wtedy jeszcze początkującym botanikiem. Jest to artykuł o objętości około 40 stron, do którego dołączone są dwie tabele. Ogólne podstawowe prawo ludzkiego umysłu — tak zaczyna swoją pracę Schleiden — prawo, które determinuje jego nieodparte pragnienie jedności w poznaniu i ustanowienia, zarówno w nauce, jak i w dziedzinie organizmów, analogii dla obu duże działy – królestwa zwierząt i roślin – wielokrotnie skłaniały do zajmowania się tym tematem. Tak wiele umysłów było nim zajętych, ale - temu nie można zaprzeczyć - wszystkie dotychczasowe próby w tym zakresie zawiodły i były złudzeniami. Przyczyna leży w fakcie, że pojęcie jednostki, w takim sensie, w jakim stosuje się je w przyrodzie zwierzęcej, nie ma zastosowania w świecie roślinnym. Najwspanialszą rzeczą, jaką można powiedzieć o jednostce w tym sensie, są najniższe rośliny, niektóre glony i grzyby, składające się tylko z jednej komórki. Ale każda bardziej rozwinięta roślina jest zbiorem całkowicie zindywidualizowanych zamkniętych indywidualności… które są komórkami” (s. 137). Celowo to przynieśliśmy długi cytat, który jest początkiem artykułu Schleidena, aby pokazać, jak obca była mu idea jedności struktury mikroskopowej zwierząt i roślin, wyrażonej w strukturze komórkowej. Tymczasem to właśnie ten pomysł jest kamieniem węgielnym teorii komórkowej, za jednego ze współautorów uważa się zazwyczaj Schleidena.
Aby właściwie ocenić pracę Schleidena, należy pamiętać o pozycji teorii komórkowej w botanice do 1837 r., kiedy Schleiden zakończył swoją pracę. Całkowicie fałszywy pomysł, że Schleiden udowodnił uniwersalne rozmieszczenie komórek w roślinach, a nawet odkrył komórki w roślinach. To jest zakłamywanie rzeczywistości. rozwój historyczny Nauki. Na początku lat trzydziestych ubiegłego wieku w botanice powstawała kompletna koncepcja komórki jako struktury elementarnej. flora; Schleiden w swojej pracy przyjmuje to stanowisko jako niewzruszoną konkluzję. Nawet takie niekomórkowe części roślin, jak się wcześniej wydawało, jako wodonośne naczynia z drewna, do tego czasu są uważane za zmodyfikowane, osobliwie zróżnicowane i połączone komórki. Schleiden nie musiał ustalać powszechnego rozmieszczenia komórek w roślinach: ustalenie tego stanowiska było, jak widzieliśmy, zbiorowym sukcesem wielu prac licznej galaktyki botaników z pierwszej ćwierci ubiegłego wieku.
K. A. Timiryazev (1920) słusznie napisał o wyrażeniu „odkrycie komórki”: „Ale faktem jest, że komórki nikt nie odkrył” (s. 79), podkreślając tym samym, że „odkrycie” komórki nie jest zasługą pewien naukowiec. Nie jest również prawdą, że Schleiden, jak pisze Aschoff (1938), rozwinął doktrynę „wszechogarniającej konstrukcji z komórek wszystkich roślin” (s. 177). I w tym względzie rację ma K. A. Timiryazev, który napisał: „Schleiden jest powszechnie uważany za twórcę tej doktryny komórki, która okazała się tak bogata w najbardziej owocne uogólnienia. Ale to niesprawiedliwe… Schleiden, elokwentny, żarliwy przeciwnik rutyny i stagnacji, mógł słusznie powiedzieć o sobie, jak kiedyś Bacon, że jest trębaczem, heroldem, buccynatorem, obwieszczającym powstanie tej doktryny, ale fakty potwierdzające, że istniały już wcześniej…” (s. 75). Charakterystyczne jest, że Unger (Unger, 1846) w swoich podstawach botaniki, nakreślając pozycję komórki jako uniwersalnej elementarnej struktury organizmów, wskazuje w odnośniku literackim na Schwanna i Kölliker, nie wspominając nawet o Schleidenie w tym aspekcie.
Sama koncepcja komórki u Schleidena nie różni się od idei, które ukształtowały się wcześniej i znalazły odzwierciedlenie w podręczniku Meyena (1830) jeszcze zanim Schleiden zaczął studiować botanikę. Na poziomie tych idei Schleiden wziął komórkę za bańkę lub komorę odgraniczoną muszlą, wewnątrz której może się znajdować zawartość. Ta „zawartość” komórki (przyszła protoplazma!) zwróciła też uwagę Meyena, który poświęcił jej wiele badań, nie rozumiejąc jednak znaczenia tego głównego składnika komórek. Schleiden widział także protoplazmę komórek roślinnych, ale nie rozumiał znaczenia „zawartości” komórki. Dla niego jest to guma (Gummi) lub galaretka (Gallerte). Część protoplazmy Schleiden przypisana do ściany komórkowej. Ta ostatnia jego zdaniem składa się z dwóch warstw, pomiędzy którymi znajduje się jądro komórkowe – „ditoblast”, który nigdy nie leży wewnątrz komórki, ale zawsze jest zamknięty w ścianie komórkowej „w taki sposób, że ściana komórkowa dzieli się na dwie płytki, z których jedna przechodzi na zewnątrz, a druga wewnątrz cytoblastu. Ten, z którym idzie wewnątrz, zwykle bardziej miękkie i galaretowate” (s. 142). Z rysunków Schleidena widać, jaka to „warstwa wewnętrzna”. Ściana komórkowa» wziął warstwę ciemieniową protoplazmy komórek roślinnych.
Jakie zadanie postawił Schleiden w swojej pracy? „Każda komórka – pisze – prowadzi podwójne życie: zupełnie niezależne, związane jedynie z własnym rozwojem, i drugie zależne, gdyż jest integralną częścią rośliny. Łatwo jednak zauważyć, że zarówno dla fizjologii roślin, jak i dla fizjologii porównawczej procesy życiowe poszczególnych komórek powinny być generalnie na pierwszym miejscu, powinny stanowić nieuniknioną podstawę, iw tym przypadku pytanie jest stawiane przede wszystkim : jak to osobliwe mały organizm, komórka?" (str. 138). To zadanie, geneza komórki, jest podstawą artykułu Schleidena. Moment genetyczny w tym znaczeniu został wysunięty już wcześniej, ale nie można zaprzeczyć, że Schleiden, zgodnie ze swoim czasem, postawił ten problem jaśniej niż jego poprzednicy.
Odpowiadając na postawione pytanie, Schleiden rozwija swoją teorię powstawania komórek. W tej teorii centralną rolę w rozwoju nowych komórek przypisuje się jądru. Jak widzieliśmy, została odkryta na długo przed pracą Schleidena, ale nie doczekała się żadnej definitywnej interpretacji. Według Schleidena jądro jest "cytoblastem" - poprzednikiem komórki. Teorię tworzenia komórek rozwiniętą przez Schleidena można krótko scharakteryzować w następujący sposób.
W dziąśle przylegającym od wewnątrz do ścian wcześniej istniejących komórek powstają ziarna; Schleiden nazywa je śluzem i uważa, że ziarna te w wyniku kondensacji tworzą jąderka, a następnie tworzy się jądro, które pojawia się jako ziarnisty osad wokół jąderka. Z jednej strony na powierzchni jądra z „śluzu” ponownie tworzy się skorupa; ogranicza cytoblast iw ten sposób powstaje otoczona ścianą przestrzeń, w której jądro jest zamknięte w grubości ściany. Ta przestrzeń to nowa komórka. Dlatego według Schleidena komórki potomne powstają wewnątrz komórek macierzystych. Schleiden nie omawia liczby nowych komórek, które mogą rozwinąć się w jednej matczynej komórce, ani losu tej matczynej komórki edukacyjnej.
To jest istota teorii powstawania komórek, istota „doskonałych badań Schleidena, które rzuciły tak wiele światła na ten obszar” – charakterystyka pracy Schleidena podana przez Theodora Schwanna. Jak się wkrótce okazało, teoria Schleidena opiera się na źle zinterpretowanych obserwacjach. To właśnie ta błędna teoria powstawania komórek została przejęta od Schleidena przez jego przyjaciela Schwanna i była to najsłabszy punkt w nauczaniu Schwanna. Sachs w swojej historii botaniki charakteryzuje teorię Schleidena następującymi ostrymi słowami: „Teoria Schleidena o tworzeniu komórek powstała z niezrozumiałego zbiegu niejasnych obserwacji i z góry przyjętych opinii, co więcej, bardzo przypomina głównie stare teorie Sprengla i Treviranus” ( s. 76). Sam Schleiden uparcie bronił swojej teorii cytogenezy i cytował ją nawet w 4. wyd. „Podstawy botaniki naukowej” (1861).
W swoim artykule Schleiden, oprócz rozważanej teorii powstawania komórek, zajmuje się rozwojem zgrubień na ścianach naczyń spiralnych oraz rozwija teoretyczne argumenty dotyczące pracy roślin. Praca Schleidena nie zawiera w tej sekcji niczego zasadniczo nowego, a ponieważ ta część artykułu nie jest bezpośrednio związana z naszym tematem, nie ma potrzeby się nad nią rozwodzić.
Jaką ocenę należy wystawić aspekt historyczny Rola Schleidena w rozwoju nauki o komórkach? Martin Heidenhain (M. Heidenhain, 1899) pod koniec ubiegłego wieku zauważył błędność idei równoważnej wartości w historii komórkowej teorii Schleidena i Schwanna. Później kwestia ta została ponownie podniesiona z całą stanowczością na podstawie krytycznej analizy literatury czeskiego histologa Studnichki (1933), wielkiego znawcy historii teorii komórkowej. Rzeczywiście, tradycyjne porównanie nazwisk Schleiden i Schwann, zwykle przedstawianych jako „współautorzy” teorii komórkowej, nie znajduje uzasadnienia w uważnej analizie źródeł. Schleiden nie był współautorem teorii komórki; on, jak widzieliśmy, był całkowicie obcy głównej idei tej teorii - jedności mikroskopijnej elementarnej struktury zwierząt i roślin; nie jest twórcą doktryny komórkowej w dziedzinie botaniki, ponieważ główne założenia tej doktryny zostały opracowane przed nim. Należy to podkreślić, gdyż w literaturze, zarówno zagranicznej, jak i naszej, wokół nazwiska Schleiden narosła „legenda”, której w historii nauki jest tak wiele z powodu niedostatecznej znajomości oryginałów. Studnichka w cytowanym wyżej artykule o Schleidenie przytoczył fragmenty kilkudziesięciu zagranicznych podręczników z zakresu histologii i biologii, a nawet specjalnych artykułów z historii nauki o komórkach, gdzie rola Schleidena jest całkowicie fałszywie przedstawiona i powtarzana jest legenda, że nauka zawdzięcza odkrycie Schleidenowi struktura komórkowa w roślinach, że Schleiden i Schwann stworzyli teorię komórkową itp. Do listy nieuzasadnionych, a czasem wręcz śmiesznych stwierdzeń o roli Schleidena, na które powoływał się Studnich, można niestety dodać pokaźną listę cytatów z nowszych podręczników i nawet dzieła specjalne, w tym pisma, które twierdzą, że są dziełami historycznymi, zarówno w naszym, jak iw naszym kraju literatura zagraniczna. Historyczna rola pracy Schleidena jest niezaprzeczalna, ale ta rola różni się od tego, co zwykle jest omawiane. Schleidenowi przypisuje się wprowadzenie podejścia genetycznego do teorii tkanek i komórek. Próby podobnego podejścia podejmowano przed Schleidenem (Wolf, Mirbel, Sprengel, Treviranus; w histologii zwierząt – Valentin), ale w tym czasie nie mogły one być tak skuteczne jak prace Schleidena, które pojawiły się, gdy idea komórki jako podstawowa budowa roślin była już powszechna. Bez podejścia genetycznego Schwann nie mógłby stworzyć spójnej teorii komórkowej, popartej przekonującymi danymi z tamtych czasów. Dopiero odwołując się do historii rozwoju tkanek i komórek, Schwann potrafił wykazać „zgodność” różnych struktur elementarnych, mógł udowodnić ich homologię. Prace Schleidena odegrały oczywiście znaczącą rolę w skierowaniu myśli Schwanna na podobną ścieżkę badań.
Ale to nie wszystko. Aby móc w przekonujący sposób pokazać ich homologię, odwołując się do historii rozwoju struktur elementarnych, konieczne było znalezienie cechy przewodniej i biorąc ją za wiodące ogniwo, rozwikłać plątaninę skomplikowanych relacji między strukturami elementarnymi u zwierząt tkanki. Schwann nauczył się tej przewodniej cechy od Schleidena. To jest rdzeń. Komórki w różne tkaniny mogą na zewnątrz bardzo się od siebie różnić, ale podobieństwo jąder jest uderzające, pomagając w homologacji pozornie odmiennych formacji. Jądro było znane zarówno w komórkach roślinnych, jak iw strukturach zwierzęcych przed Schleidenem. Ale dopiero w jego pracy jądro nabrało znaczenia głównej cechy rozwijającej się komórki. Ten znak służył Schwannowi jako dźwignia, chwytając go, był w stanie stworzyć teorię komórkową.
Takie jest znaczenie Schleidena w historii nauki komórkowej. Nie można go stawiać obok Schwanna, nie był współtwórcą teorii komórkowej, ale jego praca była niezbędnym ogniwem w łańcuchu badań przygotowujących materiał, bez którego geniusz Schwanna byłby być może bezsilny dokonać uogólnień, które sformułował w formie teorii komórkowej. Virchow (1859) trafnie to ujął, wskazując, że Schwann stał „na ramionach” Schleidena.
Jeśli znajdziesz błąd, zaznacz fragment tekstu i kliknij Ctrl+Enter.
Schemat jądra komórkowego B komórka biologiczna, rdzeń(łac. jądro) - organelle komórkowe występujące w większości komórek eukariotycznych i zawierające geny jądrowe, które tworzą bardzo materiał genetyczny. Jądro pełni dwie główne funkcje: kieruje reakcjami chemicznymi w cytoplazmie i przechowuje informacje niezbędne do podziału komórki.
Jądro zostało odkryte około 1833 roku przez botanika Roberta Browna.
Oprócz genomu komórkowego jądro zawiera pewne białka, które regulują odczytywanie informacji genetycznej. Odczyt genów na poziomie jądrowym przyciąga złożone procesy transkrypcja, obróbka pierwotnego mRNA i eksport dojrzałego mRNA do cytoplazmy.
Rdzeń ma zwykle średnicę 8-25 mikrometrów. Otoczona jest podwójną błoną zwaną otoczką jądrową. Pory jądrowe przechodzą przez wewnętrzną i zewnętrzną membranę w pewnych odstępach czasu. Otoczka jądrowa reguluje i ułatwia transport między jądrem a cytoplazmą poprzez rozdzielanie reakcje chemiczne zachodzących w cytoplazmie z reakcji zachodzących w jądrze. Błona zewnętrzna jest ciągła z szorstkim retikulum endoplazmatycznym (RER) i może mieć związane rybosomy. Przestrzeń między dwiema membranami (zwana „przestrzenią okołojądrową”) jest ciągła z prześwitami RER. Strona nuklearna otoczka jądrowa otoczone siecią włókien zwaną laminatami jądrowymi.
Wewnętrzna część Jądro zawiera jedno lub więcej jąderek otoczonych macierzą zwaną nukleoplazmą. Nukleoplazma to żelowata ciecz (podobna pod tym względem do cytoplazmy), w której rozpuszczonych jest wiele substancji. Substancje te obejmują trifosforan nukleotydu, cząsteczki sygnałowe, DNA, RNA i białka (enzymy i włókna).
Jądro i układ błony wewnętrznej Materiał genetyczny występuje w jądrze w postaci chromatyny, czyli kompleksu białka i DNA. DNA występuje jako cała linia dyskretne molekuły zwane chromosomami. Istnieją dwa rodzaje chromatyny: euchromatyna i heterochromatyna. Euchromatyna jest mniej zwartą formą DNA, regiony DNA w postaci euchromatyny zawierają geny, które często są odczytywane przez komórkę.
W heterochromatynie DNA jest upakowane bardziej zwięźle. Regiony DNA w postaci heterochromatyny zawierają geny, które nie są odczytywane przez komórkę na tym etapie rozwoju (ten typ heterochromatyny jest znany jako heterochromatyna fakultatywna) lub są to regiony tworzące telomery i centromery chromosomów (ten typ heterochromatyny jest znana jako konstruktywna heterochromatyna). Na Organizmy wielokomórkowe, komórki są niezwykle wyspecjalizowane do wykonywania określonych funkcji, więc potrzebne i odczytywane są różne zestawy genów. Dlatego regiony DNA, które są w postaci heterochromatyny, zależą od typu komórki.
Jąderko jest gęstą strukturą w jądrze, w której gromadzą się elementy rybosomu. Jądro może mieć jedno lub więcej jąder.
III. Jądro komórkowe.
Jądro zostało odkryte i opisane w 1831 roku przez angielskiego botanika R. Browna.
Jest charakterystyczny dla wszystkich komórek eukariotycznych, z wyjątkiem dojrzałych erytrocytów ssaków i roślinnych rurek sitowych. Komórki mogą być jednojądrzaste (zwykle); wielojądrzaste (komórki wątroby i mięśni u ludzi i ssaków, grzyby itp.). Kształt i wielkość jądra komórkowego jest bardzo zmienna i zależy od typu organizmu, a także od rodzaju, wieku i stan funkcjonalny komórki. W komórkach okrągłych lub wielokątnych jest zwykle kulisty, w wydłużonych komórkach ma kształt pręta lub owalny, w leukocytach ma kształt łapy lub nawet wielopłatowy (w komórkach gruczołów pajęczynówki niektórych owadów i pająków). Średnica jądra od 3 do 10 mikronów, (5 - 20 mikronów)
Przez skład chemiczny jądro różni się od reszty komórki wysoka zawartość DNA (15 - 30%) i RNA (12%). 99% DNA komórki koncentruje się w jądrze, gdzie wraz z białkami (histonami i niehistonami) tworzy kompleksy - dezoksyrybonukleoproteiny (DNP).
Funkcje jądra:
1) przechowywanie i odtwarzanie informacji genetycznej;
2) regulacja procesów metabolicznych zachodzących w komórce.
Cykl życia komórki (cykl komórkowy) obejmuje dwa okresy:
1) podział, w wyniku którego powstają dwie komórki potomne;
2) okres między dwoma podziałami, który nazywa się - interfaza.
Podczas interfazy komórka przygotowuje się do podziału: komórka rośnie, następuje replikacja DNA i różne substancje niezbędna do rozszczepienia, energia jest magazynowana. W trakcie podziału struktury komórkowe rdzenie ulegają znaczącym zmianom.
W jądrze międzyfazowym znajdują się: błona jądrowa; sok jądrowy; jąderka i chromatyna.
otoczka jądrowa(cardiolemma) jest reprezentowana przez dwie elementarne membrany, pomiędzy którymi się znajduje przestrzeń okołojądrowa . Zewnętrzna błona błony jądrowej, granicząca z hialoplazmą, ma złożoną pofałdowaną strukturę, w niektórych miejscach połączoną z kanałami EPS. Przestrzeń okołojądrowa i forma ER pojedynczy system kanały komunikacyjne. Rybosomy znajdują się na zewnętrznej błonie jądrowej, wewnętrzna błona jest gładka. Błona jądrowa jest przesiąknięta wieloma porami, których średnica wynosi około 30 - 100 nm. Ich liczba zmienia się w zależności od czynności funkcjonalnej jądra. W młodych komórkach zawsze jest więcej porów niż w starych.
Funkcja powłoki jądrowej:
1. regulacja metabolizmu między jądrem a cytoplazmą;
2. funkcja ochronna.
Wymiana substancji między jądrem a cytoplazmą odbywa się na kilka sposobów:
1. Przez pory zapewniające selektywną przepuszczalność;
Przykład: uwolnienie mRNA i podcząstek rybosomu do cytoplazmy lub wejście do jądra rybosomalnych białek, nukleotydów i cząsteczek regulujących aktywność DNA.
2. Przez sznurowanie wypustek i wypukłości otoczki jądrowej.
3. Przez dyfuzję przez otoczkę jądrową.
Pomimo aktywnej wymiany substancji między jądrem a cytoplazmą, błona jądrowa oddziela zawartość jądrową od cytoplazmy, umożliwiając istnienie specjalnego środowiska wewnątrzjądrowego, różniącego się od otaczającej cytoplazmy.
sok nuklearny(nukleoplazma; karioplazma) jest jednorodną masą wypełniającą przestrzeń między strukturami jądra (chromatyną i jąderkiem). Składa się z białek (enzymów), nukleotydów, aminokwasów i Różne rodzaje RNA (mRNA, tRNA, rRNA) i DNA oraz pośrednie produkty przemiany materii. Sok jądrowy realizuje związek struktur jądrowych i wymianę z cytoplazmą komórki.
chromatyna reprezentuje dezoksyrybonukleoproteina(DNP), wykrywany pod mikroskopem świetlnym w postaci cienkich pasm (fibryli) małych granulek lub grudek. Podstawą chromatyny są nukleoproteiny - długie nitkowate cząsteczki DNA (około 40%), połączone ze specyficznymi białka - histony(40%) zawiera duża liczba lizyna i arginina.
W procesie rozszczepienia jądrowego nukleoproteiny spiralizują się, skracają, w ten sposób zagęszczając (100-500 razy) w zwarte chromosomy w kształcie prętów.
Chromosomy(grecki chrom – farba i soma – ciało) zostały tak nazwane w związku ze zdolnością do intensywnego barwienia. Chromosomy zawierają również RNA, kwaśne białka, lipidy i minerały(jony Ca i Mg), a także enzym polimeraza DNA, niezbędny do replikacji DNA. Podczas interfazy chromosomy są widoczne tylko pod spodem mikroskop elektronowy i jest DNP zwanym chromatyną (stan despiralizacji chromosomów).
Chromosom metafazowy składa się z dwóch podłużnych pasm DNP - chromatyd, połączonych ze sobą w obszarze pierwotnego przewężenia - centromery . W centralnej części centromeru znajdują się kinetochory do którego przyczepiają się mikrotubule włókien wrzeciona rozszczepienia podczas mitozy. Niektóre chromosomy mają zwężenie wtórne, niezwiązane z mocowaniem gwintu wrzeciona. Ten region chromosomów kontroluje syntezę jąderka (organizatora jąderka) i nazywany jest satelitą. Centromer dzieli chromosom na dwa ramiona.
Położenie centromeru determinuje 3 główne typy chromosomów:
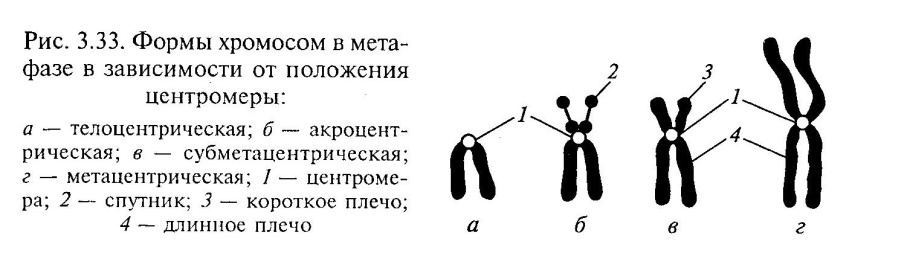 równe ramię
(metacentryczny) - z ramionami równej długości;
równe ramię
(metacentryczny) - z ramionami równej długości;
Submetacentryczny lub nierówne ramiona - ramiona o nierównej długości;
w kształcie pręta- z jednym długim i drugim bardzo krótkim ramieniem (akrocentrycznym).
Struktura chromosomów jest wyraźnie widoczna na etapie metafazy mitozy. Badanie chromosomów pozwoliło ustalić następujące fakty:
ü We wszystkich komórkach somatycznych dowolnego organizmu roślinnego lub zwierzęcego liczba chromosomów jest taka sama;
ü Komórki płciowe zawsze zawierają o połowę mniej chromosomów niż komórki somatyczne danego typu organizmu;
ü Wszystkie organizmy należące do tego samego gatunku mają taką samą liczbę chromosomów w komórkach.
Przykład: glisty końskie - 2; Mucha Drosophila - 8; Szympansy - 48; Popiół - 46; Mężczyzna - 46; Gołąb - 80; Karp - 104; Radiolaria (protista) - 1000 - 1600.
Liczba chromosomów nie zależy od wysokości organizacji i nie zawsze wskazuje na pokrewieństwo filogenetyczne: taką samą liczbę można znaleźć u gatunków systematycznie bardzo odległych od siebie i znacznie różniących się od organizmów o bliskim pochodzeniu. Liczba chromosomów nie jest zatem cechą specyficzną dla gatunku. Jednak charakterystyka zestawu chromosomów jako całości jest specyficzna dla gatunku, tj. charakterystyczne tylko dla jednego rodzaju roślin lub zwierząt.
Kariotyp- zestaw cech ilościowych (liczba i rozmiar) i jakościowych (kształt) zestawu chromosomów komórki somatycznej. Liczba chromosomów w kariotypie jest zawsze parzysta. Wynika to z faktu, że w komórkach somatycznych występują 2 chromosomy o tym samym kształcie i wielkości. Jeden pochodzi z organizmu ojca, drugi z organizmu matki.
Nazywa się chromosomy, które mają ten sam rozmiar i kształt oraz zawierają te same geny homologiczny.
Nazywa się liczbę chromosomów w dojrzałych komórkach rozrodczych haploidalny (pojedynczy) i jest oznaczony List łaciński N. Komórki somatyczne zawierają podwójną liczbę chromosomów - zestaw diploidalny - oznaczone 2n. Komórki posiadające więcej niż dwa zestawy chromosomów to tzw poliploidalny (4n, 8n, 16n itd.)
Jąderka - są zaokrąglone, silnie zwarte, nie ograniczone obszarami błony jądra komórkowego o średnicy 1–2 μm lub większej. Kształt, wielkość i liczba jąderek zależy od stanu funkcjonalnego jądra: im większe jąderko, tym wyższa jest jego aktywność. Jądro może zawierać od 1 do 10 jąderek, a czasami, na przykład, w jądrach drożdży, są one całkowicie nieobecne. Jąderko zawiera około 80% białka, 10-15% RNA, trochę DNA i inne składniki chemiczne.
Jąderka występują tylko w jądrach niedzielących się, podczas podziału zanikają, a po zakończeniu podziału ponownie tworzą się wokół pewnych odcinków chromosomów - geny zwane organizatorami jąderkowymi.
W jąderku RNA łączy się z białkami, w wyniku czego powstają rybonukleoproteiny - prekursory rybosomów. Te ostatnie przechodzą przez pory błony jądrowej do cytoplazmy, gdzie kończy się ich tworzenie. Zatem jąderko jest akumulacją rRNA i rybosomów różne etapy formacje.
Jądro zostało odkryte i opisane w 1833 roku przez Anglika R. Browna. Jądro jest obecne we wszystkich komórki eukariotyczne, z wyjątkiem dojrzałych erytrocytów i rurek sitowych roślin. Komórki zwykle mają jedno jądro, ale czasami można znaleźć komórki wielojądrzaste.
Jądro jest kuliste lub owalny kształt. Niektóre komórki mają jądra podzielone na segmenty. Wielkość jąder wynosi od 3 do 10 mikronów średnicy.
Jądro jest niezbędne do życia komórki. Reguluje aktywność komórek. Jądro przechowuje informacje dziedziczne zawarte w DNA. Ta informacja, dzięki jądru, jest przekazywana do komórek potomnych podczas podziału komórki. Jądro determinuje specyficzność białek syntetyzowanych w komórce. Jądro zawiera wiele białek niezbędnych do jego funkcji. W jądrze syntetyzowany jest RNA.
Jądro ma błonę jądrową, która oddziela je od cytoplazmy, karioplazmy (sok jądrowy), jednego lub więcej jąderek, chromatyny
Otoczka jądrowa składa się z dwóch błon. Ma pory, które się bawią ważna rola w transporcie substancji do i z cytoplazmy. Pory nie są trwałymi formacjami. Ich liczba zmienia się w zależności od czynności funkcjonalnej jądra. Liczba porów wzrasta w okresie największej aktywności jądrowej. Otoczka jądrowa jest bezpośrednio połączona z retikulum endoplazmatycznym.
NA zewnętrzna męmbrana otoczka jądrowa, z poza istnieją rybosomy, które syntetyzują określone białka, które powstają tylko na rybosomach otoczki jądrowej.
Sok jądrowy (karioplazma) - wewnętrzna zawartość jądra, jest roztworem białek, nukleotydów, jonów, bardziej lepkim niż hialoplazma. W nim
obecne są również białka fibrylarne. Karioplazma zawiera jądra i chromatynę. Sok jądrowy zapewnia normalne funkcjonowanie materiał genetyczny.
Jądra są obowiązkowym składnikiem jądra, znajdują się w jądrach międzyfazowych i są małymi ciałami o kulistym kształcie. Jąderka są gęstsze niż jądro. W jąderkach zachodzi synteza rRNA, innych rodzajów RNA oraz tworzenie podjednostek rybosomów.
Pojawienie się jąderek jest związane z pewnymi strefami chromosomów zwanymi organizatorami jąderkowymi. Liczba jąderek jest określona przez liczbę organizatorów jąderek. Zawierają geny rRNA.
Chromatyna (materiał wybarwiony) jest gęstą substancją jądra, dobrze wybarwioną barwnikami zasadowymi. Chromatyna składa się z cząsteczek DNA w kompleksie z białkami (histonami i niehistonami), RNA.
W jądrach niedzielących się (interfazowych) chromatyna może równomiernie wypełniać objętość jądra, będąc w stanie rozłożonym. Ta rozproszona chromatyna (euchromatyna) jest aktywna genetycznie. Cząsteczki DNA zawierające informacje dziedziczne są zdolne do podwojenia się podczas replikacji i możliwy jest transfer (transkrypcja) informacji genetycznej z DNA na mRNA.
Czasami w jądrach interfazowych widoczne są skupiska chromatyny, które są obszarami skondensowanej chromatyny (heterochromatyny). To są obszary nieaktywne. Na przykład w komórkach kobiece ciało, gdzie obecne są dwa chromosomy X, jeden jest w aktywnym stanie rozproszonym, a drugi w nieaktywnym, skondensowanym stanie.
Podczas podziału jądrowego chromatyna intensywniej się wybarwia, skrapla - tworząc bardziej spiralne (skręcone) nici, zwane chromosomami.
Chromosomy są syntetycznie nieaktywne. Strukturę chromosomów najlepiej badać w momencie ich największej kondensacji, czyli w metafazie i na początku anafazy mitozy.
Każdy chromosom w metafazie mitozy składa się z dwóch chromatyd powstałych w wyniku reduplikacji i połączonych centromerem (przewężenie pierwotne). W centralnej części centromeru znajdują się kinetochory, do których podczas mitozy przyczepiają się mikrotubule włókien wrzeciona (ryc. 47). W anafazie chromatydy są oddzielone od siebie. Tworzą się z nich chromosomy potomne zawierające tę samą informację genetyczną. Centromer dzieli chromosom na dwa ramiona. Chromosomy z ramionami równymi nazywane są ramionami równymi lub metacentrycznymi, z ramionami nierównej długości - ramionami nierównymi - submetacentrycznymi, z jednym krótkim, a drugim prawie niezauważalnym - pręcikowatym lub akrocentrycznym.
Niektóre chromosomy mają wtórne przewężenie, które oddziela satelitę. Wtórne przewężenia nazywane są organizatorami jąderkowymi. W nich, w interfazie, dochodzi do powstawania jąderka. Organizatorzy jąderka zawierają DNA
odpowiedzialny za syntezę rRNA. Ramiona chromosomów kończą się regionami zwanymi telomerami, które nie mogą łączyć się z innymi chromosomami.
Kinetochory znajdują się w regionie centromerowym chromosomów. / - kinetochor, 2 - wiązka mikrotubul kinetochoru; 3 - chromatyda.
Liczba, wielkość i kształt chromosomów w zestawie różne rodzaje może się różnić. Zestaw cech zestawu chromosomów nazywa się kariotypem.
Zestaw chromosomów jest specyficzny i stały dla osobników każdego gatunku. Ludzie mają 46 chromosomów, myszy mają 40 chromosomów i tak dalej.
W komórkach somatycznych, które mają diploidalny zestaw chromosomów, chromosomy są sparowane. Nazywa się je homologiczne. Jeden chromosom w parze pochodzi z organizm matki, drugi - od ojca.
Zmiany w strukturze chromosomów lub ich liczbie wynikają z mutacji.
Każda para chromosomów w zestawie jest indywidualna. Chromosomy z różnych par nazywane są niehomologicznymi.
W kariotypie wyróżnia się chromosomy płciowe (u ludzi jest to chromosom X i chromosom Y) i autosomy (cała reszta).
Komórki płciowe mają zbiór haploidalny chromosomy.
Podstawą chromosomu jest cząsteczka DNA związana z białkami (histonami itp.) w nukleoproteinę.
Główne stanowisko biologii molekularnej sformułowane przez F. Cricka głosi, że przekazywanie informacji genetycznej odbywa się:
1) z DNA na DNA przez replikację;
2) od DNA przez mRNA (mRNA) do białka.
Proces samoreprodukcji makrocząsteczek kwasów nukleinowych (replikacja) zapewnia dokładne kopiowanie informacji genetycznej i jej przekazywanie z pokolenia na pokolenie.
Zasada komplementarności leżąca u podstaw budowy cząsteczki DNA pozwala zrozumieć, w jaki sposób syntetyzowane są nowe cząsteczki w okresie syntezy interfazy koło życia komórki przed podziałem.