الميتوكوندريا.يوجد في سيتوبلازم الخلايا الحيوانية والنباتية ما يسمى بعضيات الطاقة - الميتوكوندريا (من الكلمة اليونانية "mitos" - الخيط ، "chondrion" - الحبوب). ويختلف شكل الميتوكوندريا، فهي يمكن أن تكون بيضاوية، على شكل قضيب، على شكل خيط بمتوسط قطر 1 ميكرومتر وطول 7 ميكرومتر. ويعتمد عدد الميتوكوندريا على النشاط الوظيفي للخلية ويمكن أن يصل إلى عشرات الآلاف في عضلات طيران الحشرات.
الهيكل الداخليتمت دراسة الميتوكوندريا (الشكل 11، 12، 15) باستخدام المجهر الإلكتروني. تظهر الصور المجهرية الإلكترونية أن الميتوكوندريا محاطة خارجيًا بغشاء خارجي، والذي له نفس بنية غشاء البلازما. يوجد تحت الغشاء الخارجي غشاء داخلي يشكل طيات عديدة - أعراف. يوجد داخل الميتوكوندريا الحمض النووي الريبي (RNA) والحمض النووي (DNA) والريبوسومات، والتي تختلف عن تلك الموجودة في السيتوبلازم. يتم بناء إنزيمات محددة في أغشيتها، والتي يتم من خلالها تحويل الطاقة إلى الميتوكوندريا العناصر الغذائيةإلى طاقة ATP الضرورية لحياة الخلية والجسم ككل.
أرز. 15. مخطط هيكل الميتوكوندريا
البلاستيدات.هذه هي العضيات المميزة للخلايا النباتية فقط. هناك ثلاثة أنواع من البلاستيدات: البلاستيدات الخضراء الخضراء، والبلاستيدات الملونة الملونة (ولكن ليست الخضراء)، والبلاستيدات البيضاء عديمة اللون.
البلاستيدات الخضراء(الشكل 16) على شكل قرص أو كرة يبلغ قطرها 4-6 ميكرون مع غشاء مزدوج - خارجي وداخلي. يوجد داخل البلاستيدات الخضراء الحمض النووي والريبوسومات وهياكل غشائية خاصة - جرانا، متصلة ببعضها البعض وبالغشاء الداخلي للبلاستيدات الخضراء، تحتوي كل بلاستيدات الخضراء على حوالي 50 جرانا، مرتبة في نمط رقعة الشطرنج لتحسين التقاط الضوء. تحتوي الأغشية الحبيبية على صبغة الكلوروفيل الخضراء. بفضل الكلوروفيل، يتم تحويل الطاقة إلى البلاستيدات الخضراء ضوء الشمسفي الطاقة الكيميائية للATP. يتم استخدام طاقة ATP في البلاستيدات الخضراء للتوليف مركبات العضوية، الكربوهيدرات في المقام الأول.
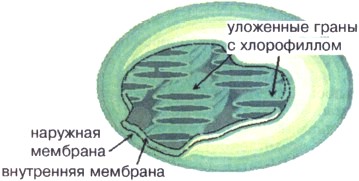
أرز. 16. رسم تخطيطي لهيكل البلاستيدات الخضراء
البلاستيدات الملونة.أصباغ حمراء و اللون الأصفر، الموجودة في البلاستيدات الملونة، تعطي أجزاء مختلفة من النباتات اللون الأحمر والأصفر. يتم تلوين جذور الجزر وثمار الطماطم بسبب الأصباغ الموجودة في البلاستيدات الملونة. مزيج البلاستيدات الملونة التي تحتوي على أصباغ مختلفة يخلق مجموعة واسعة من ألوان الزهور وثمار النباتات.
الكريات البيضهي مكان لتراكم المغذيات الاحتياطية - النشا. يوجد بشكل خاص عدد كبير من الكريات البيض في خلايا درنات البطاطس. في الضوء، يمكن أن تتحول البلاستيدات البيضاء إلى البلاستيدات الخضراء (ونتيجة لذلك تتحول درنات البطاطس إلى اللون الأخضر). في الخريف، تتحول البلاستيدات الخضراء إلى بلاستيدات ملونة وتتحول الأوراق الخضراء والفواكه إلى اللون الأصفر والأحمر.
حركة الخلايا المستمرة. عضيات الحركة.العديد من الخلايا وحيدة الخلية و الكائنات متعددة الخلايالديهم القدرة على التحرك. يشير هذا إلى حركة الخلية في الفضاء والحركة داخل الخلايا لعضياتها. في البيئة السائلة، تتم حركة الخلايا عن طريق حركة السوط والأهداب (الشكل 10، 2، 8، 11). هذه هي الطريقة التي تتحرك بها العديد من الكائنات الحية وحيدة الخلية، على سبيل المثال الحنديرة الخضراء، والسوط، والأهداب، وما إلى ذلك. كما تتحرك بعض أنواع البكتيريا بمساعدة الأسواط الطويلة والمرنة، والتي تدور بسرعة، مما يضمن تقدم الخلية. تتحرك الأميبا وبعض الكائنات الحية البسيطة الأخرى، وكذلك الخلايا المتخصصة متعددة الخلايا (على سبيل المثال، الخلايا الليمفاوية) بمساعدة النواتج المتكونة على سطح الخلايا.
الخلية موجودة حركة مستمرة. أثناء البلعمة واحتساء الخلايا، يخترق غشاء البلازما داخل الخلية، وتتحرك الجسيمات الحالة، وحويصلات مجمع جولجي، وتتحرك الميتوكوندريا، وأخيرًا يتحرك السيتوبلازم نفسه.
يتم ضمان الحركة الخلوية بواسطة هيكل خلوي يتكون من الأنابيب الدقيقة والألياف الدقيقة والألياف الدقيقة مركز الخلية. الأنابيب الدقيقة عبارة عن أسطوانات طويلة مجوفة يبلغ قطرها 25 نانومتر، وتتكون جدرانها من البروتينات. تتكون سوط وأهداب الخلايا الحيوانية والنباتية من أنابيب دقيقة متوازية. تختلف الأسواط عن الأهداب في الطول فقط. وبالتالي، فإن الحيوانات المنوية لدى الثدييات لديها سوط واحد يصل طوله إلى 100 ميكرون. الأهداب أقصر بعشر مرات من الأسواط، ويوجد عدة آلاف من الأهداب في الخلية الواحدة. الخيوط الدقيقة عبارة عن هياكل رفيعة جدًا تتكون من آلاف جزيئات بروتين الأكتين المرتبطة ببعضها البعض. في الخلايا العضلية، يتم تضمينها مع خيوط البروتين الأخرى - الميوسين، في المجمعات التي تضمن الوظيفة الانقباضية لهذه الخلايا.
يوجد في سيتوبلازم خلايا جميع الكائنات الحية، بالقرب من النواة، مركز خلوي يشارك في انقسام الخلايا. يتضمن مركز الخلية للخلايا الحيوانية وبعض النباتات مركزًا مركزيًا (الشكل 11، 3). Centriole هو تشكيل مقترن. يحتوي على اسطوانتين تتكونان من الأنابيب الدقيقة وتقعان بشكل متعامد مع بعضهما البعض.
الادراج الخلوية. وأخيرا، ينبغي أن يقال عن الادراج العديدة في السيتوبلازم. الشوائب هي هياكل غير دائمة من السيتوبلازم، والتي، على عكس العضيات، إما تظهر أو تختفي خلال حياة الخلية. تحتوي الشوائب الكثيفة على شكل حبيبات على مواد مغذية احتياطية (النشا والبروتينات والسكريات والدهون) أو نفايات الخلية، والتي لا يمكن إزالتها على الفور لسبب أو لآخر. تتمتع جميع بلاستيدات الخلايا النباتية بالقدرة على تصنيع وتجميع العناصر الغذائية الاحتياطية.
في زرع الخلايايحدث تراكم العناصر الغذائية الاحتياطية أيضًا في الفجوات - الأكياس الغشائية ذات محلول مائيالأملاح والمركبات العضوية، والتي غالبًا ما تشغل كامل حجم الخلية تقريبًا، مما يدفع النواة والسيتوبلازم إلى غشاء البلازما.
- لماذا تسمى الميتوكوندريا "محطات الطاقة" في الخلية؟
- ما هي الهياكل الخلوية التي تساهم في حركتها؟
- ما هي الادراج الخلوية؟ ما هو دورهم في الخلية؟
- وصف السمات الهيكلية للميتوكوندريا والبلاستيدات الخضراء فيما يتعلق بوظائفها في الخلية.
مركز الخلية- عضية مرئية تحت المجهر الضوئي في الخلايا الحيوانية و النباتات السفلى. يقع عادةً بالقرب من النواة أو في المركز الهندسي للخلية ويتكون من جسمين مركزيين على شكل قضيب، يبلغ حجمهما حوالي 0.3-1 ميكرومتر. تحت المجهر الإلكتروني، ثبت أن المركز المركزي عبارة عن أسطوانة، جدرانها مبنية بواسطة تسعة أزواج من الأنابيب الرفيعة جدًا. يوجد في منتصف الأسطوانة تجويف مملوء بكتلة متجانسة. زوج من المريكزات محاط بمنطقة أخف - الغلاف المركزي.
يلعب مركز الخلية دور مهمأثناء حركة الكروموسومات أثناء الانقسام. ترتبط به قدرة بعض الخلايا على التحرك بنشاط. يتم إثبات ذلك من خلال حقيقة أنه عند قاعدة السوط أو أهداب الخلايا المتحركة (الأوالي والحيوانات المنوية) توجد تكوينات لها نفس بنية مركز الخلية.
عضيات الحركة.يمكن للخلايا أن تتحرك باستخدام عضيات متخصصة، والتي تشمل الأهداب والأسواط. أهداب الخلايا دائمًا عديدة (في الأوليات يبلغ عددها المئات والآلاف) ويبلغ طولها 10-15 ميكرون. غالبًا ما يكون هناك 1-8 سوط ، طولها 20-50 ميكرون. يتشابه هيكل الأهداب والسوط في كل من الخلايا النباتية والحيوانية. تحت المجهر الإلكتروني، تم اكتشاف أن الأنابيب الدقيقة تمتد على طولها بالكامل. يقع اثنان منهم في المركز، ومن حولهم في المحيط يوجد 9 أزواج أخرى من الأنابيب الدقيقة. هذا الهيكل بأكمله مغطى تذكر الذكريات، وهو استمرار لغشاء الخلية. لا تضمن حركة السوط والأهداب حركة الخلايا في الفضاء فحسب، بل تضمن أيضًا الحركة مواد مختلفةعلى سطح الخلايا، وكذلك دخول جزيئات الطعام إلى داخل الخلية. يوجد في قاعدة الأهداب والسوط أجسام قاعدية تتكون أيضًا من الأنابيب الدقيقة. ويعتقد أن الأجسام القاعدية هي مركز تكوين الأنابيب الدقيقة من الأسواط والأهداب. غالبًا ما تنشأ الأجسام القاعدية بدورها من مركز الخلية.
لا يحتوي عدد كبير من الكائنات وحيدة الخلية وبعض الخلايا متعددة الخلايا على عضيات خاصة للحركة والتحرك بمساعدة الأرجل الكاذبة (الأرجل الكاذبة). تسمى الحركة بمساعدة الأرجل الكاذبة بالحركة الأميبية. يعتمد على حركة جزيئات بروتينات خاصة تسمى البروتينات المقلصة.
الادراجتوجد غالبًا في الخلايا النباتية وتختلف عن العضيات من حيث أنها تكوينات مؤقتة وغير دائمة. تعتمد كميتها على شدة عملية التمثيل الغذائي وحالة الجسم. وعادة ما تكون مرئية تحت المجهر الضوئي على شكل حبيبات أو قطرات ذات أحجام وأشكال مختلفة. بواسطة التركيب الكيميائيالتمييز: الكربوهيدرات والدهون والبروتينات. تبدو شوائب الكربوهيدرات والبروتين مثل الحبوب، بينما تبدو شوائب الدهون مثل القطرات. في الخلايا النباتية، غالبا ما يتم تخزين الكربوهيدرات في شكل حبيبات النشا، وفي الخلايا الحيوانية - الجليكوجين. توجد الحبوب البروتينية بكميات كبيرة في سيتوبلازم بيض الحيوانات على شكل صفار. هناك الكثير منهم في بذور النباتات. قطرات الدهون موجودة في الخلايا النسيج الضامالحيوانات وفي بذور النباتات. توجد شوائب بلورية (أملاح) في الخلايا النباتية الأحماض العضوية). في ظل ظروف معينة، يمكن للخلية استخدام جميع أنواع الادراج خلال حياتها، ثم تتراكم مرة أخرى.
توجد في سيتوبلازم العديد من الخلايا النباتية تكوينات خاصة - فجوات. وهي خزانات مملوءة بخليط من محاليل المواد الكيميائية المختلفة. تتشكل فجوات كبيرة بشكل خاص في الخلايا القديمة.
|
معايير للمقارنة |
أنواع الريبوسومات |
|
|
نوع بدائية النواة |
نوع حقيقي النواة |
|
|
العثور على في الخلايا |
ريبوسومات من البكتيريا الحقيقية والبكتيريا الزرقاء الريبوسومات البلاستيدية الميتوكوندريا الريبوسومات ريبوسومات البكتيريا العتيقة (شبيهة بالريبوسومات حقيقية النواة) |
السيتوبلازم الريبوسومات في حقيقيات النوى: الحيوانات، والنباتات |
|
نسبة الحمض النووي الريبي: البروتين | ||
|
كمية الرنا الريباسي وحدة فرعية |
طول الجزيء 1500-1600 نيوكليوتيدات (الحد الأدنى - 950 في ريبوسومات الميتوكوندريا) |
طول الجزيء الواحد حوالي 1800 نيوكليوتيدات |
|
كمية الرنا الريباسي بشكل كبير وحدة فرعية |
2 جزيئات: جزيء واحد يبلغ طوله حوالي 3000 نيوكليوتيدات، جزيء واحد يبلغ طوله حوالي 120 نيوكليوتيدات |
3 جزيئات: جزيء واحد يبلغ طوله حوالي 4-5 آلاف نيوكليوتيدات، جزيء واحد يبلغ طوله حوالي 120 نيوكليوتيدات، جزيء واحد يبلغ طوله حوالي 160 نيوكليوتيدات (يشبه هذا الجزيء أحد أقسام الرنا الريباسي النووي بدائية النواة) |
|
البروتين (جميع الجزيئات تقريبًا فريدة من نوعها) |
حوالي 20 جزيء في الوحدة الفرعية الصغيرة و30 جزيء في الوحدة الفرعية الكبيرة |
ما يقرب من 30 جزيء لكل وحدة فرعية صغيرة و 40 جزيءًا في الوحدة الكبيرة |
مركز الخلية
مركز الخلية عبارة عن عضية تتحكم في تكوين ونمو الأنابيب الدقيقة للهيكل الخلوي للخلية حقيقية النواة.(الشكل 38)
أرز. 38. مركز الخلية 1 – ثلاثة توائم من الأنابيب الدقيقة. 2 – المتحدث شعاعي. 3 – الهيكل المركزي لـ”عجلة العربة”؛ 4 – القمر الصناعي. 5 – الليزوزوم. 6 – ديكتيوسومات مجمع جولجي. 7 – فقاعة تحدها. 8 – خزان الشبكة الإندوبلازمية الحبيبية. 10 – الميتوكوندريا. 11 - الجسم المتبقي. 12 – الأنابيب الدقيقة. 13- كاريوتيكا (حسب ر. كرستيتش مع التعديلات).
تتمثل الوظائف الرئيسية لمركز الخلية في تكوين أنظمة الأنابيب الدقيقة الهيكلية الخلوية وعضيات الحركة والمغزل.
أساس مركز الخلية هو زوج من المريكزات (أحيانًا يصل عددها إلى 4-6-8 في الخلايا متعددة الصبغيات)، وتقع في وسط الخلية.
تم اكتشاف المريكزات ووصفها لأول مرة بواسطة فليمنج (1875)، ولكن تمت دراستها بالتفصيل فقط باستخدام المجهر الإلكتروني. تظهر المريكزات ارتباطًا وثيقًا بـ المغلف النووي، وفي العديد من حقيقيات النوى السفلية (الأوليات، الطحالب، الفطريات، mesokaryotes)، يتم بناء المريكزات في الغلاف النووي.
يوجد مركز الخلية دائمًا في خلايا الحيوانات متعددة الخلايا. تفتقر بدائيات النوى إلى مركز الخلية والتوبولين. في خلايا الفطريات والطحالب والحيوانات وحيدة الخلية، لا يوجد مركز الخلية دائمًا، ولكن في الخلايا نباتات أعلىلم يتم الكشف عنها (مع استثناءات نادرة). في حالة عدم وجود مركز خلية، يتم تنفيذ وظائفها في حقيقيات النوى بواسطة مركز تكوين الأنابيب الدقيقة - TsOMT.
مريكزإنها أسطوانة مجوفة يبلغ قطرها حوالي 0.15 ميكرون وطولها 0.3-0.5 ميكرون (في كثير من الأحيان - عدة ميكرونات). تتكون الجدران المركزية من 9 ثلاثة توائمأنابيب توبولين دقيقة متصلة بواسطة جسور داينين. عادة، توجد المريكزات في أزواج: أحد المريكزات هو المريكز الأم، والآخر هو المريكز الابنة؛ يقع المريكز الابنة بشكل عمودي على المريكز الأم. مثل هذا الزوج من المريكزات - ثنائي- له شكل T أو شكل L. يحيط بالدبلوسوم طبقة كثيفة من السيتوبلازم تتشكل الغلاف الجوي، أو الجسيم المركزي. أثناء انقسام الخلايا، تمتد خيوط كثيفة من الأنابيب الدقيقة من الغلاف المركزي، وتتشكل الغلاف الفلكي.
يحدث تضاعف المريكز تقريبًا في منتصف الطور البيني: ينفصل المريكز الأم عن المريكز الابنة، وينمو مريكز جديد من كل مريكز. ش مجموعات مختلفةالكائنات الحية، وهذه العملية لها خصائصها الخاصة.
بالإضافة إلى العضيات، تحتوي الخلايا على شوائب خلوية. ويمكن احتواؤها ليس فقط في السيتوبلازم، ولكن أيضًا في بعض العضيات، مثل الميتوكوندريا والبلاستيدات.
ما هي الادراج الخلوية؟
هذه تشكيلات ليست دائمة. وعلى عكس الكائنات العضوية، فهي ليست مستقرة. بالإضافة إلى ذلك، لديهم بنية أبسط بكثير وأداء وظائف سلبية، مثل النسخ الاحتياطي.
كيف يتم بناؤها؟
معظمها على شكل قطرة، لكن بعضها قد يكون مختلفًا، على سبيل المثال، يشبه اللطخة. أما بالنسبة للأحجام، فقد تختلف. يمكن أن تكون الشوائب الخلوية أصغر من العضيات، بنفس الحجم أو أكبر.
وهي تتكون أساسًا من مادة واحدة محددة، تكون عضوية في معظم الحالات. يمكن أن يكون إما دهون أو كربوهيدرات أو بروتين. 
تصنيف
اعتمادًا على مصدر المادة التي تتكون منها، هناك الأنواع التالية من الشوائب الخلوية:
- خارجي.
- ذاتية النمو؛
- منتشر.
يتم بناء الادراج الخلوية الخارجية من مركبات كيميائيةالتي دخلت الخلية من الخارج. تسمى تلك التي تتكون من مواد تنتجها الخلية نفسها بالداخلية. على الرغم من أن الادراج الفيروسية يتم تصنيعها بواسطة الخلية نفسها، إلا أن هذا يحدث نتيجة دخول الحمض النووي الفيروسي إليها. ببساطة تأخذه الخلية من أجل الحمض النووي الخاص بها وتقوم بتصنيع بروتين الفيروس منه.
اعتمادا على الوظائف التي تؤديها الادراج الخلوية، يتم تقسيمها إلى الصباغية والإفرازية والغذائية.
يتم تقسيم الشوائب أيضًا إلى أنواع اعتمادًا على المركبات الكيميائية المحددة التي تتكون منها. 
الادراج الخلوية: الوظائف
يمكن أن يكون لديهم ثلاث وظائف. دعونا ننظر إليهم في الجدول
وهذه كلها وظائف للتكوينات غير الدائمة في الخلية.
شوائب الخلايا الحيوانية
يحتوي السيتوبلازم في الحيوان على شوائب غذائية وصبغية. تحتوي بعض الخلايا أيضًا على خلايا إفرازية.
الغذائية في الخلايا الحيوانية هي إدراج الجليكوجين. لديهم شكل حبيبة بحجم حوالي 70 نانومتر. 
الجليكوجين هو المادة الاحتياطية الرئيسية للحيوان. مثل من هذه المادةيقوم الجسم بتخزين الجلوكوز. هناك نوعان من الهرمونات التي تنظم عملية التمثيل الغذائي للجلوكوز والجلوكوجين: الأنسولين والجلوكاجون. وكلاهما ينتج عن البنكرياس. الأنسولين هو المسؤول عن تكوين الجليكوجين من الجلوكوز، والجلوكاجون، على العكس من ذلك، يشارك في تخليق الجلوكوز.
تم العثور على معظم شوائب الجليكوجين في خلايا الكبد. كما أنها موجودة بكميات كبيرة في العضلات، بما في ذلك القلب. تحتوي شوائب الجليكوجين في خلايا الكبد على شكل حبيبات يبلغ حجمها حوالي 70 نانومتر. يتجمعون في مجموعات صغيرة. شوائب الجليكوجين في الخلايا العضلية ( خلايا العضلات) لها شكل دائري. وهي مفردة وأكبر قليلاً من الريبوسومات.
ومن سمات الخلايا الحيوانية أيضًا شوائب الدهون. هذه أيضًا عبارة عن شوائب غذائية يمكن للجسم من خلالها الحصول على الطاقة طارئ. وهي تتكون من الدهون ولها شكل دمعة. في الأساس، وترد هذه الادراج في خلايا الأنسجة الضامة الدهنية - الخلايا الشحمية. هناك نوعان من الأنسجة الدهنية: الأبيض والبني. تحتوي الخلايا الشحمية البيضاء على قطرة واحدة كبيرة من الدهون، بينما تحتوي الخلايا البنية على العديد من الخلايا الصغيرة.
أما بالنسبة للشوائب الصبغية، فتتميز الخلايا الحيوانية بتلك التي تتكون من الميلانين. وبفضل هذه المادة، يكون لقزحية العين والجلد وأجزاء أخرى من الجسم لون معين. كلما زاد عدد شوائب الميلانين في الخلايا، كلما كانت هذه الخلايا أغمق.
الصباغ الآخر الذي يمكن العثور عليه في الخلايا الحيوانية هو الليبوفوسين. هذه المادة لونها أصفر-بني. يتراكم في عضلة القلب والكبد مع تقدم عمر الأعضاء.
شوائب الخلايا النباتية
كما يتم تضمين الادراج الخلوية، وهيكلها ووظائفها التي ندرسها، أيضًا
الادراج الغذائية الرئيسية في هذه الكائنات هي حبوب النشا. في شكلها، تقوم النباتات بتخزين الجلوكوز. عادةً ما تكون شوائب النشا عدسية أو كروية أو بيضاوية الشكل. يمكن أن يختلف حجمها اعتمادًا على نوع النبات والعضو الذي توجد فيه الخلايا. يمكن أن تتراوح من 2 إلى 100 ميكرون.
شوائب الدهونومن خصائص الخلايا النباتية أيضًا. وهي ثاني أكثر الادراج الغذائية شيوعا. لديهم شكل كروي وغشاء رقيق. وتسمى أحيانا الأجسام الكروية.
شوائب البروتينموجودة فقط في الخلايا النباتية، فهي ليست نموذجية للحيوانات. وهي تتكون من بروتينات بسيطة - البروتينات. هناك نوعان من شوائب البروتين: حبيبات الأليرون والأجسام البروتينية. قد تحتوي حبيبات الأليورون إما على بلورات أو ببساطة على بروتين غير متبلور. لذلك، تسمى الأولى معقدة، والثانية - بسيطة. أما حبيبات الأليرون البسيطة، والتي تتكون من بروتين غير متبلور، فهي أقل شيوعًا.
أما بالنسبة للشوائب الصبغية فتتميز النباتات بـ الكريات البلاستية. تتراكم فيها الكاروتينات. هذه الادراج هي سمة من البلاستيدات.
تتكون الشوائب الخلوية، التي ندرس تركيبها ووظائفها، في الغالب من مركبات كيميائية عضوية، ومع ذلك، توجد في الخلايا النباتية أيضًا تلك التي تتشكل من لا المواد العضوية. هذا بلورات أكسالات الكالسيوم. 
وهي موجودة فقط في فجوات الخلية. يمكن أن تأتي هذه البلورات في مجموعة واسعة من الأشكال، وغالبًا ما تكون فردية لكل فرد. أنواع معينةالنباتات.
غالبًا ما تتمتع الكائنات الحية التي تتكون من خلية واحدة بالقدرة على الحركة بنشاط. آليات الحركة التي نشأت في عملية التطور متنوعة للغاية.
عضيات الحركة هي الأرجل الكاذبة (في الساركوديات وبعض الأسواط) أو الأسواط (في الأسواط) أو الأهداب (في الأهداب).
بالإضافة إلى ذلك، يمكن إجراء الحركة نتيجة لتقلص الألياف الدقيقة الخاصة - myonemes الموجودة تحت الأغطية في السيتوبلازم (في القمة)
حصلت حركة الأميبا على اسمها من أبسط كائن حي، الأميبا. (انظر الملحق رقم 6).أعضاء الحركة في الأميبا هي أرجل زائفة - أرجل كاذبة، وهي نتوءات من السيتوبلازم. يتم تشكيلها في أماكن مختلفةسطح السيتوبلازم. يمكن أن يختفوا ويظهروا في مكان آخر.
أرجل كاذبة(من الزائفة... والقيح اليوناني، gen. p. podos - الساق) (الأرجل الكاذبة)، والنمو السيتوبلازمي المؤقت في الكائنات وحيدة الخلية (جذور الأرجل، السبوروزوان، وما إلى ذلك) وفي بعض الخلايا (كريات الدم البيضاء، البلاعم، وما إلى ذلك) من الخلايا متعددة الخلايا الكائنات الحية. إنها تعمل على نقل والتقاط الطعام والجزيئات الأخرى.
تعتبر الحركة باستخدام الأسواط من سمات العديد من الطحالب أحادية الخلية (على سبيل المثال، الكلاميدوموناس)، والأوالي (على سبيل المثال، الأوجلينا الخضراء) والبكتيريا. أعضاء الحركة في هذه الكائنات هي السوط - نواتج السيتوبلازم على سطح السيتوبلازم.
السوطيات (انظر الملحق رقم 7). مقاسات من 2-5 ميكرون إلى 1 مم. وهي تتحرك بمساعدة واحد أو أكثر من الأسواط، التي قد يختلف طولها من فرد إلى آخر. فقط عدد قليل جدًا من الأنواع يمكن أن تشكل أرجلًا كاذبة. عادة ما يتكاثرون لا جنسيانتيجة للانقسام الطولي للخلايا؛ البعض لديه التكاثر الجنسي. الغالبية العظمى من السوطيات هي أنواع تعيش بحرية وتوجد في المياه البحرية والعذبة، وكذلك في التربة. تلعب أشكال العوالق دورًا مهمًا في دورة المواد؛ العديد من الأنواع هي المؤشرات البيولوجيةتلوث المياه
بنية الخلية.
الخلية هي نظام حي أولي، وهي أساس البنية والنشاط الحيوي لجميع الحيوانات والنباتات. توجد الخلايا ككائنات مستقلة (على سبيل المثال، البروتوزوا والبكتيريا) وكجزء من الكائنات متعددة الخلايا، حيث توجد خلايا جرثومية تعمل على التكاثر، وخلايا الجسم (جسدية)، مختلفة في البنية والوظيفة (على سبيل المثال، الخلايا العصبية والعظمية). ، العضلات ، الإفرازية).
تختلف أحجام الخلايا من 0.1 إلى 0.25 ميكرون (بعض البكتيريا) إلى 155 ملم (بيضة النعام في قشرتها).
خلية أي كائن حي كاملة نظام المعيشة. وهو يتألف من ثلاثة أجزاء مرتبطة بشكل لا ينفصم: الغشاء والسيتوبلازم والنواة. (انظر الملحق رقم 9).
1. غشاء الخلية.
يتفاعل غشاء الخلية بشكل مباشر مع البيئة الخارجية ويتفاعل مع الخلايا المجاورة (في الكائنات متعددة الخلايا).
غشاء الخلية لديه بنية معقدة. ويتكون من طبقة خارجية وغشاء بلازمي يقع تحتها.
تختلف الخلايا الحيوانية والنباتية في بنية طبقتها الخارجية (انظر الملحق رقم 10).في النباتات، وكذلك في البكتيريا والطحالب الخضراء المزرقة والفطريات، يوجد قذيفة كثيفة، أو جدار الخلية. في معظم النباتات يتكون من الألياف. يلعب جدار الخلية دورًا مهمًا للغاية: فهو إطار خارجي وقشرة واقية ويوفر انتفاخًا للخلايا النباتية: يمر الماء والأملاح وجزيئات العديد من المواد العضوية عبر جدار الخلية.
الطبقة الخارجية لسطح الخلايا الحيوانية، على عكس جدران الخلايا النباتية، رقيقة ومرنة للغاية. وهو غير مرئي تحت المجهر الضوئي ويتكون من مجموعة متنوعة من السكريات والبروتينات. تسمى الطبقة السطحية للخلايا الحيوانية مركب السكر.
يؤدي الكأس السكري في المقام الأول وظيفة الاتصال المباشر بين الخلايا الحيوانية والبيئة الخارجية، مع جميع المواد المحيطة بها. نظرًا لسمكها الصغير (أقل من 1 ميكرون)، فإن الطبقة الخارجية للخلايا الحيوانية لا تلعب دورًا داعمًا، وهو ما يميز جدران الخلايا النباتية. يحدث تكوين الجليكوكليكس وكذلك جدران خلايا النباتات بسبب النشاط الحيوي للخلايا نفسها.
2. السيتوبلازم.
السيتوبلازم (من السيتو... والبلازما)، مصطلح "السيتوبلازم" اقترحه إي. ستراسبرجر (1882).
تم ترسيمها من بيئة خارجيةغشاء البلازما، السيتوبلازم هو البيئة الداخلية شبه السائلة للخلايا (انظر الملحق رقم 8).في السيتوبلازم الخلايا حقيقية النواةوتقع النواة والعضيات المختلفة.
تقع النواة في الجزء المركزي من السيتوبلازم. كما أنه يحتوي على شوائب مختلفة - منتجات النشاط الخلوي، والفجوات، وكذلك الأنابيب الصغيرة والخيوط التي تشكل الهيكل العظمي للخلية.
الفجوات (انظر الملحق رقم 8)(فجوة فرنسية، من اللاتينية فراغ - فارغ)، تجاويف في الخلايا الحيوانية والنباتية أو الكائنات وحيدة الخلية. هناك فجوات هضمية ومقلصة (نابضة) تنظم الضغط الأسموزي وتعمل على إزالة الفضلات من الجسم. (انظر الملحق رقم 11).
تسود البروتينات في تكوين المادة الرئيسية للسيتوبلازم.
تتم العمليات الأيضية الرئيسية في السيتوبلازم، فهو يوحد النواة وجميع العضيات في وحدة واحدة، ويضمن تفاعلها، ونشاط الخلية كنظام حي متكامل واحد.
تجريبيًا، من الممكن الحصول على خلايا سيتوبلاستية حية خالية من الأسلحة النووية، والتي يمكنها تصنيع البروتينات والدهون وATP خلال 1-3 أيام. ثم، بالطبع، يموتون بسبب عدم القدرة على تصنيع الحمض النووي الريبي الجديد في غياب النواة.
حجم السيتوبلازم خلايا مختلفةليس هو نفسه: في الخلايا الليمفاوية يساوي تقريبًا حجم النواة، وفي خلايا الكبد يشكل السيتوبلازم 94٪ من إجمالي حجم الخلية.
رسميا، ينقسم السيتوبلازم إلى ثلاثة أجزاء: العضيات، والادراج، والهيالوبلازم.
العضيات- المكونات اللازمة لأي خلية والتي بدونها لا تستطيع الخلية أن تحافظ على وجودها.
تشمل العضيات ذات الغشاء الواحد عضيات الجهاز الفراغي: الشبكة الإندوبلازمية، والجسيمات الحالة، وجهاز جولجي، والبيروكسيسومات، والغشاء البلازمي.
العضيات ذات الغشاء المزدوج هي الميتوكوندريا والبلاستيدات. تتضمن هذه المجموعة أيضًا نواة الخلية.
الملكية المشتركةالعضيات الغشائية هي أنها مبنية من أغشية البروتين الدهني التي تنغلق على نفسها، بحيث تشكل تجاويف مغلقة وبالتالي تقسم السيتوبلازم إلى مجموعات من حجيرات مختلفة وظيفيا.
تشمل العضيات غير الغشائية الريبوسومات والجسيمات المركزية للخلايا الحيوانية.
الادراج_لا توجد دائمًا وتمثل رواسب من المواد الاحتياطية (الجليكوجين، صفار البيض) أو تراكمات المنتجات الأيضية (الأصباغ، بلورات الملح في النباتات).
الهيالوبلازما(من "هولين" - شفاف) - هذه هي البلازما الرئيسية، العصارة الخلوية، صحيح البيئة الداخليةالخلايا. تكوين الهيالوبلازم معقد للغاية، والاتساق قريب من الهلام.
المواد الهلامية- أنظمة غروانية منظمة ذات وسط سائل منتشر، تحت تأثير خارجي أو العوامل الداخليةيمكنهم تغيير حالة التجميع والانتقال إلى مرحلة أكثر سيولة - سول. يمكن أن تحدث تحولات هلامية مماثلة في السيتوبلازم تحت تأثير بروتين الأكتين، وتتغير حالته في أجزاء مختلفة من الخلية، مما يضمن حركة الخلية بأكملها. عندما يتفاعل الأكتين الليفي مع بروتينات مثل الفيمبرين، يستقر الجل، وعندما يرتبط بالبروتينات التي يعتمد نشاطها على تركيز أيونات الكالسيوم (على سبيل المثال، الجلسولين)، يتحول النظام بأكمله إلى حالة سائلة.
يمكن أن يتغير التعبير عن عناصر الهيكل العظمي للخلية، بما في ذلك مكون الأكتين، بشكل ملحوظ خلال دورة الخلية.
الأهمية الوظيفية للسيتوبلازم كبيرة جدًا.
يحتوي الهيالوبلازم، بالإضافة إلى أيونات مختلفة من المركبات غير العضوية، على إنزيمات تشارك في تخليق الأحماض الأمينية والنيوكليوتيدات، الأحماض الدهنيةالسكريات. على الريبوسومات والريبوسومات الموجودة على الأغشية، يتم تصنيع البروتينات المختلفة التي تضمن التمثيل الغذائي الخلوي.
3. الشبكة الإندوبلازمية.
تمتلئ المنطقة الداخلية بأكملها من السيتوبلازم بالعديد من القنوات والتجاويف الصغيرة، جدرانها عبارة عن أغشية مشابهة في هيكلها لغشاء البلازما. تتفرع هذه القنوات وتتصل مع بعضها البعض وتشكل شبكة تسمى الشبكة الإندوبلازمية أو الشبكة الإندوبلازمية.
الشبكة الأندوبلازمية (انظر الملحق رقم 12)غير متجانسة في بنيتها. وهناك نوعان معروفان منه: حبيبي (خشن) وناعم.
يوجد على أغشية القنوات وتجويف الشبكة الحبيبية العديد من الأجسام المستديرة الصغيرة - الريبوسومات التي تعطي الأغشية مظهرًا خشنًا.
لا تحمل أغشية الشبكة الإندوبلازمية الملساء الريبوسومات على سطحها.
تؤدي الشبكة الإندوبلازمية العديد من الوظائف المتنوعة. وتتمثل المهمة الرئيسية للشبكة الإندوبلازمية الحبيبية في المشاركة في تخليق البروتين، والذي يحدث في الريبوسومات.
يحدث تخليق الدهون والكربوهيدرات على أغشية الشبكة الإندوبلازمية الملساء. تتراكم جميع هذه المنتجات التخليقية في القنوات والتجاويف، ثم يتم نقلها إلى عضيات مختلفة في الخلية، حيث يتم استهلاكها أو تتراكم في السيتوبلازم كشوائب خلوية.
تربط الشبكة الإندوبلازمية العضيات الرئيسية للخلية (انظر الملحق رقم 8).
4. الأغشية البيولوجية.
الأغشية البيولوجية ("الغشاء" اللاتيني - الجلد والفيلم) عبارة عن أفلام بروتينية دهنية رقيقة (لا يزيد سمكها عن 10 نانومتر) تتكون من طبقة مزدوجة من جزيئات الدهون، والتي تتضمن جزيئات من بروتينات مختلفة. تقع على سطح الخلايا (غشاء البلازما) والجزيئات داخل الخلايا (النوى، الميتوكوندريا، الخ). من حيث الوزن، اعتمادًا على نوع الغشاء، تبلغ نسبة الدهون 25-60٪. ونسبة البروتينات 40-75%. تحتوي العديد من الأغشية على كربوهيدرات يمكن أن يصل حجمها إلى 2-10٪.
تعتمد صحة الخلية وعمرها إلى حد كبير على حالة الغشاء.
أ) خصائص الأغشية.
نفاذية كاملة للماء. يسمح الغشاء دائمًا بمرور الماء داخل الخلية أو خارجها، اعتمادًا على المكان الذي يكون فيه تركيز الماء أكبر. تسمى حركة المادة من منطقة ذات تركيز عالٍ إلى منطقة ذات تركيز أقل بالانتشار. انتشار المادة لا يتطلب طاقة.
الموصلية الانتقائية للمواد المذابة:
تخترق الجزيئات ذات الشحنة السالبة الغشاء بشكل أسرع وأسهل. تخترق المواد القابلة للذوبان في الدهون الغشاء بسهولة أكبر من المواد القابلة للذوبان في الماء. تخترق الجزيئات الصغيرة الغشاء بسهولة أكبر من الجزيئات الكبيرة.
ب) النقل النشط للمواد.
بعض المواد قادرة على اختراق الغشاء في الاتجاه المعاكس لانتشارها، أي من مكان منخفض إلى مكان أعلى تركيز عالي. عن طريق النقل النشط، تتم إزالة أيونات الصوديوم والهيدروجين والكلور الزائدة باستمرار من الخلية. على العكس من ذلك، فإن الفوسفات والجلوكوز والأحماض الأمينية تخترق بنشاط السيتوبلازم. يتضمن النقل النشط دائمًا استهلاك الطاقة.
قد يُفقد الغشاء جزئيًا أثناء حياة الخلية نتيجة لتكوين فجوات هضمية (فقاعات). يتم استعادة الغشاء بانتظام نتيجة لعمل عضيات خاصة. توليف فجوات الغشاء. تلتصق هذه الفجوات في أي مكان على غشاء الخلية، وتستعيد حجمها وخصائصها السابقة.
ج) الدهون الغشائية.
تكوين الدهون الأغشية البيولوجيةمتعدد جدا. ممثلين مميزينالدهون الموجودة في أغشية الخلايا هي الدهون الفوسفاتية والسفينغوميلين والكوليسترول (الدهون الستيرويدية).
ميزة مميزةالدهون الغشائية هي تقسيم جزيئها إلى جزأين مختلفين وظيفيًا: ذيول غير قطبية وغير مشحونة تتكون من أحماض دهنية، ورؤوس قطبية مشحونة.
تحمل الرؤوس القطبية رسوم سلبيةأو قد تكون محايدة. يفسر وجود ذيول غير قطبية حسن ذوبان الدهون في الدهون والمذيبات العضوية. في إحدى التجارب، من خلال خلط الدهون المعزولة من الأغشية مع الماء، يمكن الحصول على طبقات أو أغشية ثنائية الجزيئية يبلغ سمكها حوالي 7.5 نانومتر، حيث تكون المناطق الطرفية للطبقة عبارة عن رؤوس قطبية محبة للماء، والمنطقة المركزية هي ذيول الدهون غير المشحونة. جزيئات. جميع أغشية الخلايا الطبيعية لها نفس البنية.
تختلف أغشية الخلايا بشكل كبير عن بعضها البعض في تكوين الدهون. على سبيل المثال، أغشية البلازماالخلايا الحيوانية غنية بالكوليسترول (تصل إلى 30٪) ومنخفضة بالليسيثين، في حين أن أغشية الميتوكوندريا غنية بالفوسفوليبيد وفقيرة بالكوليسترول.
يمكن لجزيئات الدهون أن تتحرك على طول الطبقة الدهنية، ويمكن أن تدور حول محورها، وتنتقل أيضًا من طبقة إلى أخرى. تتمتع البروتينات العائمة في البحيرة الدهنية أيضًا ببعض الحركة الجانبية. يختلف تكوين الدهون على جانبي الغشاء، مما يحدد عدم التماثل في بنية الطبقة الصفراوية.
د) البروتينات الغشائية.
جزيئات البروتين، وهي أثقل وأكبر بكثير من جزيئات الدهون، تكون مدمجة في الطبقة الصفراوية للغشاء. تكون معظم جزيئات البروتين مغمورة جزئيًا في الغشاء وترتبط بالدهون من خلال تفاعلات كارهة للماء. هذه هي ما يسمى البروتينات المتكاملة.
ترتبط البروتينات الأخرى فقط مع "النهايات" القطبية لجزيئات الدهون وتقع على سطح الطبقة الصفراوية، بينما تخترق البروتينات الأخرى الغشاء مباشرة. يتجلى عدم التماثل في بنية الأغشية بشكل واضح فيما يتعلق بالبروتينات المتكاملة. إنها موجهة بشكل صارم: نهاياتها الطرفية N تواجه تجويف الفجوة أو البيئة خارج الخلية.
تختلف الأغشية بشكل كبير في تركيب البروتينات، والتي تحدد في الواقع خصائصها الوظيفية. يمكن تقسيم جميع بروتينات الغشاء إلى ثلاث مجموعات: الإنزيمات وبروتينات المستقبلات والبروتينات الهيكلية.
تحتوي الأغشية المختلفة على مجموعة مميزة من الإنزيمات. على سبيل المثال، يتم تحديد ATPase المعتمد على K-Na، والذي يشارك في نقل الأيونات، في غشاء البلازما. ترتبط بروتينات المستقبلات على وجه التحديد بمواد معينة، كما كانت، "تتعرف عليها": وهي بروتينات مستقبلات للهرمونات، للتعرف على سطح الخلايا المجاورة، والفيروسات، وما إلى ذلك.
توفر البروتينات الهيكلية قوة الغشاء وترتبط بمجموعة متنوعة من الهياكل البروتينية في السيتوبلازم. على سبيل المثال، في الخلايا الظهارية، ترتبط بروتينات غشاء البلازما الخاصة بعناصر الهيكل الخلوي وتشارك في تكوين عدد من الوصلات بين الخلايا، مثل الديسموسومات والوصلات اللاصقة وما إلى ذلك.
ه) الكربوهيدرات الغشائية.
ترتبط بشكل رئيسي بالأغشية البروتينات السكرية - جزيئات البروتين المرتبطة تساهمياً بسلاسل الكربوهيدرات. عادة، توجد جزيئات الكربوهيدرات في الطبقات الخارجية للأغشية. غالبًا ما تكون هذه سلاسل خطية أو متفرعة قصيرة، والتي تشمل الجالاكتوز، والمانيوز، والفركتوز، والسكروز، وN-أسيتيل الجلوكوزامين. أرابينوز. الزيلوز ، إلخ.
و) تركيب أغشية الخلايا.
جميع أغشية الخلايا (باستثناء أغشية الميتوكوندريا والبلاستيدات) مبنية في الشبكة الإندوبلازمية الحبيبية. تنفصل عنه فجوات غشائية صغيرة تندمج مع أغشية جهاز جولجي. تُستخدم الحويصلات الغشائية، التي ينتجها جهاز جولجي، في بناء الغشاء البلازمي والفجوات الإفرازية وجميع مكونات الغشاء الأخرى للخلية.
5. غشاء البلازما.
غشاء بلازمي (انظر الملحق رقم 8)(البلازما، غشاء الخلية)، تحت الجليكالوكسي و جدار الخليةيوجد غشاء بلازمي يحيط ببروتوبلازم الخلايا النباتية والحيوانية. ويبلغ سمك الغشاء البلازمي حوالي 10 نانومتر، ولا يمكن دراسة بنيته ووظائفه إلا باستخدام المجهر الإلكتروني.
إنه لا يعمل كحاجز ميكانيكي فحسب، بل الأهم من ذلك أنه يحد من التدفق الحر في اتجاهين للمواد ذات الجزيئات المنخفضة والعالية داخل وخارج الخلية. علاوة على ذلك، تعمل البلازما بمثابة هيكل "يتعرف" على العديد من الأشياء المواد الكيميائيةوتنظيم النقل الانتقائي لهذه المواد إلى داخل الخلية. وهي مثل أغشية الخلايا الأخرى، تنشأ وتتجدد بسبب النشاط الاصطناعيالشبكة الإندوبلازمية ولها بنية مشابهة لها.
أ) دور حاجز النقل للبلازما.
يتم تحديد الاستقرار الميكانيكي للغشاء البلازمي ليس فقط من خلال خصائص الغشاء نفسه، ولكن أيضًا من خلال خصائص الجليكالوكسي المجاور والطبقة القشرية من السيتوبلازم.
السطح الخارجي للغشاء البلازمي (انظر الملحق رقم 13).مغطاة بطبقة ليفية فضفاضة من مادة بسمك 3-4 نانومتر - الجليكالوكسي. وهو يتألف من سلاسل عديد السكاريد المتفرعة من البروتينات الغشائية المتكاملة، والتي يمكن أن يوجد بينها الجليكوليبيدات والبروتيوغليكان التي تفرزها الخلية. توجد هنا أيضًا بعض الإنزيمات المحللة الخلوية المشاركة في تحلل المواد خارج الخلية (الهضم خارج الخلية، على سبيل المثال، في ظهارة الأمعاء).
الطبقة القشرية من السيتوبلازم، التي يبلغ سمكها 0.1-0.5 ميكرومتر، لا تحتوي على الريبوسومات والهياكل الغشائية، ولكنها غنية بالألياف الدقيقة للأكتين.
غشاء البلازما، مثل أغشية الخلايا البروتينية الدهنية الأخرى، شبه منفذ. تتمتع المياه والغازات المذابة فيها بأقصى قدرة على الاختراق. يمكن أن يحدث النقل الأيوني بشكل سلبي، دون استهلاك الطاقة. في هذه الحالة، تشكل بعض بروتينات النقل الغشائي مجمعات جزيئية، وهي قنوات تمر من خلالها الأيونات عبر الغشاء عن طريق الانتشار البسيط.
وفي حالات أخرى، ترتبط بروتينات نقل الغشاء الخاصة بشكل انتقائي بأيون واحد أو آخر وتنقله عبر الغشاء. يسمى هذا النوع من النقل بالنقل النشط ويتم باستخدام مضخات أيون البروتين. على سبيل المثال، عند إنفاق جزيء واحد من ATP، يقوم نظام ضخ K-Na بضخ 3 أيونات Na من الخلية في دورة واحدة ويضخ 2 أيونات K مقابل تدرج التركيز.
بالاشتراك مع النقل الأيوني النشط، تخترق السكريات المختلفة والنيوكليوتيدات والأحماض الأمينية البلازما. الجزيئات الكبيرة، مثل البروتينات، لا تمر عبر الغشاء. يتم نقلهم، وكذلك جزيئات المادة الأكبر حجما، إلى الخلية من خلال الالتقام . أثناء الالتقام الخلوي، تلتقط منطقة معينة من البلازما المواد الموجودة خارج الخلية وتغلفها وتحيط بها في فجوة غشائية. هذه الفجوة هي إندوسوم - يندمج في السيتوبلازم مع الليزوزوم الأولي ويحدث هضم المادة الملتقطة.
ينقسم الالتقام الخلوي رسميًا إلى البلعمة (امتصاص الجزيئات الكبيرة بواسطة الخلية) واحتساء الخلايا (امتصاص المحاليل).
ويشارك الغشاء البلازمي أيضًا في إزالة المواد من الخلية بمساعدة طرد خلوي - عملية عكسية للالتقام الخلوي.
ب) الدور المستقبلي للبلازما.
البروتينات الناقلة في الغشاء الخارجي للخلية هي أيضًا مستقبلات تتعرف على أيونات معينة وتتفاعل معها. يمكن أن تعمل البروتينات الغشائية أو عناصر الجليكالوكسي كمستقبلات على سطح الخلية. وتنتشر هذه المناطق الحساسة للمواد الفردية على سطح الخلية أو تتجمع في مناطق صغيرة.
إن دور العديد من المستقبلات الخلوية ليس فقط ربط مواد معينة، ولكن أيضًا نقل الإشارات من السطح إلى الخلية. على سبيل المثال، عندما يعمل الهرمون على الخلية، تتكشف سلسلة الأحداث على النحو التالي: يتفاعل جزيء الهرمون على وجه التحديد مع بروتين مستقبل غشاء البلازما، ودون اختراق الخلية، ينشط الإنزيم الذي يصنع AMP الدوري. هذا الأخير ينشط أو يثبط إنزيمًا أو مجموعة من الإنزيمات داخل الخلايا.
إن تنوع وخصوصية مجموعات المستقبلات على سطح الخلية يؤدي إلى تكوين خلايا عصبية شديدة نظام معقدعلامات تسمح للخلايا بالتمييز بين "الذات" (لنفس الفرد أو نفس النوع) و"غير الذاتية".
ج) الاتصالات بين الخلايا.
في الكائنات متعددة الخلايا، تتشكل التجمعات الخلوية المعقدة بسبب التفاعلات بين الخلايا. عندما تكون الخلايا على مقربة من بعضها البعض، يضمن الكأس السكري التصاق الخلايا بسبب وجود بروتينات سكرية عبر الغشاء (كادهيرين). هذا هو اتصال بسيط من خلية إلى أخرى، حيث تكون الفجوة بين الخلايا 10-20 نانومتر.
في الظهارة، غالبًا ما يتم العثور على تقاطع محكم أو مقفل، حيث تكون الطبقات الخارجية من غشائي البلازما قريبة قدر الإمكان وتقع كريات بروتينات الغشاء المتكامل في نقاط اتصالها. هذا الاتصال لا يمكن اختراقه للجزيئات والأيونات، فهو يغلق التجاويف بين الخلايا.
إرساء الاتصالات، أو جهات الاتصال، لا تقوم فقط بتوصيل أغشية البلازما للخلايا المجاورة، ولكنها تتواصل أيضًا مع العناصر الليفية في الهيكل الخلوي. على سبيل المثال، تتميز الديسموسومات، التي تشبه اللويحات أو الأزرار، بوجود طبقة كثيفة من البروتينات السكرية ديسموجلين في الفضاء بين الخلايا.
على الجانب السيتوبلازمي، بجوار البلازماليما توجد طبقة من بروتين ديزموبلاكين، المرتبط بالخيوط الوسيطة للهيكل الخلوي.
تعتبر تقاطعات الفجوة تقاطعات اتصال الخلايا. في منطقة الاتصال الفجوة يمكن أن يكون هناك من 20-30 إلى عدة آلاف connexons - الهياكل البروتينية الأسطوانية ذات قناة داخليةبقطر 2 نانومتر. يتكون كل اتصال من 6 وحدات فرعية من بروتين Connectin. تلعب Connexons دور القنوات المباشرة بين الخلايا التي من خلالها يمكن للأيونات والمواد ذات الوزن الجزيئي المنخفض أن تنتشر من خلية إلى أخرى.
6. الليزوزومات.
الليزوزومات (من lyses واليونانية سوما - الجسم). وهي أجسام مستديرة صغيرة. يتم فصل كل ليسوسوم عن السيتوبلازم بواسطة غشاء. يوجد داخل الليزوزوم إنزيمات يمكنها تحطيم (أي الليزوزوم - ومن هنا الاسم) البروتينات والسكريات والببتيدات والأحماض النووية (انظر الملحق رقم 8).
هذه فئة متنوعة جدًا من الحويصلات بحجم 0.1-0.4 ميكرومتر، ويحدها غشاء واحد (سمكه حوالي 7 نانومتر)، مع محتويات غير متجانسة بالداخل. تتشكل بسبب نشاط الشبكة الإندوبلازمية وجهاز جولجي وفي هذا الصدد تشبه الفجوات الإفرازية.
دورهم الرئيسي هو المشاركة في عمليات الانهيار داخل الخلايا لكل من الجزيئات البيولوجية الخارجية والداخلية. ميزة مميزةالجسيمات الحالة هي أنها تحتوي على حوالي 40 إنزيمًا مائيًا: البروتينات، النيوكلياز، الفوسفاتيز، الجليكوسيداز، وما إلى ذلك، والتي يحدث العمل الأمثل لها عند الرقم الهيدروجيني 5.
في الليزوزومات، يتم إنشاء البيئة الحمضية بسبب وجود "مضخة" البروتون في أغشيتها، والتي تستهلك طاقة ATP.
بالإضافة إلى ذلك، يتم بناء البروتينات الحاملة في أغشية الليزوزوم لنقل منتجات التحلل المائي من الليزوزوم إلى السيتوبلازم: مونومرات الجزيئات المنقسمة - الأحماض الأمينية والسكريات والنيوكليوتيدات والدهون. من أجل عدم هضم نفسها، تتم حماية العناصر الغشائية للليزوزومات بواسطة السكريات قليلة التعدد، والتي تمنع الهيدروليزات من التفاعل معها.
من بين الجسيمات الليزوزومية ذات الأشكال المختلفة، يتم تمييز أربعة أنواع: الليزوزومات الأولية والثانوية، والجسيمات البلعمية الذاتية، والأجسام المتبقية.
الليزوزومات الأولية.
عادة ما تكون هذه حويصلات غشائية صغيرة يبلغ قطرها حوالي 100 نانومتر مع محتويات غير هيكلية تحتوي على فوسفاتيز الحمض النشط، وهو إنزيم محدد للجسيمات الحالة. لقد ثبت أن الإنزيمات الليزوزومية يتم تصنيعها، كالعادة، في الشبكة الحبيبية، ويتم تعبئتها في حويصلات غشائية في جهاز جولجي. يشبه المسار الكامل لتشكيل الليزوزومات الأولية تكوين الحبيبات الإفرازية، على سبيل المثال، في خلايا البنكرياس.
الليزوزومات الثانوية.
تندمج الفجوات البلعمية أو الفجوات الخلوية مع الليزوزومات الأولية لتكوين الليزوزومات الثانوية. تبدأ عملية تحلل الركائز التي تمتصها الخلية تحت تأثير الهيدروليزات الموجودة في الليزوزوم الأولي. يتم تقسيم المواد الحيوية إلى مونومرات، والتي يتم نقلها عبر الغشاء الليزوزومي إلى السيتوبلازم، حيث يتم إعادة استخدامها وإدراجها في مختلف التركيبات الاصطناعية والكيميائية. العمليات الأيضية. يرتبط تنوع حجم وبنية الليزوزومات الخلوية في المقام الأول بتنوع الليزوزومات الثانوية - وهي منتجات اندماج الفجوات الداخلية مع الليزوزومات الأولية.
الهيئات المتبقية.
قد لا يكتمل تحلل الجزيئات الحيوية المنشأ داخل الليزوزومات. في هذه الحالة، تتراكم المنتجات غير المهضومة في تجويف الليزوزوم، ويصبح الليزوزوم الثانوي جسمًا متبقيًا (تيلوليسوسوم). يتم ضغط محتوياتها وإعادة ترتيبها. في كثير من الأحيان في الهيئات المتبقية، لوحظ الهيكلة الثانوية للدهون غير المهضومة، والتي تشكل هياكل الطبقات المعقدة. تترسب المواد الصباغية هناك.
الجسيمات الذاتية.
وفقًا للتشكل، يتم تصنيف الجسيمات الحالة الذاتية (البلعمة الذاتية) على أنها ليسوسومات ثانوية، ولكن داخل هذه الجزيئات توجد شظايا أو حتى هياكل سيتوبلازمية كاملة، مثل الميتوكوندريا، والبلاستيدات، والريبوسومات، وعناصر الشبكة، وما إلى ذلك. ومن المفترض أن عملية البلعمة الذاتية هي المرتبطة باختيار وتدمير المكونات الخلوية المتغيرة "المكسورة".
7. جهاز جولجي.
جهاز جولجي (مجمع جولجي) اسمه C. جولجي، عضية الخلية.
وفي العديد من الخلايا الحيوانية، مثل الخلايا العصبية، تأخذ شكل شبكة معقدة تقع حول النواة. في خلايا النباتات والأوالي، يتم تمثيل جهاز جولجي بواسطة أجسام فردية على شكل منجل أو قضيب. وتتشابه بنية هذه العضية في خلايا الكائنات الحية النباتية والحيوانية، على الرغم من تنوع شكلها (انظر الملحق رقم 8).
يشتمل جهاز جولجي على: تجاويف تحدها الأغشية وتقع في مجموعات (5-10)؛ فقاعات كبيرة وصغيرة تقع في نهايات التجاويف. كل هذه العناصر تشكل مجمعا واحدا.
يقوم جهاز جولجي بالعديد من الوظائف المهمة.
يجلب البروتينات إلى شكل عملها النهائي، على سبيل المثال، "يربط" بعض البروتينات ببروتينات كبيرة مجمعات البروتين- يربط أيونات المعادن الأساسية ببعض البروتينات.
تشكل حويصلات غشائية، والتي، بعد خروجها من مجمع جولجي، إما تستعيد غشاء الخلية أو تتحول إلى ليسوسومات. الليزوزومات هي عضيات غشائية للخلية، تمثل حويصلات مجهرية مملوءة بالإنزيمات الهاضمة.
أداء وظائف الجهاز الهضمي والوقائي. على سبيل المثال، يتم تصنيع خلايا البنكرياس في الثدييات الانزيمات الهاضمة، والتي تتراكم في تجاويف العضو العضوي.
يمكنهم الالتصاق بالفجوة الهضمية عن طريق سكب الإنزيمات الهضمية فيها. عندما تتلامس خلية مع مادة غريبة أو مع خلية غريبة، تلتصق الليزوزومات غشاء الخلية، وإطلاق إنزيماتها خارج الخلية. يمكن أن تشارك إنزيمات الليزوزوم أيضًا في "الموت المبرمج" لخلية الفرد.
بفضل نشاط جهاز جولجي، يحدث تجديد ونمو غشاء البلازما.
8. الريبوسومات.
تم العثور على الريبوسومات في خلايا جميع الكائنات الحية. هذه أجسام مستديرة مجهرية يبلغ قطرها 15-20 نانومتر. يتكون كل ريبوسوم من جزيئين غير متساويين في الحجم، صغير وكبير. (انظر الملحق رقم 14).
تحتوي خلية واحدة على عدة آلاف من الريبوسومات، وهي موجودة إما على أغشية الشبكة الإندوبلازمية الحبيبية أو تقع بحرية في السيتوبلازم.
تحتوي الريبوسومات على البروتينات والحمض النووي الريبي (RNA). وظيفة الريبوسومات هي تخليق البروتين. تخليق البروتين – عملية صعبة، والتي لا يتم تنفيذها بواسطة ريبوسوم واحد، ولكن بواسطة مجموعة كاملة، بما في ذلك ما يصل إلى عدة عشرات من الريبوسومات المتحدة. تسمى هذه المجموعة من الريبوسومات متعدد الجسيمات .
تتراكم البروتينات المركبة أولاً في قنوات وتجويف الشبكة الإندوبلازمية، ثم يتم نقلها إلى العضيات ومواقع الخلايا حيث يتم استهلاكها.
الشبكة الإندوبلازمية والريبوسومات (انظر الملحق رقم 8)، الموجودة على أغشيتها، تمثل جهازًا واحدًا للتخليق الحيوي ونقل البروتينات.
9. الميتوكوندريا.
غشاء العضيات ذاتية الانقسام. يحتوي السيتوبلازم في معظم الخلايا الحيوانية والنباتية على أجسام صغيرة (0.2-7 ميكرون) - الميتوكوندريا (اليونانية "mitos" - الخيط، "chondrion" - الحبوب والحبيبات).
تظهر الميتوكوندريا بوضوح بالمجهر الضوئي، ويمكنك من خلاله فحص شكلها وموقعها وحساب عددها (انظر الملحق رقم 8).تمت دراسة البنية الداخلية للميتوكوندريا باستخدام المجهر الإلكتروني.
تتكون قشرة الميتوكوندريا من غشائين - خارجي وداخلي. الغشاء الخارجي أملس ولا يشكل أي طيات أو نتوءات. على العكس من ذلك، يشكل الغشاء الداخلي طيات عديدة يتم توجيهها إلى تجويف الميتوكوندريا. تسمى طيات الغشاء الداخلي كريستامي (لاتينية "كريستا" - ريدج، ثمرة) تحدث فيها عملية أكسدة حمض اللاكتيك، ونتيجة لذلك يتم إطلاق الطاقة وتخزينها على شكل ATP.
يختلف عدد الأعراف في الميتوكوندريا في الخلايا المختلفة. يمكن أن يكون هناك من عدة عشرات إلى عدة مئات منها، مع وجود العديد من الأعراف بشكل خاص في الميتوكوندريا في الخلايا النشطة، مثل الخلايا العضلية.
تسمى الميتوكوندريا "محطات الطاقة" للخلايا، لأن وظيفتها الرئيسية هي تخليق حمض الأدينوزين ثلاثي الفوسفوريك (ATP). يتم تصنيع هذا الحمض في الميتوكوندريا في خلايا جميع الكائنات الحية وهو مصدر عالمي للطاقة اللازمة للعمليات الحيوية للخلية والكائن الحي بأكمله.
الميتوكوندريا، مثل البكتيريا، تتكاثر عن طريق الانشطار المباشر.
10. مركز الخلوي.
يوجد في الخلايا القريبة من النواة عضية تسمى مركز الخلية. يتكون الجزء الرئيسي من مركز الخلية من جسمين صغيرين (أحياناً أكثر) - مركزيات تقع في منطقة صغيرة من السيتوبلازم الكثيف. كل مركز مركزي له شكل أسطوانة يصل طولها إلى 1 ميكرومتر، وتتكون جدرانها من أنابيب دقيقة تقع على طول محور الأسطوانة. تحتوي المريكزات على بروتينات وليست كذلك عدد كبير منالحمض النووي الريبي. هناك زوجان من المريكزات في الخلية. في كل زوج، تقع المريكزات بشكل متعامد مع بعضها البعض.
تلعب المريكزات دورًا مهمًا في انقسام الخلايا. أنها تشكل خيوط بروتينية طويلة، ما يسمى المغزل. عندما تنقسم الخلية، تتباعد المريكزات نحو قطبيها، مما يحدد اتجاه مغزل الانقسام.
11. البلاستيدات.
البلاستيدات هي عضيات خلوية مرتبطة بغشاء وتنقسم ذاتيًا. على عكس جميع العضيات التي تمت مناقشتها أعلاه، توجد البلاستيدات في الخلايا النباتية فقط.
يشبه الهيكل الميتوكوندريا: يتكون من غشائين، الخارجي الأملس والداخلي، ويشكلان العديد من النتوءات المسطحة - ثايلاكويدات . يتم ترتيب جميع الثايلاكويدات في أكوام مثل أكوام العملات المعدنية. يتم استدعاء كل مكدس وجه . يوجد بين الجرانا سائل داخلي من البلاستيد يسمى سدى . يحتوي على الحمض النووي الخاص به، والذي يشبه في تركيبه الحمض النووي البكتيري.
تتكاثر البلاستيدات مثل البكتيريا عن طريق الانقسام المباشر.
بناءً على خصائصها الهيكلية، هناك ثلاثة أنواع رئيسية من البلاستيدات (انظر الملحق رقم 15): أخضر - البلاستيدات الخضراء. الأحمر والبرتقالي والأصفر - البلاستيدات الملونة. عديم اللون - الكريات البيضاء.
أ) البلاستيدات الخضراء.
توجد هذه العضيات في خلايا الأوراق وأعضاء النباتات الخضراء الأخرى (البراعم والفواكه غير الناضجة)، وكذلك في مجموعة متنوعة من الطحالب.
أحجام البلاستيدات الخضراء هي 4-6 ميكرون، في معظم الأحيان لديهم شكل بيضاوي. في النباتات العليا، تحتوي الخلية الواحدة عادةً على عشرات من البلاستيدات الخضراء.
اللون الاخضرتعتمد البلاستيدات الخضراء على محتوى صبغة الكلوروفيل فيها.
البلاستيدات الخضراء هي العضية الرئيسية للخلايا النباتية التي يحدث فيها التمثيل الضوئي، أي تكوين المواد العضوية (الكربوهيدرات) من المواد غير العضوية (ثاني أكسيد الكربون وH20) باستخدام طاقة ضوء الشمس.
البلاستيدات الخضراء مماثلة في هيكل الميتوكوندريا (انظر الملحق رقم 16).يتم فصل البلاستيدات الخضراء عن السيتوبلازم بغشاءين - خارجي وداخلي. يكون الغشاء الخارجي أملس، بدون طيات أو نتوءات، بينما يشكل الغشاء الداخلي العديد من النتوءات المطوية الموجهة إلى البلاستيدات الخضراء. لذلك، يتركز عدد كبير من الأغشية داخل البلاستيدات الخضراء، وتشكل هياكل خاصة - جرانا. توجد جزيئات الكلوروفيل في الأغشية الحبيبية، ولهذا السبب تحدث عملية التمثيل الضوئي هنا.
يتم تصنيع ATP أيضًا في البلاستيدات الخضراء. بين الأغشية الداخلية للبلاستيدات الخضراء يوجد DNA و RNA والريبوسومات. وبالتالي، في البلاستيدات الخضراء، وكذلك في الميتوكوندريا، يحدث تخليق البروتين اللازم لنشاط هذه العضيات. تتكاثر البلاستيدات الخضراء بالتقسيم (انظر الملحق رقم 17).
ب) البلاستيدات الملونة.
البلاستيدات الملونة هي بلاستيدات تحتوي على أصباغ شبيهة بالدهون تلون الخلية باللون الأصفر والبرتقالي والأحمر. وتوجد في ثمار النباتات الناضجة مما يمنحها اللون المناسب، وكذلك في أوراق الأشجار متساقطة الأوراق في فصل الخريف.
ج) الليوكوبلاست.
الليوكوبلاست هي بلاستيدات عديمة اللون. أنها لا تحتوي على أي أصباغ على الغشاء الداخلي. في الخلية هم مسؤولون عن تخليق وتراكم السكريات (النشا). توجد بكميات كبيرة في خلايا البراعم الموجودة تحت الأرض (درنات البطاطس وخرشوف القدس) وكذلك في الفواكه والبذور.
د) تحولات البلاستيدات.
يمكن أن تتحول البلاستيدات الخضراء بسهولة إلى أنواع أخرى من البلاستيدات. نلاحظ ذلك مع اصفرار واحمرار الثمار أو الأوراق الناضجة في الخريف. في الظلام، يمكن أن يتغير لون البلاستيدات الخضراء، وتتحول إلى بلاستيدات بيضاء. ومع ذلك، فإن هذه العمليات لا رجعة فيها: لا تتحول البلاستيدات البيضاء والبلاستيدات الخضراء أبدًا إلى البلاستيدات الخضراء.
النواة (نواة الخلية) الجزء الإلزاميالخلايا في العديد من الكائنات وحيدة الخلية وجميع الكائنات متعددة الخلايا. الأحجام من 1 ميكرون (في بعض الأوليات) إلى 1 ملم (في بيض بعض الأسماك والبرمائيات). تم استخدام مصطلح "النواة" (النواة اللاتينية) لأول مرة من قبل ر. براون في عام 1833، عندما وصف الهياكل الكروية التي لاحظها في الخلايا النباتية (انظر الملحق رقم 8).
أ) الغلاف النووي.
يتم فصل الفضاء الداخلي لنواة الخلية عن السيتوبلازم بواسطة غلاف نووي يتكون من غشائين. تتشابه أغشية الغشاء النووي في تركيبها مع مكونات الغشاء الأخرى للخلية وهي مبنية على نفس المبدأ: فهي عبارة عن أغشية رقيقة من البروتين الدهني تتكون من طبقة مزدوجة من جزيئات الدهون التي تندمج فيها جزيئات البروتين.
يسمى الفراغ الموجود بين الأغشية النووية الداخلية والخارجية شبه نووي.
يوجد عادة عدد كبير من الريبوسومات على سطح الغشاء النووي الخارجي، وفي بعض الأحيان يكون من الممكن ملاحظة الانتقال المباشر لهذا الغشاء إلى نظام قناة الشبكة الإندوبلازمية الحبيبية للخلية.
ويرتبط الغشاء النووي الداخلي بطبقة بروتينية ليفية رقيقة - الصفيحة النووية ، تتكون من بروتينات اللامين. إن الشبكة الكثيفة من ألياف الصفيحة النووية قادرة على ضمان سلامة النواة، حتى بعد تفكك الأغشية الدهنية للغلاف النووي في التجربة. مع داخلوترتبط حلقات الكروماتين التي تملأ النواة بالصفيحة.
يحتوي الغلاف النووي على ثقوب يبلغ قطرها حوالي 90 نانومتر، تتشكل نتيجة اندماج الأغشية النووية الخارجية والداخلية. مثل هذه الثقوب في الغلاف النووي محاطة بهياكل بروتينية معقدة تسمى مجمع المسام النووي.
تتوضع الوحدات البروتينية الثمانية التي يتكون منها المسام النووي حول ثقب الغلاف النووي على شكل حلقات يبلغ قطرها حوالي 120 نانومتر، ويتم ملاحظتها بالمجهر الإلكتروني على جانبي الغلاف النووي. تحتوي الوحدات الفرعية البروتينية في مجمع المسام على نتوءات موجهة نحو مركز المسام، حيث تظهر أحيانًا "الحبيبة المركزية" التي يبلغ قطرها 10-40 نانومتر.
يعد حجم المسام النووية وبنيتها أمرًا قياسيًا لجميع الخلايا حقيقية النواة. يعتمد عدد المسام النووية على النشاط الأيضي للخلايا: كلما ارتفع مستوى العمليات التخليقية في الخلية، زاد عدد المسام لكل وحدة مساحة سطح نواة الخلية.
في عملية النقل السيتوبلازمي النووي، تعمل المسام النووية كنوع من المنخل الجزيئي، حيث تقوم بتمرير الأيونات والجزيئات الصغيرة (السكريات، النيوكليوتيدات، ATP، إلخ) بشكل سلبي، على طول تدرج التركيز، وتنفيذ النقل الانتقائي النشط للجزيئات الكبيرة. من البروتينات والبروتينات النووية الريبية، أي أحماض مجمعات الحمض النووي الريبي (RNA) مع البروتينات. على سبيل المثال، البروتينات المنقولة إلى النواة من السيتوبلازم، حيث يتم تصنيعها، يجب أن تحتوي على تسلسلات محددة من حوالي 50 حمضًا أمينيًا (ما يسمى بتسلسلات NLS)، "يتم التعرف عليها" بواسطة مجمع المسام النووي. في هذه الحالة، يقوم مجمع المسام النووي، الذي ينفق الطاقة على شكل ATP، بنقل البروتين من السيتوبلازم إلى النواة بشكل فعال.
ب) الكروماتين.
نواة الخلية هي مستودع لجميع المعلومات الوراثية للخلية تقريبًا، وبالتالي فإن المحتوى الرئيسي لنواة الخلية هو الكروماتين: مركب من الحمض النووي الريبي منقوص الأكسجين (DNA) وبروتينات مختلفة.
في النواة وخاصة في الكروموسومات الانقسامية، يتم طي الحمض النووي للكروماتين عدة مرات، وتعبئته بطريقة خاصة لتحقيق درجة عاليةالضغط. بعد كل شيء، يجب وضع جميع خيوط الحمض النووي الطويلة، التي يبلغ طولها الإجمالي، على سبيل المثال، في البشر حوالي 164 سم، في نواة الخلية، التي يبلغ قطرها بضعة ميكرومترات فقط. يتم حل هذه المشكلة عن طريق التعبئة التسلسلية للحمض النووي في الكروماتين باستخدام بروتينات خاصة.
الجزء الأكبر من بروتينات الكروماتين عبارة عن بروتينات هيستون، والتي تعد جزءًا من وحدات فرعية للكروماتين الكروي تسمى النيوكليوزومات . هناك 5 أنواع من بروتينات الهيستون.
النيوكليوزوم هو جسيم أسطواني يتكون من 8 جزيئات هيستون، يبلغ قطرها حوالي 10 نانومتر، حيث يتم "جرح" أقل بقليل من لفتين من خيط جزيء الحمض النووي. في ميكروسكوب الكترونييبدو مثل هذا الكروماتين المتحلل بشكل مصطنع وكأنه "خرز على خيط".
في نواة الخلية الحية، ترتبط النيوكليوزومات بإحكام بمساعدة بروتين رابط هيستون آخر، لتشكل ما يسمى بليف الكروماتين الأولي الذي يبلغ قطره 30 نانومتر. توفر البروتينات الأخرى ذات الطبيعة غير الهيستونية والتي تشكل جزءًا من الكروماتين مزيدًا من الضغط. أي طي ألياف الكروماتين التي تصل إلى مكانها القيم القصوىأثناء انقسام الخلايا في الكروموسومات الانقسامية أو الانقسامية.
يوجد الكروماتين في نواة الخلية على شكل كروماتين كثيف ومكثف، حيث تكون الألياف الأولية بطول 30 نانومتر معبأة بإحكام، وعلى شكل كروماتين منتشر متجانس. تعتمد النسبة الكمية لهذين النوعين من الكروماتين على طبيعة النشاط الأيضي للخلية ودرجة تمايزها. على سبيل المثال، تحتوي نوى كريات الدم الحمراء في الطيور، التي لا تحدث فيها عمليات النسخ والنسخ النشطة، على كروماتين مكثف كثيف فقط.
يحتفظ بعض الكروماتين بحالته المضغوطة والمكثفة طوال دورة الخلية - ويسمى هذا الكروماتين الكروماتين المغاير ويختلف عن الكروماتين الحقيقي في عدد من الخصائص.
ج) النسخ والنسخ.
تحتوي الخلايا حقيقية النواة عادة على عدة كروموسومات (من اثنين إلى عدة مئات)، والتي تفقد شكلها المضغوط في النواة (في الطور البيني، أي بين الانقسامات الانقسامية)، وتخفف وتملأ حجم النواة على شكل كروماتين. على الرغم من حالة التكثيف، فإن كل كروموسوم يحتل موقعًا محددًا بدقة في النواة ويتصل بالغلاف النووي من خلال الصفيحة. ثابت بدقة على السطح الداخليالغلاف النووي مثل هياكل الكروموسوم مثل السنترومير والتيلوميرات.
في مرحلة معينة من دورة حياة الخلية، في الفترة الاصطناعية، يحدث التضاعف، أي يتضاعف الحمض النووي للنواة بأكمله، ويصبح الكروماتين أكبر بمرتين. وتأتي البروتينات اللازمة لهذه العملية، بالطبع، من السيتوبلازم عبر المسام النووية. وهكذا، تستعد الخلية للانقسام الخلوي القادم - الانقسام، ومتى المجموعسيعود الحمض النووي الموجود في النواة إلى مستواه الأصلي.
يبدأ تنفيذ المعلومات الوراثية الموجودة في الحمض النووي في شكل جينات بالنسخ، أي بتركيب الحمض النووي الريبي المرسال (i-RNA) - نسخ دقيقة من الجينات، والتي سيتم استخدامها بعد ذلك لبناء البروتينات في السيتوبلازم على الريبوسومات.
تحدث هذه العملية في نقاط مختلفة في حجم النواة، ولا تختلف شكليا عن الكروماتين المحيط بها. في أغلب الأحيان يكون من الممكن ملاحظة النسخ المنتشر، أي. كروماتين منزوع التكثيف.
بالإضافة إلى الكروماتين الذي يشكل الكروموسومات، تحتوي نوى حقيقيات النوى عادة على نواة واحدة أو أكثر.
د) مصفوفة البروتين النووي.
ولتنفيذ عمليات النسخ والنسخ وأيضًا للحفاظ على موضع معين للكروموسومات في حجم النواة، توجد هياكل بروتينية إطارية تسمى مصفوفة البروتين النووي. تتكون هذه المصفوفة من ثلاثة مكونات مورفولوجية على الأقل:
· الطبقة الليفية المحيطية - الصفيحة.
· المصفوفة الداخلية أو المتداخلة للنواة
· ومصفوفة النواة.
تظهر الملاحظات أن مكونات المصفوفة النووية ليست هياكل جامدة ومتجمدة؛ فهي ديناميكية ويمكن أن تختلف بشكل كبير تبعًا لذلك الميزات الوظيفيةالنوى. لقد ثبت أن مصفوفة البروتين تحتوي على العديد من نقاط الارتباط القوية بالحمض النووي النووي، والذي بدوره لديه تسلسلات نيوكليوتيدات خاصة ضرورية لهذا الغرض.
13. النواة.
أ) عدد النواة في الخلية.
بدءاً من الطحالب الخضراء والفطريات والأوليات السفلى وانتهاءً بـ كائنات أعلىجميع الخلايا لها هياكل نووية إلزامية - النواة (انظر الملحق رقم 18).تحتوي هذه القاعدة على عدد كبير من الاستثناءات التي تؤكد فقط على أهمية وضرورة وجود النواة في الجسم دورة الحياةالخلايا. تشمل هذه الاستثناءات خلايا البويضات المنقسمة، حيث تكون النواة غائبة في المراحل المبكرة من التطور الجنيني، أو الخلايا التي اكتملت التطور وهي متخصصة بشكل لا رجعة فيه، على سبيل المثال، بعض خلايا الدم.
يمكن أن يختلف عدد النوى في الخلية، لكن عددها في كل نواة يعتمد على التوازن الجيني للخلية. وقد وجد أن تكوين النواة ينطوي على ذلك أماكن معينةبعض الكروموسومات، والتي يمكن تتبع ارتباطها بالنواة بوضوح في الطور النهائي والطور التمهيدي. عادة ما يكون لهذه الكروموسومات انقباضات ثانوية، ومناطقها هي الأماكن التي تتطور فيها النواة. وقد أطلق مكلينتوك (1934) على هذه المناطق الكروموسومية اسم "المنظمون النوويون".
تعتبر أماكن الانقباضات الثانوية مميزة بشكل خاص لموقع المنظمين النوويين، ولكن يمكن أن يوجد الأخير في بعض الأحيان في نهايات الكروموسومات أو في عدة أماكن على طول الكروموسوم.
الرقم الإجمالييتم تحديد النواة لكل نواة من خلال عدد المنظمات النووية وتزداد وفقًا لصيغة النواة. ومع ذلك، في كثير من الأحيان يكون عدد النوى لكل نواة عدد أقلالمنظمين النوويين. لقد ثبت أن النوى يمكن أن تندمج؛ بالإضافة إلى ذلك، يشارك في بعض الأحيان العديد من المنظمين في تكوين نواة واحدة.
حتى في أعمال M. S. Navashin (1934) فقد تبين أن موضع الكروموسومات، والذي في الظروف العاديةتشكل نواة كبيرة وتصبح غير نشطة عندما يظهر موضع "أقوى" في النواة على كروموسوم آخر بعد التهجين. تشير حقيقة أنه في ظل ظروف معينة يمكن قمع نشاط بعض المنظمات النووية أو زيادة نشاط المنظمات الأخرى التي كانت في السابق في حالة كامنة مخفية، إلى أنه يتم الحفاظ على توازن معين في كمية المادة النووية في الخلايا أو وبعبارة أخرى، يتم تنظيم المنتجات "الإجمالية" الصادرة عن النواة.
بناءً على الحقائق المذكورة أعلاه، يمكن استخلاص الاستنتاجات التالية:
Þ يرتبط تكوين النوى وعددها بنشاط مناطق كروموسوم معينة - المنظمات النووية الموجودة بالنسبة للجزء الاكبرفي مناطق الانقباضات الثانوية.
Þ تغيرات في عدد النواة في الخلايا من هذا النوعيمكن أن يحدث بسبب اندماج النوى أو بسبب التغيرات في توازن الكروموسومات في الخلية.
ب) فسيولوجيا وكيمياء النواة.
تتميز النواة، مقارنة بمكونات الخلية الأخرى، بأنها البنية الأكثر كثافة مع أعلى تركيز من الحمض النووي الريبوزي (RNA)، مع كثافة عالية للغاية. نشاط عاليفيما يتعلق بتوليف الحمض النووي الريبي.
يكون تركيز الحمض النووي الريبي (RNA) في النواة دائمًا أعلى من تركيز الحمض النووي الريبي (RNA) في المكونات الأخرى للخلية، وبالتالي فإن تركيز الحمض النووي الريبي (RNA) في النواة يمكن أن يكون أعلى بمقدار 2-8 مرات منه في النواة، وأعلى بمقدار 1-3 مرات منه في النواة. السيتوبلازم. نسبة تركيز الحمض النووي الريبي (RNA) في النواة والنواة والسيتوبلازم لخلايا كبد الفأر هي 1:7، 3:4، 1، وفي خلايا البنكرياس - 1:9، 6:6، 6.
لم يتم اكتشاف أي حمض نووي في النواة، ولكن عند فحص الخلايا الثابتة، يتم دائمًا تحديد منطقة الكروماتين حول النواة. يبدو أن هذا الكروماتين حول النواة، وفقًا للمجهر الإلكتروني، جزء لا يتجزأ بنية معقدةنوية.
تعد النواة واحدة من أكثر الأماكن نشاطًا في الخلية لدمج السلائف في الحمض النووي الريبي (RNA). الحمض النووي الريبي النووي هو مقدمة الحمض النووي الريبي السيتوبلازمي.
يتم تصنيع الحمض النووي الريبي السيتوبلازمي في النواة.
ج) الحمض النووي الريبي للنواة.
تقييم المحتوى العامفي الأجزاء النووية من البروتينات، RNA وDNA، يمكن للمرء أن يرى أن RNA يمثل حوالي 10% من الكتلة الإجمالية للنواة.
وبما أن الجزء الأكبر من الحمض النووي الريبي السيتوبلازمي هو الحمض النووي الريبي الريباسي، فيمكننا القول أن الحمض النووي الريبي النووي ينتمي إلى هذه الفئة.
تم تأكيد فكرة أن النواة هي موقع تخليق الرنا الريباسي (rRNA) وتكوين الريبوسوم من خلال حقيقة أن جزيئات RNP تم عزلها من المستحضرات النووية، والتي يمكن وصفها، سواء في تكوين الحمض النووي الريبي (RNA) (خصائص الترسيب) أو في الحجم، بأنها ريبوسومات أو سلائفها. مع معاملات الترسيب المختلفة.
د) الحمض النووي للنوى.
كشفت الدراسات البيوكيميائية عن وجود كمية معينة من الحمض النووي في النوى المعزولة، والتي يمكن التعرف عليها باستخدام الكروماتين حول النواة أو مع منظمات الكروموسوم النووي. محتوى الحمض النووي في النواة المعزولة هو 5-12% من الوزن الجاف و6-17% من إجمالي الحمض النووي للنواة.
الحمض النووي للمنظم النووي هو نفس الحمض النووي الذي يحدث عليه التخليق النووي، أي. الريبوسوم، RNA.
وهكذا، من خلال الأعمال البيوكيميائية، ظهرت فكرة أن العديد من الجينات المتطابقة لتخليق الرنا الريباسي (rRNA) متمركزة في نواة الحمض النووي. يتم تصنيع الرنا الريباسي (rRNA) من خلال تكوين سلائف ضخمة وتحولها الإضافي (النضج) إلى جزيئات RNA أقصر للوحدات الفرعية الكبيرة والصغيرة من الريبوسومات.
أثناء دراسة نواة بويضات السمندل، واجه الباحثون ظاهرة مثيرة للاهتمام - العدد الزائد من النوى. أثناء نمو البويضات، يظهر ما يصل إلى 1000 نواة صغيرة غير مرتبطة بالكروموسومات. هذه النوى هي التي عزلها O. Miller وفي نفس الوقت تزداد كمية الحمض النووي الريبوزي (rDNA) لكل نواة البويضة. وتسمى هذه الظاهرة التضخيم . إنه يكمن في حقيقة أن التكاثر الفائق لمنطقة المنظم النووي يحدث، حيث تتحرك العديد من النسخ بعيدًا عن الكروموسومات وتصبح نواة عاملة إضافية. هذه العملية ضرورية لتراكم عدد ضخم (10 12) من الريبوسومات في كل خلية بويضة، مما يضمن التطور المستقبلي للجنين عند المراحل الأولىحتى في غياب تخليق الريبوسومات الجديدة. تختفي النوى الزائدة بعد نضوج خلية البويضة.
ه) البنية التحتية للنواة.
عند الدراسة عدد كبير خلايا مختلفةتتميز الحيوانات والنباتات ببنية ليفية أو شبكية للنواة محاطة بكتلة منتشرة أكثر أو أقل كثافة. وقد تم اقتراح أسماء لهذه الأجزاء: الجزء الليفي - nucleoneme والجزء المتجانس المنتشر هو مادة غير متبلورة، أو جزء غير متبلور . كشفت الدراسات المجهرية الإلكترونية التي أجريت في وقت واحد تقريبًا عن البنية الليفية الخيطية للنوية.
ومع ذلك، لا يتم دائمًا التعبير عن هذا الهيكل الخيطي للنوية بشكل واضح. في بعض الخلايا، تندمج الخيوط الفردية من النيوكليونات، ويمكن أن تكون النواة متجانسة تمامًا.
عند الفحص الدقيق للنواة، يمكن للمرء أن يلاحظ أن الشيء الرئيسي مركبات اساسيهالنواة - حبيبات كثيفة يبلغ قطرها حوالي 15 نانومتر وألياف رفيعة يبلغ سمكها 4-8 نانومتر. في كثير من الحالات (بويضات الأسماك والبرمائيات، والخلايا المرستيمية للنباتات)، يتم تجميع المكون الليفي في منطقة مركزية كثيفة (جوهر)، خالية من الحبيبات، وتحتل الحبيبات المنطقة المحيطية للنواة. في بعض الحالات (على سبيل المثال، خلايا جذر النبات)، لم يتم ملاحظة أي بنية إضافية في هذه المنطقة الحبيبية.
وقد وجد أن المناطق غير المتبلورة من النواة غير متجانسة. يكشف هيكلها عن مناطق ملونة قليلاً - مراكز ليفية - والمناطق الداكنة المحيطة بها، والتي تحتوي أيضًا على بنية ليفية.
بالإضافة إلى هذين المكونين من النواة الموجودة في مؤخراتم إيلاء الكثير من الاهتمام لبنية الكروماتين حول النواة. هذه الشبكة من الكروماتين والحمض النووي داخل النواة هي نظام موحدوتمثل جزءا لا يتجزأ من النواة.
تتكون الحبيبات والجزء الليفي من البروتينات النووية الريبية.
وقد تبين أن المراكز الليفية الخفيفة هي التي تحتوي على الحمض النووي الريبوزي (rDNA).