^ غشاء بلازمي ، أو البلازما، تحتل مكانة خاصة بين أغشية الخلايا المختلفة. هذا هيكل محيطي سطحي يحد من الخلية من الخارج ، مما يحدد ارتباطها المباشر بالبيئة خارج الخلية ، وبالتالي مع جميع المواد والمحفزات التي تعمل على الخلية. لذلك ، فإن دور غشاء البلازما هو أن يكون حاجزًا ، حاجزًا بين محتويات الخلايا المنظمة بشكل معقد والبيئة الخارجية. في هذه الحالة ، لا تؤدي البلازمية دور الحاجز الميكانيكي فحسب ، بل والأهم من ذلك أنها تحد من التدفق الحر للمواد منخفضة وعالية الجزيئات في كلا الاتجاهين عبر الغشاء. علاوة على ذلك ، تعمل البلازما كهيكل "يتعرف" على المستقبلات والمواد الكيميائية المختلفة وينظم بشكل انتقائي نقل هذه المواد داخل وخارج الخلية. بمعنى آخر ، يؤدي غشاء البلازما الوظائف المرتبطة بالنقل الانتقائي المنظم للمواد عبر الغشاء ويعمل كمحلل خلوي أولي. في هذا الصدد ، يمكن اعتبار البلازما عضوي الخليةالمدرجة في نظام الفراغ للخلية. مثل الأغشية الأخرى لهذا النظام (أغشية الجسيمات ، الإندوسومات ، جهاز جولجي ، وما إلى ذلك) ، فإنه ينشأ ويتم تحديثه بسبب النشاط التركيبي للشبكة الإندوبلازمية وله تركيبة مماثلة. من الغريب أن غشاء البلازما يمكن تشبيهه بغشاء فجوة داخل الخلايا ، ولكن يتم قلبه من الداخل إلى الخارج: فهو غير محاط بالهيالوبلازم ، ولكنه يحيط به.
دور النقل الحاجز للبلازما
يعمل غشاء البلازما ، الذي يحيط بالخلية من جميع الجوانب ، كحاجز ميكانيكي. من أجل اختراقها باستخدام الإبر الدقيقة أو الماصات الدقيقة ، يلزم بذل الكثير من الجهد. مع ضغط إبرة مجهرية عليها ، تنحني بقوة أولاً ، وبعد ذلك فقط تخترق. الأغشية الدهنية الاصطناعية أقل استقرارًا. يمكن تحديد هذا الاستقرار الميكانيكي لغشاء البلازما من خلال مكونات إضافية مثل glycocalyx والطبقة القشرية من السيتوبلازم (الشكل 127).
مركب السكرعبارة عن طبقة خارجية لغشاء البروتين الدهني تحتوي على سلاسل عديد السكاريد من البروتينات الغشائية المتكاملة - البروتينات السكرية. تحتوي هذه السلاسل على كربوهيدرات مثل المانوز والجلوكوز و N-acetylglucosamine وحمض السياليك ، وما إلى ذلك. تشكل البوليمرات غير المتجانسة الكربوهيدراتية سلاسل متفرعة ، يمكن أن توجد بينها جليكوليبيدات وبروتيوغليكان معزولة عن الخلية. يتم تسخين طبقة glycocalyx بكثافة ، ولها تناسق يشبه الهلام ، مما يقلل بشكل كبير من معدل الانتشار في هذه المنطقة. مواد مختلفة. إنزيمات التحلل المائي التي تفرزها الخلية ، والتي تشارك في الانقسام خارج الخلية للبوليمرات (الهضم خارج الخلية) إلى جزيئات المونومر ، والتي يتم نقلها بعد ذلك إلى السيتوبلازم من خلال غشاء البلازما ، يمكن أيضًا أن "تتعثر" هنا.
في ميكروسكوب الكتروني، خاصة عندما طرق خاصةعلى النقيض من عديد السكاريد ، يكون للكلان الجلدي شكل طبقة ليفية فضفاضة ، بسمك 3-4 نانومتر ، تغطي كامل سطح الخلية. يتم التعبير عن glycocalyx جيدًا بشكل خاص في حدود الفرشاة لخلايا الظهارة المعوية الماصة (الخلايا المعوية) ، ومع ذلك ، فهي موجودة في جميع الخلايا الحيوانية تقريبًا ، ولكن درجة شدتها مختلفة (الشكل 128).
بالإضافة إلى ذلك ، يتم توفير الاستقرار الميكانيكي لغشاء البلازما من خلال بنية الطبقة القشرية المجاورة لها من جانب السيتوبلازم والهياكل الليفية داخل الخلايا. قشري(من كلمة - قشرة - لحاء ، قشر) طبقةالسيتوبلازم الكذب على اتصال وثيق مع البروتين الدهني الغشاء الخارجي، لديه عدد من الميزات. هنا ، بسماكة 0.1-0.5 ميكرون ، لا توجد ريبوسومات وحويصلات غشائية ، ولكن في بأعداد كبيرةهناك عناصر ليفية في السيتوبلازم - خيوط دقيقة وغالبًا ما تكون الأنابيب الدقيقة. المكون الرئيسي ليفية للطبقة القشرية هو شبكة من الألياف الدقيقة أكتين. يوجد هنا أيضًا عدد من البروتينات المساعدة ، وهي ضرورية لحركة أقسام السيتوبلازم (لمزيد من التفاصيل حول نظام الخلايا الهيكلية الحركية ، انظر أدناه). إن دور هذه البروتينات المرتبطة بالأكتين مهم جدًا ، لأنه يفسر مشاركتها في الارتباط ، في "تثبيت" البروتينات المتكاملة لغشاء البلازما.
في العديد من البروتوزوا ، وخاصة ciliates ، يشارك غشاء البلازما في التكوين حبيبات، وهي طبقة صلبة تحدد غالبًا شكل الخلية. يمكن أن تجاور أكياس الغشاء غشاء البلازما هنا من الداخل ؛ في هذه الحالة ، توجد ثلاث طبقات غشاء بالقرب من سطح الخلايا: غشاء البلازما نفسه وغشاءان من الحويصلات الهوائية. في أهداب الحذاء ، تشكل الحبيبات ثخانات ، تقع على شكل سداسيات ، في وسطها تظهر أهداب (الشكل 129). يمكن أيضًا أن ترتبط صلابة التكوينات الحبيبية بعناصر السيتوبلازم الكامنة وراء غشاء البلازما ، مع الطبقة القشرية. وهكذا ، في قمم قشرة الأوجلينا بالقرب من الغشاء ، بالإضافة إلى فجوات الغشاء ، توجد حزم متوازية من الأنابيب الدقيقة والألياف الدقيقة. هذا التعزيز المحيطي الليفي ، جنبًا إلى جنب مع محيط الغشاء متعدد الطبقات المطوي ، يخلق بنية حبيبية صلبة.
يتمثل دور الحاجز للبلازما أيضًا في الحد من الانتشار الحر للمواد. أظهرت التجارب النموذجية على الأغشية الدهنية الاصطناعية أنها قابلة للنفاذ إلى الماء والغازات والجزيئات الصغيرة غير القطبية للمواد القابلة للذوبان في الدهون ، ولكنها غير منفذة تمامًا للجزيئات المشحونة (الأيونات) والأجزاء الكبيرة غير المشحونة (السكريات) (الشكل 130).
تحد الأغشية الطبيعية أيضًا من معدل تغلغل المركبات ذات الوزن الجزيئي المنخفض في الخلية.
النقل عبر الغشاء للأيونات والمركبات منخفضة الوزن الجزيئي
غشاء البلازما ، مثل أغشية خلايا البروتين الدهني الأخرى ، شبه نافذ. هذا يعني أن الجزيئات المختلفة تمر من خلاله بسرعات مختلفة و حجم أكبرالجزيئات ، يكون مرورها أبطأ عبر الغشاء. تحدد هذه الخاصية غشاء البلازما كحاجز تناضحي. يتمتع الماء والغازات المذابة فيه بأقصى قدرة اختراق ، وتخترق الأيونات الغشاء ببطء أكبر (حوالي 10 4 مرات أبطأ). لذلك ، إذا تم وضع خلية ، على سبيل المثال ، كريات الدم الحمراء ، في بيئة يكون فيها تركيز الأملاح أقل مما هو عليه في الخلية (انخفاض ضغط الدم) ، فإن الماء من الخارج سوف يندفع إلى داخل الخلية ، مما يؤدي إلى زيادة حجم الخلية وتمزق غشاء البلازما ("صدمة ناقصة التوتر"). على العكس من ذلك ، عندما يتم وضع كريات الدم الحمراء في المحاليل الملحية ، يحدث المزيد تركيز عاليمما هو عليه في الخلية ، سيكون هناك خروج للمياه من الخلية إلى البيئة الخارجية. في الوقت نفسه ، سوف تتجعد الخلية ويقل حجمها.
مثل هذا النقل السلبي للماء خارج الخلية إلى داخل الخلية لا يزال يسير بمعدل منخفض. يبلغ معدل تغلغل الماء عبر الغشاء حوالي 10-4 سم / ثانية ، وهو أقل 100000 مرة من معدل انتشار جزيئات الماء عبر طبقة مائية بسمك 7.5 نانومتر. وخلص إلى أن في غشاء الخليةيوجد في طبقة البروتين الدهني "مسام" خاصة لاختراق الماء والأيونات. عددهم ليس كبيرًا جدًا: يجب أن تكون المساحة الإجمالية التي يبلغ حجم "المسام" الفردي حوالي 0.3-0.8 نانومتر 0.06٪ فقط من سطح الخلية بأكمله.
على عكس الأغشية الدهنية ثنائية الطبقة الاصطناعية ، فإن الأغشية الطبيعية ، وغشاء البلازما بشكل أساسي ، لا تزال قادرة على نقل الأيونات والعديد من المونومرات ، مثل السكريات والأحماض الأمينية ، وما إلى ذلك. نفاذية الأيونات منخفضة ، ومعدل مرور الأيونات المختلفة هو ليس نفس الشيء. معدل مرور أعلى للكاتيونات (K + ، Na +) وأقل بكثير للأنيونات (Cl -).
يحدث نقل الأيونات عبر غشاء البلازما بسبب المشاركة في هذه العملية لبروتينات نقل الغشاء - بيرميز. يمكن لهذه البروتينات نقل مادة واحدة في اتجاه واحد (uniport) أو عدة مواد في وقت واحد (symport) ، أو ، مع استيراد مادة واحدة ، إزالة أخرى من الخلية (antiport). لذلك ، على سبيل المثال ، يمكن أن يدخل الجلوكوز الخلايا بشكل متناغم مع أيون الصوديوم.
يمكن أن يحدث النقل الأيوني على طول تدرج التركيز - بشكل سلبيبدون استهلاك إضافي للطاقة. على سبيل المثال ، يدخل أيون الصوديوم إلى الخلية من بيئة خارجيةحيث يكون تركيزه أعلى منه في السيتوبلازم. في حالة النقل السلبي ، تشكل بعض بروتينات النقل الغشائي مجمعات جزيئية ، القنواتتمر من خلالها الجزيئات الذائبة عبر الغشاء عن طريق الانتشار البسيط على طول تدرج تركيز. بعض هذه القنوات مفتوحة بشكل دائم ، في حين أن الجزء الآخر يمكن أن يغلق أو ينفتح استجابة إما للارتباط بجزيئات الإشارة أو التغيرات في تركيز الأيونات داخل الخلايا. في حالات أخرى ، بروتينات غشائية خاصة - ناقلاتيرتبط بشكل انتقائي بأيون واحد أو آخر ويحمله عبر الغشاء (الانتشار الميسر) (الشكل 131).
يبدو أن وجود مثل هذه القنوات الناقلة للبروتين يجب أن يؤدي إلى توازن في تركيزات الأيونات والمواد ذات الوزن الجزيئي المنخفض على جانبي الغشاء. في الواقع ، ليس الأمر كذلك: يختلف تركيز الأيونات في سيتوبلازم الخلايا اختلافًا حادًا ليس فقط عن البيئة الخارجية ، ولكن أيضًا عن بلازما الدم التي تغمر الخلايا في جسم الحيوان. على الطاولة يوضح الشكل 14 تركيزات الأيونات داخل وخارج الخلية.
الجدول 14
| وهو | تركيز داخل الخلايا ، مم | تركيز خارج الخلية ، مم |
| نا + | 5-15 | 145 |
| ك + | 140 | 5 |
| ملغ 2 + | 30 | 1-2 |
| * Ca2 + | 1-2 | 2,5-5 |
| Cl- | 4 | 110 |
* تركيز Ca 2+ في الحالة الحرة في العصارة الخلوية للخلايا حقيقية النواة هو 10 -7 م ، وخارجها 10 -3 م.
كما يتضح ، في هذه الحالة ، يكون التركيز الكلي للكاتيونات أحادية التكافؤ داخل الخلايا وخارجها متماثلًا تقريبًا (150 مم) ، مساوي التوتر. ولكن اتضح أنه في السيتوبلازم ، يكون تركيز K + أعلى بحوالي 50 مرة ، و Na + أقل منه في بلازما الدم. علاوة على ذلك ، يتم الحفاظ على هذا الاختلاف فقط في الخلية الحية: إذا قُتلت الخلية أو تم قمع عمليات التمثيل الغذائي فيها ، فستختفي الاختلافات الأيونية على جانبي غشاء البلازما بعد فترة. يمكنك ببساطة تبريد الخلايا إلى +2 0 درجة مئوية ، وبعد فترة سيصبح تركيز K + و Na + على جانبي الغشاء كما هو. عندما يتم تسخين الخلايا ، يتم استعادة هذا الاختلاف. ترجع هذه الظاهرة إلى حقيقة وجود ناقلات بروتين غشائية في الخلايا تعمل ضد تدرج التركيز ، بينما تنفق الطاقة بسبب التحلل المائي ATP. هذا النوع من العمل يسمى النقل النشط، ويتم تنفيذه بمساعدة البروتين مضخات الأيونات. يحتوي غشاء البلازما على جزيء من وحدتين (K + + Na +) - مضخة ، وهي أيضًا ATPase. أثناء التشغيل ، تضخ هذه المضخة 3 أيونات Na + في دورة واحدة وتضخ 2 K + أيونات في الخلية مقابل تدرج التركيز. في هذه الحالة ، يتم إنفاق جزيء ATP واحد ، والذي ينتقل إلى فسفرة ATPase ، ونتيجة لذلك يتم نقل Na + عبر الغشاء من الخلية ، ويحصل K + على فرصة الارتباط بجزيء البروتين ثم يتم نقله إلى خلية (الشكل 132). نتيجة للنقل النشط بمساعدة المضخات الغشائية ، يتم أيضًا تنظيم تركيز الكاتيونات ثنائية التكافؤ Mg 2+ و Ca 2+ في الخلية ، وكذلك مع استهلاك ATP.
هذه وظيفة بدوام كامليخلق النفاث والمضخات تركيزًا ثابتًا للأيونات والمواد ذات الوزن الجزيئي المنخفض في الخلية ، ويخلق ما يسمى. التوازن ، ثبات التراكيز تناضحيًا المواد الفعالة. وتجدر الإشارة إلى أن ما يقرب من 80٪ من إجمالي ATP للخلية يتم إنفاقه على الحفاظ على التوازن.
بالاشتراك مع النقل النشط للأيونات عبر غشاء البلازما ، يتم نقل مختلف السكريات والنيوكليوتيدات والأحماض الأمينية.
وبالتالي ، فإن النقل النشط للجلوكوز ، والذي يدخل الخلية بشكل متناغم (في وقت واحد) مع تدفق أيون الصوديوم المنقول بشكل سلبي ، سيعتمد على نشاط المضخة (K + + Na +). إذا تم حظر هذه المضخة (K + -Na +) ، فحينئذٍ سيختفي الاختلاف في تركيز Na على جانبي الغشاء ، بينما سينخفض انتشار Na + في الخلية ، وفي نفس الوقت سيختفي الاختلاف في تركيز Na سيتوقف تدفق الجلوكوز في الخلية. بمجرد استعادة عمل (K + -Na +) - ATPase ويتم إنشاء اختلاف في تركيز الأيونات ، يزداد التدفق المنتشر لـ Na + على الفور وفي نفس الوقت نقل الجلوكوز. وبالمثل ، من خلال الغشاء وتدفق الأحماض الأمينية ، والتي يتم نقلها بواسطة بروتينات حاملة خاصة تعمل كنظم رمزية ، وتنقل الأيونات في نفس الوقت.
يرجع النقل النشط للسكريات والأحماض الأمينية في الخلايا البكتيرية إلى تدرج أيونات الهيدروجين.
في حد ذاته ، تشير مشاركة بروتينات غشائية خاصة تشارك في النقل السلبي أو النشط للمركبات ذات الوزن الجزيئي المنخفض إلى الخصوصية العالية لهذه العملية. حتى في حالة النقل الأيوني السلبي ، فإن البروتينات "تتعرف" على أيون معين وتتفاعل معه وترتبط على وجه التحديد وتغير شكلها ووظيفتها. لذلك ، بالفعل على سبيل المثال النقل مواد بسيطةتعمل الأغشية كمحللات وكمستقبلات. يتجلى دور المستقبل هذا بشكل خاص عندما تمتص الخلية البوليمرات الحيوية.
النقل الحويصلي: الالتقام الخلوي والإفراز الخلوي
لا تمر الجزيئات الضخمة مثل البروتينات والأحماض النووية والسكريات المتعددة ومجمعات البروتين الدهني وغيرها من خلال أغشية الخلايا ، على عكس كيفية نقل الأيونات والمونومرات. يحدث نقل الجزيئات الدقيقة ومجمعاتها والجسيمات داخل وخارج الخلية بطريقة مختلفة تمامًا - من خلال النقل الحويصلي. يعني هذا المصطلح أن الجزيئات الكبيرة أو البوليمرات الحيوية أو مجمعاتها لا يمكنها دخول الخلية عبر غشاء البلازما. وليس فقط من خلاله: أي أغشية خلوية غير قادرة على نقل البوليمرات الحيوية عبر الغشاء ، باستثناء الأغشية التي تحتوي على ناقلات بروتينية خاصة - بورينات (أغشية الميتوكوندريا والبلاستيدات والبيروكسيسومات). تدخل الجزيئات الكبيرة الخلية أو من حجرة غشائية إلى أخرى محاطة بالفجوات أو الحويصلات. هذه نقل حويصلييمكن تقسيمها إلى نوعين: طرد خلوي- إزالة المنتجات الجزيئية من الخلية الالتقام- امتصاص الخلية للجزيئات الكبيرة (الشكل 133).
أثناء الالتقام الخلوي ، يلتقط جزء معين من غشاء البلازما ، كما هو الحال ، مادة خارج الخلية ، محاطًا بها في فجوة غشائية نشأت بسبب غزو غشاء البلازما. في مثل هذه الفجوة الأولية ، أو في جسيم داخلي، يمكن لأي بوليمرات حيوية أو مجمعات جزيئية كبيرة أو أجزاء من الخلايا أو حتى خلايا كاملة الدخول ، حيث تتحلل بعد ذلك ، وتتحول إلى مونومرات ، تدخل الهيالوبلازم عن طريق نقل الغشاء. الأهمية البيولوجية الرئيسية للالتقام الخلوي هو اكتساب اللبنات الأساسية من خلال الهضم داخل الخلايا، والذي يتم إجراؤه في المرحلة الثانية من الالتقام الخلوي بعد اندماج الجسيم الداخلي الأولي مع الليزوزوم ، وهو فجوة تحتوي على مجموعة من الإنزيمات المتحللة بالماء (انظر أدناه).
ينقسم الالتقام رسميا إلى كثرة الخلاياو البلعمة(الشكل 134). البلعمة - التقاط الجسيمات الكبيرة وامتصاصها بواسطة الخلية (أحيانًا حتى الخلايا أو أجزائها) - تم وصفه لأول مرة بواسطة I.I. Mechnikov. تم العثور على البلعمة ، وهي القدرة على التقاط الجزيئات الكبيرة بواسطة الخلية ، بين الخلايا الحيوانية ، سواء وحيدة الخلية (على سبيل المثال ، الأميبا ، بعض الأهداب المفترسة) والخلايا المتخصصة للحيوانات متعددة الخلايا. الخلايا المتخصصة ، البلعمية ، هي خصائص كل من اللافقاريات (الخلايا الأميبية للدم أو سائل التجويف) والفقاريات (العدلات والضامة). تم تعريف كثرة الخلايا في الأصل على أنها امتصاص الماء أو محاليل مائية مواد مختلفة. من المعروف الآن أن كلا من البلعمة والكثرة يتقدمان بشكل متشابه للغاية ، وبالتالي فإن استخدام هذه المصطلحات يمكن أن يعكس فقط الاختلافات في أحجام وكتلة المواد الممتصة. ما تشترك فيه هذه العمليات هو أن المواد الممتصة على سطح غشاء البلازما محاطة بغشاء على شكل فجوة - جسيم داخلي يتحرك داخل الخلية.
يمكن أن يكون الالتقام الخلوي ، بما في ذلك كثرة الوراثة والبلعمة ، غير محدد أو تأسيسي ، ودائمًا ومحددًا ، بوساطة مستقبلات (مستقبلات). شحم غير محدد h (كثرة الخلايا البلعمية) ، وهذا ما يسمى لأنه يستمر كما لو كان تلقائيًا ويمكن أن يؤدي في كثير من الأحيان إلى التقاط وامتصاص مواد غريبة تمامًا أو غير مبالية بالخلية ، على سبيل المثال ، جزيئات السخام أو الأصباغ.
غالبًا ما يكون الالتقام الخلوي غير النوعي مصحوبًا بالامتصاص الأولي للمادة المحبوسة بواسطة glycocalyx في plasmolemma. يحتوي الكاليكس ، بسبب المجموعات الحمضية لعديد السكاريد ، على شحنة سالبة ويرتبط جيدًا بمجموعات مختلفة من البروتينات موجبة الشحنة. مع هذا الامتصاص الالتقام غير النوعي ، يتم امتصاص الجزيئات الكبيرة والجزيئات الصغيرة (البروتينات الحمضية ، الفيريتين ، الأجسام المضادة ، الفيروسات ، الجسيمات الغروانية). يؤدي كثرة الخلايا في الطور السائل إلى الامتصاص مع الوسط السائل للجزيئات القابلة للذوبان التي لا ترتبط بالبلازما.
على الخطوة التاليةهناك تغيير في شكل سطح الخلية: إما ظهور انغالات صغيرة لغشاء البلازما ، أو الانغماس ، أو ظهور نتوءات أو ثنيات أو "رتوش" على سطح الخلية (rafl - in اللغة الإنجليزية) ، والتي ، كما كانت ، تتداخل ، تطوى ، تفصل كميات صغيرة من الوسط السائل (الشكل 135 ، 136). النوع الأول من حدوث الحويصلة الصنوبرية ، هو نموذجي لخلايا الظهارة المعوية ، البطانة ، للأميبا ، والثاني - للبلعمات والخلايا الليفية. تعتمد هذه العمليات على إمداد الطاقة: تمنع مثبطات التنفس هذه العمليات.
بعد إعادة ترتيب السطح هذه ، تتبع عملية الالتصاق والاندماج للأغشية الملامسة ، مما يؤدي إلى تكوين حويصلة قشرية (صنوبرية) ، تنفصل عن سطح الخلية وتتعمق في السيتوبلازم. يحدث كل من الالتقام غير النوعي والمستقبلات ، مما يؤدي إلى انقسام حويصلات الغشاء ، في مناطق متخصصة من غشاء البلازما. هذه هي ما يسمى ب حفر يحدها. يطلق عليهم ذلك لأنه من جانب السيتوبلازم ، فإن غشاء البلازما مغطى ، ومغطى بطبقة ليفية رقيقة (حوالي 20 نانومتر) ، والتي في أقسام رقيقة للغاية ، كما كانت ، حدود ، تغطي نتوءات صغيرة ، حفر (الشكل. 137). تحتوي جميع الخلايا الحيوانية تقريبًا على هذه الحفر ؛ فهي تحتل حوالي 2٪ من سطح الخلية. الطبقة المحيطة تتكون أساسًا من البروتين كلاذرينيرتبط بعدد من البروتينات الإضافية. تشكل ثلاثة جزيئات من الكلاذرين ، مع ثلاثة جزيئات من بروتين منخفض الوزن الجزيئي ، بنية triskelion ، التي تشبه صليبًا معقوفًا ثلاثي الشعاع (الشكل 138). تشكل ارتفاعات الكلاذرين على السطح الداخلي لحفر غشاء البلازما شبكة فضفاضة تتكون من خماسيات وسداسية الشكل تشبه السلة عمومًا. تغطي طبقة الكلاذرين كامل محيط فجوات الالتحام الأولية الفاصلة ، والتي تحدها الحويصلات.
ينتمي Clathrin إلى أحد الأنواع المزعومة. البروتينات "المبطنة" (البروتينات المغلفة COP). ترتبط هذه البروتينات ببروتينات مستقبلات متكاملة من جانب السيتوبلازم وتشكل طبقة ضماد حول محيط الصنوبر الناشئ ، الحويصلة الداخلية الأولية - الحويصلة "الحدودية". في فصل الإندوسوم الأولي ، تشارك البروتينات أيضًا - الدينامينات ، التي تتبلمر حول عنق الحويصلة الفاصلة (الشكل 139).
بعد أن تنفصل الحويصلة الحدودية عن بلازما الدم وتبدأ في الانتقال إلى عمق السيتوبلازم ، تتفكك طبقة الكلاثرين ، وتتفكك ، ويكتسب الغشاء الداخلي (الصنوبر) شكله المعتاد. بعد فقدان طبقة الكلاذرين ، تبدأ الجسيمات الداخلية في الاندماج مع بعضها البعض.
وقد وجد أن أغشية الحفر المجاورة تحتوي على نسبة قليلة نسبيًا من الكوليسترول ، والتي يمكن أن تحدد انخفاض تصلب الأغشية وتساهم في تكوين الفقاعات. قد يكون المعنى البيولوجي لظهور "غلاف" الكلاذرين على طول محيط الحويصلات هو أنه يوفر التصاق الحويصلات المجاورة بعناصر الهيكل الخلوي ونقلها اللاحق في الخلية ، ويمنعها من الاندماج مع بعضها البعض .
يمكن أن تكون شدة كثرة الخلايا غير النوعية في المرحلة السائلة عالية جدًا. لذا فإن الخلية الظهارية الأمعاء الدقيقةتشكل ما يصل إلى 1000 pinosomes في الثانية ، وتشكل الضامة حوالي 125 pinosomes في الدقيقة. حجم الصنوبر صغير ، الحد الأدنى لها هو 60-130 نانومتر ، ولكن وفرتها تؤدي إلى حقيقة أنه أثناء الالتقام الخلوي ، يتم استبدال البلازما الليفية بسرعة ، كما لو كانت "تنفق" على تكوين العديد من الفجوات الصغيرة. لذلك في البلاعم ، يتم استبدال غشاء البلازما بالكامل في غضون 30 دقيقة ، في الخلايا الليفية - في غضون ساعتين.
قد يكون المصير الإضافي للاندوسومات مختلفًا ، فقد يعود بعضها إلى سطح الخلية وتندمج معه ، ولكن معظميدخل في عملية الهضم داخل الخلايا. تحتوي الإندوسومات الأولية في الغالب على جزيئات غريبة محاصرة في الوسط السائل ولا تحتوي على إنزيمات متحللة للماء. يمكن أن تندمج الجسيمات الداخلية مع بعضها البعض مع زيادة الحجم. ثم يندمجون مع الجسيمات الأولية (انظر أدناه) ، والتي تدخل الإنزيمات في تجويف الجسيم الداخلي الذي يحلل البوليمرات الحيوية المختلفة. يتسبب عمل هذه الهيدرولات الليزوزومية في حدوث هضم داخل الخلايا - تحلل البوليمرات إلى مونومرات.
كما ذكرنا سابقًا ، خلال عملية البلعمة والكثافة ، تفقد الخلايا مساحة كبيرة من البلازما (انظر الضامة) ، والتي ، مع ذلك ، يتم استعادتها بسرعة أثناء إعادة تدوير الغشاء ، بسبب عودة الفجوات وإدماجها في البلازما. ويرجع ذلك إلى حقيقة أن الحويصلات الصغيرة يمكن أن تنفصل عن الإندوسومات أو الفجوات ، وكذلك من الجسيمات الحالة التي تندمج مرة أخرى مع غشاء البلازما. مع إعادة التدوير هذه ، يحدث نوع من النقل "المكوكي" للأغشية: البلازما - بينوسوم - فجوة - بلازما. هذا يؤدي إلى استعادة المنطقة الأصلية لغشاء البلازما. وقد وجد أنه مع مثل هذه العودة ، إعادة تدوير الغشاء ، يتم الاحتفاظ بجميع المواد الممتصة في الجسيم الداخلي المتبقي.
محددأو مستقبلات بوساطةلدى الالتقام الخلوي عدد من الاختلافات عن غير المحددة. الشيء الرئيسي هو أن الجزيئات يتم امتصاصها حيث توجد مستقبلات محددة على غشاء البلازما مرتبطة فقط بهذا النوع من الجزيئات. غالبًا ما تسمى هذه الجزيئات التي ترتبط ببروتينات المستقبل على سطح الخلايا يجند.
تم وصف الالتقام الخلوي بوساطة المستقبلات لأول مرة في تراكم البروتينات في بويضات الطيور. يتم تصنيع بروتينات حبيبات الصفار ، فيتيلوجينين ، في الأقمشة المختلفة، ولكن بعد ذلك يدخلون المبيضين مع تدفق الدم ، حيث يرتبطون بمستقبلات غشائية خاصة للبويضات ، وبعد ذلك ، بمساعدة الالتقام الخلوي ، يدخلون إلى الخلية ، حيث يحدث ترسب حبيبات الصفار.
مثال آخر على الالتقام الخلوي الانتقائي هو نقل الكوليسترول إلى الخلية. يتم تصنيع هذا الدهن في الكبد ، وبالاقتران مع الدهون الفوسفورية الأخرى وجزيء البروتين ، يشكل ما يسمى. البروتين الدهني منخفض الكثافة (LDL) ، والذي يفرزه الكبد و نظام الدورة الدمويةينتشر في جميع أنحاء الجسم (الشكل 140). مستقبلات غشاء البلازما المخصصة منتشرة على السطح خلايا مختلفة، يتعرف على مكون البروتين في LDL ، ويشكل معقدًا محددًا لمستقبلات يجند. بعد ذلك ، ينتقل مثل هذا المركب إلى منطقة الحفر المجاورة ويتم استيعابها - محاطًا بغشاء ومغمور في أعماق السيتوبلازم. لقد ثبت أن المستقبلات الطافرة يمكن أن تربط LDL ، لكنها لا تتراكم في منطقة الحفر المجاورة. بالإضافة إلى مستقبلات LDL ، تم العثور على أكثر من عشرين مادة أخرى متورطة في الالتقام الخلوي للمستقبلات لمواد مختلفة ، وكلها تستخدم نفس مسار الاستيعاب عبر الحفر الحدودية. من المحتمل أن يكون دورهم في تراكم المستقبلات: يمكن لنفس الحفرة الحدودية أن تجمع حوالي 1000 مستقبل من فئات مختلفة. ومع ذلك ، في الخلايا الليفية ، توجد مجموعات مستقبلات LDL في منطقة الحفر المجاورة حتى في حالة عدم وجود رابط في الوسط.
المصير الآخر لجسيم LDL الممتص هو أنه يخضع للاضمحلال في التركيب ليسوسوم ثانوي. بعد الانغماس في السيتوبلازم لحويصلة حدية محملة بـ LDL ، يحدث فقدان سريع لطبقة clathrin ، وتبدأ الحويصلات الغشائية في الاندماج مع بعضها البعض ، وتشكل فجوة داخلية تحتوي على جزيئات LDL الممتصة التي لا تزال مرتبطة بمستقبلات على سطح الغشاء . ثم يحدث تفكك معقد مستقبلات ligand ، تنفصل فجوات صغيرة عن الجسيم الداخلي ، التي تحتوي أغشيةها على مستقبلات حرة. يتم إعادة تدوير هذه الحويصلات ، ودمجها في غشاء البلازما ، وبالتالي تعود المستقبلات إلى سطح الخلية. مصير LDL هو أنه بعد الاندماج مع الجسيمات الحالة ، يتم تحللها بالماء لتحرير الكوليسترول ، والذي يمكن دمجه في أغشية الخلايا.
تتميز الإندوسومات بقيمة أقل من الرقم الهيدروجيني (درجة الحموضة 4-5) ، وبيئة حمضية أكثر من فجوات الخلايا الأخرى. هذا بسبب وجود البروتينات في أغشيتها. مضخة البروتون، ضخ أيونات الهيدروجين مع الاستهلاك المتزامن لـ ATP (H + المعتمدة على ATPase). تلعب البيئة الحمضية داخل الإندوسومات دورًا مهمًا في تفكك المستقبلات والروابط. بجانب، بيئة حمضيةهو الأمثل لتنشيط الإنزيمات المتحللة للماء في تكوين الجسيمات الحالة ، والتي يتم تنشيطها عند اندماج الجسيمات الحالة مع الإندوسومات وتؤدي إلى تكوين الجسيمات الداخلية، حيث يحدث انقسام البوليمرات الحيوية الممتصة.
في بعض الحالات ، لا يرتبط مصير الترابطات المنفصلة بالتحلل المائي الليزوزومي. لذلك في بعض الخلايا ، بعد ارتباط مستقبلات البلازما ببروتينات معينة ، تغرق الفجوات المغلفة بالكالاثرين في السيتوبلازم ويتم نقلها إلى منطقة أخرى من الخلية ، حيث تندمج مرة أخرى مع غشاء البلازما ، وتنفصل البروتينات المرتبطة عن مستقبلات. هذه هي الطريقة التي يتم بها نقل بعض البروتينات من خلال جدار الخلية البطانية من بلازما الدم إلى البيئة بين الخلايا (الشكل 141). مثال آخر على انتقال الخلايا هو نقل الأجسام المضادة. لذلك في الثدييات ، يمكن أن تنتقل الأجسام المضادة للأم إلى الشبل من خلال اللبن. في هذه الحالة ، يظل معقد المستقبل والجسم المضاد دون تغيير في الجسيم الداخلي.
البلعمة
كما ذكرنا سابقًا ، فإن البلعمة هي نوع من الالتقام الخلوي وترتبط بامتصاص الخلية للتجمعات الكبيرة من الجزيئات الكبيرة حتى الخلايا الحية أو الميتة. بالإضافة إلى كثرة الخلايا ، يمكن أن يكون البلعمة غير محدد (على سبيل المثال ، امتصاص جزيئات الذهب الغرواني أو بوليمر ديكستران بواسطة الخلايا الليفية أو الضامة) ومحددة ، بوساطة مستقبلات على سطح غشاء البلازما للخلايا البلعمية. أثناء البلعمة ، تتشكل فجوات كبيرة داخلية - بلعم، والتي تندمج بعد ذلك مع الجسيمات الحالة لتشكل البلعمة.
على سطح الخلايا القادرة على البلعمة (في الثدييات ، هذه هي العدلات والضامة) ، هناك مجموعة من المستقبلات التي تتفاعل مع بروتينات الترابط. لذلك في الالتهابات البكتيريةترتبط الأجسام المضادة للبروتينات البكتيرية بالأسطح الخلايا البكتيرية، وتشكل طبقة تبدو فيها المنطقة F c للأجسام المضادة للخارج. يتم التعرف على هذه الطبقة من خلال مستقبلات محددة على سطح الضامة والعدلات ، وفي مواقع ارتباطها ، يبدأ امتصاص البكتيريا بتغليفها بغشاء البلازما للخلية (الشكل 142).
طرد خلوي
يشارك غشاء البلازما في إزالة المواد من الخلية بمساعدة طرد خلوي- عملية الالتقام العكسي (انظر الشكل 133).
في حالة الإفراز الخلوي ، يتم وضع المنتجات داخل الخلايا في فجوات أو حويصلات ومنفصلة عن الهيالوبلازم بواسطة غشاء تقترب من غشاء البلازما. عند نقاط الاتصال الخاصة بهم ، يندمج غشاء البلازما وغشاء الفجوة ، ويتم إفراغ الفقاعة في البيئة. بمساعدة الإخراج الخلوي ، تحدث عملية إعادة تدوير الأغشية المشاركة في الالتقام الخلوي.
يرتبط خروج الخلايا بإفراز مواد مختلفة يتم تصنيعها في الخلية. من خلال إفراز المواد وإطلاقها في البيئة ، يمكن للخلايا إنتاج وإطلاق مركبات منخفضة الوزن الجزيئي (أستيل كولين ، أمينات حيوية المنشأ ، وما إلى ذلك) ، وكذلك في معظم الحالات الجزيئات الكبيرة (الببتيدات ، والبروتينات ، والبروتينات الدهنية ، والببتيدوغليكان ، وما إلى ذلك). يحدث الإفراز أو الإفراز في معظم الحالات استجابة لإشارة خارجية (النبضات العصبية ، والهرمونات ، والوسطاء ، وما إلى ذلك). على الرغم من أنه في بعض الحالات يحدث خروج الخلايا باستمرار (إفراز الفبرونيكتين والكولاجين بواسطة الخلايا الليفية). وبالمثل ، تتم إزالة بعض السكريات (الهيميسليلوز) المشاركة في تكوين جدران الخلايا من سيتوبلازم الخلايا النباتية.
يتم استخدام معظم المواد التي يتم إفرازها من قبل خلايا أخرى من الكائنات متعددة الخلايا (إفراز الحليب ، عصارات الجهاز الهضمي ، الهرمونات ، إلخ). ولكن في كثير من الأحيان تفرز الخلايا مواد لاحتياجاتها الخاصة. على سبيل المثال ، يتم تنفيذ نمو غشاء البلازما عن طريق تضمين أقسام من الغشاء كجزء من فجوات خارجية ، تفرز الخلية بعض عناصر الكاليكس في شكل جزيئات بروتين سكري ، إلخ.
يمكن امتصاص إنزيمات التحلل المائي المعزولة من الخلايا عن طريق الإفراز الخلوي في الطبقة السكرية وتوفر انقسامًا خارج الخلية مرتبطًا بالغشاء من البوليمرات الحيوية المختلفة والجزيئات العضوية. الهضم غير الخلوي الغشائي له أهمية كبيرة للحيوانات. تم العثور على ذلك في ظهارة معويةتم العثور على ثدييات في منطقة ما يسمى بحدود الفرشاة لظهارة الشفط ، وهي غنية بشكل خاص بالكلان السكري. كمية كبيرةمجموعة متنوعة من الإنزيمات. بعض هذه الإنزيمات من أصل بنكرياس (الأميليز ، والليباز ، والبروتينات المختلفة ، وما إلى ذلك) ، والبعض الآخر تفرزه الخلايا الظهارية نفسها (exohydrolases ، التي تتحلل بشكل أساسي oligomers و dimers مع تشكيل المنتجات المنقولة).
دور مستقبلات البلازما
لقد التقينا بالفعل بهذه الميزة لغشاء البلازما عند التعرف على وظائف النقل الخاصة به. البروتينات والمضخات الحاملة هي أيضًا مستقبلات تتعرف على أيونات معينة وتتفاعل معها. ترتبط بروتينات المستقبلات بالرابطات وتشارك في اختيار الجزيئات التي تدخل الخلايا.
بروتينات الغشاء أو عناصر جليكوكاليكس - يمكن للبروتينات السكرية أن تعمل كمستقبلات على سطح الخلية. يمكن أن تتناثر هذه المواقع الحساسة للمواد الفردية على سطح الخلية أو يتم جمعها في مناطق صغيرة.
قد تحتوي الخلايا المختلفة للكائنات الحيوانية على مجموعات مختلفة من المستقبلات أو حساسية مختلفة لنفس المستقبلات.
لا يقتصر دور العديد من المستقبلات الخلوية على ارتباط مواد معينة أو القدرة على الاستجابة لها العوامل الفيزيائيةولكن أيضًا في نقل الإشارات بين الخلايا من السطح إلى الخلية. في الوقت الحاضر ، تمت دراسة نظام إرسال الإشارات إلى الخلايا بمساعدة هرمونات معينة ، والتي تشمل سلاسل الببتيد ، جيدًا. تم العثور على هذه الهرمونات مرتبطة بمستقبلات محددة على سطح غشاء البلازما للخلية. تقوم المستقبلات ، بعد الارتباط بالهرمون ، بتنشيط بروتين آخر موجود بالفعل في الجزء السيتوبلازمي من غشاء البلازما ، وهو adenylate cyclase. يصنع هذا الإنزيم جزيء AMP الدوري من ATP. يتمثل دور AMP الدوري (cAMP) في أنه رسول ثانوي - منشط للإنزيمات - كينازات تسبب تعديلات لبروتينات الإنزيم الأخرى. لذلك ، عندما يعمل هرمون البنكرياس الجلوكاجون ، الذي تنتجه الخلايا A لجزر لانجرهانز ، على خلايا الكبد ، يرتبط الهرمون بمستقبل معين ، مما يحفز تنشيط إنزيم الأدينيلات. ينشط cAMP المركب بروتين كيناز أ ، والذي بدوره ينشط سلسلة من الإنزيمات التي تكسر الجليكوجين (عديد السكاريد لتخزين الحيوانات) إلى الجلوكوز. عمل الأنسولين هو عكس ذلك - فهو يحفز دخول الجلوكوز إلى خلايا الكبد وترسبه في شكل الجليكوجين.
بشكل عام ، تتكشف سلسلة الأحداث على النحو التالي: يتفاعل الهرمون بشكل خاص مع جزء المستقبل من هذا النظام ، ودون اختراق الخلية ، ينشط adenylate cyclase ، الذي يصنع cAMP ، الذي ينشط أو يثبط إنزيمًا داخل الخلايا أو مجموعة من الإنزيمات . وهكذا ، فإن الأمر ، الإشارة من غشاء البلازما تنتقل داخل الخلية. كفاءة نظام محلقة الأدينيلات عالية جدًا. وبالتالي ، يمكن أن يؤدي تفاعل جزيء هرموني واحد أو عدة جزيئات هرمونية ، بسبب تخليق العديد من جزيئات cAMP ، إلى تضخيم الإشارة آلاف المرات. في هذه الحالة ، يعمل نظام cyclase adenylate كمحول للإشارات الخارجية.
هناك طريقة أخرى يتم فيها استخدام رسل ثانوي آخر - وهذا ما يسمى. مسار فوسفاتيديلينوسيتول. تحت تأثير إشارة مناسبة (بعض وسطاء الأعصاب والبروتينات) ، يتم تنشيط إنزيم فسفوليباز C ، والذي يشق فوسفاتيديلينوسيتول فوسفوليبيد ، وهو جزء من غشاء البلازما. من ناحية أخرى ، تعمل منتجات التحلل المائي لهذا الدهن على تنشيط بروتين كيناز سي الذي ينشط سلسلة كيناز مما يؤدي إلى تفاعلات خلوية معينة ، ومن ناحية أخرى يؤدي إلى إطلاق أيونات الكالسيوم التي تنظم خط كاملالعمليات الخلوية.
مثال آخر على نشاط المستقبلات هو مستقبلات الأسيتيل كولين ، وهو ناقل عصبي مهم. صدر أستيل كولين من نهاية العصب، يرتبط بالمستقبل الموجود على الألياف العضلية ، ويسبب تدفقًا اندفاعيًا لـ Na + في الخلية (إزالة استقطاب الغشاء) ، مما يفتح على الفور حوالي 2000 قناة أيونية في منطقة النهاية العصبية العضلية.
يؤدي تنوع وخصوصية مجموعات المستقبلات الموجودة على سطح الخلايا إلى إنشاء نظام معقد للغاية من العلامات التي تجعل من الممكن تمييز خلايا الفرد (من نفس الفرد أو من نفس النوع) عن خلايا الآخرين. تدخل الخلايا المماثلة في تفاعلات مع بعضها البعض ، مما يؤدي إلى التصاق الأسطح (الاقتران في البروتوزوا والبكتيريا ، وتشكيل مجمعات خلايا الأنسجة). في هذه الحالة ، يتم استبعاد الخلايا التي تختلف في مجموعة الواسمات المحددة أو التي لا تدركها من هذا التفاعل ، أو يتم تدميرها في الحيوانات الأعلى نتيجة للتفاعلات المناعية (انظر أدناه).
يرتبط غشاء البلازما بتوطين مستقبلات معينة تستجيب للعوامل الفيزيائية. لذلك ، في غشاء البلازما أو مشتقاته في بكتيريا التمثيل الضوئي والطحالب الخضراء المزرقة ، تتفاعل بروتينات المستقبل (الكلوروفيل) مع الكميات الخفيفة. يوجد في الغشاء البلازمي للخلايا الحيوانية الحساسة للضوء نظام خاص لبروتينات المستقبلات الضوئية (رودوبسين) ، والتي يتم من خلالها تحويل إشارة الضوء إلى إشارة كيميائية ، مما يؤدي بدوره إلى توليد نبضة كهربائية.
التعرف بين الخلايا
في الكائنات متعددة الخلايا ، بسبب التفاعلات بين الخلايا ، تتشكل مجموعات خلوية معقدة ، يمكن إجراء صيانتها طرق مختلفة. في الأنسجة الجرثومية والجنينية ، وخاصة على المراحل الأولىفي التطور ، تظل الخلايا متصلة ببعضها البعض بسبب قدرة أسطحها على الالتصاق ببعضها البعض. هذا العقار التصاقيمكن تحديد (اتصال ، التصاق) الخلايا من خلال خصائص سطحها ، والتي تتفاعل على وجه التحديد مع بعضها البعض. تمت دراسة آلية هذه الوصلات جيدًا ، ويتم توفيرها من خلال التفاعل بين البروتينات السكرية لأغشية البلازما. مع هذا التفاعل بين الخلايا بين أغشية البلازما ، تظل هناك دائمًا فجوة بعرض 20 نانومتر ، مليئة بالجلد السكري. علاج الأنسجة بالأنزيمات التي تنتهك سلامة جلايكوكاليكس (الغشاء المخاطي الذي يعمل مائيًا على الغشاء المخاطي ، عديدات السكاريد المخاطية) أو يتلف غشاء البلازما (البروتياز) يؤدي إلى عزل الخلايا عن بعضها البعض ، مما يؤدي إلى تفككها. ومع ذلك ، إذا تمت إزالة عامل التفكك ، يمكن للخلايا أن تتجمع وتتجمع. لذلك من الممكن فصل خلايا الإسفنج بألوان مختلفة البرتقالي والأصفر. اتضح أن هناك نوعين من الركام يتشكلان في خليط هذه الخلايا: تلك التي تتكون من خلايا صفراء فقط وخلايا برتقالية فقط. في هذه الحالة ، تقوم المعلقات الخلوية المختلطة بالتنظيم الذاتي ، واستعادة الهيكل الأصلي متعدد الخلايا. تم الحصول على نتائج مماثلة مع المعلقات الخلوية المنفصلة لأجنة البرمائيات. في هذه الحالة ، هناك فصل مكاني انتقائي لخلايا الأديم الظاهر من الأديم الباطن ومن اللحمة المتوسطة. علاوة على ذلك ، إذا تم استخدام الأنسجة للتجميع المراحل المتأخرةتطور الأجنة ، ثم تتجمع مجموعات مختلفة من الخلايا ذات خصوصية الأنسجة والأعضاء بشكل مستقل في أنبوب اختبار ، وتتشكل مجاميع ظهارية ، على غرار الأنابيب الكلوية، إلخ.
وجد أن البروتينات السكرية عبر الغشاء هي المسؤولة عن تراكم الخلايا المتجانسة. مباشرة عن الاتصال والالتصاق والخلايا مسؤولة عن جزيئات ما يسمى. بروتينات CAM (جزيئات التصاق الخلية). بعضها يربط الخلايا مع بعضها البعض بسبب التفاعلات بين الجزيئات ، والبعض الآخر يشكل اتصالات أو جهات اتصال خاصة بين الخلايا.
يمكن أن تكون التفاعلات بين البروتينات اللاصقة محبة للمثليينعندما ترتبط الخلايا المجاورة ببعضها البعض بمساعدة جزيئات متجانسة ، غير متجانسعندما تشارك أنواع مختلفة من CAM في الخلايا المجاورة في الالتصاق. يحدث الارتباط بين الخلايا من خلال جزيئات رابط إضافية.
هناك عدة فئات من بروتينات CAM. هذه هي الكاديرينات ، الغلوبولين المناعي مثل N-CAM (جزيئات التصاق الخلايا العصبية) ، سيليكتينز ، إنتغرينات.
كاديرينزهي بروتينات غشاء ليفية متكاملة تشكل متجانسات متوازية. ترتبط المجالات المنفصلة لهذه البروتينات بأيونات Ca 2+ ، مما يمنحها صلابة معينة. هناك أكثر من 40 نوعًا من الكاديرين. وهكذا ، فإن E-cadherin هو سمة من سمات خلايا الأجنة المزروعة مسبقًا والخلايا الظهارية للكائنات البالغة. يعتبر P-cadherin سمة مميزة لخلايا الأرومة الغاذية والمشيمة والبشرة ؛ يقع N-cadherin على سطح الخلايا العصبية وخلايا العدسة وعلى عضلات القلب والهيكل العظمي.
^ جزيئات التصاق الخلايا العصبية (N-CAM) تنتمي إلى عائلة الغلوبولين المناعي ، فهي تشكل روابط بين الخلايا العصبية. تشارك بعض N-CAMs في توصيل المشابك وكذلك التصاق الخلية. الجهاز المناعي.
سيليكتينزأيضًا ، تشارك البروتينات المتكاملة لغشاء البلازما في التصاق الخلايا البطانية ، في ارتباط الصفائح الدموية ، الكريات البيض.
إنتغرينزهي مقاييس غير متجانسة ، ذات سلاسل و. تربط الإنتغرينات الخلايا بشكل أساسي بركائز خارج الخلية ، لكن يمكنها أيضًا المشاركة في التصاق الخلية ببعضها البعض.
التعرف على البروتينات الأجنبية
كما ذكرنا سابقًا ، يتطور تفاعل معقد معقد على الجزيئات الكبيرة الأجنبية (المستضدات) التي دخلت الجسم - استجابة مناعية. يكمن جوهرها في حقيقة أن بعض الخلايا الليمفاوية تنتج بروتينات خاصة - أجسام مضادة ترتبط على وجه التحديد بالمستضدات. على سبيل المثال ، تتعرف البلاعم على مجمعات الأجسام المضادة مع مستقبلات سطحها وتمتصها (على سبيل المثال ، امتصاص البكتيريا أثناء البلعمة).
بالإضافة إلى ذلك ، يوجد في جسم جميع الفقاريات نظام استقبال للخلايا الأجنبية أو الخلايا الخاصة بها ، ولكن مع تغيير بروتينات غشاء البلازما ، على سبيل المثال ، عندما اصابات فيروسيةأو بطفرات ، غالبًا ما ترتبط بتنكس الخلايا الورمي.
توجد البروتينات على سطح جميع خلايا الفقاريات ، ما يسمى. مجمع رئيسية في أنسجة الجسم(معقد التوافق النسيجي الرئيسي - MHC). هذه هي بروتينات متكاملة ، بروتينات سكرية ، مغاير. من المهم جدًا أن نتذكر أن كل فرد لديه مجموعة مختلفة من بروتينات معقد التوافق النسيجي الكبير. هذا يرجع إلى حقيقة أنها متعددة الأشكال للغاية ، لأن كل فرد لديه عدد كبير من الأشكال المتناوبة من نفس الجين (أكثر من 100) ، بالإضافة إلى ذلك ، هناك 7-8 مواضع ترميز جزيئات معقد التوافق النسيجي الكبير. ينتج عن هذا كل خلية كائن معين، وجود مجموعة من بروتينات معقد التوافق النسيجي الكبير ، سيكون مختلفًا عن خلايا فرد من نفس النوع. شكل خاصتتعرف الخلايا الليمفاوية ، الخلايا الليمفاوية التائية ، على معقد التوافق النسيجي الكبير في أجسامهم ، لكن أدنى تغييرفي بنية معقد التوافق النسيجي الكبير (على سبيل المثال ، الارتباط بفيروس ، أو نتيجة طفرة في الخلايا الفردية) ، يؤدي إلى حقيقة أن الخلايا الليمفاوية التائية تتعرف على هذه الخلايا المتغيرة وتدمرها ، ولكن ليس عن طريق البلعمة. تفرز بروتينات perforin محددة من فجوات إفرازية ، والتي يتم تضمينها في الغشاء السيتوبلازمي للخلية المعدلة ، وتشكل قنوات الغشاء فيها ، مما يجعل غشاء البلازما منفذاً ، مما يؤدي إلى موت الخلية المعدلة (الشكل 143 ، 144).
اتصالات خاصة بين الخلايا
بالإضافة إلى هذه الروابط اللاصقة البسيطة نسبيًا (ولكن المحددة) (الشكل 145) ، هناك عدد من الهياكل أو جهات الاتصال أو الوصلات الخاصة بين الخلايا التي تؤدي وظائف معينة. هذه هي توصيلات القفل والتثبيت والاتصال (الشكل 146).
قفلأو اتصال محكمسمة من سمات ظهارة أحادية الطبقة. هذه هي المنطقة التي تكون فيها الطبقات الخارجية لأغشية البلازما قريبة قدر الإمكان. غالبًا ما يُرى الغشاء المكون من ثلاث طبقات في هذا التلامس: يبدو أن الطبقتين الأسموفيليتين الخارجيتين لكلا الغشاءين تندمجان في طبقة واحدة مشتركة بسماكة 2-3 نانومتر. لا يحدث اندماج الأغشية على كامل منطقة التلامس المحكم ، ولكنه عبارة عن سلسلة من نقاط التقاء الأغشية (الشكل 147 أ ، 148).
في الاستعدادات المستوية لكسور غشاء البلازما في منطقة التلامس الضيق ، باستخدام طريقة التجميد والتقطيع ، وجد أن نقاط التلامس للأغشية هي صفوف من الكريات. هذه هي بروتينات الإكلودين والكلودين ، وهي بروتينات خاصة متكاملة من غشاء البلازما ، مبنية في صفوف. يمكن أن تتقاطع هذه الصفوف من الكريات أو الشرائط بطريقة تشكل ، كما كانت ، شبكة أو شبكة على سطح الانقسام. هذا الهيكل نموذجي جدًا للظهارة ، وخاصة الغدد والأمعاء. في الحالة الأخيرة ، يشكل التلامس المحكم منطقة اندماج مستمرة لأغشية البلازما ، تحيط بالخلية في الجزء القمي (العلوي ، بالنظر إلى تجويف الأمعاء) (الشكل 148). وبالتالي ، فإن كل خلية في الطبقة ، كما كانت ، محاطة بشريط من هذا التلامس. يمكن أيضًا رؤية هذه الهياكل مع وجود بقع خاصة في المجهر الضوئي. لقد تلقوا الاسم من علماء التشكل لوحات نهاية. اتضح أنه في هذه الحالة ، فإن دور الاتصال المحكم الإغلاق ليس فقط في الاتصال الميكانيكي للخلايا مع بعضها البعض. منطقة التلامس هذه ضعيفة النفاذية للجزيئات والأيونات ، وبالتالي فهي تغلق التجاويف بين الخلايا وتعزلها (ومعها البيئة الداخلية للجسم) عن البيئة الخارجية (في هذه الحالة ، تجويف الأمعاء).
يمكن إثبات ذلك باستخدام متناقضات كثيفة الإلكترون مثل محلول اللانثانم هيدروكسيد. إذا كان تجويف الأمعاء أو قناة بعض الغدد ممتلئًا بمحلول من هيدروكسيد اللانثانم ، ثم في الأقسام الموجودة تحت المجهر الإلكتروني ، تكون المناطق التي توجد بها هذه المادة ذات كثافة إلكترون عالية وستكون مظلمة. اتضح أنه لا منطقة الاتصال الضيق ولا المساحات بين الخلايا تحتها مظلمة. في حالة تلف الوصلات الضيقة (عن طريق المعالجة الأنزيمية الخفيفة أو إزالة أيونات الكالسيوم) ، فإن اللانثانوم يخترق أيضًا المناطق بين الخلايا. وبالمثل ، فقد ثبت أن الوصلات الضيقة غير منفذة للهيموجلوبين والفيريتين في أنابيب الكلى.
وبالتالي ، فإن الوصلات الضيقة هي حواجز ليس فقط للجزيئات الكبيرة ، ولكن أيضًا غير منفذة للسوائل والأيونات.
يحدث التلامس المغلق أو الضيق بين جميع أنواع الظهارة أحادية الطبقة (البطانة ، الظهارة المتوسطة ، البطانة البطانية العصبية).
حصرهأو وصلات متشابكةأو جهات الاتصال تسمى كذلك لأنها لا تربط أغشية البلازما للخلايا المجاورة فحسب ، بل ترتبط أيضًا بالعناصر الليفية للهيكل الخلوي (الشكل 149). يتميز هذا النوع من المركبات بوجود نوعين من البروتينات. أحدها هو بروتينات (رابط) عبر الغشاء ، والتي تشارك إما في الاتصال الفعلي بين الخلايا أو في اتصال البلازما مع مكونات المصفوفة خارج الخلية (الغشاء القاعدي للظهارة ، البروتينات الهيكلية خارج الخلية للنسيج الضام).
والثاني هو البروتينات داخل الخلايا التي تربط أو تثبت الألياف السيتوبلازمية للهيكل الخلوي خلف عناصر الغشاء لمثل هذا الاتصال.
تتضمن وصلات التثبيت تقاطعات نقاط التثبيت بين الخلايا ، أو نطاقات التثبيت ، أو الوصلات البؤرية ، أو لوحات التثبيت ، وكلها ترتبط داخل الخلايا بالخيوط الدقيقة للأكتين.
مجموعة أخرى من تقاطعات الربط بين الخلايا ، الديسموسومات والهيميديسموسومات ، ترتبط بعناصر أخرى من الهيكل الخلوي ، وهي خيوط وسيطة.
تم العثور على تقاطعات دقيقة بين الخلايا في العديد من الأنسجة غير الظهارية ، ولكن تم وصف الهيكل بشكل أكثر وضوحًا. شرائط خاصة (لاصقة)في ظهارة أحادية الطبقة (الشكل 150). يحيط هذا الهيكل محيط الخلية الظهارية بالكامل ، على غرار ما يحدث في حالة التقاطع الضيق. في أغلب الأحيان ، يقع مثل هذا الحزام أو الشريط أسفل وصلة ضيقة (انظر الشكل 146). في هذا المكان ، لا يتم تجميع أغشية البلازما معًا ، بل يتم تحريكها إلى حد ما على مسافة 25-30 نانومتر ، وتظهر منطقة ذات كثافة متزايدة بينهما. هذا ليس أكثر من مواقع تفاعل البروتينات السكرية عبر الغشاء ، والتي تلتصق على وجه التحديد ببعضها البعض وتوفر اتصالًا ميكانيكيًا بين أغشية خليتين متجاورتين. تنتمي بروتينات الرابط هذه إلى E-cadherins ، وهي بروتينات توفر تمييزًا محددًا للأغشية المتجانسة بواسطة الخلايا. يؤدي تدمير هذه الطبقة من البروتينات السكرية إلى عزل الخلايا الفردية وتدمير الطبقة الظهارية. على الجانب السيتوبلازمي بالقرب من الغشاء ، يُلاحظ تراكم بعض المواد الكثيفة ، والتي تلتصق بطبقة من الخيوط الرقيقة (6-7 نانومتر) الممتدة على طول غشاء البلازما في شكل حزمة تمتد على طول محيط الخلية. الخيوط الرقيقة عبارة عن ألياف أكتينية ؛ ترتبط بغشاء البلازما عبر بروتين الكاتينين ، الذي يشكل طبقة كثيفة بالقرب من الغشاء.
تكمن الأهمية الوظيفية لوصلة الشريط هذه فقط في الالتصاق الميكانيكي للخلايا ببعضها البعض: مع تقلص خيوط الأكتين في الشريط ، يمكن أن يتغير شكل الخلية. يُعتقد أن الانكماش التعاوني لألياف الأكتين في جميع خلايا الطبقة الظهارية يمكن أن يسبب تغييرًا في هندستها ، على سبيل المثال ، الطي في أنبوب ، على غرار ما يحدث أثناء تكوين الأنبوب العصبي في أجنة الفقاريات.
^ الاتصالات البؤرية أو لويحات التصاقتحدث في العديد من الخلايا وتتم دراستها جيدًا بشكل خاص في الخلايا الليفية. هم مبنيون على أساس خطة عامةبشرائط لاصقة ، ولكن يتم التعبير عنها كمناطق صغيرة - لويحات على البلازما. في هذه الحالة ، ترتبط بروتينات الإنتجرين الرابط عبر الغشاء على وجه التحديد ببروتينات المصفوفة خارج الخلية (على سبيل المثال ، فيبرونيكتين) (الشكل 151). من جانب السيتوبلازم ، ترتبط هذه البروتينات السكرية نفسها ببروتينات الغشاء ، والتي تشمل أيضًا الفينكولين ، والذي يرتبط بدوره بحزمة من خيوط الأكتين. تكمن الأهمية الوظيفية للاتصالات البؤرية في تثبيت الخلية على الهياكل خارج الخلية وفي إنشاء آلية تسمح للخلايا بالتحرك.
اجسام رابطة، الهياكل على شكل لويحات أو أزرار تربط الخلايا ببعضها البعض (الشكل 152 ، 153 أ). في الفضاء بين الخلايا ، تظهر هنا أيضًا طبقة كثيفة ، ممثلة بتفاعل كادرينات غشائية متكاملة - ديسموجلينز ، التي تربط الخلايا ببعضها البعض. على الجانب السيتوبلازمي ، توجد طبقة من بروتين ديزموبلاكين مجاورة لبلازما الدم ، والتي ترتبط بها الخيوط الوسيطة للهيكل الخلوي. تم العثور على Desmosomes في أغلب الأحيان في الظهارة ، وفي هذه الحالة تحتوي الخيوط الوسيطة على الكيراتين. في خلايا عضلة القلب ، تحتوي خلايا عضلة القلب على ليفية desmin كجزء من الديسموسومات. في البطانة الوعائية ، تحتوي الديسموسومات على خيوط فيمينتين وسيطة.
Hemidesmosomes- من حيث المبدأ ، فهي متشابهة في التركيب مع الديسموسوم ، لكنها عبارة عن اتصال بين الخلايا والتركيبات بين الخلايا. لذلك في الظهارة ، تتفاعل البروتينات السكرية الوصلة (الإنتغرينات) للديسموسومات مع بروتينات ما يسمى. الغشاء القاعدي ، والذي يشمل الكولاجين ، اللامينين ، البروتيوغليكان ، إلخ.
يعتبر الدور الوظيفي للديسموسومات والهيميديسموسومات ميكانيكيًا بحتًا - فهي تلتصق بقوة بالخلايا مع بعضها البعض وبالمصفوفة خارج الخلية الأساسية ، مما يسمح للطبقات الظهارية بمقاومة الأحمال الميكانيكية الثقيلة. وبالمثل ، فإن الديسموسومات تربط خلايا عضلة القلب ببعضها البعض بإحكام ، مما يسمح لها بأداء حمولة ميكانيكية ضخمة مع البقاء مقيدة في هيكل مقلص واحد.
على عكس التلامس الضيق ، فإن جميع أنواع جهات الاتصال الرابطة قابلة للنفاذ إلى المحاليل المائية ولا تلعب أي دور في الحد من الانتشار.
^ اتصالات الفجوة تعتبر اتصالات اتصالات الخلايا ؛ هذه هي الهياكل التي تشارك في الإرسال المباشر مواد كيميائيةمن خلية إلى أخرى ، والتي يمكن أن تلعب دورًا فسيولوجيًا رئيسيًا ليس فقط في عمل الخلايا المتخصصة ، ولكنها توفر أيضًا تفاعلات بين الخلايا أثناء تطور الكائن الحي ، أثناء تمايز خلاياها. من سمات هذا النوع من جهات الاتصال تقارب أغشية البلازما لخليتين متجاورتين على مسافة 2-3 نانومتر (الشكل 147 ب ، 153 ب). هذا هو الظرف لفترة طويلةلا يسمح بالتمييز على أقسام سامسونج هذا النوعالاتصال من جهة اتصال عزل (إغلاق) محكم. عند استخدام هيدروكسيد اللانثانم ، لوحظ أن بعض جهات الاتصال الضيقة تتسرب من مانع التسرب. في هذه الحالة ، ملأ اللانثانم فجوة رقيقة بعرض حوالي 3 نانومتر بين أغشية البلازما المجاورة للخلايا المجاورة. كان هذا هو أصل مصطلح - فجوة الاتصال. تم تحقيق مزيد من التقدم في فك رموز هيكلها باستخدام طريقة التقطيع بالتجميد. اتضح أن مناطق تقاطع الفجوة (من 0.5 إلى 5 ميكرومتر في الحجم) على شقوق الأغشية منقطة بجسيمات قطرها 7-8 نانومتر سداسية الشكل بفترة 8-10 نانومتر ، ولها قناة بعرض حوالي 2 نانومتر في مركز. تسمى هذه الجسيمات كونيكسونس(الشكل 154). في مناطق تماس الفجوة ، يمكن أن يكون هناك من 10-20 إلى عدة آلاف من الروابط ، حسب الميزات الوظيفيةالخلايا. تم عزل Connexons مبدئيًا ؛ وهي تتكون من ست وحدات فرعية من connectin ، وهو بروتين بوزن جزيئي يبلغ حوالي 30 ألفًا. وتتحد مع بعضها البعض ، وتشكل الموصلات تكتلًا أسطوانيًا - وكونيكسون ، يوجد في وسطه قناة. يتم تضمين الروابط الفردية في غشاء البلازما بطريقة تخترقها. يتم معارضة أحد الروابط الموجودة على غشاء البلازما للخلية بدقة من خلال كونيكسون على غشاء البلازما للخلية المجاورة ، بحيث تشكل قنوات هذين المكونين وحدة واحدة. تلعب Connexons دور القنوات المباشرة بين الخلايا والتي من خلالها يمكن للأيونات والمواد ذات الوزن الجزيئي المنخفض أن تنتشر من خلية إلى أخرى. وجد أنه يمكن للكونيكسونات أن تغلق ، وتغير قطر القناة الداخلية ، وبالتالي تشارك في تنظيم نقل الجزيئات بين الخلايا.
تم فهم الأهمية الوظيفية لتقاطعات الفجوة في دراسة الخلايا العملاقة الغدد اللعابيةديبتيرا. نظرًا لحجمها ، يمكن إدخال الأقطاب الكهربائية الدقيقة بسهولة في هذه الخلايا من أجل دراسة التوصيل الكهربائي لأغشيتها. اتضح أنه إذا تم إدخال أقطاب كهربائية في خليتين متجاورتين ، فإن أغشية البلازما الخاصة بها تظهر مقاومة كهربائية منخفضة ، ويتدفق التيار بين الخلايا. علاوة على ذلك ، اتضح أنه عندما يتم حقن صبغة الفلورسنت في خلية واحدة ، يتم اكتشاف الملصق بسرعة في الخلايا المجاورة. باستخدام الفلوروكرومات المختلفة ، وجد في خلايا زراعة أنسجة الثدييات أن المواد التي لا يزيد وزنها الجزيئي عن 1-1.5 ألف وحجم لا يزيد عن 1.5 نانومتر يمكن نقلها من خلال تقاطعات الفجوة (في الحشرات ، المواد ذات الوزن الجزيئي يصل وزنها إلى 2000). من بين هذه المواد الأيونات المختلفة والأحماض الأمينية والنيوكليوتيدات والسكريات والفيتامينات والمنشطات والهرمونات و cAMP. لا يمكن للبروتينات ولا الأحماض النووية أن تمر عبر تقاطعات الفجوة.
يتم استخدام هذه القدرة على تقاطعات الفجوة لتكون بمثابة مكان لنقل المركبات ذات الوزن الجزيئي المنخفض أنظمة الخلايا، حيث يلزم النقل السريع لنبضة كهربائية (موجة إثارة) من خلية إلى أخرى دون مشاركة وسيط عصبي. وهكذا ، فإن جميع الخلايا العضلية لعضلة القلب متصلة ببعضها البعض باستخدام وصلات فجوة (بالإضافة إلى ذلك ، فإن الخلايا هناك متصلة أيضًا بوصلات لاصقة) (الشكل 147 ب). هذا يخلق شرطًا للتقليل المتزامن لعدد ضخم من الخلايا. مع نمو ثقافة خلايا عضلة القلب الجنينية (خلايا عضلة القلب) ، تبدأ بعض الخلايا في الطبقة بالتقلص تلقائيًا بشكل مستقل عن بعضها البعض مع تردد مختلف، وفقط بعد تشكيل فجوات الوصلات بينهما تبدأ في الضرب بشكل متزامن كطبقة واحدة من الخلايا المتقلصة. بنفس الطريقة ، يتم ضمان تقلص مفصل لخلايا العضلات الملساء في جدار الرحم.
يمكن أن تخدم وصلات الفجوة غرض التعاون الأيضي بين الخلايا عن طريق تبادل الجزيئات المختلفة أو الهرمونات أو cAMP أو المستقلبات. مثال على ذلك هو الزراعة المشتركة للخلايا الطافرة ثيميدين كيناز مع الخلايا الطبيعية: عندما تحدث تقاطعات فجوة بين هذه الأنواع من الخلايا ، تتلقى الخلايا الطافرة ثيميدين ثلاثي الفوسفات من الخلايا الطبيعية من خلال تقاطعات الفجوة ويمكن أن تشارك في تخليق الحمض النووي.
في أجنة الفقاريات المبكرة ، بدءًا من مرحلة الخلايا الثمانية ، ترتبط معظم الخلايا ببعضها البعض عن طريق تقاطعات الفجوة. عندما يتمايز الجنين ، تختفي فجوات الوصلات بين جميع الخلايا وتبقى فقط بين مجموعات الخلايا المتخصصة. على سبيل المثال ، أثناء تكوين الأنبوب العصبي ، يتم قطع اتصال خلايا هذا الهيكل مع بقية البشرة.
تعتمد سلامة وتشغيل تقاطعات الفجوة بشكل كبير على مستوى أيونات الكالسيوم 2+ داخل الخلية. عادة ، يكون تركيز الكالسيوم في السيتوبلازم منخفضًا جدًا. إذا تم حقن Ca 2+ في إحدى خلايا طبقة زراعة الأنسجة ، فلن تحدث زيادة في مستوى Ca 2+ في السيتوبلازم في الخلايا المجاورة ؛ الخلايا ، كما كانت ، منفصلة عن جيرانها ، تتوقف عن العمل كهرباءوالأصباغ. بعد مرور بعض الوقت ، بعد تراكم الكالسيوم الذي تم إدخاله بواسطة الميتوكوندريا ، يتم استعادة بنية ووظائف تقاطعات الفجوة. تعد هذه الخاصية مهمة للغاية للحفاظ على سلامة وتشغيل طبقة الخلايا بأكملها ، حيث لا ينتقل الضرر الذي يلحق بإحداها إلى الطبقة المجاورة من خلال تقاطعات الفجوة ، والتي تتوقف عن العمل كقنوات انتشار بين الخلايا.
^ اتصال متشابك (المشابك). هذا النوع من الاتصال هو نموذجي لـ أنسجة عصبيةويحدث بين خليتين عصبيتين وبين خلية عصبية وبعض العناصر الأخرى - مستقبل أو مستجيب (على سبيل المثال ، نهاية عصبية عضلية). نقاط الاشتباك العصبي هي مناطق تلامس بين خليتين متخصصتين في انتقال الإثارة أو التثبيط في اتجاه واحد من عنصر إلى آخر (الشكل 155). أساسًا هذا النوعالحمل الوظيفي ، يمكن أيضًا نقل النبضات بواسطة أنواع أخرى من جهات الاتصال (على سبيل المثال ، فجوة الاتصال في عضلة القلب) ، ومع ذلك ، في الاتصال المشبكي ، كفاءة عاليةفي إدراك الدافع العصبي. تتشكل المشابك العصبية على عمليات الخلايا العصبية - هذه هي الأقسام النهائية للتشعبات والمحاور. المشابك العصبية الداخلية عادة ما يكون لها امتدادات على شكل كمثرى ، لويحات في نهاية عملية الخلية العصبية. يمكن لمثل هذا الامتداد النهائي لعملية إحدى الخلايا العصبية الاتصال وتشكيل اتصال متشابك مع جسم خلية عصبية أخرى وعملياتها. تشكل العمليات المحيطية للخلايا العصبية (المحاور) اتصالات محددة مع خلايا المستجيب أو المستقبل. لذلك ، فإن المشبك هو بنية تتشكل بين مناطق خليتين (بالإضافة إلى ديسموسوم). يتم فصل أغشية هذه الخلايا عن طريق مساحة بين الخلايا - شق متشابك يبلغ عرضه حوالي 20-30 نانومتر. في كثير من الأحيان في تجويف هذا الشق ، تكون مادة ألياف دقيقة متعامدة على الأغشية مرئية. يسمى الغشاء الموجود في منطقة التلامس المشبكي لخلية واحدة قبل المشبكي ، ويسمى الآخر ، الذي يدرك النبض ، ما بعد المشبكي. في المجهر الإلكتروني ، يبدو كلا الغشاءين كثيفين وسميكين. بالقرب من الغشاء قبل المشبكي ، تم الكشف عن عدد كبير من الفجوات الصغيرة والحويصلات المشبكية المليئة بالناقلات العصبية. الحويصلات المشبكية في وقت مرور النبضة العصبية تخرج محتوياتها في الشق المشبكي. غالبًا ما يبدو الغشاء بعد المشبكي أكثر سمكًا من الأغشية العادية بسبب تراكم العديد من الألياف الرقيقة حوله من جانب السيتوبلازم.
روابط بلازمية. تم العثور على هذا النوع من الاتصال بين الخلايا في النباتات. هيولي هي قنوات أنبوبية رفيعة تربط خليتين متجاورتين. يبلغ قطر هذه القنوات عادة 20-40 نانومتر. يمر الغشاء الذي يحد من هذه القنوات مباشرة إلى أغشية البلازما للخلايا المجاورة. تمر خلويات Plasmodesmata عبر جدار الخلية الذي يفصل بين الخلايا (الشكل 156 ، 157). وهكذا ، في بعض الخلايا النباتية ، تربط plasmodesmata الهيالوبلازم للخلايا المجاورة ، لذلك لا يوجد تمييز كامل رسميًا ، أي فصل جسم خلية عن أخرى ، بل هو عبارة عن خلوي: اتحاد العديد من مناطق الخلايا بمساعدة السيتوبلازم الجسور. يمكن أن تخترق العناصر الأنبوبية الغشائية داخل plasmodesmata ، وربط صهاريج الشبكة الإندوبلازمية للخلايا المجاورة. تتشكل Plasmodesmata أثناء انقسام الخلية ، عندما يتم بناء جدار الخلية الأساسي (انظر أدناه). في الخلايا المنقسمة حديثًا ، يمكن أن يكون عدد plasmodesmata كبيرًا جدًا (يصل إلى 1000 لكل خلية) ، مع شيخوخة الخلية يتناقص عددها بسبب التمزق مع زيادة السماكة. جدار الخلية.
إن الدور الوظيفي للوسائط البلازمية كبير جدًا: بمساعدتهم ، فإن الدوران بين الخلايا للحلول المحتوية على العناصر الغذائيةوالأيونات والمركبات الأخرى. يمكن أن تتحرك قطرات الشحوم على طول plasmodesmata ، وتصاب الخلايا بالفيروسات النباتية من خلال plasmodesmata. ومع ذلك ، تظهر التجارب أن النقل المجاني عبر رابطات البيانات الوصفية يقتصر على الجسيمات التي لا تزيد كتلتها عن 800 دالتون.
> وظائف غشاء البلازما. آليات نقل المواد عبر غشاء البلازما. وظيفة المستقبل للبلازما
يؤدي غشاء البلازما عددًا من الوظائف المهمة:
1) الحاجز.تتمثل وظيفة الحاجز لغشاء البلازما في الحد من الانتشار الحر للمواد من خلية إلى أخرى ، لمنع تسرب محتويات الخلية القابلة للذوبان في الماء. ولكن نظرًا لأن الخلية يجب أن تتلقى العناصر الغذائية الضرورية ، وتحرر المنتجات النهائية لعملية التمثيل الغذائي ، وتنظم تركيزات الأيونات داخل الخلايا ، فقد تم تشكيل آليات خاصة لنقل المواد عبر غشاء الخلية.
2) النقل.وظيفة النقل هي ضمان دخول وخروج المواد المختلفة داخل وخارج الخلية. خاصية مهمةأغشية - النفاذية الاختيارية، أو نفاذية.يمر بسهولة في الماء والغازات القابلة للذوبان في الماء ويصد الجزيئات القطبية مثل الجلوكوز أو الأحماض الأمينية.
توجد عدة آليات لنقل المواد عبر الغشاء:
النقل السلبي
النقل النشط
النقل في عبوات غشائية.
النقل السلبي.انتشار -هذه هي حركة جزيئات الوسط ، مما يؤدي إلى انتقال مادة من منطقة يكون تركيزها فيها مرتفعًا إلى منطقة ذات تركيز منخفض. أثناء نقل الانتشار ، يعمل الغشاء كحاجز تناضحي. يعتمد معدل الانتشار على حجم الجزيئات وقابليتها للذوبان النسبي في الدهون. كيف أحجام أصغرالجزيئات وكلما زادت قابليتها للذوبان في الدهون (محبة للدهون) ، زادت سرعة تحركها عبر طبقة الدهون الثنائية. يمكن أن يكون الانتشار حيادي(نقل الجزيئات غير المشحونة) و وزن خفيف(بمساعدة البروتينات الحاملة الخاصة). الانتشار الميسر أسرع من الانتشار المحايد. يتمتع الماء بأقصى قوة اختراق ، حيث أن جزيئاته صغيرة وغير مشحونة. يسمى انتشار الماء عبر غشاء الخلية التنافذ.من المفترض وجود "مسام" خاصة في غشاء الخلية لاختراق الماء وبعض الأيونات. عددهم صغير ، وقطرها حوالي 0.3-0.8 نانومتر. تنتشر الجزيئات القابلة للذوبان بسهولة في طبقة ثنائية الدهون ، مثل O ، والجزيئات القطبية غير المشحونة ذات القطر الصغير (CO ، اليوريا) بسرعة أكبر عبر الغشاء.
يسمى نقل الجزيئات القطبية (السكريات والأحماض الأمينية) الذي يتم بمساعدة بروتينات النقل الغشائي الخاص نشر الميسر.توجد هذه البروتينات في جميع أنواع الأغشية البيولوجية ، وكل بروتين محدد مصمم لحمل جزيئات من فئة معينة. بروتينات النقل عبارة عن غشاء عابر ؛ تعبر سلسلة البولي ببتيد الخاصة بها طبقة ثنائية الدهون عدة مرات ، وتتشكل من خلال ممرات فيها. وهذا يضمن نقل مواد معينة عبر الغشاء دون ملامسة مباشرة له. هناك فئتان رئيسيتان من بروتينات النقل: بروتينات حاملة (ناقلات)و تشكيل القناةالبروتينات (قنوات البروتين). تحمل البروتينات الحاملة الجزيئات عبر الغشاء عن طريق تغيير تكوينها أولاً. تشكل البروتينات المكونة للقناة مسامًا مملوءة بالماء في الغشاء. عندما تكون المسام مفتوحة ، تمر عبرها جزيئات من مواد معينة (عادة أيونات غير عضوية بالحجم والشحنة المناسبين). إذا لم يكن لجزيء المادة المنقولة شحنة ، فسيتم تحديد اتجاه النقل بواسطة تدرج التركيز. إذا كان الجزيء مشحونًا ، فإن نقله ، بالإضافة إلى تدرج التركيز ، يتأثر أيضًا بالشحنة الكهربائية للغشاء (إمكانات الغشاء). الجانب الداخليعادة ما تكون شحنة البلازما سالبة الشحنة بالنسبة إلى الخارج. تسهل إمكانات الغشاء تغلغل الأيونات الموجبة الشحنة في الخلية وتمنع مرور الأيونات سالبة الشحنة.
النقل النشط.النقل النشط هو حركة المواد ضد التدرج الكهروكيميائي. يتم تنفيذه دائمًا بواسطة البروتينات الناقلة ويرتبط ارتباطًا وثيقًا بمصدر للطاقة. تحتوي البروتينات الحاملة على مواقع ارتباط بالمادة المنقولة. وكلما زاد ارتباط هذه المواقع بالمادة ، زاد معدل النقل. يسمى النقل الانتقائي لمادة واحدة uniport.يتم تنفيذ نقل العديد من المواد أنظمة النقل المشترك.إذا سارت عملية النقل في اتجاه واحد ، فهي كذلك symportإذا كان في المقابل مضاد للميناء.على سبيل المثال ، يتم نقل الجلوكوز من السائل خارج الخلية إلى الخلية بطريقة أحادية. يتم نقل الجلوكوز و Na 4 من التجويف المعوي أو أنابيب الكلى ، على التوالي ، إلى خلايا الأمعاء أو الدم بشكل متزامن ، ويتم نقل C1 ~ و HCO "مضاد للمنفذ.
مثال على البروتين الحامل الذي يستخدم الطاقة المنبعثة أثناء التحلل المائي ATP لنقل المواد نا + -ل + مضخة،وجدت في غشاء البلازما لجميع الخلايا. تعمل مضخة Na + -K على مبدأ antiport ، حيث تضخ Na "خارج الخلية و K t في الخلية مقابل التدرجات الكهروكيميائية الخاصة بهم. يخلق تدرج Na + ضغطًا تناضحيًا ويحافظ على حجم الخلية ويضمن نقل السكريات والأحماض الأمينية يتم إنفاق ثلث الطاقة على هذه المضخة الضرورية للنشاط الحيوي للخلايا. وعند دراسة آلية عمل مضخة Na + -K + ، وجد أنها إنزيم ATPase وبروتين متكامل عبر الغشاء. وجود Na + و ATP ، تحت تأثير ATPase ، يتم فصل الفوسفات الطرفي عن ATP وربطه بالبقايا حمض الأسبارتيكعلى جزيء ATPase. يتم فسفرة جزيء ATPase ، ويغير تكوينه ، ويتم إخراج Na + من الخلية. بعد إزالة Na من الخلية ، يتم دائمًا نقل K "إلى الخلية. للقيام بذلك ، يتم قطع الفوسفات المتصل سابقًا من ATPase في وجود K. ويتم إزالة الفسفرة من الإنزيم ، ويعيد تكوينه ، ويكون K 1" تضخ "في الخلية.
يتكون ATPase من وحدتين فرعيتين ، كبيرة وصغيرة. تتكون الوحدة الفرعية الكبيرة من آلاف بقايا الأحماض الأمينية التي تعبر الطبقة الثنائية عدة مرات. له نشاط تحفيزي ويمكن عكسه عن طريق الفسفرة وإزالة الفسفرة. تحتوي الوحدة الفرعية الكبيرة على الجانب السيتوبلازمي على مواقع لربط Na + و ATP وما فوق الخارج-مواقع ربط K + و ouabain. الوحدة الفرعية الصغيرة هي بروتين سكري ووظيفتها غير معروفة بعد.
مضخة Na + -K لها تأثير كهربائي. يزيل ثلاثة أيونات صوديوم موجبة الشحنة من الخلية ويدخل فيها أيونين من نوع K. ونتيجة لذلك ، يتدفق تيار عبر الغشاء ، مكونًا جهدًا كهربائيًا ذا قيمة سالبة في الجزء الداخلي للخلية نسبة إلى سطحها الخارجي . تنظم مضخة Na "-K + حجم الخلية ، وتتحكم في تركيز المواد داخل الخلية ، وتحافظ على الضغط الاسموزي ، وتشارك في تكوين إمكانات الغشاء.
النقل في عبوات غشائية. يتم نقل الجزيئات الكبيرة (البروتينات والأحماض النووية والسكريات والبروتينات الدهنية) والجزيئات الأخرى عبر الغشاء من خلال التكوين والانصهار المتسلسل للحويصلات (الحويصلات) المحاطة بالغشاء. تحدث عملية النقل الحويصلي على مرحلتين. في البداية ، يلتصق غشاء الحويصلة وغشاء البلازما معًا ثم يندمجان. بالنسبة لمسار المرحلة 2 ، من الضروري إزاحة جزيئات الماء عن طريق تفاعل طبقات ثنائية الدهون ، والتي تقترب من بعضها البعض حتى مسافة 1-5 نانومتر. يُعتقد أن هذه العملية يتم تنشيطها بواسطة خاص بروتينات الانصهار(تم عزلهم حتى الآن في الفيروسات فقط). النقل الحويصلي له ميزة مهمة- الجزيئات الكبيرة الممتصة أو المفرزة في الحويصلات عادة لا تختلط مع الجزيئات الكبيرة أو العضيات الأخرى في الخلية. يمكن أن تندمج الفقاعات مع أغشية معينة ، مما يضمن تبادل الجزيئات الكبيرة بين الفضاء خارج الخلية ومحتويات الخلية. وبالمثل ، يتم نقل الجزيئات الكبيرة من حجرة خلية إلى أخرى.
يسمى نقل الجزيئات الكبيرة والجسيمات إلى الخلية الالتقام.في هذه الحالة ، يتم تغليف المواد المنقولة بجزء من غشاء البلازما ، وتتشكل فقاعة (فجوة) تتحرك داخل الخلية. اعتمادًا على حجم الحويصلات المتكونة ، يتم تمييز نوعين من الالتقام الخلوي - كثرة الكريات والبلعمة.
كثرة الخلايايوفر امتصاص المواد السائلة والمذابة على شكل فقاعات صغيرة (د = 150 نانومتر). البلعمة -هذا هو امتصاص الجزيئات الكبيرة أو الكائنات الحية الدقيقة أو شظايا العضيات والخلايا. في هذه الحالة ، تتشكل حويصلات كبيرة أو فاجوسومات أو فجوات (d-250 نانومتر أو أكثر). في البروتوزوا ، وظيفة البلعمة هي شكل من أشكال التغذية. في الثدييات ، يتم تنفيذ وظيفة البلعمة عن طريق الضامة والعدلات ، والتي تحمي الجسم من العدوى عن طريق ابتلاع الميكروبات الغازية. تشارك البلاعم أيضًا في التخلص من الخلايا القديمة أو التالفة وشظاياها (في جسم الإنسان ، تمتص البلاعم أكثر من 100 خلية دم حمراء قديمة يوميًا). يبدأ البلعمة فقط عندما يرتبط الجسيم الممتص بسطح البلعمة وينشط خلايا المستقبل المتخصصة. يؤدي ارتباط الجسيمات بمستقبلات غشائية معينة إلى تكوين الأرجل الكاذبة ، والتي تغلف الجسيم وتندمج عند الحواف وتشكل فقاعة - بلعم.يحدث تكوين البلعمة المناسبة والبلعمة فقط إذا كان الجسيم ، أثناء عملية التغليف ، على اتصال دائم بمستقبلات البلازما ، كما لو كان "ينطلق".
ينتهي جزء كبير من المادة التي تمتصها الخلية عن طريق الالتقام الخلوي في الجسيمات الحالة. يتم تضمين الجسيمات الكبيرة في البلعمةوالتي تندمج بعد ذلك مع الجسيمات الحالة لتشكل البلعمة.يتم نقل السوائل والجزيئات الكبيرة التي يتم تناولها أثناء كثرة الخلايا في البداية إلى الإندوسومات ، والتي تندمج أيضًا مع الجسيمات الحالة لتشكيل الجسيمات الداخلية. إنزيمات التحلل المائي المختلفة الموجودة في الجسيمات الحالة تدمر بسرعة الجزيئات الكبيرة. يتم نقل منتجات التحلل المائي (الأحماض الأمينية والسكريات والنيوكليوتيدات) من الجسيمات الحالة إلى العصارة الخلوية ، حيث تستخدمها الخلية. يتم إرجاع معظم مكونات غشاء الحويصلات الداخلية من البلعمة والداخلية عن طريق الإفراز الخلوي إلى غشاء البلازما وإعادة استخدامها هناك. رئيسي الأهمية البيولوجيةالالتقام الخلوي هو اكتساب اللبنات الأساسية من خلال الهضم داخل الخلايا للجزيئات الكبيرة في الجسيمات الحالة.
امتصاص المواد في الخلايا حقيقية النواةيبدأ في مناطق متخصصة من غشاء البلازما ، ما يسمى حفر يحدها.في الصور المجهرية الإلكترونية ، تبدو الحفر كأنها غشاءات لغشاء البلازما ، حيث يكون الجانب السيتوبلازمي منها مغطى بطبقة ليفية. الطبقة ، كما كانت ، تحد حفرًا صغيرة من غشاء البلازما. تحتل الحفر حوالي 2٪ من السطح الكلي لغشاء الخلية حقيقية النواة. في غضون دقيقة ، تنمو الحفر ، وتنتشر بشكل أعمق وأعمق ، ويتم سحبها إلى الخلية ، ثم تضيق عند القاعدة ، وتنقسم ، وتشكل حويصلات ذات حدود. وقد ثبت أن ما يقرب من ربع الغشاء على شكل حويصلات متجاورة ينفصل عن غشاء البلازما للأرومات الليفية في غضون دقيقة واحدة. تفقد الحويصلات بسرعة حدودها وتكتسب القدرة على الاندماج مع الجسيم الحال.
قد يكون الالتقام غير محدد(التأسيسية) و محدد(مستقبل). في الالتقام غير النوعيتلتقط الخلية وتمتص المواد الغريبة تمامًا عنها ، على سبيل المثال ، جزيئات السخام والأصباغ. في البداية ، تترسب الجزيئات على الكُلَى السُكري في البلازما. مجموعات البروتين المشحونة إيجابياً يتم ترسيبها بشكل جيد (كثف) ، حيث أن glycocalyx يحمل شحنة سالبة. ثم يتغير شكل غشاء الخلية. يمكن أن تغرق ، وتشكل انغابات (انغالات) ، أو ، على العكس من ذلك ، تشكل نواتج تبدو وكأنها تنثني ، وتفصل أحجامًا صغيرة من الوسط السائل. يعتبر تكوين الانغابات أكثر شيوعًا بالنسبة لخلايا الظهارة المعوية والأميبات والنواتج - للخلايا البلعمية والخلايا الليفية. يمكن منع هذه العمليات عن طريق مثبطات الجهاز التنفسي. يمكن أن تندمج الحويصلات الناتجة - الإندوسومات الأولية - مع بعضها البعض ، ويزداد حجمها. بعد ذلك ، تتحد مع الجسيمات الحالة ، وتتحول إلى جسيم داخلي - فجوة في الجهاز الهضمي. شدة كثرة الخلايا غير النوعية في المرحلة السائلة عالية جدًا. تتكون البلاعم حتى 125 ، والخلايا الظهارية للأمعاء الدقيقة تصل إلى ألف صنوبر في الدقيقة. تؤدي وفرة الصنوبر إلى حقيقة أن البلازما تنفق بسرعة على تكوين العديد من الفجوات الصغيرة. تتم استعادة الغشاء بسرعة كبيرة أثناء إعادة التدوير أثناء الإفراز الخلوي بسبب عودة الفجوات وإدماجها في البلازما. في البلاعم ، يتم استبدال غشاء البلازما بالكامل في غضون 30 دقيقة ، وفي الخلايا الليفية في غضون ساعتين.
أكثر على نحو فعالالامتصاص من السائل خارج الخلية لجزيئات معينة هو الالتقام النوعي(بوساطة المستقبلات). في هذه الحالة ، ترتبط الجزيئات الكبيرة بالمستقبلات التكميلية على سطح الخلية ، وتتراكم في الحفرة المتاخمة ، ومن ثم ، لتشكيل جسيم داخلي ، يتم غمرها في العصارة الخلوية. يضمن الالتقام الخلوي للمستقبلات تراكم جزيئات معينة في مستقبلها. تسمى الجزيئات التي ترتبط بمستقبل على سطح البلازما يجند.بمساعدة الالتقام الخلوي للمستقبلات في العديد من الخلايا الحيوانية ، يتم امتصاص الكوليسترول من البيئة خارج الخلية.
يشارك غشاء البلازما في إزالة المواد من الخلية (خروج الخلايا). في هذه الحالة ، تقترب الفجوات من غشاء البلازما. عند نقاط التلامس ، تندمج البلازما وغشاء الفجوة وتدخل محتويات الفجوة إلى البيئة. في بعض الأوليات ، يتم تحديد المواقع الموجودة على غشاء الخلية لإخراج الخلايا مسبقًا. لذلك ، في غشاء البلازما لبعض ciliates هناك مناطق معينة مع الترتيب الصحيح للكريات الكبيرة من البروتينات المتكاملة. تحتوي الأكياس المخاطية والكيسات ثلاثية الشعيرات الجاهزة تمامًا للإفراز على هالة من كريات البروتين المتكاملة في الجزء العلوي من غشاء البلازما. هذه المقاطع من غشاء الأكياس المخاطية والكيسات الثلاثية على اتصال مع سطح الخلية. لوحظ خروج خلوي غريب في العدلات. إنهم قادرون ، في ظل ظروف معينة ، على إطلاق الجسيمات الحالة في البيئة. في بعض الحالات ، تتشكل نواتج صغيرة من غشاء البلازما المحتوي على الجسيمات الحالة ، والتي تنفصل بعد ذلك وتنتقل إلى البيئة. في حالات أخرى ، هناك غزو للبلازما في أعماق الخلية والتقاطها للجسيمات الحالة بعيدًا عن سطح الخلية.
تتم عمليات الالتقام الخلوي والإفراز الخلوي بمشاركة نظام المكونات الليفية من السيتوبلازم المرتبط بالبلازما.
وظيفة المستقبل للبلازما.هذه واحدة من الوظائف الرئيسية والعالمية لجميع الخلايا ، وهي وظيفة المستقبل للبلازما. يحدد تفاعل الخلايا مع بعضها البعض ومع البيئة الخارجية.
يمكن تمثيل المجموعة الكاملة من التفاعلات المعلوماتية بين الخلايا بشكل تخطيطي كسلسلة من التفاعلات المتتالية - مستقبل الإشارة - المرسل - استجابة المرسل (مفهوم استجابة الإشارة).يتم نقل المعلومات من خلية إلى أخرى عن طريق إشارات الجزيئات التي يتم إنتاجها في بعض الخلايا وتؤثر بشكل خاص على الخلايا الأخرى الحساسة للإشارة (الخلايا المستهدفة). جزيء الإشارة - وسيط أساسييرتبط بالمستقبلات الموجودة في الخلايا المستهدفة التي تستجيب فقط لإشارات معينة. جزيئات الإشارة - يجند -تقترب من مستقبلها مثل مفتاح القفل. الروابط للمستقبلات الغشائية (مستقبلات البلازما) هي جزيئات محبة للماء ، وهرمونات الببتيد ، والناقلات العصبية ، والسيتوكينات ، والأجسام المضادة ، والمستقبلات النووية - الجزيئات القابلة للذوبان في الدهون ، والستيرويد وهرمونات الغدة الدرقية ، وفيتامين د. سطح الخلية - السكريات والبروتينات السكرية. يُعتقد أن المناطق الحساسة للمواد الفردية مبعثرة على سطح الخلية أو مجمعة في مناطق صغيرة. لذلك ، يوجد على سطح الخلايا بدائية النواة والخلايا الحيوانية عدد محدود من الأماكن التي يمكن أن ترتبط بها الجزيئات الفيروسية. تتعرف بروتينات الغشاء (الناقلات والقنوات) على مواد معينة وتتفاعل معها وتحملها. تشارك مستقبلات الخلية في إرسال الإشارات من سطح الخلية إليها. يؤدي تنوع وخصوصية مجموعات المستقبلات على سطح الخلية إلى إنشاء نظام معقد للغاية من العلامات التي تجعل من الممكن تمييز خلايا المرء عن تلك الموجودة في الخلايا الأخرى. تتفاعل الخلايا المتشابهة مع بعضها البعض ، ويمكن أن تلتصق أسطحها ببعضها البعض (الاقتران في البروتوزوا ، وتشكيل الأنسجة في الخلايا متعددة الخلايا). يتم تدمير أو رفض الخلايا التي لا ترى العلامات ، وكذلك تلك التي تختلف في مجموعة العلامات المحددة. عندما يتم تكوين مركب مستقبلات ليجند ، يتم تنشيط بروتينات الغشاء: بروتين محول ، بروتين مضخم. نتيجة لذلك ، يغير المستقبل شكله ويتفاعل مع سلف المرسل الثاني الموجود في الخلية - رسول.يمكن أن يكون الرسل الكالسيوم المتأين، فسفوليباز ج ، أدينيلات سيكلاز ، غوانيلات سيكلاز. تحت تأثير الرسول ، يتم تنشيط الإنزيمات المشاركة في التوليف أحادي الفوسفات الدوري - AMPأو HMF.يغير الأخير نشاط نوعين من إنزيمات البروتين كينيز في سيتوبلازم الخلية ، مما يؤدي إلى فسفرة العديد من البروتينات داخل الخلايا.
التكوين الأكثر شيوعًا لـ cAMP ، والذي تحت تأثيره يزداد إفراز عدد من الهرمونات - هرمون الغدة الدرقية ، الكورتيزون ، البروجسترون ، وانهيار الجليكوجين في الكبد والعضلات ، وتواتر وقوة تقلصات القلب ، وهدم العظام ، والعكس يزيد امتصاص الماء في نبيبات النيفرون.
إن نشاط نظام adenylate cyclase مرتفع للغاية - يؤدي تخليق cAMP إلى زيادة الإشارة بمقدار عشرة آلاف.
تحت تأثير cGMP ، يزيد إفراز البنكرياس للأنسولين ، والهستامين بواسطة الخلايا البدينة ، ويزيد السيروتونين عن طريق الصفائح الدموية ، ويتم تقليل أنسجة العضلات الملساء.
في كثير من الحالات ، أثناء تكوين مجمع المستقبلات ، يحدث تغيير في إمكانات الغشاء ، مما يؤدي بدوره إلى تغيير في نفاذية غشاء البلازما و عمليات التمثيل الغذائيفي قفص.
توجد على غشاء البلازما مستقبلات محددة تستجيب للعوامل الفيزيائية. لذلك ، في البكتيريا الضوئية ، توجد الكلوروفيل على سطح الخلية التي تتفاعل مع الضوء. يحتوي غشاء البلازما في الحيوانات الحساسة للضوء النظام بأكملهبروتينات مستقبلات الضباب - رودوبسين ، والتي يتم من خلالها تحويل المنبهات الضوئية إلى إشارة كيميائية ، ثم نبضة كهربائية.
غشاء الخلية (البلازما) عبارة عن حاجز شبه منفذ يفصل سيتوبلازم الخلايا عن البيئة. في الوقت الحاضر ، نموذج الفسيفساء السائل للغشاء ، الذي اقترحه في السبعينيات جي سينغر وجي نيكلسون ، يعتبر أساسًا في علم الأحياء.
يعتمد هذا النموذج على عدة مبادئ أساسية:
1. يتكون الغشاء من طبقة مزدوجة من جزيئات الدهون. توجد الأجزاء القطبية المحبة للماء من الجزيئات (الرؤوس) خارج الغشاء ، والأجزاء الكارهة للماء ، والأجزاء غير القطبية (الذيل) - بالداخل.
2. بروتينات الغشاء مدمجة بشكل فسيفسائي في الطبقة الدهنية الثنائية. يمر البعض منهم عبر الغشاء (يطلق عليهم اسم متكامل) ، والبعض الآخر يقع على السطح الخارجي أو الداخلي للغشاء (يطلق عليهم اسم المحيطي).
3. تحتوي القاعدة الدهنية للغشاء على خصائص سائل (مثل زيت سائل) ويمكن أن تغير كثافته. تعتمد لزوجة الغشاء على تكوين الدهون ودرجة الحرارة. في هذا الصدد ، يمكن لبروتينات الغشاء والدهون نفسها أن تتحرك بحرية على طول الغشاء وداخله. هذه خاصية مهمة جدًا للغشاء ، والتي تضمن أن الغشاء يؤدي العديد من الوظائف (انظر أدناه)
4. معظم أغشية الخلايا عضيات الغشاءتشبه في الأساس غشاء البلازما.
5. على الرغم من التركيب المشترك لأغشية جميع الخلايا ، يختلف تكوين البروتينات والدهون في كل نوع من الخلايا وداخل الخلية. يختلف أيضًا تكوين طبقات الدهون الخارجية والداخلية.
وظائف غشاء البلازما
يؤدي الغشاء مجموعة كاملة من الوظائف في الخلية المرتبطة بالحفاظ على سلامة الخلية ، مما يضمن تبادل المواد والطاقة بين الخلية و بيئةوالتفاعل مع الخلايا الأخرى.
وظيفة النقل
يتميز غشاء البلازما بخصائص حاجز شبه منفذ يسمح بمرور مواد وجزيئات معينة فقط ولا يسمح بمرورها. هناك عدة أنواع من النقل عبر الغشاء: النقل السلبي ، النقل النشط ، النقل الأيوني ، النقل في عبوات غشائية (داخلي وإخراج الخلايا).
النقل السلبيهو نقل المواد عبر الغشاء من منطقة عالية التركيز إلى منطقة تركيز منخفض. يتم تنفيذه في شكلين: في شكل انتشار بسيط وفي شكل انتشار "سهل". كلتا العمليتين لا تتطلبان طاقة ، فهي بطيئة نسبيًا وتتوقف عندما يتساوى تركيز المواد على جانبي الغشاء. يعتمد معدل الانتشار وإمكانية نقل المواد عبر الغشاء (بالإضافة إلى التركيز) على عدد من العوامل: درجة الحرارة ، والحجم الجزيئي ، والقدرة على الذوبان في الدهون. تمر المواد القابلة للذوبان في الدهون عبر طبقات الدهون بسهولة ، والمواد القابلة للذوبان في الماء بصعوبة. هناك "مسام" دهنية وبروتينية خاصة في الغشاء يمر من خلالها الانتشار. الانتشار البسيط هو عملية بطيئة نسبيًا وقد قامت الطبيعة بتكييف بروتينات حاملة غشائية لتسريع النقل. تتحد مع المادة المنقولة وتنقلها من جانب واحد من الغشاء إلى الجانب الآخر. لكل مجموعة من المواد ، يجب أن يكون للغشاء ناقلات خاصة به. وتسمى هذه العملية "الانتشار الميسر" ، وهي تتم أسرع بعشر مرات من الانتشار البسيط.
النقل الأيوني هو نوع من النقل السلبي للأيونات المشحونة ، لكن له خصائصه الخاصة. عادةً ما تحمل البروتينات الموجودة في سيتوبلازم الخلية شحنة سالبة على سطحها ، مما يؤدي إلى تكوين خلفية كهربائية معينة في الخلية. إذا تم توجيه تدفقات الأيونات الموجبة (الكاتيونات) أو السالبة (الأنيونات) إلى الخلية ، إذن الأيونات الموجبةسوف يدخل المزيد ، نظرًا لأن جزءًا من الكاتيونات سيكون مرتبطًا بالبروتينات السيتوبلازمية ، يتم تحييده ويخلق فرقًا إضافيًا في تركيزات أيونات مختلفة الشحنة داخل وخارج الخلية. لنقل الأيونات في الغشاء ، هناك إما مسام أو ناقلات أيونية خاصة.
النقل النشط- معظم شكل مهمالنقل للخلية. يؤدي الانتشار ، إذا استمر لفترة كافية ، إلى حقيقة أن التوازن قائم على جانبي الغشاء ، وهو أمر غير مقبول للخلية - وهذا موت لها. تكوين السيتوبلازم و السائل الخلاليتختلف إلى حد كبير. لذلك ، يوجد نظام "نقل نشط" في الخلية. في هذه الحالة ، يتم نقل الجزيئات مقابل تدرج التركيز (من منطقة التركيز المنخفض إلى منطقة عالية). لهذا ، توجد مجمعات غشاء بروتينية خاصة (قنوات أيونية وجزيئية) تعمل مع استهلاك الطاقة. يذهب ما يصل إلى 40٪ من إجمالي الطاقة التي تنتجها الخلية إلى تكاليف النقل!
النقل في عبوات غشائية (داخلي وإخراج الخلايا). في كثير من الحالات ، يجب أن تنتقل الخلية عبر الغشاء ليس الجزيئات أو الأيونات الفردية ، ولكن المجمعات الجزيئية الكاملة وحتى الجسيمات. لا تمر من خلال مسام الغشاء. في هذا الصدد ، هناك نوع خاص من النقل في الخلية - النقل في عبوات الغشاء. في بعض الحالات ، يحدث نتوء في غشاء البلازما الخارجي ، ويغطي جسيمًا غريبًا (على سبيل المثال: جسيم غذائي أو بكتيريا) ، ويغلقه في حلقة غشاء ويغمر بعمق في سيتوبلازم الخلية. تم اكتشاف هذه العملية من قبل العالم الروسي الشهير الحائز على جائزة نوبل I.I. Mechnikov وأطلق عليه اسم البلعمة. تم اكتشاف هذه الظاهرة أثناء الدراسة خصائص الحمايةخلايا الدم التي تحبس البكتيريا وتبطل مفعولها. تلتقط الخلية الجزيئات الصغيرة وقطرات السائل عن طريق تشكيل غزوات الغشاء - وتسمى هذه العملية كثرة الخلايا (pinos اليونانية - السائل).
في الواقع ، تعد البلعمة والتضخم نوعين من الظاهرة الشائعة التي تتميز بها معظم الخلايا - الالتقام الخلوي. العملية العكسية للالتقام الخلوي - تسمى إزالة أي مواد ومنتجات من الخلايا بـ exocytosis. على أساس نقل الغشاء ، يتم بناء العملية الكاملة لإفراز وامتصاص الأسرار والهرمونات بواسطة الخلايا. من المهم التأكيد على أن الداخلي والإفراز الخلوي هما عمليتان مستهلكتان للطاقة ، وبالتالي ، هما نوعان من النقل النشط.
وظيفة مستقبلات الغشاء
مبادئ عمل مستقبلات الغشاء.
تتلقى الخلية باستمرار إشارات من البيئة الخارجية حول وجود جزيئات إشارات مختلفة ويجب أن تستجيب بشكل مناسب لهذه الإشارات ، وتنقل المعلومات منها إلى الخلية. لهذا الغرض ، يتم بناء مجمعات مستقبلات خاصة في غشاء البلازما. كقاعدة عامة ، هذه تكوينات معقدة للعديد من جزيئات البروتين (قد يشمل تكوين المستقبلات أيضًا دهون الغشاء والكربوهيدرات).
تحتوي جميع مستقبلات غشاء الخلية المختلفة على عدد من السمات المشتركة:
1. المستقبلات محددة - أي تلتزم فقط بمواد معينة. يتم تحديد خصوصية المستقبل من خلال بنية "الموقع النشط" في جزيئه وقدرة الجزيئات الأخرى على الارتباط بهذا الموقع النشط.
2. تتم عملية الاستقبال ونقل الإشارة إلى الغشاء أو في عمق الخلية مع إنفاق كبير من الطاقة.
3. وفقًا لمبدأ التشغيل ، يمكن تقسيم جميع المستقبلات إلى ثلاث مجموعات: المستقبلات فعل مباشر؛ المستقبلات غير المباشرة والمستقبلات التحفيزية. في الحالة الأولى ، يرتبط الجزيء بجزء المستقبل من المعقد وينقل الإشارة مباشرة إلى القناة الأيونية. في النسخة الثانية ، يرسل جزء المستقبل من المعقد إشارة إلى قناة أيونية في الغشاء أو في عمق الخلية من خلال نظام من البروتينات المساعدة يسمى "الرسل الثاني". في المتغير الثالث ، يتم تنشيط جزء المستقبل من المركب ، بعد التفاعل مع جزيء الإشارة ، ويقوم بوظائف الإنزيم ، مما يؤثر على عمل الخلية.
وظيفة العزل للغشاء. الاتصالات بين الخلايا
في متعددة الخلاياالخلايا على اتصال دائم مع بعضها البعض ، والتي تكتسب أحيانًا ميزات التفاعل الوثيق. يتضح هذا بشكل خاص في الأنسجة التي تحد البيئة الخارجية (تكامل ، أمعاء ، الخطوط الجوية) أو عندما تكون هناك حاجة للاتصال الوثيق بالخلايا من أجل العمل المنسق (العضلات الملساء والقلبية ، الخلايا العصبيةوالألياف).
هناك أربعة أنواع من الوصلات بين الخلايا: الوصلات العازلة (مفترق tigth) ، الوصلات الضيقة (مفترق الفجوة) ، الديسموسومات (desmosome) والوصلات اللاصقة (مفترق الالتصاق).
عزل جهات الاتصال ترقى إلى مستوى أسمائهم - مجمعات البروتينفي أغشية الخلايا المجاورة متصلة بطريقة لا تمر فيها جزيئات وأيونات عبر هذا التلامس. تسمح الوصلات الضيقة بمرور الجزيئات والأيونات الصغيرة ، ويمكن أن يحدث التبادل بين الخلايا من خلالها. على وجه الخصوص ، يتم تكييف التقاطعات الضيقة عن طريق العصبية و خلايا العضلاتلنقل الإشارات الكهربائية. تم تصميم Desmosomes لعقد مجموعة من الخلايا مع بعضها البعض أو لبعض الركيزة الأخرى.
يمكن أن تختفي بسرعة وتعاود الظهور. تم تصميم جهات الاتصال اللاصقة أيضًا لربط الخلايا ببعضها البعض. فهي أكثر قابلية للتغير ويمكن أن تتشكل وتختفي بسرعة حسب الحاجة.
النواة مسؤولة عن تخزين المادة الجينية المسجلة على الحمض النووي ، وتتحكم أيضًا في جميع عمليات الخلية. يحتوي السيتوبلازم على عضيات ، لكل منها وظائفها الخاصة ، على سبيل المثال ، التوليف المواد العضوية، الهضم ، إلخ. وسنتحدث عن المكون الأخير بمزيد من التفصيل في هذه المقالة.
في علم الأحياء؟
تتحدث لغة بسيطة، إنها صدفة. ومع ذلك ، فإنه ليس دائمًا غير قابل للاختراق تمامًا. يُسمح دائمًا بنقل بعض المواد عبر الغشاء.
في علم الخلايا ، يمكن تقسيم الأغشية إلى نوعين رئيسيين. الأول هو غشاء البلازما الذي يغطي الخلية. والثاني هو أغشية العضيات. هناك عضيات لها غشاء واحد أو غشاءان. تشمل الخلايا أحادية الغشاء الشبكة الإندوبلازمية والفجوات والليزوزومات. تنتمي البلاستيدات والميتوكوندريا إلى الغشاءين.
أيضا ، يمكن أن تكون الأغشية داخل العضيات. عادة ما تكون هذه مشتقات من الغشاء الداخلي للعضيات ثنائية الغشاء.
كيف يتم ترتيب أغشية العضيات ذات الغشاءين؟
للبلاستيدات والميتوكوندريا غشاءان. يكون الغشاء الخارجي لكلا العضيتين أملسًا ، لكن الغشاء الداخلي يشكل الهياكل اللازمة لعمل العضو العضوي.
لذلك ، فإن قشرة الميتوكوندريا لها نتوءات داخلية - كرستيات أو تلال. عليهم ، تحدث دورة التفاعلات الكيميائية اللازمة للتنفس الخلوي.
مشتقات الغشاء الداخلي للبلاستيدات الخضراء عبارة عن أكياس على شكل قرص - ثايلاكويدات. يتم جمعها في أكوام - حبوب. يتم دمج الجرانات المنفصلة مع بعضها البعض بمساعدة الصفائح - الهياكل الطويلة التي تشكلت أيضًا من الأغشية.
هيكل أغشية العضيات أحادية الغشاء
هذه العضيات لها غشاء واحد فقط. عادة ما يكون غشاء أملس يتكون من الدهون والبروتينات.
ملامح هيكل غشاء البلازما للخلية
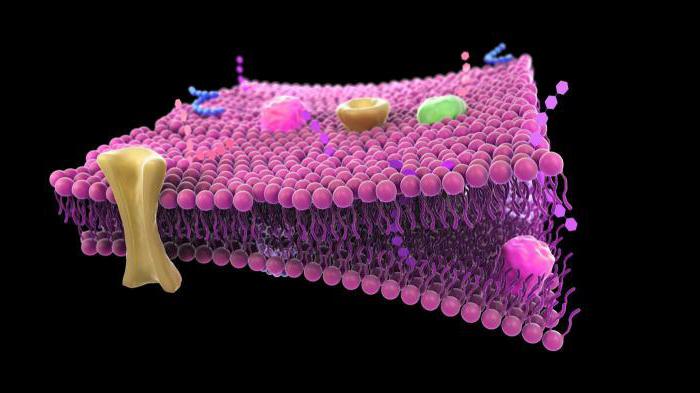
يتكون الغشاء من مواد مثل الدهون والبروتينات. يوفر هيكل غشاء البلازما سمكه من 7-11 نانومتر. يتكون الجزء الأكبر من الغشاء من الدهون. 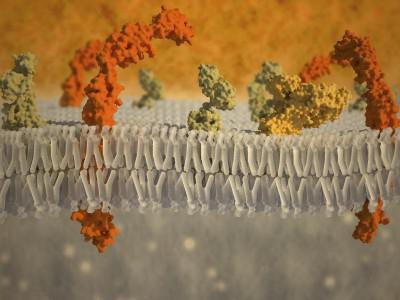
يوفر هيكل غشاء البلازما وجود طبقتين فيه. الأولى عبارة عن طبقة مزدوجة من الدهون الفسفورية ، والثانية طبقة من البروتينات.
دهون غشاء البلازما
تنقسم الدهون التي يتكون منها غشاء البلازما إلى ثلاث مجموعات: الستيرويدات ، والسفينجوفوسفوليبيد ، والجليسيروفوسفوليبيد. يحتوي جزيء الأخير في تركيبته على بقايا كحول الجلسرين ثلاثي الهيدروجين ، حيث يتم استبدال ذرات الهيدروجين لمجموعتين من الهيدروكسيل بسلاسل من الأحماض الدهنية ، ويتم استبدال ذرة الهيدروجين من مجموعة الهيدروكسيل الثالثة ببقايا حمض الفوسفوريك ، والتي بدورها ترتبط بقايا إحدى القواعد النيتروجينية.
يمكن تقسيم جزيء الغليسيروفوسفوليبيد إلى قسمين: الرأس والذيل. الرأس محبة للماء (أي أنها تذوب في الماء) ، وذيولها كارهة للماء (فهي تطرد الماء ولكنها تذوب في مذيبات عضوية). بسبب هذا الهيكل ، يمكن تسمية جزيء الجليسيروفوسفوليبيد amphiphilic ، أي كاره للماء ومحبة للماء في نفس الوقت.
تتشابه الشحميات السفينغوفوسفورية في التركيب الكيميائيلالجليسيروفوسفوليبيد. لكنها تختلف عن تلك المذكورة أعلاه في تكوينها ، فبدلاً من بقايا الجلسرين ، تحتوي على بقايا كحول سفينجوزين. ولجزيئاتها أيضًا رؤوس وذيول.
توضح الصورة أدناه بوضوح بنية غشاء البلازما.
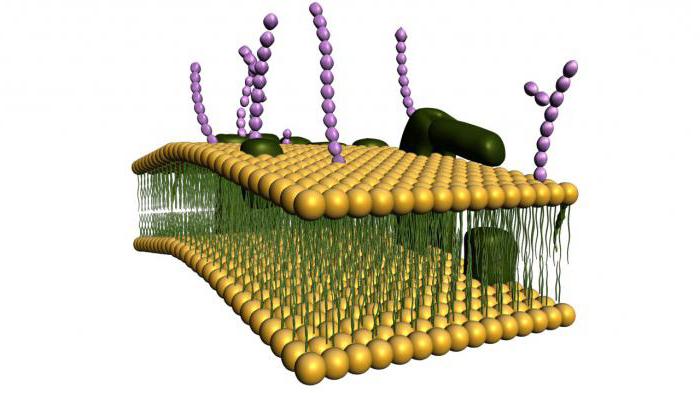
بروتينات غشاء البلازما
أما بالنسبة للبروتينات التي تتكون منها بنية غشاء البلازما ، فهي في الأساس بروتينات سكرية.
اعتمادًا على موقعها في الغلاف ، يمكن تقسيمها إلى مجموعتين: هامشية ومتكاملة. الأولى هي تلك الموجودة على سطح الغشاء ، والثانية هي تلك التي تخترق سمك الغشاء بالكامل وتوجد داخل الطبقة الدهنية.
اعتمادًا على الوظائف التي تؤديها البروتينات ، يمكن تقسيمها إلى أربع مجموعات: الإنزيمات ، والبنية ، والنقل ، والمستقبلات.
جميع البروتينات الموجودة في بنية غشاء البلازما لا ترتبط كيميائياً بالفوسفوليبيدات. لذلك ، يمكنهم التحرك بحرية في الطبقة الرئيسية من الغشاء ، والتجمع في مجموعات ، وما إلى ذلك. ولهذا السبب لا يمكن تسمية بنية الغشاء البلازمي للخلية بالثبات. إنه ديناميكي ، لأنه يتغير طوال الوقت.
ما هو دور غشاء الخلية؟
يسمح هيكل غشاء البلازما له بالتعامل مع خمس وظائف.
الأول والأهم هو تقييد السيتوبلازم. نتيجة لذلك ، تحتوي الخلية على شكل دائموالحجم. يتم ضمان هذه الوظيفة من خلال حقيقة أن غشاء البلازما قوي ومرن.
الدور الثاني هو التوفير بسبب مرونتها ، يمكن أن تشكل أغشية البلازما نتوءات وثنيات عند تقاطعاتها.
الوظيفة التالية جدار الخلية- ينقل. يتم توفيره بواسطة بروتينات خاصة. شكرا لهم المواد المناسبةيمكن نقلها إلى القفص والتخلص منها غير الضرورية.
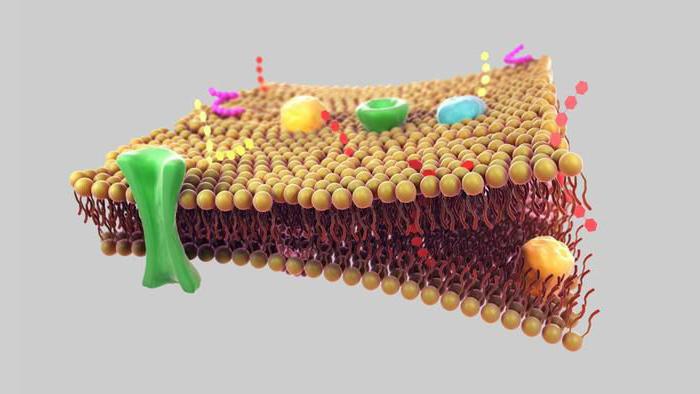
بالإضافة إلى ذلك ، يؤدي غشاء البلازما وظيفة إنزيمية. يتم تنفيذه أيضًا بفضل البروتينات.
والوظيفة الأخيرة هي إرسال الإشارات. نظرًا لحقيقة أن البروتينات تحت تأثير ظروف معينة يمكنها تغيير هيكلها المكاني ، يمكن لغشاء البلازما إرسال إشارات إلى الخلايا.
الآن أنت تعرف كل شيء عن الأغشية: ما هو الغشاء في علم الأحياء ، ما هو ، كيف يتم ترتيب غشاء البلازما والأغشية العضوية ، ما هي الوظائف التي يؤدونها.
المحاضرة رقم 4.
عدد الساعات: 2
غشاء بلازمي
1.
2.
3. الاتصالات بين الخلايا.
1. هيكل غشاء البلازما
غشاء البلازما ، أو غشاء البلازما ، هو سطح محيطي هيكل يحدالخلية من الخارج وتوفير اتصالها بالخلايا الأخرى والبيئة خارج الخلية. لها سمكحوالي 10 نانومتر. من بين أغشية الخلايا الأخرى ، فإن غشاء البلازما هو الأكثر سمكًا. كيميائيا ، غشاء البلازما مجمع البروتين الدهني.المكونات الرئيسية هي الدهون (حوالي 40٪) والبروتينات (أكثر من 60٪) والكربوهيدرات (حوالي 2-10٪).
الدهون مجموعة كبيرةالمواد العضوية التي لها قابلية ضعيفة للذوبان في الماء (مقاومة الماء) وقابلية جيدة للذوبان في المذيبات العضوية والدهون (ألفة للدهون).الدهون التمثيلية الموجودة في غشاء البلازما هي الفوسفوليبيدات والسفينجوميلين والكوليسترول. في زرع الخلايايتم استبدال الكوليسترول بالفيتوستيرول. بواسطة دور بيولوجييمكن تقسيم بروتينات غشاء البلازما إلى بروتينات الإنزيم والمستقبلات والبروتينات الهيكلية.الكربوهيدرات البلازمية هي جزء من غشاء البلازما في حالة ملزمة (الدهون السكرية والبروتينات السكرية).
هو مقبول حاليا بشكل عام نموذج فسيفساء سائل لهيكل الغشاء البيولوجي.وفقًا لهذا النموذج ، يتكون الأساس الهيكلي للغشاء من طبقة مزدوجة من الدهون الفوسفورية المغطاة بالبروتينات. تتقابل ذيول الجزيئات في طبقة مزدوجة ، بينما تظل الرؤوس القطبية بالخارج ، وتشكل أسطحًا محبة للماء. لا تشكل جزيئات البروتين طبقة مستمرة ، فهي تقع في الطبقة الدهنية ، وتغرق في أعماق مختلفة (توجد بروتينات محيطية ، وبعض البروتينات تخترق الغشاء من خلالها ، وبعضها مغمور في الطبقة الدهنية). لا ترتبط معظم البروتينات بدهون الغشاء. يبدو أنها تطفو في "بحيرة دهنية". لذلك ، فإن جزيئات البروتين قادرة على التحرك على طول الغشاء ، أو التجمع في مجموعات ، أو ، على العكس من ذلك ، تشتت على سطح الغشاء. يشير هذا إلى أن غشاء البلازما ليس تشكيلًا ثابتًا ومجمدًا.
خارج البلازما توجد طبقة الغشاء - مركب السكر. سمك هذه الطبقة حوالي 3-4 نانومتر. تم العثور على Glycocalyx في جميع الخلايا الحيوانية تقريبًا. يرتبط بغشاء البلازما مجمع البروتين السكري.تشكل الكربوهيدرات سلاسل طويلة ومتفرعة من السكريات المرتبطة بالبروتينات والدهون في غشاء البلازما. يمكن أن يحتوي جلايكوكاليكس على بروتينات إنزيمية تشارك في الانهيار خارج الخلية للمواد المختلفة. منتجات النشاط الأنزيمي (الأحماض الأمينية ، النيوكليوتيدات ، حمض دهنيإلخ) من خلال غشاء البلازما وتمتصه الخلايا.
يتم تجديد غشاء البلازما باستمرار. يحدث هذا عن طريق ربط الفقاعات الصغيرة من سطحها بالخلية ودمج فجوات من داخل الخلية في الغشاء. وهكذا ، يوجد في الخلية تدفق مستمر لعناصر الغشاء: من غشاء البلازما إلى السيتوبلازم (الالتقام)وتدفق الهياكل الغشائية من السيتوبلازم إلى سطح الخلية (طرد خلوي).في دوران الأغشية ، يتم تعيين الدور الرائد لنظام فجوات الغشاء في مجمع جولجي.
4. وظائف غشاء البلازما. آليات نقل المواد عبر غشاء البلازما. وظيفة المستقبل للبلازما
يؤدي غشاء البلازما عددًا من الوظائف المهمة:
1) حاجز.وظيفة الحاجز لغشاء البلازما هيالحد من الانتشار الحر للمواد من خلية إلى أخرى ، مما يمنعتسرب دوران محتويات الخلية القابلة للذوبان في الماء. لكن منذكيف يجب أن تتلقى الخلية العناصر الغذائية الضرورية ، أنتقسمة المنتجات النهائية الأيضية ، وتنظيم داخل الخلاياتركيزات الأيونات ، ثم شكلت آليات خاصة لنقل المواد عبر غشاء الخلية.
2) ينقل.وظيفة النقل هي ضمان دخول وخروج المواد المختلفة داخل وخارج الخلية. خاصية مهمة للغشاء النفاذية الاختيارية، أو نفاذية. يمر بسهولة في الماء وقابل للذوبان في الماءالغازات وصد الجزيئات القطبية مثل الجلوكوز أوأحماض أمينية.
توجد عدة آليات لنقل المواد عبر الغشاء:
النقل السلبي
النقل النشط
النقل في عبوات غشائية.
النقل السلبي. انتشار -هذه هي حركة جزيئات الوسط ، مما يؤدي إلى انتقالمواد من منطقة يكون تركيزها فيها مرتفعًا إلى منطقة ذات تركيز منخفضنشوئها. أثناء نقل الانتشار ، يعمل الغشاء كحاجز تناضحي. معدل الانتشار يعتمد على القيمةالجزيئات وقابليتها للذوبان النسبي في الدهون. أقل الأوقاتقياسات الجزيئات وكلما زادت قابليتها للذوبان في الدهون (محبة للدهون) ، زادت سرعة تحركها عبر طبقة الدهون الثنائية.يمكن أن يكون الانتشار حيادي(نقل غير مشحونةجزيئات) و وزن خفيف(بمساعدة بروتينات خاصةأنوف). الانتشار الميسر أسرع من الانتشار المحايد.أقصى اختراقالماء لديه القدرةكيف تكون جزيئاته صغيرة وغير مشحونة. انتشار الماء عبر الخلايايسمى الغشاء أوسمو سمك السلورمن المفترض أن في الخليةغشاء للاختراقالماء وبعض الأيونات بشكل أساسيهناك "مسام" خاصة. عددهمصغير ، وقطرهاحوالي 0.3-0.8 نانومتر. ينتشر بسرعة أكبر من خلال الغشاء جيد ، قابل للذوبان في الدهون بسهولةطبقة ثنائية من جزيء ، على سبيل المثال O ، و جزيئات قطبية غير مشحونةلاي قطر صغير (SO ، moشيفين).
نقل الجزيئات القطبية (معالسكريات والأحماض الأمينية) المنفذةيتم تسليمها بمساعدة النقل الغشائي الخاصالبروتينات تسمى نشر الميسر.هذه البروتيناتتوجد في جميع أنواع الأغشية البيولوجية ولكل منها محدد تم تصميم بروتين نيويورك لنقل جزيئات فئة معينةسا. بروتينات النقل عبارة عن غشاء عابر ، حيث تعبر سلسلة البولي ببتيد الخاصة بها طبقة ثنائية الدهون عدة مرات ، وتشكل من خلال الممرات. هذا يضمن نقل محددالمواد من خلال الغشاء دون اتصال مباشر به.هناك فئتان رئيسيتان من بروتينات النقل: السناجب- الناقلون (الناقلون) و تشكيل القناةبروتينات (بيضاءقنوات كي). تحمل البروتينات الحاملة الجزيئات عبر الغشاء عن طريق تغيير تكوينها أولاً.تشكل البروتينات المكونة للقناة أغشية ممتلئة ماء المسام. عندما تكون المسام مفتوحة ، جزيئات من مواد معينة(عادة أيونات غير عضوية ذات حجم وشحنة مناسبة) تمر عبرها. إذا لم يكن لجزيء المادة المنقولة شحنة ، إذن يتم تحديد اتجاه النقل بواسطة تدرج التركيز.إذا كان الجزيء مشحونًا ، فعند نقله ، بالإضافة إلى التدرج ، يخدع التركيز ، تؤثر الشحنة الكهربائية للغشاء أيضًا على (الغشاءمحتمل). عادة ما يتم شحن الجانب الداخلي للبلازما منسلبي فيما يتعلق بالخارج. تسهل إمكانات الغشاء تغلغل الأيونات الموجبة الشحنة في الخلية و يمنع مرور الأيونات سالبة الشحنة.
النقل النشط. النقل النشط هو حركة المواد ضد التدرج الكهروكيميائي. يتم تنفيذه دائمًا عن طريق البروتينات المتحولة.الحمالين وثيقا الصلة زان بمصدر للطاقةجي. في نقل البروتينات chiki لها مؤامرات ملزمة مع النقلمادة tiable.أكثر من هذا القبيل يتواصل tkov مع الأشياءكلما زادت السرعةنمو النقل. يسمى النقل الانتقائي لمادة واحدة uniport. يتم تنفيذ نقل العديد من المواد كوتران أنظمة رياضية. إذا سارت عملية النقل في اتجاه واحد -هذا symportإذا كان في المقابل مضاد للميناء.لذا،على سبيل المثال ، يتم نقل الجلوكوز من السائل خارج الخلية إلى الخلية بطريقة أحادية. نقل الجلوكوز ونا 4 من الأمعاء أونبيبات الكلى ، على التوالي ، في خلايا الأمعاء أو الدم تتم بشكل متوازي ، ويكون نقل C1 ~ و HCO "مضادًا للمنفذ. يفترض من المفترض أنه أثناء النقل ، قابل للانعكاسالتغييرات في الناقل ، مما يسمح بتحريك المواد المتصلة به.
مثال على بروتين ناقل يستخدم للنقلالمواد ، الطاقة المنبعثة أثناء التحلل المائي لـ ATP هينا + -مضخة K + ، وجدت في غشاء البلازما لجميع الخلايا. Na + -K تعمل المضخة على مبدأ الضخ المضاد للمنافذواي نا "من الخلية إلى داخل الخلية مقابل الكهروكيميائية التدرجات. الانحدارنا + يخلق ضغطًا تناضحيًا ويحافظ على حجم الخلية ويضمن نقل السكريات والأحماض الأمينيةnoacids. يستهلك تشغيل هذه المضخة ثلث الطاقة اللازمة للنشاط الحيوي للخلايا.عند دراسة آلية العمل Na + - K + تم تركيب المضخةمن المعروف أنه إنزيم ATPase وإنزيم عبر الغشاء بروتين تيغرال. في وجودنا + و ATP تحت تأثير ATP-يُفصل الفوسفات الطرفي عن ATP ويرتبط بالبقاياحمض الأسبارتيك على جزيء ATPase. جزيء Phos ATPasephorylated ، يغير تكوينه و Na مشتق من الخلايا. بعد الانسحابنا يتم دائمًا نقل K "من الخلية إلى الخلية. للقيام بذلك ، يتم قطع الفوسفات المتصل سابقًا من ATPase في وجود K. ويتم إزالة الفسفرة من الإنزيم ، ويعيد تكوينه ، ويتم" ضخ "K 1 في الخلية.
يتكون ATPase من وحدتين فرعيتين ، كبيرة وصغيرة.تتكون الوحدة الفرعية الكبيرة من آلاف بقايا الأحماض الأمينية ،عبور الطبقة الثنائية عدة مرات. لها عامل حفاز النشاط وقادر على أن تكون فسفرة عكسية و dephosphoمثقوب. وحدة فرعية كبيرة على الجانب السيتوبلازميليس لديها مواقع ملزمةنا + و ATP ، وفي الخارج -مواقع التجليد K + و ouabain. الوحدة الفرعية الصغيرة هيبروتين سكري ووظيفته غير معروفين بعد.
Na + -K المضخة لها تأثير كهربائي. يزيل ثلاثةأيون موجب الشحنةنا و خارج الخلية ويقدم اثنينونتيجة لذلك ، يتدفق تيار عبر الغشاء مكونًا إلكترونًاric بقيمة سالبة في الجزء الداخلي للخلية بالنسبة لسطحها الخارجي.نا "-K + المضخة تنظم حجم الخلية ، وتتحكم في تركيز الموادداخل الخلية ، يحافظ على الضغط الاسموزي ، ويشارك في إنشاء إمكانات الغشاء.
النقل في عبوات غشائية. نقل عبر غشاء الجزيئات الكبيرة (البروتينات والأحماض النووية)يتم تنفيذ الكثير ، والسكريات ، والبروتينات الدهنية) والجزيئات الأخرى من خلال التكوين المتسلسل والاندماج المحيطحويصلات مرتبطة بالغشاء (حويصلات). عملية النقل الحويصلييمر بمرحلتين. في البدايةغشاء الحويصلة و plasmalemmaتلتصق ببعضها البعض ثم تندمج.من أجل مرور المرحلة 2 ، من الضروريdimo بحيث تكون جزيئات الماء أنتمزدحمة من خلال تفاعل طبقات ثنائية الدهون ، والتي تقترب من بعضها البعض حتى مسافة 1-5 نانومتر.يعتبر أن هذه العملية مفعلةخاص بروتينات الانصهار(هممعزولة حتى الآن فقط من الفيروسات). النقل الحويصلي لهميزة مهمة - الجزيئات الكبيرة الممتصة أو المفرزة ،الموجودة في الحويصلات ، عادة لاقابل للمزج مع macromoles الأخرىculae أو عضيات الخلية. بويمكن أن تندمج الحويصلات مع أنواع محددة الأغشية التي توفرتبادل شيفايت للجزيئات الكبيرة بيندو الفضاء خارج الخلية ومحتويات الخلية. بصورة مماثلةيتم نقل الجزيئات الكبيرة من حجرة خلية إلى أخرى.
يسمى نقل الجزيئات الكبيرة والجسيمات إلى الخلية إندو خلوي.في هذه الحالة ، يتم تغليف المواد المنقولة في الشاييتكون غشاء البلازما ، حويصلة (فجوة) ، والتيالذي ينتقل إلى الخلية. حسب حجم الصورةالفقاعات ، هناك نوعان من الالتقام الخلوي - كثرة الخلايا والبلعمة.
كثرة الخلايايوفر امتصاص السوائل والمذابةمواد على شكل فقاعات صغيرة (د = 150 نانومتر). البلعمة -هو امتصاص الجزيئات الكبيرة والكائنات الحية الدقيقةاستدعاء أو شظايا من العضيات والخلايا. في نفس الوقت يتشكلونحويصلات كبيرة أو فاجوسومات أو فجوات (د -250 نانومتر أو أكثر). في وظيفة البلعمة الأولية - شكل من أشكال التغذية. في الثدييات ، يتم تنفيذ وظيفة البلعمة بواسطة الضامة و neutملامح تحمي الجسم من العدوى عن طريق امتصاص الميكروبات الغازية. وتشارك الضامة أيضًا في الاستخدامالخلايا القديمة أو التالفة وشظاياها (في الجسمتستوعب البلاعم البشرية أكثر من 100 إريثريتيد قديم يوميًاالبويضات). يبدأ البلعمة فقط عند تناول الجسيميلتصق بسطح البلعمة وينشط التخصصخلايا مستقبلات ناي. ربط الجسيمات بعناصر محددةتتسبب مستقبلات الغشاء في تكوين الأرجل الكاذبة ، والتييغلف الجاودار الجسيمات ويندمج عند الحواف ويشكل فقاعة -بلعم.تشكيل البلعمة والبلعمة المناسبةيمشي فقط إذا كان في عملية تغليف الجسيماتعلى اتصال دائم بمستقبلات البلازما ، كما لو كانت "راكدةوميض البرق ".
جزء كبير من المادة التي تمتصها الخلية بواسطة endocytosis ، ينتهي في الجسيمات الحالة. بما في ذلك الجسيمات الكبيرةنأمل في البلعمةوالتي تندمج بعد ذلك مع الجسيمات الحالة لتشكل البلعمة.يمتص السائل والجزيئات الكبيرة أثناءكثرة الخلايا ، يتم نقلها في البداية إلى الإندوسومات ، والتيتندمج مع الجسيمات الحالة لتشكيل الجسيمات الداخلية. انا حاضر العديد من الإنزيمات المتحللة للماء الموجودة في الجسيمات الحالةريال عماني تدمير الجزيئات الكبيرة. منتجات التحلل المائي (الأحماض الأمينيةالكثير ، السكريات ، النيوكليوتيدات) من الجسيمات الحالة إلى العصارة الخلوية ،حيث يتم استخدامها من قبل الخلية. معظم مكونات الغشاء تعود الحويصلات الداخلية من البلعوم والداخلية عن طريق خروج الخلايا إلى غشاء البلازما وهناك تختفي مرة أخرىليز. الأهمية البيولوجية الرئيسية للالتقام الخلويهناك إيصال لبنات البناء بسبب داخل الخلايا هضم الجزيئات الكبيرة في الجسيمات الحالة.
يبدأ امتصاص المواد في الخلايا حقيقية النواة فيمناطق cialized من غشاء البلازما ، ما يسمى بنحن X حفر يحدها.على الصور المجهرية الإلكترونيةتبدو الحفر وكأنها غزوات غشاء البلازما ، السيتوبلازمجانب الحصيرة مغطى بطبقة ليفية. طبقة مثلمن شأنه أن يحد ساحة حفر صغيرة malemmas. تحتل الحفر حوالي 2٪ منفوق سطح غشاء الخليةنحن حقيقيات النوى. في غضون دقيقة الحفرة تنمو أعمق وأعمقشيا ، يتم سحبها إلى القفص وبعد ذلك ، يضيق في القاعدة ، ينفصل ،تشكيل حويصلات مهدبة.لقد ثبت أن منغشاء ليفي ماتيtov في غضون دقيقة واحدة تقشرحوالي ربعأغشية على شكل pu يحدهازيركوف. تخسر الفقاعات بسرعة حدودهم والحصول على وسيلةالقدرة على الاندماج مع الجسيم.
قد يكون الالتقام غير محدد(تأسيسي)و محدد(مستقبل).في الالتقام غير النوعيالخلية تتولىتمتص المواد الغريبة تمامًا عنها ، على سبيل المثال ، جزيئات السخام ،الأصباغ. أولاً ، تترسب الجزيئات على الكاليكس البلازما. يعجل بشكل جيد (كثف) علىمجموعات من البروتينات موجبة الشحنة ، حيث يحمل جلايكوكاليكس شحنة سالبة. ثم يتغير شكل الخليةأغشية. يمكن أن تغرق ، وتشكل الغزوات(الانغلاف) ، أو العكس ، شكل النواتج ،التي يبدو أنها تتراكم ، وتفصل الأحجام الصغيرة بيئة سائلة. تشكيل الغزوات هو أكثر مميزةلخلايا الظهارة المعوية والأميبات والنمو - للبلعمات و الليفية. يمكن منع هذه العمليات عن طريق المثبطاتعمليه التنفس. الحويصلات الناتجة هي جسيمات داخلية أولية يمكن أن تندمجبعضها البعض ، تتزايد في الحجم. في المستقبل سوف ينضمون تختلط مع الجسيمات الحالة ، وتتحول إلى جسيم داخلي - هضمفجوة جديدة. شدة كثرة الخلايا غير النوعية في المرحلة السائلة تصل إلىعالية بشكل كبير. تشكل البلاعم ما يصل إلى 125 ، والخلايا الظهارية ناعماالأمعاء الدقيقة تصل إلى ألف دبوس في الدقيقة. تؤدي وفرة الصنوبر إلى حقيقة أن البلازما تنفق بسرعة على تكوين متعددةوجود فجوات صغيرة. انتعاش الغشاء سريع جدا.Tro أثناء إعادة التدوير في عملية طرد الخلايا بسبب عودة واkuoles وإدماجها في البلازما. في الضامة ، كل البلازمايتم استبدال الغشاء الكلاسيكي في غضون 30 دقيقة ، وفي الخلايا الليفية في غضون ساعتين.
طريقة أكثر فاعلية للامتصاص من السوائل خارج الخليةالجزيئات الكبيرة الخاصة بالعظام هي en محدد docytosis(بوساطة المستقبلات). الجزيئات الكبيرة في هذاترتبط بالمستقبلات التكميلية على السطحتتراكم الخلايا في الحفرة المتاخمة ، ثم تشكل جسيمًا داخليًا ، تنغمس في العصارة الخلوية. يضمن الالتقام الخلوي للمستقبلات تراكم جزيئات معينة في مستقبلها.الجزيئات التي ترتبط على سطح البلازما بالمستقبلطارة تسمى يجند. بمساعدة المستقبل يعتبر الالتقام الخلوي في العديد من الخلايا الحيوانية هو الامتصاصالكوليسترول خارج الخليةبيئة.
يشارك غشاء البلازما في إزالة المواد من الخلية (خروج الخلايا). في هذه الحالة ، تقترب الفجوات من غشاء البلازما. عند نقاط التلامس ، تندمج البلازما وغشاء الفجوة وتدخل محتويات الفجوة إلى البيئة.في بعض الأوليات ، يتم تحديد المواقع الموجودة على غشاء الخلية لإخراج الخلايا مسبقًا. لذلك ، في غشاء البلازما تحتوي بعض الشركات الهدبية على مناطق معينة ذات الترتيب الصحيح للكريات الكبيرة من البروتينات المتكاملة. فيالأكياس المخاطية و trichocysts من ciliates جاهزة تمامًا للإفراز ، في الجزء العلوي من plasmalemma يوجد كورولا من الكريات المتكاملةالبروتينات. هذه المقاطع من غشاء الأكياس المخاطية و trichocysts soprikaتلتصق بسطح الخلية.لوحظ خروج خلوي غريب في العدلات. هم spoقادرة على إطلاقها في البيئة في ظل ظروف معينةتفجير الجسيمات الحالة الخاصة بك. في بعض الحالات ، تتشكل نواتج صغيرة من غشاء البلازما المحتوي على الجسيمات الحالة ، والتي تنفصل بعد ذلك وتنتقل إلى البيئة. في حالات أخرى ، هناك غزو للبلازما في أعماق الخلية والتقاطها للجسيمات الحالة الموجودةلا شيء بعيدًا عن سطح الخلية.
تتم عمليات الالتقام الخلوي والإفراز الخلوي بمشاركة نظام المكونات الليفية من السيتوبلازم المرتبط بالبلازما.
وظيفة المستقبل للبلازما. هذا من الرئيسي ، الشامل لجميع الخلايا ، هو reوظيفة المستقبل للبلازما. يحدد التفاعلالخلايا مع بعضها البعض ومع البيئة.
يمكن تمثيل المجموعة الكاملة من التفاعلات المعلوماتية بين الخلايا بشكل تخطيطي كسلسلة متتاليةتفاعلات استجابة المرسال الثانية لمستقبل الإشارة (مفهوم إشارة الاستجابة).تنقل الإشارات المعلومات من خلية إلى أخرىجزيئات nye التي يتم إنتاجها في بعض الخلايا وخاصةيؤثر جسديًا على الآخرين الحساسين للإشارة (الخلايا مي شيني). جزيء الإشارة - وسيط أساسيربطتتفاعل etsya مع المستقبلات الموجودة على الخلايا المستهدفة فقط لإشارات معينة. جزيئات الإشارة - يجند-تقترب من مستقبلها مثل مفتاح القفل. يجند-mi لمستقبلات الغشاء (مستقبلات البلازما) هوالجزيئات المحبة للماء ، وهرمونات الببتيد ، والوسائط العصبيةتوري ، السيتوكينات ، الأجسام المضادة ، والمستقبلات النووية - الدهون الجزيئات الذائبة ، الستيرويد وهرمونات الغدة الدرقية ، فيتامين دكمستقبلات على القمةيمكن أن تعمل الخلايا كبروتيناتأغشية أو عناصر تحلل المخاليطكاليفورنيا - السكريات والبروتينات السكرية.ويعتقد أن حساسة للفصل مناطق المواد ، مبعثرسان على طول سطح الخلية أوفي شكل شرائط صغيرة. نعم ، فيسطح الخلايا بدائية النواةوالخلايا الحيوانية هناك حدودعدد ثابت من الأماكن التي يمكنهم من خلالهاربط الجزيئات الفيروسية. ميميالسناجب (الناقلون والقاناly) تعلم وتفاعل ونقلارتداء مواد معينة فقط.مستقبلات الخلايا متورطة فيانتقال الإشارات من سطح الخلية إلى الداخل.التنوع والخصوصيةخندق من المستقبلات على سطح الخليةيؤدي إلى نظام معقد للغايةنحن علامات للتمييزالخلايا الخاصة من الغرباء. خلايا مماثلةتتفاعل مع بعضها البعض ، يمكن أن تلتصق أسطحها ببعضها البعض (الاقتران فيالبروتوزوا ، تكوين الأنسجة في الكائنات متعددة الخلايا). الخلايا لا تدركعلامات مشتركة ، وكذلك تلك التي تختلف فيالبورون من المحدداتممزقة أو مرفوضة.عند تكوين مركب مستقبلات يجند ،بروتينات الغشاء: محول البروتين ، محسن البروتين.نتيجة لذلك ، يغير المستقبل شكله وتفاعلهلا مع السلائف الخلوية للرسول الثانيكا - رسول.يمكن أن تكون الرسل مؤينة الكالسيوم والفوسفوليبلـ C ، adenylate cyclase ، guanylate cyclase. تأثر الرسولتنشيط الإنزيمات المشاركة في التوليف أحادي الفوسفات الدوري - AMPأو HMF.هذا الأخير يغير الأصلوجود نوعين من إنزيمات البروتين كينيز في سيتوبلازم الخلية ، مما يؤدي إلى فسفرة العديد من البروتينات داخل الخلايا.
تشكيل cAMP الأكثر شيوعًا ، تحت تأثيريزيد إفراز عدد من الهرمونات - هرمون الغدة الدرقية ، والكورتيزون ، والبروجسترون ، ويزيد انهيار الجليكوجين في الكبد والعضلات ،معدل ضربات القلب والقوة ، تآكل العظم ، عكس امتصاص الماء في أنابيب النيفرون.
إن نشاط نظام adenylate cyclase مرتفع للغاية - يؤدي تخليق cAMP إلى زيادة الإشارة بمقدار عشرة آلاف.
تحت تأثير cGMP ، إفراز البنكرياس للأنسولين ، الهيستامين بواسطة الخلايا البدينة ، السيروتونينالبيدات ، يتم تقليل أنسجة العضلات الملساء.
في كثير من الحالات ، عند تكوين مجمع مستقبلات يجندهناك تغيير في إمكانات الغشاء ، والذي بدوره يؤدي إلى تغيير في نفاذية غشاء البلازما والتمثيل الغذائيبعض العمليات في الخلية.
يحتوي غشاء البلازما على مستقبلات محددة توري الاستجابة للعوامل الفيزيائية. لذلك ، في البكتيريا الضوئية ، توجد الكلوروفيل على سطح الخلية ،يتفاعل مع الضوء. في الحيوانات الحساسة للضوء في البلازمايوجد نظام كامل لبروتينات مستقبلات الفوجور في الغشاء الكاوي -رودوبسين ، بمساعدة من المنبه الضوئي يتحولإلى إشارة كيميائية ثم نبضة كهربائية.
3. الاتصالات بين الخلايا
في الحيوانات متعددة الخلايا ، تشارك البلازما في التكوين اتصالات بين الخلاياتوفير تفاعلات بين الخلايا. هناك عدة أنواع من هذه الهياكل.
§ اتصال بسيط.تم العثور على اتصال بسيط بين غالبية الخلايا ذات الأصول المختلفة المجاورة لبعضها البعض. يمثل تقارب أغشية البلازما للخلايا المجاورة على مسافة 15-20 نانومتر. في هذه الحالة ، يحدث تفاعل طبقات الكاليكس في الخلايا المجاورة.
§ اتصال ضيق (مغلق). مع مثل هذا الاتصال ، تكون الطبقات الخارجية لغشاءتي البلازما قريبة قدر الإمكان. التقارب كثيف لدرجة أن هناك نوعًا من دمج أقسام من أغشية البلازما لخليتين متجاورتين. لا يحدث اندماج الأغشية على كامل منطقة التلامس المحكم ، ولكنه عبارة عن سلسلة من نقاط التقاء الأغشية. يتمثل دور الاتصال الوثيق في توصيل الخلايا ببعضها البعض ميكانيكيًا. هذه المنطقة غير قابلة للاختراق بالنسبة للجزيئات الكبيرة والأيونات ، وبالتالي فهي تغلق وتحدد الفجوات بين الخلايا (ومعها البيئة الداخلية للجسم) من البيئة الخارجية.
§ رقعة التصاق ، أو ديسموسوم. الديسموسوم هو مساحة صغيرة يصل قطرها إلى 0.5 ميكرومتر. في منطقة الديسموسوم على جانب السيتوبلازم ، توجد منطقة من الألياف الرقيقة. يتمثل الدور الوظيفي للديسموسومات بشكل أساسي في الاتصال الميكانيكي بين الخلايا.
§ اتصال الفجوة ، أو الرابطة. مع هذا النوع من التلامس ، يتم فصل أغشية البلازما للخلايا المجاورة بفجوة من 2-3 نانومتر على مسافة 0.5-3 ميكرومتر. في هيكل البلازمات توجد مجمعات بروتينية خاصة (connexons). يتم معارضة أحد الروابط الموجودة على غشاء البلازما للخلية بدقة من خلال كونيكسون على غشاء البلازما للخلية المجاورة. نتيجة لذلك ، يتم تشكيل قناة من خلية إلى أخرى. يمكن أن تتقلص Connexons ، وتغير قطر القناة الداخلية ، وبالتالي تشارك في تنظيم نقل الجزيئات بين الخلايا. تم العثور على هذا النوع من الاتصال في جميع مجموعات الأنسجة. يتمثل الدور الوظيفي لمفرق الفجوة في نقل الأيونات والجزيئات الصغيرة من خلية إلى أخرى. لذلك ، في عضلة القلب ، ينتقل الإثارة ، التي تعتمد على عملية تغيير نفاذية الأيونات ، من خلية إلى أخرى عبر الرابطة.
§ اتصال متشابك ، أو المشبك. نقاط الاشتباك العصبي هي مناطق تلامس بين خليتين متخصصتين في انتقال الإثارة أو التثبيط في اتجاه واحد من عنصر إلى آخر. هذا النوع من الاتصال هو سمة من سمات النسيج العصبي ويحدث بين اثنين من الخلايا العصبية وبين الخلايا العصبية وبعض العناصر الأخرى. يتم فصل أغشية هذه الخلايا عن طريق مساحة بين الخلايا - شق متشابك يبلغ عرضه حوالي 20-30 نانومتر. يسمى الغشاء الموجود في منطقة التلامس المشبكي لخلية واحدة قبل المشبكي ، بينما يسمى الآخر - بعد المشبكي. بالقرب من الغشاء قبل المشبكي ، تم الكشف عن عدد كبير من الفجوات الصغيرة (الحويصلات المشبكية) التي تحتوي على الناقل العصبي. في وقت مرور النبض العصبي ، تقوم الحويصلات المشبكية بإخراج الناقل العصبي إلى الشق المشبكي. يتفاعل الوسيط مع مواقع المستقبلات لغشاء ما بعد المشبكي ، مما يؤدي في النهاية إلى انتقال النبضات العصبية. بالإضافة إلى نقل النبضات العصبية ، توفر المشابك اتصالًا صارمًا بين أسطح خليتين متفاعلتين.
§ المتصورة.تم العثور على هذا النوع من الاتصال بين الخلايا في النباتات. هي عبارة عن قنوات أنبوبية رفيعة تربط خليتين متجاورتين. يبلغ قطر هذه القنوات عادة 40-50 نانومتر. تمر جزيئات Plasmodesmata عبر جدار الخلية الذي يفصل الخلايا. في الخلايا الشابة ، يمكن أن يكون عدد البيانات الوصفية عالية جدًا (يصل إلى 1000 لكل خلية). مع شيخوخة الخلايا ، ينخفض عددها بسبب التمزق مع زيادة سمك جدار الخلية. يتمثل الدور الوظيفي للوسائط البلازمية في ضمان الدوران بين الخلايا للمحاليل التي تحتوي على العناصر الغذائية والأيونات والمركبات الأخرى. تصيب Plasmodesmata الخلايا بفيروسات النبات.
الهياكل المتخصصة لغشاء البلازما
تشكل البلازما في العديد من الخلايا الحيوانية نواتج من هياكل مختلفة (ميكروفيلي ، أهداب ، سوط). غالبًا ما توجد على سطح العديد من الخلايا الحيوانية ميكروفيلي.هذه النتوءات من السيتوبلازم ، التي تحدها البلازما ، لها شكل أسطوانة ذات قمة مستديرة. Microvilli هي خاصية مميزة للخلايا الظهارية ، ولكنها توجد أيضًا في خلايا الأنسجة الأخرى. يبلغ قطر الميكروفيلي حوالي 100 نانومتر. عددهم وطولهم يختلفون عن أنواع مختلفةالخلايا. تكمن أهمية الميكروفيلي في زيادة كبيرة في مساحة سطح الخلية. هذا مهم بشكل خاص للخلايا المشاركة في الامتصاص. لذلك ، في ظهارة الأمعاء ، يوجد ما يصل إلى 2x10 8 ميكروفيلي لكل 1 مم 2 من السطح.
[المحاضرة السابقة] [جدول المحتويات] [المحاضرة التالية]